








-
Iron (Fe) is one of the key micronutrients required for plants. It plays an important role in various physiological pathways, such as chlorophyll synthesis and photosynthesis[1−3]. Fe exists as insoluble hydroxides and oxides in most soils, which greatly reduces its availability for plants[3,4]. Fe deficiency therefore has become a widespread issue in agriculture, decreasing crop yield[5,6]. It also severely affects the growth of apple trees, resulting in decreased yield and poor quality of apple[7].
To cope with Fe deficiency, two major efficient mechanisms have evolved to absorb and utilize Fe in plants: the reduction mechanism (named strategy I) used mainly by dicots and non-grass monocots, and the chelation mechanism (named strategy II) used mainly by grasses[8,9]. Apple trees use strategy I to initiate the response to Fe deficiency, in which H+-ATPases-mediated protons are extruded from the root cell to acidify the rhizosphere and subsequently solubilize iron for absorption[9,10]. In modern orchards, the application of a Fe nutrition agent is not efficient to relieve apple plants from Fe-deficiency stress, even at the cost of environmental pollution[7]. Therefore, it is important to find an alternative way to improve Fe tolerance in apple.
Approximately 90% of total Fe in leaves exists in the chloroplasts, which is necessary for maintaining function[11−13]. The chloroplast is the vital site for chlorophyll synthesis and photosynthesis in plants. Fe deficiency directly induces excessive reactive oxygen species (ROS) production, and consumes antioxidant enzymes in plants, such as superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), etc, which causes the oxidative damage of the chloroplast and subsequently photosynthesis in plants[14,15]. Consequently, the chlorosis of leaves as a result of chlorophyll degradation is a typical symptom of Fe deficiency stress, which unavoidably leads to decreased photosynthetic efficiency[16,17]. Therefore, protecting chloroplasts from excessive ROS-mediated oxidative damage is critical to relieve plants from Fe deficiency.
ROS is known as an important second messenger for signal transduction that regulates many physiological processes, in the early response to various stresses. But severe stress-mediated ROS burst in plants, if not scavenged timely and properly, will trigger oxidative damage and even programed cell death[18,19]. To protect plants from oxidative damage under different stresses, two antioxidant mechanisms have evolved to balance the in vivo level of ROS, including the antioxidant mechanism and antioxidant enzyme mechanism[20,21]. Antioxidants include tocopherol, ascorbic acid, carotenoids, glutathione and antioxidant enzymes include SOD, POD, CAT, which orchestrate the oxidative protection of plants under various stresses[18].
Melatonin, a well-known antioxidant for efficient ROS-scavenging, was first reported in plants in 1995[22,23]. Different from other antioxidants, melatonin possesses a amphiphilic feature, which ensures that it can move freely between all cellular compartments, including chloroplasts, mitochondria, cytosol and so on. Additionally, both melatonin and its metabolites can scavenge different kinds of ROS including superoxide anion (O2•−) and hydrogen peroxide (H2O2), which evokes the cascade amplification effect of melatonin on the ROS-scavenging[24]. Therefore, it is reported as the strongest ROS scavenger compared to other antioxidants. Melatonin is proven to protect plants against various abiotic and biotic stresses, through scavenging ROS timely and properly[25−28]. Four enzymes catalyze its classic biosynthetic pathway in plants sequentially, including tryptophan decarboxylase (TDC), tryptamine 5-hydroxylase (T5H), serotonin N-acetyltransferase (SNAT/AANAT), and N-acetylserotonin O-methyltransferase (ASMT). Among them, ASMT and SNAT are known as the possible rate-limiting enzymes catalyzing melatonin synthesis in plants. They can catalyze melatonin synthesis in alternate order[24]. Our previous research has proved that the chloroplast is the major organelle synthesizing melatonin in plant[29]. Melatonin protects the chloroplast via scavenging ROS in situ and therefore improves the efficiency of photosynthesis[29,30]. Apple melatonin synthetase ASMT9 is localized in the chloroplast. It is speculated that melatonin might promote resistance to Fe deficiency stress through scavenging ROS and protecting the chloroplast, which remains unknown in apple.
In this study, Malus baccata (Linn.) Borkh. (M. baccata), an Fe-deficiency sensitive apple rootstock, was used. The Fe deficiency stress-induced ROS burst in M. baccata increased melatonin level and decreased the activities of antioxidant enzymes. Exogenous melatonin could decrease leaf chlorosis in M. baccata under Fe deficiency, reduce chlorophyll degradation and therefore stabilise photosynthetic rate, simultaneously increasing Fe content in leaves. Meanwhile, exogenous melatonin induced the increased endogenous melatonin level and the enhanced activities of antioxidant enzymes, which resulted in the efficient scavenging of ROS under Fe deficiency. Interestingly, exogenous melatonin induced the expression of melatonin synthetase gene in M. baccata, but the protein level of synthetase declined after melatonin treatment, inferring post-transcriptional regulation for synthetase in apple. This study contributes to the regulation mechanism of melatonin biosynthesis in response to stress, and provides a novel way to improve Fe tolerance for apple production.
