









-
Corticioid and hydnoid fungi represent important groups of wood-inhabiting fungi belonging to Basidiomycota[1]. The basidiomata of corticioid fungi are thin, crust-like, or membranaceous in appearance, soft, leathery, or hard in texture, and variable in color. This group of fungi typically grows on wood or soil, and is widely distributed in forest ecosystems from tropical to temperate regions worldwide. Most corticioid species are classified into the orders Agaricales, Atheliales, Auriculariales, Cantharellales, Corticiales, Gloeophyllales, Hymenochaetales, Polyporales, Russulales, Thelephorales, and Trechisporales[2−7]. The morphological characteristics of hydnoid fungi are highly diverse, ranging from resupinate, pileate to stipitate. However, they share a common morphological feature: their basidiomata possess odontioid to tooth-like spines[1], which facilitates spore release. Most hydnoid fungi are found in the orders Auriculariales, Cantharellales, Hymenochaetales, Polyporales, Russulales, and Thelephorales[1,7−9].
Thelephorales was established by Oberwinkler in 1976 and, for a long time, consisted of two families: Thelephoraceae and Bankeraceae[10−13]. Thelephoraceae was introduced by Chevallier in 1826 in the context of the regional flora of Paris, with the type genus Thelephora[14]. It was defined as encompassing fungi that possess resupinate, effuse-reflexed, or coralloid basidiomata, hyaline to coloured basidiospores with warts or echinuli[8]. Donk[10] originally separated Bankeraceae from Thelephoraceae based on spore ornamentation and distinctive odour. Species of Bankeraceae are characterized by stipitate, hydnoid or poroid basidiomata, as well as hyaline to brown basidiospores with warts[15,16]. Furthermore, certain species within Thelephorales exhibit considerable macroscopic polymorphism, which is strongly influenced by environmental factors, or results from typically growing around obstacles, resulting in highly variable and irregular shapes[16]. Therefore, relying solely on morphological characteristics is far from sufficient for distinguishing and identifying species of Thelephorales. Based on multi-gene phylogenetic analyses, Song et al. proposed that Thelephorales is divided into six clades at the family level: Bankeraceae, Lenzitopsidaceae, Polyozellaceae, Sarcodonaceae, Thelephoraceae, and Tomentellopsidaceae[13]. These families include 11 genera: Amaurodon, Boletopsis, Hydnellum, Lenzitopsis, Neosarcodon, Odontia, Phellodon, Polyozellus, Sarcodon, Thelephora, and Tomentellopsis[13]. Additionally, Corneroporus was excluded from the phylogenetic analyses due to the lack of available sequence data[13]. However, based on its segregation from Boletopsis[17], it was retained as a distinct genus within Sarcodonaceae[13]. Notably, Jülich[12] was the first to establish two families: Lenzitopsidaceae (type genus: Lenzitopsis), and Boletopsidaceae (type genus: Boletopsis). Subsequently, Singer et al.[18] proposed the family Sarcodontaceae, with Sarcodon designated as its type genus. Although Bankeraceae[10] predates Boletopsidaceae[12], the name Bankeraceae is currently applied to a clade comprising Phellodon and Amaurodon[13]. In accordance with the priority principle stipulated in the International Code of Nomenclature for algae, fungi, and plants (ICN), Boletopsidaceae is the only valid and correct name for the family that includes Boletopsis—a conclusion derived directly from the ICN's emphasis on taxonomic priority. The Sarcodontaceae can only be legitimately adopted if Boletopsis is explicitly excluded from the family. Correspondingly, Lenzitopsidaceae Jülich represents the correct name for this family.
The order Hymenochaetales was established with the type family Hymenochaetaceae[19]. Species within Hymenochaetales exhibit high morphological diversity. The first comprehensive molecular study on homobasidiomycetes indicated that the Hymenochaetales proposed by Oberwinkler required a broader interpretation[20]. Since then, the taxonomic status of Hymenochaetales has been thoroughly investigated through molecular phylogenetic and morphological studies[21−31]. The familial classification of Hymenochaetales has been continually emended, with a total of 14 family names successively proposed within this order[32,33]. At the generic level, Hymenochaetales comprises 84 genera, among which approximately 20 genera have no confirmed position in any family[31−33]. Many families within Hymenochaetales are small in terms of species numbers, with several being monogeneric[34].
Corticioid and hydnoid fungi belonging to Hymenochaetales and Thelephorales exhibit diverse trophic modes, including saprotrophy, parasitism, and symbiosis, and play vital ecological roles in forest ecosystems[33,35−38]. The Thelephorales are globally distributed, with a particular prevalence in temperate to tropical forests. Within these habitats, they primarily function as ectomycorrhizal fungi or saprobes associated with both gymnosperms and angiosperms, typically occurring on the ground, underneath fallen trunks, and branches of woody plants[39−51]. Wood-decaying fungi can decompose cellulose, hemicellulose and lignin in the plant cell walls, releasing nutrients and facilitating the recycling of elements such as carbon, nitrogen, and phosphorus[52−54]. Ectomycorrhizal fungal species form symbiotic associations with plants by developing ectomycorrhiza, which enables nutrient exchange[44,45,47,50]. As a result, these fungi play a crucial role in energy flow and nutrient cycling in forest ecosystems. Certain corticioid and hydnoid fungi, particularly ectomycorrhizal species, are sensitive to environmental pollution and soil nutrient status. Declines in their abundance have been observed in some European localities[55−60]. Consequently, hydnoid fungi have been designated as priority conservation targets in numerous countries, with several species now listed on national Red Lists[61−63].
In Southwest China, species within Thelephorales and Hymenochaetales, such as Hydnellum, Inonotus, Phellinus, Sarcodon, and Thelephora, hold significant economic value due to their edibility and medicinal properties[64,65]. Market statistics indicate that thousands of tons of Sarcodon spp. and Hydnellum spp. are sold in the free markets in Sichuan Province (China) each year[64]. With the deepening understanding of hydnoid fungi, the research on the medicinal value and active substances of certain species has gradually attracted more attention[66−75]. Some species of Thelephorales not only have edible value but also possess important medicinal functions, including cholesterol-lowering, antioxidant, anti-inflammatory, antitumor and immune enhancement, etc.[76,77]. Certain species within Hymenochaetales also exhibit medicinal properties[78−84].
Southern China presents a geographically intriguing area for research, as it encompasses several regions with distinct geologic histories, and biogeographically belongs to tropical and subtropical zones. This region features a pronounced topographical gradient, descending from elevated western terrains to lower eastern landscapes. The western part is dominated by plateaus and basins, while the eastern part consists predominantly of plains, low mountains, and hills, resulting in an overall topography with a distinct stepped distribution. Notably, the southern region of the Qinghai-Xizang Plateau is geographically located in southern China, and its significant altitudinal variations contribute to an exceptionally diverse range of climatic types[85,86]. China's tropical and subtropical forests harbor unique and rich biodiversity while providing vital ecosystem services[87]. The Qinghai-Xizang Plateau exerts a dual influence on East Asian climate systems: it obstructs westerly atmospheric circulation and amplifies monsoon intensity, which has facilitated the development of characteristic subtropical rainforest ecosystems across extensive areas south of the Qinling Mountains-Huaihe River line in China's subtropical zone. Additionally, this region encompasses a biodiversity hotspot (i.e., the mountains of southwest China), and one of the three major karst regions globally[87,88]. The abundant vegetation and diverse geographical environment create favorable conditions for the growth of corticioid and hydnoid fungi. In recent years, several studies have been conducted on corticioid and hydnoid fungi in subtropical and southern temperate forests in China, leading to the publication of numerous species[35,36,38,64,89−101]. However, the taxonomic and phylogenetic relationships among taxa of corticioid and hydnoid fungi remain insufficiently understood.
During the investigation of corticioid and hydnoid fungal specimens from the subtropical to temperate plateau region of China, a significant number of valuable specimens were collected. In this study, these specimens were analyzed using macro-morphology, microscopic examinations, ultrastructural analyses, and multi-gene molecular phylogenetic analysis (including ITS, nLSU, nSSU, and mtSSU). Based on these analyses, 30 new species are proposed, belonging to the genera Hydnellum, Hymenochaete, Lyomyces, Neosarcodon, Odontia, Peniphorella, Phellodon, Sarcodon, Thelephora, and Xylodon. Three keys to the species of Hydnellum, resupinate and upright Thelephora from China were provided.
-
Fresh basidiomata of the corticioid and hydnoid fungi were collected from Chongqing Municipality, Fujian Province, Guangdong Province, Guangxi Zhuang Autonomous Region, Guizhou Province, Hunan Province, Sichuan Province, Xizang Autonomous Region, Yunnan Province, and Zhejiang Province in China (Fig. 1). The specimens were photographed in situ, and detailed macroscopic characteristics of the fresh samples were recorded. They were then dried using an electric food dehydrator at 40 °C, sealed in envelope bags and deposited in the Herbarium of the Institute of Applied Ecology, Chinese Academy of Sciences (IFP).
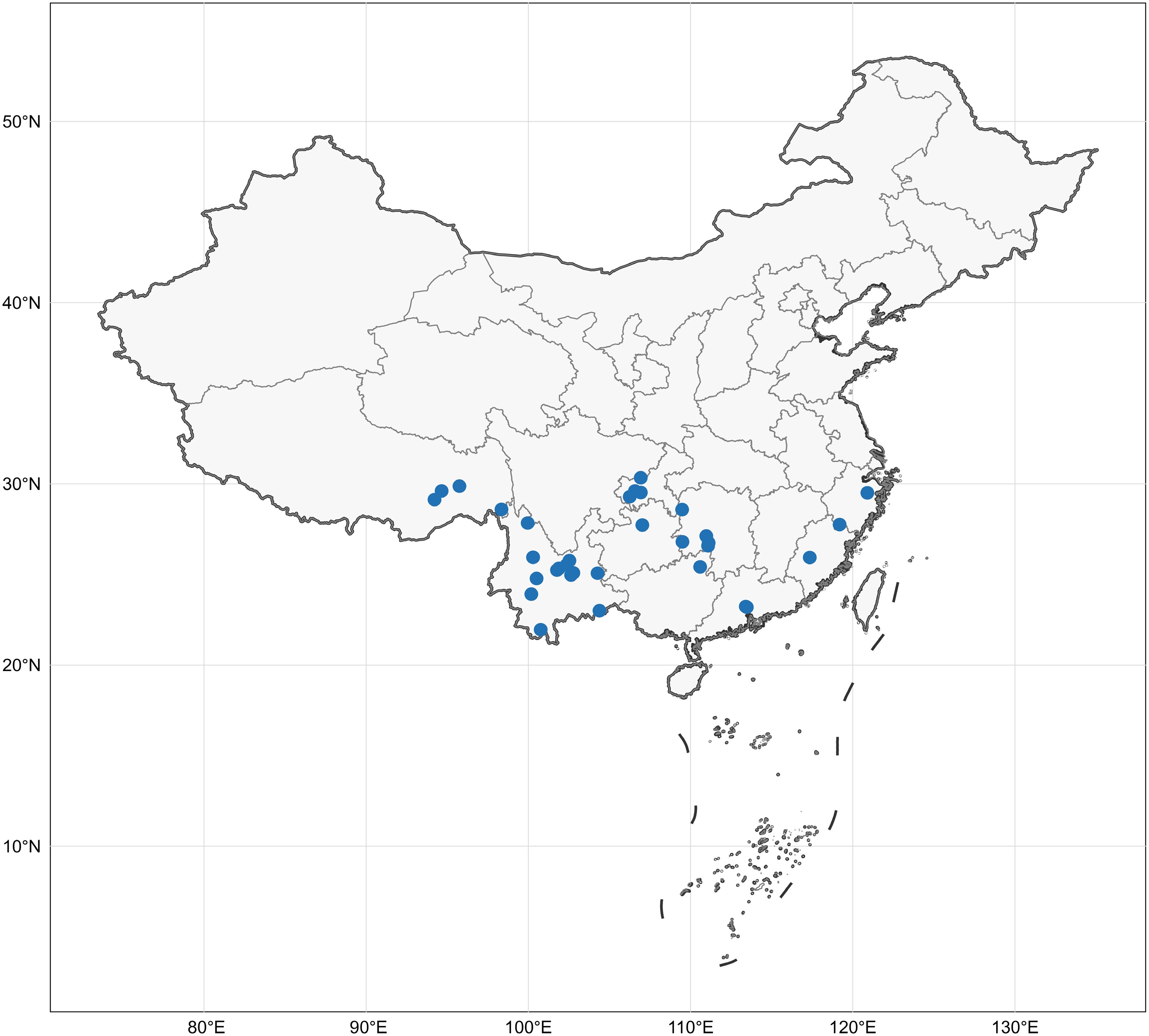
Figure 1.
The collection localities of all specimens in this study. Map source: Standard map approved by the Ministry of Natural Resources of China, Approval No. GS(2024)0650.
Morphological studies
-
Macroscopic descriptions of collected specimens were based on fresh basidiomata. Microscopic procedures followed the method described by Yuan et al.[102]. Dried material was mounted in 5% aqueous KOH, and Melzers reagent to test for any amyloid and/or dextrinoid reactions (Melzer's reagent: 1.5 g KI [potassium iodide], 0.5 g I [crystalline iodine], 22 g chloral hydrate, distilled water 20 mL). The following abbreviations are used in the text: KOH = 5% potassium hydroxide; L = mean spore length (arithmetic average of all spores); W = mean spore width (arithmetic average of all spores); Q = variation in the ratios of L/W between specimens studied, and n = total number of spores measured from a given number of specimens. Sections were examined at magnifications up to × 1,000 using a Nikon Eclipse E600 microscope (Tokyo, Japan) with phase-contrast illumination, and dimensions were estimated with an accuracy of 0.1 μm. Microscopic drawings were prepared with the aid of a drawing tube. Spore measurements excluded the apiculus, and 5% of the measurements at each end of the range are given in parentheses. The spore measurements were made with a Nikon SMZ 645 compound microscope. Colour codes refer to Kornerup & Wanscher[103].
DNA extraction, amplification, and sequencing
-
Phire Plant Direct PCR Kit (Thermo Fisher Scientific, Waltham, MA, USA) procedures were used to extract total genomic DNA from the basidiomata. Polymerase chain reactions (PCR) were performed on a Bio-Rad T100TM Thermal Cycler (Bio–RAD Inc., Hercules, CA, USA). The internal transcribed spacer region (ITS) was amplified with primer pairs ITS1 and ITS4[104]; for the large subunit of nuclear ribosomal RNA gene (nLSU), LROR and LR7 were used[105]. The small subunit of nuclear ribosomal RNA gene (nSSU) was amplified with primer pairs NS1 and NS4[104]. The mitochondrial SSU (mtSSU) region was amplified with primer pairs MS1 and MS2[104].
The final PCR volume was 25 μl; each tube contained 1 μl each primer, 1 μl extracted DNA, 10 μl ddH2O, and 12.5 μl T5 Super PCR Mix (containing Taq polymerase, dNTP and Mg2+, Beijing Tisingke Biotech Co., Ltd., Beijing, China). The PCR procedure for ITS was: initial denaturation at 95 °C for 3 min, followed by 34 cycles of denaturation at 94 °C for 40 s, annealing at 54 °C for 45 s, and extension at 72 °C for 1 min, and a final extension at 72 °C for 10 min. The PCR procedure for nLSU was: initial denaturation at 94 °C for 1 min, followed by 34 cycles of denaturation at 94 °C for 30 s, annealing at 50 °C for 1 min, and extension at 72 °C for 1.5 min, and a final extension at 72 °C for 10 min. The PCR procedure for nSSU was: initial denaturation at 95 °C for 3 min, followed by 34 cycles of denaturation at 94 °C for 40 s, annealing at 55 °C for 45 s, and extension at 72 °C for 1 min, and a final extension at 72 °C for 10 min. The PCR procedure for and mtSSU was an initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 1 min, annealing at 55 °C for 2 min, and extension at 72 °C for 1.5 min, and a final extension at 72 °C for 10 min. All amplified PCR products were estimated visually with 1.4% agarose gels stained with ethidium bromide and sequenced at the Beijing Genomics Institute (BGI) with the same primers. All new sequences generated in this study were submitted to GenBank. Duplicate PCR and sequencing were performed for all new species described from a single specimen, which effectively negated the possibility of sequencing errors.
Phylogenetic analyses
-
Sequences for phylogenetic analysis were obtained from GenBank (
www.ncbi.nlm.nih.gov/genbank ) using the BLAST option and downloaded (Supplementary Table S1). The sequences were aligned with MAFFT v.7 (https://mafft.cbrc.jp/alignment/server/index.html ), after which the alignments were manually corrected using MEGA v. 7.0[106,107]. The dataset was initially aligned and later, ITS, nLSU, nSSU, and mtSSU sequences were combined using using BioEdit v. 7.0.9[108].Phylogenetic analyses for each dataset were conducted using Bayesian Inference (BI) analysis and Maximum Likelihood (ML) methods. All characters were weighted, and gaps were treated as missing data. BI analysis with MrBayes v. 3.2.7[109] implemented the Markov Chain Monte Carlo (MCMC) technique. Four simultaneous Markov chains were run with 15, 10, 5, 5 million generations for the four datasets respectively, starting from random trees and keeping one tree every 100th generation until the average standard deviation of split frequencies was below 0.01. The value of burn-in was set to discard 25% of trees when calculating the posterior probabilities. Bayesian Posterior Probabilities (BPP) were obtained from the 50% majority rule consensus of the trees kept. An ML analysis used the same datasets as the BI analysis, and was performed in RAxML v. 8.2.4[110]. The best tree was obtained by performing 1,000 rapid bootstrap inferences, followed by a thorough search for the most likely tree[111]. Phylogenetic trees were visualised with FigTree v. 1.4.2 (
http://tree.bio.ed.ac.uk/software/figtree , accessed on 20 April, 2025), and additional layout was carried out using Adobe Illustrator CS v. 5. -
Thelephorales Corner ex Oberw.
Bankeraceae Donk
Phellodon phylogeny based on combined ITS, nLSU, and nSSU sequence data (Fig. 2)
The aligned dataset comprised 79 specimens, representing 42 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for the combined ITS + nLSU + nSSU (Fig. 2) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 3,545 characters, of which 2,872 characters are constant, 105 are variable and parsimony uninformative, and 568 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the four genes; the models for ITS, nLSU, and nSSU were GTR + I + G, with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.004752. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.
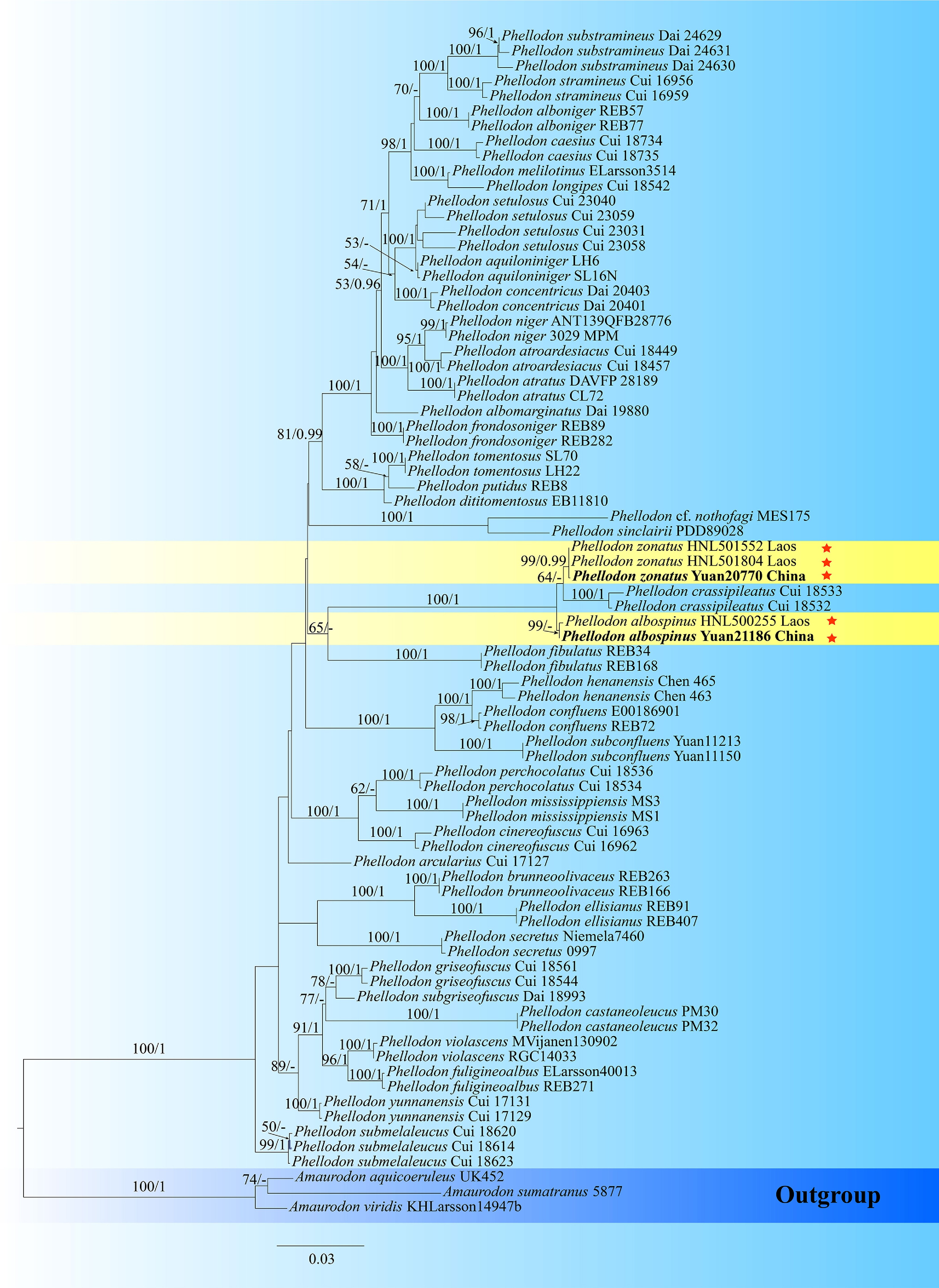
Figure 2.
Maximum likelihood tree illustrating the phylogeny of Phellodon and related genera in Thelephorales based on ITS + nLSU + nSSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 50%, and Bayesian posterior probabilities more than 0.95 respectively. Specimens examined are in bold, and new species are marked with red stars.
Boletopsidaceae Bondartsev & Singer ex Jülich
Hydnellum phylogeny based on combined ITS, nLSU, and nSSU sequence data (Fig. 3)
The aligned dataset comprised 187 specimens, representing 86 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for combined ITS + nLSU + nSSU (Fig. 3) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 3,715 characters, of which 2,520 characters are constant, 167 are variable and parsimony uninformative, and 1,028 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the four genes; the models for ITS, nLSU, and nSSU were GTR + I + G, with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.038592. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.
Neosarcodon and Sarcodon phylogeny based on combined ITS, nLSU, and nSSU sequence data (Fig. 4)
The aligned dataset comprised 86 specimens, representing 55 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for combined ITS + nLSU + nSSU (Fig. 4) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 3,379 characters, of which 2,349 characters are constant, 220 are variable and parsimony uninformative, and 810 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the four genes; the models for ITS, nLSU and nSSU were GTR + I + G, with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.006577. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.
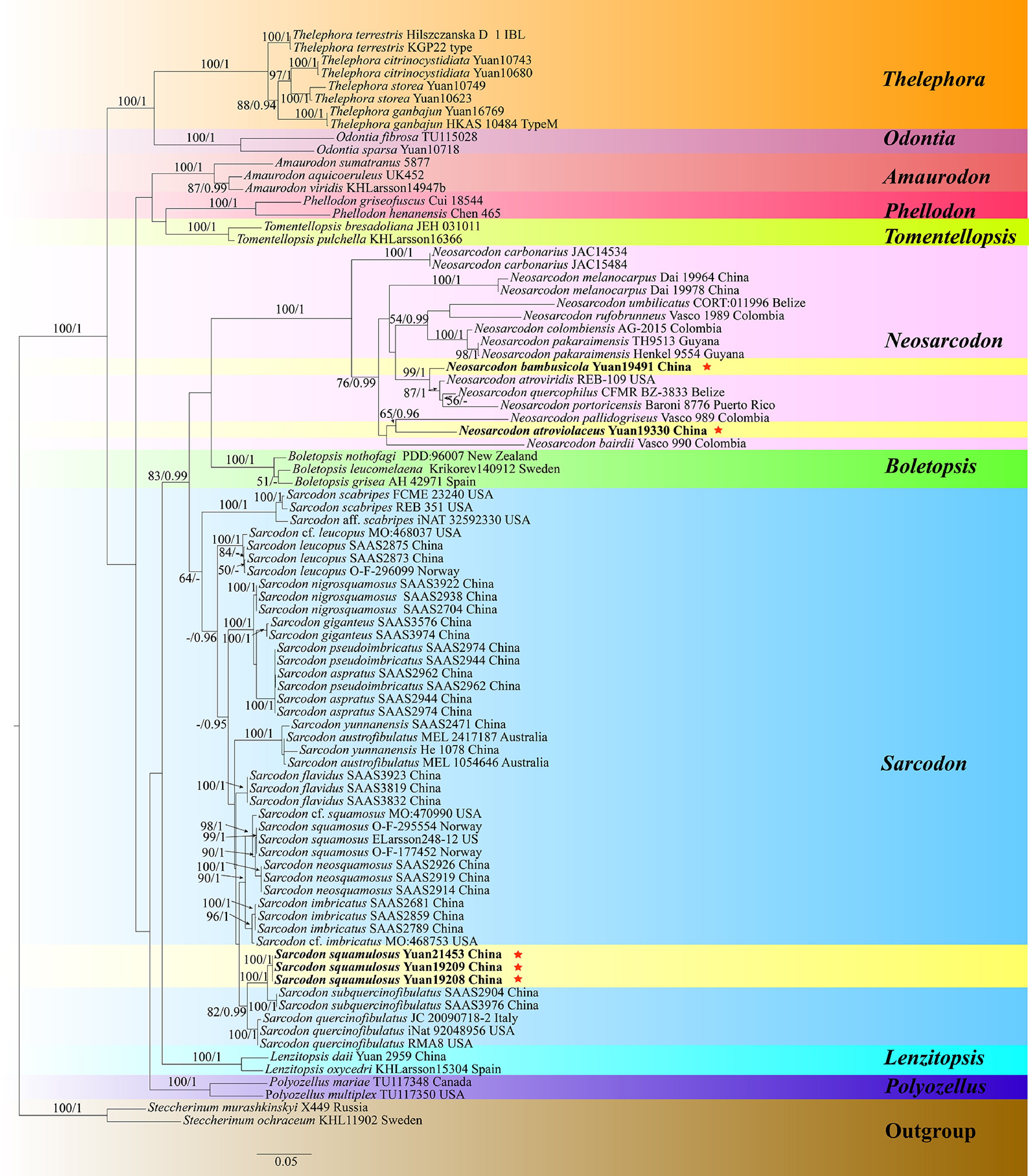
Figure 4.
Maximum likelihood tree illustrating the phylogeny of Neosarcodon, Sarcodon, and related genera in Thelephorales based on ITS + nLSU + nSSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 50%, and Bayesian posterior probabilities more than 0.95 respectively. Specimens examined are in bold, and new species are marked with red stars.
Thelephoraceae Chevall.
Odontia phylogeny based on combined ITS, nLSU, and nSSU sequence data (Fig. 5)
The aligned dataset comprised 38 specimens, representing 28 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for combined ITS + nLSU + nSSU (Fig. 5) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 3,306 characters, of which 2,515 characters are constant, 223 are variable and parsimony uninformative, and 568 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the three genes; the models for ITS, nLSU, and nSSU were GTR + I + G with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.003309. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.
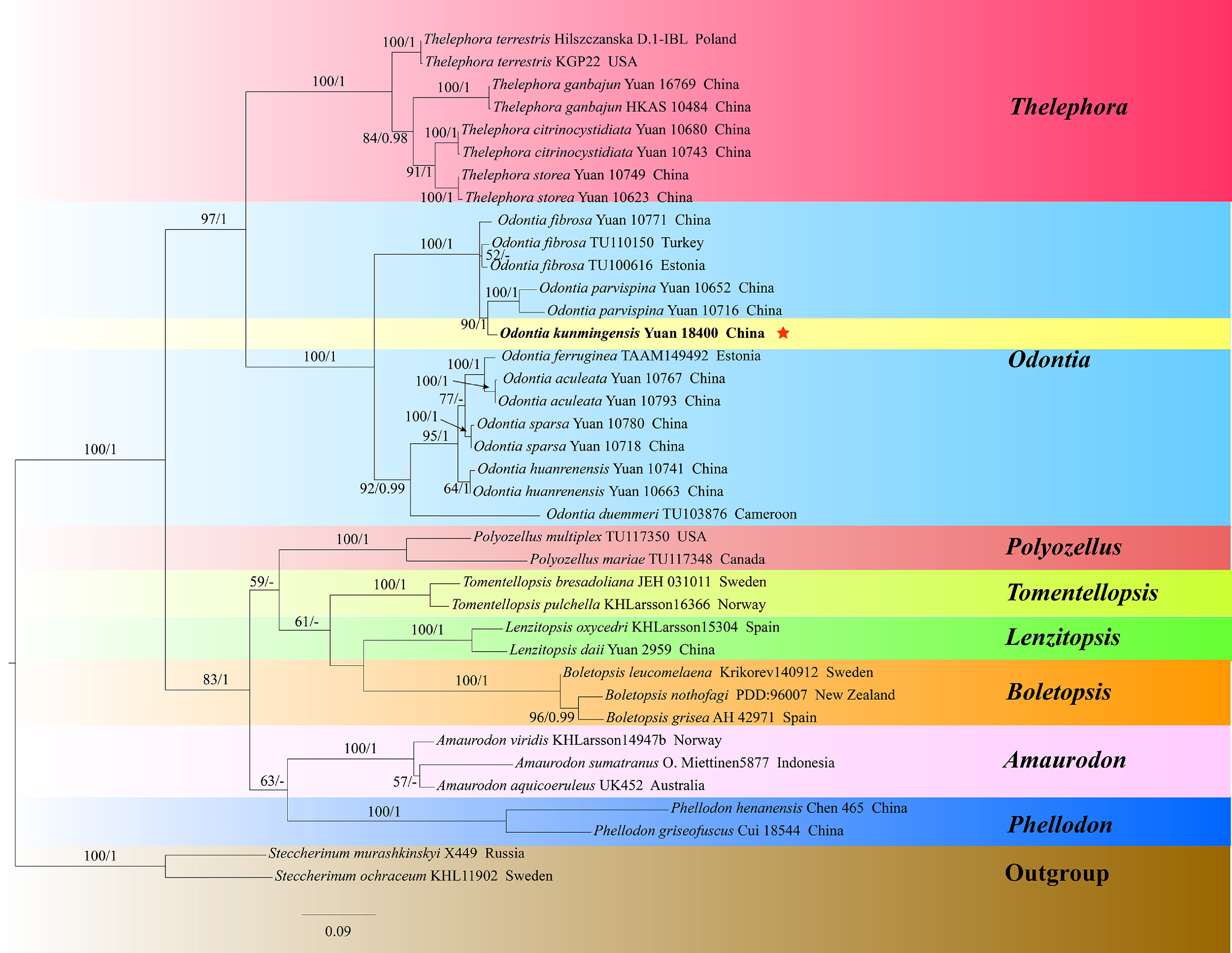
Figure 5.
Maximum likelihood tree illustrating the phylogeny of Odontia, and related genera in Thelephorales based on ITS + nLSU + nSSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 50%, and Bayesian posterior probabilities more than 0.95 respectively. Specimens examined are in bold, and new species are marked with red stars.
Thelephora phylogeny based on combined ITS, nLSU, nSSU, and mtSSU sequence data (Fig. 6)
The aligned dataset comprised 248 specimens, representing 193 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for combined ITS + nLSU + nSSU + mtSSU (Fig. 6) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 4,595 characters, of which 2,698 characters are constant, 659 are variable and parsimony uninformative, and 1,238 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the four genes; the models for ITS, nLSU, nSSU, and mtSSU were GTR + I + G, with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.031686. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.
Hymenochaetales Oberw.
Hymenochaetaceae Donk
Hymenochaete phylogeny based on combined ITS and nLSU sequence data (Fig. 7)
The aligned dataset comprised 123 specimens representing 105 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for combined ITS + nLSU (Fig. 7) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 2,713 characters, of which 1,602 characters are constant, 276 are variable and parsimony uninformative, and 835 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the four genes; the models for ITS and nLSU were GTR + I + G, with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.044018. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.

Figure 7.
Maximum likelihood tree illustrating the phylogeny of Hymenochaete, and related genera in Hymenochaetales based on ITS + nLSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 50%, and Bayesian posterior probabilities more than 0.95 respectively. Specimens examined are in bold, and new species are marked with red stars.
Peniophorellaceae L.W. Zhou, X.Wei Wang & S.L. Liu
Schizoporaceae Jülich
Peniophorella - Xylodon phylogeny based on combined ITS, nLSU, and nSSU sequence data (Fig. 8)
The aligned dataset comprised 150 specimens representing 150 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for combined ITS + nLSU + nSSU (Fig. 8) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 3,743 characters, of which 2,172 characters are constant, 436 are variable and parsimony uninformative, and 1,135 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the four genes; the models for ITS, nLSU, and nSSU were GTR + I + G, with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.023398. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.
Schizoporaceae Jülich
Lyomyces phylogeny based on combined ITS and nLSU sequence data (Fig. 9)
The aligned dataset comprised 70 specimens, representing 48 species. Four Markov chains were run for two runs from random starting trees, each for one million generations for combined ITS + nLSU (Fig. 9) data set with trees and parameters sampled every 1,000 generations. The dataset had an aligned length of 2,159 characters, of which 1,371 characters are constant, 220 are variable and parsimony uninformative, and 568 are parsimony informative. The best-fit evolutionary models applied in Bayesian analyses were selected by MrModeltest v. 2.3 for each region of the four genes; the models for ITS and nLSU were GTR + I + G, with equal frequency of nucleotides. Bayesian analysis resulted in a topology similar to that from ML analysis, with an average standard deviation of split frequencies = 0.007485. The ML topology was presented and annotated with ML bootstraps ≥ 50%, and BI bootstraps ≥ 0.95.
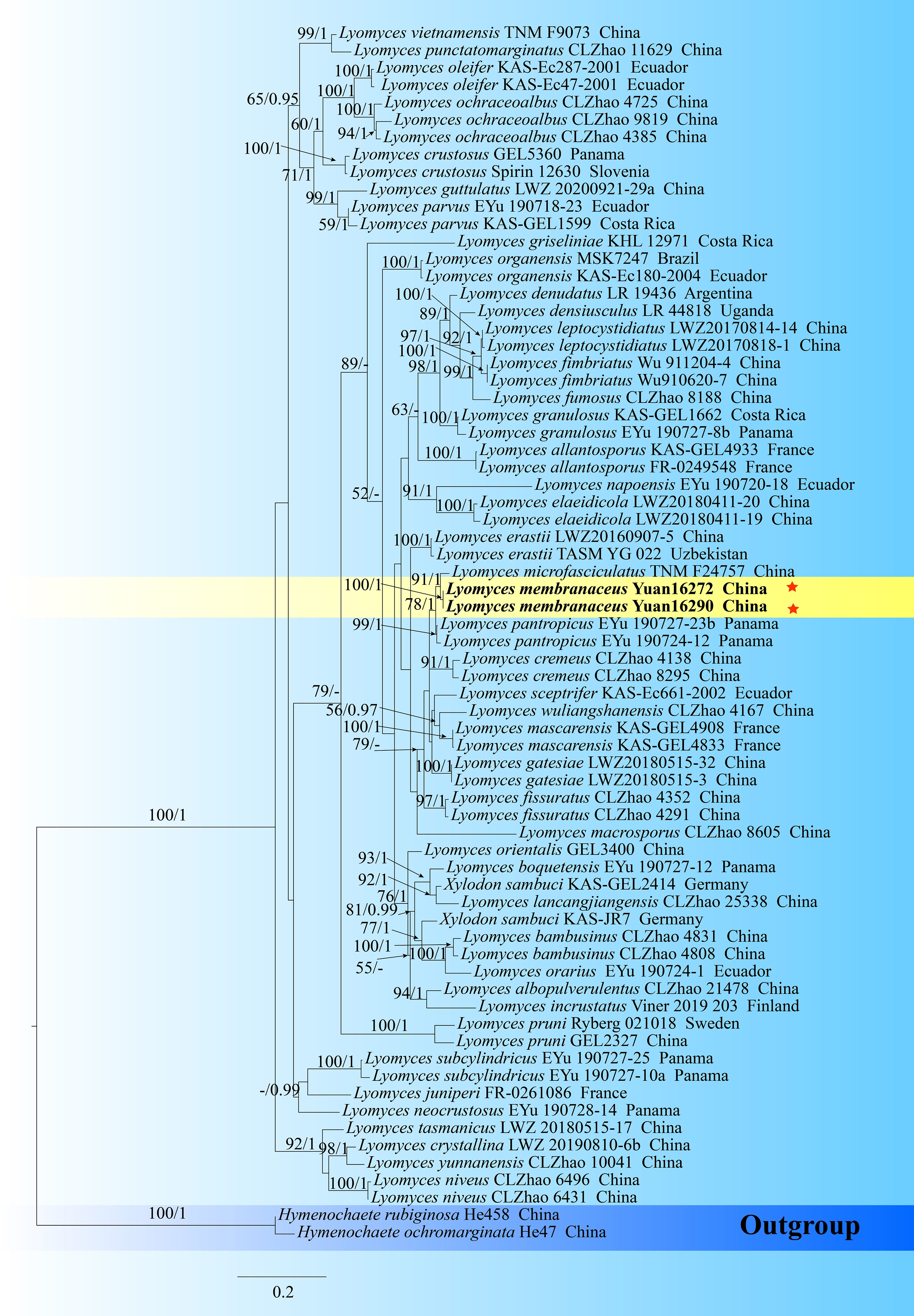
Figure 9.
Maximum likelihood tree illustrating the phylogeny of Lyomyces, and related genera in Hymenochaetales based on ITS + nLSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 50%, and Bayesian posterior probabilities more than 0.95 respectively. Specimens examined are in bold, and new species are marked with red stars.
Taxonomy
-
Thelephorales Corner ex Oberw.
Index Fungorum number: IF 90575
Bankeraceae Donk
Index Fungorum number: IF 80513
Type genus – Phellodon P. Karst.
Phellodon P. Karst.
Index Fungorum number: IF 18247
Type species – Phellodon niger (Fr.) P. Karst.
Notes – Phellodon was established by Petter Adolf Karsten in 1881 to distinguish it from other hydnoid genera with fleshy basidiomata, such as Hydnum and Sarcodon. The generic name is derived from the 'phellos' (cork), and 'odous' (tooth), referring to the cork-like texture of the pileus and hydnoid hymenophoral surface. The genus originally comprised three species, namely Ph. niger, Ph. cyathiformis and Ph. melaleucus. Phellodon niger, originally described by Fries as Hydnum nigrum[112], was later designated as the type species by Banker[113]. Subsequently, Maas Geesteranus[114] recognized only four Phellodon species in Europe, namely Ph. confluens, Ph. melaleucus, Ph. niger, and Ph. tomentosus. Later, Niemelä et al.[115] described a new species from pine forests in Finland. Recent molecular taxonomic studies on Phellodon have focused primarily on China, Europe, and the USA. In these investigations, 11 distinct taxa were identified in the southeastern USA[13,116], 13 in Europe[117], and 17 new species were documented from China[13,91−93,118]. In the present study, two new species of Phellodon from China are described, based on morphological characteristics and phylogenetic analyses inferred from ITS, nLSU, and nSSU sequences (Fig. 2).
Phellodon albospinus Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 10 and 11
Fungal Names number: FN 573093

