








-
Prunus mume, known as the Japanese apricot, Chinese plum, or Mei, is a deciduous tree. It is valued for both its beauty and fruit. Originating in China, it has been grown in East Asia for centuries[1]. This tree is popular for its fragrant blossoms that appear in mid- to late-winter. Its flowers add much-needed colour when other plants are dormant. Japanese apricot typically blooms from late January to March, making it one of the earliest flowering ornamental trees[2−6]. Many cultivars exist, featuring single and double flowers in white, pink, and red, for aesthetic diversity[1,3,7]. The tree’s graceful branches make it ideal for gardens and bonsai art. Its fruit, a small yellow-green to yellow drupe, ripens in early summer. The fruit is used in traditional foods, drinks, and medicine throughout East Asia[1,8]. The timing of flowering in Prunus mume is tightly linked to both fruit yield and fruit quality. Early blooming can enhance ornamental beauty but may expose flowers to late winter frosts. This can lower fruit set and quality[9,10]. Genetic studies have identified key regulators that control flowering time, dormancy release, and chilling requirements, which affect yield through proper timing[11−14]. Cultivars suited to different climates show various chilling and heat requirements[15]. These are measurable and linked to stable yields across environments[15−17]. The early flowers attract visual interest and have cultural meanings tied to seasonal change, particularly in traditional gardens and public spaces. The length and abundance of blooms depend on the tree’s genetics and climate adaptation, influencing winter and spring aesthetics[18,19]. Breeding efforts often focus on creating cultivars that match local phenology. This approach ensures maximum ornamental impact year after year while enhancing fruit production, as the edible fruits are highly valued in East Asia[20].
-
The development of flower buds progresses through distinct stages, though these phases are not as easily visible. Flowering is primarily external and readily observable. These two developmental mechanisms are associated with seasonal dormancy in perennial deciduous trees like Prunus mume[3,21−23]. Dormancy enables floral buds to endure harsh winter conditions and to bloom only when spring weather becomes favourable. It is typically divided into four phases: paradormancy, endodormancy, ecodormancy, and dormancy release. Dormancy generally begins in early autumn, around late September to October, when bud growth ceases, and metabolic activity slows. During endodormancy, buds do not respond to external growth signals and require extended chilling for release[23−25]. The transition from endodormancy to ecodormancy happens once the chilling requirement is satisfied, which can vary from 26.3 to 75.7 chill units depending on the genotype[17,26]. After that, heat accumulation and hormonal regulation control the dormancy phases that lead to flowering. These processes work together with a complex genetic network to guide bud development toward blooming.
Hormonal shifts, particularly involving ABA and GA, regulate both the induction and release of dormancy[27,28]. At the genetic level, key regulators MCM1-AGAMOUSDEFICIENS Serum Response Factor box (MADS-box) gene families play a central role in coordinating floral organ differentiation and the cycling of dormancy in response to environmental signals and internal rhythms[29,30]. During dormancy, bud metabolic activity is significantly reduced, with declines in glycolysis, sucrose degradation, and cell wall metabolism; these processes are reversed upon dormancy release[31]. The upregulation of DAM genes is associated with the maintenance of dormancy. In contrast, increased expression of SVP genes is linked to dormancy release and the initiation of floral differentiation[32,33].
The initiation of floral buds in Prunus mume follows a defined sequence from bud setting to full bloom, governed by both genetic and physiological factors. This process encompasses several discrete developmental stages, significantly modulated by the interaction between dormancy regulators and environmental cues such as temperature and photoperiod[34,35]. Flower bud differentiation in Prunus mume occurs sequentially from late spring to early summer, with this differentiation concluding before the onset of deep dormancy[1,3].
This development can be categorized into eight identifiable stages: S1, the undifferentiated stage where the bud remains vegetative; S2, flower primordium formation, establishing floral meristem identity; S3, sepal initiation, when sepal primordia differentiate; S4, petal initiation, marking the beginning of floral organogenesis; S5, stamen initiation, with the stamen primordia emerging; S6, pistil initiation, characterised by the formation of carpel and pistil primordia; S7, ovule development, during which ovule differentiation occurs within the pistil; and S8, anther development, finalising the process with anther and pollen formation before dormancy (Fig. 1).
