









-
Apple is a widely consumed fruit worldwide. Roots are undoubtedly fundamental organs of any plant. They can absorb nutrients, minerals, and water, as well as anchor the plant in the soil, which is ultimately essential for plant growth and development. Roots obtained from the embryo are called embryonic roots, and roots that arise from non-root organs or existing roots are called post-embryonic roots[1]. The post-embryonic roots emerging from existing roots are characterized as LRs, whereas roots arising from non-root organs are titled adventitious roots (ARs)[1]. To manage the needs of developing shoots, roots must constantly expand their surface area to fulfill the requirements of aboveground tree portions, replace older malfunctioning roots, and discover new nutrient and water sources. Moreover, apple root morphology is strictly associated with their functions. Generally, roots with a diameter < 2 mm are considered as fine roots, which actively perform absorption and are equipped to control the hormone synthesis process and allocation [2]. Therefore, fine roots are necessary to maintain plant physiology[3]. B9 is a superior apple rootstock because of its strong tree architecture; good fruiting ability; resistance to pests, diseases, and other environmental challenges; and high production[4]. However, B9 is sensitive to soil environmental changes, which restricts its use. Furthermore, studies on rooting molecular biology preliminary focused on Arabidopsis[5−8], while some reports have been published on apple rooting[2]. Therefore, it is essential to identify the problems related to LR formation to better understand and address the difficulties associated with LR formation in apple rootstocks.
The development of apple LRs comprises three stages, including the initiation of LR, the development of LR primordia, and the emergence of LR[9]. According to Dubrovsky et al.[10], LRs are initiated from pericycle cells inside a xylem pole, which act as founder cells. Then, unique signals are obligatory to trigger these founder cells to undertake cell division to produce LR primordia, leading to LR formation[9].
Nitrate serves as an essential nitrogen source and is involved as a critical signaling molecule for modulating flowering time, regulating root architecture and leaf growth, and prompting auxin-related gene expression[11−13]. Moreover, the formation of LRs and nitrate provision are firmly linked. The soil nitrate level is usually low because of its high solubility, leaching ability, and rapid acquisition by bacteria and fungi[14]. There are two nitrate transportation mechanisms in higher plants, the high-affinity transport system (HATS) and the low-affinity transport system (LATS), based on the absorption, distribution, and storage of nitrate[15]. The nitrate supply is quickly and intensely sensed by the plant cells. After that, the nitrate signaling pathway alters the expression of different gene sets to regulate cell and organ metabolism. NITRATE TRANSPORTER gene (NRT1.1), a dual affinity nitrate transporter, functions in the auxin-mediated nitrate signaling pathway along with modulating root architecture[16]. NRT2.1, a HATS type gene, played an important role in regulating root development at low nitrate concentrations and its expression was inhibited by high nitrate[17,18]. After transport to the leaves, nitrate is reduced to ammonium by nitrate reductase (NR, NIA1, and NIA2). ANR1 is a nitrate-inducible gene that reveals a high involvement in nitrate-stimulated root elongation[19].
Endogenous hormones are primarily tied to the LR formation via affecting the signaling system inside the plant and controlling cell fate and specialization. Auxin is a vital hormone for LR formation, modulating numerous physiological processes within the plant body. In Arabidopsis, LR initiation was stimulated by auxin[20], and its sole application seems to be adequate for LR initiation[21,22]. In apples, the general mechanism through which auxin promotes LR formation and development has been published[23]; however, little is known about how auxin and other hormones interact to regulate LRs. Furthermore, abscisic acid (ABA) and cytokinin (CTK) are known to be negative regulators of LR formation[24,25].
