









-
With societal and economic developments, the demand for urban garden plants with leaves of various colors has increased. Traditional plant materials used for landscaping are often monotonous and far from meeting the societal needs. Therefore, the breeding of garden plants with colorful leaves has recently gained momentum. Plants with colorful leaves can be as colorful as flowers. They can enrich urban landscapes; thus, they have become new favorites for garden greening. Among plants with colorful leaves, the yellow-leaf ornamental plants are the brightest and most eye-catching[1]. They also have ornamental value, making them suitable for the landscaping of gardens and courtyards.
Genetic engineering is an effective way to change the leaf color of plants. For example, the overexpression of MYB transcription factors can activate the expression of genes involved in anthocyanin biosynthesis, leading to the increased redness of leaves or fruit peels. Specifically, the overexpression of Myb119 in Poplus trichocarpa (P. trichocarpa) significantly increased the anthocyanin level, resulting in red leaves[2].
Golden2-like (GLK), a member of the GARP transcription factor family, widely exists in plants[3]. In plants, the GLKs mainly regulate chloroplast development. They are also involved in biotic and abiotic stress responses, senescence, and hormone signaling[4]. Studies have shown that overexpression of AtGLK1 in Arabidopsis thaliana can promote the development of chloroplasts in some non-photosynthetic organs such as roots. On the contrary, the glk1glk2 double mutant of Arabidopsis exhibited light green leaves, abnormal chloroplast development and decreased accumulation of chlorophyll precursors[3,5,6,7]. The GLK family is diverse in different species. The Arabidopsis thaliana and Oryza sativa genomes each contain two GLKs[3]. Recent studies have revealed that members of the GLK family may be functionally redundant while exerting different functions in different tissues[5]. For example, ZmGLK1 and ZmGLK2 in maize specifically regulate the development of sheath and mesophyll cells, respectively[8]. SlGLK1 in Solanum lycopersicum and CaGLK1 in Capsicum annuum are mainly expressed in roots, whereas SlGLK2 and CaGLK2 are mainly expressed in fruits[9]. A recent study has reported that the birch genome (Betula platyphylla Suk.) contains only one GLK gene[10], and the repression of BpGLK1 affects chloroplast development, thereby leading to yellow-green leaves. GLK family also plays an important role in response to adversity stress. Studies have shown that Arabidopsis overexpressing AtGLK1 has enhanced resistance to the Fusarium graminearum and Botrytis cinerea[11,12].
The hybrid poplar Populus alba × P. berlinensis is a major broadleaf tree species in Northeast China. It has been widely used for landscaping in urban roads, parks, courtyards, and residential areas because of its straight trunk, gray green bark, and beautiful appearance. In addition, P. alba × P. berlinensis is a triploid male polar species, which exhibits multiple advantages for landscaping[13]. For example, it does not generate catkins similar to other poplar species, and its male infertility can prevent gene drifting, making it an ideal material for molecular breeding. In this study, we identified GLKs that are highly expressed in P. alba × P. berlinensis leaves, and then employed gene silencing technology to generate GLK RNA interference (RNAi) transgenic lines with significantly reduced GLK expression levels. The transgenic lines exhibited aesthetically appealing and eye-catching yellow-green leaves. Since they are infertile triploids, they can satisfy the demands of various landscaping purposes after a safety assessment.
-
The GLK genes are involved in chloroplast development, and the repression of GLKs has been reported to cause yellow leaves in multiple species[2]. In this study, we intended to generate a golden-yellow poplar varieties using gene silencing technology and targeted the GLK genes in a male triploid hybrid poplar, P. alba × P. berolinensis, for silencing. The annotated P. trichocarpa GLKs were used as queries to identify PabGLKs from the unigenes we assembled from the sequence reads from the cDNA libraries of the triploid. Finally, a total of five candidate PabGLKs were obtained and designated PabGLK1 to PabGLK5.
