









-
Cassava (Manihot esculenta Crantz) originates from the Amazon region and is primarily cultivated in tropical areas. It is one of the world's starch-rich crops, serving as a staple food in tropical regions like Africa[1]. Additionally, it is utilized as feed and as a raw material in various industries[2]. Cassava bacterial blight (CBB), caused by Xanthomonas phaseoli pv. manihotis (Xpm) stands out as the most destructive disease affecting cassava production[3]. CBB played a significant role in the famine in Zaire between 1970 and 1975, leading to a staggering loss of up to 80% of cassava production in Central Africa[3]. Identification and characterization of resistant genes responsive to Xpm infection in cassava will provide valuable genetic resources for developing stress-resistant cassava cultivars.
Stress-associated proteins (SAPs) are typical A20/AN1 zinc-finger proteins with physiological functions in regulating abiotic stress signaling and defense responses to disease in plants[4−6]. There are 14 genes in Arabidopsis, 18 in rice, 17 in Pinus massoniana, 17 in potato, and 27 in sunflower[7−11]. Based on the number and structure of zinc-finger motifs, Arabidopsis SAP proteins can be classified into three groups (I-III). Group I comprises of a A20 zinc finger domain characterized by multiple Cys2-Cys2 finger motifs, while Group II includes an AN1 zinc finger domain containing multiple Cys2-His2 finger motifs, or both. Group III consists of proteins with additional Cys2-His2 RING motifs at the C-terminus[10]. Previous studies have shown that SAPs enhance the ability of plants to respond to abiotic stress. Overexpressing ZmSAP8 enhanced the tolerance to drought stress in Arabidopsis, with higher seed germination and longer root length[12]. Overexpression of ZmSAP1 or ZmSAP7 significantly enhances salt stress tolerance in Arabidopsis and rice[13]. Overexpression of NtSAP9 significantly enhances freezing tolerance in tobacco, whereas knockout of NtSAP9 resulted in reduced freezing tolerance[14]. Meanwhile, the SAP gene family participates in regulating plant growth and development. For example, in Artemisia annua, AaSAP1 positively regulated the development of glandular trichomes, thereby increasing the production of artemisinin[15]. Oryza sativa OsZFP185 regulates plant growth and stress responses by affecting GA and ABA biosynthesis in rice[16]. Moreover, studies have shown that SAPs play important roles in plant immunity. For instance, overexpression of tomato SlSAP3 led to increased accumulation of reactive oxygen species (ROS) upon flg22 treatment, accompanied by elevated expression of immune marker genes triggered by PAMP, such as SlPTI5 and SlLRR22[17]. Further research revealed that SlSAP3 positively regulated tomato resistance to the Pseudomonas syringae pv. tomato DC3000 (Pst DC3000) pathogen through the SA signal[17]. Overexpression of rice OsSAP1 enhanced plant resistance against virulent bacterial pathogens, and upregulated the expression of defense genes in tobacco[18]. Silencing or knocking out of SAP12 in tobacco and Arabidopsis increased plant susceptibility to Meloidogyne incognita[5]. Therefore, investigating the SAP gene family is crucial for understanding its functional diversity across multiple plant species.
However, the molecular mechanisms underlying SAP-mediated stress tolerance and pathogen resistance remain incompletely understood. Multiple regulatory pathways appear to be involved in SAP function. First, SAP proteins participate in signal transduction through protein-protein interactions with RLCKs under abiotic stress[19]. For instance, OsSAP11 interacts with OsRLCK253 via its A20 zinc-finger domain, thereby enhancing water-deficit and salt stress tolerance in transgenic Arabidopsis, and modulating the expression of endogenous stress-response genes[20]. Additionally, post-translational modifications of SAP proteins, particularly phosphorylation and ubiquitination play critical roles in regulating their function. In Arabidopsis, AtSAP9 interacts with RADIATION SENSITIVE23d (Rad23d) to target ubiquitinated proteins for proteasome degradation, thereby fine-tuning abiotic stress responses and homeostasis of stress-related proteins[21]. Furthermore, SAP proteins act as transcriptional regulators by binding to cis-elements in the promoter regions of stress-related genes[19]. GaZnF, a potential SAP-family transcription factor, binds to MYB-box elements in the GUSP1 promoter, suggesting that SAP proteins can directly or indirectly regulate stress-related gene expression in response to drought, salt, and heavy metal stresses[22]. MiPDI1 interacts with SlSAP12, an ortholog of the redox-regulated AtSAP12, and played a significant role in plant responses to abiotic and biotic stresses[5]. Despite extensive characterization of SAP functions in various plant-pathogen interactions, the molecular mechanisms by which SAP proteins contribute to resistance in cassava against Xpm have not yet been elucidated.
In the present study, we identified and characterized the SAP gene family containing A20/AN1 zinc-finger domains from the cassava genome and assessed their transcriptional responses to various abiotic and biotic stresses. Identification, characterization, and elucidation of evolutionary patterns of the cassava SAP gene family, along with analyzing their involvement in diverse stress responses could substantially advance the development of strategies to enhance stress tolerance and pathogen resistance in cassava. Additionally, our research specifically focuses on the characterization of MeSAP10 and its role in response to the Chinese strain of Xpm (XpmCHN11). This investigation offers valuable insights for the functional characterization of SAP gene family members in cassava.
-
Cassava cultivar South China 8 (SC8) obtained from the National Cassava Germplasm Nursery and the Chinese Academy of Tropical Agricultural Sciences, was used in this study. SC8 was cultivated as previously described[23]. Briefly, SC8 stems were planted in a substrate composed of a 1:1 mixture of nutrient soil and vermiculite in a greenhouse. The plants were grown under controlled conditions with a 16 h light and 8 h dark cycle at 28 °C with a relative humidity maintained between 60% and 80%.
Hormone treatment and XpmCHN11 inoculation
-
For phytohormone analysis, the callus was cultured in MS medium, supplemented with 100 μM ABA and JA, respectively. For H2O2 treatments, the callus was cultured in MS medium supplemented with 100 mM H2O2. Samples were collected at 0, 2, 6, 12, and 24 h after treatments. For salinity and drought treatments, the callus tissues grown in pots was subjected to 150 mM NaCl and 20% PEG6000, respectively. Samples were collected at 0, 2, 6, 12, 24, and 48 h after treatments. The SC8 seedlings with consistent growth conditions were inoculated with XpmCHN11 following a previously reported method[23]. The 3rd and 4th leaves were sampled at 0, 3, 6 h, and 1, 3, 6 d after inoculation. All samples were quickly frozen in liquid nitrogen for subsequent RNA isolation.
RNA extraction and quantitative real-time PCR (qRT-PCR) assay
-
Total RNA was extracted from leaves using the RNAplant plus kit (TIANGEN, China), and the RNA was used for cDNA synthesis with SPARKscript ll RT Plus Kit (with gDNA eraser) (Sparkjade, China). qRT-PCR was performed by using ChamQ Universal SYBR qPCR Master Mix (Vazyme, China) following protocols on analytikjena qTOWER3 84G (Jena, Germany). The cycling program was 95 °C for 5 s, 55 °C for 15 s, and 68 °C for 20 s, for 40 cycles. MeTubulin (Manes. 08G061700) and MeUBQ10 (Manes. 07G019300) were selected as the internal reference gene. Relative expression levels of genes were calculated using the 2−ΔΔCᴛ method[24]. Primer sequences are listed in Supplementary Table S5.
Identification and phylogenetic analysis of SAP genes in cassava
-
Cassava SAP genomic, coding, and amino acid sequences were retrieved from Phytozome v12.1 (
https://phytozome.jgi.doe.gov/pz/portal.html ). Conserved domains were identified using HMMER against the Pfam database[25,26]. Putative cassava SAP proteins were identified by BLASTP using the amino acid sequences of Arabidopsis SAP (TAIR 10, E-value ≤ 1e–10) as queries, and A20/AN1 domains were further confirmed using NCBI CDD (www.ncbi.nlm.nih.gov/cdd ). Physicochemical properties, amino acid length/composition, isoelectric points (pIs), and molecular weights (MWs), were calculated using the ExPASy website (https://web.expasy.org ), and subcellular localization was predicted using PSORT (www.genscript.com/psort.html ). Phylogenetic analysis was performed in MEGA 7.0 using the Neighbor-Joining method with 1,000 bootstrap replicates. SAP protein sequences from Arabidopsis, rice, and cassava were downloaded from the Phytozome database (https://phytozome.jgi.doe.gov ).Chromosomal distribution, gene structure, and evolutionary relationship of cassava SAP genes
-
The chromosomal locations of cassava SAP genes were plotted using MapInspect software following the procedure described (
http://mapinspect.software.informer.com ). The gene structures of SAPs were based on gene models downloaded from the Phytozome 12.1 website as described above, and the conserved motifs of SAP proteins were predicted using MEME (http://meme-suite.org/tools/meme ). These characteristics were visualized on the phylogenetic tree using ITOL 4 (https://itol.embl.de )[27]. Gene duplication analysis of cassava SAP gene family and collinearity analysis of orthologs between M. esculenta and A. thaliana as well as between M. esculenta and O. sativa, were performed using MCScanX with default parameters, and the results were visualized using Circos[28,29].Cis-elements analysis of cassava SAP genes
-
To investigate the promoter regions of the SAP genes, the 2 kb genomic DNA upstream sequences of 16 SAP family genes were extracted and subjected to TSSP (
www.softberry.com )[30].Expression patterns of MeSAP genes
-
To analyze MeSAP gene expression across cassava tissues, and after Xpm infection, RNA-seq datasets (Arg7, KU50, and wild W14) were downloaded from the NCBI (Supplementary Tables S2−S4). Reads were mapped using HISAT2[31], and transcripts were assembled and quantified as FPKM (Fragments per kilobase of transcript per million mapped reads) with StringTie using default settings[32]. Differentially expressed genes in mock- and Xpm-infected samples were identified using the DESeq2 R package with a false discovery rate < 0.001, and at least a two-fold expression difference[33]. Heatmaps were generated in TBtools using log2-transformed FPKM values[34].