With regards to the auxin signaling pathways, auxin resistant/auxin (AUX/IAA) protein degradations stimulated auxin response factors (ARFs), which activate the expression of auxin-responsive genes[26,27]. In addition, IAA14 interacted with the ARF7 and ARF19[28], which shows that LR initiation is stimulated by auxin in a cell-independent manner by the signaling module of SLR/IAA14-ARF7-ARF19 in Arabidopsis. However, the underlying molecular mechanism of auxin-induced LRs and signal transduction in apple has yet to be studied. In Arabidopsis, the expression of WUSCHEL RELATED HOMEOBOX genes (WOX5 and WOX11), LATERAL ORGAN BOUNDARIES DOMAIN genes (LBD16 and LBD29), and CYCLIN D3;2 (CYCD3;2) were induced by auxin[29−31]; however, the specific role of these genes in apple, remains to be clarified. WOX11 and LBDs (16 and 29) have been involved in maintaining stem cell renewal by the upregulation of cell cycle-related genes[32,33]. Therefore, we can assume that MdWOX11, MdLBD16, and MdLBD29 promote LR growth by the upregulation of cell cycle-related genes, which might be key players during LR formation.
In view of the above-mentioned relationships between nitrate, hormones, and related genes in the pathway of LR formation, we studied the molecular and physiological mechanisms that caused these morphological changes in B9 apple rootstock, by the exogenous application of nitrate treatments. We also examined the hormonal content and gene expression levels of nitrate transporters and assimilation, IAA-synthesis, transport and signal transduction, and root development and cell cycle-related genes by nitrate treatment. According to the results of this study, we explained how nitrate regulated LR growth by regulating hormones and gene expression levels. The ultimate objective of this study was to investigate the effects of nitrate treatment on LR formation and development from physiological and molecular approaches and to provide the foundation for apple LR regulation.
-
To determine the appropriate amount of nitrate, we used five different nitrate treatments. The phenotype of the control group was compared with the nitrate-treated groups to compare shoot and root morphological parameters shown in Fig. 1. The nitrate treatments stimulated the nursery plant's growth. The root growth is related to aboveground tree growth; therefore, plant height (cm) and stem diameter (mm) were measured following nitrate treatments. After carefully analyzing the data, it was found that the nursery plant heights and stem diameters were increased by increasing nitrate supply compared to the control group, where control nursery plant heights measured 19.63 cm, 23.5% lower than that of the T5 nursery plants. A similar pattern was observed for stem diameter (Fig. 1b). Moreover, the number of roots in control nursery plants was only 51.33, whereas T5 nursery plants developed 92.66 roots, which was significantly higher than the control group and other treatments (Fig. 1b). The highest root length, 2637.8 cm, was measured in the T5 plants, which was 38.9% higher than in the control nursery plants (Fig. 1b). Interestingly, the root projection area shared similar statistical results with the root surface area, where T5 appeared with a maximum projection area of 102 cm3; however, the lowest was observed in the control group (61 cm3). Moreover, the highest root volumes were observed in the T3, T4, and T5 plants, which had 3.05, 3.09, and 3.11 cm3, respectively, compared with the control, T1, and T2 nursery plants. (Fig. 1b).

Figure 1. Effect of nitrate treatments on the phenotype of roots, scale bar = 1 cm. (a) The root numbers, root length, root volume, root surface area, and root projection area were measured after five weeks of nitrate treatment. (b) B9 apple rootstock nursery plants grown on 1/2 strength Hoagland solution without nitrate served as control plants, for treatment, the amount of nitrate are as a follows: T1, 2.25 mM L−1; T2, 9 mM L−1; T3, 18 mM L−1; T4, 36 mM L−1; and T5, 72 mM L−1. Error bars refer to the average value ± SD from three biological replicates. Asterisks indicate significant differences at * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Root thickness is directly related to the absorption of nutrients. In this study, the roots were classified into four groups (< 0.5 mm, 0.5−2 mm, 2−4.5 mm, and > 4.5 mm) based on their diameter. Root length (cm), surface area (cm2), and volume (cm3) of the control and nitrate-treated nursery plants are shown in Table 1. We found that the categories of 0−0.5 mm and 0.5−2 mm had the highest root length and surface area for all treatment categories, whereas the category > 4.5 mm contained few roots in all measured parameters. Furthermore, T5 nursery plants showed that the values of most measured parameters were higher than that of the control nursery plants ( Table 1). From the results, we can conclude that nitrate-treated nursery plants showed superior roots (particularly T5) over the control group, so we can consider that T5 was an adequate amount of nitrate for LR development in B9.
