









-
Orchidaceae, the largest group of monocotyledons, comprises approximately 25,000 species across 900 genera[1]. Cymbidium species, commonly known as Chinese orchids, are highly valuable in horticulture as potted plants and have garnered considerable attention owing to their exceptional ornamental traits and remarkable species diversity[2,3]. Deciphering the functions of key genes underlying these core ornamental traits, including their flowering time, diverse floral morphology, distinctive fragrance, flower color, and unique leaf variegation, is a crucial prerequisite for enhancing the ornamental value and breeding efficiency of Cymbidium[4]. Recent advances in multi-omics technologies have enabled large-scale genomic, transcriptomic, proteomic, and metabolomic analyses in Cymbidium, leading to the identification of numerous candidate genes implicated in these key ornamental traits[3,5,6]. These investigations have laid a solid foundation for elucidating the genetic basis of the traits and facilitating breeding-based improvement of Cymbidium. However, the prolonged breeding cycle of Cymbidium and the absence of a stable transformation system have severely impeded functional genetic studies[1,7,8]. Consequently, there is a pressing need to develop efficient and reliable tools for functional analysis of the genes.
Transient transformation technology has emerged as powerful alternative, enabling rapid gene silencing or overexpression in nonmodel plants without the need for stable transformation and thereby providing an efficient approach for analyses of gene function[9−11]. Among these techniques, virus-induced gene silencing (VIGS) is recognized as a fundamental reverse genetics approach that is widely used for transient gene silencing[1]. This technology exploits the plant's innate RNA interference (RNAi) machinery by introducing a mild viral vector that carries a fragment of the target gene, thereby specifically silencing the post-transcriptional expression of the target genes[12]. Compared with stable genetic modification techniques such as gene editing and Ethyl methane sulfonate (EMS) mutagenesis, VIGS does not require plant regeneration through tissue culture or the generation of stable transgenic lines, offering distinct advantages such as operational simplicity and shorter experimental timelines[13]. In 1995, the Tobacco mosaic virus (TMV)–phytoene desaturase (PDS) vector was first constructed, based on the TMV, pioneering the application of VIGS technology[14,15]. Subsequently, a VIGS system mediated by Tobacco rattle virus (TRV) was successfully established, and its effectiveness was validated by silencing the PDS gene in tobacco[14,15]. The TRV-VIGS system, in particular, has been successfully implemented in a broad range of dicot and monocot species owing to its mild infection symptoms, sustained silencing effect, and wide host range[16,17]. PDS, a crucial enzyme in carotenoid production, serves as a visual indicator for assessing the effectiveness of VIGS[18]. Its silencing leads to a characteristic photobleaching phenotype[18,19]. To date, the TRV-VIGS system has been widely applied in various crops, including pepper (Capsicum annuum)[20], barley (Hordeum vulgare)[16], and cotton (Gossypium hirsutum)[21]. Furthermore, its application has been successfully extended to numerous ornamental species, such as Gerbera[22], Rosa[12], Phalaenopsis[7], Narcissus[15], Gladiolus[17], and Lilium[23]. However, the establishment of a TRV-VIGS system in Cymbidium has been hindered by the orchid's unique biological constraints, such as a fleshy rhizome structure and a slow growth cycle. This technical barrier has significantly impeded functional gene studies in this genus, creating a pressing need for the development of efficient genetic tools for functional analysis.
In parallel, Agrobacterium-mediated transient transformation has become one of the key tools for gene function validation in nonmodel plants[10]. Compared with stable transformation, this technique offers distinct advantages such as simple operation, short cycle, and high-level transient expression without genomic integration[9]. The recently developed Ruby reporter system enables the production of a visible red pigment in transformed tissues, providing a facile and visual method for monitoring transformation efficiency[24]. This reporter has been successfully applied to establish transient expression systems across diverse plant species[25]. The Agrobacterium-mediated transient transformation technique has proven effective in a broad spectrum of crops and ornamentals[26], including Populus euphratica[27], Brassica campestris[28], Paeonia lactiflora[9], Rosa[29], Dendrobium[30], Gerbera[31], and Anthurium[32]. These successful applications provide strong support for rapid gene function analysis and trait improvement in horticultural species. However, the application of Agrobacterium-mediated transient expression systems in Cymbidium species remains limited, underscoring the urgent need to develop an efficient and stable transient overexpression platform for this genus.
To address this technological gap, we aimed to develop and optimize two efficient transient systems for gene function analysis in Cymbidium: A TRV-based VIGS system and a Ruby-based Agrobacterium-mediated transient overexpression system (Fig. 1a). Using a TRV vector carrying green fluorescent protein (GFP), we first confirmed systemic viral infection and movement within Cymbidium plants. Subsequently, the CsPDS gene fragment was cloned into the pTRV2 vector to generate the pTRV2-CsPDS construct, which induced noticeable photobleaching in the leaves. The silencing of the endogenous PDS gene was then confirmed by quantitative real-time polymerase chain reaction (qRT-PCR). Furthermore, we established an efficient Agrobacterium-mediated transient expression system using the Ruby reporter in the fleshy rhizomes and petals of Cymbidium, and then systematically optimized the key infiltration parameters for both systems. The establishment of these versatile tools provides reliable technical support for rapid characterization of gene function and will significantly accelerate the molecular breeding of Cymbidium orchids.