Construction and disease resistance analysis of CsCMV-mediated MeSAP10 gene-edited plants
-
For construction of the pCsCMV-MeSAP10 vector, the MeSAP10 target fragment was predicted using the VIGS tool (
https://vigs.solgenomics.net ) and cloned into the pCsCMV-NC vector. For pCsCMV2-MeSAP10, the coding sequence (CDS) of MeSAP10 was amplified from cDNA prepared from SC8 cassava leaves and inserted into the pCsCMV2-NC vector[35,36]. Primers are listed in Supplementary Table S5. For inoculation, pCsCMV-NC, pCsCMV-MeSAP10, pCsCMV2-NC, and pCsCMV2-MeSAP10 were introduced into Agrobacterium tumefaciens GV3101 together with the p19 plasmid. Subsequently, agrobacterium was collected and resuspended in infiltration buffer (1 M MgCl2, 0.5 M 2-morpholinoethanesulfonic acid hydrate, and 150 mM acetosyringone, pH 5.6) to an OD600 = 0.6, and then infiltrated into leaves of 4-week-old cassava plants as previously described.Statistical analysis
-
All statistical analyses were conducted using IBM SPSS Statistics version 20 software. One-way ANOVA and Student's t-test were used to determine significant differences, with p < 0.05 considered statistically significant. GraphPad Prism version 8.0.2 was used for generating graphs.
-
Using amino acid sequences of Arabidopsis SAP family proteins as query sequences (E-value ≤ 1e–10) (Supplementary Fig. S1), 16 SAP family proteins were identified from the cassava genome via BLASTP, and their corresponding coding genes were named MeSAP1–MeSAP16 (Fig. 1). Detailed information regarding the MeSAP family genes are shown in Supplementary Table S1. Overall, the length of the amino acid sequences for these SAPs ranged from 125 (MeSAP2) to 293 (MeSAP10). Predictions of isoelectric point (pI) and protein molecular weight (MW) for MeSAPs revealed that pI values ranged from 7.99 (MeSAP4) to 9.37 (MeSAP15), while protein MWs varied between 13.66 kDa (MeSAP2) and 32.42 kDa (MeSAP10), respectively. Predicted subcellular localization indicated that all SAP members were localized in the nucleus (Supplementary Table S1).
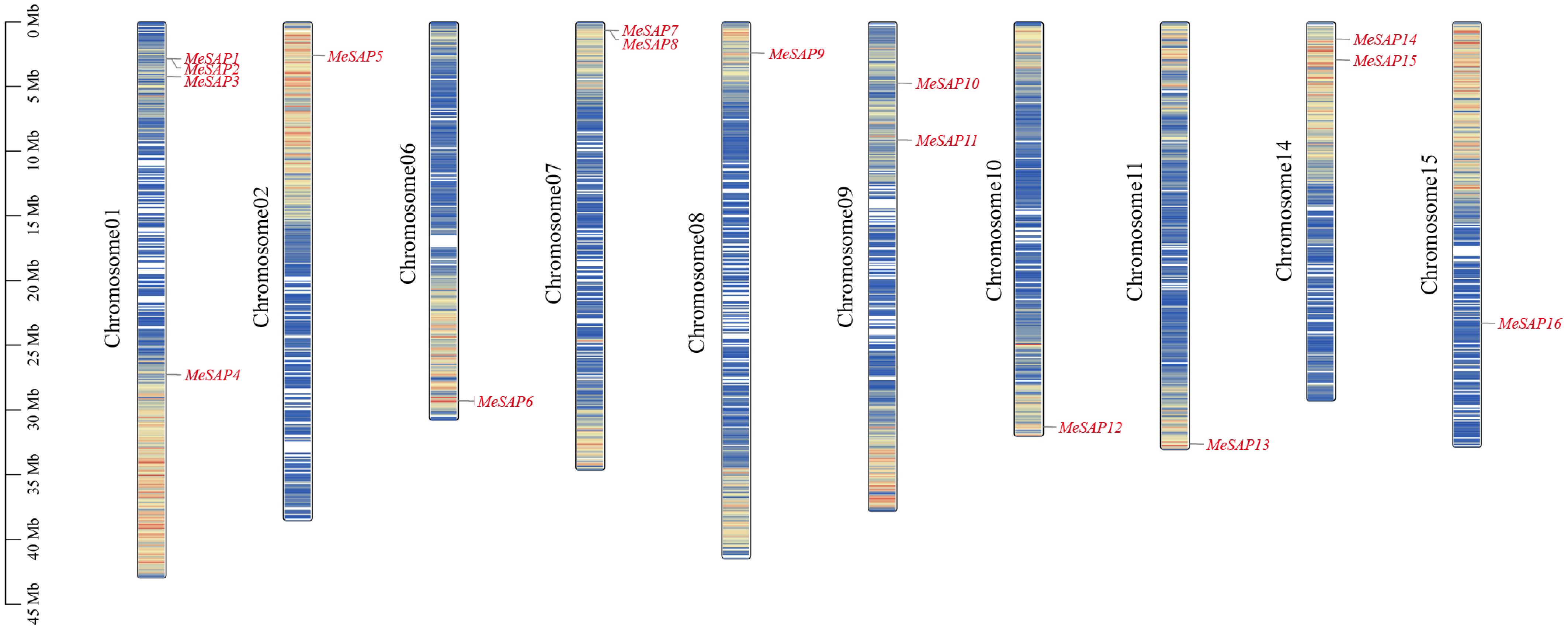
Figure 1.
Distribution of the SAP genes among 10 cassava chromosomes. The scale of the left side represents the length of chromosome displayed in megabase (Mb).
MeSAP1 to MeSAP16 were unevenly distributed across the 10 cassava chromosomes (Chr.). The number of MeSAPs located on a single chromosome varied from one to four, with Chr. 2, 6, 8, 10, 11, and 15 containing one gene each, Chr. 7, 9, and 14 containing two genes, and only Chr. 1 containing four genes. There was no evident positive correlation between chromosome length and the number of SAP family genes.