A phylogenetic tree was constructed to reveal the genetic relationships of GLKs from P. alba × P. berolinensis, P. trichocarpa, and Arabidopsis. The results indicate that the identified PabGLKs could be divided into three major groups (Fig. 1a). Groups I and II each contained two PabGLKs; namely, PabGLK1 and PabGLK5 were in Group I, whereas PabGLK2 and PabGLK3 were in Group II. Group III had only one member, PabGLK4. A multiple sequence alignment of the five PabGLKs revealed that the nucleotide sequences of the two PabGLKs, PabGLK1 and PabGLK5, shared the highest similarity of 98%, while PabGLK4 and PabGLK3, shared the lowest similarity of 91% (Fig. 1b and d). We then measured the total expression levels of PabGLKs in the leaves, stems, and roots of P. alba ×
P. berolinensis using qPCR. The PabGLKs were expressed most highly in leaves, followed by stems. Roots had the lowest levels of PabGLK trasncripts, which were approximately 10 times lower than those of their counterparts in leaves (Fig. 1c). 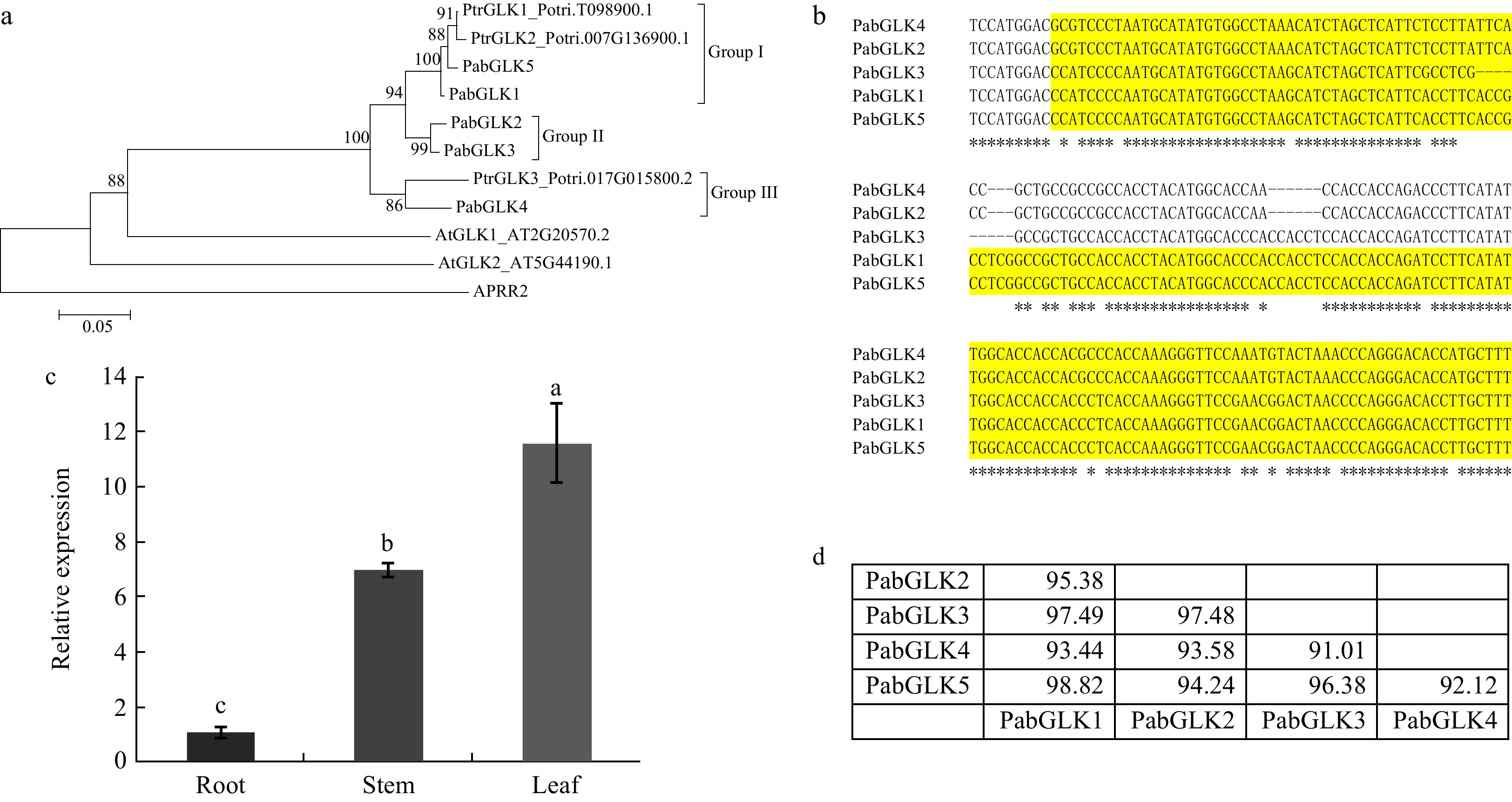
Figure 1. Identification of GLKs that are expressed in P. alba × P. berolinensis. (a): Phylogenetic relationship among GLKs from P. alba × P. berolinensis, P. trichocarpa, and Arabidopsis. (b): A multiple sequence alignment of the identified PabGLKs. The colored region was used for RNAi. (c): Tissue specific expression levels of the PabGLKs in P. alba ×
P. berolinensis quantified with qRT-PCR. 18S rRNA and α-Tubulin were used as endogenous control. (d): Nucleotide sequence similarities among differnt PabGLKs in P. alba × P. berolinensis. Generation of PaGLK repressed poplar
-
GLKs have been reported to be functionally redundant in many species. In P. alba × P. berolinensis, a total of five expressed GLKs were identified in leaves. Based on the results of multiple sequence alignment, we used a 200-bp region of coding region that is conserved among all PabGLKs for RNA silencing (Fig. 1b). An RNAi vector (p35S::GLK-RNAi) was designed to simultaneously silence all PabGLKs by targeting the selected region. The resulting RNAi construct was delivered into P. alba × P. berolinensis by Agrobacterium-mediated genetic transformation and three PabGLKs repression lines, designated RE1, RE2, and RE3, were obtained. In addition to the three PabGLK RNAi lines, we also generated three PabGLK5 overexpression lines, OE1, OE2, and OE3.