-
It is well-known that the chlorosis of leaves is a typical symptom for Fe deficiency stress in plants. To observe the effects of Fe deficiency on apple plants, the M. baccata plants were supplemented with 4 μM (Fe deficiency group) and 40 μM (normal group, named as CK) iron nutrient solution, respectively. The results showed that, before Fe deficiency treatment, the growth condition of M. baccata plants were similar, and all of the leaves were green. After 14 d of treatment, the apple leaves turned yellow under Fe deficiency, and the leaves in the normal group were still green (Fig. 1a). Furthermore, to explore the effects of melatonin on the tolerance to Fe deficiency in apple, both the Fe deficient and control groups were treated with 50 μM, 75 μM and 100 μM melatonin. Similar phenotypes were observed, 75 μM melatonin concentration was selected for further experiments. The results showed that under normal growth conditions, the MT-treated plants of M. baccata grew faster and had more leaves than the control (Fig. 1a). Compared with the normal plants which had leaf chlorosis under Fe deficiency, the MT-treated leaves were still green (Fig. 1a). In addition, the plants with melatonin grew better than those without melatonin under Fe deficiency (Fig. 1a). It is therefore suggested that melatonin enhances the tolerance to Fe deficiency in M. baccata.
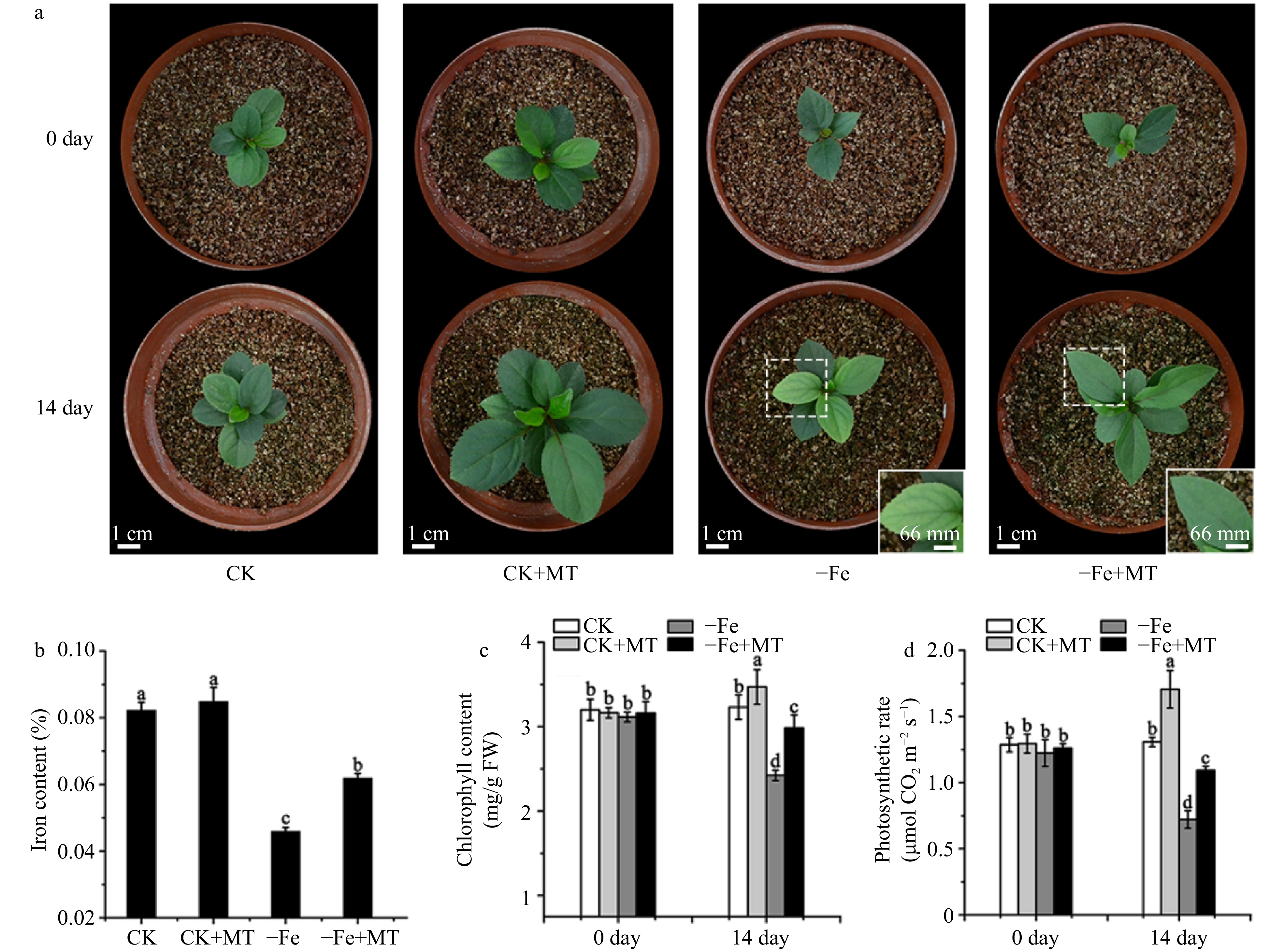
Figure 1. Phenotypes of M. baccata apple plants under Fe deficiency or normal growth conditions with and without melatonin. (a) The growth conditions of M. baccata plants at 0 and 14 d, respectively. Bar: 1 cm or 66 mm, respectively. (b) The Fe content in leaves under Fe deficiency for 14 d. (c) The chlorophyll content in leaves at 0 and 14 d, respectively. (d) The photosynthetic rate of M. baccata apple plants at 0 and 14 d, respectively. CK: plants under normal growth conditions (40 μM Fe); −Fe: plants under Fe deficient conditions (4 μM Fe); MT: melatonin. Data are means ± SD of three independent experiments. Lowercase letters: significant differences based on LSD multiple comparison (p < 0.05).