Figure 10.
Basidiomata of Phellodon albospinus (holotype IFP 020036).
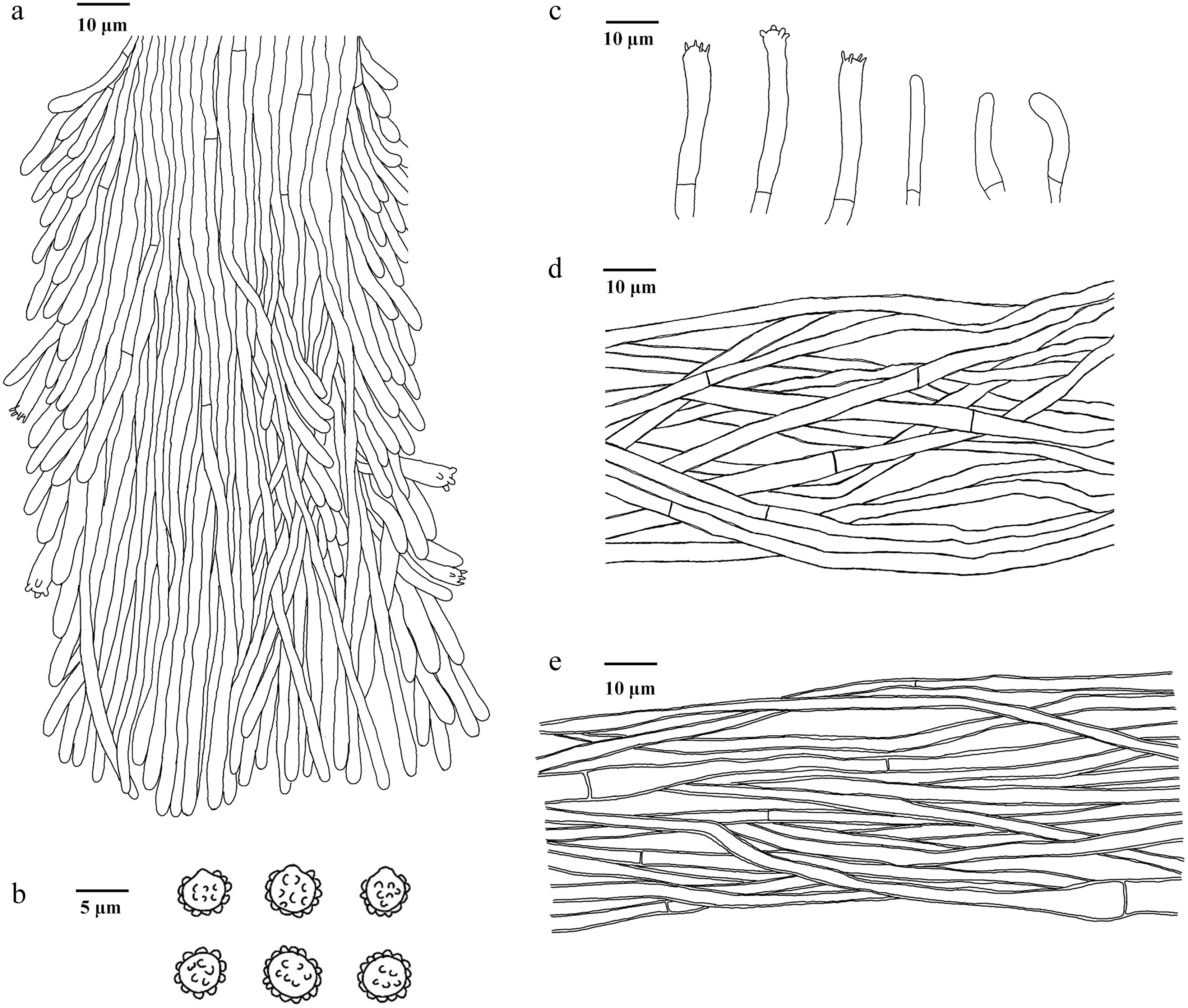
Figure 11.
Microscopic structures of Phellodon albospinus (drawn from the holotype IFP 020036). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Phellodon albospinus is characterized by the white spines when fresh, white to dark brown pileal surface, generative hyphae with long-cell and colorless to pale yellow basidiospores.
Etymology – Albospinus (Lat.): referring to the white spines.
Type – China, Yunnan Province, Lufeng City, Gaofeng Township, Beidala Village, GPS coordinates 25°20′4″ N, 101°52′57″ E, altitude 1,950 m, ground in mixed forest, 9 September, 2024, Yuan 21186 (IFP 020036, GenBank ITS: PQ803282; LSU: PV257916).
Description – Basidiomata terrestrial, stipitate, annual, solitary, soft and leathery when fresh, becoming woody and light in weight upon drying; taste mild, mild odor when dry. Pileus applanate to flabelliform, smooth, to 30 mm diam and 2–6 mm thick at center. Pileal surface white (–A1), light orange (5A4) to dark brown (8F8), zonate, glabrous when fresh, becoming glabrous, rugose when dry; margin white (–A1) when fresh, brownish orange (6C4) when dry, even to wavy, sometimes lobed. Spine surface white (–A1) when fresh, light brown (6D6) to black when dry; spines up to 2 mm long, base up to 0.2 mm diam, conical, 3–5 per mm, strongly decurrent on stipe, without spines at pileus margin, brittle when dry. Stipe lateral, 25–29 mm long, and 4–7 mm diam., leathery when fresh, woody upon drying, light brown (6D6) to dark brown (8F8), solid inner, cylindrical to flat, or attenuate downwards with a bulbous base.
Hyphal structure – Hyphal system monomitic, thin to thick-walled, CB+ in thick-walled hyphae, IKI–, tissues turned light yellow-green to olive green in KOH.
Pileus – Generative hyphae with simple-septa, slightly thick-walled, colorless, rarely branched, uninflated, long-cell, parallel, 3–4.5 μm diam.
Spines – Generative hyphae with simple-septa, thin-walled, colorless, unbranched, uninflated, interwoven in subsurface layer to parallel below, long-cell, straight, 2–3 μm diam.
Stipe – Generative hyphae with simple-septa, slightly thick to thick walled, colorless, unbranched, occasionally inflated, long-cell, straight, 2–5 μm diam.
Basidia – Clavate, with four sterigmata, and a basal simple-septa, 15‒43 × 3‒5 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subglobose or apple-like, colorless to pale yellow, thin-walled, tuberculate, tuberculi usually isolated, less than 1.0 μm long, (3.8‒)4‒5.1(–5.9) × (3.3–)3.5‒4.5(–4.9) μm, L = 4.36 μm, W = 4.08 μm, Q = 1–1.22 (n = 30/1), CB–, IKI–.
Notes – The new species Phellodon albospinus is classified within Phellodon (Fig. 2), and forms a clade with Ph. crassipileatus and Ph. zonatus. Ph. albospinus resembles Ph. crassipileatus in sharing the dark brown pileal surface and white spines when fresh. However, Ph. crassipileatus differs from Ph. albospinus due to its the special odors when dry and absent of clamp connections[92]. Ph. albospinus resembles Ph. subgriseofuscus in having white spines. However, Ph. subgriseofuscus differs from Ph. albospinus due to its fenugreek odor when dry, and wider basidia (5–7 µm)[93].
The phylogenetic analyses based on sequence data revealed a new distribution record for this species from Laos in the UNITE database, thereby expanding its known geographic range.
Phellodon zonatus L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 12,13
Fungal Names number: FN 572441

Figure 12.
Basidiomata of Phellodon zonatus (holotype IFP 020035).
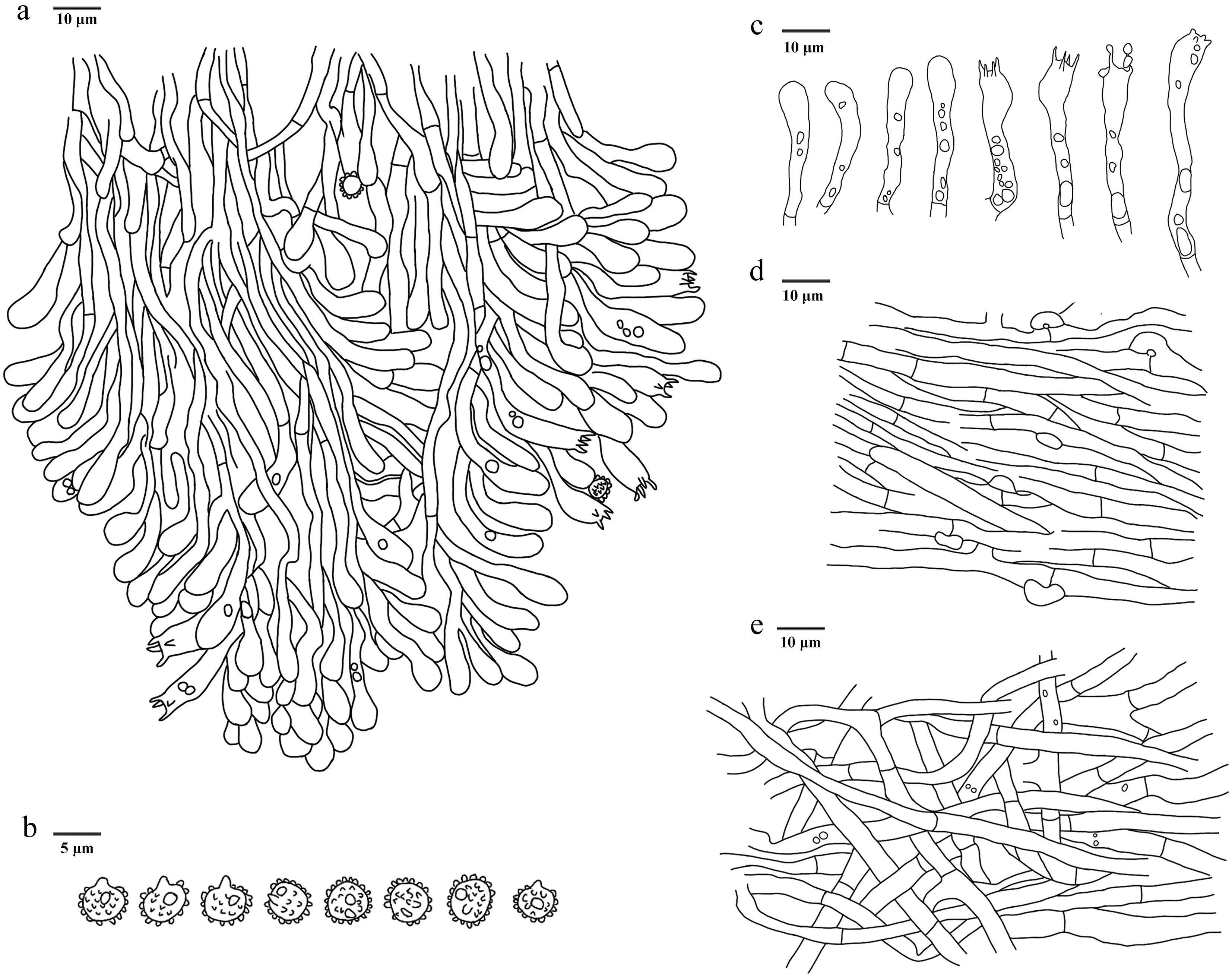
Figure 13.
Microscopic structures of Phellodon zonatus (drawn from the holotype IFP 020035). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Phellodon zonatus is characterized by the structural complexity basidiomata with a light fenugreek odor, flabellate to semicircle pileus, clavate or sinuous basidia and basidiospores measuring (3.7–)3.8‒4(–4.2) × (3‒)3.1‒3.8(–4) μm.
Etymology – Zonatus (Lat.): referring to the zonate pileus surface.
Type – China, Yunnan Province, Lincang City, Wulaoshan National Forest Park, GPS coordinates 23°54′46″ N, 100°10′51″ E, altitude 2,480 m, ground in mixed forest, 24 August, 2024, Yuan 20770 (holotype: IFP 020035, GenBank ITS: PQ803281; LSU: PV257915; SSU: PV257941).
Description – Basidiomata terrestrial, stipitate, annual, solitary, structural complexity may be increased further by developing secondary pilei from upper pilei surfaces, usually incorporates individual litter, releasing a light fenugreek odor when dry. Pileus flabellate to semicircle, smooth, to 80 mm broad, and 8 mm thick when dry, corky, and crisp with irregularly rounded, and undulating to distinctly lobate margins which, remain white when in active growth, around the center the texture is bumpy, moving away from the center there may be concentric corrugations and color zonation may be weak to strong, usually some shade of dark yellow (4C8) or tan (6D5–6E6) when fresh, dark gray (–F1) to dark brown (6F4–9F8) in the center, and pale brown (5D4–7D8) at the edge when dry. Spines less than 2 mm long, white (–A1) when fresh, later to yellowish white (1A2–4A2) to orange white (5A2–6A2), dark color at the basal. Stipe cylindrical or basally tapered with a smooth texture, 26–30 mm long, and 5–10 mm across, lacking a distinct woolly tomentum but can be grooved, flattened or slightly velvety in places, with decurrent rudimentary or entire spines towards the apex, reddish brown (8D4–9F8) when fresh, later to pale brown (5D4–7D8).
Hyphal structure – Hyphal system monomitic, thin-walled, CB–, IKI–, tissues having a weakly greenish-grey color in KOH.
Pileus – Generative hyphae mostly with simple-septa, rarely with clamp connections, thin-walled, colorless, unbranched, uninflated, parallel, 2–6 μm diam.
Spines – Generative hyphae with simple-septa, thin-walled, colorless, unbranched, uninflated, interwoven in subsurface layer to parallel below, long-cell, straight, 2–4 μm diam.
Stipe – Generative hyphae with simple-septa, thin-walled, colorless, rarely branched, uninflated, long-cell, straight, 2–5 μm diam.
Basidia – Clavate or sinuous, with 4 sterigmata and a basal simple-septa, the apex is swollen and constricted in the middle with oil droplets, 30‒47 × 3‒6.5 μm, CB–, IKI–.
Cystidia – Absent.
Spores – Basidiospores subglobose or apple-like, colorless, thin-walled, with oil-droplet inside, tuberculate, tuberculi usually isolated, less than 1.0 μm long, (3.7–)3.8‒4(–4.2) × (3‒)3.1‒3.8(–4) μm, L = 3.91 μm, W = 3.42 μm, Q = 1–1.15 (n = 30/1), CB–, IKI–.
Notes – The new species Phellodon zonatus is classified within Phellodon (Fig. 2), and forms a clade with Ph. crassipileatus. Ph. zonatus resembles Ph. crassipileatus in sharing the special odors when dry, the color of spines. However, Ph. crassipileatus differs from Ph. zonatus due to its shorter stipe (up to 1.5 cm), the apex shape of basidia and larger basidiospores ([3.5–]4–5 × 4–5 µm)[92]. Ph. zonatus resembles Ph. perchocolatus and Ph. subgriseofuscus in having white spines. However, Ph. perchocolatus differs from Ph. zonatus due to its larger basidiospores (4–5[–5.5] × [3.5–]4–4.5[–5] μm), and its longer size of spines (up to 3 mm)[92]. Ph. subgriseofuscus distinguish from Ph. zonatus in its longer basidiospores (4–5 × [3–]3.2–4.8 μm), and black pileal surface[93].
Two closely related sequences from Laos were retrieved from the UNITE database. Phylogenetic analyses demonstrated that these sequences cluster within the same clade as the specimens described in this study, thereby extending the distribution range of Phellodon zonatus to Laos.
Boletopsidaceae Bondartsev & Singer ex Jülich
Index Fungorum number: IF 81724
Type genus – Boletopsis Fayod
Hydnellum P. Karst.
Index Fungorum number: IF17781
Type species – Hydnellum suaveolens (Scop.) P. Karst.
Notes – The early stipitate hydnoid fungi were initially placed in the genus Hydnum by Fries[119]. Karsten[120] established the genus Hydnellum, typified by Hydnellum suaveolens, and distinguished it from the Hydnum based on the texture of the basidiomata, the morphological characteristics of stipes and spines. Subsequently, the stipitate hydnoid species have continued to attract considerable systematic research[114,121−124]. Donk[10] proposed the family Bankeraceae, including Bankera and Phellodon, while placing Boletopsis, Hydnellum and Sarcodon in the Thelephoraceae. Jülich[12] broadened the concept of Bankeraceae to encompass stipitate and hydnoid basidiomata, and redefined the family based on spore color and morphology, placing Hydnellum and Sarcodon together in the Bankeraceae. With the advent of molecular systematics, studies have shown that Hydnellum and Sarcodon are closely related phylogenetically, and neither genus forms a strictly monophyletic group[25,125,126]. Larsson et al.[127] redefined the generic boundaries between Hydnellum and Sarcodon based on molecular evidence. Subsequently, numerous multi-gene phylogenetic studies and new species descriptions have been published, further enhancing the species diversity of Hydnellum[36,64,94]. Currently, approximately 80 species have been described and assigned to the genus Hydnellum according to records in Index Fungorum. In the present study, seven new species—Hyd. carnosum, Hyd. hydrangeoides, Hyd. infundibuliforme, Hyd. liantaishanense, Hyd. porphyreum, Hyd. testaceum and Hyd. tomentosum—are described based on the ITS, nLSU, and nSSU data (Fig. 3).
Hydnellum carnosum L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 14,15
Fungal Names number: FN 572433

Figure 14.
Basidiomata of Hydnellum carnosum (holotype IFP 020026). Photo by Xue-Lian Gao.
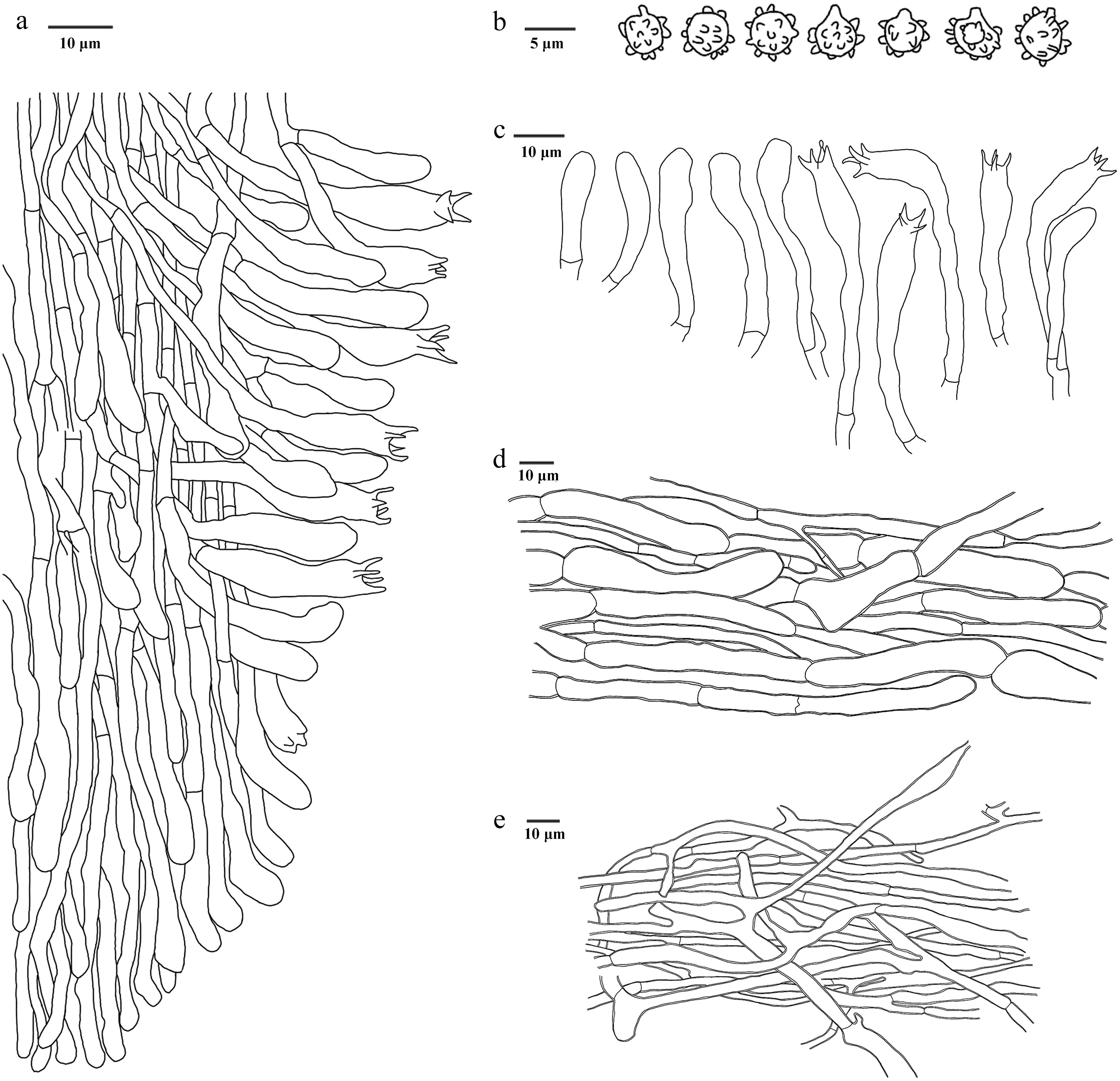
Figure 15.
Microscopic structures of Hydnellum carnosum (drawn from the holotype IFP 020026). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Hydnellum carnosum is characterized by the deep orange to brown basidiomata, basidiomata with a bitter taste, stipe completely covered with spines and ellipsoid to subglobose basidiospores.
Etymology – Carnosum (Lat.): referring to the carnose basidiomata.
Type – China, Yunnan Province, Chuxiong Yi Autonomous Prefecture, Lufeng City, Guangtong Town, Shanjianshan, GPS coordinates 25°14′24″ N, 101°45′52″ E, altitude 2,300 m, ground in mixed forest, 31 August, 2023, GXL 0391 (holotype: IFP 020026, GenBank ITS: PQ349955; LSU: PV257895; SSU: PV257935).
Description – Basidiomata terrestrial, stipitate, annual, gregarious, soft and fleshy when fresh, becoming firm, brittle, and light in weight upon dry, taste bitter, odour farinaceous when dry. Pileus subcircular, bumpy and uneven, pubescent, upwarp scales, incurved margins, moving away from the center radiating projecting fibrils, involuted, and rarely lobed, deep orange (5A8–6A8) to brown (6D4–7F8) when fresh, light-brown (5D4–7D8) to brown (6D4–7F8) when dry. Spines conical, tenuous, less than 2 mm long, decurrent on stipe, and nearly to the ground, spines at pileus margin, white (–A1) when fresh, beige (4C3) at the apex and brown (6D4–7F8) at the basal when dry, brittle. Stipe central, clavate to cylindrical, completely covered with spines, reddish brown (16A4–19A5) when fresh, dark yellow (4C8) to light-brown (5D4–7D8) when dry, sunken, rugous.
Hyphal structure – Hyphal system monomitic, generative hyphae with simple-septa, colorless, thin- to slightly thick-walled, CB+, IKI–, tissues olivaceous in KOH.
Pileus – Generative hyphae slightly thick-walled, sparsely branched, uninflated, partly shrinking at the septate, unequal septate, interwoven, 3–15 μm diam.
Spines – Generative hyphae thin-walled, infrequent branched, more or less parallel along spines, long-cell, straight, 2–3 μm diam.
Stipe – Generative hyphae slightly thick-walled, frequent branched, inflated, long-cell, more or less flexural, 2–5 μm diam.
Basidia – Clavate, thin-walled, smooth, colorless, 4 sterigmata and up to 5 μm, with simple-septa or branch at base, 27–58 × 5–7 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores ellipsoid to subglobose, colorless, thin-walled, tuberculate, tuberculi usually isolated, up to 1.0 μm long, (3.5–)3.7–4 × 3–3(–3.5) μm, L = 3.97 μm, W = 3.81 μm, Q = 1–1.14 (n = 30/1), CB–, IKI–.
Notes – The new species Hydnellum carnosum is classified within Hydnellum (Fig. 3), and forms a clade within subg. Scabrosum[36]. This subgenus is characterized planar to depressed and brown pileus, azonate pileal surface with scabrosity, variously brown spines, not duplex and yellow to orange context, inflated and unclamped generative hyphae, and irregularly ellipsoid to globose basidiospores[36]. Similarly, Hyd. carnosum has bumpy and uneven pileus with tomentum, unclamped generative hyphae, and ellipsoid basidiospres. However, Hyd. carnosum has deeper color of context (brown). Hyd. carnosum resembles Hyd. fagiscabrosum in having fleshy basidiomata, stipe covered with spines, and pileus with scales. However, Hyd. fagiscabrosum differs from Hyd. carnosum due to its brown basidiospores, white pileus margins, and longer stipe[128].
Hydnellum hydrangeoides L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 16,17
Fungal Names number: FN 572435

Figure 16.
Basidiomata of Hydnellum hydrangeoides (holotype IFP 020029).
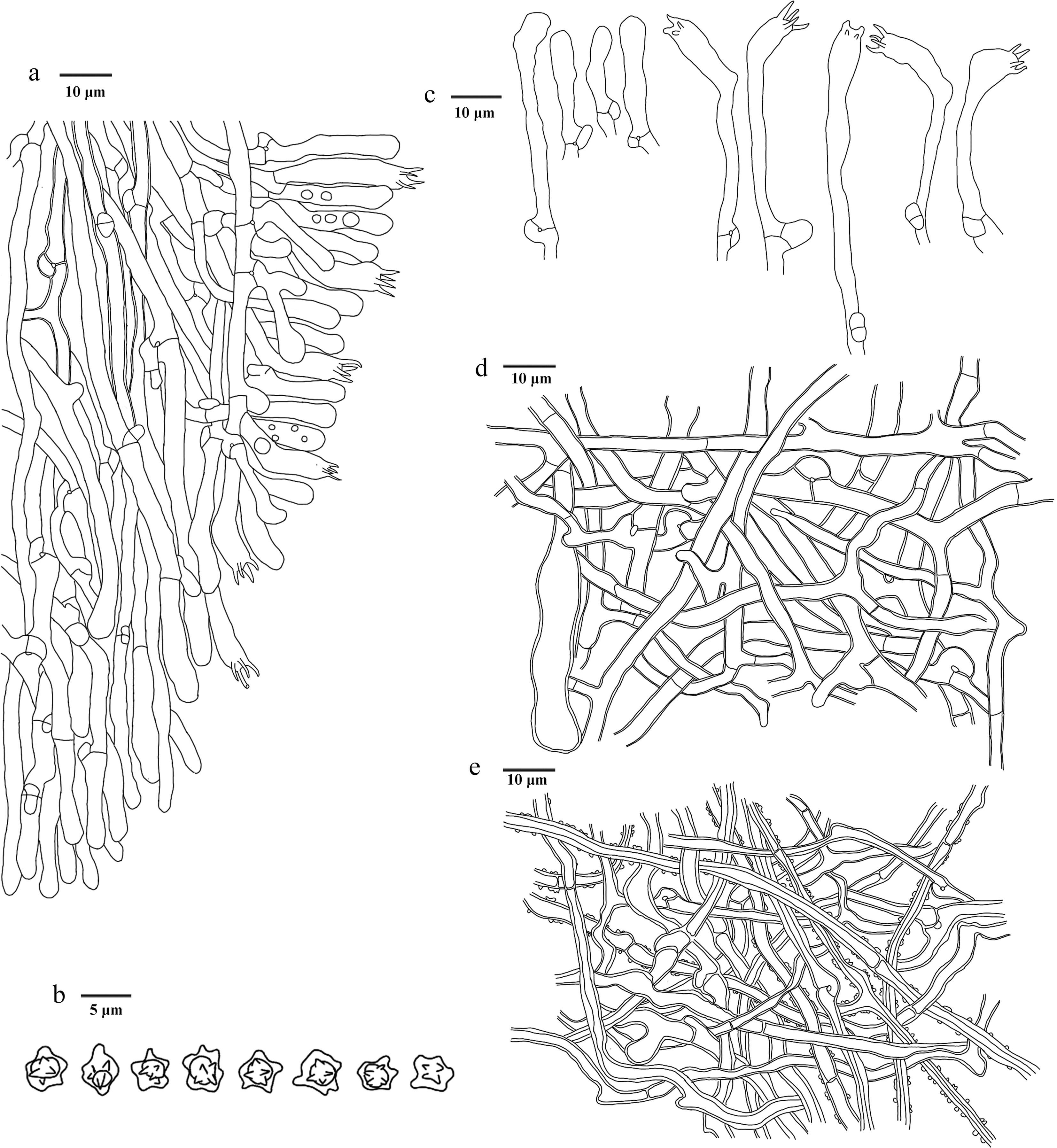
Figure 17.
Microscopic structures of Hydnellum hydrangeoides (drawn from the holotype IFP 020029). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Hydnellum hydrangeoides is characterized by the broccoli-like basidiomata with a typical smell of dry mushrooms, tomentose pileal surfaces, isolated or multiplex spines, and basidiospores with oily-like contents.
Etymology – Hydrangeoides (Lat.): referring to basidiomata similar to the hydrangea.
Type – China, Yunnan Province, Kunming City, Luquan Yi, and Miao Autonomous County, Tuanjie Town, GPS coordinates 25°45′43″ N, 102°31′54″ E, altitude 1,800 m, ground in mixed forest, 11 October, 2023, Yuan 19215 (holotype: IFP 020029, GenBank ITS: PQ805351; LSU: PV257890; SSU: PV257929).
Description – Basidiomata terrestrial, stipitate, annual, solitary, fleshy, broccoli-like, becoming hard and brittle upon dry, taste mild, releasing a typical smell of dry mushrooms (odor fragrant). Pileus hemispherical, from many repeated small pilei, up to 6 cm tall, individuals more than 5 mm thick, often incurved margins, irregularly flabelliform, hard, brittle, and tenacious, usually uneven and bumpy, surface with tomentose, greyish orange (5B3–6B6) to brownish orange (6C3–7C8) when fresh, olive brown (4D3–4F8) to purplish grey (13B2–14F2) when dry. Spines conical, more than 2 mm long, isolated or multiplex, usually differentiated into 2–3, without decurrent on stipe, violet grey (15B2–18F2) when fresh, surface light violet (16A4–19A5) and inner deep violet (15D8–18E8) when dry, brittle. Stipe clavate, 70 × 30 mm, middle, smooth, uneven and bumpy, unincorporates litter, white (–A1) when fresh, pale violet (15A3–19A3) when dry, hard.
Hyphal structure – Hyphal system monomitic, colorless, thin- to slightly thick-walled, CB+, IKI–, tissues olivaceous in KOH.
Pileus – Generative hyphae mostly with simple-septa, rarely with clamp connections, slightly thick-walled, smooth, frequent branched, irregularly arranged, flexuous, rarely inflated, and up to 9 μm wide, 2–6 μm diam.
Spines – Generative hyphae with clamp connections, thin- to slightly thick-walled, smooth, frequent branched, parallel along spines, uninflated, long cell, unequal septate, 2–5 μm diam.
Stipe – Generative hyphae mostly with simple-septa, rarely with clamp connections, thick-walled, smooth to adhesion crystals, frequent branched, flexuous, irregularly arranged, inflated, and up to 10 μm wide, long-cell, unequal septate, flexuous, 2–6 μm diam.
Basidia – Clavate, thin-walled, smooth, colorless, with 4 sterigmata and a basal clamp connection, 26‒59 × 3‒5 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subglobose to globose, with oily-like contents, colorless, thin-walled, tuberculate, tuberculi usually isolated, less than 1.0 μm long, (2.8–)2.9–4.0 × 2.5‒3.0 μm, L = 3.1 μm, W = 3.0 μm, Q = 1–1.33 (n = 30/1), CB–, IKI–.
Notes – In the phylogenetic tree (Fig. 3), the new species Hydnellum hydrangeoides is revealed as a sister to Hyd. scleropodium and Hyd. cyanopodium. They exhibit some similar characteristics: spines length (2–2.5 mm long), and tuberculate basidiospores[122]. However, Hyd. hydrangeoides can be distinguished from Hyd. scleropodium by its narrower basidia and shorter spines[122]. Hyd. hydrangeoides differs from Hyd. cyanopodium due to its tomentose pileus surface, and smaller stipe[122]. Hyd. hydrangeoides resembles Hyd. bomiense and Hyd. yunnanense in having a tomentose pileal surface. However, Hyd. bomiense differs from Hyd. hydrangeoides due to its grayish yellow to dark brown pileus, shorter spines (< 1.1 mm), and basidia (15–42 × 4–7 µm)[36]. Hyd. yunnanense distinguish from Hyd. hydrangeoides by its shorter stipe (< 4 cm), and basidia (13–28 × 4–7 µm)[36].
Hydnellum infundibuliforme L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 18,19
Fungal Names number: FN 572431

Figure 18.
Basidiomata of Hydnellum infundibuliforme (holotype IFP 020021).
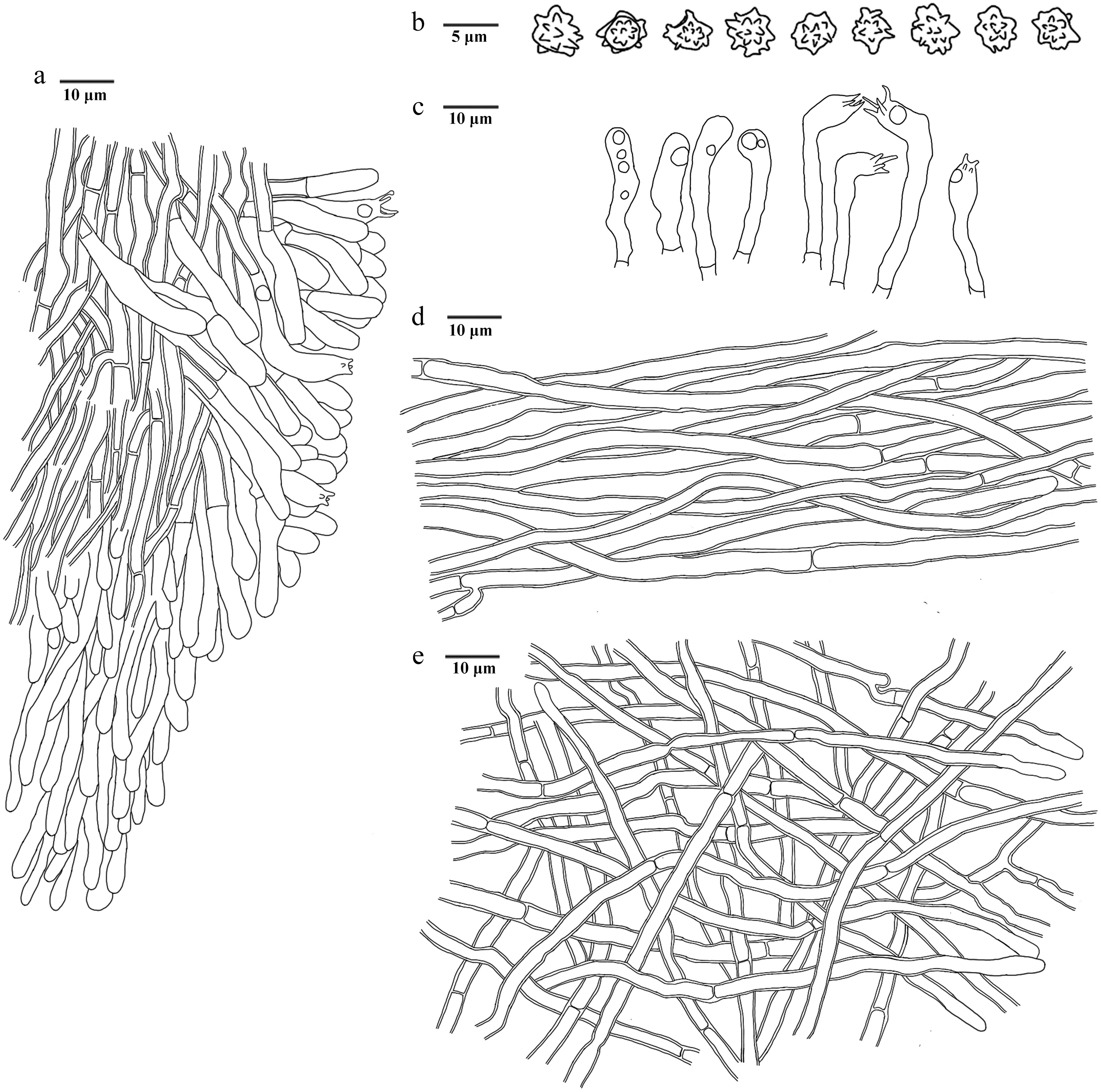
Figure 19.
Microscopic structures of Hydnellum infundibuliforme (drawn from the holotype IFP 020021). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Hydnellum infundibuliforme is characterized by the odorless basidiomata with a woody taste, uneven and bumpy pileal surfaces, tomentose stipe surfaces and ellipsoid to subglobose basidiospores.
Etymology – Infundibuliforme (Lat.): referring to the infundibuliform pileus.
Type – China, Yunnan Province, Lufeng City, Gaofeng Township, Beidala Village, GPS coordinates 25°20′4″ N, 101°52′57″ E, altitude 1,950 m, ground in mixed forest, 9 September, 2024, Yuan 21180 (holotype: IFP 020021, GenBank ITS: PQ805356).
Description – Basidiomata terrestrial, stipitate, annual, gregarious, fleshy when fresh, becoming hard and brittle upon drying, taste mild like woody, not releasing a smell. Pileus irregularly flabelliform to subcircular or splintered, 20–35 mm long, and 15–30 mm across, occasionally adheres to forming complexes, often incurved margins, corky, stiff, and brittle, usually uneven and bumpy, concentrically zonate, invagination at the center, moving away from the center radiating tufts and projecting fibrils and the color becoming lighter, beige at the growth active areas, reddish brown (8D4–9F8) to dark brown (6F4–9F8) when fresh, light brown (5D4–7D8) when dry, and light yellow (1A4–4A5) crystals precipitated. Spines conical, tenuous, solitary, up to 3 mm long, decurrent on stipe, light brown (5D4–7D8) when fresh, brown (6D4–7F8) when dry, brittle. Stipe clavate, 22–57 mm long, and 3–7 mm across, middle, surface with tomentose, rarely producing branched, occasionally incorporates litter, inner corky, reddish brown (8D4–9F8) to dark brown (6F4–9F8) when fresh, resilient, reddish brown (8D4–9F8) when dry, brittle.
Hyphal structure – Hyphal system monomitic, generative hyphae with simple-septa, smooth, colorless, thin- to slightly thick-walled, CB+, IKI–, tissues indigoticus in KOH.
Pileus – Generative hyphae slightly thick-walled, sparsely branched, parallel, regularly arranged, uninflated, occasionally flexuous, 2–5 μm diam.
Spines – Generative hyphae thin- to slightly thick-walled, infrequent branched, parallel along spines, uninflated, long-cell, straight, 1.5–4 μm diam.
Stipe – Generative hyphae slightly thick-walled, sparsely branched, uninflated, long-cell, straight, unequal septate, occasionally flexuous,1.5–3.5 μm diam.
Basidia – Clavate, thin-walled, smooth, four sterigmata, constricted at the base, with oily-like contents, occasionally, 23‒41 × 3‒5 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores ellipsoid to subglobose, colorless, thin-walled, tuberculate, tuberculi usually isolated, less than 1.0 μm long, 3.0–4.0(‒4.2) × (2.5‒)2.9‒3.3(‒3.5) μm, L = 3.4 μm, W = 3.0 μm, Q = 1–1.33 (n = 60/2), CB–, IKI–.
Material examined (paratypes) – China, Yunnan Province, Lufeng City, Gaofeng Township, Beidala Village, GPS coordinates 25°20′4″ N, 101°52′57″ E, altitude 1,950 m, ground in mixed forest, 9 September, 2024, Yuan 21185 (IFP 020022, GenBank ITS: PQ805357; SSU: PV257933); ground in mixed forest, 9 September, 2024, Yuan 21220 (IFP 020023, GenBank ITS: PQ805358; SSU: PV257934).
Notes – The new species Hydnellum infundibuliforme is grouped within Hydnellum, forming a monophyletic lineage, and is closely associated with Hyd. cumulatum (Fig. 3). Hyd. infundibuliforme resembles Hyd. cumulatum in sharing the similar size of spine (3 mm). However, Hyd. cumulatum differs from Hyd. infundibuliforme due to larger pileus (7 cm broad), wider basidia (24.0–32.5 × 5.0–5.8[–6.4] µm), and larger basidiospores ([3.7–]4.0–5.0× 3.5–4.5 µm)[122]. Hyd. infundibuliforme resembles Hyd. auratile, Hyd. brunneorubrum and Hyd. rubidofuscum in having reddish brown pileus. However, Hyd. auratile differs from Hyd. infundibuliforme due to deeply split margin, and tawny to purplish brown spines[124]. Hyd. brunneorubrum differs from Hyd. infundibuliforme by its longer spines (up to 4 mm), wider stipes (up to 10 mm), and larger basidiospores ([4–]4.1–5.1[–5.2] × [3.1–]3.2–4.6[–4.8] µm)[36]. Hyd. rubidofuscum differs from Hyd. infundibuliforme by its larger pileus (up to 70 mm), scrupose, fibrillose to virgate pileal surface. and grayish brown spines[36].
Hydnellum liantaishanense L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 20,21
Fungal Names number: FN 572432