Table 1. Root length, root surface area, and root volume in control and nitrate-treated B9 apple nursery plants.
Treatments The length of different diameters (cm) The surface area of different diameters (cm2) The volume of different diameters (cm3) 0−0.5 mm 0.5−2 mm 2−4.5 mm > 4.5 mm 0−0.5 mm 0.5−2 mm 2−4.5 mm > 4.5 mm 0−0.5 mm 0.5−2 mm 2−4.5 mm > 4.5 mm CK 1721 ± 129.54 164.4 ± 20.25 5.85 ± 0.12 0.18 ± 0.007 107.06 ± 8.66 45.81 ± 4.60 2.63 ± 0.09 0.52 ± 0.19 0.68 ± 0.06 1.33 ± 0.15 0.20 ± 006 0.07 ± 0.03 T1 1828 ± 50.98 263.7 ± 21.88 6.17 ± 1.54 1.30 ± 0.26 130.64 ± 4.79* 72.27 ± 6.94* 6.08 ± 1.45 2.51 ± 0.59 0.90±0.03 1.87 ± 0.21* 0.48 ± 0.11 0.41 ± 0.11 T2 1904 ± 107.52 235.6 ± 11.41 5.91 ± 0.87 1.16 ± 0.24 134.29 ± 6.13** 63.33 ± 3.44 5.73±0.82 2.37 ± 0.47 0.92 ± 0.03 1.62±0.10 0.45 ± 0.06 0.39 ± 0.07 T3 1986 ± 72.02** 352.1 ± 25.95 7.49 ± 1.08 1.33 ± 0.18 151.05 ± 6.66**** 93.27 ± 7.25**** 7.49 ± 1.06 2.71 ± 0.37 1.11 ± 0.05 2.38 ± 0.15**** 0.61 ± 0.08 0.47 ± 0.07 T4 2049 ± 81.10** 337.7 ± 33.50 6.90 ± 1.33 0.88 ± 0.35 158.60 ± 0.35**** 89.56 ± 9.62**** 6.83 ± 1.30 1.59 ± 0.70 1.15 ± 0.05 2.23 ± 0.26*** 0.553 ± 0.10 0.23 ± 1.11 T5 2306 ± 89.85**** 332.3 ± 16.40 6.03 ± 1.70 0.45 ± 0.14 171.80 ± 8.09**** 87.44 ± 5.61**** 5.64 ± 1.68 0.95 ± 0.36 1.21 ± 0.06* 2.15 ± 0.19*** 0.43 ± 0.13 0.13 ± 0.04 Roots were classified into four different size groups based on their diameter: < 0.5 mm, 0.5−2 mm, 2−4.5 mm, and > 4.5 mm. Values are the means ± SD of three biological replicates. Effect of nitrate treatment on the phenotype of the roots
-
We compared the control nursery plants with the T5 nursery plants in terms of phenotype and morphological parameters throughout this study (0 d, 7 d, 14 d, 21 d, and 28 d) (Fig. 2a, b). After carefully observing the obtained data, it was noted that the root numbers were continuously increasing from 0 d to 28 d in both groups. On 0 d, the LR numbers were recorded at 28.7 in both groups, and on day 28, it was 84.3 in T5 nursery plants, which was 56.1% higher than the control nursery plants (Fig. 2b). A similar trend and pattern was also recorded for root length (cm), with roots being longer in T5 nursery plants at all time points compared with the control group (Fig. 2b). Root volume (cm3) increased from 0 d to 14 d, and a decrease occurred at 21 d in both groups; after that, both increased to 28 d. In addition, root surface area (cm2) and root projection area (cm2) both have similar statistical values and patterns (Fig. 2b).