Application of melatonin improved Fe content, reduced chlorophyll degradation and maintained photosynthesis in M. baccata under Fe deficiency
-
To further confirm the effects of melatonin on the resistance to Fe deficiency, the Fe content of the leaves was analysed. The results showed that under normal growth conditions, the Fe content of leaves with melatonin and without melatonin had no significant change. But under Fe deficiency stress, melatonin treatment significantly improved the Fe content of leaves (Fig. 1b). The chlorosis of leaves results from the decreased chlorophyll level, which further triggered the reduced photosynthetic rate in plants, so chlorophyll level and photosynthetic rate were measured in the leaves of M. baccata. The results showed that prior to Fe deficiency or melatonin treatment, chlorophyll content and photosynthetic rate in four groups of M. baccata had no obvious differences. After 14 d, the chlorophyll content (2.4 mg/g FW) in the Fe-deficiency group was lower than that in the normal group (3.2 mg/g FW), suggesting Fe-deficiency induced the degradation of chlorophyll. But when treated with melatonin, the chlorophyll content of leaves (2.95 mg/g FW) was increased dramatically compared with that without melatonin (2.4 mg/g FW) under Fe deficiency (Fig. 1c). Further, the photosynthetic rate in the Fe-deficiency group (0.7 μmol·m−2·s−1) was significantly lower than that in the normal group (1.2 μmol·m−2·s−1) after 14 d. However, photosynthetic rate was still kept at 1.1 μmol·m−2·s−1 with exogenous melatonin under Fe deficiency (Fig. 1d). It implied that exogenous melatonin might protect the chloroplasts under Fe deficiency by reducing degradation of chlorophyll and consequently maintaining photosynthesis.
Melatonin alleviated oxidative damage induced by Fe deficiency
-
It is known that Fe deficiency induces oxidative damage in plants. The results showed that there were little O2•− and H2O2 production in leaves under normal growth conditions, but under Fe deficiency, O2•− and H2O2 production in leaves was significantly increased (Fig. 2a). This suggests that Fe deficiency induces ROS burst in M. baccata. With melatonin treatment, the intensities of histochemical staining in leaves were obviously weaker, suggesting melatonin dramatically reduces the production of O2•− and H2O2 under Fe deficiency (Fig. 2a). In addition, compared with the normal growth conditions, Fe deficiency dramatically induces the malondialdehyde (MDA) level in leaves of M. baccata. MDA levels indicate the degree of oxidative damage in plants. It suggested that Fe deficiency induces severe oxidative damage in M. baccata. But exogenous melatonin dramatically reduces the MDA level of leaves under Fe deficiency and normal growth conditions (Fig. 2b). The results suggest that exogenous melatonin treatment alleviates oxidative damage induced by Fe deficiency, through scavenging O2•− and H2O2 and consequently decreasing MDA content.

Figure 2. ROS and MDA level in M. baccata plants under Fe deficiency or normal growth conditions with and without melatonin. (a) The O2•− and H2O2 levels in leaves. (b) The MDA levels in leaves. CK: plants under normal growth conditions (40 μM Fe); −Fe: plants under Fe deficient conditions (4 μM Fe); MT: melatonin. Data are means ± SD of three independent experiments. Lowercase letters: significant differences based on LSD multiple comparison (p < 0.05).
Exogenous melatonin promoted the level of melatonin and the activities of antioxidant enzymes in M. baccata plants
-
To determine the change of endogenous melatonin level, melatonin content in M. baccata plants was detected by HPLC as described by Zheng et al.[30]. Before Fe deficiency or melatonin treatment, the melatonin level in four groups had no obvious differences. Under Fe deficiency for 14 d, the melatonin level in M. baccata plants was increased by approximately three times that of those under normal growth conditions (Fig. 3a). It suggested that Fe deficiency stress could induce the synthesis of endogenous melatonin, which might result from the ROS burst triggered by Fe deficiency. After treatment with exogenous melatonin for 14 d, the melatonin content was increased about 2.4 times of that without melatonin under normal growth conditions. And exogenous melatonin treatment slightly improved the endogenous melatonin level under Fe deficiency (Fig. 3a). The activities of antioxidant enzymes (SOD, POD and CAT) were also measured in M. baccata plants. The results showed that the activities of SOD, POD and CAT were dramatically decreased under Fe deficiency compared with that under normal growth conditions, but their activities were increased significantly when treated with melatonin under Fe deficiency or normal growth conditions (Fig. 3b−3d). The results suggest that exogenous melatonin treatment improves the in vivo level of melatonin and the activities of antioxidant enzymes, which might further promote the scavenging of ROS under Fe deficiency in M. baccata.