Figure 20.
Basidiomata of Hydnellum liantaishanense (holotype IFP 020024).
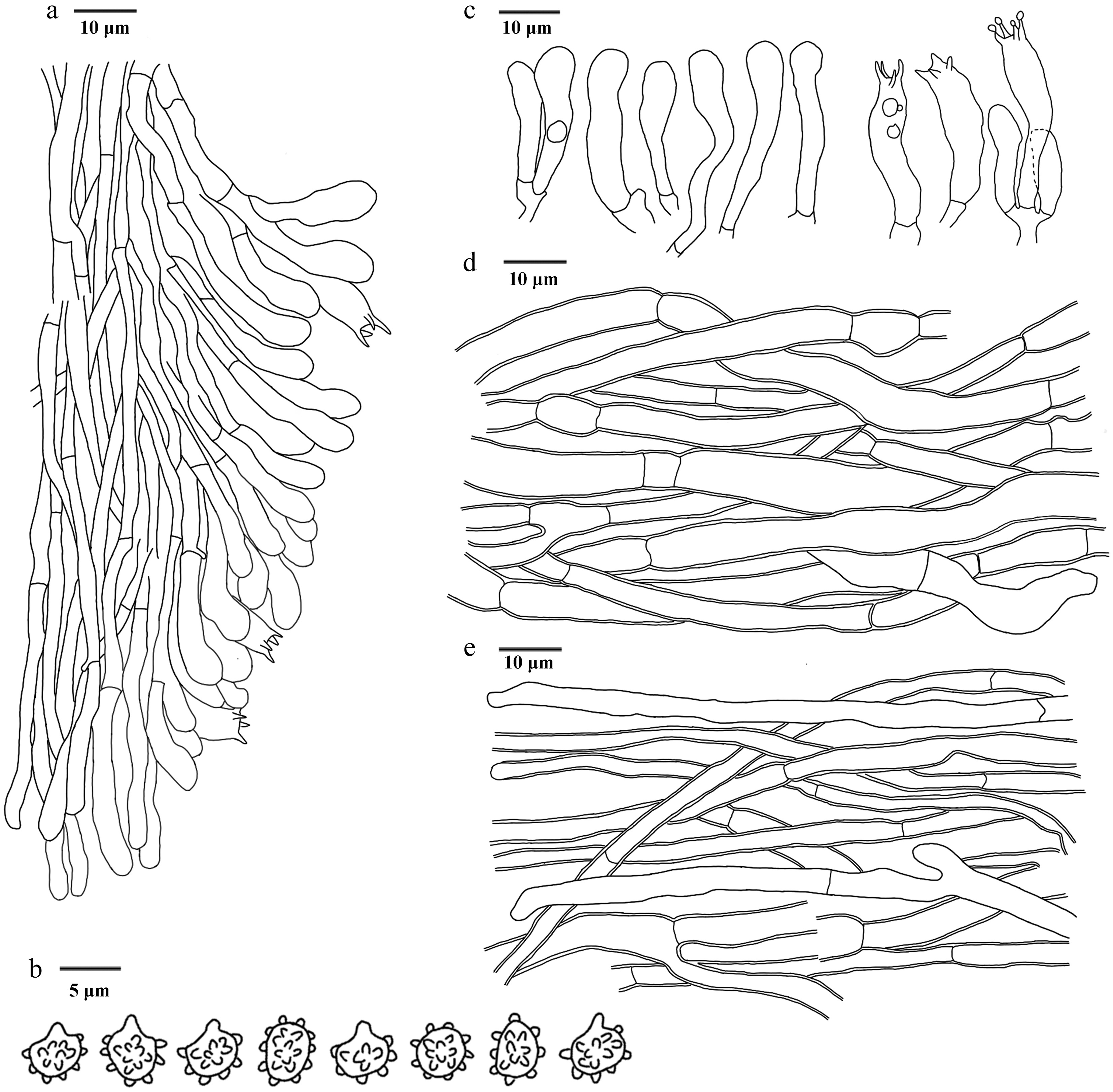
Figure 21.
Microscopic structures of Hydnellum liantaishanense (drawn from the holotype IFP 020024). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Hydnellum liantaishanense is characterized by the unclosed round to circular pileus with upwarp scales, uneven and bumpy pileal surfaces, spines growing near the soil, and clavate or sinuous basidia.
Etymology – Liantaishanense (Lat.): referring to the type from Liantaishan Mountain.
Type – China, Yunnan Province, Dali Bai Autonomous Prefecture, Dali City, Shuanglang Town, Liantai Mountain, GPS coordinates 25°56′50″ N, 100°17′38″ E, altitude 2,988 m, ground in mixed forest, 7 September, 2024, Yuan 21089 (holotype: IFP 020024, GenBank ITS: PQ805355; LSU: PV257894).
Description – Basidiomata terrestrial, stipitate, annual, solitary, fleshy when fresh, becoming hard and brittle upon drying, taste mild, not releasing a smell. Pileus unclosed round to circular, irregularly, up to 60 mm, often incurved margins, moving away from the center there are radiating tufts and projecting fibrils, hard and brittle, usually uneven and bumpy, concentrically zonate, reddish brown (8D4–9F8) to dark brown (6F4–9F8) when fresh, light brown (5D4–7D8) when dry, rugose, with upwarp scales, and not easily dislodged. Spines conical, solitary, less than 3 mm long, decurrent on stipe, surface dark violet (14F3–14F8) when fresh, brown (6D4–7F8) when dry, brittle. Stipe cylindrical to clavate, 25–70 mm long, and 6–10 mm across, middle, smooth, fleshy when fresh, brittle when dry, some of the spines grow near the soil, reddish white (7A2–13A2) to orange red (8A6–8B8).
Hyphal structure – Hyphal system monomitic, generative hyphae with simple-septa, thin- to slightly thick-walled, CB+, IKI–, tissues olivaceous in KOH.
Pileus – Generative hyphae colorless, sparsely branched, regularly arranged, inflated, sometimes flexuous, 3–10 μm diam.
Spines – Generative hyphae thin-walled, colorless, sparsely branched, parallel along spines, uninflated, straight, 2–4 μm diam.
Stipe – Generative hyphae colorless, sparsely branched, uninflated, long-cell, straight, 2–8.5 μm diam.
Basidia – Clavate or sinuous, thin-walled, smooth, 4 sterigmata, constricted at the base, with oily-like contents, occasionally, 22‒35 × 3‒5 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subglobose to globose, apple-like, colorless, thin-walled, tuberculate, tuberculi usually isolated, less than 1.0 μm long, (3.7‒)3.8–4.1(‒4.5) × (2.9‒)3‒3.4(‒3.7) μm, L = 4.0 μm, W = 3.0 μm, Q = 1.21–1.33 (n = 60/2), CB–, IKI–.
Material examined (paratype) – China, Yunnan Province, Dali Bai Autonomous Prefecture, Dali City, Shuanglang Town, Liantai Mountain, GPS coordinates 25°56′50″ N, 100°17′38″ E, altitude 2,988 m, ground in mixed forest, 7 September, 2024, Yuan 21027 (IFP 020025, GenBank ITS: PQ805354; LSU: PV257893; SSU: PV257932).
Notes – In this study, the new species Hydnellum liantaishanense is placed within Hydnellum (Fig. 3), and it forms a clade with Hyd. fuscoindicum and Hyd. porphyreum. Hyd. liantaishanense resembles Hyd. porphyreum in having clavate stipe and simple-septate hyphal system. However, Hyd. liantaishanense can be delimited from Hyd. porphyreum by its narrower basidia and shorter basidiospores. Hyd. liantaishanense resembles Hyd. atrospinosum and Hyd. squamulosum in having circular pileus. Hyd. atrospinosum differs from Hyd. liantaishanense in its larger pileus (up to 75 mm), shorter spines (< 2.5 mm), and basidia (17–45 µm)[36]. Hyd. squamulosum distinguish from Hyd. liantaishanense by its smaller pileus (< 35 mm), shorter spines (< 2 mm), and stipes (< 4 cm)[36].
Hydnellum porphyreum L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 22,23
Fungal Names number: FN 572434

Figure 22.
Basidiomata of Hydnellum porphyreum (holotype IFP 020027). Photo by Yan-Yan He.
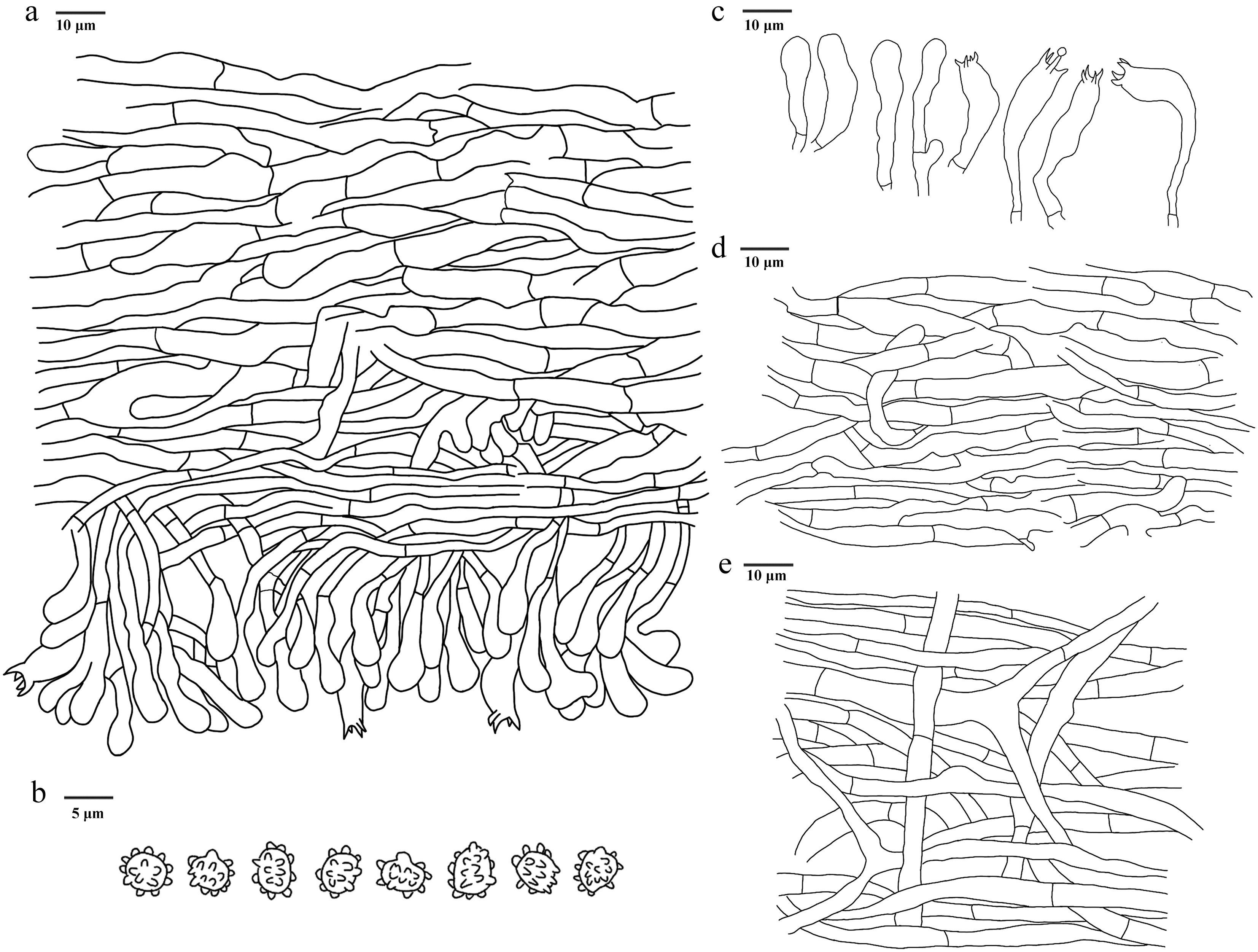
Figure 23.
Microscopic structures of Hydnellum porphyreum (drawn from the holotype IFP 020027). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Hydnellum porphyreum is characterized by the purplish-pink to light-violet pileus, stipe with decurrent spines, curved and fleshy stipes and ellipsoid to ovoid basidiospores.
Etymology – Porphyreum (Lat.): referring to the purple basidiomata.
Type – China, Guangxi Zhuang Autonomous Region, Guilin City, Xing'an County, Gaoshang Town, GPS coordinates 25°24′58″ N, 110°35′30″ E, altitude 1,100 m, ground in mixed forest, 3 April, 2024, Yuan 19292 (holotype: IFP 020027, GenBank ITS: PQ805352; LSU: PV257891; SSU: PV257930).
Description – Basidiomata terrestrial, stipitate, annual, solitary to gregarious, soft and fleshy when fresh, becoming firm and light in weight upon dry, taste none, odour farinaceous when dry. Pileus planar, round, up to 21 mm across and 4–7 mm thick at the centre, incurved, rarely lobed, surface azonate, pubescent, floccose to felted, purplish-pink (14A5) to light-violet (15A5–19A5) when fresh, later to light-brown (5D4–7D8) when dry, margin light-yellow (1A4–4A5) to greyish violet (17D5) when fresh, dark violet (15F3–18F8) with age. Spines conical, up to 2.6 mm long, base up to 0.4 mm diam., 3–4 per mm, decurrent on stipe, without spines at pileus margin, brittle when dry, surface white (–A1) to pale violet (15A3–19A3) when fresh, brown (6E5–7F8) when dry. Stipe central, clavate, curved, fleshy, 1.5 cm long, and 0.8 cm diam., light violet (16A4–19A5) to violet brown (10F7) when fresh, light brown (6D8) to dark brown (7F7) when dry, sunken, rugous.
Hyphal structure – Hyphal system monomitic, generative hyphae with simple-septa, CB–, IKI–, tissues having a dark greenish-blue color in KOH.
Pileus – Generative hyphae thin-walled, colorless, rarely branched, inflated, interwoven, mostly 4–7.5 μm diam.
Spines – Generative hyphae thin-walled, colorless, partially crystalline, unbranched, more or less parallel along spines, long-cell, straight, 2–4 μm diam.
Stipe – Generative hyphae thin-walled, colorless, rarely branched, inflated, long-cell, straight, 3.5–8 μm diam.
Basidia – Clavate, thin-walled, smooth, colorless, with four sterigmata, and a basal simple-septa, 21–31 × 6–7 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores ellipsoid to ovoid, brown, thin-walled, tuberculate, tuberculi usually isolated, less than 1.0 μm long, (3.5–)4–4.5 × 3–3.5(–4) μm, L = 4.09 μm, W = 3.07 μm, Q = 1.14–1.50 (n = 60/2), CB–, IKI–.
Material examined (paratype) – China, Guangxi Zhuang Autonomous Region, Guilin City, Xing'an County, Gaoshang Town, GPS coordinates 25°24′58″ N, 110°35′30″ E, altitude 1,100 m, ground in mixed forest, 3 April, 2024, Yuan 19293 (IFP 020028, GenBank ITS: PQ805353; LSU: PV257892; SSU: PV257931).
Notes – In the phylogenetic tree (Fig. 3), the new species Hydnellum porphyreum is grouped into Hydnellum, and is clustered with Hyd. fuscoindicum and Hyd. liantaishanense. Hyd. porphyreum resembles Hyd. fuscoindicum in having ellipsoid, tuberculate basidiospores[122]. However, Hyd. fuscoindicum differs from Hyd. porphyreum due to its broader pileus (3–13 cm), longer stipe (2–5 μm), and wider basidia (7–8 μm)[122]. Hyd. porphyreum resembles Hyd. bomiense and Hyd. yunnanense in having similar-sized pileus. However, Hyd. bomiense differs from Hyd. porphyreum due to grayish yellow to dark brown pileal surface, longer stipes (up to 2 cm), and spines (up to 1.1 mm)[36]. Hyd. yunnanense differs from Hyd. porphyreum by its grayish red to dark brown pileal surface, longer stipes (up to 4 cm) and larger basidiospores ([4.1–]4.2–5.1[–5.3] × [3.4–]3.5–4.5[–5] µm)[36].
Hydnellum testaceum L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 24,25
Fungal Names number: FN 572436

Figure 24.
Basidiomata of Hydnellum testaceum (holotype IFP 020030).
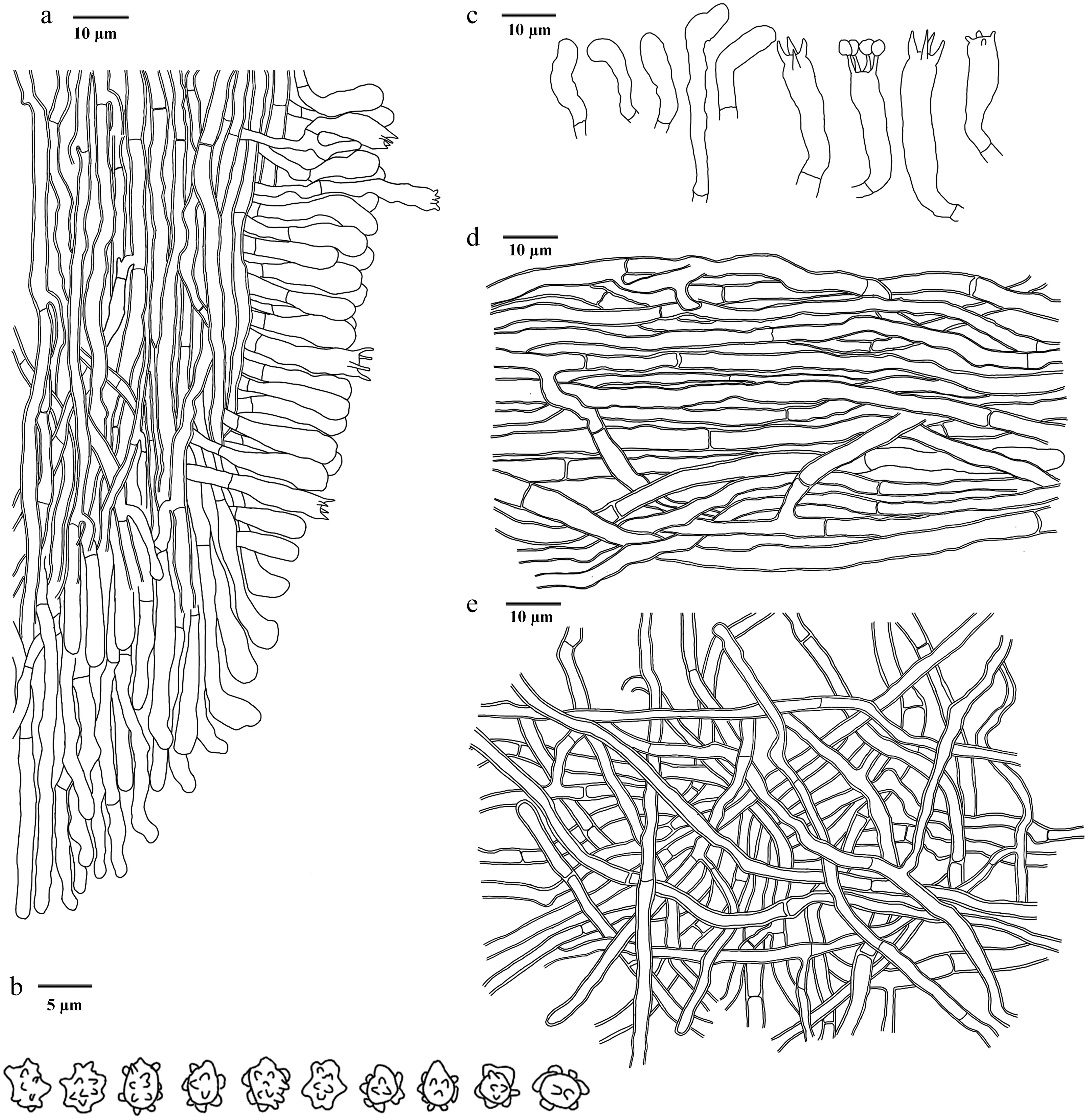
Figure 25.
Microscopic structures of Hydnellum testaceum (drawn from the holotype IFP 020030). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Hydnellum testaceum is characterized by the choanoid, ellipsoid to subcircular pileus, vivid red to brick red pileal surfaces, spines sometimes in groups of three, and ellipsoid to subglobose basidiospores.
Etymology – Testaceum (Lat.): referring to the basidiomata being brick-red when fresh.
Type – China, Zhejiang Province, Shaoxing City, Xinchang county, GPS coordinates 29°30′11″ N, 120°53′59″ E, altitude 300 m, ground in Pinus spp. forest, 21 April, 2024, Yuan 19305 (holotype: IFP 020030, GenBank ITS: PQ805346; LSU: PV257887; SSU: PV257927).
Description – Basidiomata terrestrial, stipitate, annual, gregarious, fleshy when fresh, becoming hard and brittle when dry, corky, taste mild, not releasing a smell. Pileus choanoid, ellipsoid to subcircular, 40–50 mm long, and 30–45 mm across, occasionally duplex, often incurved margins, hard and brittle, usually uneven and bumpy, concentrically zonate, surface covered with velvety to tomentose, and becoming crateriform with age, away from the center, and becoming lighter in color, vivid red (9A8–11A8) to brick red (7D7) when fresh, brown (6D4–7F8) when dry. Spines conical, solitary, more than 2 mm long, sometimes in groups of three, decurrent on stipe, brick red (7D7) when fresh, brown (6D4–7F8) when dry, brittle. Stipe clavate, middle, surface with tomentose, inside corky, usually incorporates individual litter, brick red (7D7) when fresh, brown (6D4–7F8) when dry, hard and brittle.
Hyphal structure – Hyphal system monomitic, generative hyphae with simple-septa, colorless, smooth, thin- to slightly thick-walled, CB–, IKI–, tissues black in KOH.
Pileus – Generative hyphae thin- to slightly thick-walled, infrequent branched, regularly arranged, parallel, uninflated, sometimes flexuous, long-cell, unequal septate, 2–5 μm diam.
Spines – Generative hyphae thin- to slightly thick‐walled, frequent branched, parallel along spines, uninflated, long-cell, unequal septate, sometimes flexuous, 2–3 μm diam.
Stipe – Generative hyphae slightly thick-walled, long-cell, unequal septate, irregularly interwove, infrequent branched, uninflated, sometimes flexuous, 2–4 μm diam.
Basidia – Clavate, thin-walled, colorless, smooth, four sterigmata, constricted at the basal, 21‒34 × 4‒6 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores ellipsoid to subglobose, colorless, thin-walled, tuberculate, tuberculi usually isolated, sometimes in groups of two, up to 1 μm long, (2.5‒)3.0–4 × 2.5‒3.5 μm, L = 3.6 μm, W = 3.0 μm, Q = 1–1.38 (n = 30/1), CB–, IKI–.
Notes – In the phylogenetic tree (Fig. 3), the new species Hydnellum testaceum is grouped into Hydnellum, forming a monophyletic lineage closely associated with Hyd. atrorubrum. Hyd. testaceum shares similarities with Hyd. atrorubrum characterized by the monomitic hyphal system with simple-septate generative hyphae and tomentose stipe surface. However, Hyd. atrorubrum can be delimited from Hyd. testaceum by its darker color of pileus surface (surface light brown to dark ruby), and larger basidiospores ([4.1–]4.5–6[–6.1] × [3.2–]3.9–5.1[–6] μm)[36]. Hyd. testaceum shares similarities with Hyd. squamulosum and Hyd. yunnanense showing red pileal surface with white margin. However, Hyd. squamulosum can be distinguished from Hyd. testaceum in its smaller pileus (< 35 mm), shorter spines (< 2 mm), and larger basidiospores ([4–]4.1–5[–5.1] × [3.2–]3.3–4.1[–4.2] µm)[36]. Hyd. yunnanense differs from Hyd. testaceum by its shorter spines (< 1.5 mm), and basidia (13–28 μm)[36].
Hydnellum tomentosum L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 26,27
Fungal Names number: FN 572430

Figure 26.
Basidiomata of Hydnellum tomentosum (holotype IFP 020019).
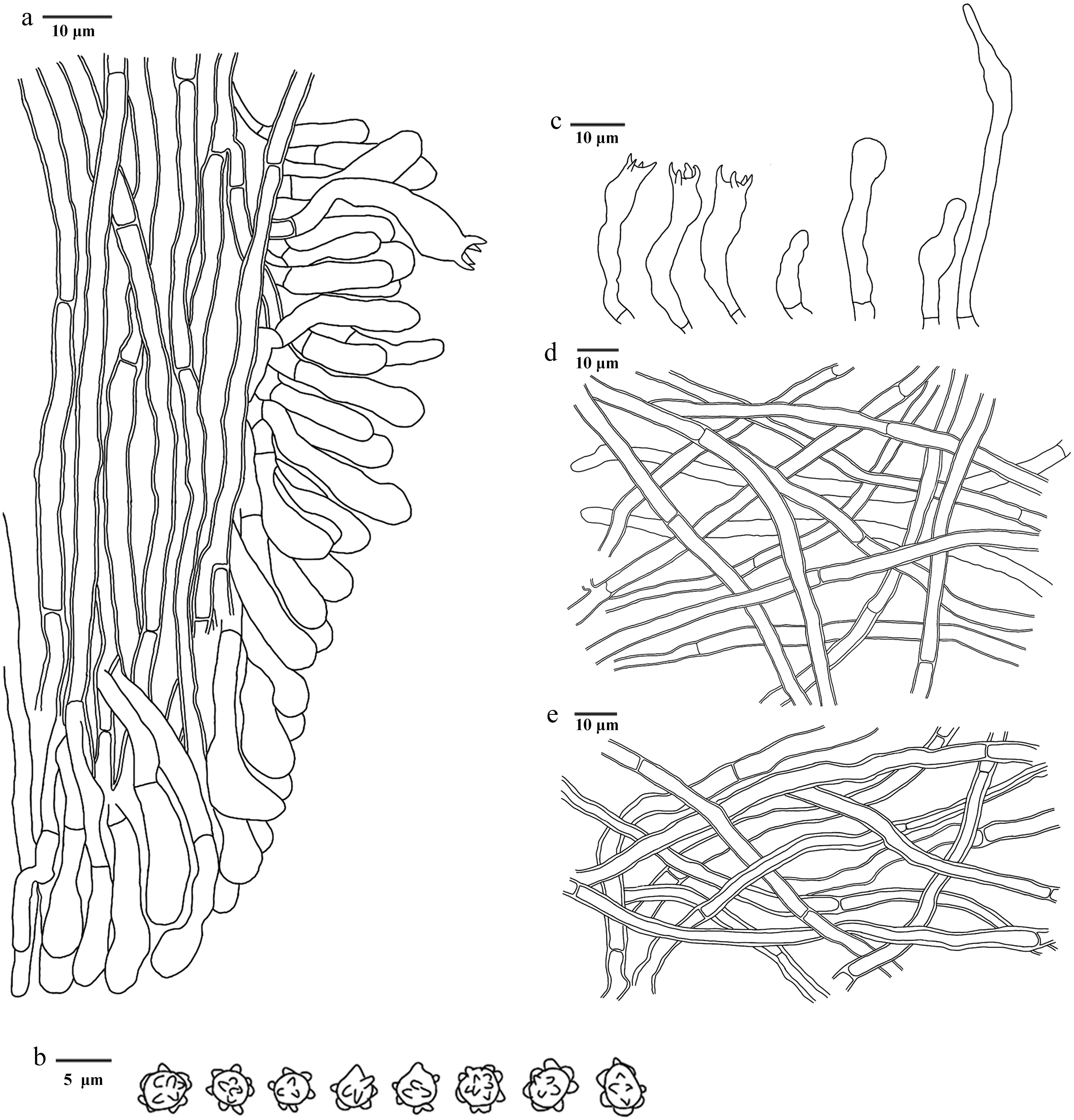
Figure 27.
Microscopic structures of Hydnellum tomentosum (drawn from the holotype IFP 020019). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Hydnellum tomentosum is characterized by the flabelliform to subcircular pileus, pinkish white to brown pileal surfaces, tomentose stipes, and clavate or sinuous basidia.
Etymology – Tomentosum (Lat.): referring to the pileus covered with tomentum.
Type – China, Yunnan Province, Dali Bai Autonomous Prefecture, Dali City, Shuanglang Town, Liantai Mountain, GPS coordinates 25°56′37″ N, 100°17′18″ E, altitude 2,988 m, ground in mixed forest, 7 September, 2024, Yuan 21017 (holotype: IFP 020019, GenBank ITS: PQ805350; LSU: PV257889).
Description – Basidiomata terrestrial, stipitate, annual, gregarious, fleshy when fresh, becoming hard and brittle when drying, taste mild, not releasing a smell. Pileus flabelliform to subcircular, 15–50 mm long, and 11–37 mm across, occasionally duplex, often incurved margins, stiff and brittle, usually uneven and bumpy, concentrically zonate, surface covered with velvety to tomentose, and becoming crateriform with age, away from the center, and becoming lighter in color, pinkish white (10A2) to violet brown (10E4–11F8) when fresh, pale red (7A3–12A3) to brown (6D4–7F8) when dry. Spines conical, solitary, more than 3 mm long, decurrent on stipe, brittle when dry, flesh light brown (5D4–7D8), surface pastel red (7A4–10A4) at basal and white at apex when fresh, greyish ruby (12C3–12E7) to dark brown (6F4–9F8) when dry. Stipe clavate, 16–50 mm long, and 5–13 mm across, middle, surface with tomentose, inside corky, usually incorporates individual litter, hard and brittle when dry, brown (6D4–7F8) when fresh and dry.
Hyphal structure – Hyphal system monomitic, generative hyphae with simple-septa, thin- to thick-walled, CB+, IKI–, tissues yellowish in KOH.
Pileus – Generative hyphae thin- to slightly thick-walled, colorless, sparsely branched, irregularly arranged, uninflated, sometimes flexuous, long-cell, 2–6 μm diam.
Spines – Generative hyphae slightly thick-walled, colorless, infrequent branched, parallel along spines, uninflated, long-cell, straight, 2–4 μm diam.
Stipe – Generative hyphae thick-walled, colorless, long-cell, irregularly arranged, unbranched, uninflated, sometimes flexuous, the septate unequal, 3–6 μm diam.
Basidia – Clavate or sinuous, thin-walled, smooth, colorless, four sterigmata, constricted at the basal, 19‒29 × 3‒4 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subglobose to globose, colorless, thin-walled, tuberculate, tuberculi usually isolated, sometimes in groups of two, up to 1 μm long, 3.0–4 × (2.6‒)2.8‒3.0(‒3.2) μm, L = 3.3 μm, W = 3.0 μm, Q = 1–1.33 (n = 60/2), CB–, IKI–.
Material examined (paratype) – China, Yunnan Province, Dali Bai Autonomous Prefecture, Nanjian Yi Autonomous County, Lingbao Mountain National Forest Park, GPS coordinates 24°46′37″ N, 100°30′23″ E, altitude 2,500 m, ground in mixed forest, 19 September, 2019, Yuan 14387 (IFP 020020, GenBank ITS: MW579970; LSU: MW579908; SSU: MW579934).
Notes – The new species Hydnellum tomentosum is identified as a sister to Hyd. melanocarpum (Fig. 3). Hyd. tomentosum resembles Hyd. melanocarpum in sharing the odorless basidiomata. However, Hyd. melanocarpum differs from Hyd. tomentosum due to its glabrous stipe, wider basidia (18–38 × 5–7 µm) and smaller basidiospores (4.5–5.5[–6] × [3.5–]3.8–5.1 µm)[94]. Hyd. tomentosum shares similarities with Hyd. atrorubrum and Hyd. brunneorubrum showing brownish red pileus with white margin. However, Hyd. atrorubrum can be distinguished from Hyd. tomentosum in its larger basidia (20–48 × 5–8 µm), and basidiospores ([4.1–]4.5–6[–6.1] × [3.2–]3.9–5.1[–6] µm)[36]. Hyd. brunneorubrum differs from Hyd. tomentosum by its longer spines (up to 4 mm), shorter stipe (< 3 cm), and larger basidia (12–50 × 3–7 µm)[36].
Neosarcodon Xiao L. He, Di Wang & W.H. Peng
Index Fungorum number: IF 849989
Type species – Neosarcodon pakaraimensis (A.C. Grupe & T.W. Henkel) Xiao L. He, Di Wang & W.H. Peng
Notes – Larsson et al.[127] identified a clade of stipitate Thelephorales consisting of neotropical Sarcodon species, proposing that it might represent an undescribed genus, temporarily referred to as 'Neosarcodon'. Neosarcodon was later formally described by Wang et al.[64]. Neosarcodon has terrestrial, stipitate-pileate basidiomata. Pileus conic, surface smooth to fibrillose. Stipe hollow to solid, concolorous with pileus or slightly paler. Spines adnate, white or pallid at first, later with some shade of brownish gray. Context fleshy, soft, brittle, whitish to pale grayish. Odor indistinct. Hyphae inflated and thin-walled, frequent clamp connections. Basidiospores generally subglobose, tuberculate, brown in mass. Cystidia absent. In this study, species diversity of Neosarcodon in South China was analyzed based on morphological and molecular evidence (ITS, nLSU, and nSSU) (Fig. 4), two new species N. atroviolaceus and N. bambusicola are introduced.
Neosarcodon atroviolaceus L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 28,29
Fungal Names number: FN 572437

