









-
Apple (Malus domestica Borkh.) fruit are the final products of the fusion between two gametic nuclei: the male pollen grain and the female egg-cell. After the pollen grain is deposited onto the flower's stigma, it germinates and grows a pollen tube down the style to fertilize the egg-cell. The pollen can come from the same plant or from a different one. However, a broadly distributed, pre-zygotic and genetic mechanism called gametophytic self-incompatibility (GSI) prevents the self-fertilization of closely related individuals, promoting out-crossing and thereby maintaining genetic diversity. GSI is found in many angiosperms, including the Solanaceae, Rosaceae, and Plantaginaceae families[1,2].
Apple has a homomorphic GSI mechanism where inhibition of self-fertilization occurs through genetic or biochemical mechanisms that function regardless of flower morphology[3] operating in a reproductive system, which has two different and tightly linked components (S-genes). One is located in the pistil and the other is specific to the pollen[1,4]. The pistil component is an extracellular ribonuclease (S-RNase) that inhibits self-pollen tube growth[5]. The pollen-specific component is controlled by multiple genes called SFBBs (i.e. S-locus F-box brothers) that interact with the S-RNase in an allele-specific way[6]. Both components work in a collaborative manner to control the single, multi-genic and multi-allelic S-locus. Each SFBB interacts with S-RNases, such that non-self-RNases are degraded allowing pollen tube growth[7] (Fig. 1). The selective pressures underlying this collaborative recognition mechanism generate a lower diversity of the S-pollen genes than is found on the S-pistil locus, which shows a higher degree of allelic polymorphism[8]. However, these multi-genic S-haplotypes are inherited as single segregating units keeping their functionality across generations[9,10].
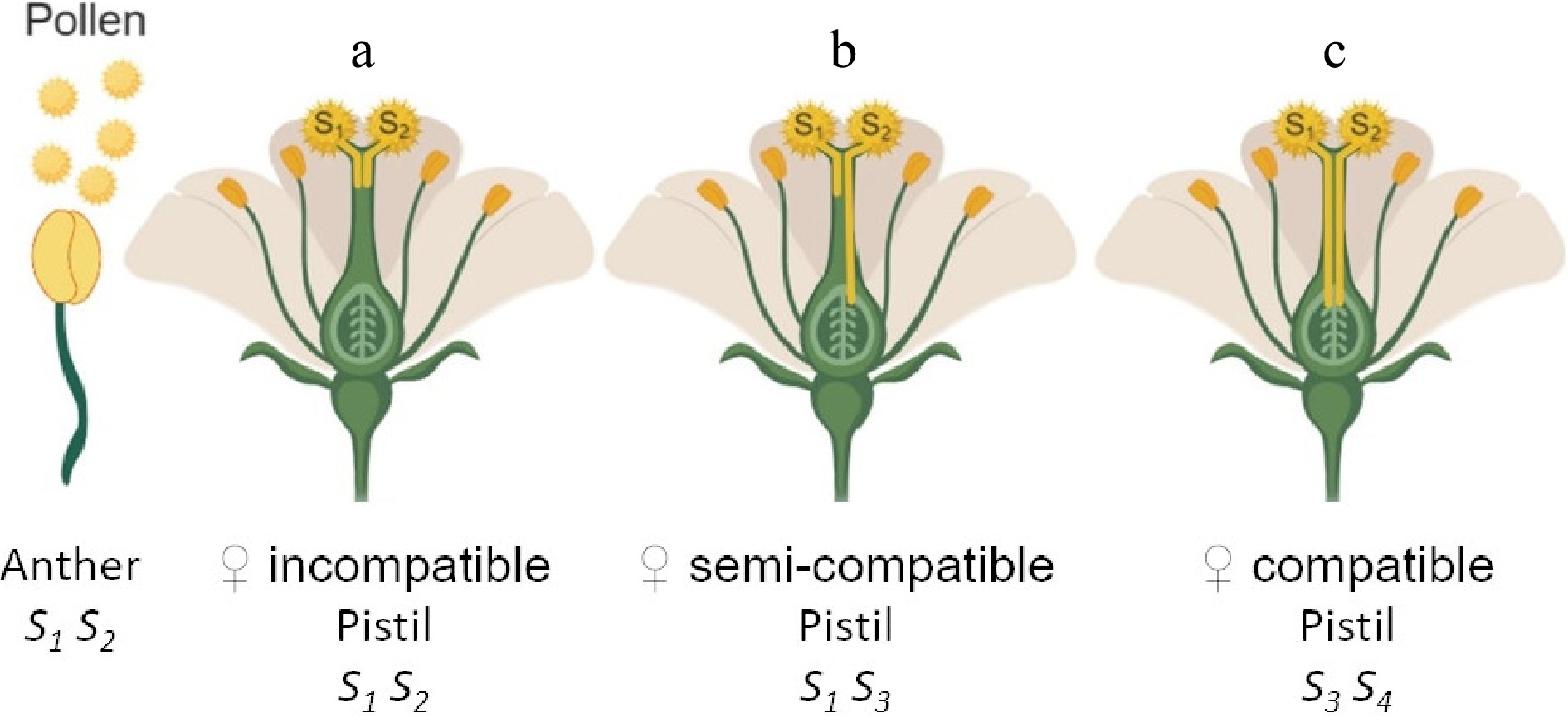
Figure 1. Genetic control of gametophytic self-incompatibility (GSI) in Malus. The S-locus is composed of two tightly linked components, found in the pollen and pistil respectively. In GSI, the pollen self-incompatibility phenotype is controlled gametophytically, i.e., the genotype of the haploid pollen itself (gametophyte) determines its incompatibility type. For example, the pollen composition of a certain pollen donor plant is phenotypically half S1 and half S2. In the female parent, two alleles are co-dominant and both are expressed in the pistil. Pollen inhibition occurs when there is a match between the donor pollen S-haplotype and either of the two haplotypes present in the pistil, producing an incompatible reaction that inhibits the growth of the 'self' pollen tube growth. Three types of reactions can occur during a cross: (a) incompatible; neither of the two gametes will germinate, (b) semi-compatible; half the donor pollen will be inhibited and the other half will germinate and grow normally, (c) compatible; all pollen will germinate and grow normally.