Next, the expression levels of PabGLK5 and the total expression levels of all five PabGLKs in the leaves of different RE and OE lines relative to the WT were measured using qPCR. The total transcript levels of PabGLKs in the leaves of RE1, RE2, and RE3 lines were reduced by 74.10%, 71.10%, and 60.70%, respectively, compared with the WT. By contrast, the PabGLK5 transcript levels in the leaves of OE1, OE2, and OE3 lines increased 220.61%, 61.00%, and 51.80%, respectively (Fig. 2).
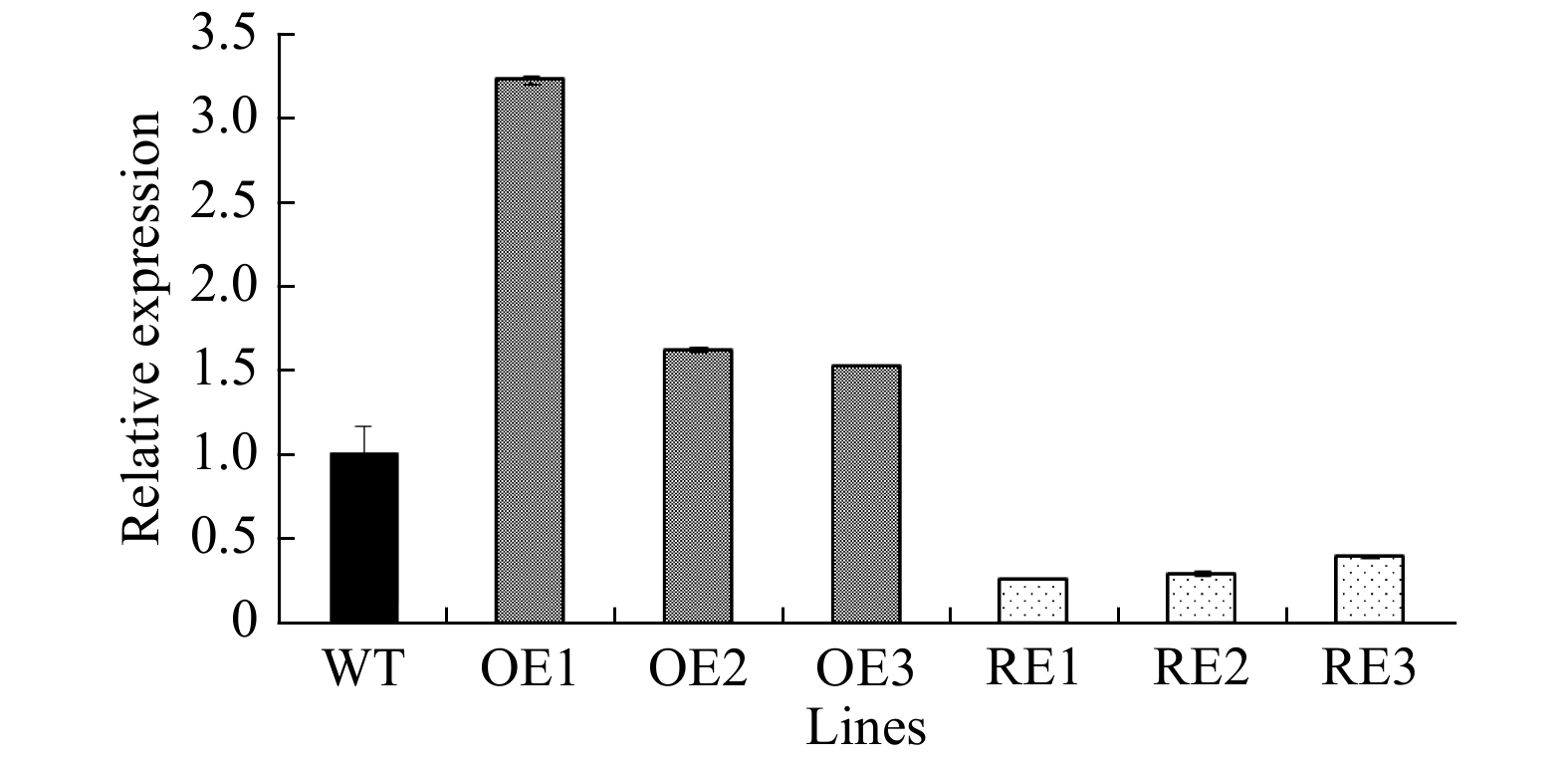
Figure 2. Relative PabGLK5 transcript levels in PabGLK5 overexpression lines and the total transcript levels of all five PabGLKs in PabGLK RNAi lines. OE, overexpression lines of PabGLK5; RE, repression lines of PabGLKs; WT, wildtype.