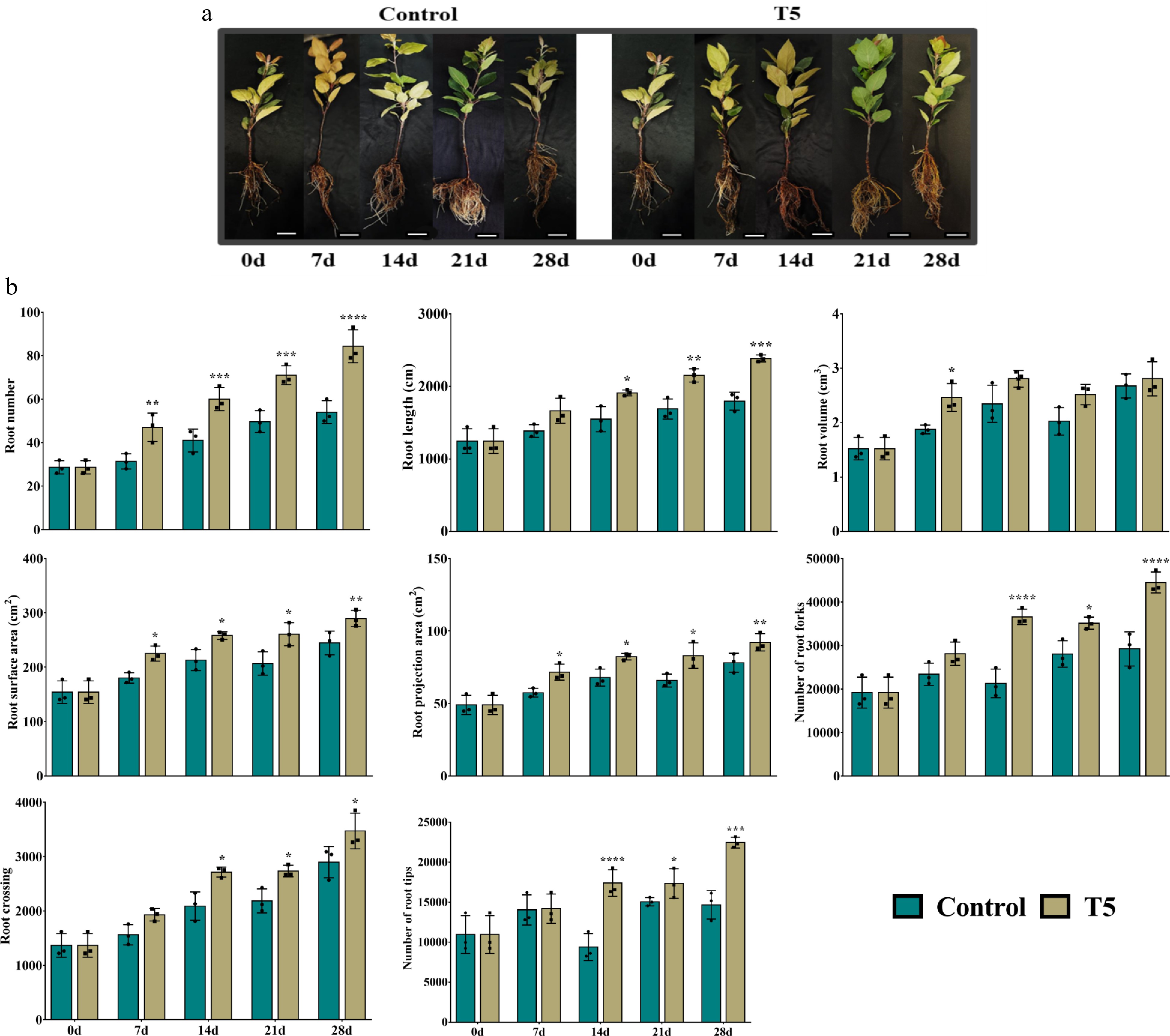
Figure 2. Effect of nitrate treatment (control and T5, 72 mM L-1) on the phenotype of roots, scale bar = 1 cm. (a) The root numbers, root length, root volume, root surface area, root projection area, number of root forks, root crossing, and number of root tips were measured at different time points (0 d, 7 d, 14 d, 21 d, and 28 d). Asterisks indicate significant differences at * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Effect of nitrate treatment on hormone content
-
Hormonal content is directly related to root formation and development. To identify how hormones are altered by nitrate and play crucial roles in developing LRs, we measured the endogenous hormone levels of IAA, GA, ABA, GA3, JA, and ZR in both groups (control and T5) at all time points (0 d, 7 d, 14 d, 21 d, and 28 d) (Fig. 3). After rigorous examination, we found a high rate of IAA, which is generally considered a key hormone for the formation and development of roots. Its high rate is associated with a high rooting rate. From the results, the endogenous concentration of IAA increased steadily in both groups from 0 d towards 28 d (except control at 28 d, where it had decreased in comparison with 21 d) (Fig. 3). The highest concentration of GA was observed at 7 d in T5 nursery plants, which was 49.2% higher than that of control nursery plants; however, it was lower at 14 d and 21 d than control nursery plants, and then a slight increase occurred at 28 d. The ABA content was higher at 0 d and decreased toward 7 d and 14 d in both groups (Fig. 3). The