Figure 28.
Basidiomata of Neosarcodon atroviolaceus (holotype IFP 020031). Photo by Jian-Feng Tan.
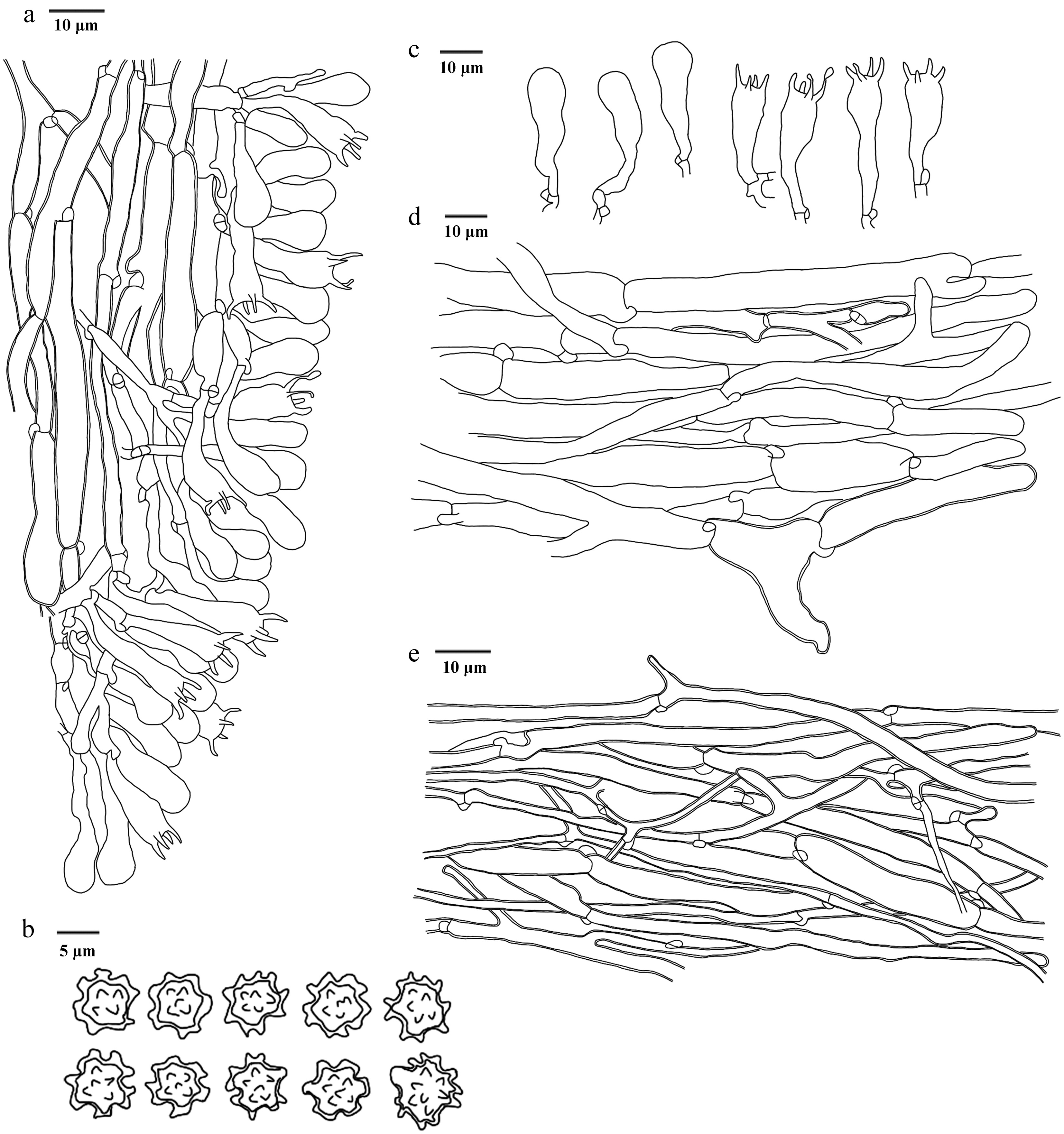
Figure 29.
Microscopic structures of Neosarcodon atroviolaceus (drawn from the holotype IFP 020031). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from context. (e) Hyphae from stipe.
Diagnosis – Neosarcodon atroviolaceus is characterized by the dark violet pileus when fresh, spines growing to the pileus margin, stipe covered with reticulated fiber and epllisoid to globose basidiospores.
Etymology – Atroviolaceus (Lat.): referring to the basidiomata is dark violet when fresh.
Type – China, Guangdong Province, Guangzhou City, Huangpu District, Tianlu Lake, GPS coordinates 23°13′49″ N, 113°24′20″ E, altitude 116 m, ground in bamboo groves, 13 May, 2024, Yuan 19330 (holotype IFP 020031, GenBank ITS: PV221948; SSU: PV440981).
Description – Basidiomata terrestrial, stipitate, annual, gregarious, soft and fleshy when fresh, becoming brittle and light in weight upon dry, taste mild, slight smell. Pileus subcircle to circle, 30–50 mm long, and 30–45 mm across, smooth, glossy, thin, hard, brittle, often incurved margins, dark violet (15F3–18F8) when fresh, yellowish brown (5D4–5F8) when dry. Spines conical, stocky, less than 3 mm long, isolated, growing to the edge of pileus, without decurrent on stipe, schizogenous, deep violet (15D8–18E8) when fresh, reddish brown (8D4–9F8) to dark brown (6F4–9F8) when dry, brittle. Stipe clavate to cylindrical, 40–55 mm long, and 5–10 mm across, middle, smooth, covered with reticulated fiber, unincorporates litter, deep violet (15D8–18E8) when fresh, olive brown (4D3–4F8) when dry, hard.
Hyphal structure – Hyphal system monomitic, generative hyphae with clamp connections, colorless, thin- to slightly thick-walled, CB+, IKI–, tissues black in KOH.
Pileus – Generative hyphae thin- to slightly thick-walled, smooth, colorless, moderately branched, uninflated, long cell, unequal septate, parallel interwoven, 4–13 μm diam.
Spines – Generative hyphae thin- to slightly thick-walled, smooth, colorless, sparsely branched, parallel interwoven along spines, straight, long cell, unequal septate, 3–14 μm diam.
Stipe – Generative hyphae slightly thick-walled, smooth, colorless, frequently branched, parallel interwoven, more or less flexous, 2–8.5 μm diam.
Basidia – Clavate, thin-walled, smooth, colorless, 4 sterigmata, sterigmata up to 6 μm, inflated at the apex, with a clamp connection at the base, 24–37 × 7–10 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores epllisoid to globose, colorless, slightly thick-walled, tuberculate, tuberculi usually isolated or grouped in 2, less than 2.0 μm long, 5–6 × (4.5–)5–5.5(–6) μm, L = 5.2 μm, W = 5.0 μm, Q = 1–1.1 (n = 30/1), CB+, IKI–.
Notes – In the phylogenetic tree (Fig. 4), the new species Neosarcodon atroviolaceus is closely associated with N. pallidogriseus. However, N. pallidogriseus can be delimited from N. atroviolaceus by its larger basidia ([30−]34−41[−44] × 10−13[−15] μm), narrower pileus (8−16 mm broad), and stipe (2−5 mm broad)[129]. N. atroviolaceus is similar to N. quercophilus in having deep violet pileus, spine, and stipe[130]. However, N. quercophilus differs from N. atroviolaceus by its wider basidiospores (7–9 μm), and longer basidia (10–16 μm)[130].
Neosarcodon bambusicola L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 30,31
Diagnosis – Neosarcodon bambusicola is characterized by the greyish green to brown pileus, stipe covered with reticulated fiber and epllisoid to subglobose basidiospores.
Fungal Names number: FN 572439

Figure 30.
Basidiomata of Neosarcodon bambusicola (holotype IFP 020033). Photo by Zhong-Ping Feng.
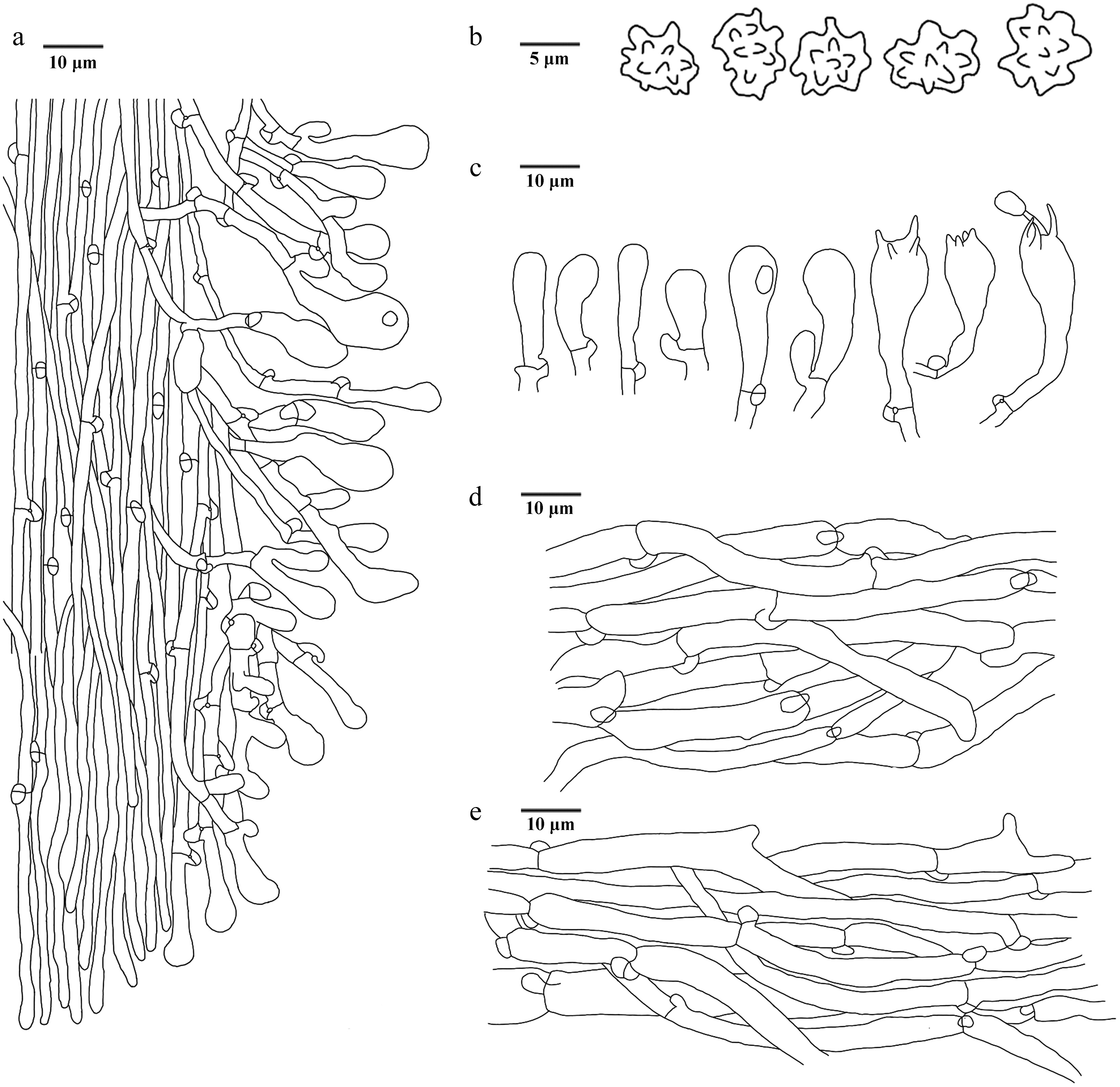
Figure 31.
Microscopic structures of Neosarcodon bambusicola (drawn from the holotype IFP 020033). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Etymology – Bambusicola (Lat.): referring to this species exclusively growing in bamboo groves.
Type – China, Sichuan Province, Guang'an City, Linshui County, Dingping Town, GPS coordinates 30°20′10″ N, 106°55′42″ E, altitude 351 m, ground in bamboo groves (Phyllostachys edulis), 2 July, 2024, Yuan19491 (holotype: IFP 020033, GenBank ITS: PV221949; SSU: PV440979).
Description – Basidiomata terrestrial, stipitate, annual, solitary to gregarious, soft and fleshy when fresh, becoming brittle and light in weight upon dry, taste mild, not releasing a smell. Pileus subcircle to circle, 30–55 mm long, and 25–50 mm across, hard, brittle, smooth, surface without tomentose, often incurved margins, greyish green (1C3–1D7) to brown (6D4–7F8) when fresh, greyish green(1C3–1D7) to olive green (2F6) when dry, involuted. Spines conical, up to 2 mm long, isolated, without decurrent on stipe, violet grey (15B2–18F2) when fresh, olive green (2F6) when dry, brittle. Stipe clavate, 50–55 mm long, and 15–20 mm across, middle, smooth, covered with reticulated fiber, unincorporates litter, dark violet (15F3–18F8) when fresh, olive green (2F6) when dry, hard.
Hyphal structure – Hyphal system monomitic, generative hyphae with clamp connections, thin-walled, CB–, IKI–, tissues black in KOH.
Pileus – Generative hyphae thin-walled, smooth, colorless, sparsely branched, uninflated, unequal septate, parallel interwoven, 4–8 μm diam.
Spines – Generative hyphae thin-walled, smooth, colorless, sparsely branched, parallel along spines, straight, 1.5–3 μm diam.
Stipe – Generative hyphae thin-walled, smooth, colorless, unbranched, parallel interwoven, more or less flexous, 3–8 μm diam.
Basidia – Clavate, thin-walled, smooth, colorless, 4 sterigmata and with a simple-septa at the base, inflated at the apex, 18–29 × 6–10 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores epllisoid to subglobose, colorless, thin-walled, tuberculate, tuberculi usually isolated or grouped in twos, less than 2.0 μm long, (4–)4.5–5 × 3–4.5 μm, L = 4.86 μm, W = 3.98 μm, Q = 1.11–1.43 (n = 30/1), CB–, IKI–.
Note – The new species Neosarcodon bambusicola is grouped within Neosarcodon, and clusters with N. atroviridis, N. portoricensis, and N. quercophilus (Fig. 4). N. atroviridis can be delimited from N. bambusicola by its longer basidia ([25.0–]30.0–50.0 × [6.0–]7.5–11.0 μm) and larger basidiospores (7.0–8.5[–9.0] × 6.0–7.5 μm)[131]. N. portoricensis differs from N. bambusicola by its grayish brown pileus surface, smaller stipe (22–48 × 5–7 mm) and basidia ([30–]33–44[–48] × 8–13 µm)[130]. N. quercophilus differentiates from N. bambusicola in its narrower pileus (18–33 mm broad), slenderer stipe (40–75 × 4–8 mm) and larger basidiospores (5–7 × 7–9 µm)[130]. N. bambusicola resembles N. bairdii in having greyish to olive pileus[129]. However, N. bairdii differs from N. bambusicola by its grayish brown spines, shorter stipes (30–50 mm) and larger basidiospores ([5–]6–7 × 7–8[–9] µm)[129].
Sarcodon Quél. ex P. Karst.
Index Fungorum number: IF 18501
Type species – Sarcodon imbricatus (L.) P. Karst.
Notes – Sarcodon was established by Finnish mycologist Petter Adolf Karsten[132], with Sarcodon imbricatus designated as the type species. Karsten[133] assigned it in the Hydnaceae within the Polyporales based on the characteristic of hydnoid hymenophore. However, the taxonomic position of Sarcodon has long been a subject of debate. Banker[134] maintained its placement in the Hydnaceae, while other studies suggested a closer affinity with the Thelephoraceae. As mycological taxonomy advanced, the taxonomic position of Sarcodon gradually became clearer. Maas Geesteranus[124] conducted a systematic morphological study of Sarcodon species in Europe, describing several new species and revising the boundaries of the genus. Phylogenetic analyses have shown that the genus Sarcodon belongs to the order Thelephorales, and is closely related to the genera Hydnellum and Neosarcodon[35,64,126,135]. In this study, one new species of Sarcodon is introduced from China based on morphological characteristics and phylogenetic analyses inferred from ITS, nLSU, and nSSU sequences (Fig. 4).
Sarcodon squamulosus L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 32,33
Fungal Names number: FN 572444

Figure 32.
Basidiomata of Sarcodon squamulosus (holotype IFP 020045). Photo by Yang-Ling Deng.
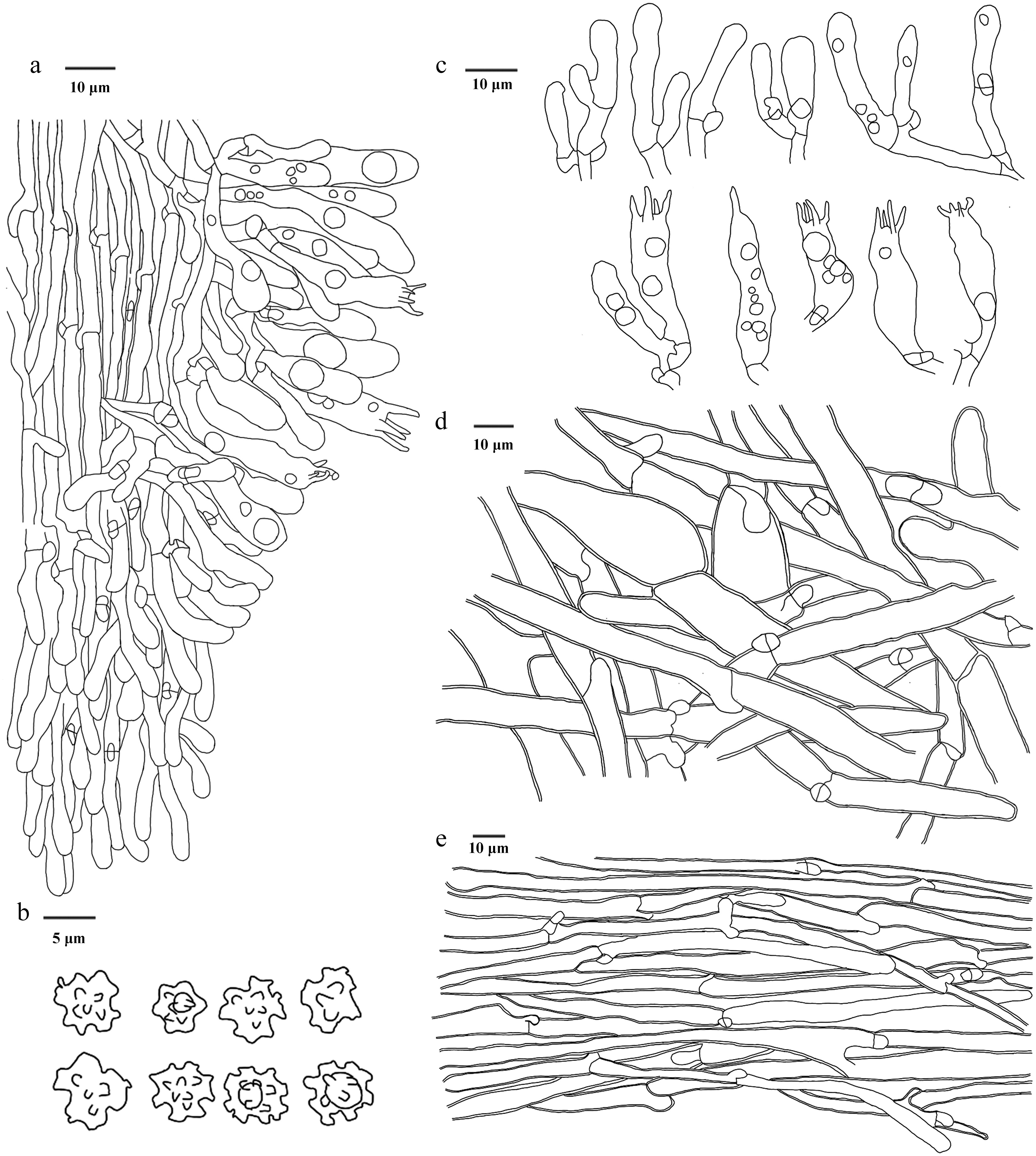
Figure 33.
Microscopic structures of Sarcodon squamulosus (drawn from the holotype IFP 020045). (a) Section through spines. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphae from pileus. (e) Hyphae from stipe.
Diagnosis – Sarcodon squamulosus is characterized by the cream basidiomata when fresh, tomentose stipe surface, decurrent spines on the stipe, and basidiospores with oily-like contents.
Etymology – Squamulosus (Lat.): referring to the pileal surface covered with small scales.
Type – China, Hunan Province, Shaoyang City, Xinning County, Huilongsi Town, GPS coordinates 26°44′33″ N, 111°6′53″ E, altitude 300 m, ground in mixed forest, 28 September, 2024, Yuan 21453 (holotype: IFP 020045, GenBank ITS: PQ805359; LSU: PV257883; SSU: PV257923).
Description – Basidiomata terrestrial, stipitate, annual, solitary to gregarious, soft and fleshy when fresh, becoming brittle and light in weight upon dry, taste slightly bitter, not releasing a smell. Pileus planar, ellipsoid to circle, 45–60 mm long, and 35–50 mm across, often incurved margins, involuted after drying, radiating projecting fibrils aggregate into bundles becoming dark brown (6F4–9F8) scales-like, beige when fresh and later to black, cream (4A3) when fresh, greyish yellow (4B3–4B6) to light brown (5D4–7D8) when dry, rarely lobed. Spines conical, up to 2 mm long, isolated, decurrent on stipe, cream (4A3) when fresh, surface beige (4C3) to light brown (5D4–7D8), inner beige (4C3) when dry, brittle. Stipe clavate, middle, surface with tomentose, becoming fibrils when dry covered with surface, usually unincorporates litter, cream (4A3) when fresh, pale yellow (1A3–4A3) when dry, brittle.
Hyphal structure – Hyphal system monomitic, generative hyphae with clamp connections, colorless, smooth, thin- to slightly thick-walled, CB+, IKI–, tissues unchanged in KOH.
Pileus – Generative hyphae slightly thick-walled, sparsely branched, inflated, and up to 21 μm, long cell, unequal septate, irregularly interwoven, 3–14 μm diam.
Spines – Generative hyphae thin-walled, infrequently branched, parallel along spines, straight, 2–4 μm diam.
Stipe – Generative hyphae slightly thick-walled, infrequently branched, parallel, straight, 3–11 μm diam.
Basidia – Clavate, smooth, colorless, thin-walled, four sterigmata, and with a clamp connection at the base, 21–48 × 5–9 μm, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subglobose to globose, irregular, more or less with oily-like contents, colorless, thin-walled, tuberculate, tuberculi usually isolated or grouped in two or more, bi- to trifurcate-like in shape, up to 2.0 μm long, (3–)4–5 × 3–4.5(–5) μm, L = 4.5 μm, W = 3.8 μm, Q = 1–1.33 (n = 60/2), CB–, IKI–.
Material examined (paratypes) – China, Chongqing City, Banan District, Fengsheng Town, GPS coordinates 29°31′18″ N, 106°56′5″ E, altitude 550 m, ground in Pinus spp. and Quercus spp. forests, 12 October, 2023, Yuan 19208 (IFP 020046, GenBank ITS: PQ805361), Yuan 19209 (IFP 020047, GenBank ITS: PQ805360).
Notes – In this study, the new species Sarcodon squamulosus is placed within Sarcodon (Fig. 4), and it forms a clade with S. quercinofibulatus. S. squamulosus resembles S. quercinofibulatus by the monomitic hyphal system with clamped generative hyphae. However, S. quercinofibulatus differs from S. squamulosus by its wider basidia (30–40 µm vs 8–10 µm), larger basidiospores ([6–]6.5–7.4[–8] µm vs [5–]5.4–6.4[–7] µm) and larger pileus (14 cm)[136]. S. squamulosus is similar to S. leucopus in sharing off-white pileus, and fleshy basidiomata[64]. However, S. leucopus can be distinguished from S. squamulosus by its smooth pileal surface, shorter spines (< 1.5 mm), and presence of cystidia[64,137]. S. squamulosus is similar to S. pseudoimbricatus in sharing pileus with scales. However, S. pseudoimbricatus differs from S. squamulosus by its longer spines (up to 0.7 cm), larger basidiospores (6–7.5×4.5–5.5[–6] μm), and the presence of cystidia[64].
Sequence alignment analysis indicated that the ITS sequence of this species shared high similarity with the corresponding sequence in the UNITE database, with only one nucleotide difference identified between them. This sequence was obtained from a sample collected in Chuxiong (Yunnan Province, China), representing a new distribution site of this species in China.
Thelephoraceae Chevall.
Index Fungorum number: IF 81463
Type genus – Thelephora Ehrh. ex Willd.
Odontia Pers.
Index Fungorum number: IF 790576
Type species – Odontia ferruginea Pers.
Notes – Odontia was established by Persoon in 1794, with O. ferruginea designated as the type species. Historically, Odontia has been treated as a synonym of Tomentella. However, the phylogenetic analyses have revealed that Odontia constitutes a monophyletic lineage related to Tomentella. Notably, species within Odontia exhibit a saprotrophic lifestyle, distinct from the ectomycorrhizal habit typical of Tomentella[49,138]. Morphologically, Odontia is characterized by resupinate, arachnoid basidiomata that are easily separable from the substrate; a hymenophoral surface ranging from smooth to granulose or hydnoid; a dimitic hyphal system in rhizomorphs; a monomitic or dimitic hyphal system in the subiculum; clavate basidia and irregular ellipsoid to globose basidiospores with verruculose ornamentation[43,139,140]. Species of Odontia have been reported across temperate regions of Eurasia, North America, Africa, France, Australia, New Zealand, China, and Turkey[43,49,138,139,141−143]. In the present study, a new species of Odontia from China is described, based on morphological characteristics, and phylogenetic analyses inferred from ITS, nLSU, and nSSU sequences (Fig. 5).
Odontia kunmingensis Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 34,35
Fungal Names number: FN 572440

Figure 34.
Basidiomata of Odontia kunmingensis (holotype IFP 020034).
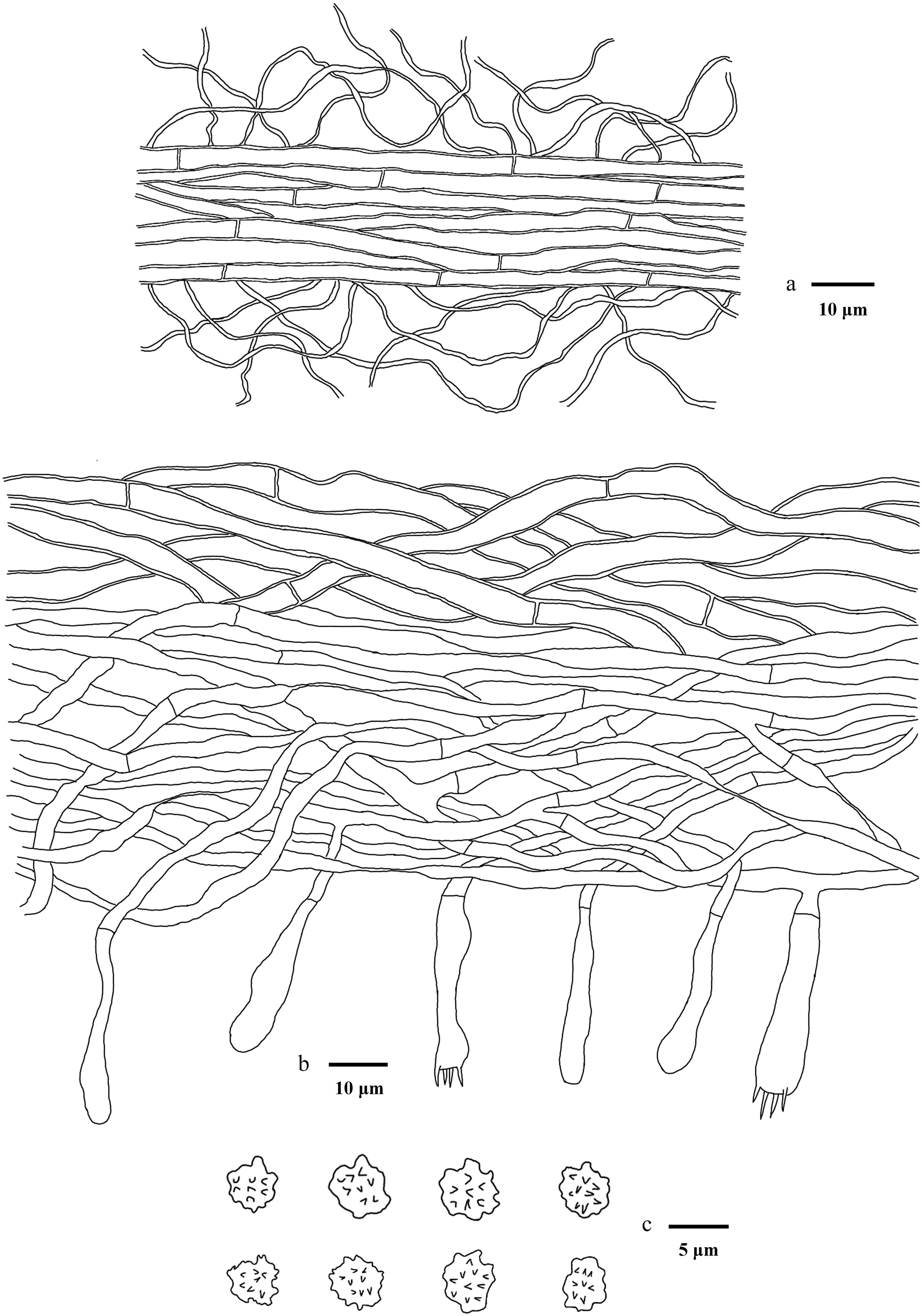
Figure 35.
Microscopic structures of Odontia kunmingensis (drawn from the holotype IFP 020034). (a) Section through rhizomorph. (b) Section through basidiomata. (c) Basidiospores in frontal and lateral view.
Diagnosis – Odontia kunmingensis is characterized by the type C rhizomorphs, yellowish brown to brown hymenophoral surface, and irregular subglobose to ellipsoid basidiospores.
Etymology – Kunmingensis (Lat.): referring to the Kunming City, where the type is collected.
Type – China, Yunnan Province, Kunming City, the Golden Hall Scenic Area, GPS coordinates 25°5′4″ N, 102°46′19″ E, altitude 2,000 m, on fallen angiosperm branch, 17 August, 2023, Yuan 18400 (holotype: IFP 020034, GenBank ITS: PV259700; LSU: PX024029; SSU: PV505424).
Description – Basidiomata, annual, resupinate, easily separable from the substrate, arachnoid, continuous, soft coriaceous to leathery, and without odor or taste when fresh, becoming cottony upon drying, up to 0.2 mm thick. Hymenophoral surface yellowish brown to brown (5D8–6F8) when dry, smooth, turning lighter or concolorous with subiculum. Subiculum mostly brown. Sterile margin determinate, byssoid, concolorous, with hymenophore.
Rhizomorphs – Present in subiculum and margins, 20–50 μm diam, rhizomorph surface rather smooth, hyphal structure in rhizomorphs dimitic, differentiated, of type C (according to Agerer 1987–2008), compactly arranged, generative hyphae in the center of the rhizomorph with simple-septate, thick-walled, 2–3 μm diam, colorless in KOH, skeletal hyphae at outer part of rhizomorph thin-walled, 1 μm diam, grayish yellow in KOH, CB+, IKI–.
Hyphal structure – Hyphal system monomitic, generative hyphae with simple-septa, thick-walled, 4–6 μm diam, without encrustation, grayish yellow in KOH, CB+, IKI–.
Subhymenium – Generative hyphae with simple-septa, thin-walled, 2–4 μm diam, occasionally collapsed, hyphal cells more or less uniform, colorless in KOH, CB–, IKI–.
Basidia – 35–40 μm long and 4–7 μm diam at apex, 2–4 μm at base, clavate, not stalked, not sinuous, grayish yellow in KOH and in distilled water, 4 sterigmata and with a simple-septa at the base, sterigmata 2–4 μm long, 0.5–1 μm diam at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores irregular subglobose to ellipsoid in frontal face and irregular subglobose in lateral face, oildrops absent, light brown in KOH, and in distilled water, thin-walled, verruculose, warts usually grouped in two or more, alike in shape, 0.5 μm long, (5.9–)6.2–8.2(–8.9) × (5.4–)5.8–7.5(–7.8) μm, L = 7.18 μm, W = 6.51 μm, Q = 1–1.3 (n = 30/1), CB–, IKI–.
Notes – The new species Odontia kunmingensis forms a clade with O. fibrosa and O. parvispina (Fig. 5). They share similar morphological and anatomical characteristics: annual, resupinate, arachnoid basidiomata, brown subiculum, the presence of rhizomorphs with a dimitic hyphal system, simple-septate generative hyphae in the subiculum, and clavate basidia[139,140]. However, O. fibrosa can be differentiated by thicker basidiomata (up to 0.4 mm vs up to 0.2 mm in O. kunmingensis), dark brown and granulose hymenophoral surface when dry, longer spines (up to 1 mm), and larger basidiospores (7.1–9 × 6.1–8 μm vs 6.2–8.2 × 5.8–7.5 μm in O. kunmingensis). O. parvispina differs from O. kunmingensis by its hydnoid hymenophoral surface (spines up to 0.06 mm long), narrower generative hyphae (3–4 μm vs 4–6 μm in O. kunmingensis), and smaller basidiospores (6–7 × 5–6.4 μm vs 6.2–8.2 × 5.8–7.5 μm in O. kunmingensis)[140].
Thelephora Ehrh. ex Willd.
Index Fungorum number: IF 18640
Type species – Thelephora terrestris Ehrh.
Notes – Thelephora Ehrh. ex Willd. was established by Willdenow, with Thelephora terrestris Ehrh. designated as the type species[144−146]. In 1794, Persoon discovered and described the first villaceous fungus, Odontia ferruginea Pers, which was subsequently transferred to Tomentella as Tomentella ferruginea (Pers.) Pat. by Patouillard in 1887. Since then, taxonomic studies on Tomentella have gradually advanced, revealing that the genus exhibits diverse and complex morphological features, including basidiomata with various colors, basidiospores with diverse shapes and ornamentations, and hymenophoral surfaces ranging from smooth to granulose[43,142]. Thelephora and Tomentella are taxonomically closely related, typically forming a paraphyletic group within the same evolutionary clade[37,38,147−149]. They share several morphological similarities, such as a monomitic hyphal system with clamped generative hyphae and ornamented (warted to echinulate) basidiospores. However, Tomentella is distinguishable by its resupinate, effused, and tightly adherent basidiomata[8,43,149,150]. In their latest study, Kõljalg et al.[151] revised the classification of Tomentella, merging it with Thelephora. In this study, 193 species were included to construct the phylogenetic tree (Fig. 6), and 12 new species are described and illustrated, based on morphological characteristics, and phylogenetic evidence.
Thelephora angusta Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 36,37
Fungal Names number: FN 572908

Figure 36.
Basidiomata of Thelephora angusta (holotype IFP 020075).
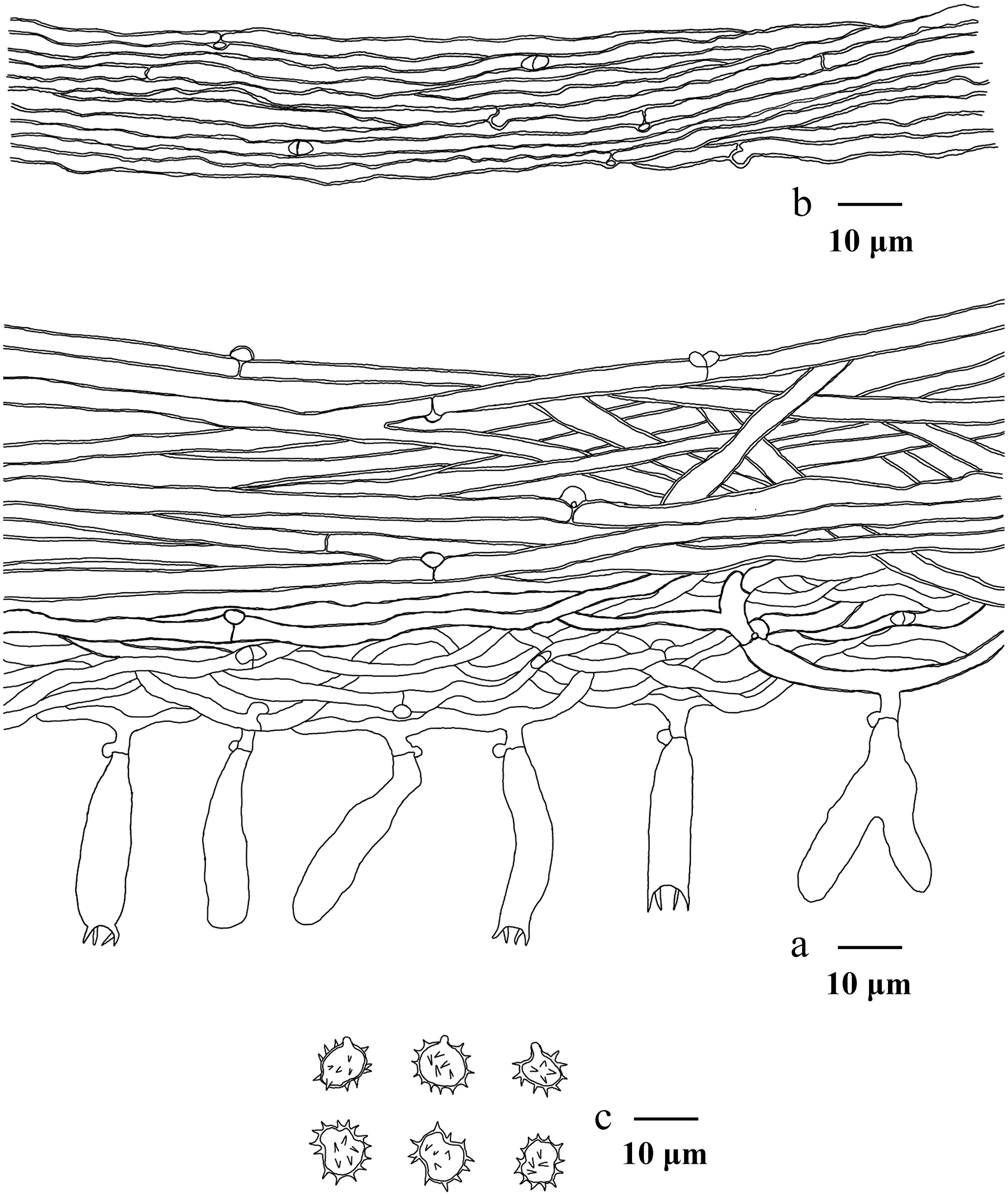
Figure 37.
Microscopic structures of Thelephora angusta (drawn from the holotype IFP 020075). (a) Section through rhizomorph. (b) Section through basidiomata. (c) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora angusta is characterized by the type B rhizomorphs, light brown to reddish grey hymenophoral surface, subglobose to ellipsoid (irregular lobed) basidiospores.
Etymology – Angusta (Lat.): referring to the narrow rhizomorphs of the type.
Type – China, Yunnan Province, Diqing Tibetan Autonomous Prefecture, Shangri-La City, Potatso National Park, GPS coordinates 27°50′12″ N, 99°57′27″ E, altitude 3,655 m, on bark of Abies sp., 15 August, 2023, Yuan 18145 (holotype: IFP 020075, GenBank ITS: PV843164; LSU: PV843171; mtSSU: PV856499).
Description – Basidiomata annual, resupinate, separable from the substrate, mucedinoid, without odor or taste when fresh, 0.4–0.7 mm thick, continuous. Hymenophoral surface smooth, light brown to reddish grey (7D6–7B2), and turning darker than subiculum. Sterile margin often determinate, byssoid, whitish, paler than hymenophore.
Rhizomorphs – Present in subiculum and margins, 15–50 μm diam, rhizomorphic surface more or less smooth, hyphae in rhizomorph monomitic, undifferentiated, of type B, compactly arranged and of uniform, single hyphae with clamp connections and simple-septa, slightly thick- to thick-walled, unbranched, 1–3.5 μm diam, pale brown in KOH, CB+, IKI–.
Subiculum – Hyphal system monomitic, generative hyphae with clamp connections and rarely simple-septa, slightly thick- to thick-walled, branched, 3–6 µm diameter, occasionally collapsed, without encrustation, pale brown in KOH, and distilled water, CB+, IKI–.
Subhymenium – Generative hyphae clamped, slightly thick- to thin-walled, occasionally branched, 1.5–4 µm diameter, without encrustation, hyphal cells: more or less uniform, grayish yellow in KOH and in distilled water, CB+, IKI–.
Basidia – 25–45 µm long and 4–6.5 µm diameter at apex, 3–4 µm at base, with a clamp connection at the base, clavate, not stalked, sinuous, without transverse septa, pale brown in KOH, and distilled water, four sterigmata, sterigmata 2.5–3.5 µm long, and 0.8–1µm diameter at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores ellipsoid (irregular lobed) in frontal and ellipsoid (irregular lobed) in lateral face (in small amounts also ellipsoid to subglobose), pale brown in KOH, and distilled water, thick-walled, echinulate to aculeate, echinuli or aculei usually isolated, sometimes grouped in 2, up to 1.5 µm long, (6.8–)7–8.5(–9.1) × (5.1–)5.6–7.5(–8.2) µm, L = 7.79 µm, W = 6.53 µm, Q = 1–1.48 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Yunnan Province, Diqing Tibetan Autonomous Prefecture, Deqin County, Baima Snow Mountain National Nature Reserve, GPS coordinates 28°20′4″ N, 99°5′49″ E, altitude 3,500 m, fallen angiosperm branch, 13 August, 2023, Yuan 17935 (IFP 020076, GenBank ITS: PV843165; LSU: PV843172; mtSSU: PV856500).
Notes – In this study (Fig. 6), the new species Thelephora angusta is grouped with T. viridula, T. pyrolae, T. subtestacea, T. galzinii, and T. tenuifarinacea. They exhibit some similar characteristics: resupinate basidiomata. However, T. pyrolae differs from T. angusta by its longer basidia (50–65 µm), larger basidiospores (7.5–11 µm across), and the absences of rhizomorphs[142]. T. subtestacea can be distiniguished from T. angusta by its narrower subicular hyphae, absences of rhizomorphs and presences of cystidia[43]. T. galzinii differentiates from T. angusta by its wider subhymenophoral hyphae, and utriform basidia[43]. T. tenuifarinacea differs from T. angusta by its wider basidia, subglobose to globose, and nodulose to verrucose basidiospores[99].
Thelephora bomiensis Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 38,39
Fungal Names number: FN 572449