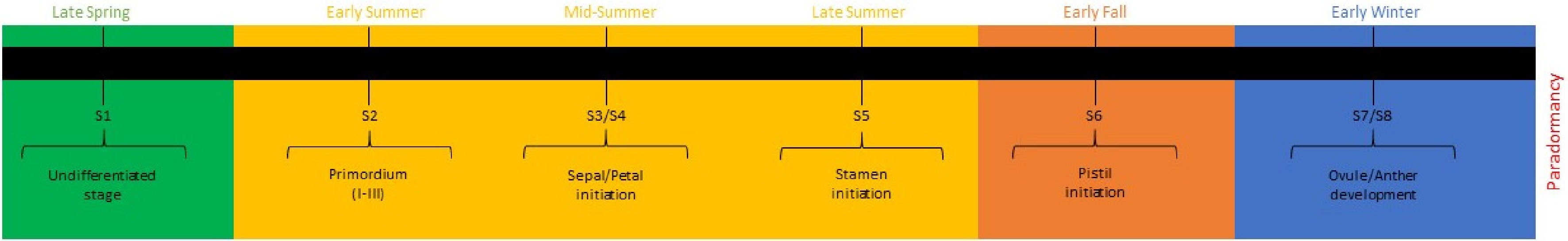
Figure 1.
Timeline of bud initiation and development in Prunus mume.
-
Prunus mume is highly valued for its flowers and fruit, regardless of the cultivation purpose. The timing of flowering is a key parameter as it directly impacts fruit production, enhances ornamental value, and supports breeding programs aimed at improving desirable traits. Growers often encounter challenges related to climate variability, particularly when fluctuating or unpredictable temperatures disrupt chill accumulation during dormancy, or expose blooms to damaging late frosts[36,37]. Early blooming cultivars may suffer flower damage if a late frost occurs, resulting in reduced fruit set and economic loss. Additionally, mismatches between flowering time and optimal pollination periods, caused by asynchronous flowering or environmental stress, can hinder effective fruit production[38−41].
Pistil abortion is another issue, leading to underdeveloped or absent pistils, which reduce fruit set and quality. Incomplete flowers can result in yield losses of 50%–80% and poorer fruit texture[4,42,43]. Furthermore, susceptibility to pathogens during flowering and leaf development complicates crop management[44,45]. Therefore, careful cultivar selection, breeding for resilience to climatic stresses, and improved management practices are essential to mitigate frost damage and ensure synchronised flowering and pollination for the horticultural industry.
-
Research on flowering in Prunus mume employs a variety of complementary approaches, ranging from field observations to advanced molecular biology techniques[46]. Each method offers valuable insights into the complex regulation of flowering time and its environmental interactions. Long-term field observations and phenological data collection have been extensively used to monitor flowering timing and developmental stages under natural conditions[47]. These records provide invaluable baseline information on the variability of blooming times across cultivars and environments, revealing patterns linked to climate factors. Such empirical data have facilitated the categorisation of early, mid, and late-blooming groups, guiding breeding and cultivation strategies. Studies have shown that floral buds respond to temperature during dormancy, correlating bloom times with climate adaptation[48]. Controlled environment experiments, whether in growth chambers or greenhouses, allow for precise manipulation of abiotic factors like temperature, photoperiod, and chilling duration to examine their effects on dormancy release and flower bud development. Research on cold response proteins and metabolic shifts during flowering at low temperatures in Prunus mume illustrates how controlled conditions can reveal adaptation mechanisms[49]. Additionally, advancements in molecular approaches are enhancing our understanding of the genetic and regulatory networks that control flowering. Genome-wide association studies (GWAS) on numerous cultivars have identified quantitative loci (QTL) associated with flowering time[6,50,51]. Transcriptomic profiling has revealed candidate genes and regulatory pathways, while epigenetic and metabolomic analyses have provided further insights into colour formation and developmental regulation[52,53]. Combining these approaches with bioinformatics enhances our understanding and prediction of flowering phenology. Computational modelling that integrates temperature accumulation with dormancy phases improves bloom date forecasts, aiding orchard management and addressing industry challenges.