concentrations of GA3 and ZR were higher at 7 d in T5 nursery plants and lower at other time points than in control nursery plants. The JA content was higher at all time points in the control group (except at 14 d) than in the T5 nursery plants (Fig. 3). These results indicate that nitrate treatment dramatically influences the formation and development of LRs at all time points by changing hormonal content.
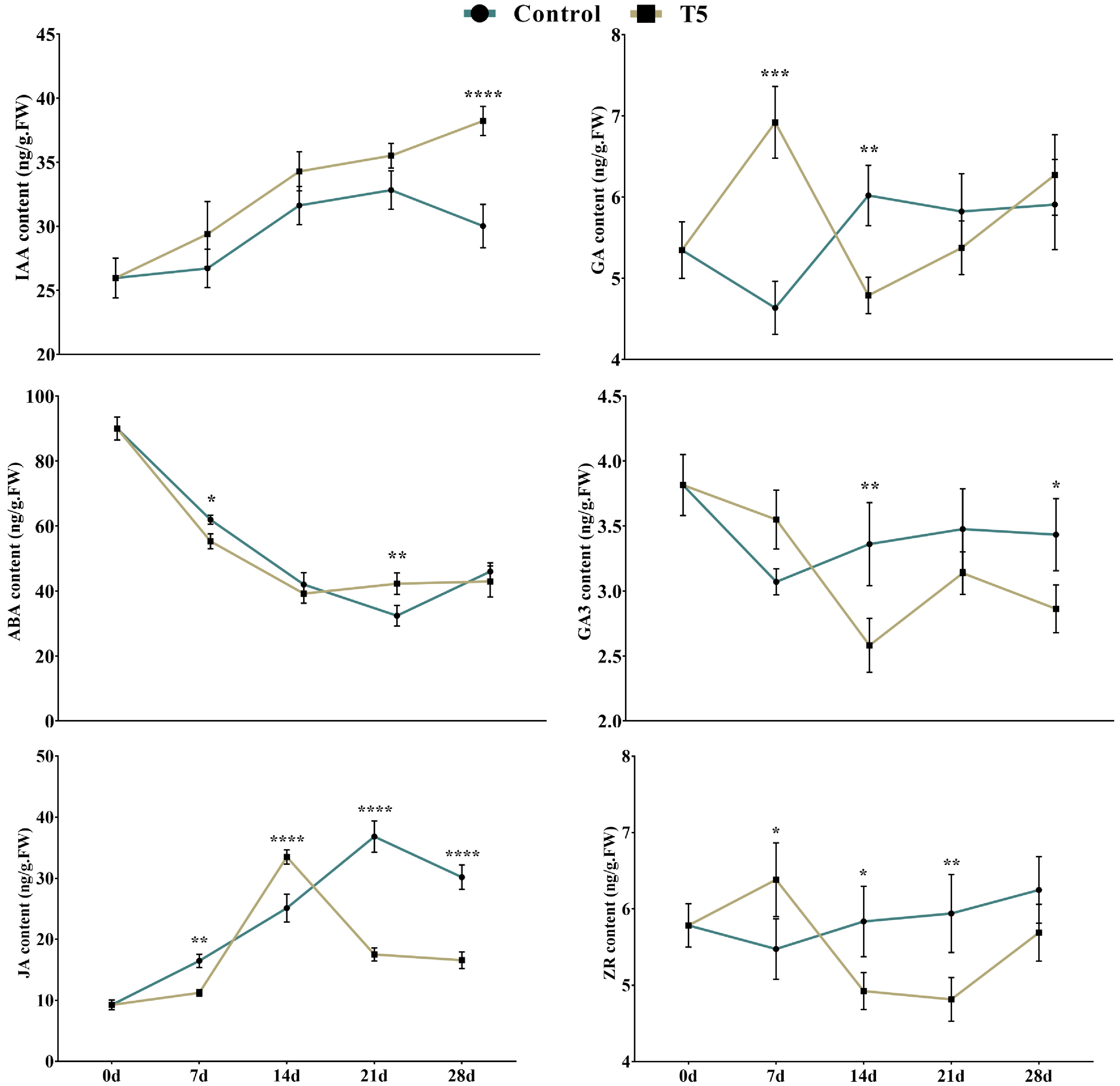
Figure 3. Effect of nitrate treatment (control and T5, 72 mM L-1) on the IAA: indole acetic acid, GA: gibberellic acid, ABA: abscisic acid, GA3: gibberellic acid 3, JA: jasmonic acid. ZR: zeatin riboside. Error bars denote an average value of ± SD from three biological replicates. Asterisks indicate significant differences at * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Effect of nitrate treatment on the relative expression of nitrate-related genes
-
We determined the relative expression levels of a nitrate transporter and nitrate assimilation-related genes using RT-qPCR during the period of this study (Fig. 4). The transcripts of MdNRT1.1, MdNRT2.1, and MdNRT2.4 were upregulated in the control group (except MdNRT2.1 at 28 d) compared with T5 nursery plants (Fig. 4). The expression level of MdNRT2.5 was higher at all time points (except at 28 d) in T5 nursery plants than in the control nursery plants. The transcript abundances of MdNRT3.1 and MdANR1 were higher at 7 d, 14 d, and 21 d in response to the control group (Fig. 4). The expression of MdATG18a seemed higher at 7 d, 14 d, and 28 d in T5 nursery plants compared with that in control nursery plants. Interestingly, the transcripts of MdNIA2 and MdNR were higher at all time points in T5 nursery plants (except MdNR at 28 d) than in the control nursery plants.