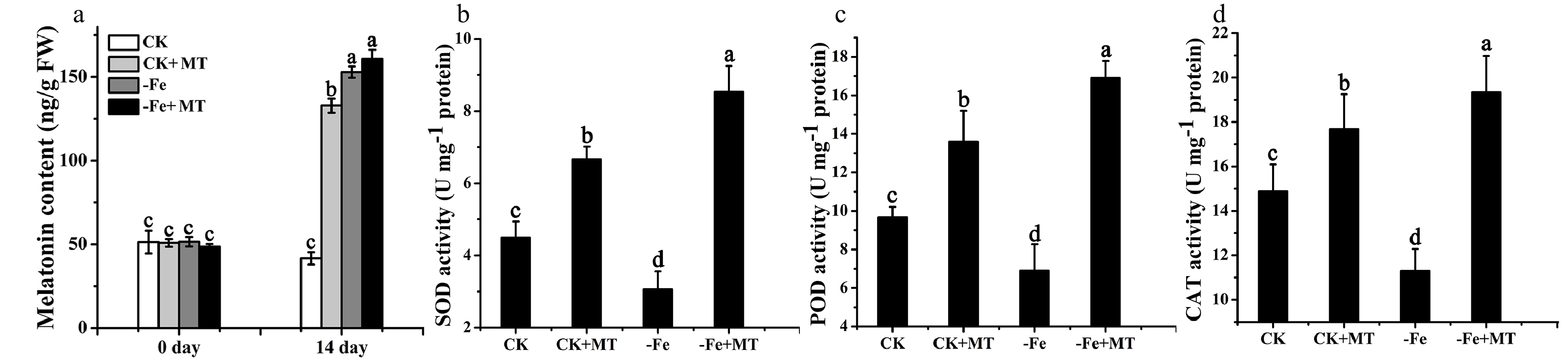
Figure 3. Melatonin content and activities of antioxidant enzymes in M. baccata plants under Fe deficiency or normal growth conditions, with and without melatonin. (a) Melatonin content of M. baccata plants detected by HPLC. The activities of SOD (b), POD (c) and CAT (d) in leaves. CK: plants under normal growth conditions (40 μM Fe); −Fe: plants under Fe deficiency conditions (4 μM Fe); MT: melatonin. Data are means ± SD of three independent experiments. Lowercase letters: significant differences based on LSD multiple comparison (p < 0.05).
Exogenous melatonin improved the expression of melatonin synthetase genes, but reduced protein levels in M. baccata plants
-
To further investigate the effects of melatonin treatment on the synthesis of endogenous melatonin, the expression of melatonin synthetase was detected in M. baccata plants. The results showed that the expression of synthetase genes (MbASMT9, MbAANAT3 and MbTDC1) was increased significantly under Fe deficiency, which was in line with the change in melatonin. And exogenous melatonin treatment improved the expression of synthetase genes under Fe deficiency or normal growth conditions (Fig. 4a). Interestingly however, the protein level of MbASMT9 declined after exogenous melatonin treatment under Fe deficiency and normal growth conditions (Fig. 4b), suggesting the existance of post-transcriptional regulation for ASMT9 in M. baccata. The results implied that there is likely a complicated mechanism in M. baccata, which could balance the synthesis of endogenous melatonin regulated by exogenous melatonin.
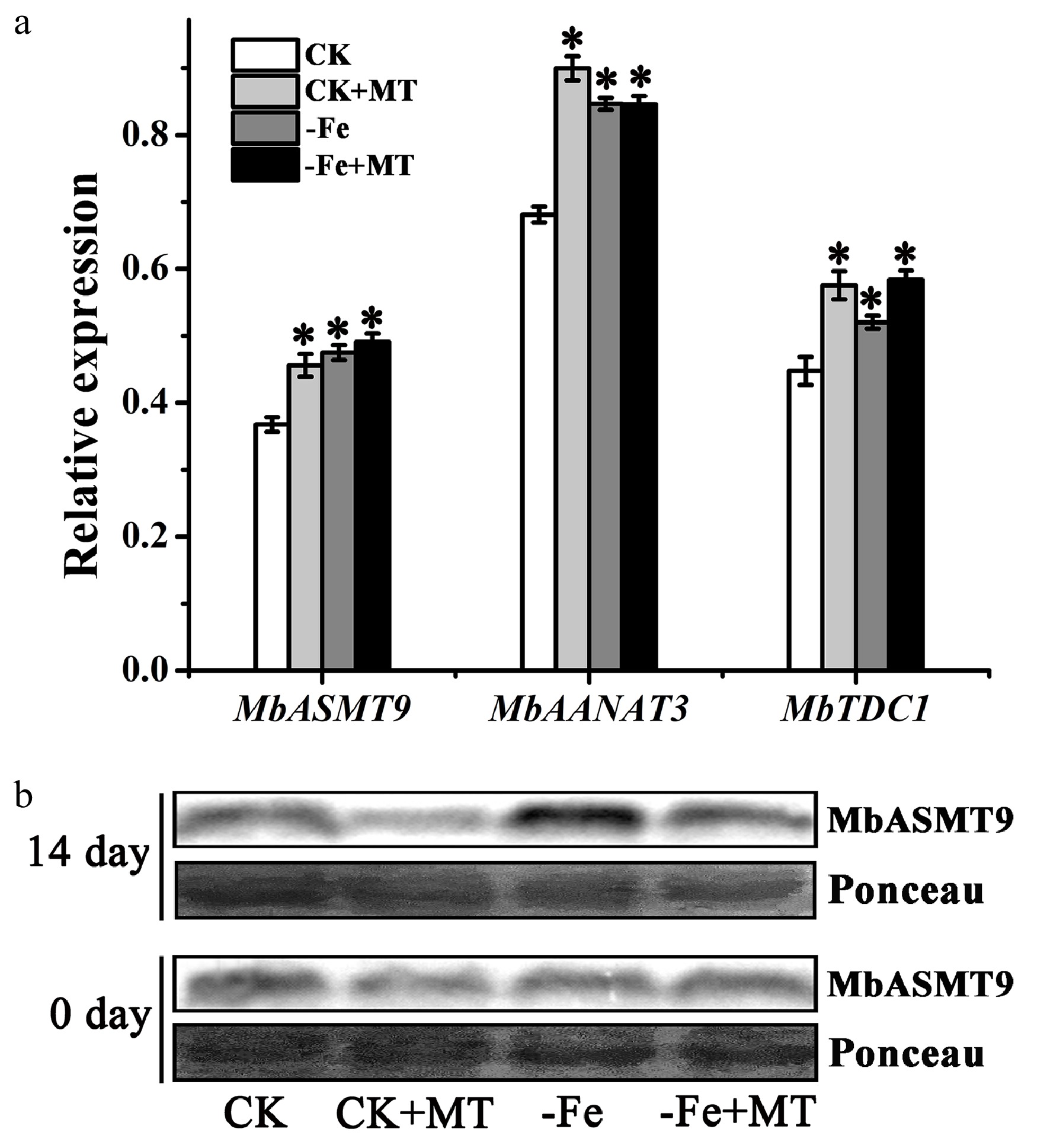
Figure 4. Expression of melatonin synthetase in M. baccata plants under Fe deficiency or normal growth conditions, with and without melatonin. (a) Relative expression of MbASMT9, MbAANAT3 and MbTDC1 genes detected by qPCR after 14 d. (b) Protein level of MbASMT9 detected by western blot. Ponceau stained bands indicated the quantity loading of total protein. Data are means ± SD of three separate experiments. * p < 0.05 (Student’s t-test).
-
Fe deficiency has become a severe problem that affects the yield and quality of apple due to the poor availability caused by its insolubility in most soils[5−7]. Currently, there is a lack of efficient and environmently-friendly ways to cope with Fe deficiency in a modern orchard. Hence, it is important to find a way to improve Fe deficiency resistance to improve apple yield and quality in cultivated land.