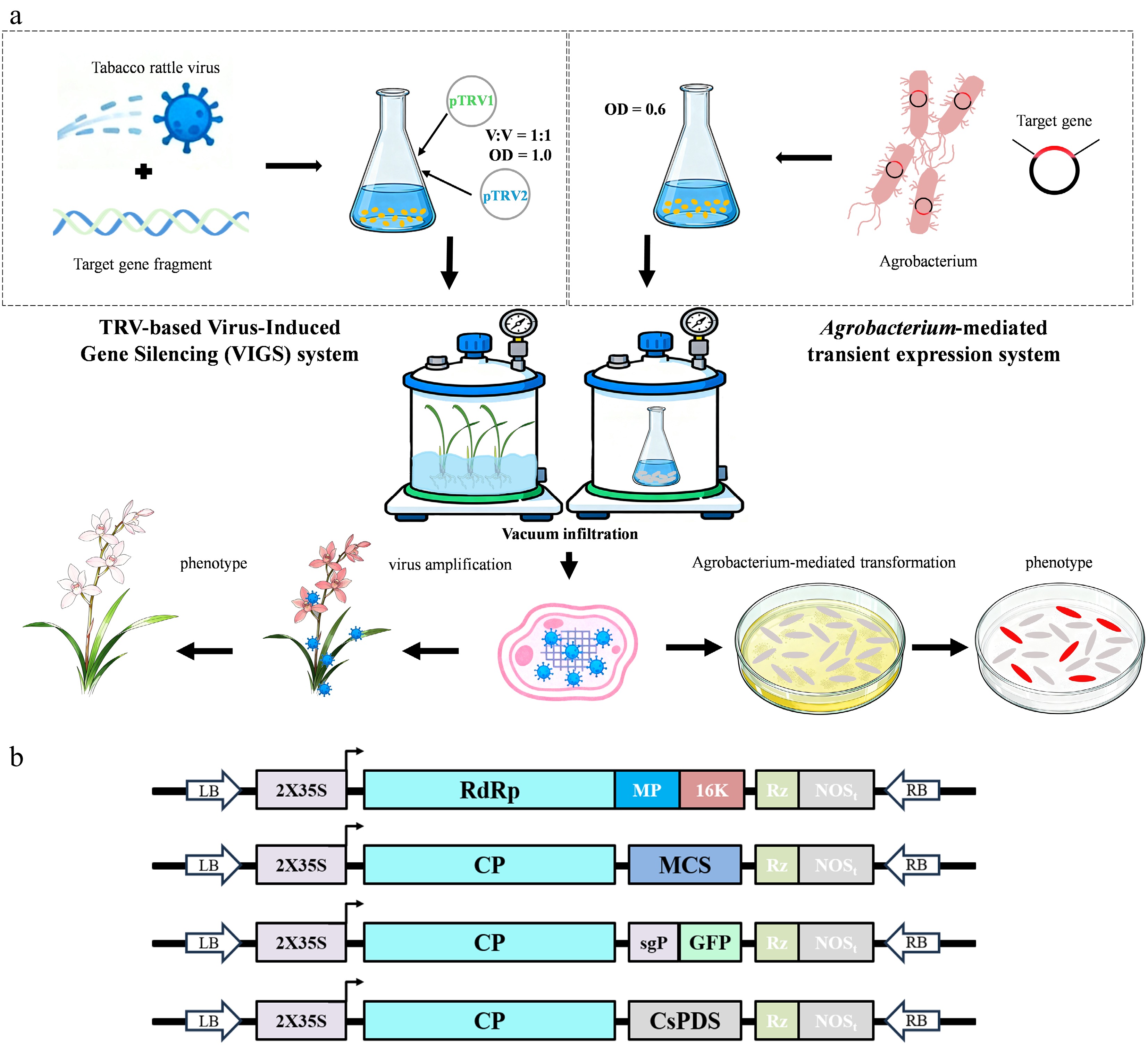
Figure 1.
Virus-induced gene silencing (VIGS) and Agrobacterium-mediated transient expression in Cymbidium. (a) A workflow for VIGS and Agrobacterium-mediated transient expression infiltration in Cymbidium. (b) Schematics of pTRV1, pTRV2, pTRV-GFP, and pTRV2-CsPDS. RdRp, RNA-dependent RNA polymerase; MP, movement protein; 16k, 16-kD protein; CP, coat protein; sgP, subgenomic promoter; MCS, multiple cloning site; LB, left border; RB, right border; Rz, self-cleaving ribozyme; NOSt, NOS terminator.
-
The Cymbidium sinense cultivar 'Xiao Xiang' was used in this study. Mature plants were cultivated under uniform conditions in a greenhouse of the Institute of Environmental Horticulture, Guangdong Academy of Agricultural Sciences, China. All plants were cultivated under consistent conditions, including pot type, fertilization, irrigation, and disease control protocols, as previously described[2]. Infection assays were initiated when the plants reached the developmental stage of flower and leaf bud emergence.
Total RNA extraction and identification of the CsPDS gene
-
Total RNA was isolated from Cymbidium leaves with the RNAprep Pure Plant Kit (TIANGEN, China) and was subsequently reverse-transcribed into first-strand complementary DNA (cDNA) following a previously described method[33]. To identify the PDS homologous gene in Cymbidium, full-length PDS protein sequences from model species, including Arabidopsis thaliana, Oryza sativa, and other closely related orchid species, were retrieved from the National Center for Biotechnology Information (NCBI,
www.ncbi.nlm.nih.gov ). To identify homologous sequences, a BLASTp analysis was carried out against the local Cymbidium species genomes using these sequences as input, using a threshold of 1e–5 for the E-value and 60% for sequence identity[34]. Conserved structural domains of the candidate PDS proteins were predicted using the Conserved Domain Database. Ultimately, PDS homologous gene of Cymbidium was identified, which contains the conserved PDS functional domains.Phylogenetic analysis and sequence alignment
-
Multiple sequence alignment of the complete amino acid sequences was performed using MEGA software (version 12), utilizing the ClustalW algorithm with its default settings for phylogenetic reconstruction. The alignment sequences were provided by the NCBI. A maximum likelihood (ML) phylogenetic tree was constructed with the aid of IQ-TREE software, version 1.6.12.[34]. The robustness of the phylogenetic tree was tested with 1,000 ultrafast bootstrap replicates. The resulting phylogenetic tree was visualized and annotated using Evolview (version 3). A separate multiple sequence alignment of CsPDS and its orthologs from other plant species was generated using DNAMAN software (version 5.0) under the default settings.
Vector construction
-
To construct the cloning vector, the CsPDS DNA fragment was first amplified from Cymbidium sinense 'Xiao Xiang' cDNA using high-fidelity Taq DNA polymerase (Vazyme, China) with the primers CsPDS-RT-F and CsPDS-RT-R (Supplementary Table S1). The amplified product was then purified, ligated into the pCE2 TA/Blunt-Zero vector (Vazyme, China), and transformed into Escherichia coli DH5α competent cells (Tiangen, China). Recombinant plasmid DNA was isolated using the FastPure® Plasmid Mini Kit (Vazyme, China). For the pTRV2-CsPDS vector construct, a 300-bp fragment of CsPDS with the EcoRI and BamHI restriction sites was amplified using the gene-specific primers (Supplementary Table S1). Then the 300-bp consensus CsPDS fragment was selected for insertion into the TRV2 vector. The pTRV1, pTRV2, pTRV-GFP, and pTRV-NtPDS vectors were obtained from Wuhan Boyuan Biotechnology Co., Ltd. Schematic representations of these vector constructs are shown in Fig. 1b. For the Ruby reporter construct, three crucial betalain biosynthesis genes, namely BvCYP76AD1S (GenBank: ON600884.1), BvDODA1S (GenBank: ON600882.1), and cDOPA5GT(GenBank: ON600883.1), were cloned into the binary vector pCAMBIA3301 under the control of the cauliflower mosaic virus (CaMV) 35S promoter to generate the Ruby plasmid. Empty vectors were used as negative controls in the subsequent experiments.