Phenotypic analysis of PaGLK transgenic poplars
-
The L* value can be used as an indicator of the brightness of leaf color in the CIELab color system, and a greater L* value indicates higher brightness. In addition, the b* value reflects the ratio of yellow/blue, and a greater b* value reveals increased yellow or decreased blue color. A colorimeter detected significantly reduced L* and b* values for PabGLK5 OE lines or showed no significant difference compared with those of different growing periods. By contrast, the L* and b* values for PabGLK RNAi lines were significantly higher than those of the WT. In particular, on July 15, the mean values of L* and b* for the three PabGLKs RNAi lines were 15.40% and 22.40% higher than those for the WT, respectively (Fig. 3a−c), suggesting that repressing the expression of PabGLKs not only turned P. alba × P. berolinensis leaves yellow but also enhanced the brightness, as measured by the CIELAB system.
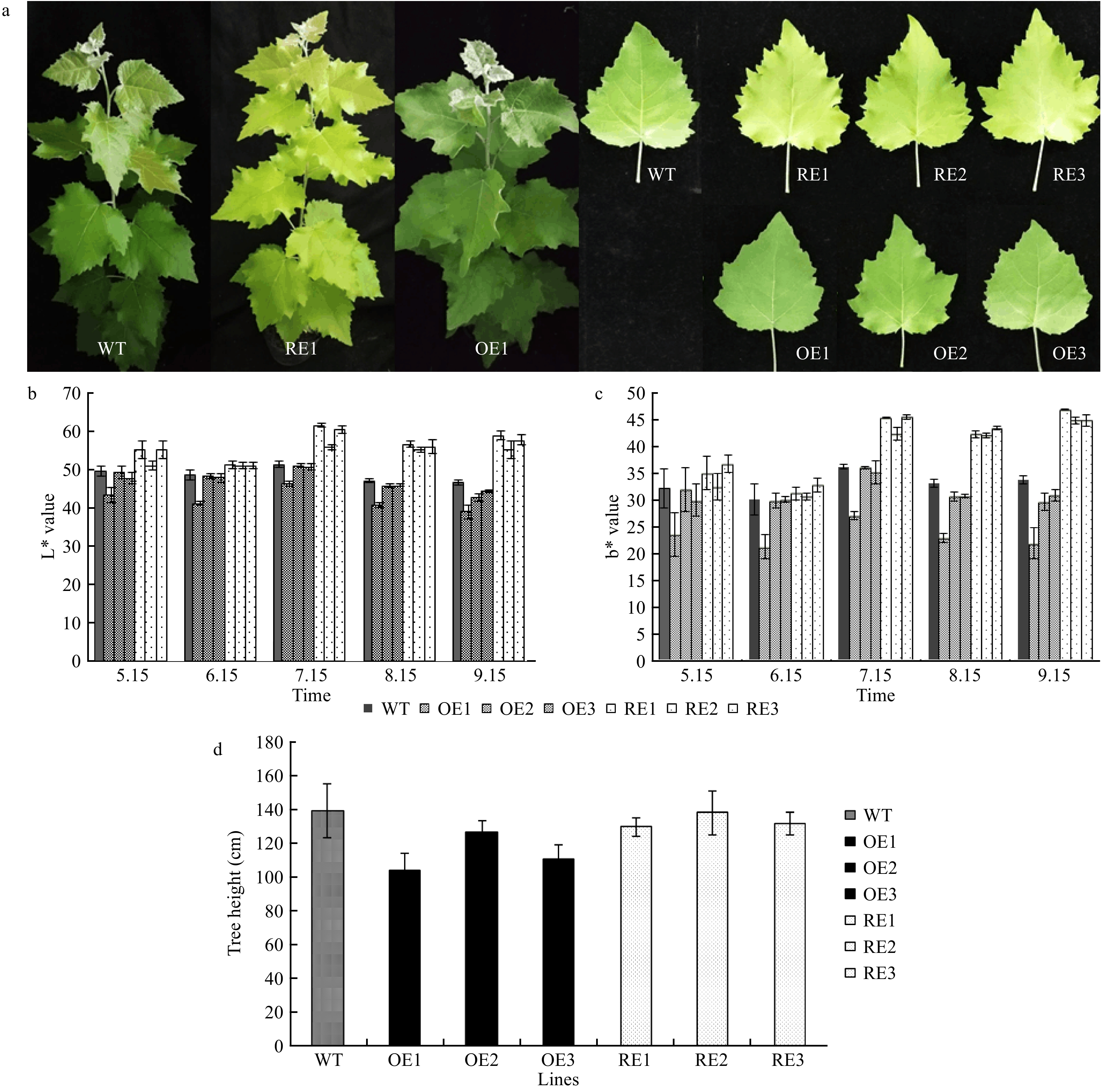
Figure 3. Phenotypic characterization of the PabGLK transgenic lines. (a): The yellow-green leaf phenotype of the PabGLK RNAi lines. (b) and (c): L* and b* values of the transgenic lines and WT measured by the CIELab color system. (d): Plant heights of one-year-old WT and transgenic lines. OE, overexpression lines of PabGLK5; RE, repression lines of PabGLKs; WT, wildtype.