Based on previous research, melatonin, as an efficient ROS scavenger, had been applied to improve resistance to various stresses such as drought, salt and waterlogging in apple[30−32]. For the first time, we found that melatonin enhances the tolerance to Fe deficiency stress in M. baccata, an Fe-deficiency sensitive apple rootstock. Exogenous melatonin relieved apple plants from Fe deficiency stress, which was indicated by increased Fe content, decreased chlorophyll degradation, reduced leave chlorosis, and subsequently maintained photosynthesis (Fig. 1). As melatonin may inhibit Fe oxidation in soil (due to its strong reducibility), which results in poor levels of Fe available to roots, exogenous melatonin wasn’t sprayed on the soil in this study. Similarly, there were reports regarding the protective function of melatonin on chlorophyll and photosynthesis under drought, salt and waterlogging stresses in apple[30,32−34]. The results implied that exogenous melatonin might protect chloroplasts under Fe deficiency by reducing degradation of chlorophyll and consequently maintaining photosynthesis, which results in the enhanced tolerance to Fe deficiency in apple.
As a well-known ROS scavenger, the involvement of melatonin in timely migration of excessive ROS under Fe deficiency was detected. The results showed that Fe deficiency significantly induced the excessive production of O2•− and H2O2 in leaves of M. baccata, and consequently the MDA level, as an important indicator of oxidative damage, were increased under Fe deficiency (Fig. 2a and b). But exogenous melatonin dramatically reduced the levels of O2•−, H2O2 and MDA in leaves of M. baccata (Fig. 2a and 2b). In addition, the trend of melatonin level, which contained the absorbed-melatonin from exogenous melatonin treatment and the endogenous synthetic-melatonin, was contrary to the change trend of ROS and oxidative damage under Fe deficiency (Fig. 2 and Fig. 3a). This suggests that melatonin alleviated the oxidative damage induced by Fe deficiency through efficiently scavenging ROS. Plants have already evolved the systems of antioxidants and antioxidant enzymes to eliminate excessive ROS induced by various stresses[24]. Therefore, we also detected the activities of antioxidant enzymes in M. baccata. We found that Fe deficiency reduced the activities of antioxidant enzymes (SOD, POD and CAT), but exogenous melatonin significantly improved their activities under Fe deficiency in M. baccata (Fig. 3b−3d), which further promoted the scavenging of ROS and consequently decreased the oxidative damage under Fe deficiency. Under other stresses, melatonin has been also proved to promote the activity of antioxidant enzyme systems in apple[30−32,34]. The results suggested that exogenous melatonin played important roles in the protection of essential antioxidant enzyme systems in vivo. Except for their activities, we speculated that melatonin might affect the synthesis of the antioxidant enzymes, which might result in their increased total protein level. This requires further research.