Apple breeding relies on compatible and productive cross-pollination. A breeder needs information about the self-incompatibility genotypes (S-genotypes) of both parents to execute successful crosses and facilitate the selection of individuals carrying a combination of desirable traits. Traditionally, incompatibility was determined using time-consuming cross-pollination experiments, where successful fruit set was measured over many combinations of parents. Recently, less time consuming and more cost-effective molecular markers have been implemented to replace such field experiments, using either allele-specific markers amplifying a single S-RNase allele[11−18] or markers based on restriction enzyme digestion of polymerase chain reaction (PCR) products (cleaved amplified polymorphic sequences CAPS or PCR-RFLP). Simple Sequence Repeat (SSR) markers have also been used for screening S-alleles[14,15,19]. CAPS and SSR markers have helped determine most of the S-genotypes for common commercial apple varieties. However, most of the existing assays involve the use of restriction enzymes after PCR reactions and the visualization of the products on agarose gels, making them very laborious and time-consuming when handling large numbers of samples. This can be problematic evidenced when new seedlings/selections, that are potential parents in a breeding programme, need to be checked for compatibility each season.
The objective of this study was to develop and validate a high-throughput and practical method to identify 13 different apple S-RNase alleles (S1, S2, S3, S5, S7, S8, S9, S10, S20, S23, S24, S25 and S28). Our method is composed of seven allele-specific High-Resolution Melting (HRM) and four multi-allelic SSR markers, which do not require post-PCR restriction enzyme digestions or agarose gels for allele-scoring analysis. We used as controls 70 out of a total of 86 commercial apple cultivars for which S-genotypes had already been reported, to validate the accuracy of our markers for identifying the correct S-alleles. We then demonstrate the usefulness of these assays by genotyping 183 genotypes representing some of the most valuable parents within the PFR breeding program.
-
The four developed SSR markers Myb110a1_PFR, Myb110a2_PFR, Myb110b_PFR and GSI_SSR_PFR amplify polymorphic PCR products associated with 13 different S-alleles (S1, S2, S3, S5, S7, S8, S9, S10, S20, S23, S24, S25 and S28) (Table 1). All four primer pairs amplify PCR products linked to the S1, S3, S5, S7, S24 and S25 S-alleles. Both Myb110b_PFR and GSI_SSR_PFR can be used to distinguish the S10 S-allele. Myb110b_PFR exclusively identifies S2 and S9, whilst GSI_SSR_PFR amplifies a PCR product linked to the S28 S-allele. All four primer pairs, except Myb110b_PFR, amplify PCR products linked to S20 and all four but Myb110a1_PFR can identify the S23 S-allele.
Table 1. Primer sequences of quantitative real-time PCR and SSR-based markers.