Virus-induced gene silencing assay
-
Agrobacterium tumefaciens strain EHA105 was transformed individually with the following plasmids: pTRV1, pTRV2, pTRV-GFP, pTRV-NtPDS, and pTRV2-CsPDS. Each transformation was cultured as previously described[33]. Subsequently, cultures of Agrobacterium were first cultivated for 16 h at 28 °C in Luria-Bertani (LB) medium containing 50 μg/mL kanamycin and 25 μg/mL rifampicin with shaking at 200 revolutions per minute (rpm). Following this, the cells were pelleted by centrifugation at 5,000 rpm for 7 min and finally resuspended in the infiltration buffer, which consisted of 10 mM 2-(N-morpholino)ethanesulfonic acid (MES), 100 µM acetosyringone, and 10 mM MgCl2 (pH adjusted to 5.6). The Agrobacterium cultures were normalized to different levels of optical density at 600 nm (OD600) for the ensuing optimization assays, with the target values set at 0.5, 1.0, and 1.5. For VIGS inoculation, a bacterial suspension containing pTRV1 was mixed with an equal volume (1:1 ratio) of a suspension containing one of the pTRV2-derived constructs (pTRV2, pTRV2-GFP, pTRV2-NtPDS, or pTRV2-CsPDS). The mixed cultures were incubated at 23 °C in the dark for 3 h to induce expression of the virulence genes. Prior to inoculation, the pseudobulb tissues of Cymbidium plants were gently wounded with a sterile needle, taking care to avoid the meristematic regions of the flower and leaf buds to prevent growth inhibition. The wounded plant tissues were soaked in their respective Agrobacterium suspension mixtures, followed by vacuum infiltration treatment. For optimization, different negative pressures (0.03, 0.06, and 0.09 MPa) were applied for a duration of 10 min. After infiltration, the plants were maintained in darkness at 25 °C for a period of 24 h. The transient transformation of tobacco was performed according to a previously described method[18]. Subsequently, the plants were transplanted back to their original pots and maintained under a temperature of 25 °C, with the light intensity set to 10,000 lux and the relative humidity maintained at 50%. Each experimental treatment consisted of 10 independent Cymbidium plants, which were considered to be biological replicates.
Agrobacterium-mediated transient transformation
-
The pCAMBIA3301-RUBY construct and the empty pCAMBIA3301 vector were introduced into Agrobacterium tumefaciens strain EHA105 using the freeze–thaw method. A single transformed colony was inoculated into 1.5 mL of LB liquid medium containing 50 μg/mL kanamycin and 25 μg/mL rifampicin, and cultured at 28 °C with shaking at 200 rpm for 24 h. Subsequently, 1 mL of the turbid bacterial culture was transferred into 100 mL of a fresh LB medium containing the same antibiotics for secondary growth. Bacterial cultures were harvested by centrifugation at 5,000 rpm for 7 min when the OD600 reached a value of 1.0. The bacterial pellets were resuspended in an infiltration buffer composed of Murashige and Skoog (MS) basal salts, 10 mM MES, 10 mM MgCl2, 100 μM acetosyringone, and 20 g/L sucrose (pH 5.6). The OD600 of the suspensions was adjusted to specific values (0.2, 0.4, 0.6, or 0.8) for optimization. Cymbidium rhizomes were immersed in the bacterial suspension within a narrow-mouth Erlenmeyer flask and subjected to vacuum infiltration. For optimization, different infiltration durations (5, 10, 15, and 20 min) were tested under a constant pressure of 0.1 MPa. For each treatment condition, 10 rhizomes were used as biological replicates. Following infiltration, the rhizomes were incubated with the bacterial suspension at 28 °C with shaking at 200 rpm for 2 h. The rhizomes were then rinsed three times with sterile water and transferred to a co-cultivation medium comprising 0.5× MS salts, 0.1 mg·L−1 naphthaleneacetic acid (NAA), 4 mg·L−1 6-benzylaminopurine (6-BA), 0.6% (w/v) agar, and 0.01% (v/v) Tween-20 (pH 5.4). Co-cultivation was carried out in the dark for 7 d to assess the optimal transformation efficiency. The plants infiltrated with the empty pCAMBIA3301 vector served as the negative control. Transient transformation of the petals was performed according to a previously reported method[35] with modifications. Briefly, petals were immersed in an Agrobacterium suspension (OD600 = 1.0) and subjected to vacuum infiltration at 0.01 MPa for 10 min. The treated petals were then placed on Petri dishes containing moist filter paper supplemented with 1% (w/v) sucrose and incubated at 25 °C for 7 d. The experiment included three biological replicates, each consisting of ten petals collected from five independent flowers.
Phenotypic analysis and polymerase chain reaction identification of infected plants
-
Transiently transformed Cymbidium protoplasts and TRV-infected rhizomes were examined for GFP fluorescence using an LSM710 confocal laser scanning microscope (Zeiss, Germany). To confirm TRV infection, total RNA was extracted from the rhizomes and leaves of plants from the following groups: Untreated controls, mock-treated (pTRV2) plants, and those infiltrated with pTRV2-GFP or pTRV2-CsPDS. qRT-PCR amplification was performed using specific primers targeting TRV RNA1 and RNA2 (pTRV1-F/R and pTRV2-F/R; Supplementary Table S1). Leaf phenotypes from all treatment and control groups were monitored daily for 45 d post-inoculation (dpi) for the systematic documentation of photobleaching and other phenotypic alterations.
Real-time quantitative polymerase chain reaction
-
At 30 dpi, newly emerged leaves were collected from untreated control plants, mock-treated (pTRV2) plants, and pTRV2-CsPDS infiltrated plants exhibiting the photobleaching phenotype. Total RNA was isolated from these samples and reverse-transcribed into cDNA. RT-qPCR was performed to analyze the relative expression levels of the target genes using the housekeeping gene β-actin (Mol013347) as an internal reference control, according to the method previously described[36]. The relative expression levels were calculated using the 2−ΔΔCq method, and statistical significance was assessed with three technical replicates per biological sample.
Statistical analysis
-
Statistical analyses and graphing were performed in GraphPad Prism 9.0. Data represent the mean ± standard error (SE) (n = 3 biological replicates with technical triplicates). Group differences (≥ 3 groups) were assessed by one-way analysis of variance (ANOVA) and Duncan's multiple range test, with significance determined at p < 0.05.
-
To assess the susceptibility of Cymbidium to TRV and establish a visual VIGS system, we inoculated the fleshy rhizomes of Cymbidium plants with Agrobacterium tumefaciens cultures harboring the pTRV1 and pTRV2-GFP vectors (Fig. 1b). At 7 dpi, clear GFP fluorescence was detected in the infiltrated rhizomes upon microscopic examination (Fig. 2a), confirming successful delivery of the TRV-GFP vector. Notably, GFP fluorescence was also observed in newly differentiated buds (red arrows), indicating systemic movement and stable maintenance of the expression vector after inoculation. To further corroborate TRV infection, PCR analysis was conducted on rhizome tissues exhibiting positive GFP signals. PCR amplification of TRV-specific RNA1 and RNA2 fragments confirmed the presence of the TRV vector in the inoculated rhizomes (Fig. 2b). To further assess the vectors' expression efficiency, pTRV1 and pTRV2-GFP were co-transfected into Cymbidium protoplasts using an efficient Cymbidium protoplast transfection system. At 20 h post-transformation (hpt), strong GFP fluorescence was observed in the transfected protoplasts (Fig. 2c). Collectively, these findings indicate that the TRV vector can replicate and mediate systemic spread within Cymbidium plants.