Figure 38.
Basidiomata of Thelephora bomiensis (holotype IFP 020056).
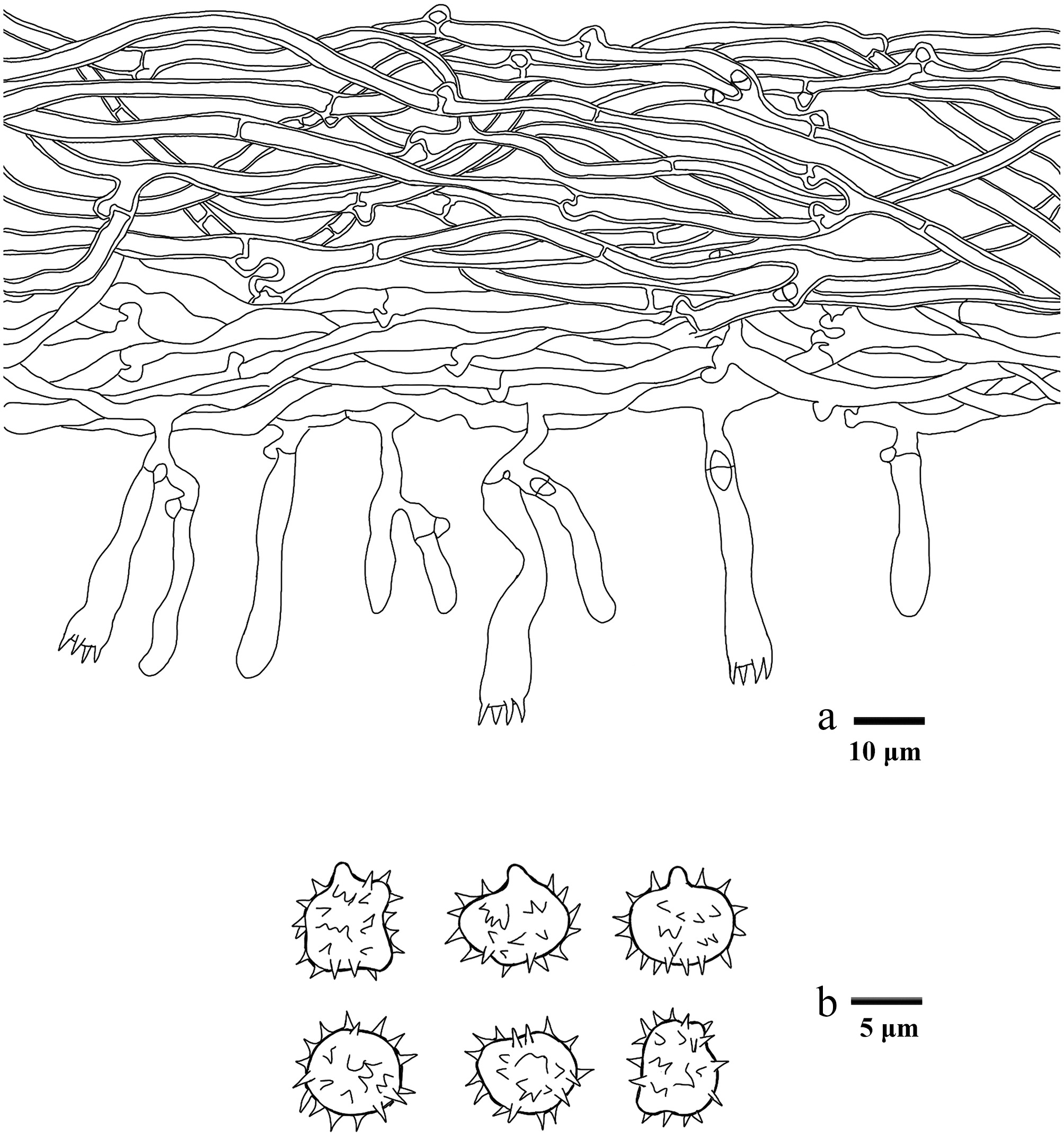
Figure 39.
Microscopic structures of Thelephora bomiensis (drawn from the holotype IFP 020056). (a) Section through basidiomata. (b) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora bomiensis is characterized by the grayish brown hymenophoral surface, long sterigmata (4–7 µm), and irregular ellipsoid to subglobose (irregularly lobed) basidiospores.
Etymology – Bomiensis (Lat.): referring to the locality (Bomi County) of the type.
Type – China, Xizang Autonomous Region, Linzhi City, Bomi County, GPS coordinates 29°52′25″ N, 95°45′5″ E, altitude 2,730 m, on angiosperm wood debris, 17 July, 2019, Yuan 13610 (holotype: IFP 020056, GenBank ITS: PQ656819; SSU: PV440978).
Description – Basidiomata annual, resupinate, adherent to the substrate, arachnoid, without odor or taste when fresh, 0.1 mm thick, uncontinuous. Hymenophoral surface smooth, grayish brown (5D3) and concolorous with the subiculum. Sterile margin often indeterminate, byssoid, concolorous with hymenophore.
Rhizomorphs – Absent.
Subiculum – Hyphal system monomitic, generative hyphae with clamp connections and rarely simple-septa, thick-walled, occasionally branched, 3–5 µm diameter, without encrustation, pale brown in KOH, and distilled water, CB+, IKI–.
Subhymenium – Generative hyphae with clamp connections, thin-walled, occasionally branched, 2.5–4 µm diameter, without encrustation, hyphal cells more or less uniform, grayish yellow in KOH and in distilled water, CB–, IKI–.
Basidia – 25–45 µm long, and 5–7 µm diameter at apex, 3–6 µm at base, with a clamp connection at the base, clavate, not stalked, sinuous, without transverse septa, pale brown in KOH, and distilled water, four sterigmata, sterigmata 4–7 µm long, and 1–2 µm diameter at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores ellipsoid (irregularly lobed) in frontal (in small amounts also subglobose), and ellipsoid (irregularly lobed) in lateral face (in small amounts also subtriangular), pale brown in KOH, and distilled water, slightly thick-walled, echinulate to aculeate, echinuli or aculei usually isolated, sometimes grouped in two or more, up to 1.5 µm long, (5.2–)6.2–8.2(–8.9) × (4.5–)4.9–7.6(–8) µm, L = 7.27 µm, W = 6.22 µm, Q = 1.01–1.32 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Xizang Autonomous Region, Linzhi City, Chayu County, Rangshequ 3rd Middle Bridge, GPS coordinates 28°35′23″ N, 98°20′36″ E, altitude 3,550 m, on fallen branch of Pinus sp., 13 July, 2019, Yuan 13506 (IFP 020057, GenBank ITS: PQ656818; LSU: PV257905; SSU: PV440977).
Notes – In this study (Fig. 6), the new species Thelephora bomiensis is grouped within Thelephora, forming a monophyletic lineage, and is closely associated with T. brevis. T. bomiensis resembles T. brevis by sharing uncontinuous basidiomata, the color of hymenophoral surface, and the shape of basidiospores[102]. However, T. bomiensis differs from T. brevis by its longer basidia, larger basidiospores and the absences of rhizomorphs. T. bomiensis is similar to T. griseocastanea and T. olivaceobrunnea in sharing grayish brown hymenophoral surface[102]. However, T. griseocastanea differs from T. bomiensis by its wider subhymenophoral hyphae (4–6 μm) and smaller basidiospores ([5–]5.5–6.5[–7] × [4–]5–5.5[–6] μm)[102]. T. olivaceobrunnea differentiates from T. bomiensis in its granulose hymenophoral surface and wider subhymenophoral hyphae (4–7 μm)[102].
Thelephora cacao Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 40,41
Fungal Names number: FN 572909

Figure 40.
Basidiomata of Thelephora cacao (holotype IFP 020077).
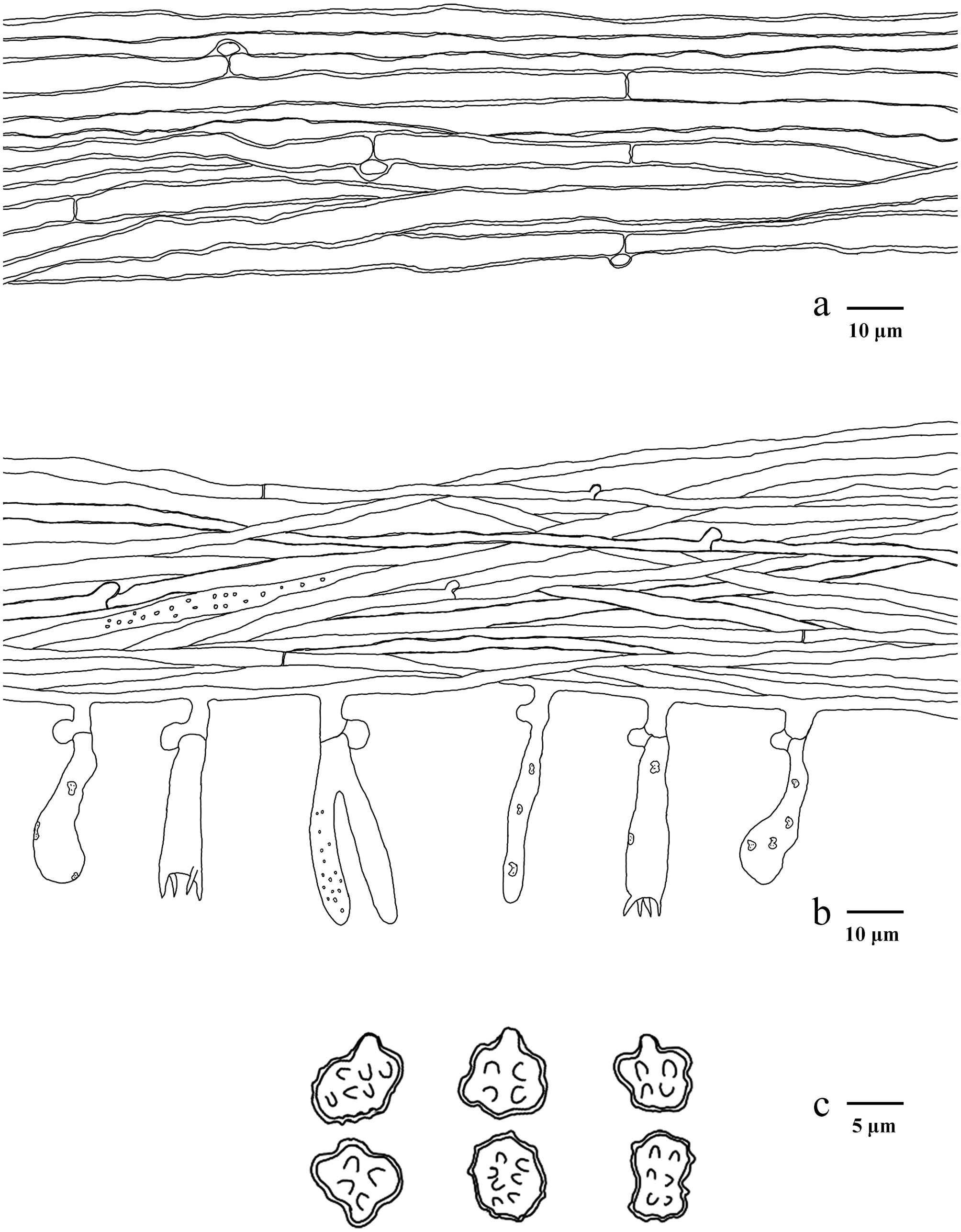
Figure 41.
Microscopic structures of Thelephora cacao (drawn from the holotype IFP 020077). (a) Hyphae from pileal context. (b) Section of hymenium and subhymenium. (c) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora cacao is characterized by the imperceptibly rugose hymenophoral surface, chocolate brown stipe, and lacunary, sulcate, irregularly ridged abhymenial face.
Etymology – Cacao (Lat.): referring to the dark brown hymenophoral surface.
Type – China, Hunan Province, Shaoyang City, Xinning County, Huilongsi Town, GPS coordinates 26°34′48″ N, 111°4′48″ E, altitude 500 m, ground in mixed forest, 5 June, 2024, Yuan 19363 (holotype: IFP 020077, GenBank ITS: PV843166; LSU: PQ803816; SSU: PV856501; mtSSU: PV856546).
Description – Basidiomata upright, up to 13 cm wide, and 3 cm high, solitary to gregarious, humid, and leathery when fresh, corky to hard and brittle when dry, pungent odor when dry. Pileus flat, flabelliform, petaloid to applanate-lobate, usually with multiple pilei arising from a shared stipe or center, and arranged in imbricate, rosette to flabelliform, margin 2–3 mm thick, nearly entire, rarely lacerate, imperceptibly wavy. Abhymenial face lacunary, sulcate, irregularly ridged, zonate, yellowish brown to grayish brown (5E6–5E) at the center then gradually becoming paler towards margin, but reddish blond (5C4) at the margin. Hymenophoral surface imperceptibly rugose, dark brown to brown (7F8–7F5) at the center, and becoming pale brown towards the margin, but brownish orange (5C3) at the margin. Stipe central, 20–30 × 10–25 mm, chocolate brown, subconical to broadened or flatted towards base, surface rugose, glabrous, dark brown (7F8–7F5).
Hyphal structure – Hyphal system monomitic, generative hyphae commonly with clamp connections and rarely simple-septa, CB+, IKI–, tissues turned brown in KOH.
Pileus – Generative hyphae slightly thick to thick-walled (< 1 μm), hyaline, rarely branched, parallel to subparallel, 2.5–5 μm.
Subhymenium – Generative hyphae slightly thick to thin-walled (< 1 μm), hyaline, occasionally branched, often covered by dense crystals, subparallel, up to 1–3 μm.
Basidia – Utriform to clavate, thin-walled, with a clamp connection at the base, multi-guttulate content sometimes very dense, occasionally covered by dense crystals, 20–45 × 4–7 μm, four sterigmata, sterigmata 5–6 μm long, and 1.5–2 μm diam. at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subtriangular to ellipsoid (irregularly lobed) in frontal view and ellipsoid to pyriform (irregularly lobed) in lateral view, brown in KOH, and in distilled water, thick-walled (< 1 μm), tuberculate, tuberculi usually isolated, sometimes in groups of two or more, 6.1–8.3(–8.5) × (5–)5.2–7.1 μm, L = 7.27 μm, W = 6.25 μm, Q = 1–1.41 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Hunan Province, Xiangxi Tujia and Miao Autonomous Prefecture, Huayuan County, GPS coordinates 28°34′48″ N, 109°28′48″ E, altitude 700 m, ground in mixed forest, 30 July, 2023, Yuan 17361 (IFP 020078, GenBank ITS: PQ656836; LSU: PQ803815; SSU: PV856502; mtSSU: PV856547).
Notes – In the present study (Fig. 6), the new species Thelephora cacao reveals it is clustered with T. bubalinomarginata and T. petaloides. In morphology, T. cacao resembles T. bubalinomarginata in having single to concrescent basidiomata, and strong fenugreek odor. However, T. bubalinomarginata can be delimited from T. cacao by its circular to irregular pilei, cylindrical stipes, and cylindrical basidiospores[13]. T. cacao shares similarities with T. petaloides in having flabelliform to applanate-lobate branches, sulcate or irregularly ridged abhymenial face, and irregularly cylindrical to flatted stipes[97]. However, T. petaloides can be distinguished from T. cacao by its imbricate to umbrella basidiomata, subglobose to subellipsoid basidiospores, and higher basidiomata[97].
Thelephora chayuensis Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 42,43
Fungal Names number: FN 572453

Figure 42.
Basidiomata of Thelephora chayuensis (holotype IFP 020064).
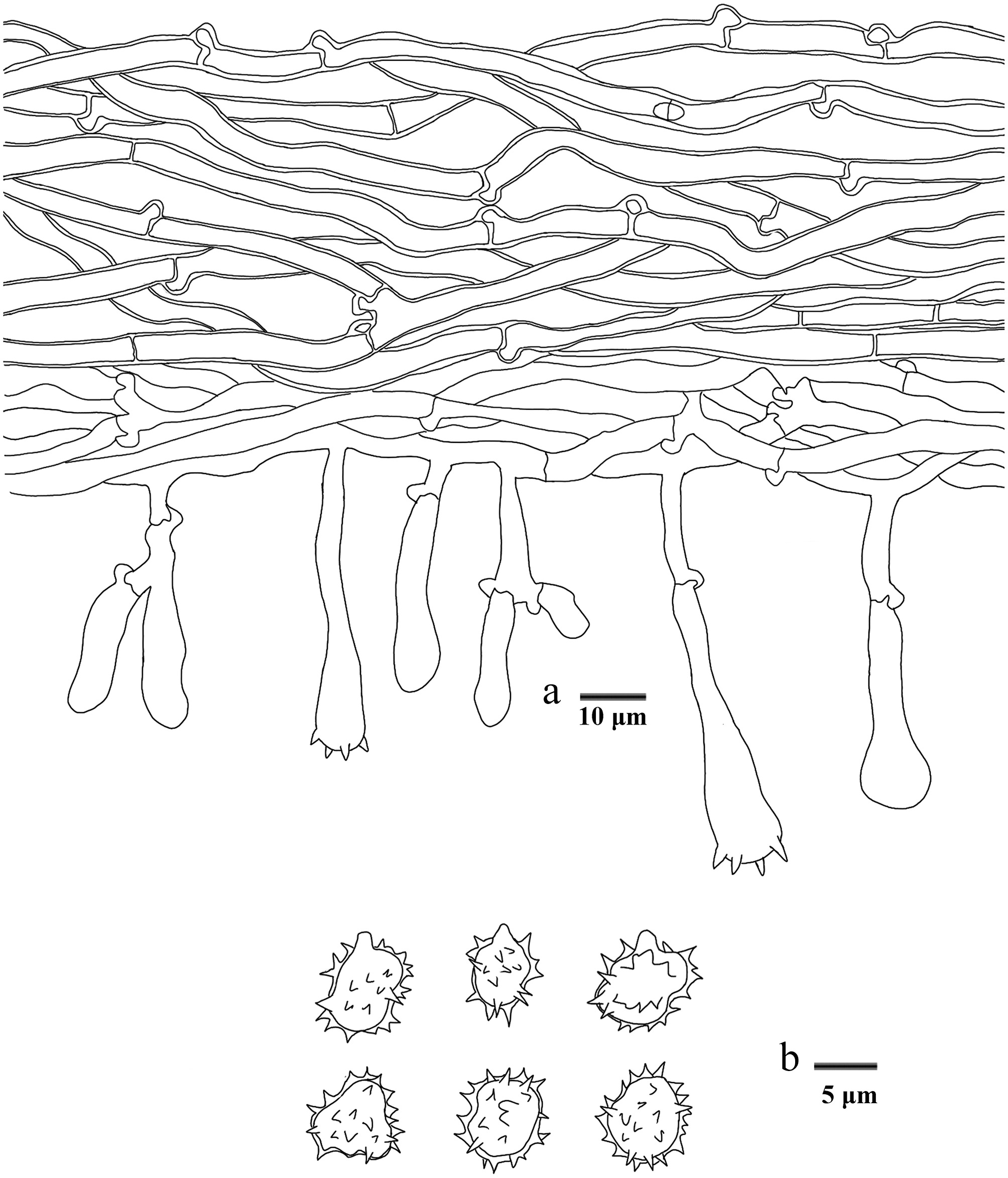
Figure 43.
Microscopic structures of Thelephora chayuensis (drawn from the holotype IFP 020064). (a) Section through basidiomata. (b) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora chayuensis is characterized by the brown to dark brown hymenophoral surface, stalked basidia, and echinulate basidiospores (4.9–)5.6–7.9(–8.6) × (4.8–)5–6.8(–6.9) µm.
Etymology – Chayuensis (Lat.): referring to the locality (Chayu County) of the type.
Type – China, Xizang Autonomous Region, Linzhi City, Chayu County, Rangshequ 3rd Middle Bridge, GPS coordinates 28°35′23″ N, 98°20′36″ E, altitude 3,550 m, on fallen angiosperm branch, 13 July, 2019, Yuan 13502 (holotype: IFP 020064, GenBank ITS: PQ656831; LSU: PV257906).
Description – Basidiomata annual, resupinate, separable from the substrate, mucedinoid, without odor or taste when fresh, 0.4–0.8 mm thick, continuous. Hymenophoral surface smooth, brown to dark brown (6E7–6F7), and concolorous with the subiculum. Sterile margin often indeterminate, byssoid, concolorous with hymenophore.
Rhizomorphs – Absent.
Subiculum – Hyphal system monomitic, generative hyphae with clamp connections, and rarely simple-septa, thick-walled, 3–5 µm diameter, without encrustation, pale brown in KOH, and distilled water, CB+, IKI–.
Subhymenium – Generative hyphae with clamp connections, thin-walled, 3–4 µm diameter, without encrustation, hyphal cells more or less uniform, pale brown in KOH, and in distilled water, CB–, IKI–.
Basidia – 20–50 µm long and 5–9 µm diameter at the apex, 2–4 µm at base, with a clamp connection at the base, clavate, stalked, without transverse septa, pale brown in KOH, and distilled water, four sterigmata, sterigmata 2–3.5 µm long, and 1–2.5 µm diameter at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subtriangular to ellipsoid (irregularly lobed) in frontal (half also ellipsoid), and ovoid to ellipsoid (irregularly lobed) in lateral face, pale brown in KOH, and distilled water, thick-walled, echinulate, echinulati usually isolated, sometimes grouped in two or more, up to 1 mm long, (4.9–)5.6–7.9(–8.6) × (4.8–)5–6.8(–6.9) µm, L = 6.48 µm, W = 6.1 µm, Q = 1.05–1.15 (n = 60/2), CB+, IKI–.
Material examined (paratypes) – China, Xizang Autonomous Region, Linzhi City, Bomi County, GPS coordinates 29°52′25″ N, 95°45′5″ E, altitude 2,730 m, on fallen branch of Pinus sp., 19 July, 2019, Yuan 13716 (IFP 020065, GenBank ITS: PQ656832; LSU: PV257907); Yunnan Province, Kunming City, Xishan District, Xishan Scenic Area, GPS coordinates 24°58′ N, 102°38′ E, altitude 1,900 m, on fallen angiosperm branch, 14 August, 2019, Yuan 13990 (IFP 020066, GenBank ITS: PQ656833); Chuxiong Autonomous Prefecture, Wuding County, Shizi Mountain, GPS coordinates 25°32′11″ N, 102°22′24″ E, altitude 1,800 m, on fallen angiosperm branch, 27 September, 2019, Yuan 14494 (IFP 020067, GenBank ITS: PQ656834).
Notes – The new species Thelephora chayuensis is grouped with the Thelephora species (Fig. 6), and is revealed as a sister to T. linzhiensis and T. liaoningensis. T. chayuensis resembles T. liaoningensis in having mucedinoid basidiomata, smooth hymenophoral surface, the absences of rhizomorphs, and the size of basidia. However, T. chayuensis differs from T. liaoningensis due to its narrower basidiospres, and dark brown to chestnut hymenophoral surface[102]. T. chayuensis is similar to T. coffeae and T. fuscofarinosa in having dark brown hymenophoral surface[102]. However, T. coffeae differs from T. chayuensis by its adherent basidiomata, and larger basidiospores ([6.5–]7–9[–9.5] × [6.5–]6.8–8.3[–8.5] μm)[102]. T. fuscofarinosa differentiates from T. chayuensis in its adherent basidiomata, and wider subhymenophoral hyphae (5–11 μm)[102].
Thelephora fasciculata Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 44,45
Fungal Names number: FN 572447

Figure 44.
Basidiomata of Thelephora fasciculata (holotype IFP 020052).
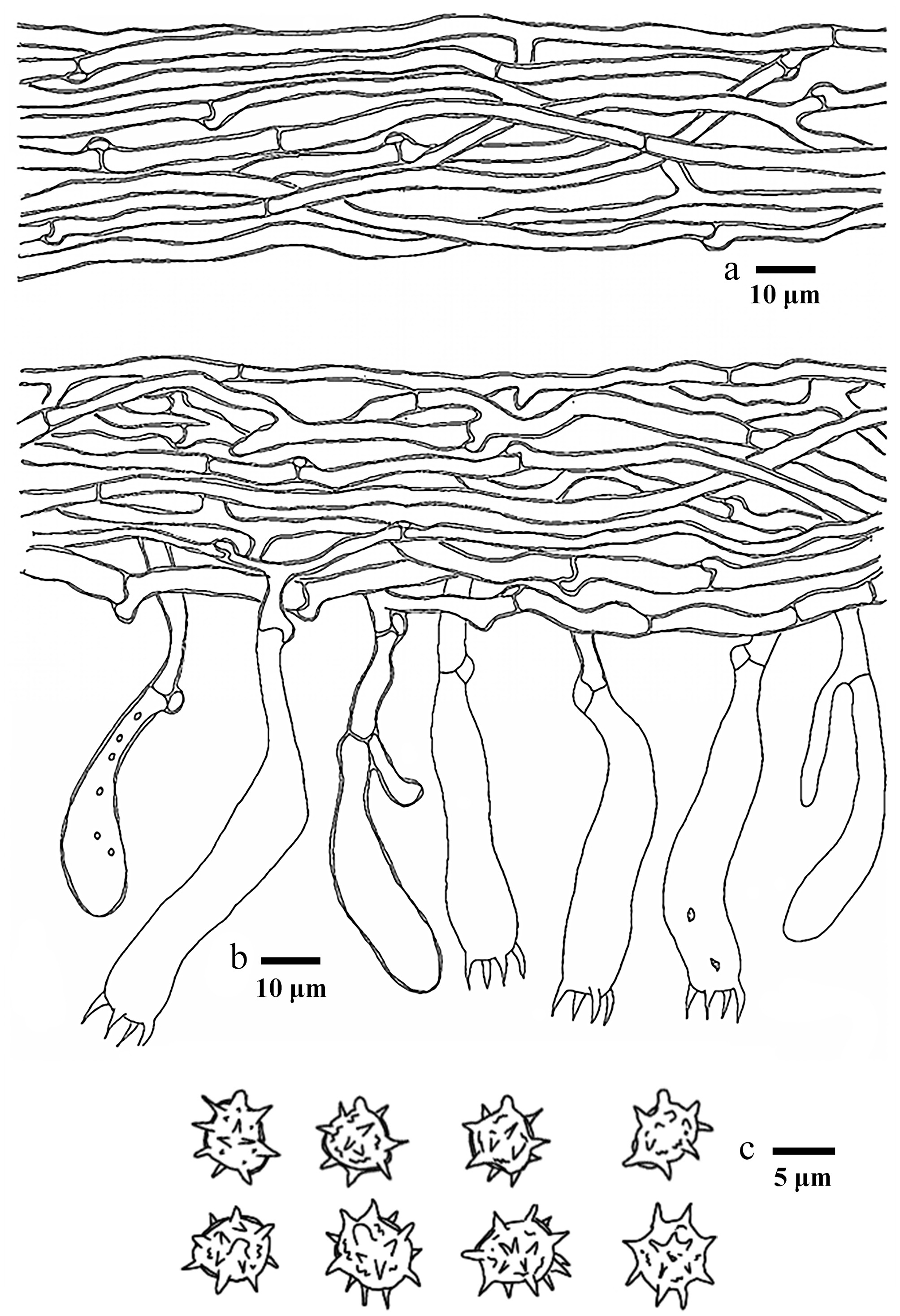
Figure 45.
Microscopic structures of Thelephora fasciculata (drawn from the holotype IFP 020052). (a) Hyphae from pileal context. (b) Section of hymenium and subhymenium. (c) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora fasciculata is characterized by the infundibuliform to ligulate pileus, dark violet basidiomata, white pileal margin, and clavillose to flatted or broadened stipe.
Etymology – Fasciculata (Lat.): referring to the fasciculate basidiomata.
Type – China, Yunnan Province, Shizong County, Junzishan National Forest Park, GPS coordinates 24°64′ N, 104°16′ E, altitude 2,409 m, ground in broad-leaved forest, 8 August, 2019, Yuan 13893 (holotype: IFP 020052, GenBank ITS: PQ656842; LSU: PQ803811; mtSSU: PV440375).
Description – Basidiomata upright, solitary, gregarious to caespitose, small-sized, up to 2.5 cm high, and 2 cm wide, coriaceous when fresh, taste mild, almost no odor when dried. Pileus infundibuliform to ligulate, imbricate, usually with multiple pilei arising from a central stipe, margin slightly thin (0.1 mm), irregularly lobed to wavy. Abhymenial surface somewhat wrinkled, zonate, dark violet (18F5) near the center then violet grey (18B2) gradually towards margin, violet white (18A2) at the margin. Hymenophoral surface dark violet (18F5) gradually becoming deep violet to violet white (18E8–18A2) towards the margin. Stipe short and central, up to 0.5 cm long, dark violet (18F5), glabrous, clavillose to flatted or broadened.
Hyphal structure – Hyphal system monomitic, generative hyphae commonly with clamp connections and rarely simple-septa, CB+, IKI–, tissues turned black in KOH.
Pileus – Generative hyphae slightly thick-walled (< 1 μm), hyaline, moderately branched, sometimes flexuous and collapsed, subparallel to loosely interwoven, 3–5 μm diam.
Subhymenium – Generative hyphae slightly thick-walled (< 1 μm), hyaline, frequently branched often near the clamp connections, slightly inflated, loosely interwoven, up to 3–7 μm diam.
Basidia – Clavate to subcylindrical, thin- to slightly thick-walled (< 1 μm), with a clamp connection at the base, multi-guttulate content sometimes very dense, 40–80 × 10–12 μm, four sterigmata, sterigmata 7–11 μm long, and 2–3 μm diam. at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores globose to elliptic in frontal and lateral face, yellowish brown to pale green in KOH and in distilled water, slightly thick-walled (< 1 μm), echinulate to aculeate, echinuli or aculei usually isolated, sometimes in groups of two or more, up to 1.5 μm long, (7.5–)8–9.5(–10) × (5.5–)6–7(–8) μm, L = 8.66 μm, W = 6.85 μm, Q = 1.23–1.26 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Fujian Province, Sanming City, Yongan City, GPS coordinates 25°56′ N, 117°21′ E, ground in broad-leaved forest, 2 July, 2024, Yuan 19490 (IFP 020053, GenBank ITS: PQ656843; LSU: PQ803812; SSU: PV257942; mtSSU: PV440376).
Notes – In this study, the new species Thelephora fasciculata is classified within Thelephora (Fig. 6), and is clustered with T. palmata, T. regularis and T. yunnanensis. They have common characteristics features: pileus clustered pseudoclavate or cuneate branch arising from a central common base, absence of cystidia and basidiospores with echinulate ornamentation[99,152]. However, T. palmata can be distinguished from T. fasciculata by its deep purple to grayish-black pileus surface, larger basidia (70–100 × 9–12 µm), and purplish basidiospores[152]. T. fasciculata can be distinguished from T. regularis by black to purplish-red pileus surface, hymenophoral surface with 1–1.5 mm white aculeolus, and larger basidiospores (7.5–10 × 5.5–8 µm in T. fasciculata vs 6–8 × 4.5–6.5 μm in T. regularis)[152].
Thelephora fasciculata shares similarly morphological characteristics with the tropical species T. versatilis, such as the pileus clustered pseudocoralloid branches, subhymenophoral hyphae frequent branching, and the basidiospores with echinulate ornamentation. However, T. versatilis can be distinguished from T. fasciculata by dark brown to yellowish-brown pileus surface, and smaller basidiospores (6–7 × 5–6 µm in T. versatilis vs 7.5–10 × 5.5–8 μm in T. fasciculata)[148].
Thelephora latihypha Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 46,47
Fungal Names number: FN 572910

Figure 46.
Basidiomata of Thelephora latihypha (holotype IFP 020079).
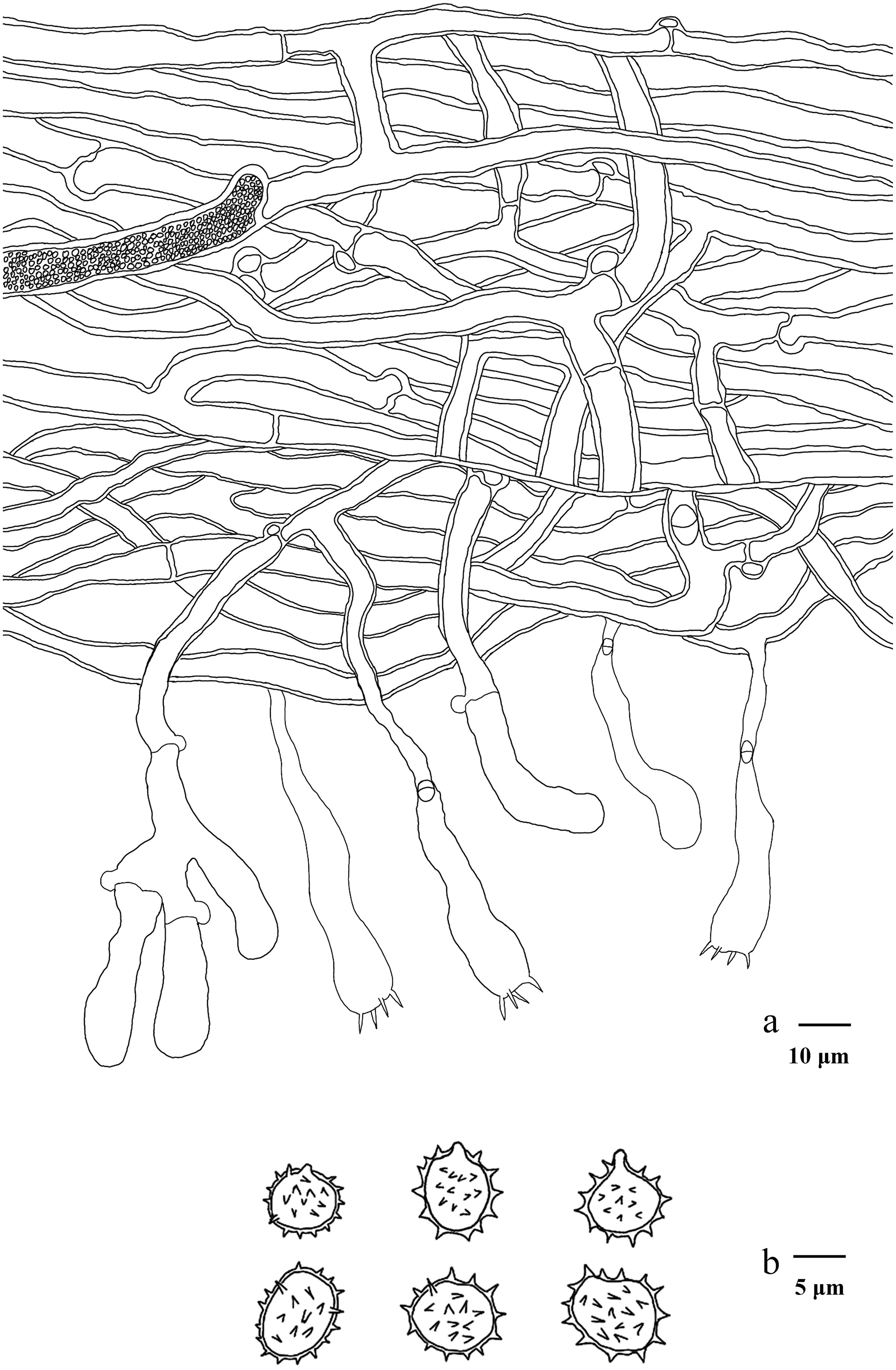
Figure 47.
Microscopic structures of Thelephora latihypha (drawn from the holotype IFP 020079). (a) Section through basidiomata. (b) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora latihypha is characterized by the grey hymenophoral surface, broad subicular hyphae (5–8.5 µm), and subglobose to ellipsoid basidiospores (6.9–)7.1–9.2(–10) × (6.3–)6.6–8.2(–8.5) µm.
Etymology – Latihypha (Lat.): referring to the broad hyphae.
Type – China, Yunnan Province, Diqing Tibetan Autonomous Prefecture, Shangri-La City, Potatso National Park, GPS coordinates 27°50′12″ N, 99°57′27″ E, altitude 3,655 m, fallen branch of Abies sp., 15 August, 2023, Yuan 18172 (holotype: IFP 020079, GenBank ITS: PV843167).
Description – Basidiomata annual, resupinate, adherent to the substrate, mucedinoid, without odor or taste when fresh, 1–2 mm thick, continuous. Hymenophoral surface smooth, grey (B1–D1), and concolorous with hymenophore. Sterile margin often indeterminate, byssoid, paler than hymenophore.
Rhizomorphs – Absent.
Subiculum – Hyphal system monomitic, generative hyphae with clamp connections and simple-septa, thick-walled, frequently branched, 5–8.5 µm diameter, with encrustation, pale brown in KOH and distilled water, CB+, IKI–.
Subhymenium – Generative hyphae with clamp connections, and simple-septa, thick-walled, frequently branched, 3–5 µm diameter, without encrustation, hyphal cells more or less uniform, pale brown in KOH, and in distilled water, CB+, IKI–.
Basidia – 30–50 µm long, and 8.5–10 µm diameter at apex, 5–7 µm at base, with a clamp connection at the base, utriform, stalked, sinuous, without transverse septa, pale brown in KOH, and distilled water, four sterigmata, sterigmata 4–7 µm long, and 1–1.5 µm in diameter at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospore ellipsoid in frontal face, and subglobose to ellipsoid in lateral face, pale brown in KOH, and distilled water, thick-walled, echinulate, echinuli usually isolated, usually 1 µm long, sometimes up to 1.5 µm long, (6.9–)7.1–9.2(–10) × (6.3–)6.6–8.2(–8.5) µm, L = 8.1 µm, W = 7.3 µm, Q = 1.02–1.3 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Yunnan Province, Diqing Tibetan Autonomous Prefecture, Shangri-La City, Potatso National Park, GPS coordinates 27°50′12″ N, 99°57′27″ E, altitude 3,655 m, fallen trunk of Abies sp., 13 August, 2023, Yuan 18228 (IFP 020080, GenBank ITS: PV843168; LSU: PV843173; SSU: PV856503).
Notes – In this study (Fig. 6), the new species Thelephora latihypha is grouped within Thelephora, and is closely associated with T. lapida. T. latihypha resembles T. lapida by sharing continuous arachnoid basidiomata, smooth hymenophoral surface, and the shape of basidia[43]. However, T. lapida differs from T. latihypha by its tomentose basidiomata, rusty brown to fuscous hymenophoral surface, and globose basidiospores[153]. T. latihypha is similar to T. cinereobrunnea and T. longiechinula in sharing grayish hymenophoral surface[37]. However, T. cinereobrunnea differs from T. latihypha by its separable basidiomata, narrower subicular hyphae (4–7 mm), and shorter basidia[37]. T. longiechinula differentiates from T. latihypha in its separable basidiomata, and wider subhymenophoral hyphae (4–7 mm)[37].
Thelephora linzhiensis Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 48,49
Fungal Names number: FN 572450