Marker name Type Primer sequences '5 - 3' Physical location Genbank locus S-Rnase alleles and product sizes (bp) S1_apple_PFR HRM Forward ACAGGCCACTGGTGGA not found in reference genome MG598487.1:1981−1996 S1, S20, S24 (38) Reverse ATTGCGTATGGCATTTTCAAT Chr17:30844510−30844530 MG598487.1:1998−2018 S2_apple_PFR HRM Forward TTGAACAAATATTATTCAATGGGGA Chr17:31240988−31240964 MG598488.1:860−884 S2 (54) Reverse CATCGTAACTATATATACCATCCGCGTA Chr17:31240964−31240943 MG598488.1:886−913 S5_apple_PFR HRM Forward AATTTATAAAACACGTGATCA not found in reference genome MG598491.1:326−346 S5 (43) Reverse GCTCCTATTGATCGATCAT not found in reference genome MG598491.1:350−370 S8_apple_PFR HRM Forward TTCGATTATTTTCAATTTACGCTT Chr17:31240889−31240871 MG598494.1 :1159−1182 S8 (162) Reverse ATTTAAGGTTGTTTCTTTGCAATAC not found in reference genome MG598494.1 :1296−1320 S9_apple_PFR HRM Forward GCTCAGGAAATGACCCAATATAC not found in reference genome MG598495.1:1284−1306 S9 (61) Reverse AATATTACCTTAGTAGAATTCATGGTTGT not found in reference genome MG598495.1:1315−1344 S23_apple_PFR HRM Forward TTTATGGCCTTCAAACTGGAA not found in reference genome MG598501.1:1065−1085 S23 (42) Reverse CAGAAGATTGGGTCGGGT not found in reference genome MG598501.1:1089−1106 S28_apple_PFR HRM Forward TGCCTCGCTCTTGAACAAA not found in reference genome MG598505.1:782−800 S28 (47) Reverse CCCCGTAATTCCCATTGAATAATA not found in reference genome MG598505.1:805−828 Myb110a1_PFR SSR Forward TCTCCCTCATCCCAGAACA Chr17:32151473−32151491 S1 (166), S3 (184), S5 (180), S7 (170), S20 (158), S24 (176), S25 (188) Reverse CGAGCCAAACAAAATTGGA Chr17:32151642−32151624 Myb110a2_PFR SSR Forward CTCTCCCTCATCCCAGAACA Chr17:32151472−32151491 S1 (325), S3 (343), S5 (339), S7 (314), S20 (317), S23 (309), S24 (320), S25 (347) Reverse TCCTACTCGGCTCGACAATC Chr17:32151800−32151781 Myb110b_PFR SSR Forward CTTCGGGCTTATTTGGGTTT Chr17:32187809−32187790 S1 (202), S2 (233), S3 (214), S5 (209), S7 (191), S9 (247), S10 (216), S23 (239), S24 (217), S25 (238) Reverse TTTGCCCCTTCAAAGATCAG Chr17:32187616−32187635 GSI_SSR_PFR SSR Forward GCCCCTTACATTCCTTTTCTTT Chr17:31704109−31704130 S1 (314), S3 (338), S5 (335), S7 (324), S10 (328), S20 (317), S23 (322), S24 (217), S25 (329), S28 (352) Reverse CAATCTTGAGTTGTCGTTGGAG Chr17:31704430−31704409 Additionally, we developed seven quantitative-PCR allele-specific markers. Six of these markers (S1, S2, S5, S9, S23 and S28_apple_PFR) amplify specific single nucleotide polymorphisms (SNPs) identified by a pairwise alignment of the coding sequences of 25 S-RNase alleles, previously published by De Franceschi et al.[20]. The seventh marker for S8 was adapted to work with our HRM methodology, by modifying the forward primer from the previously published primers pairs (Larsen et al.[19]) (Fig. 2). When used on their own, without the SSR markers, these S-allele-specific qPCR markers can identify eight S-alleles in total. The S1_apple PFR marker can resolve the S20, S24 and S1 S-alleles as separate melting curves.

Figure 2. High resolution melting (HRM) curve profiles of seven S-allele-specific markers. Amplification curves of real-time PCR marker assays (left panels), HRM difference plots, where the derivative fluorescence signal (dF/dT) is plotted as a function of temperature (right panels). Each colour represents a specific S-genotype as shown by the legends. Light grey represents samples that were not amplified in the real-time PCR.
Validation of the S-RNase allele genotyping method on a large set of apple commercial cultivars
-
The S-allele genotyping of 86 apple accessions (81 different cultivars, counting a red mutant sport of 'Fuji' and four 'Gala' mutant sports) – including 70 traditional varieties with previously reported S-alleles, five varieties with unknown S-alleles and ten newer cultivars arose from the PFR breeding programme – was undertaken using the new set of molecular markers. The S-genotypes of 59 of the 70 traditional apple varieties were in complete agreement with previous reports and the S-genotypes of their respective parents. For ten accessions, our results for one of the two S-alleles disagreed with those published previously (Supplemental Table S1): 'Abbondanza' had been reported as (S3 S5)[20], while (S3 S7) S-alleles were detected in the present study. 'Antonovka' was reported (S8 S32)[20], while we detected S8 and other allele sizes that could equally be linked to S3, S7 or S20. 'Priscilla' was reported as (S3 S9)[21] or (S9 S20)[22] or (S7 S10)[20], while we detected (S7 S28) alleles. 'Ingrid Marie' was reported as (S5 S43)[23], while our most likely observed S-genotype was (S3 S5), 'James Grieve' was reported as (S5 S8)[23], while we detected (S5 S20). 'Ben Davis' was reported as (S5 S23) while we detected (S7 S23); 'Jonathan' was reported previously as (S7 S9)[24−28]; however, we detected (S9 S23). 'Early Cortland' has been reported as (S5 S28)[29,30]; however, we report it here as (S1 S28)[29]. 'Yellow Transparent' has been reported as (S1 S5)[31,32], while we identified (S1 S7 S9 S24). Finally, 'McIntosh' was reported as (S10 S25)[11,23], while we found (S2 S25).