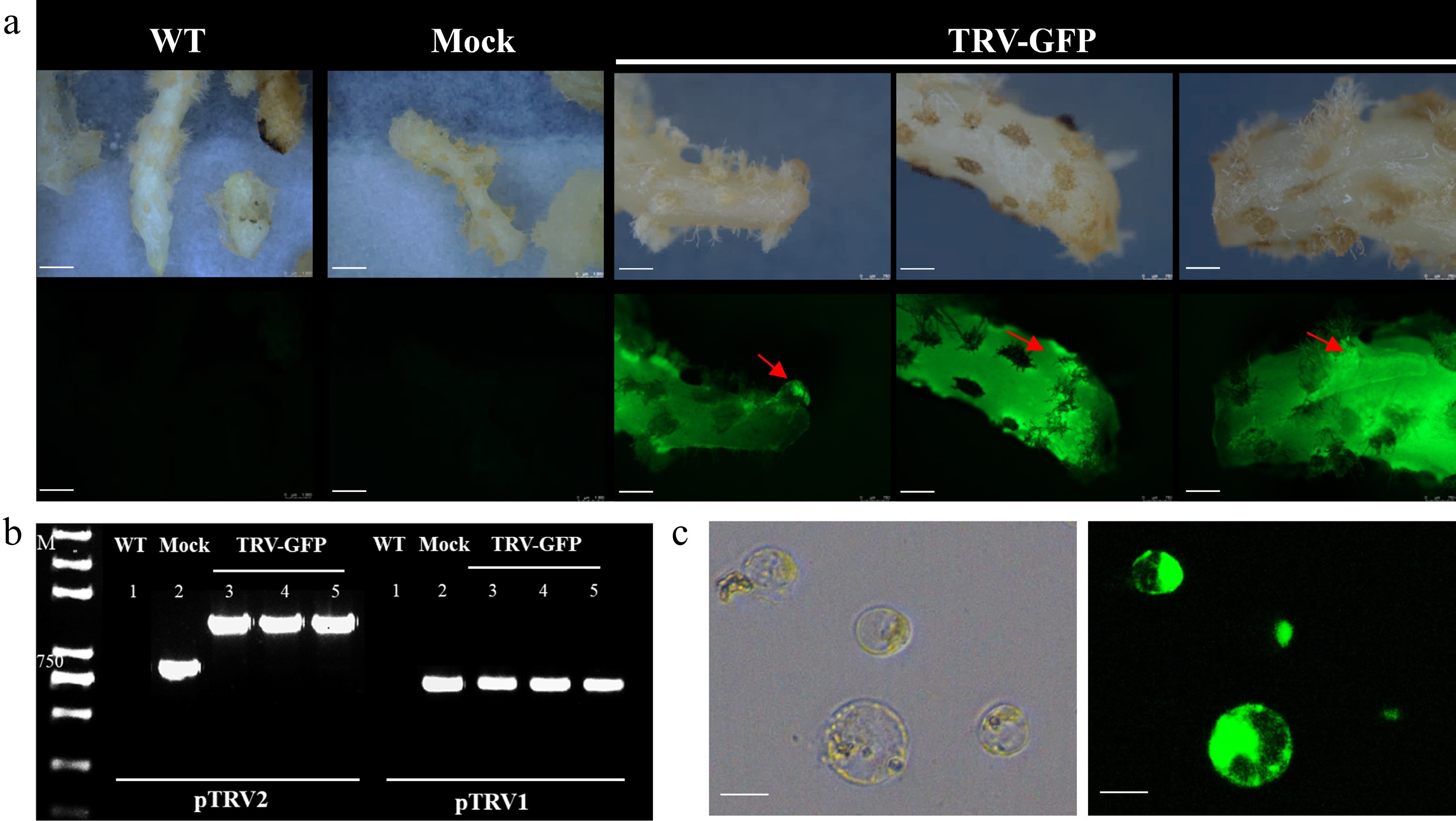
Figure 2.
Tobacco rattle virus (TRV) infection and systemic movement in Cymbidium tissues. (a) VIGS of the rhizomes of Cymbidium with pTRV2-GFP. Green fluorescent protein was detected in the infected rhizomes, and this fluorescence was transmitted to the uninfected newly emerging bud sites. WT, wild-type; Mock, rhizomes injected with pTRV1 + pTRV2; TRV-GFP, rhizomes injected with pTRV1 + pTRV2-GFP. Scale bar: 1,000 μm. (b) RT-PCR detection of TRV2 and TRV2-GFP vectors in infected tissues. M, marker. (c) Transient GFP expression in Cymbidium protoplasts transfected with pTRV2-GFP. Cells were examined 20 h after inoculation under the fluorescence microscope. Scale bar: 50 μm.
Genomewide identification of CsPDS in C. sinense
-
PDS, a critical enzyme in carotenoid biosynthesis, serves as an effective visual marker for assessing VIGS efficiency because of the photobleaching phenotype resulting from its suppression. Within the C. sinense genome, we identified a PDS homolog, designated CsPDS. Its open reading frame (ORF) is 1,749 bp, encoding a 583-amino acid protein with a predicted molecular weight (MW) of 65.29 kDa and a theoretical isoelectric point (PI) of 7.51 (Supplementary Table S2). Prediction of its subcellular localization indicated that the CsPDS protein localizes to the chloroplast, consistent with its role in carotenoid biosynthesis. An analysis of conserved motifs in the CsPDS protein was performed using the MEME suite, which led to the discovery of 10 motifs, subsequently labeled as Motif 1 to Motif 10 (Fig. 3a and Supplementary Table S3). We performed a phylogenetic analysis of PDS sequences from C. sinense and diverse plants, including Oryza sativa, Arabidopsis thaliana, Dendrobium catenatum, Oncidium, Phalaenopsis equestris, and other species, to investigate the evolutionary relationships of CsPDS. Phylogenetic analysis revealed that PDS in Cymbidium aligned most closely with PDS from Oncidium, D. catenatum, and P. equestris, suggesting a conserved evolutionary trajectory among orchidaceous species (Fig. 3b). Multiple sequence alignment of CsPDS with homologous PDS proteins from various plant species revealed a high degree of sequence conservation, particularly among orchids (Fig. 3c). The sequence alignment showed that CsPDS shares high identity with its orthologs, showing 99.66% identity with PDS from Cymbidium ensifolium, 96.74% with the Cymbidium goeringii protein, and 92.78% with the Oncidium ortholog (Fig. 3c, d), These results exhibited remarkable evolutionary conservation within the PDS-specific functional domains across plant lineages.