-
Phenological models are essential for predicting bud break and flowering times in stone fruit, based on environmental cues such as temperature and photoperiod[14,20]. Several models have been developed for Prunus species such as peach, sweet cherry, almond, and Japanese apricot as well[54−57]. Commonly used models include chilling requirement models that quantify the cold exposure necessary to break dormancy, followed by forcing models that accumulate temperature units (growing degree days/hours) to predict the timing of bud break and flowering. Dynamic chill models have gained importance as they account for chilling negation caused by warm spells, leading to more accurate dormancy break estimations[58,59]. In Prunus mume, these models have effectively captured general trends in blooming dates across different cultivars and climates[17]. However, challenges remain, such as an incomplete understanding of endodormancy break mechanisms and variability among genotypes and microclimates that the models may not fully address. The sensitivity of flower bud development to insufficient winter chilling and spring heat spells can impact model accuracy. Additionally, genetic controls and environmental plasticity add complexity that existing temperature-based models might not adequately consider. Recent advancements highlight the importance of incorporating multiple interacting factors, including chilling, forcing, and photoperiod, while also accounting for genetic variation and phenotypic plasticity[20]. Experimental studies have shown species- and provenance-specific responses to these factors, which can enhance parameterization[60]. Models are increasingly integrating genetic and transcriptomic data, allowing for predictions at both molecular and population scales. Climate change projections indicate that shifts in winter chilling and forcing conditions will alter phenology and suitable cultivation zones of Prunus mume and related species[37,48,61]. Some models simulate range shifts and phenological stages to inform breeding and adaptation strategies. Efforts are also underway to improve predictions through non-linear heat accumulation and dynamically updating chilling metrics to bolster robustness in warming scenarios[61−65].
-
Chilling, heat accumulation, and photoperiod are the primary environmental regulators that facilitate the transition from dormancy to flowering in Prunus mume[66]. Flower buds require adequate exposure to low winter temperatures to release endodormancy and allow the initiation of flowering. Research shows that chilling induces molecular changes, including the inhibition and degradation of the PmFRL3 protein and modulation of MADS-box genes, which coordinate dormancy release and bud break[15,67]. Transcriptome analyses further reveal that cold exposure upregulates CBF expression while repressing flowering inhibitors, thereby activating floral developmental pathways. Cultivars exhibit differing chilling requirements, contributing to variations in flowering time and adaptation to climatic conditions[68,69]. Once dormancy is released, heat accumulation accelerates floral bud development, leading to blooming. GA signalling plays a central role in this process, with REPRESSOR OF GA-Like 2 (RGL2) interacting with flowering regulators to facilitate the transition[70,71]. While initial low temperature exposure can delay flowering, subsequent warming stimulates growth and activates pathways and metabolic processes necessary for flower maturation[15,72]. Molecular studies indicate that alterations in sugar metabolism and hormone regulation during warming periods support both flower development and the production of aroma compounds[30,73]. Photoperiod serves as an additional regulatory signal, influencing the timing of flowering through its effects on circadian and flowering-related gene expression. Although less extensively studied in Prunus mume compared to other Prunus species, photoperiod sensitivity likely interacts with temperature cues to refine flowering time. SQUAMOSA PROMOTER BINDING PROTEIN (SBP), FT, and the NAC have been implicated in this response, integrating environmental signals to promote the flowering transition[74,75]. Other environmental factors, such as water availability, carbon dioxide concentration, and abiotic stress, also affect flowering by altering physiological processes and stress responses. Water stress can delay flowering and reduce flower quality, while elevated CO2 may influence photosynthesis and carbohydrate partitioning. Furthermore, studies indicate that UDP-glycosyltransferase genes in Prunus mume respond to abiotic stress, suggesting molecular connections between stress adaptation and floral development. Light intensity and spectral quality further modulate leaf physiology, thereby indirectly affecting the capacity of the plant to flower[76].