Gene structure and conserved motifs of MeSAPs
-
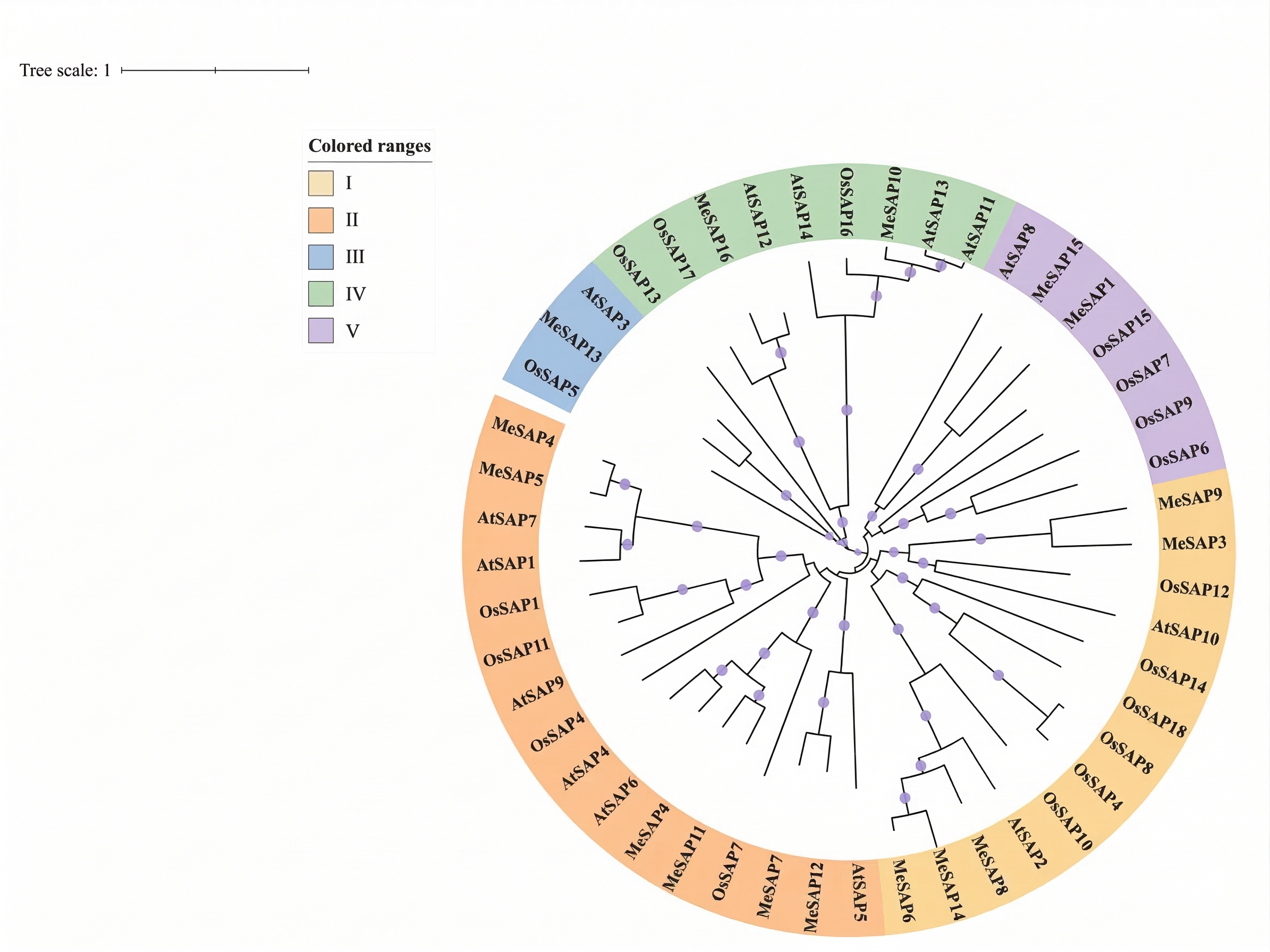
To elucidate the phylogenetic relationship and classification of MeSAPs, a neighbor-joining (NJ) phylogenetic tree was constructed. The tree revealed that the 16 MeSAP proteins clustered into five groups (Supplementary Fig. S1). Among these proteins, 5 SAP proteins were classified into Group I, 6 into Group II, whereas Group III contained only SAP13. Group IV comprised SAP10 and SAP16, and Group V included SAP1 and SAP15. To explore the structural diversity of the cassava SAP family genes, gene structure analyses of the 16 MeSAP genes were conducted based on their evolutionary relationships. Remarkably, all genes were found to lack introns. 14 SAP proteins shared a common structural feature: a highly conserved AN1 and A20 domain, whereas MeSAP10 and MeSAP16 possessed two AN1 motifs (Fig. 2, Supplementary Fig. S2). Interestingly, SAPs containing only a single A20 domain were not identified in cassava, consistent with observations in Arabidopsis, maize, and tomato, but not rice[11,37]. In agreement with alignment and phylogenetic analysis, members of each group exhibited similar patterns of conserved motifs.
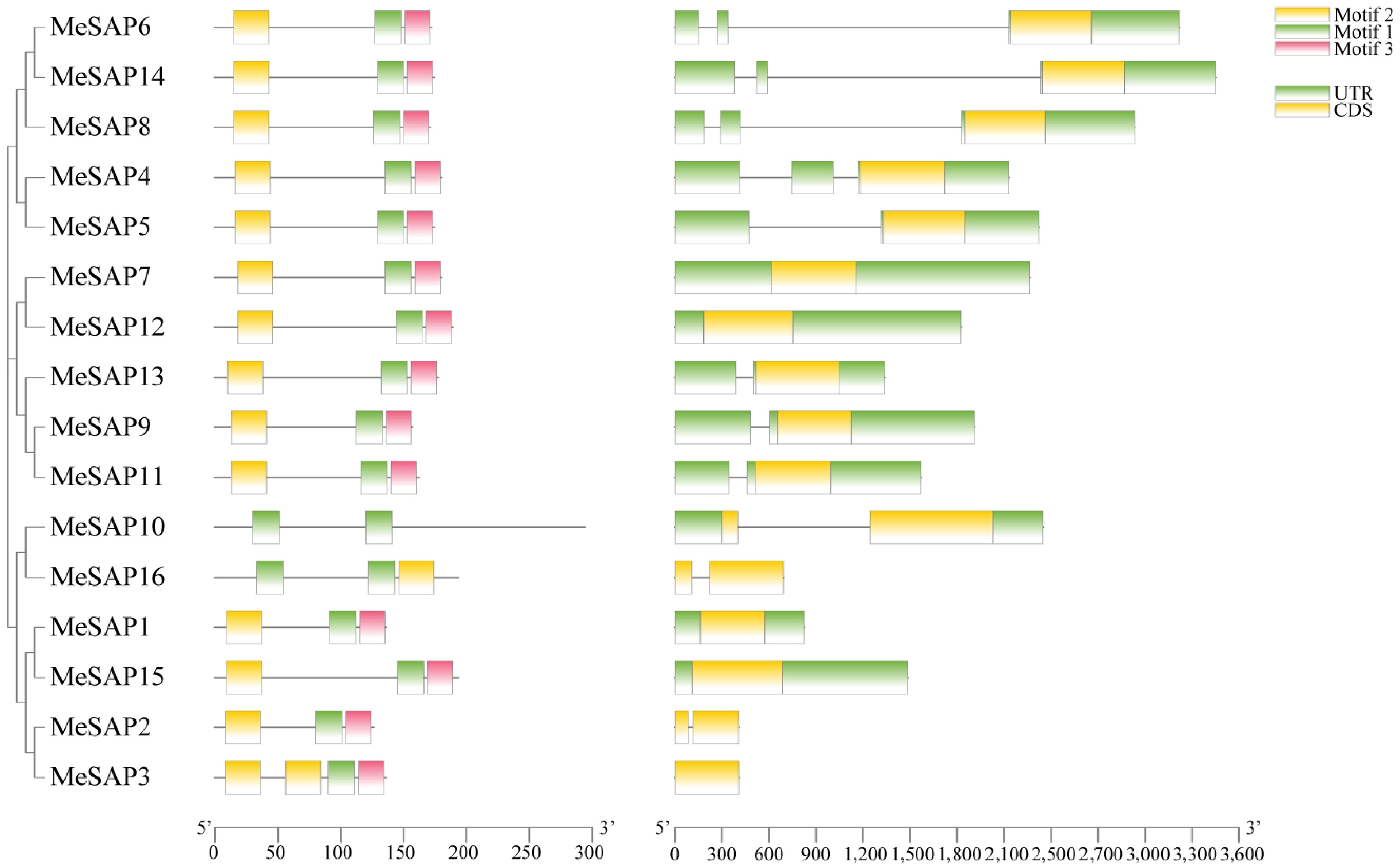
Figure 2.
The structure, clustering, and conserved motif analysis of cassava MeSAP proteins. The left side illustrates the gene structure of the cassava MeSAP gene family. The right side displays the conserved motifs of cassava MeSAP proteins.
Evolutionary analysis of MeSAPs between cassava and other species
-
To further investigate the evolutionary relation of the cassava MeSAP gene family through duplication events, a homologous-block map was constructed (Fig. 3a). The results revealed that 10 MeSAP genes are located in orthologous blocks with corresponding homologous MeSAP genes. These duplicated MeSAP genes were distributed across cassava Chr. 1, 2, 6, 7, 8, 9, 10, 11, and 14. The 12 pairs of MeSAP genes included MeSAP4 and MeSAP5, MeSAP4 and MeSAP15, MeSAP5 and MeSAP14, MeSAP6 and MeSAP7, MeSAP6 and MeSAP12, MeSAP6 and MeSAP14, MeSAP7 and MeSAP12, MeSAP7 and MeSAP14, MeSAP9 and MeSAP10, MeSAP9 and MeSAP13, MeSAP10 and MeSAP13, and MeSAP12 and MeSAP14. MeSAP6, MeSAP7 and MeSAP14 were associated with three syntenic gene pairs, suggesting their potential importance in the evolution of the MeSAP gene family. To further elucidate the phylogenetic mechanisms of the cassava SAP gene family, syntenic maps were constructed using cassava, Arabidopsis, and rice (Fig. 3b, c). The results revealed eight gene pairs with collinearity between cassava and rice, and sixteen gene pairs with collinearity between cassava and Arabidopsis. These results suggested a closer relationship between cassava SAP genes and Arabidopsis compared to rice. Among them, MeSAP6, MeSAP9, MeSAP13 and MeSAP14 had orthologous genes in both rice and Arabidopsis, indicating the existence of these orthologous pairs prior to ancestral divergence. However, orthologous genes of MeSAP4, MeSAP5 and MeSAP10 were only found in Arabidopsis, while orthologous genes of MeSAP11 and MeSAP12 were only found in rice. This result suggests that these orthologous pairs may form after the divergence of dicotyledonous and monocotyledonous plants.