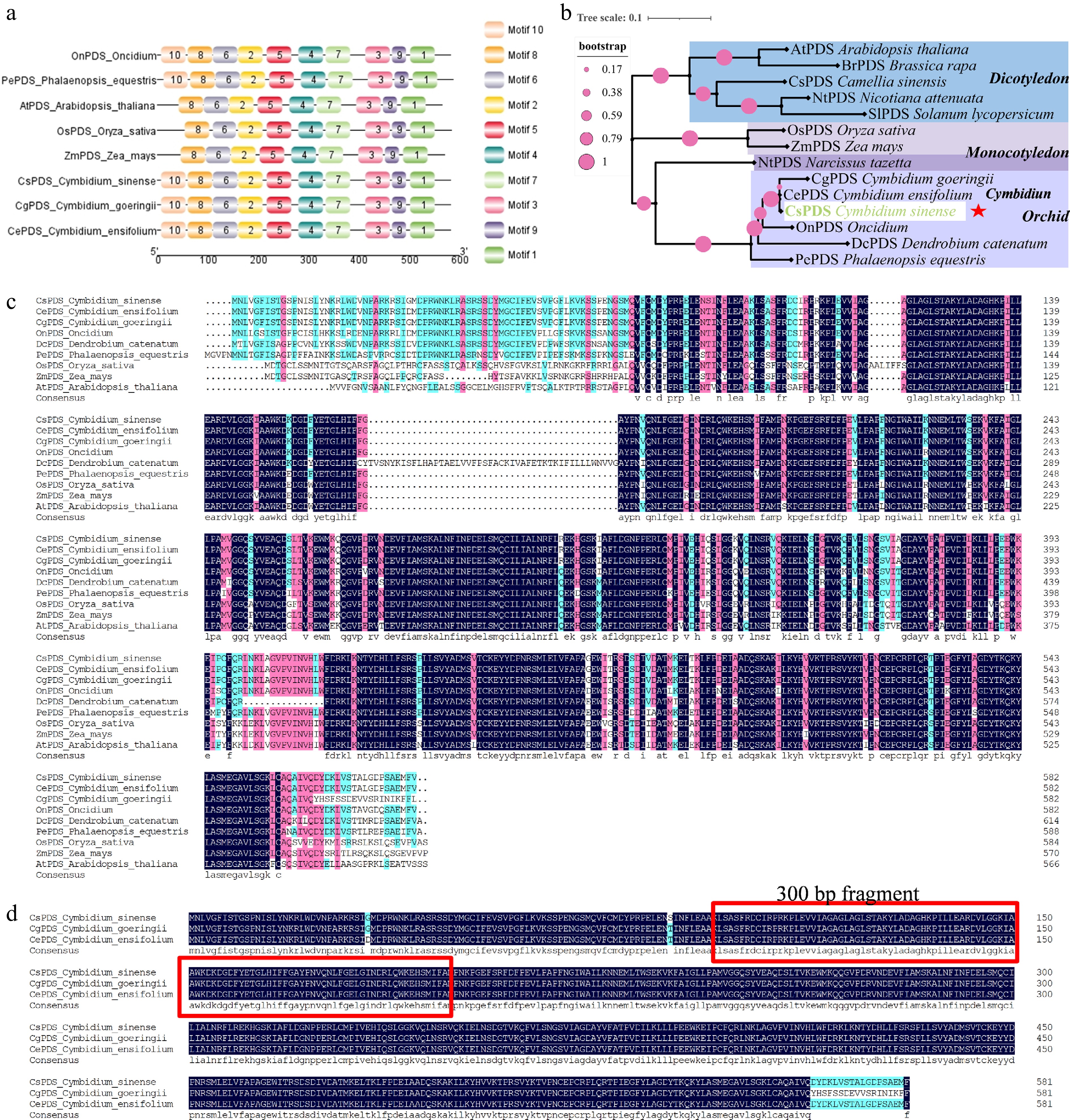
Figure 3.
Identification of the PDS gene in C. sinense. (a) Conserved protein motifs identified in CsPDS and orthologs from other plant species. (b) Maximum likelihood phylogenetic tree of PDS proteins from C. sinense and selected angiosperms. The CsPDS clade is highlighted with a red star. (c) Multiple sequence alignment of PDS protein sequences from C. sinense and other plants. (d) Protein sequence alignment of PDS from three Cymbidium species.
pTRV2-CsPDS vector construction and silencing of CsPDS in C. sinense
-
To establish a TRV-based VIGS system in Cymbidium, a 300-bp conserved fragment of CsPDS was inserted into the pTRV2 vector to generate the pTRV2-CsPDS construct (Figs. 1b and 3d). The pTRV2-NtPDS construct served as a positive control, and its infiltration into tobacco (Nicotiana benthamiana) plants resulted in distinct photobleaching in newly developed leaves (Fig. 4a), confirming the vector's functionality. Similarly, infiltration of pTRV2-CsPDS into Cymbidium stems induced visible photobleaching in emerging leaf buds at 45 dpi, whereas control plants treated with empty pTRV2 or untreated plants remained phenotypically normal (Fig. 4b and Supplementary Fig. S1a). RT-PCR analysis confirmed the presence of TRV1 and TRV2 sequences in plants exhibiting photobleaching and in empty vector-treated plants, but not in uninoculated controls (Fig. 4c), indicating successful viral infection. To determine whether the photobleaching phenotype was mediated by CsPDS silencing, we performed RT-qPCR analysis on newly developed leaf tissues. The results showed that the endogenous CsPDS transcript levels in pTRV2-CsPDS-treated plants were significantly reduced by 37.53%–72.40% compared with the controls, whereas mock-treated plants showed no notable decrease in CsPDS expression (Fig. 4d). These findings confirm that TRV-mediated silencing of CsPDS is responsible for the observed photobleaching phenotype in Cymbidium plants. To optimize VIGS efficiency, we evaluated different Agrobacterium densities (OD600 = 0.5, 1.0, 1.5) and various intensities of negative pressure (0.03, 0.06, and 0.09 MPa). Plants' survival and silencing efficiency were recorded 1 month after infiltration (Supplementary Table S4 and Fig. S1b). As shown in Fig. 4e, f, at an Agrobacterium density of OD600 = 0.5, the plant survival rates were 86.90% and 85.24% under 0.03 MPa and 0.06 MPa, respectively, but decreased to approximately 75.17% at 0.09 MPa. In contrast, the silencing efficiency remained below 44.44% across all negative pressure conditions. At OD600 = 1.0, survival rates ranged from 68.88% to 73.89% across all pressure conditions, with the highest silencing efficiency (69.44%) achieved at 0.06 MPa. At OD600 = 1.5, survival rates fell below 35.56%, despite the higher silencing efficiency, which ranged from 66.67% to 83.33%. Collectively, these results identify the optimal conditions for VIGS in Cymbidium as an OD600 of 1.0 under a negative pressure of 0.06 MPa, which balanced high silencing efficiency with acceptable plant survival.