The average heights of one-year-old PabGLK RNAi lines were comparable to that of the WT, whereas those of PabGLK5 overexpression lines (G line) were 18.23% lower than the WT (P < 0.05, Fig. 3d).
Determination of pigment contents and photosynthetic and fluorescence parameters
-
The contents of chlorophyll a, chlorophyll b, and carotenoids significantly increased by 28.5%, 65.9%, and 28.3%, respectively, in PaGLK5 overexpression lines compared with the WT. By contrast, the contents of chlorophyll a, chlorophyll b, and carotenoids in the PaGLK RNAi lines significantly decreased by 22.3%, 21.9%, and 21.7%, respectively, compared with the WT (Fig. 4a−c). The net photosynthesis (Pn) of PaGLK5 overexpression lines significantly decreased by 20.83% compared with the WT (except for OE3), whereas that of the PaGLK RNAi lines were significantly higher than the WT by 29.74% (P < 0.05, Fig. 4d). In contrast to the changes in pigment contents and Pn, none of the transgenic lines showed changes in Fv/Fm (Fig. 4e). Taken together, these results indicate that despite the reduction in the contents of photosynthetic pigments, the photosynthetic efficiency and the maximum photochemical quantum efficiency of PS II were not affected in PaGLK RNAi lines.
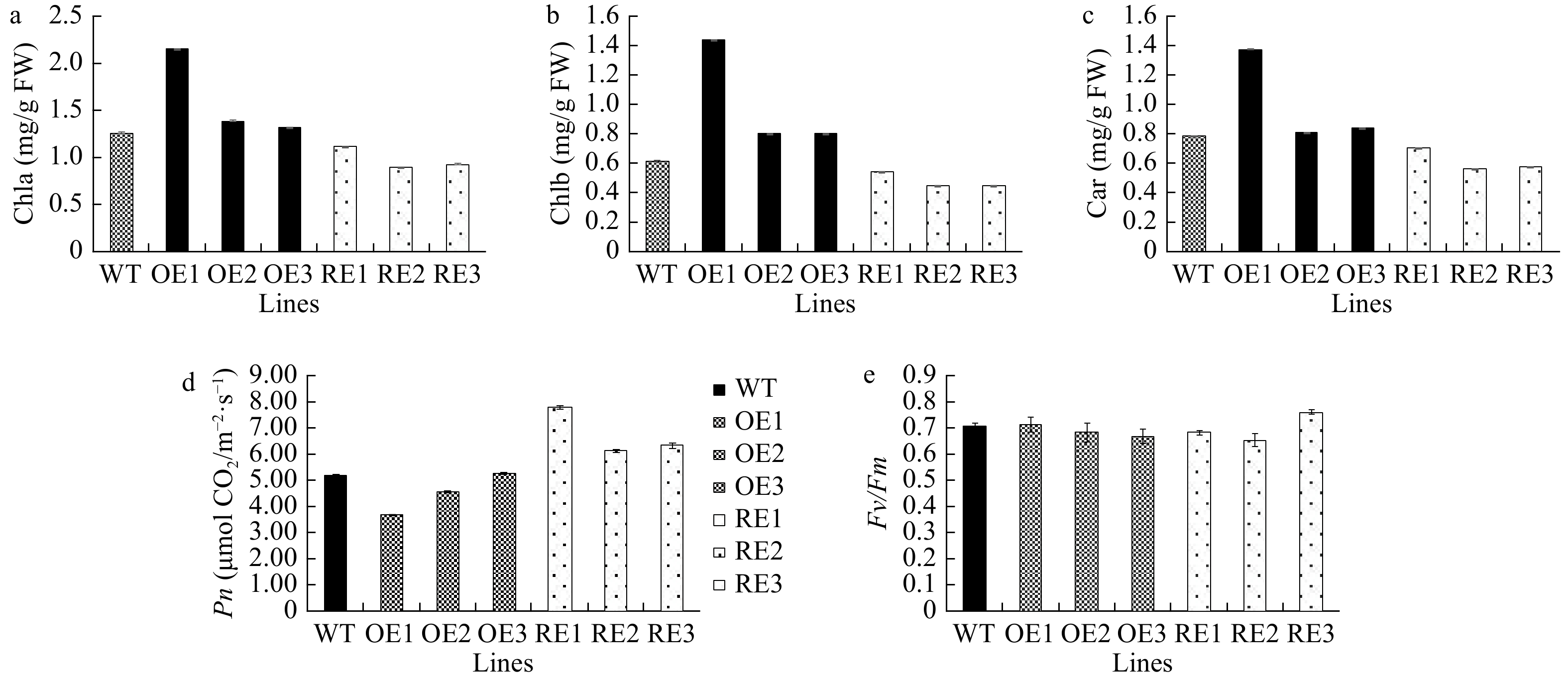
Figure 4. Pigment contents and photosynthetic and fluorescence parameters of the WT and transgenic lines. OE, overexpression lines of PabGLK5; RE, repression lines of PabGLKs; WT, wildtype. (a), (b) and (c) show the contents of chlorophyll a, chlorophyll b, and carotenoid in the WT and transgenic lines, respectively. (d) and (e) show photosynthetic parameters Pn and Fv/Fm, respectively.