Furthermore, based on the effects of exogenous melatonin on Fe deficiency tolerance, it was inferred that the change of melatonin level from external absorption and internal synthesis might play a role in the Fe deficiency response. Therefore, the underlying mechanism of melatonin on the response to Fe deficiency was further explored in vivo. We found that Fe deficiency significantly induced the expression of melatonin synthetase genes MbASMT9, MbAANAT3 and MbTDC1. Exogenous melatonin treatment also improved their expression in M. baccata (Fig. 4a), which was in line with the change in melatonin. Our previous research had already proved that ASMT9 is the key melatonin synthetase localized in the chloroplast[29]. Further, the protein level of ASMT9 was also detected in M. baccata. Interestingly, Fe deficiency stress induced its protein level, but exogenous melatonin treatment reduced its protein level under Fe deficiency or normal growth conditions (Fig. 4b), implying post-transcriptional regulation may exist for ASMT9. It was speculated that when exogenous melatonin could meet the requirements of plants, the post-transcriptional regulation trend was to save energy to avoid extra melatonin production. A similar result was previously reported that the post-transcriptional regulation of ASMT9 under waterlogging stress in apple exists[30], which was further verified by our results. It suggested that a complicated mechanism in M. baccata may exist
, which could balance the synthesis of endogenous melatonin regulated by exogenous melatonin to ensure the balance of melatonin levels under different stresses. The findings contributed to the application of melatonin to enhance the tolerance of apple to various stresses. Taken together, we summarized a model for the protective functions of melatonin against Fe deficiency stress in apple (Fig. 5). Fe deficiency induced the ROS burst, which resulted in the degradation of chlorophyll, decreased photosynthesis, and consequently the chlorosis of leaves in apple. Furthermore, the ROS burst triggered by Fe deficiency induced the synthesis of endogenous melatonin. Exogenous melatonin increased the synthesis of endogenous melatonin and the activities of antioxidant enzymes, which further alleviated the oxidative damage triggered by Fe deficiency, through efficiently scavenging ROS. Interestingly, melatonin induced the expression of melatonin synthetase genes, but reduced the protein level of synthetase, suggesting that post-transcriptional regulation may exist, which balances the melatonin level to ensure the efficient scavenging of ROS and consequently protect chloroplasts in apple. Hence, the leaves of apple were still green under Fe deficiency because of melatonin. This work not only revealed the mechanism of melatonin on Fe deficiency tolerance, but also provided a potential way of relieving plants of Fe deficiency stress for apple production.
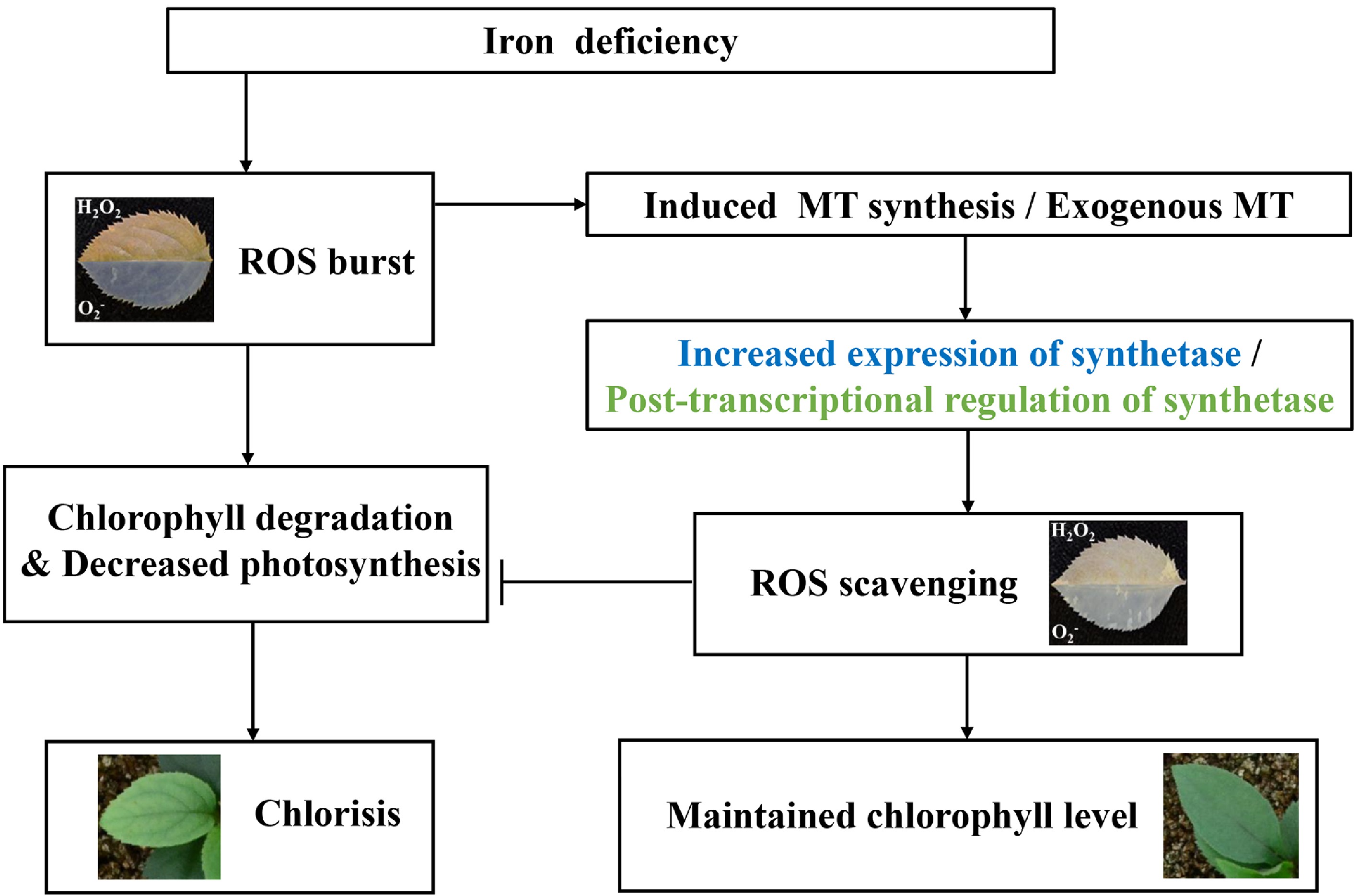