-
Environmental signals influence flowering externally, while internal physiological regulation is primarily driven by hormones. Hormonal signalling, interactions among phytohormones, and corresponding metabolic changes collectively determine the timing of floral induction and development. GA were among the first hormones recognized as promoters of flowering, aiding in bud break and floral development[27]. Its action is antagonised by aspartic acid–glutamic acid–leucine–leucine–alanine (DELLA) proteins, which act as its repressors[70]. Conversely, ABA primarily maintains dormancy, and its reduction under chilling conditions is crucial for releasing dormancy and initiating flowering. CK encourages cell proliferation and floral meristem activity, stimulating shoot growth and blooming by upregulating flowering-related transcription factors during the inductive phase. IAA governs polarity and vascular development in floral organs, shapes inflorescence architecture, and indirectly supports flowering by influencing apical dominance and resource allocation[28,77]. The flowering regulation involves extensive hormone interactions, notably ABA and GA antagonism, which facilitates floral induction. Ethylene increases ABA sensitivity during dormancy, while cytokinins work synergistically with GA to enhance cell cycle progression in developing floral buds. Auxin also plays a role in regulating vascular connections and resource mobilization, often collaborating with cytokinins during the transition from vegetative to reproductive growth. Additionally, auxin signalling through auxin response factors (ARFs) and brassinosteroid pathways involving lateral organ boundaries domains (PmLBD3) fine-tunes growth and developmental transitions[78,79]. These hormonal networks and their interactions are supported by precise gene regulation to ensure the transition from vegetative to reproductive growth. The florigen gene, conserved across herbaceous and perennial species, acts as a central mobile signal that moves from leaves to the shoot apex to promote floral induction by activating downstream targets. SOC1 integrates various pathways, including vernalization and gibberellin signalling, to upregulate floral meristem identity genes, with PmSOC1-like homologs promoting early flowering when overexpressed in model systems[80]. LFY serves as a master regulator of floral meristem identity, directly binding to the promoters of organ identity genes to initiate inflorescence development[81]. Its expression is upregulated in response to FT and other pathway integrators, a process also observed in perennial trees like Prunus mume. APETALA1 (AP1) specifies floral organ identity within the classical ABC model, promoting sepal and petal formation while repressing vegetative traits. The activation of AP1 signifies a commitment to the floral program, often occurring alongside LFY and SOC1 as pathways converge. These key players are conserved across angiosperms but exhibit species-specific adaptations in Prunus mume, where genome-wide transcriptome analyses highlight their role in early blooming phenotypes. Recent studies further indicate that PmNAC32 antagonizes PmNAC90 through transcriptional repression and protein–protein interaction, while PmNAC90 delays floral induction by activating PmTFL1 and the auxin glycosyltransferase PmUGT84B[74,75]. Collectively, these findings suggest that a complex regulatory module involving NAC transcription factors, floral integrators, and hormonal signalling precisely tunes the timing of floral transition in Prunus mume, providing a mechanistic explanation for its characteristic early blooming behaviour (Fig. 2).