Figure 48.
Basidiomata of Thelephora linzhiensis (holotype IFP 020058).
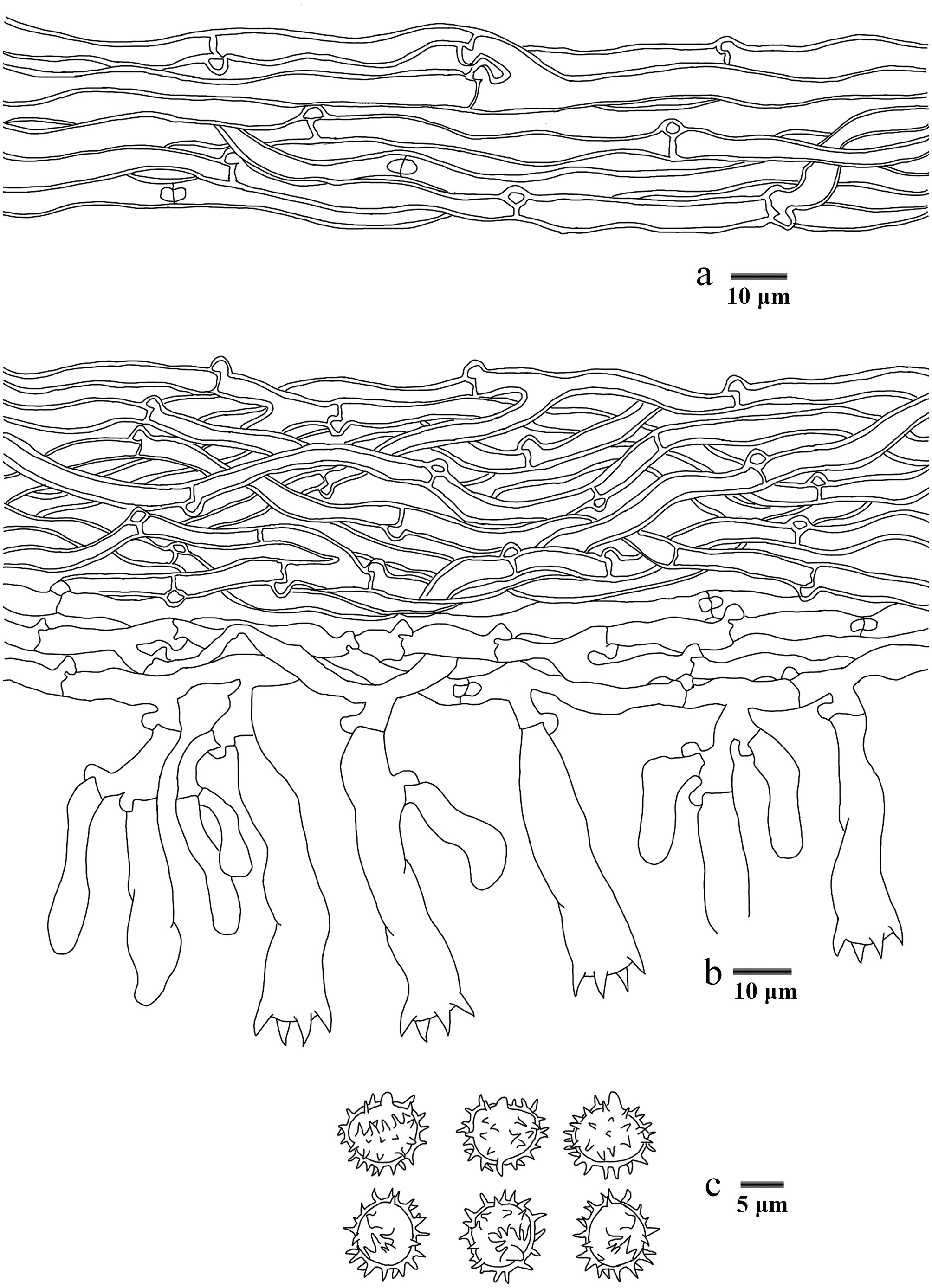
Figure 49.
Microscopic structures of Thelephora linzhiensis (drawn from the holotype IFP 020058). (a) Hyphae from a rhizomorph. (b) Section through basidiomata. (c) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora linzhiensis is characterized by the type B rhizomorphs, brownish yellow to golden brown hymenophoral surface, and utriform basidia.
Etymology – Linzhiensis (Lat.): referring to the locality (Linzhi City) of the type.
Type – China, Xizang Autonomous Region, Linzhi City, Milin County, Nanyigou Nature Reserve, GPS coordinates 29°7′41″ N, 94°12′52″ E, altitude 2,500 m, on fallen angiosperm branch, 22 October, 2021, Yuan 16203 (holotype: IFP 020058, GenBank ITS: PQ656815; LSU: PV257904).
Description – Basidiomata annual, resupinate, adherent to the substrate, arachnoid, without odor or taste when fresh, 0.1 mm thick, uncontinuous. Hymenophoral surface granulose or smooth, brownish yellow to golden brown (5C7–5D7), and concolorous with the subiculum. Sterile margin often indeterminate, byssoid, concolorous with hymenophore.
Rhizomorphs – Present in subiculum and margins, 10–55 µm diameter, rhizomorphic surface more or less smooth, hyphae in rhizomorph monomitic, undifferentiated, of type B (according to Agerer, 1987–2008), compactly arranged and uniform, single hyphae with clamp connections, thick-walled, occasionally branched, 4–7 µm diameter, pale brown to brown in KOH, CB+, IKI–.
Subiculum – Hyphal system monomitic, generative hyphae with clamp connections and rarely simple-septa, slightly thick to thick-walled, occasionally branched, 4–6 µm diameter, without encrustation, pale brown in KOH, and distilled water, CB+, IKI–.
Subhymenium – Generative hyphae with clamp connections, thin-walled, occasionally branched, 2.5–5 µm diameter, without encrustation, hyphal cells short and inflated, pale brown in KOH, and in distilled water, CB–, IKI–.
Basidia – 20–55 µm long, and 5–10 µm diameter at apex, 3–5 µm at base, with a clamp connection at the base, utriform, not stalked, sinuous, without transverse septa, pale brown in KOH, and distilled water, four sterigmata, sterigmata 4–5 µm long, and 2–3 µm diameter at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subglobose to globose in frontal and lateral face, pale brown in KOH, and distilled water, slightly thick-walled, echinulate to aculeate, echinuli or aculei usually isolated, up to 2 µm long, (6.8–)8.2–10.3(–11) × (6.2–)7.5–10(–10.8) µm, L = 9.44 µm, W = 8.63 µm, Q = 1–1.36 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Xizang Autonomous Region, Linzhi City, Milin County, Nanyigou Nature Reserve, GPS coordinates 29°7′41″ N, 94°12′52″ E, altitude 2,500 m, on fallen angiosperm branch, 22 October, 2021, Yuan 16200 (IFP 020059, GenBank ITS: PQ656814; LSU: PV257903; SSU: PV440976).
Notes – Based on the phylogenetic tree (Fig. 6), the new species Thelephora linzhiensis is grouped with Thelephora species, and is revealed as a sister to T. chayuensis and T. liaoningensis. T. linzhiensis resembles T. chayuensis in having indeterminate and byssoid sterile margin, and basidiopoers with aculeate. However, T. linzhiensis differs from T. chayuensis due to its arachnoid basidiomata, granulose hymenophoral surface, and presence of the rhizomorphs. T. linzhiensis is similar to T. aurantispora and T. kanasensis in having yellowish brown hymenophoral surface[154]. However, T. aurantispora differs from T. linzhiensis by its wider subhymenophoral hyphae (3–7 µm), and narrower basidiospores ([6.5–]7–9[–9.5] × [5.5–]6–8[–8.5] µm)[154]. T. kanasensis differentiates from T. linzhiensis in its wider subhymenophoral hyphae (4–7 µm), and basidia (8–11 µm)[154].
Thelephora longicystidiata Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 50,51
Fungal Names number: FN 572911

Figure 50.
Basidiomata of Thelephora longicystidiata (holotype IFP 020081).
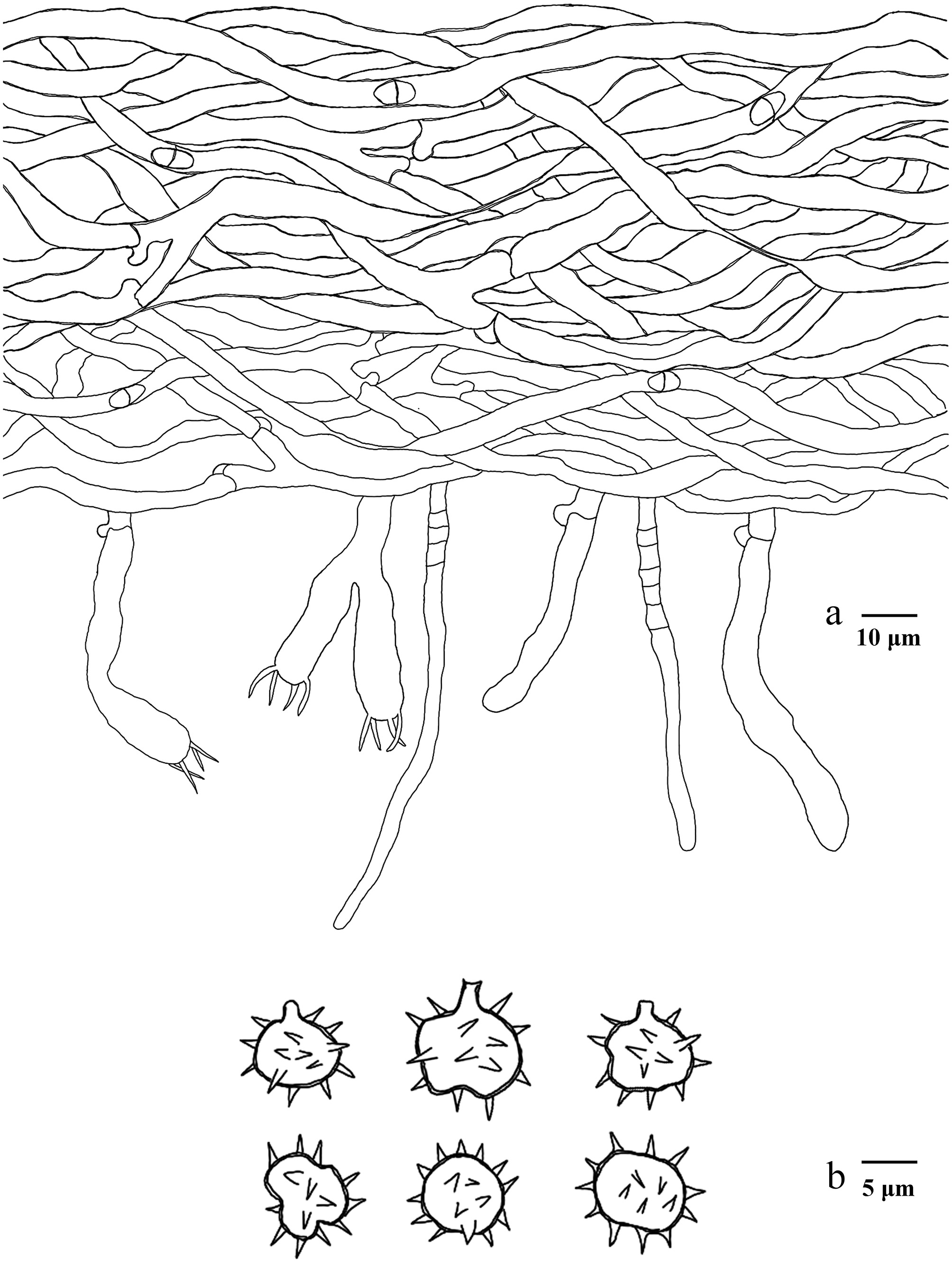
Figure 51.
Microscopic structures of Thelephora longicystidiata (drawn from the holotype IFP 020081). (a) Section through basidiomata. (b) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora longicystidiata is characterized by the brownish orange to orange grey hymenophoral surface, clavate cystidia, and subtriangular, ellipsoid to globose (irregularly lobed) basidiospores.
Etymology – Longicystidiata (Lat.): referring to the long cystidia of the type.
Type – China, Yunnan Province, Yuxi City, Xinping Autonomous County, Gasa Town, GPS coordinates 21°57′11″ N, 100°45′42″ E, altitude 2,300 m, on angiosperm trunk, 6 August, 2023, Yuan 17428 (holotype: IFP 020081, GenBank ITS: PV843170).
Description – Basidiomata annual, resupinate, adherent to the substrate, mucedinoid, without odor or taste when fresh, 0.1–1 mm thick, continuous. Hymenophoral surface smooth, brownish orange to orange grey (5C3–5B2), and turning darker than subiculum. Sterile margin often indeterminate, farinaceous, paler than hymenophore.
Rhizomorphs – Absent.
Subiculum – Hyphal system monomitic, generative hyphae with clamp connections, slightly thick-walled, branched, 4–6 µm diameter, occasionally collapsed, without encrustation, pale brown in KOH, and distilled water, CB+, IKI–.
Subhymenium – Generative hyphae with clamp connections, thin-walled, occasionally branched, 2.5–4 µm diameter, occasionally collapsed, without encrustation, hyphal cells more or less uniform, pale brown in KOH, and in distilled water, CB–, IKI–.
Basidia – 25–55 µm long, and 7–9 µm diameter at apex, 5–7 µm at base, with a clamp connection at the base, clavate, not stalked, sinuous, without transverse septa, pale brown in KOH, and distilled water, four sterigmata, sterigmata 5–7 µm long, and 1–1.5 µm diameter at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Clavate, arising from subhymenophoral hyphae, thin-walled, 55–95 µm long, 3–4 µm at the base, and 3–6 µm at the apex, projecting outside of the hymenium, septate, without encrustation, pale brown in KOH, and distilled water.
Spores – Basidiospores subtriangular to ellipsoid (irregularly lobed) (half also globose to oblong-ellipsoid) in frontal, and ellipsoid (irregularly lobed) in lateral face, pale brown in KOH, and distilled water, slightly thick-walled, aculeate, aculei usually isolated, up to 2 µm long, (4.4–)5.4–8.5(–9.2) × (3.6–)4.5–7.5(–8.1) µm, L = 6.64 µm, W = 5.81 µm, Q = 1.02–1.31 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Yunnan Province, Yuxi City, Xinping Autonomous County, Gasa Town, GPS coordinates 21°57′11″ N, 100°45′42″ E, altitude 2,300 m, dead branch of living tree, 6 August, 2023, Yuan 17424 (IFP 020082, GenBank ITS: PV843169; SSU: PV856510).
Notes – In this study (Fig. 6), the new species Thelephora longicystidiata is grouped within T. albomarginata, T. castanea, and T. sublilacina. They share common characteristics: the adherent basidiomata and smooth hymenophoral surface[43,142,155]. T. longicystidiata is similar to T. albomarginata sharing irregular basidiospores, and clavate basidia. However, T. albomarginata differs from T. longicystidiata by its longer basidia (40–60 µm), larger basidiospores (7–9.5 µm across), and the absences of cystidia[142]. T. longicystidiata resembles T. sublilacina in having sinuous basidia. However, T. sublilacina differs from T. longicystidiata by its echinulate basidiospores, utriform basidia, and wider subhymenophoral hyphae[43].
Thelephora microcarpa Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 52,53
Fungal Names number: FN 572448

Figure 52.
Basidiomata of Thelephora microcarpa (paratype IFP 020055). Photo by Yu-Rong Liang.
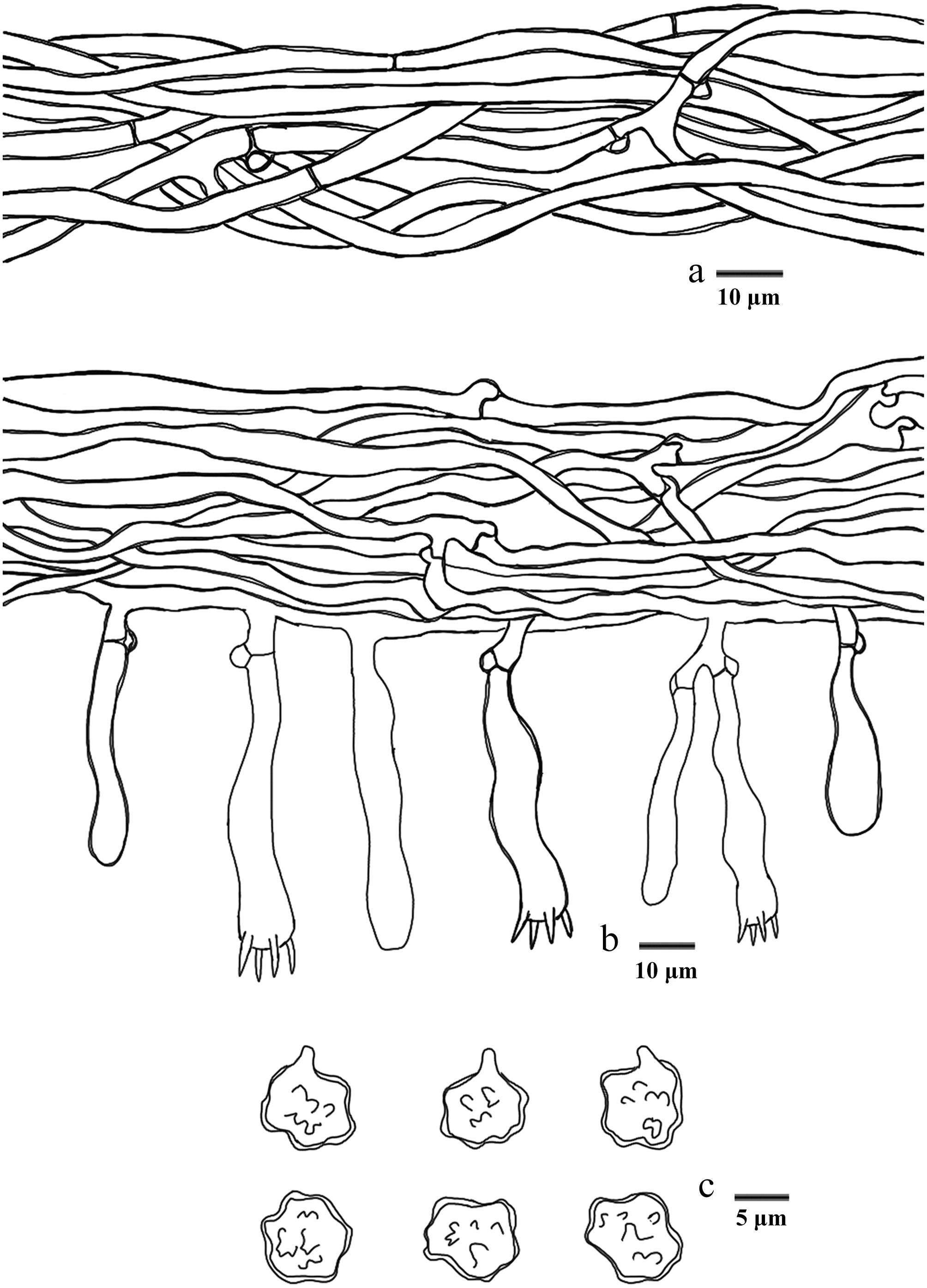
Figure 53.
Microscopic structures of Thelephora microcarpa (drawn from the holotype IFP 020054). (a) Hyphae from pileal context. (b) Section of hymenium and subhymenium. (c) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora microcarpa is characterized by the dark violet basidiomata, clavate to coralloid pileus, white pileal margin, and utriform to subcylindrical basidia.
Etymology – Microcarpa (Lat.): referring to the small basidiomata.
Type – China, Hunan Province, Huitong County, Huitong Ecological Station, GPS coordinates 26°48′ N, 109°30′ E, altitude 500 m, growing in soil under Acacia auriculiformis trees, 1 October, 2021, Yuan 16658 (holotype: IFP 020054, GenBank ITS: PQ656855; LSU: PQ803813; SSU: PV257943; mtSSU: PV440377).
Description – Basidiomata upright, solitary to concrescent, small-sized, up to 1 cm high, and 1.5 cm wide, coriaceous when fresh, taste mild, almost no odor when dried. Pileus clavate to coralliform, coralloid, usually with multiple pilei arising from a central stipe, margin slightly thin (0.1 mm), irregularly lobed to wavy. Abhymenial surface somewhat wrinkled, zonate, dark violet (18F2) near the center then greyish violet (18C4) gradually towards margin, violet white (18A2) to white (A1) at the margin. Hymenophoral surface concolorous or deeper. Stipe short and central, up to 0.5 cm long, brown, glabrous, clavillose to flatted or broadened.
Hyphal structure – Hyphal system monomitic, generative hyphae commonly clamp connections and simple-septa, CB+, IKI–, tissues turned black in KOH.
Pileus – Generative hyphae slightly thick-walled (< 1 μm), hyaline, moderately branched, sometimes flexuous and collapsed, subparallel to loosely interwoven, 3–6 μm diam.
Subhymenium – Generative hyphae slightly thick-walled (< 1 μm), hyaline, frequently branched often near the clamp connections, distinctly inflated, loosely interwoven, up to 3–8 μm diam.
Basidia – Utriform to subcylindrical, thin- to slightly thick-walled (< 1 μm), with a clamp connection at the base, 45–60 × 9–15 μm, four sterigmata, sterigmata 3–7 μm long, and 1.5–3 μm diam at base, CB+, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores globose to subglobose (irregularly lobed) in frontal view and subglobose to ellipsoid (irregularly lobed) in lateral view, yellowish brown to pale green in KOH, and in distilled water, slightly thick-walled (< 1 μm), tuberculate, tuberculi usually isolated, sometimes in groups of two or more, (5.5–)6–7.5(–8) × (4.2–)5–6.5(–7) μm, L = 6.93 μm, W = 5.81 μm, Q = 1.15–1.19 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Guangdong Province, Guangzhou city, Huangpu District, Meihua Ridge, GPS coordinates 23°11′19″ N, 113°28′5″ E, altitude 212 m, ground in sand under mixed forest, 26 May, 2024, Yuan 19344 (IFP 020055, GenBank ITS: PQ656856; LSU: PQ803814).
Notes – With reference to the phylogenetic tree (Fig. 6), the new species Thelephora microcarpa is grouped into Thelephora, and is clustered with T. pinnatifida. T. microcarpa resembles T. pinnatifida in having clavarioid to coralloid pileus. However, T. pinnatifida can be distinguished from T. microcarpa by its longer basidia (47–90 × 10–11.5 µm), and larger basidiospores ([5.5–]6–9.5 × 5–8.5 µm)[97]. T. microcarpa is similar to T. dactyliophora and T. pinnatifida in having clavarioid to coralloid pileus and white margin[97]. However, T. dactyliophora differs from T. macrocarpa by its brownish gray to gray abhymenial surface, narrower basidia (6–9 μm), and basidiospores (4–6.5 μm)[97]. T. pinnatifida can be distinguished from T. macrocarpa in its brownish orange to brown surface, longer stipe (up to 15 mm), and longer basidia (47–90 μm)[97].
Thelephora nanyigouensis Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 54,55
Fungal Names number: FN 572451

Figure 54.
Basidiomata of Thelephora nanyigouensis (holotype IFP 020060).
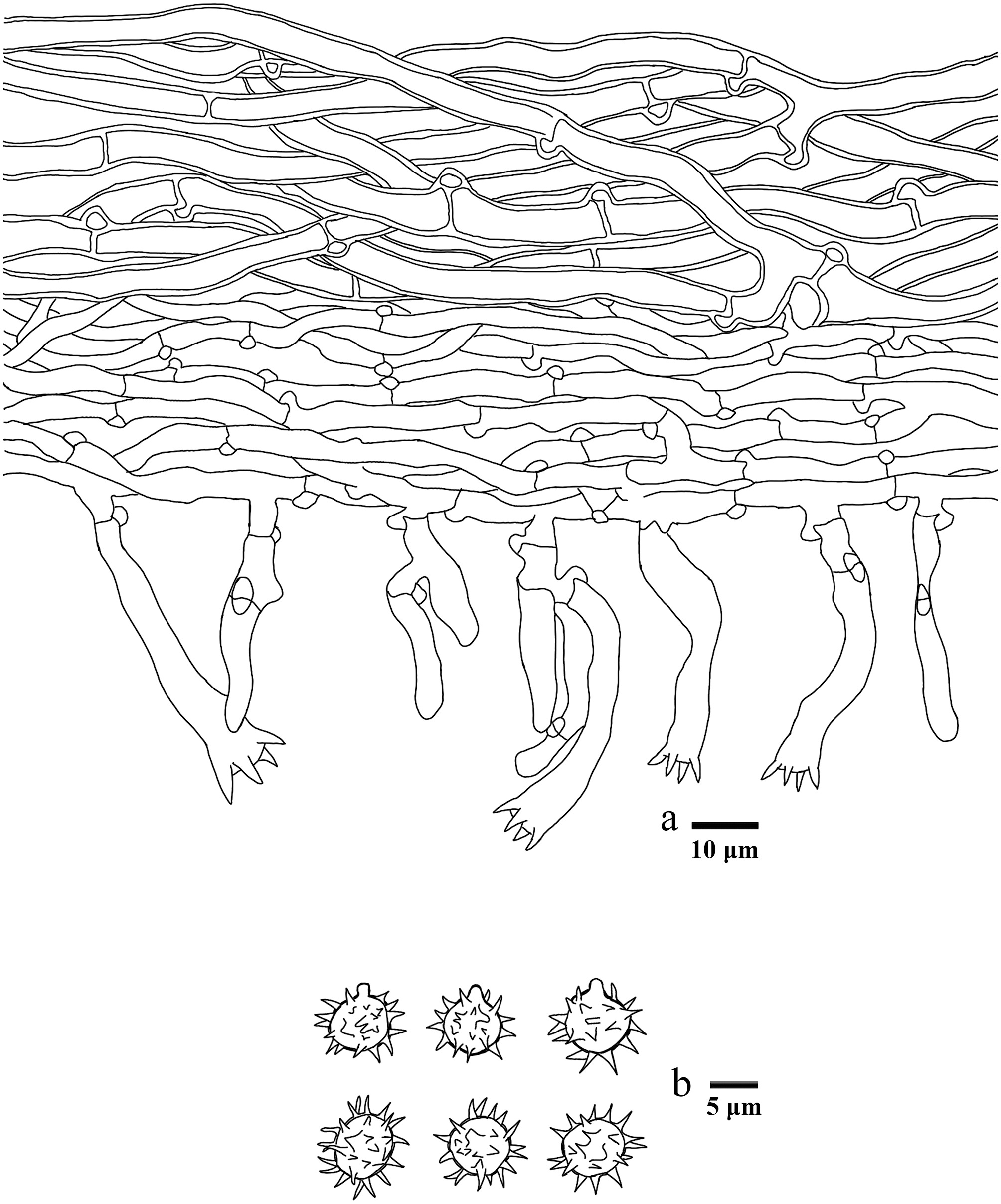
Figure 55.
Microscopic structures of Thelephora nanyigouensis (drawn from the holotype IFP 020060). (a) Section through basidiomata. (b) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora nanyigouensis is characterized by the greyish brown to reddish brown hymenophoral surface, utriform basidia (sometimes with sharp head), and aculeate basidiospores (aculei up to 2.5 µm).
Etymology – Nanyigouensis (Lat.): referring to the locality (Nanyigou Nature Reserve) of the type.
Type – China, Xizang Autonomous Region, Linzhi City, Milin County, Nanyigou Nature Reserve, GPS coordinates 29°7′41″ N, 94°12′52″ E, altitude 2,500 m, on fallen branch of Abies sp., 22 October, 2021, Yuan 16222 (holotype: IFP 020060, GenBank ITS: PQ656813; LSU: PV257902; SSU: PV257946).
Description – Basidiomata annual, resupinate, separable from the substrate, mucedinoid, without odor or taste when fresh, 0.5–1 mm thick, continuous. Hymenophoral surface smooth, greyish brown to reddish brown (9D3–9E4), and concolorous with the subiculum. Sterile margin often determinate, arachnoid, concolorous with hymenophore.
Rhizomorphs – Absent.
Subicular – Hyphal system monomitic, generative hyphae with clamp connecitons and rarely simple-septa, thick-walled, occasionally branched, 5–7 µm diameter, without encrustation, pale brown in KOH, and distilled water, CB+, IKI–.
Subhymenium – Generative hyphae with clamp connecitons and rarely simple-septa, thin-walled, frequently branched, 3–6 µm diameter, without encrustation, hyphal cells short, pale in KOH, and in distilled water, CB–, IKI–.
Basidia – 20–45 µm long, and 4–9 µm diameter at apex, 4.5–5.5 µm at base, with a clamp connection at the base, utriform sometimes with sharp head, not stalked, sinuous, without transverse septa, pale brown in KOH and distilled water, four sterigmata, sterigmata 3–5 µm long, and 1.5–2 µm diameter at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subglobose to globose in frontal and lateral face, pale brown in KOH, and distilled water, slightly thick-walled, aculeate, aculei usually isolated, up to 2.5 µm long, (9.4–)10.1–12(–12.7) × (9.1–)9.6–11.1(–12.1) µm, L = 10.98 µm, W = 10.2 µm, Q = 1.02–1.19 (n = 60/2), CB+, IKI–.
Material examined (paratype) – China, Xizang Autonomous Region, Linzhi City, Milin County, Nanyigou Nature Reserve, GPS coordinates 29°7′41″ N, 94°12′52″ E, altitude 2,500 m, on fallen branch of Picea sp., 22 October, 2021, Yuan 16162 (IFP 020061, GenBank ITS: PQ656812; LSU: PV257901; SSU: PV257945; SSU: PV440379).
Notes – Based on the phylogenetic tree (Fig. 6), the new species Thelephora nanyigouensis is grouped into Thelephora, and is clustered with T. longiechinula, T. cinereoumbrina, and T. badia. Their common features are the grayish brown to dark brown hymenophoral surface, utriform basidia, and the absence of rhizomorphs and cystidia. However, T. nanyigouensis differs from T. badia, T. cinereoumbrina, and T. longiechinula by the clamped hyphae[37,43]. T. badia is differentiated from T. nanyigouensis by its shorter basidiospores (8–11 µm) with shorter echinuli (1–1.5 µm)[43]. T. cinereoumbrina differs from T. nanyigouensis by the crustose basidiomata[37].
Thelephora nigromarginata Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 56,57
Fungal Names number: FN 572912

Figure 56.
Basidiomata of Thelephora nigromarginata (holotype IFP 020083).
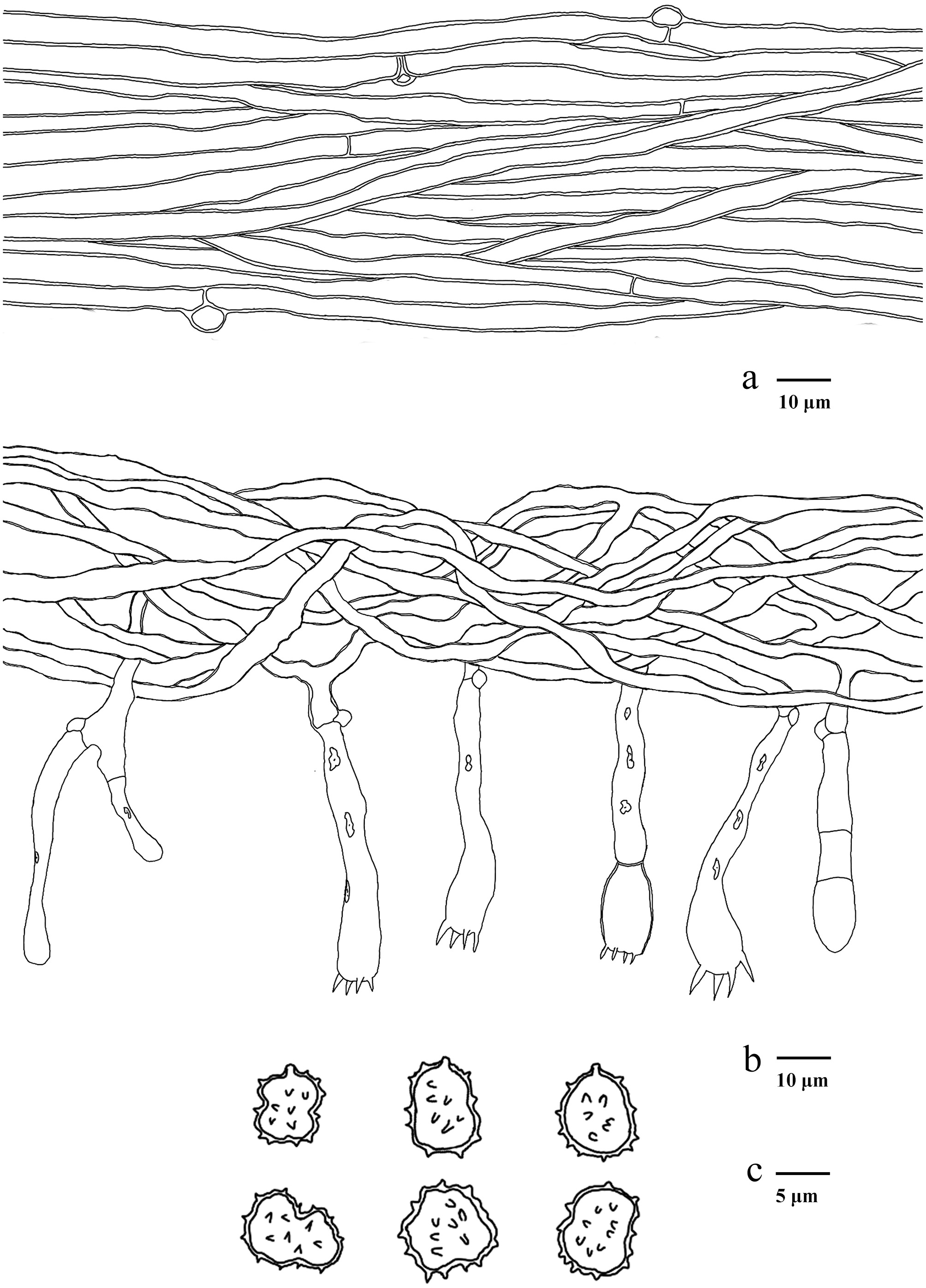
Figure 57.
Microscopic structures of Thelephora nigromarginata (drawn from the holotype IFP 020083). (a) Hyphae from pileal context. (b) Section of hymenium and subhymenium. (c) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora nigromarginata is characterized by the brownish to dark brown basidiomata, white to black pileal margin, rugulose or wrinkled abhymenial surface, and subtriangular to ellipsoid (irregularly lobed) basidiospores.
Etymology – Nigromarginata (Lat.): referring to the black pileus margin.
Type – China, Chongqing City, Jiangjin District, GPS coordinates 29°17′24″ N, 106°15′00″ E, altitude 500 m, ground in bamboo groves, 27 June, 2024, Yuan 19458 (holotype: IFP 020083, GenBank ITS: PQ656847; LSU: PQ803806; SSU: PV856507; mtSSU: PV856549).
Description – Basidiomata upright, solitary to concrescent, small to medium-sized, up to 6 cm high, and 2–3 cm wide, coriaceous when fresh, hard, and light in weight when dried, taste mild, faint odor when dry. Pileus coralloid, usually with multiple branches arising from a central common stipe or base, branch multiple ranks, flat, finger-like to narrow flabelliform, margin thin (0.1–0.2 mm), deeply lacerate. Abhymenial surface slightly rugulose or wrinkled, non-zonate, brownish to dark brown (6E5–6F1), but chalky white (1A1) to black at the margin. Hymenophoral surface concolorous with abhymenial surface, rugulose, and non-zonate. Stipe central, up to 1 cm long, brown to brownish gray (6E5–6F2), irregularly cylindrical to flatted or broadened at base, surface smooth to slightly rugose.
Hyphal structure – Hyphal system monomitic, generative hyphae commonly with clamp connections, and rarely simple-septa, CB+, IKI–, tissues turned brown in KOH.
Pileus – Generative hyphae thick-walled (< 1 μm), hyaline, occasionally branched, subparallel to loosely interwoven, without covered by dense crystals, 3–5 μm diam.
Subhymenium – Generative hyphae slightly thick-walled, hyaline, occasionally branched, loosely interwoven, up to 3–4.5 μm diam.
Basidia – Utriform to clavate, thin- to slightly thick-walled (< 1 μm), with a clamp connection at the base, multi-guttulate content sometimes very dense, 60–80 × 8–12 μm, four sterigmata, sterigmata 4–6 μm long, and 1.5–2.5 μm diam at base, CB–, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores subtriangular to ellipsoid (irregularly lobed) in frontal face, and ellipsoid (irregularly lobed) in lateral face, brown in KOH, and in distilled water, thick-walled (< 1 μm), tuberculate, tuberculi usually isolated, sometimes in groups of two or more, (6.2–)7–8.2(–9) × (4.9–)5.9–7.5(–8.1) μm, L = 7.63 μm, W = 6.63 μm, Q = 1.01–1.5 (n = 60/2), CB+, IKI–.
Material examined (paratypes) – China, Guizhou Province, Zunyi City, Xinpu New District, GPS coordinates 27°43′31″ N, 107°1′36″ E, altitude 900 m, ground in broad-leaved forest, 22 March, 2024, Yuan 19353 (IFP 020084, GenBank ITS: PQ656844; LSU: PQ803804; SSU: PV856508; mtSSU: PV856550); Hunan Province, Shaoyang City, Longhui County, GPS coordinates 27°7′12″ N, 110°58′12″ E, altitude 500 m, ground in Quercus spp. forest, 5 June, 2024, Yuan 19359 (IFP 020085, GenBank ITS: PQ656845; LSU: PQ803805; SSU: PV856509; mtSSU: PV856551).
Notes – In the present study (Fig. 6), the new species Thelephora nigromarginata, together with T. fasciculata, T. macrospora, T. palmata, T. regularis, and T. yunnanensis clustered in a clade with strong support. Morphologically, a special characteristic of T. nigromarginata is the black abhymenial surface margin when mature, which makes it distinct from other species in the genus. Furthermore, T. nigromarginata resembles T. fasciculata in having solitary to concrescent basidiomata, absence of cystidia, same diameter of context hyphae, and tuberculate basidiospores. However, T. fasciculata differs from T. nigromarginata by infundibuliform to ligulate pilei and larger basidiospores. T. nigromarginata and T. macrospora share some common features, including the upright basidiomata, similar size of basidia, and brown pilei surface. However, T. macrospora can be delimited from T. nigromarginata by its echinulate basidiospores and present of cystidia[13].
Sequence alignment analysis indicated that the ITS sequence of this species shared high similarity with the corresponding sequence in the NCBI database, almostly no nucleotide difference identified between them[156,157]. These sequences were obtained from a sample collected in Yunnan Province, China, representing a new distribution site of this species in China.
Thelephora scopiformis Y.Q. Zhu, L.J. Zhou & H.S. Yuan, sp. nov. Figs 58,59
Fungal Names number: FN 572913