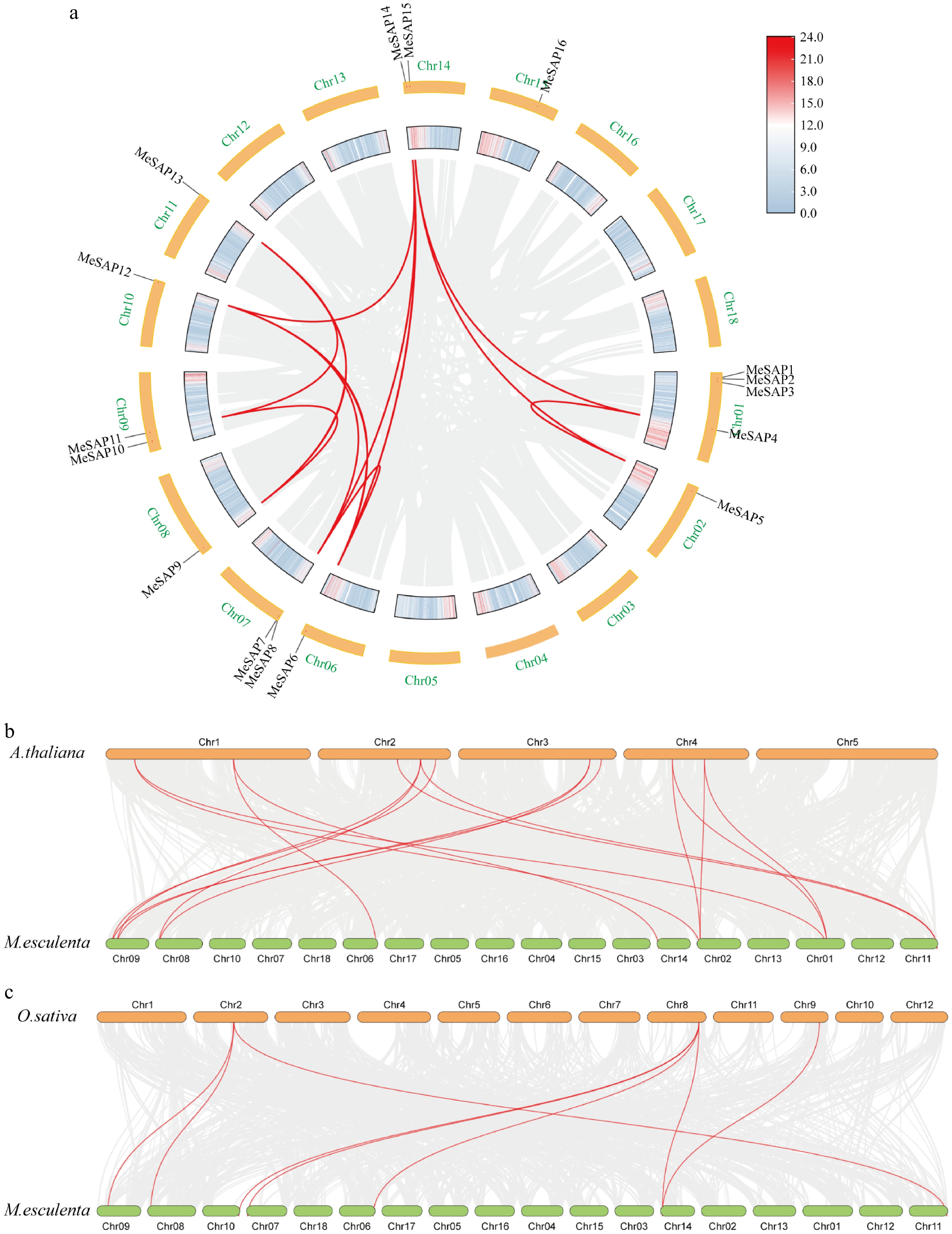
Figure 3.
Duplication and collinearity analysis of SAP genes between cassava, Arabidopsis, and rice. (a) Duplicated MeSAP genes on the cassava chromosomes. (b) Duplicated SAP genes on the cassava and rice chromosomes. (c) Duplicated SAP genes on the cassava and Arabidopsis chromosomes. Gray lines indicate all synteny blocks in the cassava genome, and the red lines indicate duplicated SAP gene pairs.
Analysis of cis-elements in the promoter regions of cassava MeSAP genes
-
To investigate the regulatory mechanisms governing the expression of cassava SAP genes, putative cis-elements within their promoter regions were predicted using the Softberry website (
www.softberry.com ) (Fig. 4). Eight cis-elements associated with stress tolerance, namely MYB element, anaerobic induction element (ARE), wound response elements (WUN-motif), G-box, F-box, MYB binding sites (MBS), WRKY binding sites (W-box), and stress-responsive elements (TC-rich repeats), were identified in the promoter regions of MeSAP genes. Notably, the MYB element was prominently present in most promoters, except for MeSAP13. The ARE, WUN-motif, and G-box were observed in the promoters of the majority of SAP genes, indicating their potential involvement in stress responses, particularly osmotic stresses. However, F-box, MBS, W-box, and TC-rich repeats were detected in only a few cassava SAP gene promoters. Moreover, eight phytohormone-responsive elements were detected in the promoters, including ABRE elements, ERE elements, and CGTCA motifs, which mediate plant responses to abscisic acid (ABA), ethylene (Eth), and methyl jasmonate (MeJA), respectively. Additionally, TCA elements, TGA elements, and GARE-motifs were found in a subset of SAP genes, potentially mediating plant responses to salicylic acid (SA), and gibberellin (GA). These findings suggest that cassava SAP genes may play diverse roles in hormone responses, highlighting their multifunctional nature in regulating stress and hormonal signaling pathways.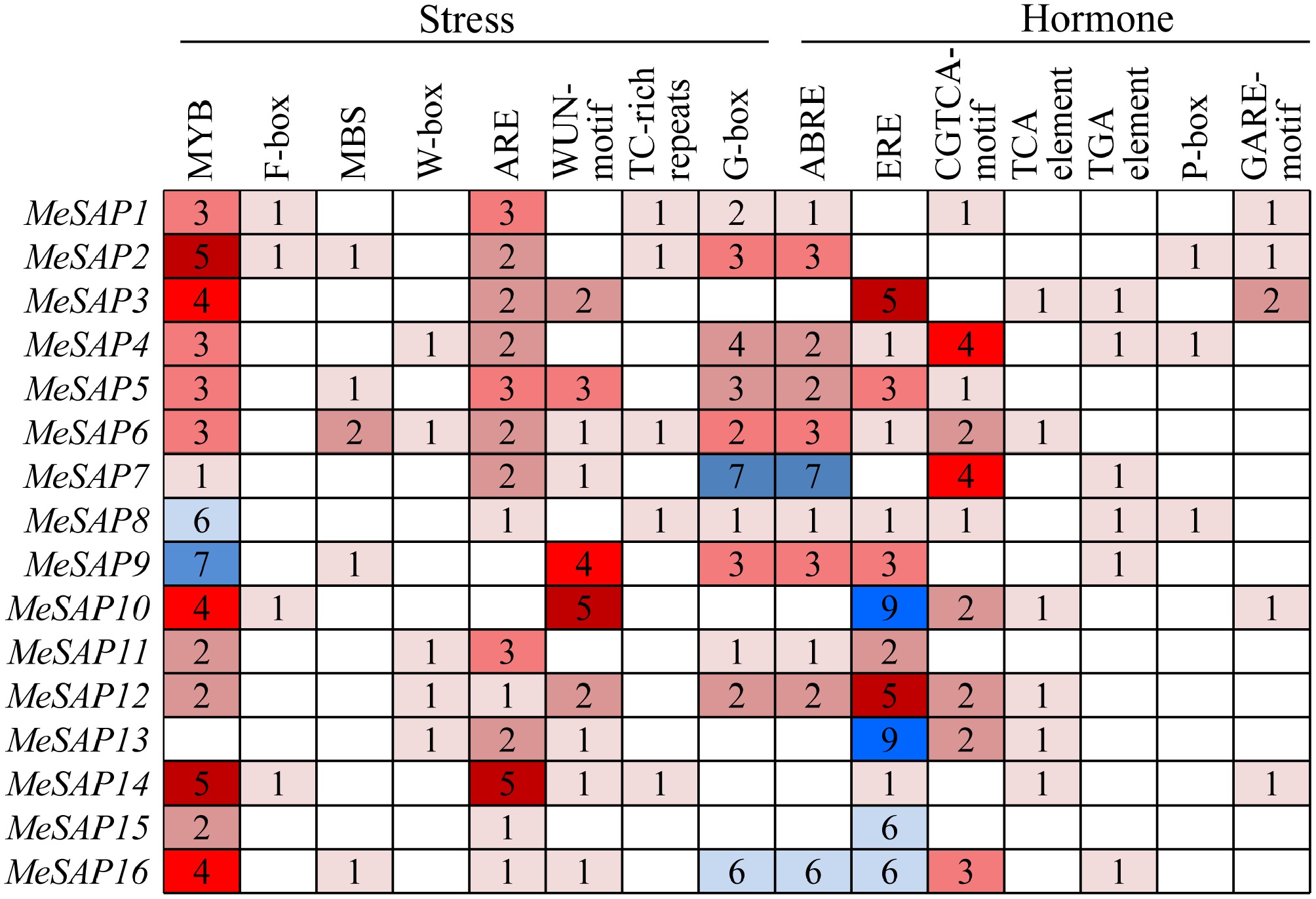
Figure 4.