The new markers also resolved both S-alleles for three out of four traditional varieties with previously undetermined S-genotypes: 'Red Dougherty' (S1 S7), 'Pinkie' (S2 S3) and 'Merton Russet' (S5 S24). We identified only the S25 allele for 'Paulared'. The ten cultivars that have recently arisen from the PFR breeding programme were successfully typed for their S-alleles: 'Scired' (S2 S9), 'PremA093' (S2 S9), 'Scifresh' (S2 S24), 'PremA153' (S2 S24), 'PremA34' (S2 S3), 'Scilate' (S5 S9), 'PremA96' (S5 S9), 'PremA17' (S5 S24), 'PremA280' (S5 S24) and 'PremA129' (S9 S24). For the Canadian variety 'Sunrise' we identified S-alleles (S3 S24).
For a remaining set of five cultivars, just one S-allele could be determined: 'Hetlina' and 'Geheimrat Dr Oldenburg' were reported as (S1 S16b)[12] and (S3 S28)[20], respectively. The respective S1 and S3 S-alleles were detected in our study; however, we did not identify allele sizes that could be linked to either S16b or S28 using the new markers. For 'Benoni', reported as (S5 S11)[33], we detected the S5 allele, as well as other marker alleles not linked to the expected S11. 'Regent' was reported as (S3 S10)[34]; we identified the S10 allele, but no S3-linked alleles were observed. Instead, an allele linked to S25 was detected. Finally, 'Panenské České' was reported as (S7 S10)[20], but only the S7 allele was detected using three SSR markers. Marker alleles linked to S10 were not found, rather the Myb110b marker detected alleles linked to S3 and S24.
Diversity of S-RNase alleles in the PFR apple cultivar breeding programme
-
When the new markers were screened over 183 genotypes, including some of the most valuable parents within the PFR apple cultivar breeding programme, we were able to determine their S-genotypes. These genotypes are the seedlings of 76 biparental families (Supplemental Table S2). For 32 of these families (from a total of 132 selections), the S-genotypes of their parental pedigrees could be verified. The frequency of S-RNases alleles found among this pool of genotypes is shown in Fig. 3. Among the 183 genotypes screened, S2 was the most common S-allele, present in 21.3% of the samples, followed by S3 (19.9%), S24 (18.6%), S5 (17.5%), S23 (13.7%), S9 (5.7%) and S7 (0.8%). Rare S-alleles were S28 (0.5%), S25, S1 and S20 (0.3% each). Only 2.2% of the S-alleles could not be assigned. The most prevalent genotypes were: (S3 S24), (S2 S24), (S2 S5), (S2 S23), (S3 S23), (S3 S5) and (S5 S23) observed at frequencies of: 7.1, 7.1, 6.3, 5.2, 4.4, 4.1 and 3%, respectively. Other less prevalent genotypes were: (S5 S24), (S5 S9), (S3 S9), (S2 S3), (S9 S24), (S2 S9) and (S23 S24), which were observed at frequencies of 2.2, 1.6, 1.4, 1.4, 1.4, 1.1 and 0.8%, respectively. The following rare genotypes were each found at a frequency of 0.3%: (S1 S3), (S2 S7), (S5 S7), (S7 S9), (S20 S28) and (S3 S28). For 1.1% of the selections, just one allele was identified (S3 ?) in 0.8% and (S23 ?) in 0.3%).

Figure 3. Frequency of S-alleles and S-genotypes of the 183 apple advanced selections of the PFR’s breeding programme. Inner plot shows the percentage frequency distribution of S-alleles from the total 366 alleles observed among the 183 genotypes tested. Outer plot represents the absolute frequency of each S-genotype. All outer slices not showing a percentage value in the figure represent 0.3% respectively.
-
A high-throughput method to identify the S-genotypes of apples was developed and validated in this study. This will help to inform the selection of compatible parental combinations when designing a crossing programme. We present a new high-throughput marker set based on four multi-allelic SSR and seven allele-specific qPCR markers. The four SSR markers can identify 13 S-RNase-alleles (S1, S2, S3, S5, S7, S8, S9, S10, S20, S23, S24, S25 and S28) and the seven HRM markers allow the identification of eight S-RNase-alleles (S1, S2, S5, S8, S9, S20, S23 and S28). The identification of the 13 S-alleles can be achieved economically and efficiently by employing three PCR reactions, using two (Myb110b_PFR and GSI_SSR_PFR) of the four SSR markers and the S8_apple_PFR marker (with the addition of an M13-tail on the 3'-end). These three markers can be multiplexed by using different fluorescent labels that can be simultaneously separated and scored on a capillary electrophoresis instrument.
Alternatively, for laboratories with access to a real-time qPCR system, as well as a capillary electrophoresis instrument, the seven qPCR allele-specific markers and one of the SSR markers (either Myb110b_PFR or GSI_SSR_PFR) will be sufficient to identify and resolve the whole set of 13 S-alleles.