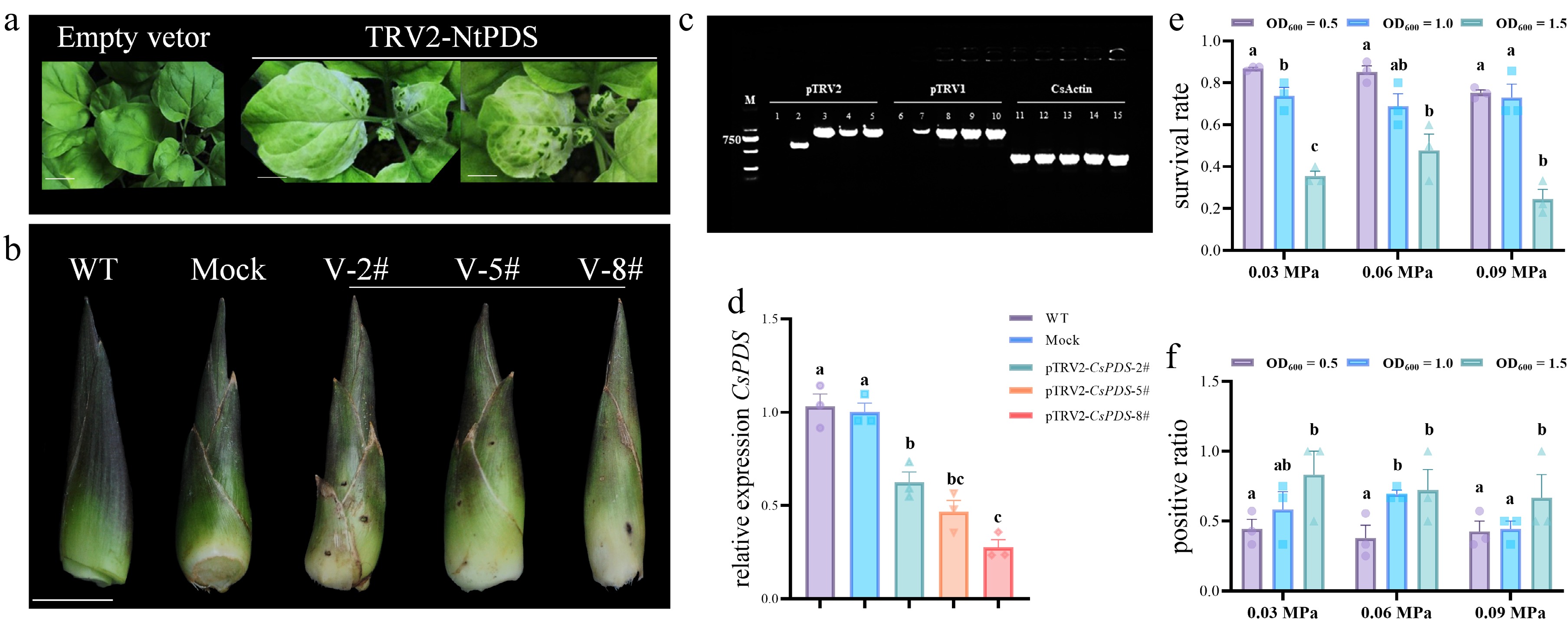
Figure 4.
Functional analysis of CsPDS silencing in C. sinense. (a) Positive control for the VIGS system: Nicotiana benthamiana leaves infiltrated with pTRV2-NtPDS exhibit photobleaching. Scale bar: 10 mm. (b) Photobleaching phenotype in C. sinense leaf buds after treatment with pTRV1 + pTRV2-CsPDS. (c) PCR-based tracking of TRV vectors confirms successful infection and replication in C. sinense tissues. WT: 1, 6, and 11; Mock: 2, 7, and 12; pTRV2-CsPDS: 3, 4, 5, 8, 9, 10, 13, 14, and 15. (d) Expression of CsPDS was downregulated in photobleached leaf buds in C. sinense. Significance was evaluated by one-way analysis of variance. The survival rates (e) and positive rates (f) of the plants in the infection solutions with different levels of negative pressure and different OD600 values after infection. Data represent the mean ± standard error of the mean (SEM) of three biological replicates. The transient transformation rate = number of positive plants / total number of infiltrated plants.
Agrobacterium-mediated transient transformation using the Ruby reporter in Cymbidium
-
To establish an Agrobacterium-mediated transient expression system in Cymbidium, we utilized the betalain-producing Ruby cassette as a visual marker (Fig. 5a). We first validated the system using Phalaenopsis 'Ama' petals, where distinct red-purple pigmentation was observed exclusively in infiltrated areas, although no coloration was detected in the empty vector-inoculated control group (Fig. 5b). This result confirmed the efficacy of the Ruby reporter in Orchidaceae. We then applied this system to Cymbidium petal tissues. As shown in Fig. 5c, discrete red-purple spots were observed on the infiltrated petals, whereas no such pigmentation was detected in the control group. We further systematically optimized the transformation protocol for Cymbidium rhizomes by testing four Agrobacterium concentrations (OD600 = 0.2, 0.4, 0.6, and 0.8) and four durations of vacuum infiltration (5, 10, 15, and 20 min) under a constant vacuum pressure of 0.01 MPa. Both transformation efficiency and survival rate were assessed for each condition (Fig. 5d–h). At OD600 = 0.2, a 15-min infiltration yielded the highest transformation efficiency (18.89%) but with a low survival rate of 34.44% (Fig. 5e); at OD600 = 0.4, the maximum transformation efficiency (18.89%) was achieved with a 10-min infiltration, and the survival rate was elevated to 68.89% (Fig. 5f); at OD600 = 0.6, a 10-min infiltration resulted in the highest transformation efficiency across all conditions (27.78%) while maintaining a favorable survival rate of 68.89% (Fig. 5g); at OD600 = 0.8, the maximum transformation efficiency was 18.89% with a 10-min infiltration, accompanied by a sharp decline in the survival rate to 31.11% (Fig. 5h). Collectively, these results demonstrate that lower Agrobacterium concentrations or shorter vacuum infiltration durations resulted in reduced transformation efficiency, whereas excessive bacterial concentration or prolonged infiltration caused irreversible tissue damage, leading to rhizome browning and, consequently, sharply reduced survival rates (Fig. 5d). Our comprehensive evaluation identified the optimal conditions for transient transformation of Cymbidium rhizomes as an OD600 of 0.6 combined with a 10-min vacuum infiltration, as this combination balances high transgene expression with minimal tissue damage.
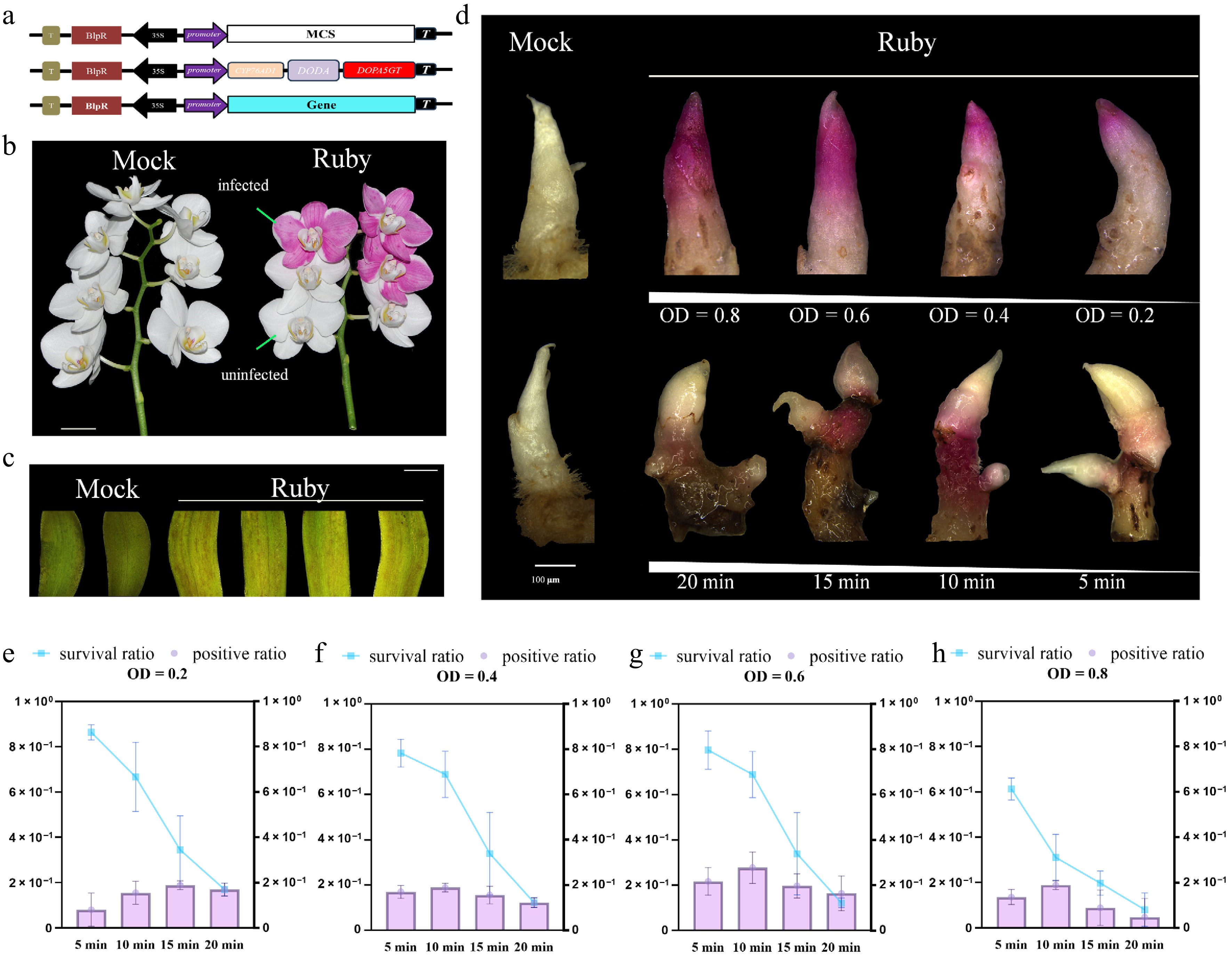
Figure 5.