The cis-elements type and numbers distribution in the promoter regions of cassava MeSAP genes.
Expression profiles of MeSAPs in different tissues
-
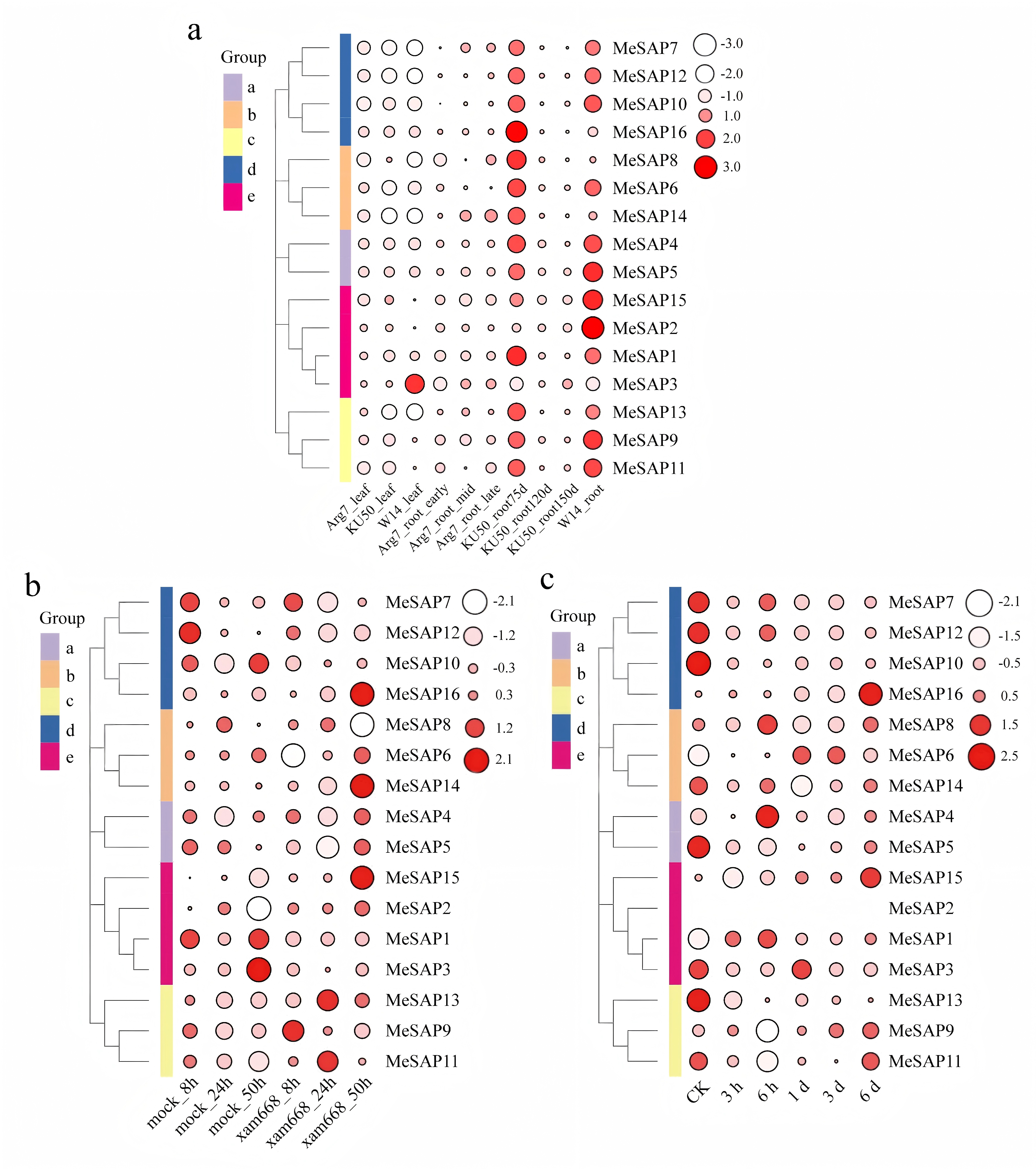
To understand the potential functions of MeSAP genes during cassava growth and development, the expression patterns of cassava SAP genes in leaves and different stages of roots were extracted for analysis based on transcriptome data from cassava varieties W14, KU50, and Arg7 (Supplementary Table S2). The analysis revealed that most MeSAP genes, except MeSAP3, exhibited low expression levels in the leaves of all varieties and in the roots of Arg7 and KU50, at 120 and 150 d. Notably, MeSAP genes, excluding MeSAP2 and MeSAP3 showed high expression during the early stages of root development in KU50. Additionally, MeSAP3, MeSAP8, MeSAP14 and MeSAP16 displayed lower expression levels in the roots of W14 compared with other tissues at different stages of root development (Supplementary Fig. S3a). However, 12 MeSAP genes exhibited higher expression levels in the roots of W14. These findings align with previous studies conducted on other plants, which have reported lower transcription levels of most SAP genes under normal conditions[25,37].
Expression profiles of MeSAPs under different pathogen strains
-
To explore the functional divergence of SAP genes, transcriptome data from cassava treated with different pathogen strains were obtained from the National Center for Biotechnology Information (NCBI) (Supplementary Tables S3, S4). The Fragments Per Kilobase of transcript per million mapped reads (FPKM) values of MeSAP genes infected with Xam668 were utilized to generate a heat map depicting the transcription levels of MeSAP genes[38]. The heat map results revealed that MeSAP14, MeSAP15, and MeSAP16 exhibited higher expression at 50 h post Xam668 inoculation, while MeSAP11 and MeSAP13 displayed high levels of expression at 24 h post-inoculation, which decreased in later stages. Furthermore, MeSAP7 and MeSAP9 were induced by Xam668 in the early stages but exhibited lower expression in later stages (Supplementary Fig. S3b). The expression levels of these genes under Xanthomonas phaseoli pv. manihotis (Xpm) infection was also assessed. MeSAP1, MeSAP4, and MeSAP8 displayed peak expression at 6 h after Xpm inoculation, while MeSAP3 and MeSAP6 showed elevated expression at 1 and 3 d after inoculation. MeSAP15 and MeSAP16 exhibited high expression at 6 d after inoculation. In contrast, MeSAP5, MeSAP7, MeSAP10, MeSAP12, and MeSAP13 remained at low expression levels following Xpm inoculation (Supplementary Fig. S3c).