Expression profile of photosynthesis-related genes
-
The expression profiles of genes encoding the light-harvesting complex components (PabLhcb1, PabLhcb2, PabLhcb4, and PabLhca3) and genes involved in chlorophyll synthesis (PabCHLD1, PabCHLD2) in the PabGLK RNAi lines and PabGLK5 overexpression lines were investigated using qPCR. The results showed that all the tested genes were significantly down-regulated in PabGLK RNAi lines but up-regulated in PabGLK5 overexpression lines compared with the WT (Fig. 5).
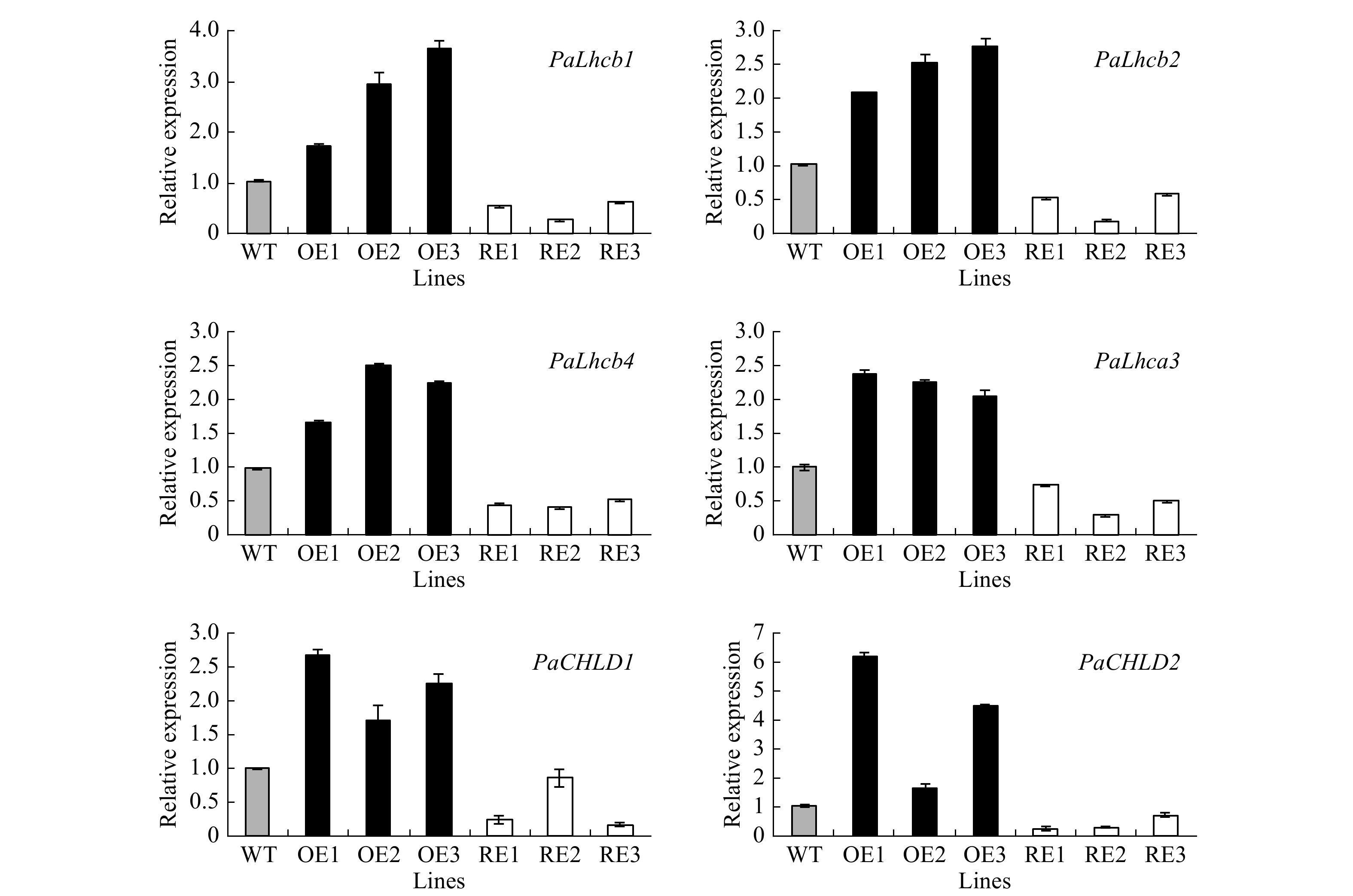
Figure 5. Expression profiles of photosynthesis-related genes in transgenic poplar. OE, overexpression lines of PabGLK5; RE, repression lines of PabGLKs; WT, wildtype.