Establishing the Ruby reporter for transient transformation in Cymbidium. (a) Schematics of the Ruby expression vector. (b) Phenotype of the Ruby reporter in the petals of Phalaenopsis. Scale bar: 10 mm. (c) Agrobacterium-mediated transient expression of the Ruby reporter in the petals of Cymbidium. Scale bar: 5 mm. (d) Distinct red-purple pigmentation was observed on Cymbidium rhizomes following infiltration with different Agrobacterium concentrations (upper). The growth phenotype of rhizomes was also recorded after various vacuum infiltration durations (lower). Scale bar: 100 μm. (e)–(h) Survival rates and positive rates of Cymbidium rhizomes in the infection solutions with different times of vacuum infiltration and different OD600 values after infection. Data represent the mean ± SEM of three biological replicates. The transient transformation rate = number of positive plants/total number of infiltrated plants.
-
Cymbidium species, commonly known as Chinese orchids, hold significant economic and horticultural value as ornamental plants[4]. In recent years, the rapid advancement of multi-omics technologies has identified numerous gene sequences linked to key agronomic traits in Cymbidium, creating an urgent need for efficient functional validation tools[37]. However, as a typical nonmodel plant, Cymbidium shares inherent biological constraints with other bulbous and woody plants, such as an extended juvenile period and low genetic transformation efficiency[33]. These limitations have severely restricted the application of conventional gene function validation approaches, resulting in a significant gap between the discovery of genes and their functional characterization. This has greatly hindered the progress of Cymbidium research.
Virus-induced gene silencing has emerged as a powerful tool for rapid functional genomics in plants, owing to its efficiency and high-throughput capability[8,27]. Among various viral vectors, TRV is widely adopted for VIGS applications owing to its broad host range, mild infection symptoms, and sustained silencing efficacy[38,39,40]. In Orchidaceae, although a robust VIGS system based on Cymbidium mosaic virus (CyMV) has been established in Phalaenopsis[7], this system may lack universal applicability across all orchid species. Systemic viral infection is crucial for successful VIGS. TRV is one of the susceptible viral vectors of Orchidaceae[38], and its utility has been validated by previous studies. For instance, TRV-mediated silencing of the DoERF5 gene significantly reduced protocorm-like body regeneration rates in Dendrobium[41]. Furthermore, TRV-VIGS systems have been successfully implemented in various ornamental plants through leaf infiltration—the most common delivery method[23]. In orchids such as Phalaenopsis and Dendrobium, this typically involves syringe infiltration of the abaxial leaf surface near emerging inflorescences. However, VIGS protocols require species-specific optimization. The unique rhizome structure, leathery leaves, and distinct flower development cycle of Cymbidium render conventional leaf infiltration ineffective. To address this challenge, we used vacuum infiltration targeting the meristems of leaf buds and floral buds on the pseudobulb of Cymbidium (Fig. 1a), an approach adapted from methods used in bulbous species such as Lilium[23], Paeonia lactiflora[42], and Gladiolus gandavensis[17]. This method effectively overcame the penetration barriers presented by tough, leathery tissues of Cymbidium. Notably, although vacuum infiltration effectively penetrates tissue barriers, it can cause more structural damage or necrosis compared with syringe injection[12]. Therefore, optimizing parameters such as bacterial density, culture conditions, and vacuum pressure is essential for balancing infection efficiency with plant survival. For example, optimal silencing in Phalaenopsis requires an OD600 of 1.0 for leaves and 1.5–2.0 for floral tissues, with incubation at 28 °C[1,12], confirming the species-specificity of VIGS parameters. Through systematic optimization, we established that vacuum infiltration with Agrobacterium at an OD600 of 1.0 under a negative pressure of 0.06 MPa, followed by dark incubation at 25 °C for 24 h prior to normal culture, achieved optimal silencing efficiency and survival rates in Cymbidium (Fig. 4). However, to achieve more stable and efficient gene silencing, future studies could focus on optimizing additional parameters, including the plants' developmental stage, the size of the target gene fragment, and infiltration duration. Previous research in Iris japonica, Phalaenopsis, and Dendrobium has demonstrated that both plant age and the length of the TRV-carried fragment significantly affect silencing efficiency[1,7,8,12]. Specifically, plants older than 1 year often show reduced susceptibility to VIGS, whereas shorter TRV-carried fragments can yield higher specificity and efficiency. Implementing VIGS at appropriate developmental stages can also help avoid silencing essential genes required for plant growth and development. The deployment of short RNA inserts marks a pivotal advancement in VIGS technology[43]. Prospective strategies that integrate multi-targeting paradigms—enabled by clustered regularly interspaced short palindromic repeat (CRISPR)-mediated gene editing—to meticulously refine the targeting specificity of these inserts while concurrently broadening their applicability to a wider spectrum of nonmodel crops across diverse taxonomic families are positioned to elevate VIGS as a high-throughput platform for functional genomics and a versatile modality for crop improvement in agricultural biotechnology.