The usefulness of the new markers was validated in over 59 of 70 well-established apple cultivars with known S-genotypes. Ten of the discrepancies with previous assays are supported by our results from parental pedigree allele analysis, acknowledging that some might be mistakes with labelling, or incorrect germplasm harvest: 'Priscilla' has been reported as (S3 S9)[21], (S9 S20)[22] or (S7 S10)[20], while we detected (S7 S28), with S28 as probably coming from 'Starking Delicious', which is reported as (S9 S28)[29]. 'Ingrid Marie' was reported as (S5 S43)[23], while our most likely observed S-genotype was (S3 S5), where S5 is derived from 'Cox's Orange Pippin' (S5 S9). However, neither S43 nor S3 has been reported in 'Cox's Pomona' S-genotype (S1 S34)[19]. 'Early Cortland' has been reported as (S5 S28)[29,30], which is consistent with its parentage: 'Cortland' (S5 S25)[29] and 'Lodi' (S1 S28)[30]; however, we reported it here as (S1 S28)[29]. 'Abbondanza' was reported as (S3 S5)[20], while (S3 S7) S-alleles were detected here. 'Antonovka' was reported (S8 S32)[20], while we detected allele S8; however, we observed different allele sizes for our SSR markers that we could link to either S3, S7 or S20. There are different 'Antonovka' accessions[35], so it is probable that they have different S-genotypes. 'James Grieve' was reported as (S5 S8)[23], while we detected (S5 S20), although the Myb110b_PFR marker showed an additional 202 bp allele, which is linked to S1 S-RNase, but did not exhibit any other allele sizes linked to S1 in any of the other SSR or qPCR markers. Then, 'Ben Davis' was reported as (S5 S23), while we detected (S7 S23), with the same allele sizes found and expected for 'Lady Williams' (S7 S23). 'Jonathan' was reported previously as (S7 S9)[24−28]; however, we characterised it as (S9 S23), but note that we do have molecular and phenotypic indicators suggesting this could be an incorrectly identified accession in the PFR germplasm (Vincent Bus, pers. comm.). Finally, 'Yellow Transparent' is reported as (S1 S5)[31,32], while we identified (S1 S7 S9 S24), which is consistent with this cultivar being a tetraploid sport mutant[36].
We demonstrated the usefulness of the markers by determining the S-genotypes of ten newer cultivars arising from the PFR breeding programme ('Scired', 'PremA093', 'Scifresh', 'PremA153', 'PremA34', 'Scilate', 'PremA96', 'PremA17', 'PremA280' and 'PremA129'). The S-genotype information for such new cultivars is valuable information for growers, enabling them to plant compatible pollenisers in commercial orchards. Despite the high diversity of S-RNase alleles that have been characterized in Malus (at least 35 different S-alleles were found among cultivars in Matsumoto's database[29]), the common worldwide practise of using a relatively small pool of cultivars that combine premium fruit quality as well as resistance to pests and environmental stresses in breeding programmes leads to new cultivars with restricted allelic combinations. Among the 183 PFR apple genotypes tested here, a pool of only 11 S-alleles was found (S1, S2, S3, S5, S7, S9, S20, S23, S24, S25, and S28). This is not surprising given that all the S-alleles of the main founders of PFR's cultivar breeding programme [namely 'Splendour' (S2 S9), 'Cox's Orange Pippin' (S5 S9), 'Red Delicious' (S9 S28), 'Golden Delicious' (S2 S3), 'Red Dougherty' (S1 S7), 'Worcester Pearmain' (S2 S24), 'Jonathan' (S23 S9), 'Fuji' (S1 S9), 'Braeburn' (S9 S14), 'Granny Smith' (S3 S23), 'James Grieve' (S5 S20), 'Wagener' (S3?), 'Cripp's Pink' (S2 S23) and 'Akane' (S7 S24)[34]] have ten of these 11 S-alleles. However, it is possible that the wider parental pool also has undetected S-alleles beyond these 11, as some breeding parents, not represented in the 183 genotypes tested here, are also derived from minor founders.
The most frequently observed allele was S2 (21.3%), which is one of the two alleles carried by 'Royal Gala' (S2 S5), a parent or grandparent in pedigrees of most of the PFR genotypes. For instance, 'Scired', 'Sciros', 'Scilate', and 'Sciray' were used as the pollen parents for many crosses and they all are progeny of a cross between 'Gala' (S2 S5) and 'Splendour' (S2 S9). 'Gala's' parentage is 'Golden Delicious' (S2 S3) and 'Kidd's Orange Red' (S5 S9), so 'Golden Delicious' is the source of this allele in 'Gala' or 'Royal Gala' crosses.
S3 was the second most abundant allele (19.9%), being present in crosses of genotypes with 'Pinkie' in their parentage. 'Pinkie' likely inherited the allele from 'Granny Smith' (S3 S23), although we do not know the S-alleles carried by its other parent A679-2. Also, crosses produced using 'Fiesta' (S3 S5), have inherited the S3 allele from 'Idared' (S3 S7), which has 'Wagener' (S3 ?) as a parent.