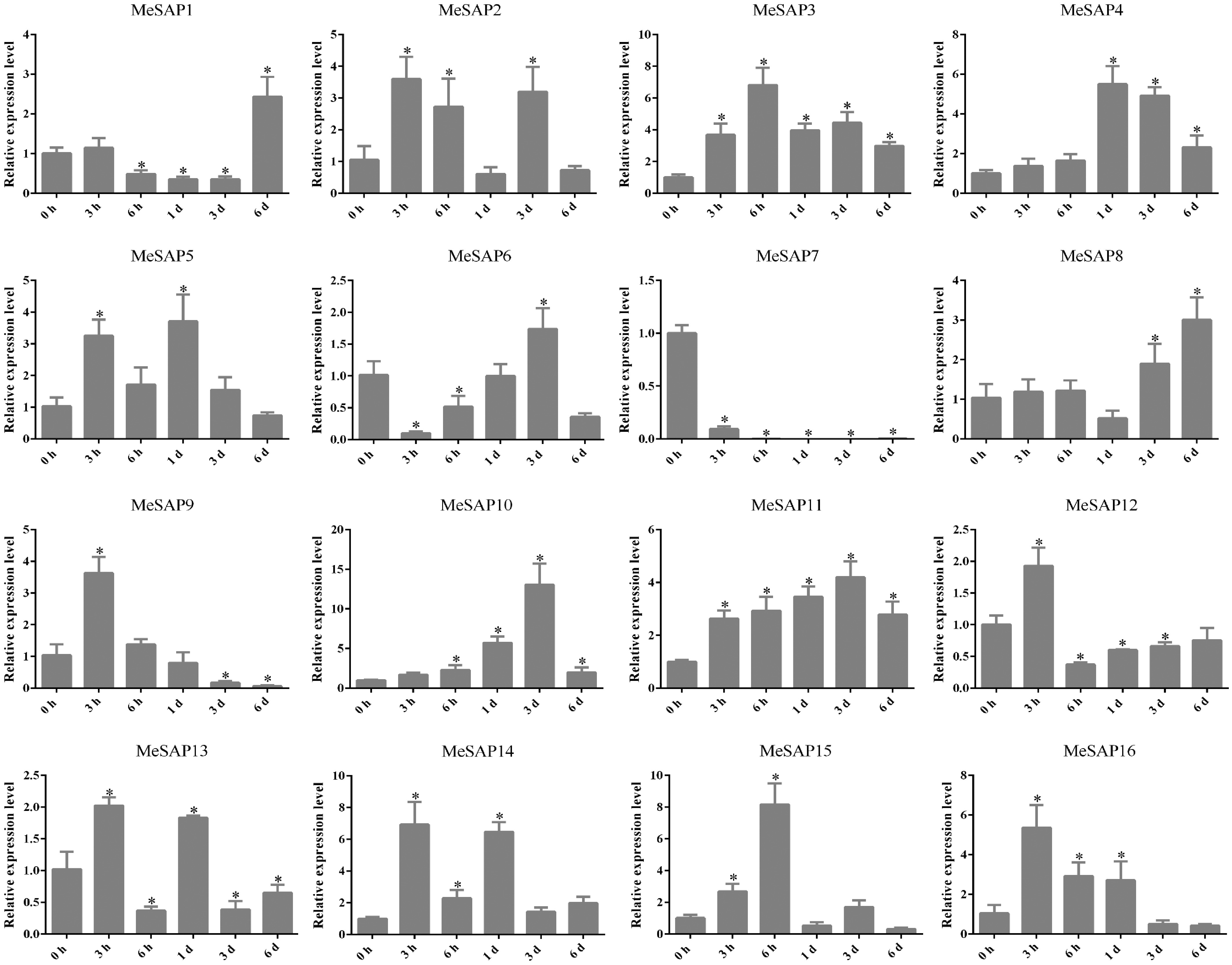
Figure 5.
Expression of cassava SAP genes induced with XpmCHN11. Data is mean ± SE pooling from three biological replicates. * Indicates a significant difference (p < 0.05) between the control and treatments.
qPCR validation of expression profiles of MeSAPs under abiotic stresses
-
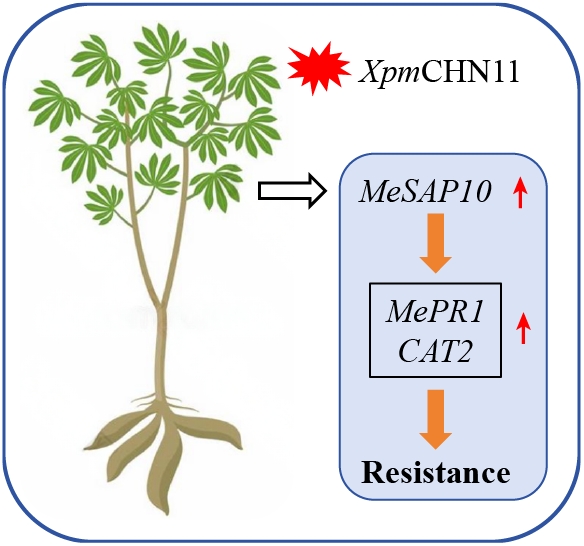
To examine the effects of abiotic stresses and hormonal treatments on MeSAP gene expression, the expression patterns of 13 MeSAP genes were analyzed using qRT-PCR under various conditions, including ABA, H2O2, NaCl, PEG6000, and cold treatments (Fig. 6). Among them, MeSAP10 was notably induced by ABA, H2O2, NaCl, PEG6000 and low temperature, but downregulated by JA treatment. In response to XpmCHN11 infection, MeSAP10 was significantly upregulated, suggesting its potential key role in cassava disease resistance (Fig. 5). These results indicate that MeSAP10 is a key gene involved in cassava responses to both abiotic and biotic stresses.
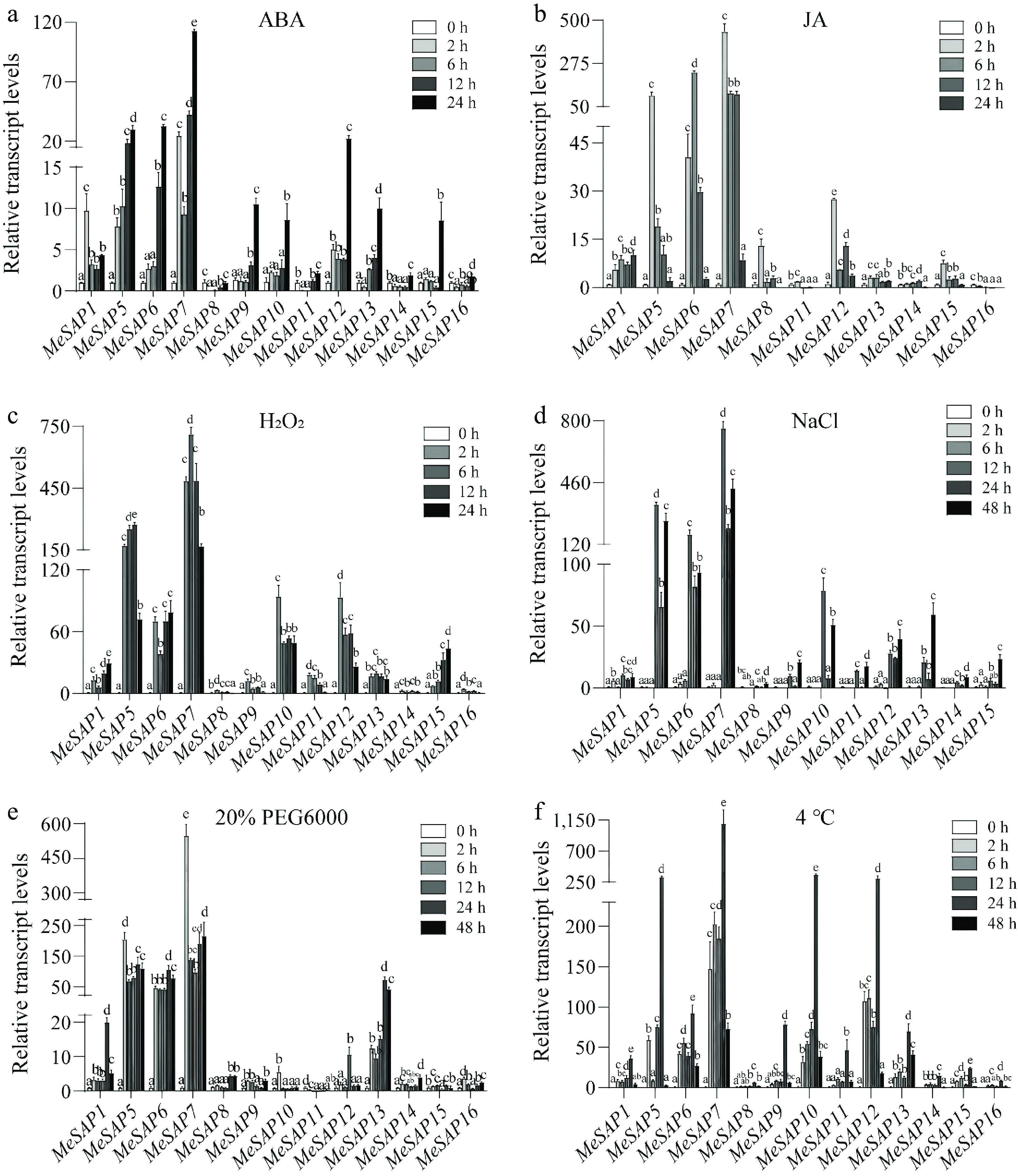
Figure 6.