Figure 58.
Basidiomata of Thelephora scopiformis (holotype IFP 020086).
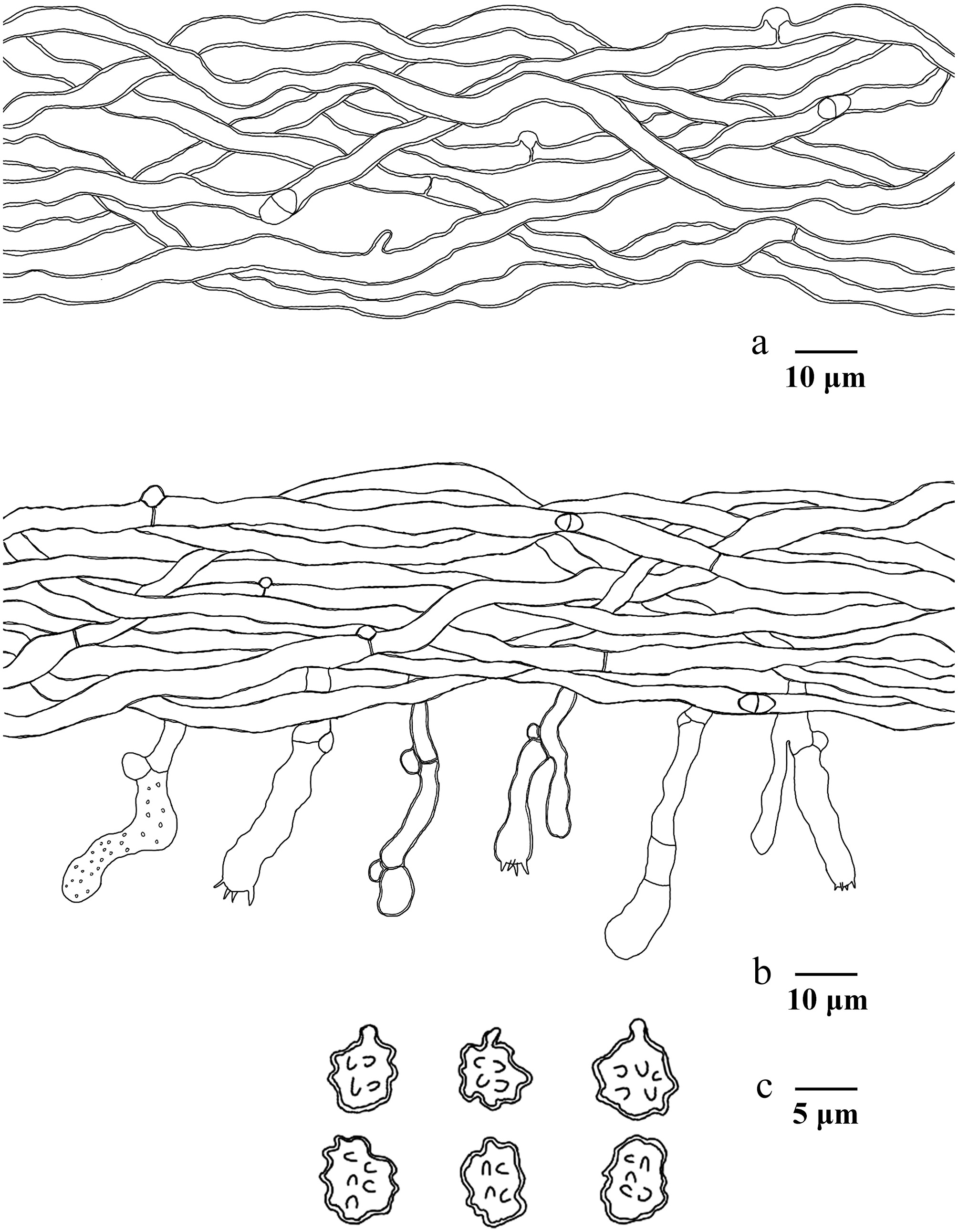
Figure 59.
Microscopic structures of Thelephora scopiformis (drawn from the holotype IFP 020086). (a) Hyphae from pileal context. (b) Section of hymenium and subhymenium. (c) Basidiospores in frontal and lateral view.
Diagnosis – Thelephora scopiformis is characterized by the branch with tips needle-like, chalky white or black pileal margin, utriform to subcylindrical basidia, and pyriform to ellipsoid (irregularly lobed) basidiospores.
Etymology – Scopiformis (Lat.): referring to the scopiform basidiomata.
Type – China, Hunan Province, Shaoyang City, Xinning County, Huilongsi Town, GPS coordinates 26°34′48″ N, 111°4′48″ E, altitude 500 m, ground in mixed forest, 5 June, 2024, Yuan 19364 (holotype: IFP 020086, GenBank ITS: PQ656860; LSU: PQ803801; SSU: PV856504; mtSSU: PV856548).
Description – Basidiomata 35–65 mm high, 15–50 mm wide, gregarious to caespitose, humid, and leathery when fresh, corky to hard and brittle when dry, faint odor when dry. Pileus coralloid, usually with multiple pilei arising from a shared stipe or center, and arranged in coralloid shape, branch clavate to pinnatifid or ramiform with tips needle-like. Abhymenial face sulcate, non-zonate, visibly ribbed, brownish (7E5) to black, but chalky white (1A1) or black at margin, margin 0.1–0.5 mm thick, deeply lacerate, velvety. Hymenophoral surface concolorous with abhymenial surface, rugulose, and non-zonate. Stipe central, 10–15 × 10–20 mm, irregularly cylindrical to flatted or broadened at base, surface smooth to slightly rugose, brownish gray (6F3).
Hyphal structure – Hyphal system monomitic, generative hyphae commonly with clamp connections and rarely simple-septa, CB+, IKI–, tissues turned brown-black in KOH.
Pileus – Generative hyphae thick-walled (< 1 μm), hyaline, rarely branched, 3–5 μm diam.
Subhymenium – Generative hyphae slightly thick-walled (< 1 μm), hyaline, rarely branched, sometimes inflated, loosely interwoven, up to 2–3.5 μm diam.
Basidia – Utriform to subcylindrical, thin- to slightly thick-walled (< 1 μm), with a clamp connection at the base, occasionally covered by dense crystals, occasionally with septate, 15–40 × 4–7 μm, four sterigmata, sterigmata 4–5.5 μm long, and 1–2 μm at base, CB+, IKI–. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores ellipsoid (irregularly lobed) in frontal face, and pyriform to ellipsoid (irregularly lobed) in lateral face, pale brown in KOH, and in distilled water, slightly thick-walled (< 1 μm), tuberculate, tuberculi usually isolated, sometimes in groups of two or more, (5–)5.4–7.6(–7.9) × (4.8–)5–6.7(–7) μm, L = 6.46 μm, W = 5.67 μm, Q = 1.15–1.25 (n = 60/2), CB+, IKI–.
Material examined (paratypes) – China, Chongqing City, Yubei District, GPS coordinates 29°36′23″ N, 106°34′19″ E, altitude 290 m, ground in Pinus spp. forest, 15 June, 2024, Yuan 19430 (IFP 020087, GenBank ITS: PQ656863; LSU: PQ803802; SSU: PV856505); Sichuan Province, Guangan City, Linshui County, Chengbei Town, GPS coordinates 30°21′00″ N, 106°55′48″ E, altitude 400 m, ground in mixed forest, 25 June, 2024, Yuan 19460 (IFP 020088, GenBank ITS: PQ656866; LSU: PQ803803; SSU: PV856506).
Notes – In the present study (Fig. 6), the new species Thelephora scopiformis closely relates to T. anthocephala and T. scissilis. Morphologically, T. scopiformis shares similar features with T. anthocephala by the white abhymenial margin and gregarious basidiomata. Nevertheless, T. anthocephala can be differentiated by its subinfundibuliform branches, ferruginous to fuscous ferruginous or purplish abhymenial face, and smooth hymenophoral surface[145]. T. scopiformis and T. scissilis share common features including the gregarious basidiomata and glabrous stipes. However, the diagnostic feature to distinguish the new species is the smooth hymenophoral surface, castaneous stipes, and larger basidiospores[145].
Hymenochaetales Oberw.
Hymenochaetaceae Donk
Index Fungorum number: IF 536196
Type genus – Hymenochaete Lév.
Hymenochaete Lév.
Index Fungorum number: IF 17816
Type species – Hymenochaete rubiginosa (Dicks.) Lév.
Note – Hymenochaete Lév. was erected in 1846, with Hym. rubiginosa (Dicks.) Lév. designated as the type species. Patouillard[158] systematically organized the Hymenochaete and proposed classification criteria based on the setae, spore morphology and hyphal system. Subsequently, Burt[159], Cunningham[160], Escobar[161], and Léger[162] subdivided the Hymenochaete into four groups or sections—Hymenochaete, Fultochaete, Gymnochaete, and Paragymnochaete—based on the presence or absence of different layers in the context of the basidiomata. Parmasto[163] conducted a cladistic analysis of all Hymenochaete species using macro-morphological and micro-morphological characters. In the comprehensive survey of Hymenochaetales, Larsson et al.[164] provided support for a monophyletic Hymenochaetaceae clade. The family Hymenochaetaceae comprises poroid and non-poroid morphological groups. In recent years, scholars have systematically compiled and taxonomically revised the global diversity of these two groups for the first time[31,165]. In the present study, based on the ITS and nLSU data (Fig. 7), the genus Hymenochaete nested in Hymenochaetaceae, in which one new species Hymenochaete baishanzuensis is reported.
Hymenochaete baishanzuensis L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 60,61
Fungal Names number: FN 572424

Figure 60.
Basidiomata of Hymenochaete baishanzuensis (holotype IFP 020012).
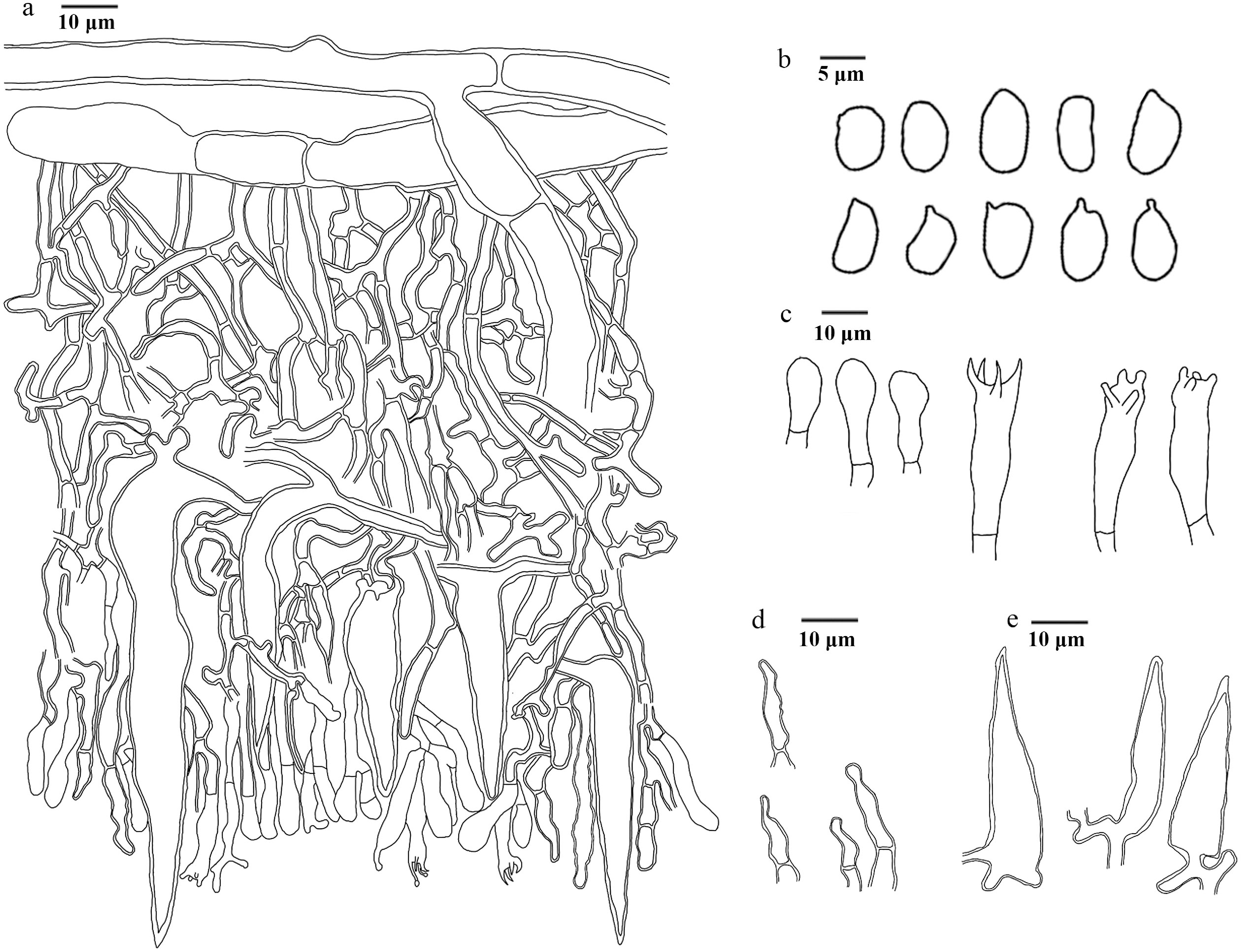
Figure 61.
Microscopic structures of Hymenochaete baishanzuensis (drawn from the holotype IFP 020012). (a) Section of hymenium. (b) Basidiospores. (c) Basidia and basidioles. (d) Hyphidia. (e) Setae.
Diagnosis – Hymenochaete baishanzuensis is characterized by the smooth to tuberculate hymenophoral surface, hyphidia with an obtuse apex and constricted, reniform to ellipsoid spores.
Etymology – Baishanzuensis (Lat.): referring to the locality of the type.
Type – China, Zhejiang Province, Lishui City, Baishanzu National Nature Reserve, GPS coordinates 27°45′11″ N, 119°11′20″ E, altitude 1,515 m, on fallen branch of angiosperm, 18 July, 2023, Wei 11406 (holotype: IFP 020012, GenBank ITS: PV883104).
Description – Basidiomata annual, resupinate, adnate, uneasily separable, coriaceous and corky when fresh, hard brittle when dry, without odor, up to 1 mm thick, hymenophoral surface smooth to tuberculate, yellowish brown (5D4–5F8) to brown (6D4–7F8) when fresh, no significant change after drying. Sterile margin indistinct.
Hyphal structure – Hyphal system monomitic, generative hyphae simple-septa, thin- to thick-walled, CB+, IKI–, tissues unchanged in KOH.
Subiculum – Generative hyphae thick-walled, smooth, dark yellow, moderately branched, parallel interwoven, 7–13 µm in diam, CB+, IKI–.
Subhymenium – Generative hyphae thin- to slightly thick-walled, smooth, dark yellow, frequently branched, more or less flexous, interwoven, 2–4 µm in diam, CB+, IKI–.
Hyphidia – Thick-walled, smooth, colorless, simple-septate, with an obtuse apex and constricted, moderately, 9–20 × 2–3 µm, CB+, IKI–, tissues unchanged in KOH.
Setae – Subulate, thick-walled, smooth, ferruginous, straight, with a sharp apex, frequently, 25–90 × 7–12 µm, arising from the subhymenium, extending through hymenium, CB+, IKI–, tissues unchanged in KOH.
Basidia – Clavate, thin-walled, smooth, colorless, with four sterigmata, and a simple-septa at the base, sterigmata up to 7 µm, slightly constricted at the base, more or less flexous, 15–20 × 3–4 µm, CB–, IKI–, tissues unchanged in KOH. Basidioles similar to basidia.
Cystidia – Absent.
Spores – Basidiospores reniform to ellipsoid, colorless, thin-walled, smooth, 3.5–4 × 2–3 µm, L = 4.0 µm, W = 2.54 µm, Q = 1.33–2 (n = 60/2), CB–, IKI–.
Material examined (paratype) – China, Zhejiang Province, Lishui City, Baishanzu National Nature Reserve, GPS coordinates 27°45′11″ N, 119°11′20″ E, altitude 1,515 m, on fallen branch of angiosperm, 18 July, 2023, Wei 11432 (IFP 020013, GenBank ITS: PV883105).
Notes – The new species Hymenochaete baishanzuensis is classified within Hymenochaete (Fig. 7), and forms a clade with Hym. megaspora. Hym. baishanzuensis resembles Hym. megaspore by sharing smooth to tuberculate hymenophoral surfaces, similar texture of basidiomata and hyphidia width (2–3 µm). However, Hym. megaspora differs from Hym. baishanzuensis by its larger setae ([80–]90–120[–140] × 8–13[–14] µm), basidia (25–32 × 5–8 µm) and baidiospores ([7–]7.5–10[–11] × 5–7 µm)[89]. Hym. baishanzuensis is similar to Hym. bambusicola, and Hym. colliculosa in having brown hymenophore surface[166]. However, Hym. bambusicola differs from Hym. baishanzuensis by its smooth hymenophore surface, dimitic hyphal system, and longer basidiospores (4.5–6 µm)[166]. Hym. colliculosa can be distinguished from Hym. baishanzuensis in its thicker basidiomata (500 µm), larger basidia (20–24 × 5–6 µm), and basidiospores (5–7 × 3.5–5 µm)[166].
Peniophorellaceae L.W. Zhou, X.Wei Wang & S.L. Liu
Index Fungorum number: IF 900252
Type genus – Peniophorella P. Karst.
Peniophorella P. Karst.
Index Fungorum number: IF 18202
Type species – Peniophorella pubera (Fr.) P. Karst.
Note – Peniophorella, typified by Peniophorella pubera, was first described as a monotypic genus by Karsten[167]. With the advancement of molecular systematics, Larsson[168] reinstated the generic independence of Peniophorella from Hyphoderma within the Hymenochaetales. Justo et al.[169] revised the family-level classification of Polyporales (Basidiomycota), and suggested that Peniophorella —accommodating 20 species—should be placed in the family Hyphodermataceae (Hymenochaetales). To date, the number of known species in Peniophorella has increased to approximately 30[170]. However, the familial placement of Peniophorella within Hymenochaetales remains controversial: while some studies have assigned it to Rickenellaceae[28,171], its definitive family-level taxonomic position requires further clarification through more extensive phylogenetic analyses, which extends beyond the scope of the current study. In the present study, Pe. alba is described based on phylogenetic analyses of ITS, nLSU, and nSSU data (Fig. 8).
Peniophorella alba L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 62,63
Fungal Names number: FN 572425

Figure 62.
Basidiomata of Peniophorella alba (holotype IFP 020014).
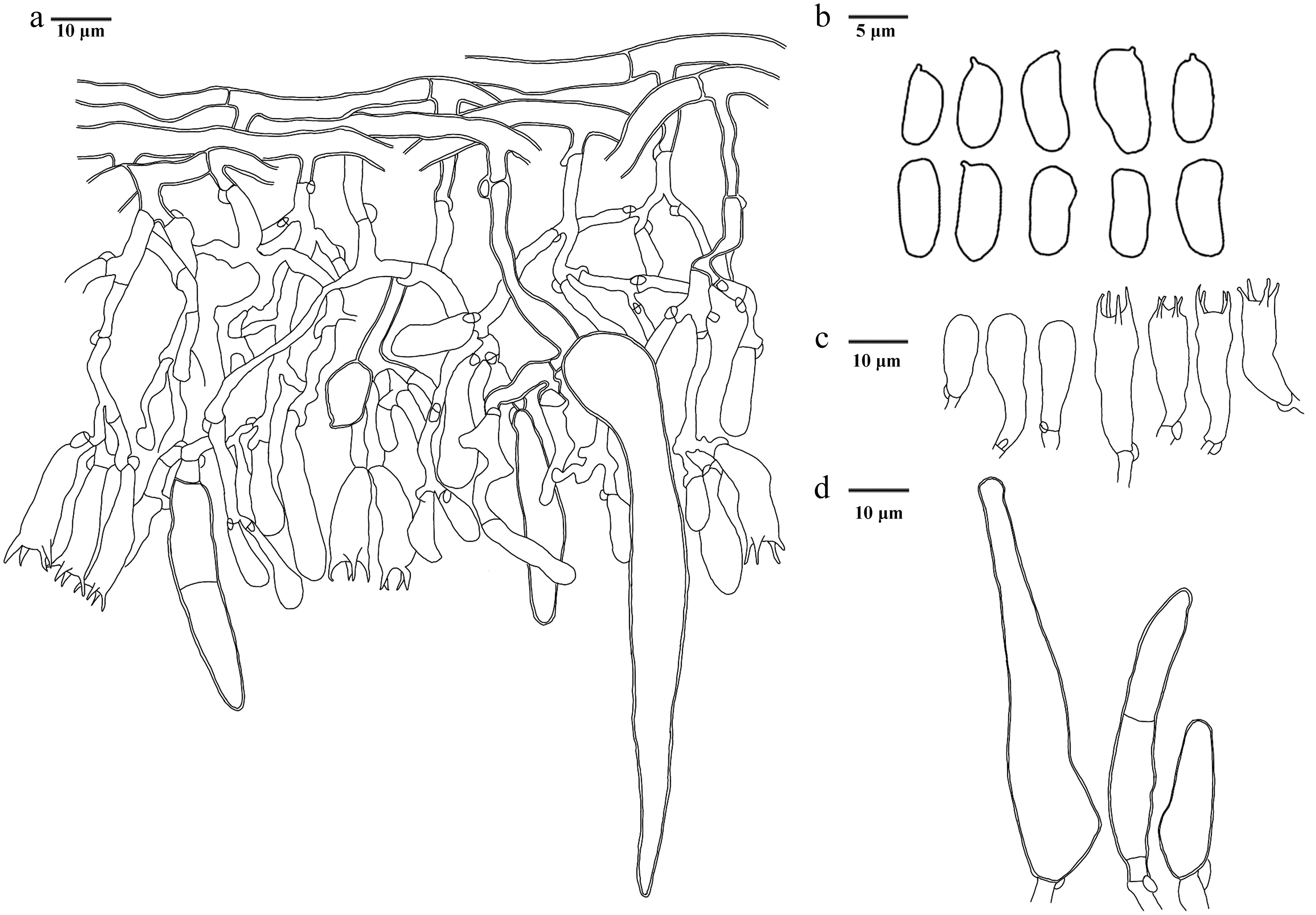
Figure 63.
Microscopic structures of Peniophorella alba (drawn from the holotype IFP 020014). (a) Section of hymenium. (b) Basidiospores. (c) Basidia and basidioles. (d) Cystidia.
Diagnosis – Peniophorella alba is characterized by the smooth hymenophoral surface, vertical subicular hyphae at the branches, cystidia with a sharp apex, and reniform to subcylindrical spores.
Etymology – Alba (Lat.): referring to the white basidiomata.
Type – China, Yunnan Province, Lincang City, Wulaoshan National Forest Park, GPS coordinates 23°54′49″ N, 100°10′55″ E, altitude 2,400 m, on dead branch of angiosperm tree, 9 August, 2023, Yuan 17692 (holotype: IFP 020014, GenBank ITS: PV883103).
Description – Basidiomata annual, resupinate, ceraceous, adnate, uneasily separable, soft leather when fresh, hard brittle when dry, without odor, 50–100 µm thick. Hymenophoral surface smooth, white (–A1) when fresh, later unchanged. Sterile margin indistinct.
Hyphal structure – Hyphal system monomitic, generative hyphae with clamp connections, thin- to thick-walled, CB+, IKI–, tissues unchanged in KOH.
Subiculum – Generative subicular hyphae thick-walled, smooth, colorless, vertical at the branches, frequently, more or less flexous, parallel interwoven, 3–5 µm in diam, CB+, IKI–.
Subhymenium – Generative hyphae thin-walled, smooth, colorless, frequently branched, more or less flexous, interwoven, 2–3 µm in diam, CB–, IKI–.
Basidia – Clavate, thin-walled, smooth, colorless, 4 sterigmata, sterigmata up to 5 µm, with a clamp connection and constriction at the base, more or less flexous, 17–27 × 5–8 µm, CB–, IKI–, tissues unchanged in KOH. Basidioles similar to basidia.
Cystidia – Thick-walled, smooth, colorless, with a clamp connection and constriction at the base, with a sharp apex, inflated near the bottom, frequently, 27–98 × 7–13 µm, CB+, IKI–, tissues unchanged in KOH.
Spores – Basidiospores reniform to subcylindrical, colorless, thin-walled, smooth, (6–)7–8(–9) × 3–4 µm, L = 7.60 µm, W = 3.20 µm, Q = 2–2.67 (n = 30/1), CB–, IKI–.
Notes – Peniophorella alba is grouped into Peniophorella based on the phylogenetic tree (Fig. 8), and is revealed as a sister to Pe. subpraetermissa. They exhibit some similar characteristics: white to milk white hymenophoral surface, and similar thick basidiomata (50–120 µm)[172]. However, Pe. alba can be delimited from Pe. subpraetermissa by its wider subicular hyphae (4–7 µm), and subymenial hyphae (2–6 µm). Peniophorella alba differs from Pe. subpraetermissa due to its smaller basidia (20–35 × 6–8µm)[172]. Pe. alba is similar to Pe. aspersa and Pe. crystallifera in having white hymenophore surface[170]. However, Pe. aspersa differs from Pe. alba by its warted hymenophore surface, presence of hyphidia, stephanocysts, and leptocystidia[170]. Pe. crystallifera can be distinguished from Pe. alba in its grandinioid hymenophore surface, and presence of heavily encrusted cystidia[170].
Schizoporaceae Jülich
Index Fungorum number: IF 80038
Type genus – Schizopora Velen.
Lyomyces P. Karst.
Index Fungorum number: IF18000
Type species – Lyomyces sambuci (Pers.) P. Karst.
Note – The genus Lyomyces is an important group of corticioid fungi within the Schizoporaceae (Hymenochaetales), and it was first established by Karsten[133] with Lyomyces sambuci designated as the type species. Members of the genus are representative white-rot fungi among corticioid fungi. Their basidiomata are typically white to cream, thin, resupinate to effused, and adherent to the substrate. The hymenophoral surface is smooth to slightly farinaceous, and the texture is delicate, and easily overlooked in the field. Due to the lack of molecular data, it has been difficult to delimit closely related genera within Schizoporaceae; therefore, Hyphodontia s. l. has been widely adopted[173,174]. Within Hyphodontia s. l., Lyomyces is nested in the Xylodon–Lyomyces–Rogersella clade, one of the six major lineages recognized in Hyphodontia s.l.[175]. Subsequent taxonomic revisions by Riebesehl & Langer[176] refined this framework and demonstrated that Hyphodontia s.l. should be segregated into six distinct genera: Hastodontia, Hyphodontia, Kneiffiella, Lagarobasidium, Lyomyces, and Xylodon. In the present study, based on combined analyses of ITS and nLSU sequences data (Fig. 9), a new species is described—Lyomyces membranaceus.
Lyomyces membranaceus L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 64,65
Fungal Names number: FN 573099

Figure 64.
Basidiomata of Lyomyces membranaceus (holotype IFP 020089).
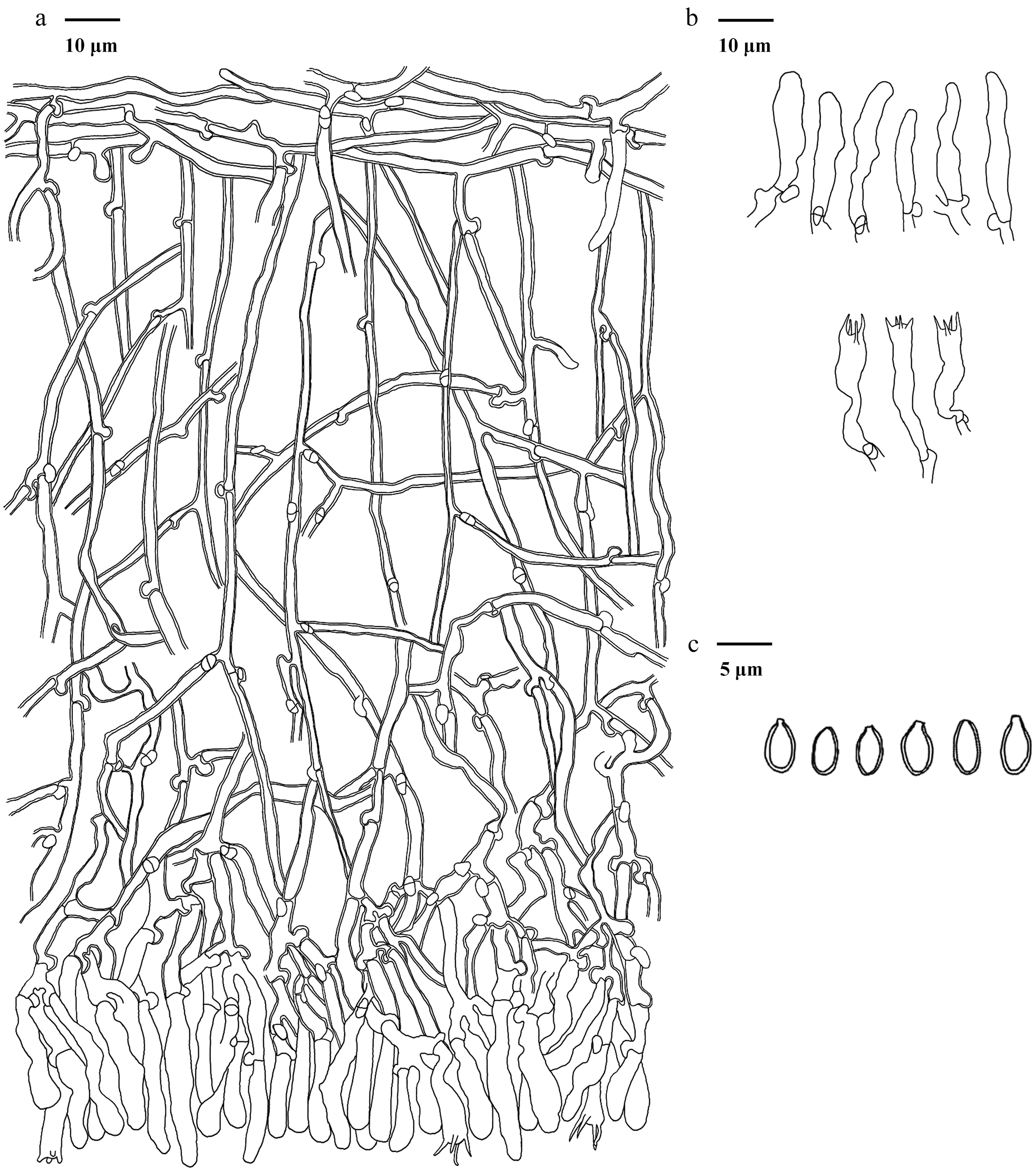
Figure 65.
Microscopic structures of Lyomyces membranaceus (drawn from the holotype IFP 020089). (a) Section of hymenium. (b) Basidia and basidioles. (c) Basidiospores.
Diagnosis – Lyomyces membranaceus is characterized by the smooth hymenophoral surface, subfusiform to subcylindrical cystidia and ellipsoid to subglobose basidiospores.
Etymology – Membranaceus (Lat.): referring to having membranaceous basidiomata.
Type – China, Xizang Autonomous Region, Bomi County, Yigong Tea Farm, GPS coordinates 30°07'55" N, 95°01'05" E, altitude 2,160 m, on fallen angiosperm branch, 24 October, 2021, Yuan 16290 (Holotype IFP 020089, GenBank ITS: PV919702; LSU: PX518154).
Description –Basidiomata annual, resupinate, adnate, membranaceous, adherent to the substrate and uneasy separable, up to 50–150 μm thick. Hymenophoral surface smooth, with cracked, white (–A1) to yellowish white (1A2–4A2) when dry. Sterile margin indistinct.
Hyphal structure – Hyphal system monomitic, generative hyphae with clamp connections, slightly thick-walled, CB+, IKI–, tissues unchanged in KOH.
Subiculum – Generative hyphae slightly thick-walled, smooth, colorless, moderately branched, parallel interwoven, 2–3 μm diameter.
Subhymenium – Generative hyphae slightly thick-walled, smooth, colorless, frequently branched, irregularly interwoven, 1–3 μm diameter.
Cystidia – subfusiform to subcylindrical, thin-walled, smooth, colorless, slightly constricted at the end, sometimes flexuous, 17–30 × 3–5 µm, CB–, IKI–.
Basidia – clavate, thin-walled, smooth, colorless, four sterigmata, sterigmata up to 5 µm long, with a clamp connection at the base, sometimes flexuous, 20–27 × 3–4 µm, CB–, IKI–. Basidioles similar to basidia.
Spores – Basidiospores ellipsoid to subglobose, thick-walled, smooth, colorless, (2.8–)3–4(–4.2) × (1.5–)1.7–2.8(–3.0) µm, L = 3.49 µm, W = 2.19 µm, Q = 1.28–2.0 (n = 60/2), CB+, IKI–. Chlamydospores and anamorpha absent.
Material examined (paratype) – China, Xizang Autonomous Region, Linzhi City, Bayi District, Pailong Town, 30°03′ N, 95°00′ E, altitude 3,416 m, on dead bamboo, 23 October, 2021, Yuan 16272 (Holotype IFP 020090, GenBank ITS: PV919703).
Notes – Based on the ITS + nLSU data (Fig. 9), the new species Lyomyces membranaceus is grouped with Lyomyces species, and is revealed as a sister to L. microfasciculatus. L. microfasciculatus can be distinguished from L. membranaceus by its warted to minutely odontioid hymenophoral surface, capitate cystidia and shorter basidia (14–15.5[–18.5] µm long)[175]. L. membranaceus resembles L. albopulverulentus, L. daweishanensis, and L. pantropicus in having a smooth hymenophoral surface. However, L. albopulverulentus distinguish from L. membranaceus in its longer capitate cystidia (37–54 × 5–9 µm), and wider basidia (7–9 µm wide)[177]. L. daweishanensis differs from L. membranaceus due to its two types cystidia (clavate or capitate), wider basidia (6.5–8.5 µm wide), and present of numerous crystals[178]. L. pantropicus differs from L. membranaceus by its four types cystidia, shorter basidia (16–17[–22] µm long), and oblong basidiospores[179].
Xylodon (Pers.) Gray
Index Fungorum number: IF 18771
Type species – Xylodon quercinus (Pers.) Gray
Note – The genus Xylodon was established by Gray in 1821, with Xylodon quercinus designated as its type species. Species of Xylodon were generally classified under Hyphodontia J. Erikss. It was not until Hjortstam & Ryvarden[180,181] reorganised Hyphodontia s.l. into different genera based on morphological features. Phylogenetic studies have demonstrated that Hyphodontia s.l. is a polyphyletic group, within which Xylodon and Kneiffiella P. Karst. contain the largest number of species[173,176,182]. Molecular systematic research has identified two major clades: the Xylodon-Lyomyces-Rogersella clade, and the Xylodon-Schizopora-Palifer clade, with species of Lyomyces, Palifer, Rogersella, Schizopora, and Xylodon intermingled within these clades[183]. The study included representative sequences and taxa of Hyphodontia s.l., such as Lyomyces, Palifer, Rogersella, Schizopora, and Xylodon. Notably, Riebesehl & Langer[176] found that distinguishing between Xylodon and Schizopora based on morphological and phylogenetic data was challenging, and thus suggested that Schizopora species should be merged into Xylodon. Additionally, the phylogenetic relationship between Xylodon species was clarified, with molecular data from the ITS and nLSU regions confirming that the genera Lagarobasidium and Xylodon are synonymous. In the present study, based on the ITS, nLSU, and nSSU data (Fig. 8), Xylodon is grouped with Lyomyces, Fasciodontia, Hyphodontia, and Kneiffiella. Furthermore, two new species—X. albus and X. bicystidiatus—are described.
Xylodon albus L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 66,67
Fungal Names number: FN 572427