Allele S24 was observed in 18.6% of the genotypes, those arising from crosses with 'Braeburn' (S9 S24) as one of the parents: 'Scifresh' is a progeny of the cross 'Braeburn' 'Royal Gala'; while 'PremA153' is derived from a 'Gala' × 'Braeburn' cross. 'PremA129' has 'Braeburn' as a grandparent, being a progeny of 'PremA280' × 'Scired', with 'PremA280' having 'Gala' and 'Braeburn' as parents. 'PremA17' (S5 S24) also has this allele, presumably from 'Braeburn': this cultivar was derived from a cross between genotypes A045R13T007 × A020R02T167, which unfortunately are no longer available in the orchard. Another source of this allele is 'Akane' (S7 S24)[34], which inherited it from 'Worcester Pearmain' (S2 S24).
The origin of S5 (17.5%) in our breeding programme is 'Cox's Orange Pippin' (S5 S9). Crosses that involve 'Fiesta' or 'James Grieve' have 'Cox's Orange Pippin' as a grandparent. It is also a great-grandparent in crosses that have 'Gala' as a parent and a great-great-grandparent in crosses that include cultivars such as 'Sweetie', 'PremA17' and 'PremA96' as parents.
The S23 allele is present (13.7%) in seedlings derived from our A068 family; however, we still need to confirm the source of the allele as we do not know the S-genotypes of grand-parents. S9 is found in 5.7% of seedlings derived from crosses where one of the parents was 'Scired', or alternatively with 'Splendour' as one of the grandparents or great-grandparents. Other important sources of the S9 allele are 'Cox Orange Pippin' (S5 S9) and 'Braeburn' (S9 S24).
At the other end of the scale, the S7 allele was only found in 4.8% progeny of crosses with 'Red Free' (S3 S7) as a parent or grandparent and in some crosses using 'Akane' (S7 S24)[34], which has 'Jonathan' (S7 S9) as the likely parental source of this allele. The alleles S25 and S28 occur rarely in the PFR breeding programme. They probably come from 'McIntosh' (S10 S25) and 'Delicious' (S9 S28), respectively; however, the pedigree of the few selections with these alleles is not complete: further information is needed to confirm this hypothesis.
According to Sheick et al.[18], 11 S-alleles (S1, S2, S3, S5, S9, S10, S20, S23, S24, S25, and S28) are represented among the U.S. industry's most produced apples. These are predominantly coming from 'Red Delicious' (S9 S28), 'Gala' (S2 S5), 'Granny Smith' (S3 S23), 'Fuji' (S1 S9), 'Golden Delicious' (S2 S3), 'Honeycrisp' (S2 S24), 'McIntosh' (S10 S25), 'Rome' (S20 S24), 'Cripps Pink' (S2 S23), and 'Empire' (S10 S28). These same 11 S-alleles but S10 are represented in the New Zealand PFR breeding programme. Instead of S10, we have S7 included in our pool of S-alleles, which is represented in crosses having 'Jonathan' (S7 S9) and 'Red Free' (S3 S7) in their pedigree. Another recent study by Lays Brancher et al.[37] identified 11 S-alleles (S1, S2, S3, S5, S7, S9, S10, S19, S20, S23, and S24) among 42 apple genotypes, including cultivars, advanced selections and accessions of the Apple Germplasm Bank of Epagri (Caçador, Santa Catarina, Brazil). The S3 and S5 alleles were most frequent (30.2% and 18.6%, respectively). The higher frequency of these alleles can be explained as 26 of the 42 accessions tested were direct or indirect descendants from the cultivars Imperatriz (S3 S5), Golden Delicious (S2 S3) and/or Gala (S2 S5), which have served as the basis for the crosses of the Epagri Apple Breeding Program.
A Danish study by Larsen et al.[19] found 25 S-alleles (S1, S2, S3, S4, S5, S6, S7, S8, S9, S10, S11, S16b, S16c, S20, S21, S23, S24, S25, S26, S28, S31, S33, S34, S36 and S40) in 432 Malus accessions including a selection of M. domestica cultivars of mainly Danish origin (402 accessions), as well as a selection of other Malus species (30 accessions). Among the 402 Danish accessions the allele S3 (28 %) was the most common followed by S1 and S7 (both 27 %). Previous studies[16,38] using cultivars from Northern Europe and the Carpathian basin found similar results where S3 and S7 were the two most common S-alleles.
Although selections used as parents in breeding programmes around the world are different due to consumer preferences, climate conditions, resistances to pest and diseases, etc., there is a common set of cultivars among worldwide breeding programmes[39]. These are 'Golden Delicious' (S2 S3), 'Braeburn' (S9 S24), 'Fuji' (S1 S9), 'Gala' (S2 S5), 'Granny Smith' (S3 S23), 'Idared' (S3 S7), 'Jonathan' (S7 S9) and 'Red Delicious' (S9 S28). The S-alleles carried by these cultivars are also the most common among the total 183 advanced selections tested within PFR's breeding programme. The S3 allele is the most common S-allele worldwide as seen in the previous studies mentioned here and among other older studies including European, American and Japanese cultivars[12,23,40]. This low diversity of S-RNase alleles highlights the need of introducing breeding cultivars with some of the less common S-alleles into breeding programmes to increase mate compatibility among parental selections.