Expression profiles of the MeSAP gene family under abiotic stresses. Expression patterns of the MeSAP gene family under (a) ABA treatment, (b) JA treatment, (c) H2O2 treatment, (d) NaCl treatment, (e) 20% PEG6000 treatment, (f) 4 °C treatment. Data is mean ± SE pooling from three biological replicates. Statistical significance was assessed by One-way ANOVA. Letters indicate significant differences between groups (p < 0.05).
MeSAP10 improves cassava resistance to XpmCHN11
-
To explore the role of MeSAP10 in response to XpmCHN11 in cassava, we generated MeSAP10-silenced plants using CsCMV-mediated gene silencing technology, with plants infected with the pCsCMV-NC empty vector as a negative control. After 4 weeks, qRT-PCR analysis revealed that the expression level of MeSAP10 in pCsCMV-SAP10 plants was reduced to 20% of that in pCsCMV-NC plants (Fig. 7b). Subsequently, leaves of pCsCMV-NC and pCsCMV-MeSAP10 plants were inoculated with XpmCHN11. The results revealed that silencing of MeSAP10 in cassava plants increased susceptibility to XpmCHN11 infection compared to pCsCMV-NC plants leaves, as evidenced by larger lesions in MeSAP10-silenced plants at 6 d post-inoculation (dpi) (Fig. 7a, c). Moreover, XpmCHN11 growth in MeSAP10-silenced leaves was significantly higher than in pCsCMV-NC plants (Fig. 7d). Additionally, the expression of MeSAP10, MePR1, and MeCAT2 was significantly downregulated in MeSAP10-silenced plants compared to pCsCMV-NC plants (Fig. 7e).
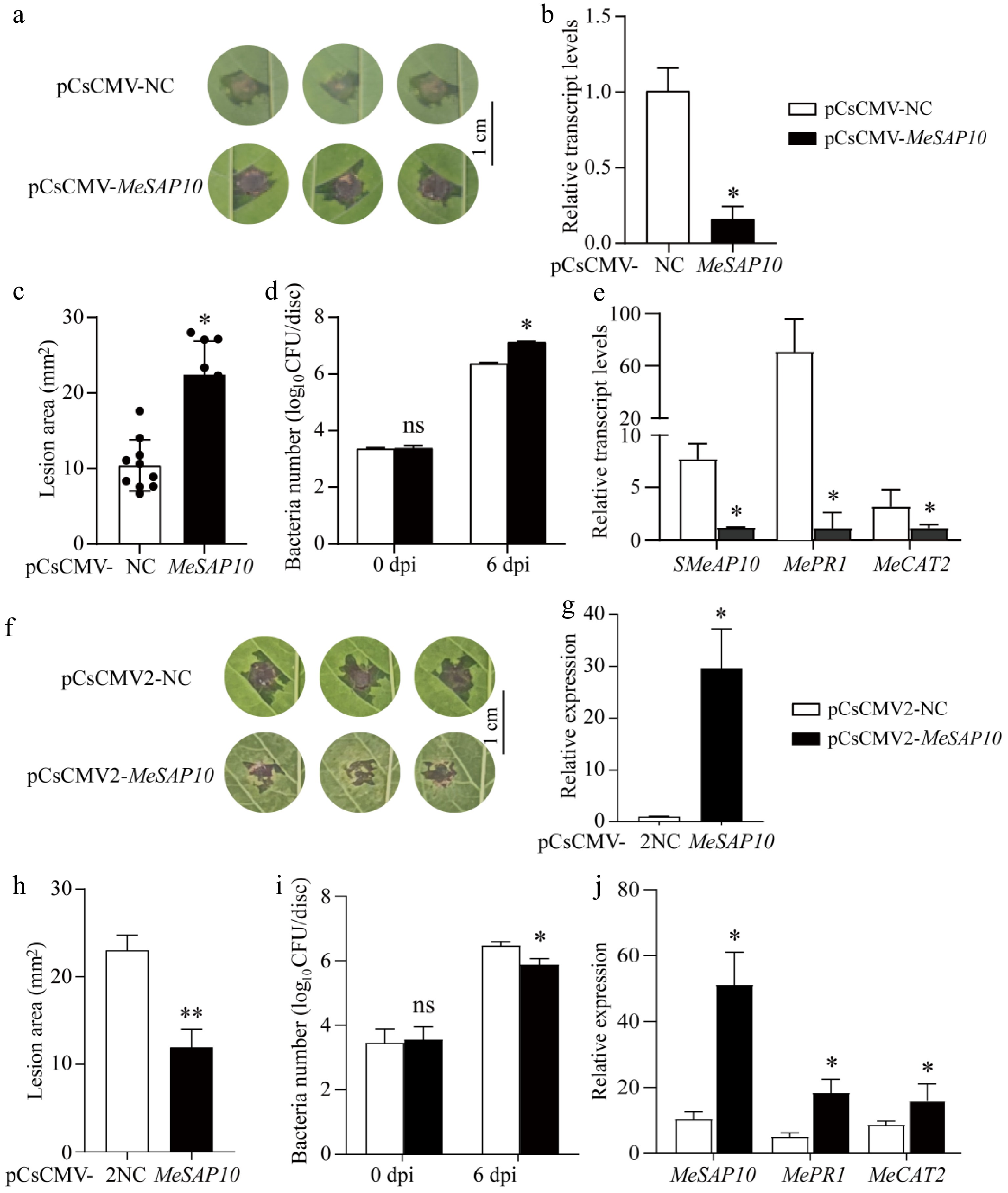
Figure 7.
Functional analysis of MeSAP10 in response to XpmCHN11. (a) Lesion phenotypes of pCsCMV-NC and pCsCMV-MeSAP10 plants after inoculation with XpmCHN11 at 6 d post-inoculation (dpi), scale bar = 1 cm. (b) Silencing efficiency of SAP10 in pCsCMV-NC and pCsCMV-MeSAP10 plants. (c) Lesion area of pCsCMV-NC and pCsCMV-MeSAP10 plants after inoculation with XpmCHN11 at 6 dpi. (d) Bacteria number in leaves of pCsCMV-NC and pCsCMV-MeSAP10 plants after inoculation with XpmCHN11 at 6 dpi. (e) Expression levels of MeSAP10, MePR1, and MeCAT2 in leaves of pCsCMV-NC and pCsCMV-MeSAP10 plants at 3 dpi. (f) Lesion phenotypes of pCsCMV2-NC and pCsCMV2-MeSAP10 plants after inoculation with XpmCHN11 at 6 d post-inoculation (dpi), scale bar = 1 cm. (g) Expression levels of MeSAP10 in pCsCMV2-NC and pCsCMV2-MeSAP10 plants. (h) Lesion area of pCsCMV2-NC and pCsCMV2-SAP10 plants after inoculation with XpmCHN11 at 6 dpi. (i) Bacteria number in leaves of pCsCMV2-NC and pCsCMV2-MeSAP10 plants after inoculation with XpmCHN11 at 6 dpi. (j) Expression levels of MeSAP10, MePR1, and MeCAT2 in leaves of pCsCMV2-NC and pCsCMV2-MeSAP10 plants at 3 dpi. Statistical significance was assessed by Student's t-tests. * Represents significant differences between pCsCMV-NC and pCsCMV-MeSAP10 plants, and between pCsCMV2-NC and pCsCMV2-MeSAP10 plants (* p < 0.05, ** 0.001 ≤ p < 0.01).
Furthermore, we generated MeSAP10-overexpressing plants (designated as pCsCMV2-MeSAP10) via CsCMV-mediated gene overexpression system, with pCsCMV2-NC plants used as the control. qRT-PCR analysis revealed that the expression level of MeSAP10 in pCsCMV2-MeSAP10 plants was 29.68-fold higher than that in pCsCMV2-NC plants (Fig. 7g). Subsequently, leaves of pCsCMV2-NC and pCsCMV2-MeSAP10 plants were inoculated with XpmCHN11. The results demonstrated that, compared with pCsCMV2-NC leaves, SAP10-overexpressing plants exhibited enhanced resistance to XpmCHN11 at 6 dpi, which was manifested by a smaller lesion area on the leaves (Fig. 7f, h). In addition, the content of XpmCHN11 in SAP10-overexpressing plants leaves was significantly lower than that in pCsCMV2-NC leaves at 6 dpi (Fig. 7i). Consistently, the expression levels of MeSAP10, MePR1, and MeCAT2 were significantly upregulated in MeSAP10-overexpressing leaves compared to those in pCsCMV2-NC leaves (Fig. 7j). These results collectively demonstrate that MeSAP10 plays an important role in plant immune responses by positively regulating cassava resistance to XpmCHN11.
-
Proteins containing the A20/AN1 zinc finger domain play crucial roles in plant responses to biotic and abiotic stresses[6,39]. While SAPs are known to function in stress tolerance as ubiquitin ligases, redox sensors, or gene expression regulators, the precise mechanism remains elusive. Recent studies have shown that constitutive expression of SAP genes in various plant species, including Arabidopsis, rice, maize, and soybean enhances tolerance to multiple abiotic stresses[10,16,37,40]. However, SAP genes in cassava have not been fully identified, and their functions in cassava remain poorly understood.