-
Generation of the first transgenic plant in 1983 has brought about broad prospects for plant genetic engineering[14]. However, the application of transgenic plants has become increasingly controversial, especially with the drift of transgenic pollen, which is a major aspect for the ecological safety of transgenic plants[15]. The plant material used in this study is a triploid male clone selected from the cross between P. alba Linn and P. berolinensis Dipp. It has a straight trunk, gray bark, beautiful tree posture, and displays male infertility. The unique characteristics of this hybrid poplar make it an ideal material for urban gardening[16]. Gene drifting to non-transgenic and wild varieties can be avoided in transgenic poplar generated using this variety because of its male infertility. Thus, the golden leaves of the poplar generated here by genetic engineering are ideal for urban landscaping.
T-DNA insertion sites in transgenic plants are usually random, sometimes resulting in differences in phenotypes between transgenic lines[17]. Therefore, the generated transgenic lines often need to be selected for several generations, and only those that exhibit the desired phenotypes can be applied. Here, the plant height and growth rate of the PabGLK RNAi lines were not affected; the average height of one-year-old PabGLK RNAi plants was 132.58 cm, comparable to that of the WT. By contrast, the plant heights of one-year-old PabGLK5 overexpression lines were significantly lower than that of the WT, which may have been a result of the increased contents of photosynthetic pigments in the OE lines. The increased amount of pigments in the leaves of PabGLK5 OE lines may have absorbed excessive light energy, leading to light inhibition.
Studies of Arabidopsis and Betula platyphylla have shown that GLKs mainly affects the synthesis of photosynthetic pigments and the development of chloroplasts by regulating the expression of genes encoding light harvesting antenna proteins and photosystem complex proteins, as well as those involved in chlorophyll synthesis[10,3]. Grana are the thylakoid structures accumulated in the chloroplasts of terrestrial plants. Photosystem II and light-harvesting complex II (LHCII) are mainly concentrated in the stacked grana. Photosystem I and light-harvesting complex I (LHCI) are mainly concentrated in the stroma lamella. If the protein content of the grana lamella and stromal lamella is reduced in the chloroplasts, it affects the development of the chloroplasts[18]. The green-yellow leaf phenotypes caused by GLK repression are known to perform stably in the field in many species. Therefore, the GLK genes are top choice candidate genes for the generation of species with different leaf colors in tree breeding.
-
In this study, the 2 month-old tissue-cultured plantlets of P. alba × P. berlinensis were used for RNA extraction for GLK cloning and gene transformation. The plantlets were vegetatively cultured from apical meristems and stem segments excised from P. alba × P. berlinensis triplod plants that had been maintained in tissue culture room for multiple generations. The medium used was the half-strength Murashige and Skoog (1/2 MS) (Phytotech) containing 20 g/L sucrose with pH 5.8 before adding 3 g l-1 Gelrite (Duchefa). The karyotype analysis of this P. alba × P. berlinensis triploid, which had been planted in the Northeast region of China for many years, was conducted earlier[19].
Identification of PabGLKs
-
Total RNA was extracted from the leaves of the P. alba × P. berlinensis triploid plantlets using the CTAB method[20]. An RNA sequencing library was constructed using the NEBNext RNA Library Prep Kit for Illumina (New England Biolabs, Ipswich, MA, USA). The cDNA library was then sequenced on an Illumina sequencing platform (HiSeq™ 4 000). The resulting reads were qualified using FastQC and assembled into contigs using Trinity[21] with default parameters. The assembled contigs were BLASTed[22] against the transcripts of P. trichocarpa[23] to identify expressed GLKs in the leaves of the triploid poplar. A multiple sequence alignment of GLKs from P. trichocarpa, P. alba × P. berlinensis, and Arabidopsis was performed using MUSCLE program[24] and a neighbor-joining phelogenetic tree was constructed using a bootstrap of 1 000 replicates in MEGA6[25].