-
Leaves from 86 apple cultivars were collected at PFR, Havelock North, New Zealand, and Washington State University, Pullman, WA, USA. Total genomic DNA was extracted using the cetyltrimethyl ammonium bromide (CTAB) method[41]. This DNA was used as a set for evaluating the new markers (Supplemental Table S1). Additionally, leaves from 183 apple genotypes, from 76 biparental families, were collected from trees in PFR's elite parental apple collections, to identify their S-genotypes.
SSR-based S-allele markers
-
Three new primer pairs were designed around two single sequence repeats (SSR) linked to the Myb110a and Myb110b genes[42] and named Myb110a1_PFR, Myb110a2_PFR and Myb110b_PFR, which are closely linked to the S-locus on apple chromosome 17. A fourth primer pair was designed for a SSR located within the S-locus (GSI_SSR_PFR) (Table 1). Design of the primer pairs was based on the GDDH13v1.1[43] apple genome as a reference and employed using the Krait software[44]. The M13 sequence TGTAAAACGACGGCCAGT was added to the 5′ end of the forward primer to enable the use of Schuelke's[45] approach to fluorescent labelling. PCR was performed in a 15 µL reaction mixture containing 1.5 mM MgCl2, 200 uM dNTPs, 13 nM of forward primer, 200 nM of reverse primer, 8.33 µL DNA-free water and 1× PCR Buffer (-MgCl2) and 0.5 U of Platinum™ Taq DNA polymerase (Thermo Fisher Scientific, 10966034). The conditions of the touchdown PCR included an initial denaturing at 94 °C for 2 min, then five cycles (94 °C for 55 s, 65 °C for 55 s (decreased by 1 °C each cycle), 72 °C for 1 min and 39 s), then 35 cycles (94 °C for 55 s, 55 °C for 55 s and 72 °C for 1 min and 39 s) and a final extension at 72 °C for 10 min. The final amplicons were subjected to capillary electrophoresis using an ABI 3500 DNA sequence analyser (Applied Biosystems, Foster City, USA) and sized using GenScan™ 500 LIZ Size Standard (Applied Biosystems). SSR allele profiles were analysed using GeneMarker™[46] version 2.20 (SoftGenetics LLC®, State College, PA, USA, www.softgenetics.com).
Quantitative real-time PCR-based markers
-
SNPs were identified by performing a multiple sequencing progressive pairwise alignment[47] of the coding sequences of 25 S-RNase alleles previously published by De Franceschi et al.[20] in Geneious version 10.0.9 (https://www.geneious.com), with the following parameters: global alignment with free end gaps algorithm, 70% similarity cost matrix, gap open penalty of 11.9, gap extension penalty of 2 and 2 refinement iterations.
Seven S-RNase allele specific primer pairs named Sx_apple_PFR (x being: 1, 2, 5, 8, 9, 23 or 28 alleles) were designed to amplify a single product of 250 bp or less. These primer pairs can be used on a conventional PCR machine or by employing the High Resolution Melting (HRM) methodology[48] on a quantitative PCR instrument. The primer pairs for S8_apple_PFR marker were modified from the ones previously published by Larsen et al.[19].
Conventional PCR reactions were carried out in 15 µL volume containing 1× PCR buffer mix (Invitrogen), 200 μM of each dNTP, 1.5 mM MgCl2, 3 μM each primer, 0.1 U Platinum™ DNA polymerase (Thermo Fisher Scientific, 10966034) and 20 ng template DNA. Amplifications were carried out on a MasterCycler ProS thermocycler (Eppendorf). The conditions of the touchdown PCR included an initial denaturing at 95 °C for 5 min, then ten cycles (94 °C for 30 s, 60 °C for 30 s (decreasing 1 degree in each cycle) and 72 °C for 45 s), then forty cycles (94 °C for 30 s, 50 °C (for S5_apple_PFR primer pair) or 55 °C (for S1, 2, 9, 23, 28_apple_PFR primer pairs) for 30 s and 72 °C for 45 s) and a final extension at 72 °C for 5 min. PCR products were then visualized on a 2% agarose gel stained with RedSafe™ 20000x (ChemBio, UK) after 1 h of electrophoresis at 100 V.
Quantitative PCR reactions were performed in a total volume of 10 µL containing 20 ng of template DNA, 2.5 mM MgCl2, 200 nM forward and reverse primers and 1× HRM master mix (Roche Applied Science). PCR and HRM were performed on a LightCycler® 480 (Roche Diagnostics). The PCR parameters were an initial denaturation step of 95 °C for 5 min, followed by 40 cycles of 95 °C for 10 s, 55 °C for 20 s, and 72 °C for 20 s. Following amplification, the samples were heated to 95 °C for 1 min and then cooled to 40 °C for 1 min. Melting curves were generated with continuous fluorescence acquisition during a final ramp from 65 °C to 95 °C at 4.8 °C/s, followed by a final cooling step of 40 °C for 30 s. The resultant fluorescence data were processed using the LightCycler® 480 software (version 1.5; Roche Applied Science). Primer sequences, fragment sizes and their respective associated S-RNase alleles are shown in Table 1.