We identified 16 MeSAP genes in cassava, which were categorized into three subgroups through whole-genome screening. Among them, MeSAP10 and MeSAP15 contained two AN1 domains, while the remaining 12 genes had one A20 domain and one AN1 domain. This finding suggests that the A20/AN1 type is the predominant arrangement among zinc-finger family members in cassava, consistent with observations in tomato and Dendrobium officinale[41,42]. All the SAP proteins contained the A20/AN1 type domain, except for MeSAP14, which only had one AN1 domain in cassava. Previous research has indicated that lower organisms, such as Saccharomyces cerevisiae and Aspergillus fumigatus, possess only AN1 zinc finger domains, suggesting the ancient origin of the AN1 zinc finger domain compared to the A20 domain[41,43]. During the evolution of living organisms, gene duplication occurs and is considered a major cause of genome complexity and functional expansion[44]. In cassava, gene duplication mainly occurs on Chr. 1, 2, 6, 7, 10, and 14.
In Arabidopsis, SAP genes have been found to confer defense against pathogens in plants. For instance, AtSAP9 was induced by pathogens, PAMP molecules, and phytohormones, leading to increased susceptibility to non-host pathogen Pseudomonas syringae pv. phaseolicola, indicating its crucial role in basal resistance[21]. Furthermore, an orchid SAP protein, Pha13, and its Arabidopsis homologue, AtSAP5, was reported to serve as an important regulatory hub in plant antiviral immunity[45]. Transcriptome data revealed that eight MeSAP family members were regulated by Xpm, with their expression profiles differing under high-virulence Xpm compared to low-virulence Xpm. Some MeSAP genes, such as MeSAP3, MeSAP15, and MeSAP16, responded to Xam668 and XpmCHN11 inoculation. Notably, MeSAP4 was only induced by XpmCHN11 and not Xam668 (Supplementary Fig. S3c). Conversely, MeSAP9, MeSAP11, MeSAP13, MeSAP14, MeSAP15, and MeSAP16 were markedly induced by Xam668. MeSAP4, MeSAP8, MeSAP15, and MeSAP16 responded to XpmCHN11. In contrast, MeSAP7 was downregulated under Xam668 and XpmCHN11 treatment. We further validated these results using qPCR in cassava leaves treated with XpmCHN11, which showed consistency with the qRT-PCR results, indicating that SAP family members responded to XpmCHN11 infection and may play crucial roles in pathogen defense. Furthermore, some MeSAP family members may respond differently to pathogens. For example, MeSAP10 did not respond to Xam668 but was significantly induced by XpmCHN11 (Fig. 5). Integration of results from different phytohormone and biotic stress assays suggested that MeSAP10 may play an important role in plant resistance, possibly through the H2O2 pathway in response to XpmCHN11.
Previous studies have emphasized the pivotal roles of phytohormones in regulating plant tolerance and resistance. It was observed that the promoters of all cassava SAP genes contain cis-acting elements related to stress and hormone responses, such as drought, low temperature, defense, and ABA responses (Fig. 4). Several SAP genes, including MeSAP4, MeSAP6, MeSAP7, MeSAP10, MeSAP12, MeSAP13, and MeSAP16 were found to contain more CGTCA elements, indicating the presence of cis-elements related to phytohormones. Interestingly, promoters of the SAP genes also contained ERE motifs responsive to ethylene, such as MeSAP3, MeSAP10, MeSAP12, MeSAP13, MeSAP15, and MeSAP16, containing more ERE elements. However, the relationship between SAP family gene expression and ethylene could not be confirmed through qRT-PCR analysis. Furthermore, MeSAP5, MeSAP9, and MeSAP10 contained more WUN motifs, which respond to wounding stress. The potential defense mechanisms of SAP genes against phytohormone stresses and their involvement in pathogen defense through hormone signal transduction warrant further investigation. Subsequent studies are needed to validate these hypotheses and elucidate the regulatory mechanisms underlying the expression of SAP genes under hormone stresses.
In conclusion, we conducted a genome-wide analysis of SAP family members in cassava, identifying a total of 16 SAP genes. A comprehensive characterization of SAP genes was performed, including analyses of gene structures, characteristic domains, and phylogenetic relationships. Moreover, we found that 16 cassava SAP genes identified from cassava were induced by abiotic stresses and pathogen treatments through RNA-seq and qRT-PCR analysis. Additionally, we found that MeSAP10 strongly responded to XpmCHN11 infection, and silencing of MeSAP10 in cassava enhanced its susceptibility to XpmCHN11, while overexpression of MeSAP10 significantly enhances its resistance (Fig. 7). All these results may help us further understand the mechanisms of cassava defense to stress conditions and provide candidate genes for breeding stress-resistant cultivars. Future research should be conducted to study the function of plant SAP family genes under stress conditions.
-
The authors confirm their contributions to the paper as follows: writing−original draft: Wang Y, Li K; visualization: Wang Y, Li K, Niu X, Yang L, Wang H; validation: Wang Y; data curation: Chen Y, Yu X; writing−review and editing: Zheng L, Yang L, Lin M; supervision, conceptualization, project administration: Chen Y. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available from the corresponding author upon reasonable request.
-
Thank you for the support of the National Natural Science Foundation of China (3226150099, 32260468), the Key Research and Experimental Development Program of Hainan province (ZDYF2023XDNY179-7), China Agriculture Research System (CARS-11-HNCYH), and the International Science and Technology Cooperation Program of Hainan Province (GHYF2022005). Thank Professor Wentao Shen from Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences for providing the vector pCsCMV-NC and pCsCMV2-NC.
-
The authors declare that they have no conflict of interest.
-
Received 15 January 2026; Accepted 3 March 2026; Published online 18 March 2026
-
# Authors contributed equally: Yixuan Wang, Mingzhi Lin, Ke Li
- Supplementary Fig. S1 Phylogenetic tree of cassava, Arabidopsis and rice SAP gene families encoded A20/AN1 zinc-finger proteins. The cassava SAP proteins could be classified into five distinct groups (I–IV).
- Supplementary Fig. S2 Multi-sequence alignment of cassava SAP proteins.
- Supplementary Fig. S3 Expression profiles of SAPs in RNA-seq. (a) Expression profiles of SAPs in different tissues based on RNA-seq. (b) Expression profiles of SAPs under Xam668 infection based on RNA-seq. (c) Expression profiles of SAPs under Xam infection based on RNA-seq. Color and circle size indicates the expression patterns of the candidate genes.
- Supplementary Table S1 The information of cassava SAP genes.
- Supplementary Tables S2 Description of transcript datasets of different tissue used for expression analysis.
- Supplementary Tables S3 Description of transcript datasets of cassava under Xam668 infection used for expression analysis.
- Supplementary Table S4 Description of transcript datasets of cassava under Xam infection used for expression analysis.
- Supplementary Table S5 Primers used in this study.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Y, Lin M, Li K, Chen Y, Zheng L, et al. 2026. MeSAP10 plays an important role in cassava disease resistance via integrated genomic screening and functional analysis. Tropical Plants 5: e007 doi: 10.48130/tp-0026-0007 

MeSAP10 plays an important role in cassava disease resistance via integrated genomic screening and functional analysis
- Received: 15 January 2026
- Revised: 01 March 2026
- Accepted: 03 March 2026
- Published online: 18 March 2026
Abstract: Stress-associated proteins (SAPs) are A20/AN1 zinc-finger proteins that participate in plant growth and development, and in responses to diverse stresses. Although SAP genes have been studied in many plants, their functions in cassava remain largely unexplored, particularly those involved in pathogen resistance. In this study, 16 MeSAPs with conserved zinc-binding motifs were identified in cassava. The phylogenetic analysis of MeSAP proteins and their orthologs from Arabidopsis and rice indicated that they can be classified into five groups. Analyses of gene structure, conserved motifs, and chromosomal distribution indicated relatively high conservation within the MeSAP gene family. Numerous elements related to stress and plant hormones were identified through prediction of the cis-acting elements in the MeSAP gene promoters. Moreover, transcriptome sequencing and quantitative real-time PCR demonstrated that MeSAP10 was significantly upregulated under XpmCHN11 infection. Functional characterization demonstrated that silencing MeSAP10 increased cassava susceptibility to XpmCHN11, accompanied by downregulation of pathogenesis-related genes MePR1 and MeCAT2. In contrast, MeSAP10 overexpression conferred enhanced disease resistance with simultaneous upregulation of the defense-related genes, MePR1 and MeCAT2. These findings provide new insights into the SAP gene family in cassava, and identify MeSAP10 as a valuable target for breeding disease-resistant varieties.
{kind=link}
{kind=link}