Figure 66.
Basidiomata of Xylodon albus (holotype IFP 020016).
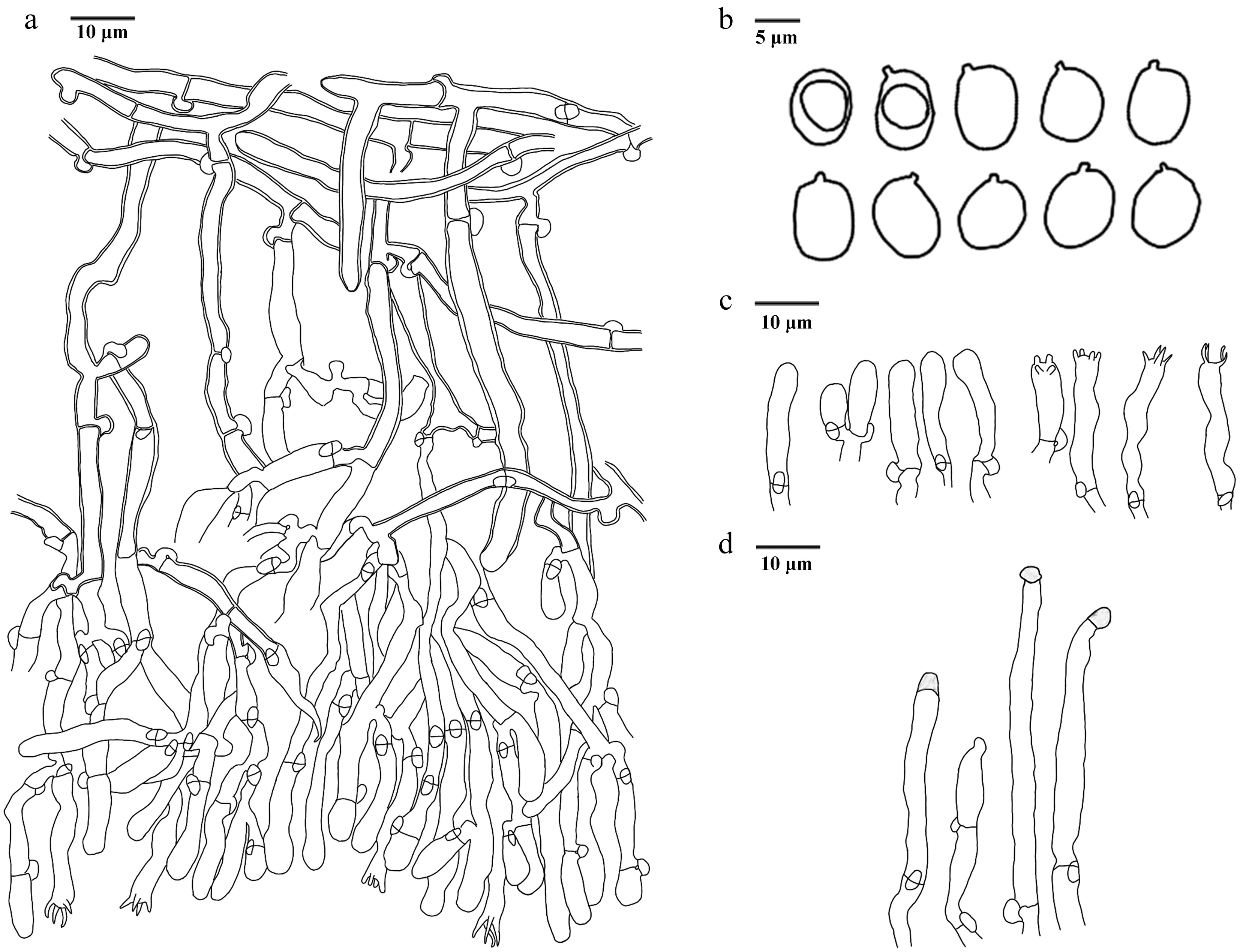
Figure 67.
Microscopic structures of Xylodon albus (drawn from the holotype IFP 020016). (a) Section of hymenium. (b) Basidiospores. (c) Basidia and basidioles. (d) Cystidia.
Diagnosis – Xylodon albus is characterized by the echinulate hymenophoral surface, narrowly clavate to clavate basidia, clavate to cylindrical cystidia, and ovoid to subglobose basidiospores.
Etymology – Albus (Lat.): referring to the white basidiomata (when fresh).
Type – China, Yunnan Province, Lincang City, Wulaoshan National Forest Park, GPS coordinates 23°54′49″ N, 100°10′55″ E, altitude 2,400 m, on fallen trunk of Pinus sp., 9 August, 2023, Yuan 17677 (holotype: IFP 020016, GenBank ITS: PV883101).
Description – Basidiomata annual, resupinate, separable, no releasing a smell, coriaceous when fresh, hard brittle when dry, up to 10 cm long, 5 cm wide, 50–200 µm thick. Hymenophoral surface echinulate, white (–A1) when fresh, pale yellow (1A3–4A3) when drying. Sterile margin indistinct.
Hyphal structure – Hyphal system monomitic, generative hyphae with clamp connections, thin- to thick-walled, IKI–, tissues unchanged in KOH.
Subiculum – Generative hyphae thick-walled, smooth, colorless, frequently branched, vertical at the branched, parallel interwoven, more or less flexous, 2–8.5 µm in diam, CB+.
Subhymenium – Generative hyphae thin-walled, smooth, colorless, frequently branched, more or less flexous, irregularly interwoven, 2–4 µm in diam., CB–.
Cystidia – Clavate to cylindrical, thin-walled, smooth, colorless, with a blunt apex, with a clamp connection at the base, rarely, 33–53 × 3–5 µm.
Basidia – Narrowly clavate to clavate, thin-walled, smooth, colorless, four sterigmata, sterigmata up to 5 µm, with a clamp connection at the base and slightly constricted, more or less flexous, with oily content, 14–23 × 3–4 µm, CB–, IKI–, tissues unchanged in KOH. Basidioles similar to basidia.
Spores – Basidiospores ovoid to subglobose, colorless, thin-walled, smooth, occasionally with oily content, 4–4.5(–5) × 3–3.5 µm, L = 4.10 µm, W = 3.02 µm, Q = 1.14–1.5 (n = 30/1), CB–, IKI–.
Notes – In this study, the new species Xylodon albus is clustered with X. acystidiatus and X. brevisetus without support (Fig. 8). X. albus resembles X. acystidiatus in its shared common color of basidiomata. However, X. acystidiatus can be distinguished from X. albus by its smooth hymenophoral surface, absence of cystidia, and larger basidia (20–25 × 4–4.5 µm)[29]. Xylodon albus shares similarities with X. brevisetus by echinulate hymenophoral surface and the same size of basidospores (4–5 × 3–3.5 µm). X. brevisetus can be delimited from X. albus by its torulose cystidia and larger basidia (20–25 × 4–5 µm)[184].
Xylodon bicystidiatus L.J. Zhou, Y.Q. Zhu & H.S. Yuan, sp. nov. Figs 68,69
Fungal Names number: FN 572428

Figure 68.
Basidiomata of Xylodon bicystidiatus (holotype IFP 020017).
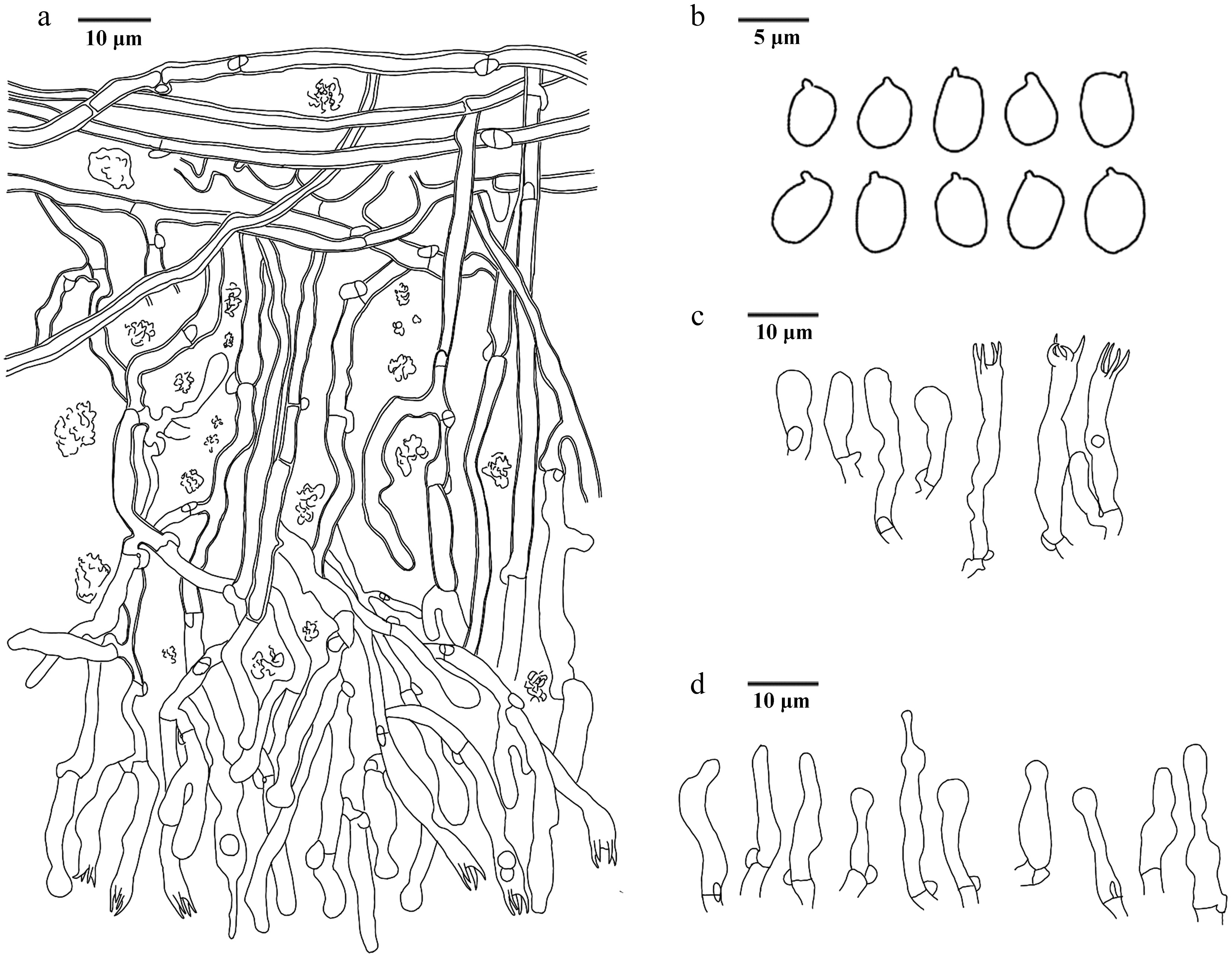
Figure 69.
Microscopic structures of Xylodon bicystidiatus (drawn from the holotype IFP 020017). (a) Section of hymenium. (b) Basidiospores. (c) Basidia and basidioles. (d) Cystidia.
Diagnosis – Xylodon bicystidiatus is characterized by the tuberculate to echinulate hymenophoral surface, ellipsoid to ovoid basidiospores, and two types cystidia.
Etymology – Bicystidiatus (Lat.): referring to having two types of cystidia.
Type – China, Yunnan Province, Diqing Tibetan Autonomous Prefecture, Shangri-La City, Potatso National Park, GPS coordinates 27°50′12″ N, 99°57′27″ E, altitude 3,655 m, on fallen trunk of Abies sp., 15 August, 2023, Yuan 18206 (holotype: IFP 020017, GenBank ITS: PV883100; LSU: PV257909; SSU: PV257948).
Description – Basidiomata annual, resupinate, separable, soft leather when fresh, hard brittle, cracked when dry, without odor, up to 15 cm long, 2 cm wide, 250 µm thick. Hymenophoral surface tuberculate to echinulate, pale yellow (1A3–4A3) to light yellow (1A4–4A5) when fresh, greyish yellow (2B3–3B7) when drying. Sterile margin indistinct.
Hyphal structure – Hyphal system monomitic, generative hyphae with clamp connections, thin- to thick-walled, CB+, IKI–, tissues unchanged in KOH.
Subiculum – Generative hyphae thick-walled, smooth, colorless, sparsely branched, parallel interwoven, 2–2.5 µm in diam.
Subhymenium – Generative hyphae thin- to slightly thick-walled, smooth, colorless, moderately branched, more or less flexous, interwoven, irregular crystals, 2–3.5 µm in diam.
Cystidia – Two types: (1) capitate cystidia, with a clamp connection at the base, thin-walled, smooth, colorless, slightly constricted at the neck, with a globose head, moderately of occurrence, 14–24 × 3–4 µm; (2) tapered cystidia, with a clamp connection at the base, thin-walled, smooth, colorless, subulate with a blunt apex, moderately of occurrence, 17.5–29 × 3–4 µm, CB–, IKI–, tissues unchanged in KOH.
Basidia – Narrowly clavate to clavate, thin-walled, smooth, colorless, four sterigmata, sterigmata up to 6 µm, with a clamp connection at the base and slightly constricted, or with oily content, 19–32 × 3–5 µm, CB–, IKI–, tissues unchanged in KOH. Basidioles similar to basidia.
Spores – Basidiospores ellipsoid to ovoid, colorless, thin-walled, smooth, (4–)4.5–5(–5.5) × 3–4 µm, L = 4.92 µm, W = 3.52 µm, Q = 1.25–1.67 (n = 30/1), CB–, IKI–.
Notes – The new species Xylodon bicystidiatus is classified within Xylodon (Fig. 8) and forms a clade with X. attenuates. X. bicystidiatus resembles X. attenuates by sharing cystidia and basidiospores of similar size[185]. However, X. attenuates can be distinguished from X. bicystidiatus by smaller basidia ([12.2–]13.0–18.1[–20] × [3.1–]3.2–4.6[–4.9] μm), wider subymenial hyphae (2.4–4.6 µm), and subicular hyphae (2.7–4.6 µm). X. bicystidiatus is similar to X. luteodontioides and X. wumengshanensis in having yellow hymenophoral surface[186]. However, X. luteodontioides differs from X. bicystidiatus by its presence of schizopapillate cystidia, and larger basidiospores (3.5–4.5 × 2.5–3.5 µm)[186]. X. wumengshanensis can be distinguished from X. bicystidiatus by its presence of fusoid cystidia and larger basidiospores ([4.5–] 5–6.5[–7] × 4–5.5 µm)[186].
Key to species of Hydnellum from China. 1 Basidiospores hyaline 2 1 Basidiospores brown 12 2 Basidia clavate or sinuous 3 2 Basidia clavate 4 3 Pileus round to circular Hydnellum liantaishanense 3 Pileus flabelliform to subcircular Hydnellum tomentosum 4 Generative hyphae with mostly simple-septa, occasionally clamped 5 4 Generative hyphae with simple-septa 7 5 Stipe central 6 5 Stipe central to lateral Hydnellum melanocarpum 6 Pileus infundibuliform Hydnellum concentricum 6 Pileus irregularly flabelliform Hydnellum hydrangeoides 7 Stipe central 8 7 Stipe lateral 10 8 Pileal margin white Hydnellum testaceum 8 Pileal margin not white 9 9 Taste mild Hydnellum infundibuliforme 9 Taste bitter Hydnellum carnosum 10 Basidiospores subglobose to ellipsoidal 11 10 Basidiospores subglobose to globose Hydnellum chocolatum 11 Pilieal surface smooth Hydnellum crassipileatum 11 Pilieal surface fibrillose Hydnellum radiatum 12 Generative hyphae with simple-septa 16 12 Generative hyphae not with simple-septa 13 13 Basidiomata fleshy Hydnellum versipelle 13 Basidiomata woody 14 14 Stipe central 15 14 Stipe lateral Hydnellum atrospinosum 15 Pilieal surface glabrous Hydnellum diabolus 15 Pilieal surface velutinous to tomentose Hydnellum suaveolens 16 Generative hyphae only with simple-septa 20 16 Generative hyphae mostly with simple-septa, occasionally clamped 17 17 Stipe surface glabrous Hydnellum cinnamomea 17 Stipe surface not glabrous 18 18 Stipe surface plushy Hydnellum caeruleum 18 Stipe surface tomentose 19 19 Taste mild Hydnellum fibulatum 19 Taste acrid Hydnellum peckii 20 Pileal surface scaled 21 20 Pileal surface not scaled 28 21 Taste none 22 21 Taste like something 24 22 Pileus depressed Hydnellum subscabrosellum 22 Pileus planar 23 23 Pileus circular Hydnellum subailaoensis 23 Pileus ellipsoid to round Hydnellum grosselepidotum 24 Taste mild Hydnellum edulium 24 Taste bitter 25 25 Pileal margin white Hydnellum fagiscabrosum 25 Pileal margin not white 26 26 Spines up to 5 mm long 27 26 Spines up to 1 mm long Hydnellum lidongensis 27 Pileus planar to plano-convex Hydnellum illudens 27 Pileus depressed Hydnellum glaucopus 28 Stipe lateral 29 28 Stipe central or central to lateral 30 29 Pileal margin incurved, occasionally incised Hydnellum ailaoense 29 Pileal margin lobed Hydnellum sulcatum 30 Stipe central 31 30 Stipe central to lateral 39 31 Pilieal surface glabrous 32 31 Pilieal surface not glabrous 34 32 Stipe surface tomentose to matted 33 32 Stipe surface spongy Hydnellum succulentus 33 Pilieal surface azonate Hydnellum coactum 33 Pilieal surface zonate Hydnellum ferrugineum 34 Pilieal surface azonate 35 34 Pilieal surface zonate 36 35 Taste bitter Hydnellum atrorubrum 35 Taste none Hydnellum porphyreum 36 Taste disagreeable 37 36 Taste none 38 37 Spines up to 5 mm Hydnellum aurantiacum 37 Spines up to 9 mm Hydnellum pineticola 38 Stipe surface subtomentose to matted Hydnellum chrysinum 38 Stipe surface glabrous Hydnellum tarda 39 Pileal margin white 44 39 Pileal margin not white 40 40 Taste none 41 40 Taste bitter 42 41 Basidiospores irregularly Hydnellum martioflavum 41 Basidiospores subglobose Hydnellum earlianum 42 Stipe rugose 43 42 Stipe velutinous to matted Hydnellum spongiosipes 43 Spines up to 4 mm Hydnellum inflatum 43 Spines up to 2 mm Hydnellum granulosum 44 Basidiospores subglobose to globose 45 44 Basidiospores irregular 46 45 Spines up to 2 mm Hydnellum xanthopus 45 Spines up to 4 mm Hydnellum nitida 46 Basidiomata fleshy Hydnellum subalpinum 46 Basidiomata woody 47 47 Stipe surface tomentose 49 47 Stipe surface not tomentose 48 48 Taste acrid Hydnellum bomiense 48 Taste none Hydnellum qinghaiense 49 Taste none Hydnellum squamulosum 49 Taste mild 50 50 Pileal margin lobed 51 50 Pileal margin eroded Hydnellum yunnanense 51 Pileus infundibuliform to flabelliform Hydnellum brunneorubrum 51 Pileus depressed to flabelliform or irregularly circular Hydnellum rubidofuscum Key to resupinate species of Thelephora from China. 1 Rhizomorphs present 2 1 Basidiomata absent 29 2 Rhizomorphs differentiated (with a distinct central core and outer layer) 3 2 Rhizomorphs undifferentiated (uniform structure) 8 3 Rhizomorphs dimitic 4 3 Rhizomorphs monomitic 5 4 Rhizomorphs type G; central hyphae usually simple septate Thelephora dimidiata 4 Rhizomorphs type G; central hyphae clamped Thelephora incrustata 5 Cystidia present 6 5 Cystidia absent 7 6 Spores aculeate (up to 2 5 µm long) Thelephora cystidiata 6 Spores echinulate to aculeate (up to 1 5 µm long) Thelephora citrinocystidiata 7 Basidiomata arachnoid, continuous Thelephora qingyuanensis 7 Basidiomata mucedinoid, discontinuous Thelephora brevis 8 Rhizomorphs of type A 9 8 Rhizomorphs of type B 11 9 Rhizomorph hyphae thin-walled Thelephora tenuirhizomorpha 9 Rhizomorph hyphae thick-walled 10 10 Subicular hyphae thick-walled, 3.5–6.5 μm Thelephora fuscocrustosa 10 Subicular hyphae slightly thick- to thick-walled Thelephora pallidomarginata 11 Hyphal septation in rhizomorphs exclusively by simple-septa 12 11 Hyphal septation in rhizomorphs with clamp connections (or both clamps and simple-septa) 14 12 Rhizomorph hyphae ≥ 3 µm 13 12 Rhizomorph hyphae < 3 µm Thelephora efibulata 13 Subicular hyphae thick-walled, smooth, 3–5 μm Thelephora efibulis 13 Subicular hyphae slightly thick walled, with encrustation, 4–8 µm Thelephora olivaceomarginata 14 Rhizomorph hyphae with both clamp connections and simple-septa 15 14 Rhizomorph hyphae possessing exclusively clamp connections 20 15 Spores thick-walled 16 15 Spores slightly thick-walled 17 16 Basidiomata mucedinoid Thelephora angusta 16 Basidiomata arachnoid Thelephora fuscoaraneosa 17 Spores ≤ 7 µm Thelephora aureomarginata 17 Spores > 7 µm 18 18 Sterigmata > 5 µm Thelephora separata 18 Sterigmata ≤ 5 µm 19 19 Cystidia present Thelephora capitatocystidiata 19 Cystidia absent Thelephora brunneoflava 20 Basidiomata discontinuous 21 20 Basidiomata continuous 22 21 Basidia utriform, not stalked Thelephora linzhiensis 21 Basidia clavate, stalked Thelephora interrupta 22 Spores wall-thined Thelephora casiae 22 Spores wall-thicked 23 23 Subglobose to bi-, tri- or quadra-lobed 24 23 Irregularly globose or lobed 25 24 Spores echinulate, up to 1 μm long Thelephora flavidobadia 24 Spores echinulate to aculeate, up to 2 µm long Thelephora guiyangensis 25 Cystidia present Thelephora gloeocystidiata 25 Cystidia absent 26 26 Basidia not stalked Thelephora inconspicua 26 Basidia stalked 27 27 Rhizomorph ≥ 20 µm Thelephora fuscogranulosa 27 Rhizomorph ≤ 20 µm 28 28 Basidia not sinuous, Subhymenial frequently branched Thelephora olivacea 28 Basidia sinuous, Subhymenial occasionally branched Thelephora parvispora 29 Cystidia present Thelephora longicystidiata 29 Cystidia absent 2 30 Spore thin or thin- to slightly thick-walled 3 30 Spore slightly thick-walled or thick-walled 5 31 Spore thin-walled Thelephora rotundata 31 Spore thin- to slightly thick-walled 4 32 Hyphal septation with both simple-septa and clamp connections Thelephora coffeae 32 Hyphal septation exclusively with clamp connections Thelephora globospora 33 Spores slightly thick-walled 6 33 Spores thick-walled 16 34 Spore ornamentation ≥ 2 µm long 7 34 Spore ornamentation < 2 µm long 11 35 Spores > 10 µm long Thelephora nanyigouensis 35 Spores ≤ 10 µm 8 36 Basidia utriform 9 36 Basidia clavate 10 37 Spores with aculeate (isolated), basidia not sinuous Thelephora griseofusca 37 Spores with echinulate (sometimes grouped in 2 or more), basidia sinuous Thelephora segregata 38 Basidia stalked, sterigmata > 10 µm long, subicular hyphae with crystal Thelephora longiaculeifera 38 Basidia not stalked, sterigmata < 10 µm long, subicular hyphae smooth Thelephora duplexa 39 Spores > 7 µm Thelephora bomiensis 39 Spores ≤ 7 µm 12 40 Basidia utriform Thelephora schrenkiana 40 Basidia clavate 13 41 Basidiomata granulose 14 41 Basidiomata smooth 15 42 Sterigmata ≤ 5 µm long Thelephora griseocastanea 42 Sterigmata > 5 µm long Thelephora olivaceobrunnea 43 Spores > 6 long and > 5 µm wide Thelephora griseomarginata 43 Spores ≤ 6 long and ≤ 5 µm wide Thelephora pallidocastanea 44 Spore ornamentation wart-like (verrucose or nodulose) 17 44 Spore ornamentation spine-like (echinulate or aculeate) 21 45 Spore broadly ellipsoid Thelephora velutina 45 Spore subglobose to globose 18 46 Hyphal > 5 µm in diam 19 46 Hyphal ≤ 5 µm in diam 20 47 Basidiospores membranaceous Thelephora wumenshanensis 47 Basidiospores farinaceous Thelephora tenuifarinacea 48 Basidia clavate (< 50 µm long) Thelephora olivaceobasidiosa 48 Basidia cylindrical to subclavate (> 50 µm long) Thelephora yunnanensis 49 Basidia clavate 22 49 Basidia utriform 25 50 Basidiospores > 7 µm long, > 6 µm wide 23 50 Basidiospores < 7 µm long, < 6 µm wide 24 51 Basidiomata arachnoid, continuous; subicular hyphae frequently branched Thelephora asiae-orientalis 51 Basidiomata mucedinoid, uncontinuous; subicular hyphae occasionally branched Thelephora aurimucida 52 Subicular hyphae frequently branched
> 5 μm; hymenophoral surface farinoseThelephora farinosa 52 Subicular hyphae rarely branched
≤ 5 μm; hymenophoral surface granuloseThelephora storea 53 Spore spine-like ornamentation > 1.5 µm long 26 53 Spore spine-like ornamentation ≤ 1.5 µm long 34 54 Spore echinuli usually isolated 27 54 Spore echinuli usually isolated, sometimes grouped in two or more 29 55 Spore echinuli subglobose to bi-lobed Thelephora longiechinuli 55 Spore echinuli subglobose to globose 28 56 Sterigmata > 10 μm long Thelephora conclusa 56 Sterigmata < 10 μm long Thelephora atrocastanea 57 Sterigmata > 6 μm long 30 57 Sterigmata ≤ 6 μm long 32 58 Subicular hyphae occasionally branched
≤ 5 μmThelephora fuscopelliculosa 58 Subicular hyphae frequently branched
> 5 μm31 59 Basidiomata dark blonde to yellowish brown Thelephora stipitata 59 Basidiomata brownish grey to dark brown Thelephora brunneogrisea 60 Hyphal ≤ 5 μm in diam Thelephora exiguelata 60 Hyphal > 5 μm in diam 33 61 Spore > 9 μm long > 8 μm wide Thelephora megaspora 61 Spore < 9 μm long < 8 μm wide Thelephora pallidobrunnea 62 Basidiomata granulose Thelephora fuscofarinosa 62 Basidiomata smooth 35 63 Subhymenial hyphae slightly thick-walled or thick-walled 36 63 Subhymenial hyphae thin-walled 38 64 Basidiomata crustose Thelephora atrobadia 64 Basidiomata mucedinoid 37 65 Subhymenial hyphae slightly thick-walled, occasionally branched > 5 μm Thelephora pertenuis 65 Subhymenial hyphae thick-walled, frequently branched ≤ 5 μm Thelephora latihypha 66 Spore globose or lobed 39 66 Spore subglobose to globose 40 67 Spore irregularly globose or lobed, aculeate, up to 1.5 μm long Thelephora liaoningensis 67 Spore subglobose to bi-, tri-, or quadra-lobed, echinulate, up to 1 μm long Thelephora chayuensis 68 Sterigmata > 5.5 μm Thelephora kanasensis 68 Sterigmata ≤ 5.5 μm 41 69 Spore ornamentation echinulate to aculeate, up to 1 µm long Thelephora aurantispora 69 Spore ornamentation echinulate, up to 1.5 μm long Thelephora changbaiensis Key to upright species of Thelephora from China. 1 Cystidia present 2 1 Cystidia absent 4 2 Cystidia one type 3 2 Cystidia two types Thelephora grandinioides 3 Cystidia clavate Thelephora ganbajun 3 Cystidia tubular Thelephora wuliangshanensis 4 Sterigmata 2–4 5 4 Sterigmata 4 9 5 Hyphae commonly clamped and
simple-septateThelephora austrosinensis 5 Hyphae clamped 6 6 Basidiospores umber purple Thelephora caryophyllea 6 Basidiospores fuscous purple 7 7 Basidiomata imbricate 8 7 Basidiomata clavarioid Thelephora palmata 8 Pileus spathulate to flabelliform Thelephora japonic 8 Pileus dimidiate or spuriously
infundibuliformThelephora terrestris 9 Hyphae clamped 10 9 Hyphae clamped and simple-septate 16 10 Basidiospores lobed 12 10 Basidiospores not lobed 11 11 Basidia subclavate Thelephora subtropica 11 Basidia barreled Thelephora yunnanensis 12 Hymenial surface concolorous with
abhymenial surface13 12 Hymenial surface not concolorous with
abhymenial surface14 13 Pileus spathulate to narrow petaloid Thelephora dactyliophora 13 Pileus clavate to pinnatifid or ramiform Thelephora pinnatifida 14 Smell strong Thelephora aurantiotincta 14 Smell not strong 15 15 Basidiospores umber brown Thelephora penicillata 15 Basidiospores olive buff Thelephora vialis 16 Stipe central 18 16 Stipe central to lateral 17 17 Basidiospores bluish green Thelephora pseudoganbajun 17 Basidiospores yellowish brown Thelephora petaloides 18 Basidiospores hyaline Thelephora sikkimensis 18 Basidiospores not hyaline 19 19 Abhymenial surface azonate 20 19 Abhymenial surface zonate 22 20 Pileus imbricate Thelephora lacunosa 20 Pileus coralloid 21 21 Pileal margin deeply lacerate Thelephora nigromarginata 21 Pileal margin tips needle-like Thelephora scopiformis 22 Smell mild 23 22 Smell no odor 25 23 Stipe subconical to broadened or flatted Thelephora cacao 23 Stipe clavillose to flatted or broadened 24 24 Pileus infundibuliform to ligulate Thelephora fasciculata 24 Pileus clavate to coralliform Thelephora microcarpa 25 Abhymenial surface radially rugulose or
wrinkled26 25 Abhymenial surface wrinkled Thelephora nebula 26 Pileal margin imperceptibly wavy Thelephora glaucoflora 26 Pileal margin irregularly lobed to wavy Thelephora aquila -
In the present study, taxonomic and phylogenetic analyses were conducted on corticioid and hydnoid fungi belonging to the orders Thelephorales and Hymenochaetales, which were collected from subtropical and southern temperate forests in China. The internal transcribed spacer (ITS) regions, along with the large and small subunit nuclear ribosomal RNA genes (nLSU and nSSU), were used to identify the new species and analyze the phylogenetic relationships within Thelephorales and Hymenochaetales, and complemented by small subunit mitochondrial rRNA gene sequences (mtSSU) for the genus Thelephora.
The basidiomata of Thelephorales are primarily characterized by two morphological types: resupinate (flat and appressed), and erect (upright). In classical taxonomy, these morphological characteristics (erect or resupinate basidiomata) were widely used as key criteria for genus delineation. However, with the advancement of modern molecular phylogenetics, accumulating evidence has revealed that species with markedly divergent basidiomata (erect vs resupinate) may share close evolutionary relationships. In recent years, progress has been made in investigating the diversity and phylogeny of Thelephora/Tomentella, yet their phylogenetic relationships remain unclear[38,147,151]. Micromorphologically, the two genera share similar characteristics, including a monomitic hyphal system with clamp connections in generative hyphae and warted or echinulate basidiospores. Macromorphologically, members of Tomentella possess resupinate basidiomata—either separable from or adherent to the substrate—with hymenophoral surfaces that are smooth or granulose, and exhibit various colors. In contrast, Thelephora is characterized by the diverse shapes of basidiomata (stereoid, imbricate, rosette, infundibuliform, or coralloid), abhymenial surfaces ranging from glabrous to strigose or wrinkled, and hymenophoral surfaces that are smooth, slightly rugose, or warted. Ecologically, Thelephora basidiomata are typically epigeous (growing on the ground), while those of Tomentella are often found on decaying wood or soil, etc., indicating distinct ecological niches between the two genera. Phylogenetically, the two genera form a paraphyletic group[138,147]. Based on previous studies, Kõljalg et al.[151] proposed merging Tomentella into Thelephora but noted that their phylogenetic relationships require further clarification using additional genetic loci or whole-genome data. In this study, phylogenetic analyses of 193 species revealed a strongly supported monophyletic clade (100% ML, 1.00 BPP, Fig. 6) encompassing all Thelephora samples, consistent with the taxonomic framework established by Kõljalg et al.[151].
The stipitate hydnoid genera within Thelephorales include Hydnellum, Neosarcodon, Phellodon, and Sarcodon. In the present study, the phylogenetic trees (Figs 2–4) clarified the phylogenetic relationships among members of Hydnellum, Phellodon, Sarcodon, and Neosarcodon at both the generic and species levels. Three genera, Hydnellum, Neosarcodon, and Sarcodon formed a distinct clade with high support (76% ML, 0.99 BPP, Fig. 6). Collectively, Hydnellum, Neosarcodon, and Sarcodon form a highly supported clade within Thelephorales. These results are consistent with the latest taxonomic framework for genera in Thelephorales[13]. However, Boletopsis (Boletopsidaceae) is closely related to Lenzitopsis (Lenzitopsidaceae) (Fig. 6). This finding conflicts with previous studies on intergeneric phylogenetic relationships[13]. The species diversity of these relatively undersampled genera requires further investigation together with other taxa within Thelephorales. Additionally, incorporating more gene fragments and whole-genome sequencing data into phylogenetic analyses may help clarify the intergeneric phylogenetic relationships. However, no whole-genome sequences are currently available for any species of Amaurodon or Tomentellopsis. Hence, it is necessary to carry out more in-depth phylogenetic analyses for these genera with more samples and sequences worldwide.
The stipitate hydnoid species within the order Thelephorales are the ectomycorrhizal fungi. Records of basidiomata for many stipitate hydnoid species are declining across numerous countries, with some even disappearing entirely[58,187−189]. For instance, in countries such as the Netherlands, Germany, Poland, Belgium, Sweden, and Czechoslovakia, species of Boletopsis, Hydnellum, Phellodon, and Sarcodon have undergone significant population declines or face extinction risks[56,58,60]. Members of the stipitate hydnoid fungal genera are considered endangered and have been included in the Red Data Lists of several European countries, apparently due to soil nitrification[58,59,190−194]. Following the publication of the International Union for the Conservation of Nature (IUCN) Red List update 2023.1, five stipitate hydnoid fungi species have been officially assessed as threatened on a global scale. In the present study, 11 new species of stipitate hydnoid fungi (Hydnellum, Neosarcodon, Phellodon, and Sarcodon) are described based solely on their holotypes. Given that some stipitate hydnoid species are known to produce relatively few basidiomata[48,195−197], we speculate that the new species not only exhibit an extremely restricted distribution but also produce basidiomata at low abundance in the wild. This phenomenon indicates parallel conservation crises between China and European nations, with stipitate hydnoid fungi undergoing dramatic declines that may approach regional extinction. Meanwhile, the significant decline in stipitate hydnoid fungi abundance serves as a critical bioindicator of diminishing basidiomata among ectomycorrhizal fungi worldwide, underscoring the increasing urgency of forest ecosystem protection. The identification of previously unknown or misclassified taxa in China will facilitate more comprehensive assessments of their distribution patterns, rarity, habitat preferences, and conservation status. This, in turn, can inform potential management strategies where necessary.
Although high species diversity among Hymenochaetales has been discovered in recent years[29,33,198−201], exploring species resources still holds great significance in China. The present study focuses on the corticioid fungi in Hymenochaetales characterized by a smooth to tuberculate hymenophore, viz. Hymenochaete (Hymenochaetaceae), Peniophorella (Peniophorellaceae), Lyomyces (Schizoporaceae), and Xylodon (Schizoporaceae). Five new species from these four genera are described, all of which are wood-inhabiting fungi. Although diverse hymenophoral configurations may exist even within the same genus of Hymenochaetales (e.g., Hymenochaete and Xylodon), such morphological variations do not impede species delimitation. For instance, the basidiomata of Hymenochaete species can exhibit forms ranging from effused-reflexed to pileate, or resupinate, as observed in the newly described species, Hymenochaete baishanzuensis.
Three genera—Fasciodontia, Lyomyces, and Xylodon—previously classified under Hyphodontia sensu lato, form a strongly supported clade, which is consistent with findings from previous studies[29,32,177]. The genus Xylodon exhibits remarkable diversity in hymenophore surface morphology, including smooth to grandinioid, tuberculate, raduloid, distinctly hydnoid and poroid. Notably, both newly described species of Xylodon described in this study are characterized by tuberculate hymenophoral surfaces. The phylogenetic analysis reveals that the new species, Lyomyces membranaceus, is most closely related to Lyomyces microfasciculatus (Fig. 9). Additionally, the present study indicates that Chinese specimens identified as L. microfasciculatus may represent a species complex, which includes L. membranaceus. The findings not only elucidate the taxonomic complexities surrounding the genus Lyomyces in China but also suggest it harbors considerable undiscovered diversity, meriting in-depth phylogenetic and taxonomic investigations. Peniophorella occupies an independent position relative to other known genera and families in Hymenochaetales, which is consistent with previous research[32]. To date, 39 species are accepted in Peniophorella, although the phylogenetic placement of certain taxa—such as Pe. pallida (Fig. 8)—requires further investigation.
In this study, 30 new species within the Thelephorales and Hymenochaetales are described, thereby enhancing the corticioid and hydnoid species diversity in southern China, and facilitating their resource utilization. However, within the Thelephorales, the phylogenetic relationships between families remain ambiguous, and their boundaries are still not clearly delineated. Genomic data from more species of Thelephorales will help determine the current taxonomic framework.
-
Not applicable.
-
The authors confirm contribution to the paper as follows: conceptualization, supervision, funding acquisition, and writing–review: Yuan H; investigation: Yuan H, Wei Y, Zhu Y, Zhou L, Liu J, Yuan Y; data collection: Wei Y, Zhu Y, Zhou L, Liu J, Yuan Y; analysis and interpretation of results: Zhu Y, Zhou L; draft manuscript preparation: Zhu Y, Zhou L. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available in the National Center for Biotechnology Information (NCBI) databases, the UNITE database, and the Fungal Names repository.
-
We are grateful to Xue-Lian Gao, Yan-Yan He, Xiang-Hua Wang, Yang-Ling Deng, Jian-Feng Tan, and Zhong-Ping Feng for collecting specimens. This study was financed by the National Natural Science Foundation of China (Project Nos U2102220 & 32570013), the Fund of CAS Key Laboratory of Forest Ecology and Silviculture, Institute of Applied Ecology, Chinese Academy of Sciences (No. KLFES-2028), and the Fund of Biological Resources Programme, Chinese Academy of Sciences (CAS-TAX-24-027).
-
The authors declare that there is no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/mycosphere-0026-0001.
-
#Authors contributed equally: Yaquan Zhu, Linjiang zhou
- Supplementary Table S1 A list of species, specimens and GenBank accession numbers of the sequences used in this study.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhu Y, Zhou L, Liu J, Yuan Y, Wei Y, et al. 2026. Thirty new species in Thelephorales and Hymenochaetales from the Eastern Himalayas and Southern China. Mycosphere 17: e001 doi: 10.48130/mycosphere-0026-0001 

Thirty new species in Thelephorales and Hymenochaetales from the Eastern Himalayas and Southern China
- Received: 31 July 2025
- Revised: 17 December 2025
- Accepted: 25 December 2025
- Published online: 28 February 2026
Abstract: Corticioid and hydnoid fungi play crucial roles in forest ecosystems, particularly in wood degradation and the formation of symbiotic relationships with plants, and exhibit extremely high species diversity in China. However, the taxonomic and phylogenetic relationships among taxa of corticioid and hydnoid fungi remain insufficiently understood. In this study, an extensive survey and sampling was conducted for Thelephorales and Hymenochaetales in subtropical and southern temperate forests of China. The internal transcribed spacer (ITS) regions, along with the large and small subunit nuclear ribosomal RNA genes (nLSU and nSSU), were used to identify the species and analyze the phylogenetic relationships within different genera, and complemented by a small subunit of mitochondrial rRNA gene sequences (mtSSU) for the genus Thelephora. Morphological and molecular analyses revealed 30 new species across ten genera of Thelephorales and Hymenochaetales. The novel species described in this paper include Hydnellum carnosum, Hyd. hydrangeoides, Hyd. infundibuliforme, Hyd. liantaishanense, Hyd. porphyreum, Hyd. testaceum, Hyd. tomentosum, Hymenochaete baishanzuensis, Lyomyces membranaceus, Neosarcodon atroviolaceus, N. bambusicola, Odontia kunmingensis, Peniophorella alba, Phellodon albospinus, Ph. zonatus, Sarcodon squamulosus, Thelephora angusta, T. bomiensis, T. cacao, T. chayuensis, T. fasciculata, T. latihypha, T. linzhiensis, T. longicystidiata, T. microcarpa, T. nanyigouensis, T. nigromarginata, T. scopiformis, Xylodon albus, and X. bicystidiatus. Descriptions and illustrations of these newly proposed species are provided, along with notes discussing their distinguishing characters and those of their allied taxa in this paper. Additionally, the delimitation and diversity of these taxa in terms of morphology, ecology, and phylogeny are explored. Three keys to the species of Hydnellum, resupinate and upright Thelephora from China are provided.
-
Key words:
- Basidiomycota /
- Corticioid and hydnoid fungi /
- Hymenochaetales /
- Thelephorales /
- Molecular phylogeny /
- Taxonomy