S-genotyping Malus cultivars and breeding seedlings
-
The four SSR-based markers were initially screened using a DNA set from 70 out of a total of 86 apple cultivars with known S-genotypes, based on previously reported CAPS or PCR-RFLP detection methods (as referenced in Supplemental Table S1). Following this, the seven HRM assays were screened over the same cultivars to validate the allele-specificity of each primer pair (Table 1).
Following the screening of the first DNA set, all 11 markers for S-genotype were further validated using 183 apple genotypes from the PFR breeding programmes. The S-alleles were confirmed by verifying the S-genotype composition within each family and by examining their pedigree composition up to the grandparent level. For the PFR breeding populations, a summary of the S-genotype composition of the tested seedlings within families was made.
Data availability
-
Raw data and R script for statistical analysis are available at link https://github.com/hrpelg/Rnotebook_Self-incompatibility
-
We demonstrated the efficiency of a set of markers for the S-locus in a Malus domestica germplasm set with known S-genotypes and we determined the S-genotypes of uncharacterized cultivars, with an emphasis on new commercial releases. We showed the S-genotyping efficacy of this method on a large sample of advanced apple genotypes from the PFR breeding programme, where S-genotypes were concordant with their parental pedigree.
This robust, reproducible, simple and cost-efficient S-RNase-genotyping method is an alternative to the present molecular approaches. The existing molecular methods employ single allele specific markers per every single S-allele or use marker based restriction enzyme digestions of PCR products to distinguish among few S-alleles needing to be visualized on agarose gels. The flexibility of our method permits to know 13 different S-alleles by employing just three different PCR reactions in a laboratory provided with a capillary electrophoresis instrument. These three PCR reactions can be multiplexed in a single electrophoresis run by using three different fluorescent colours. Alternatively, if a qPCR instrument is also available, this can be done using seven different HRM-markers and a single SSR marker. The use of a qPCR instrument allows the analysis of 384 samples per run or the multiplexing of four markers per PCR for every 96 samples.
This method is provided to scientists, breeders and growers to select compatible pollenisers and to develop new cultivars. The benefits of knowing the S-alleles that each parental selection carries are: pollination success between compatible parental pollen and pistil, higher yields of orchards planted with compatible varieties and possible parentage identificationof unknown seedlings' descent due to undesired open pollination.
- We would like to gratefully thank Stijn Vanderzande for providing the US plant material and Susan E. Gardiner for helpful advice and review of the manuscript. This work was funded by Prevar Limited (https://prevar.co.nz).
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 List of 86 commercial apple cultivars used as a validation set in this study, allele sizes (bp) per marker, observed and expected (by previous published assays) S-genotypes and parental pedigrees. a S-genotypes in disagreement with previous reports, * S-genotypes not reported prior this study and (?) Unknown S-allele or unconfirmed pedigree. Each different S-allele and its associated allele sizes are highlighted in a different colour.
- Supplemental Table S2 List of 183 apple genotypes from the PFR cultivar breeding programme grouped by family, allele sizes (bp) per marker and observed S-genotypes.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
López-Girona E, Bowatte DR, Smart MEM, Alvares S, Brancher TL, et al. 2021. A high-throughput S-RNase genotyping method for apple. Fruit Research 1: 10 doi: 10.48130/FruRes-2021-0010

A high-throughput S-RNase genotyping method for apple
- Received: 06 August 2021
- Accepted: 08 October 2021
- Published online: 25 October 2021
Abstract: Knowledge of the genotypes for the self-incompatibility locus (S-locus) in apple varieties and in genotypes being used as parents is critical for breeding and commercial production. We present a high-throughput set of molecular markers for the identification of 13 common S-RNase alleles (S1, S2, S3, S5, S7, S8, S9, S10, S20, S23, S24, S25 and S28). This set is composed of seven allele-specific quantitative PCR-based High-Resolution Melting assays and four multi-allelic SSR markers. Validation of these markers was performed using 86 apple accessions, including cultivars with known S-genotypes and recent commercial varieties arising from the Plant & Food Research (PFR) cultivar breeding programme. We also characterized the S-genotypes of 183 genotypes representing some of the most valuable parents within PFR’s cultivar breeding programme. The results of this work demonstrate the practical usefulness of this marker set to provide accurate cross-compatibility information to optimise choice of pollenisers in commercial apple orchard design, and to identify compatible parents and guide parental selection when executing apple breeding programmes, to optimise fruit crop yield and quality.
-
Key words:
- Malus /
- Self-incompatibility /
- S-allele /
- S-RNase /
- Allele-specific PCR /
- S-genotyping