Figure 5. The speculated model regarding the protective functions of melatonin against Fe deficiency stress in apple. Fe deficiency induced the ROS burst, which resulted in the degradation of chlorophyll and decreased photosynthesis. It led to the chlorosis of leaves in apple. Furthermore, the ROS burst triggered by Fe deficiency induced the synthesis of endogenous melatonin. Endogenous or exogenous melatonin alleviated the oxidative damage triggered by Fe deficiency, through scavenging ROS and consequently protecting chloroplasts in apple. Hence, the leaves of apple remained green under Fe deficiency. Arrows: the stimulation; Even bars: the inhibition.
-
After vernalization, seeds of apple (M. baccata) were sowed according to Zheng et al.[30]. Two weeks later, the seedlings were supplied with half-strength Hoagland’s nutrient solution. A week later, they were watered with complete nutrient solution[30]. The plants were grown in a culture chamber in 16/8 h light/dark cycle at 23 ± 2 °C, with a light intensity of 100 μmol·m−2·s−1.
Treatment of Fe deficiency and exogenous melatonin
-
One-month old M. baccata plants were divided into two groups, each of which had 12 seedlings. The plants in group I (Fe-deficienct group) and group II (normal group as a control) were watered with 4 μM and 40 μM iron nutrient solution every 3 d, respectively. Samples were collected before the Fe deficiency treatment (namely 0 d) and 14 d after the Fe deficiency treatment for further study, respectively.
For melatonin treatment, the stock solution was prepared as described by Zheng et al.[30], which was diluted with deionized water into 75 μM to spray the M. baccata leaves. Fe-deficient and normal groups were divided into two groups, respectively: one group was sprayed with 75 μM melatonin, and another was sprayed with water containing the same dilution ratio of ethanol. The leaves were sprayed every 2 d, and sampled at 0 and 14 d, respectively. The experiment was repeated independently in triplicate.
Measurement of Fe content
-
After 14-d treatment, the collected leaves were washed using distilled water several times to remove impurities. After dehydration at 105 °C for 30 min and continuously at 80 °C for 72 h, they were ground to powder and digested in 12 mL HNO3 and HClO4 (HNO3 : HClO4 = 5:1). The extraction was diluted with deionized water to 25 mL[35]. The Fe content was detected as described by Su et al.[36]. Each experiment was conducted independently in triplicate.
Determination of chlorophyll (Chl) and photosynthetic rate (Pn)
-
The extraction and detection of Chl was conducted as described by Zheng et al.[30]. A total of 0.2 g fresh leaves were sampled and chopped to homogenize in 3 mL 80% acetone. The homogenate was collected through filtration for diluting with 80% acetone to 25 mL. The concentration of Chl was determined based on the absorbance at 645 nm and 663 nm, respectively[30].
The photosynthetic rate was measured according to Su et al.[36]. The light intensity was set at 800 μmol·m−2·s−1. The CO2 concentration was controlled at 400 ppm using an automatic CO2 controller. Each experiment was conducted in triplicate.