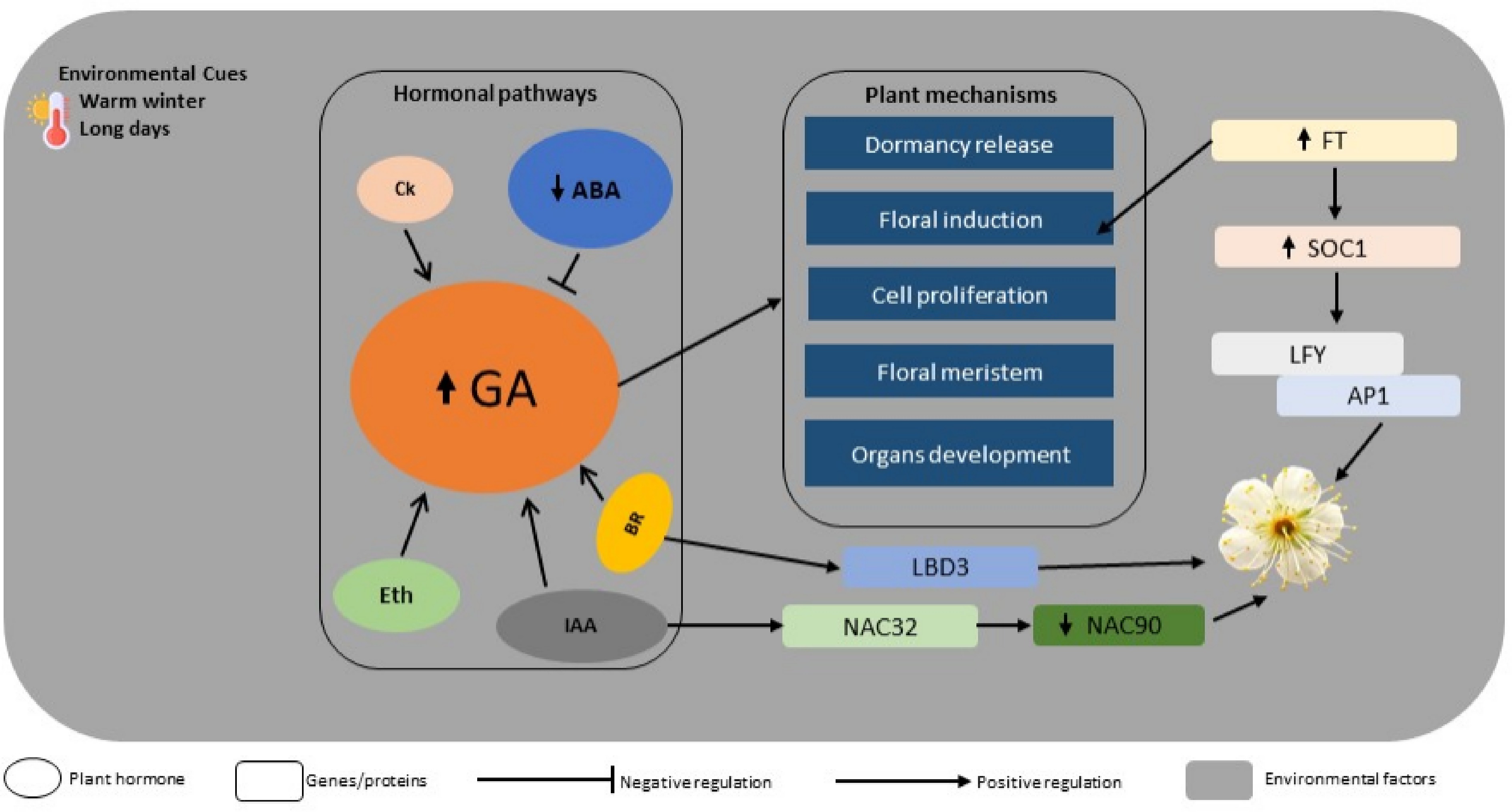
Figure 2.
Molecular regulation of flowering in Prunus mume.
-
Over the past 15 years, significant research has advanced our understanding of flowering time regulation in Prunus mume, encompassing the molecular and physiological processes that govern the transition from dormancy release to bloom. Key molecular pathways have been identified, including the roles of MADS-box genes like PmSVPs and PmDAM in integrating environmental and hormonal signals for bud dormancy regulation and floral induction. Genome-wide and transcriptomic studies have identified numerous candidate genes and quantitative trait loci linked to blooming phenology, facilitating molecular breeding strategies for both ornamental and fruit cultivars. The diversity in chilling requirements and the genetic framework underlying bloom time have been clarified, supporting climate adaptation and sustainable cultivation. Despite these advancements, several knowledge gaps persist. The precise integration of hormonal crosstalk, metabolic pathways, and epigenetic regulation during dormancy-to-flowering remains incompletely characterized. Functional validation of many candidate genes is still needed, and the mechanisms underlying genotype-environment interactions are only partially understood. While transcriptomic and association studies have identified key players, the connections between physiological processes and environmental cues at the whole-plant and field scales require further exploration.
This review focuses on Prunus mume as a model deciduous tree, while also briefly comparing it to evergreen fruit species to highlight differences in regulatory mechanisms. Unlike deciduous trees, evergreen fruit trees like litchi (Litchi chinensis) do not experience true endodormancy, which means they lack the strict chilling requirements typical of temperate deciduous perennials. Litchi serves as a valuable example, as recent studies have identified MADS-box genes that share functional similarities with those in Prunus mume. The litchi LcSVP2 gene, a homolog of the SVP/DAM family, is highly expressed in terminal buds during growth cessation and downregulated during active growth, similar to the expression patterns of PmDAM genes in plum[82]. Overexpressing LcSVP2 in Arabidopsis delays flowering, suggesting conserved repressor functions across both deciduous and evergreen systems. Hormonal regulation in litchi reveals both similarities and key differences. Like Prunus mume, ABA promotes dormancy and enhances LcSVP2 expression, while gibberellin suppression through paclobutrazol (PBZ) treatment encourages flowering by shifting the GA/ABA balance[82,83]. In contrast to deciduous trees, where chilling is a primary factor, litchi flowering is initiated by cold winter temperatures and water stress, leading to a dormancy-like quiescence without true endodormancy[84,85]. This distinction highlights the evolutionary adaptations of Prunus mume and similar deciduous trees, which have developed robust chilling-dependent dormancy mechanisms to endure winter and synchronize flowering with favourable spring conditions.