Tissue specific expression profiles of PabGLKs
-
Total RNA was extracted from the roots, stems, and leaves of two-month old P. alba × P. berlinensis triploid plantlets using the CTAB method[26,27], and cDNA was synthesized using the ReverTra Ace qPCR RT Master Mix with gDNA Remover (Toyobo, Osaka, Japan). qPCR was performed using SYBR Green Real-Time PCR Master Mix Plus (Toyobo) following the manufacturer’s recommendations on an ABI PRISM 7 500 Real-Time PCR system (Applied Biosystems, Foster City, CA, USA). The relative transcript levels of the target genes were normalized to those of 18S rRNA and α-Tubulin, by the 2−ΔΔCT method[28].
Generation of transgenic poplar
-
To repress the expression of PabGLKs, a 200-bp fragment of PabGLK5 was cloned from leaves of P. alba × P. berlinensis and inserted into the RNAi vector pFGC5941 in forward and reverse directions. Primers used for RNAi vector construction were listed in Table 1. To overexpress PabGLK5, the full-length cDNA sequence of PabGLK5 was cloned into the plant expression vector pCAMBIA1300. The resulting RNAi and overexpression constructs were introduced into the P. alba × P. berlinensis genome through Agrobacterium-mediated transformation to generate PabGLK repression and PabGLK5 overexpressing lines. Genetic transformation was performed as described previously[22].
Table 1. Primers used for RNAi vector construction.
Primers Sequences (5'-3') PaGLK-RNAi-Cis-NcoI CATGCCATGGCCATCCCCAATGCATATGTG PaGLK-RNAi-Cis-AscI TTGGCGCGCCGCAGGAAATCTAGTTGCCAGT PaGLK-RNAi-Anti- XbaI GCTCTAGACCATCCCCAATGCATATGTG PaGLK-RNAi-Anti-BamHI CGCGGATCCGCAGGAAATCTAGTTGCCAGT Determination of pigment contents and photosynthetic parameters
-
Chl and carotenoids (Car) were extracted with 80% acetone at 4 °C for 24 h in the dark and measured at 470, 645, and 663 nm with a UV-Vis spectrophotometer (TU-1901, PERSEE, Suzhou, China). Three individual plants were sampled for each line, and three leaves per plant were measured. The photosynthetic and kinetic parameters of Chl fluorescence were measured for the mature leaves of the WT and transgenic lines using a portable photosynthesis system (Li-6400, LI-COR, Nebraska USA).
Measurement of leaf colors of transgenic lines
-
A spectrocolorimeter (KONICA MINOLTA CR-400) was used to measure the leaf colors of the transgenic lines. For each line, 30 different individual plants were measured, with one leaf per plant. The measurement starts on 15 May and ends on 15 September, and the measurement was performed once in every 30 days. The measurement results were analyzed by the CIELab color system, where L* represents the brightness (Lumination) value between black and white, with a value in the range of 0−100, while b* represents blue-yellow opponents with positive numbers toward yellow and negative numbers toward blue (yellow (+b) blue (−b)).
Statistical analysis
-
SPSS statistical analysis software and Excel software were used for variance analysis and multiple comparisons.
-
This study was supported by the Applied Technology Research and Development Planning Program of Heilongjiang Province (GA19B201), the Innovation Project of the State Key Laboratory of Tree Genetics and Breeding (Northeast Forestry University) (2014A02), the Heilongjiang Touyan Innovation Team Program (Tree Genetics and Breeding Innovation Team), and the 111 Project (B16010).
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Y, Gu C, Gang H, Zheng Y, Liu G, et al. 2021. Generation of a Golden Leaf Triploid Poplar by Repressing the Expression of GLK Genes. Forestry Research 1: 3 doi: 10.48130/FR-2021-0003

Generation of a Golden Leaf Triploid Poplar by Repressing the Expression of GLK Genes
- Received: 18 December 2020
- Accepted: 07 January 2021
- Published online: 29 January 2021
Abstract: Poplar trees are excellent varieties widely used for gardening and greening. However, their single color and floating fluffy seeds are major disadvantages. Plant species or varieties with variegated leaves are desperately needed to meet various demands for gardens, urban greening and landscape decoration, as they produce rich foliage colors that are aesthetically pleasing and functional. In this study, we generated a golden leaf triploid poplar (P. alba × P. berlinensis) by repressing the expression of GLK (Golden2-like) genes in leaves. The triploid golden leaf poplar had reduced chlorophyll content but almost no change in the growth rate. It has great potential in landscaping once it passes safety assessments.
-
Key words:
- GLK /
- Poplar /
- Colorful leaf /
- Yellow green leaf