Transient gene expression represents a powerful alternative for functional gene studies in plants and has been widely adopted in various crops and ornamental species. However, transient gene expression systems for Cymbidium remain underdeveloped. Although a highly efficient protoplast transformation system for Cymbidium was established in our previous study[44], its application remains constrained by the technically demanding protoplast isolation process and stringent requirements for protoplast and plasmid DNA quality[10,45], which limits its large-scale application. In contrast, Agrobacterium-mediated transient expression can effectively circumvent these technical bottlenecks, offering the advantages of simplicity and broad applicability. In this study, we successfully established an Agrobacterium-mediated transient overexpression system in both the petals and rhizomes of Cymbidium using the visual Ruby reporter (Fig. 1a). Furthermore, the optimal transformation conditions for rhizomes were identified as an Agrobacterium suspension with an OD600 of 0.6 and a 10-minute vacuum infiltration (Fig. 5). Notably, we adapted the infiltration method to accommodate the unique morphology of Cymbidium. Unlike the syringe infiltration used in Phalaenopsis petals[46], we used vacuum infiltration, a strategy similar to that used in rose (Rosa spp.)[47]. This approach effectively addressed the challenges posed by the extended floral development period of Cymbidium. However, we observed that an excessive Agrobacterium concentration or prolonged infiltration induced tissue browning and softening in Cymbidium rhizomes (Fig. 5d). These rhizomes failed to recover normal growth, making them unsuitable for subsequent studies, which were consistent with the observed reduction in transformation efficiency and plant survival rate (Fig. 5e–h). Future optimization could focus on the developmental stage of Cymbidium rhizomes, the infiltration parameters, vacuum pressure intensity, and rhizomes' compatibility across different Cymbidium cultivars. Previous studies have demonstrated that external treatments including vacuum infiltration, cotyledon piercing, and surfactant application can significantly enhance the efficiency of transient transformation[48,49]. For instance, appropriate vacuum intensity enhanced transient transformation efficiency in Paeonia lactiflora[9], whereas younger tissues exhibited higher transformation rates in hybrid poplar (Populus spp.)[48]. These findings provide valuable guidance for the future optimization of the transient expression system in Cymbidium.
-
Although stable genetic transformation of Orchidaceae has been documented for decades[50,51] and VIGS was implemented early in some orchids[1,7], successful stable genetic transformation remains restricted to a limited number of laboratories to date[37] and lacks broad applicability across different genera. In this study, we established a functional transient expression system for Cymbidium using protocols distinct from those applied in related orchids such as Phalaenopsis and Dendrobium. The systems developed here provide efficient and practical platform for the rapid functional characterization of candidate genes. This methodological advancement is particularly valuable, given the extensive genetic resources now available for Cymbidium, which include high-quality genome sequences[5,52], comprehensive transcriptomic datasets[6,34], and detailed proteomic and metabolomic profiles[2]. The integration of our transient expression platforms with these multi-omics resources will significantly accelerate the identification and validation of genes that regulate key ornamental traits, thereby advancing molecular breeding programs for Chinese Cymbidium.
-
The authors confirm their contributions to the paper as follows: oversaw the experimental design: Yang F, Wu Z, Zhu G; conducted the experiments: Lin Z; data analysis and figure assembly: Lin Z, Lu C; responsible for manuscript writing: Yang F, Wu Z, P Delaplace, Lin Z. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available in the NCBI repository. All experimental data obtained and examined in this study are contained within supplementary information files.
-
This study acknowledges the financial support from the National Key Research and Development Program of China (2023YFD2300904), the Science and Technology Planning Project of Guangdong Province (2024B1212060012), the Seed Industry Revitalization Project of the Special Fund for the Rural Revitalization Strategy of Guangdong Province (2024-NPY-00-035), the Operation and Maintenance of Guangdong Famous and Excellent Flowers Germplasm Resource Nursery (2023-NBH-00-009), the Special Funding for the Construction of the High-Level Academy of Agricultural Sciences (NYQS202611), the Construction of an Integrated Industry System for Breeding, Propagation and Promotion of Chinese Orchids (2025-NQD-06-001), and the Innovation Team of Modern Agriculture Industry Technology System in Guangdong Province (2026CXTD12). Support was also received from the Ex Situ Conservation and Artificial Propagation of National Key Protected Orchids and Ferns (2025010), the project for the Guangdong Basic and Applied Basic Research Foundation (2024A1515011604, 2024A1515013187), the Modern Seed Industry Innovation Capability Enhancement Project of Guangdong Academy of Agricultural Sciences, the Special Foundation for Introduction of Scientific Talents of GDAAS (R2021YJ-XD001), and the Guangdong Academy of Agricultural Sciences Project (CYZX202406, R2023PY-JG023, XTXM202201, R2020PY-JX018, and XT202212).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/opr-0026-0015.
-
# Authors contributed equally: Zengyu Lin, Chuqiao Lu
- Supplementary Table S1 Primers used in this study.
- Supplementary Table S2 Information of PDS gene family members in C. sinense.
- Supplementary Table S3 Conserved motifs identified in PDS from different orchid species by MEME analysis.
- Supplementary Table S4 Survival rates of plants infected at different OD concentrations and different intensities of negative pressure.
- Supplementary Fig. S1a (A) Photobleaching phenotype in C. sinense leaf buds after treatment with pTRV1 + pTRV2-CsPDS. (B) PCR-based detection of TRV vectors verified successful infection and replication in C. sinense tissues, and illustrates the infection positive rate under various OD concentrations and negative pressure intensities.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lin Z, Lu C, Zhu G, Delaplace P, Wu Z, et al. 2026. Development of virus-induced gene silencing and Agrobacterium-mediated transient expression systems for gene function analysis in Chinese Cymbidium. Ornamental Plant Research 6: e024 doi: 10.48130/opr-0026-0015 

Development of virus-induced gene silencing and Agrobacterium-mediated transient expression systems for gene function analysis in Chinese Cymbidium
- Received: 29 November 2025
- Revised: 08 April 2026
- Accepted: 20 April 2026
- Published online: 23 June 2026
Abstract: Chinese orchids (Cymbidium spp.) are high-value ornamental plants, renowned for their elegant flowers, leaf variegation, and distinctive fragrance. However, their low transformation efficiency and prolonged regeneration periods have hindered the functional characterization of key genes in Cymbidium. To address these challenges, we developed a Tobacco rattle virus (TRV)-mediated virus-induced gene silencing (VIGS) system and a highly efficient Agrobacterium-mediated transient expression system using the Ruby reporter. These systems facilitate efficient transient gene silencing and overexpression in Cymbidium, enabling functional characterization of target genes. Phytoene desaturase (PDS), a visual marker commonly used for VIGS, and the Ruby reporter were selected to validate the system's efficiency. Infiltration with pTRV2-GFP confirmed that TRV can establish systemic infection and movement within Cymbidium plants. Furthermore, inoculation with pTRV2-CsPDS induced distinct photobleaching in leaves, and quantitative real-time polymerase chain reaction analysis confirmed the successful silencing of the endogenous PDS gene. We further optimized the VIGS conditions and identified the optimal parameters as an optical density at 600 nm (OD600) of 1.0 under a negative pressure of 0.06 MPa, followed by dark incubation at 25 °C for 24 h, which yielded the highest infiltration efficiency and plant survival rate. Using the Ruby reporter system, we also demonstrated efficient transient overexpression in Cymbidium petals and fleshy rhizomes. Additionally, we established that the optimal infiltration conditions for rhizomes were an Agrobacterium density of OD600 = 0.6 and a 10-min infiltration period. Collectively, the TRV-based VIGS system and the Ruby-based transient expression system offer reliable tools for comparative functional studies, enabling the dissection of molecular mechanisms that underlie key biological processes in Cymbidium.