While comparative analyses with evergreen trees or other perennials could provide broader insights into different regulatory strategies, such comparisons are beyond the scope of this review, which aims to consolidate knowledge specific to Prunus mume as a basis for future research. Future studies should incorporate physiology, high-resolution genomics, epigenetics, and systems biology to unravel the complex regulatory networks governing flowering. Comparative research between deciduous and evergreen perennials could shed light on how conserved genetic modules, such as MADS-box genes, have been adapted for various strategies. This work would enhance our understanding of dormancy evolution and inform breeding programs aimed at developing cultivars with tailored phenology for changing climates. In summary, flowering in Prunus mume is intricately regulated by a range of environmental cues, where chilling plays a critical role in dormancy release, warm temperatures promote bud growth, photoperiod provides regulatory inputs, and additional stress factors influence developmental and metabolic pathways. Understanding these factors at both the molecular and physiological levels is vital for breeding and managing cultivars adapted to changing climates.
-
The authors confirm their contributions to the paper as follows: contributed to conceptualization by developing and discussing the overall concept of the manuscript: Segbo S, Gao Z; writing − original draft by preparing the initial draft, carried out writing − reviewing and editing and project administration by assembling the final version of the manuscript: Segbo S; contributed to writing − review and editing through detailed and critical revision of subsequent drafts: Gao F, Shi T, Gao Z. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
-
This study was supported by the National Natural Science Foundation of China (32372670), the 'JBGS' Project of Seed Industry Revitalization in Jiangsu Province (JBGS [2021] 019), the Postgraduate Research and Practice Innovation Program of Jiangsu Province, China (KYCX23_0798), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Segbo S, Gao F, Shi T, Gao Z. 2026. From dormancy release to bloom in Prunus mume. Fruit Research 6: e023 doi: 10.48130/frures-0026-0015 

From dormancy release to bloom in Prunus mume
- Received: 12 January 2026
- Revised: 17 April 2026
- Accepted: 11 May 2026
- Published online: 12 June 2026
Abstract: Widely cultivated for ornamental and fruit tree purposes in East Asia, Prunus mume Sieb. et Zucc. is best known for its early flowering in late winter to early spring. The timing of flowering is an interesting parameter that affects fruit yield, fruit quality, and ornamental value, but it is strongly influenced by environmental changes. This review brings together recent progress in understanding how flowering time in Prunus mume is controlled by environmental signals, plant hormones, and gene regulation. Bud dormancy, which ensures survival through winter, moves through several stages before flowering, and is primarily regulated by chilling accumulation and subsequent heat requirements, while photoperiod plays a secondary modulatory role. Hormones such as gibberellins (GA) and abscisic acid (ABA) play opposite roles in controlling bud break and dormancy, while cytokinins (CK), auxin (IAA), and ethylene coordinate floral development. At the molecular level, flowering regulation involves several critical genes, including DORMANCY-ASSOCIATED MADS-box (DAM) and SHORT VEGETATIVE PHASE (SVP) genes that maintain dormancy, and FLOWERING LOCUS T (FT), SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 (SOC1), LEAFY (LFY), and APETALA1 (AP1) genes that promote flowering transition. Recent studies have also identified NAC (NAM, ATAF1/2, and CUC2) transcription factors that fine-tune early flowering in Prunus mume. Advances in genomics, transcriptomics, and phenological models have improved knowledge of flowering time control, although more work is needed to connect molecular mechanisms with field adaptation. This understanding is important for breeding cultivars suited to future climates.
-
Key words:
- Prunus mume /
- Bud dormancy /
- Dormancy release /
- Flowering time /
- Hormonal regulation /
- Environmental cues