Detection of O2•−, H2O2, malondialdehyde (MDA) and melatonin level
-
To detect O2•− and H2O2 levels, fresh leaves in each group were collected. The test was performed according to Zheng et al.[30]. For O2•− detection, the leaves were infiltrated in 25 mM K-HEPES solution (pH 7.9) dissolving 0.1 mg·mL−1 nitroblue tetrazolium for 40 min in a vacuum pump. After histochemical staining, the leaves were placed in the dark at 25 °C for 4 h. For H2O2 measurement, the leaves were infiltrated in 50 mM Tris-acetate (pH 3.8) containing 0.1 mg·mL−1 3, 3’-diaminobenzidine for 30 min in a vacuum pump. The leaves were kept in the dark at 25 °C for 24 h. After dark treatment, the leaves were soaked in 80% ethanol at 80 °C for 10 min, this was repeated until the green color of the leaves was removed. The leaves were observed and photographed.
MDA detection in leaves was conducted according to Zhao et al.[37]. 1 g leaves were ground to powder, homogenized in 10 mL 10% trichloroacetic acid and centrifuged. 2 mL of supernatant was collected, mixed with 2 mL 0.6% thiobarbituric acid, and heated at 100 °C for 20 min. The samples were immediately cooled and centrifuged, the supernatant was collected for the measurement of absorbance and the calculation of MDA content as described by Zhao et al.[37].
For melatonin measurement, around 1 g of ground leaves was ultra-sonicated (80 Hz) in 10 mL methanol at 45 °C for 35 min. After centrifugation, the supernatants were collected for purification as described by Zhao et al.[37]. The melatonin level was detected by HPLC.[37]. The experiments were conducted independently in triplicate.
Quantitative reverse transcription PCR (qPCR) analysis
-
Preparation of total RNA and cDNA was performed according to Zheng et al.[30]. The primers were designed using Primer 5 software according to the candidates MbASMT9, MbAANAT3, and MbTDC1 gene sequences, respectively (Primers of MbASMT9 and MbAANAT3 were referred to Zheng et al.[30]; MbTDC1 Forward Primer 5’-TCACGCTGTGGTTGGAGGT-3’, Reverse Primer 5’-CTGCATGCTCCTGAACCAAC-3’). The qPCR assay was conducted with Actin gene as an internal control as described by Zheng et al.[38]. The experiment was repeated in triplicate.
Test of antioxidant enzyme activities
-
Around 0.3 g of leaves from M. baccata plants from each group were ground, homogenized in 8 mL pre-cooled 50mM phosphate buffer (pH 7.8), and centrifuged at 4 °C for 15 min. Supernatants were collected to detect the activities of SOD, POD and CAT according to Zheng et al.[30]. The experiment was repeated independently in triplicate.
Western blot analysis of MbASMT9 protein level
-
Total protein in leaves from every treatment group was extracted as described by Zheng et al.[30]. The MbASMT9 protein level was analyzed by western blot using the MbASMT9 specific anti-body, which was prepared according to Zheng et al.[30]. The signals of chemiluminescence were detected by an ECL detection kit (Thermo Scientific, USA).
Statistical analysis
-
The data were analyzed using ANOVA based on Student’s t-test or Fisher’s least significant difference (LSD) assay on SPSS software (IBM, Armonk, NY, USA) according to Zheng et al.[38].
- This research was supported by the National Key Research & Development Program of China (Project No. 2019YFD1000104 and 2018YFD1000303), the National Natural Science Foundation of China (No. 31772279) and the Construction of Beijing Science and Technology Innovation and Service Capacity in Top Subjects (CEFF-PXM2019_014207_000032).
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng X, Wang L, Zhou J, Shan D, Wang N, et al. 2021. Melatonin enhances the tolerance to iron deficiency stress through scavenging ROS in apple. Fruit Research 1: 9 doi: 10.48130/FruRes-2021-0009

Melatonin enhances the tolerance to iron deficiency stress through scavenging ROS in apple
- Received: 31 July 2021
- Accepted: 09 September 2021
- Published online: 26 September 2021
Abstract: Iron (Fe) is an essential micronutrient for plants. Fe deficiency, resulting from poor availability induced by its insolubility in most soils, severely affects the growth and yield of apple (Malus domestica Borkh.). Except for the application of an Fe nutrition agent, there is a lack of efficient and environmently-friendly ways to relieve plants from Fe deficiency in a modern orchard. Here, we observed that melatonin (MT), as an efficient reactive oxygen species (ROS) scavenger, could alleviate the chlorosis of leaves in apple under Fe deficiency, following increased Fe content, decreased chlorophyll degradation and stable photosynthetic rate. Exogenous melatonin induced the synthesis of endogenous melatonin and improved the activities of antioxidant enzymes, which decreased the oxidative damage induced by Fe deficiency through properly scavenging O2•− and H2O2. In addition, exogenous melatonin promoted the expression of its synthetase genes, but decreased their protein level, inferring post-transcriptional regulation. In brief, melatonin improved the tolerance to Fe deficiency through ROS migration and chloroplast protection in apple. This work uncovered the role of melatonin on Fe deficiency tolerance and also provided a potential way to relieve plants from Fe deficiency stress for apple production.
-
Key words:
- apple /
- melatonin /
- Fe deficiency /
- ROS /
- chlorophyll degradation /
- photosynthetic rate