Figure 4. Effect of nitrate treatment (control and T5, 72 mM L-1) on the relative expression of nitrate-related genes over the study period (0 d, 7 d, 14 d, 21 d, and 28 d) during LR development. Error bars refer to the average value ± SD from three biological replicates. Asterisks indicate significant differences at * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Effect of nitrate treatment on the relative expression of IAA-related genes
-
The effects of nitrate treatment on the expressions of IAA-synthesis, transport, and signal transduction-related genes were measured (Fig. 5). This section offers different transcript patterns of IAA-related genes at diverse time points during this study. From our outcomes, it seemed that the relative expression of MdPIN1 was higher at 7 d and 21 d in T5 nursery plants and lower at 14 d and 28 d than in the control nursery plants. The transcript abundance of MdPIN2 was upregulated at 7 d and 14 d in response to T5 compared with the control nursery plants (Fig. 5). MdARF7 and MdARF19 exhibited higher levels at all time points (except MdARF7 at 21 d) in T5 nursery plants. MdAUX1 seemed higher at 7 d in T5 nursery plants. In addition, a high transcript abundance of MdIAA5 was noticed at 7 d, 14 d, and 21 d; however, MdIAA14 was highly expressed at 7 d and 14 d in response to T5 compared with the control group (Fig. 5).
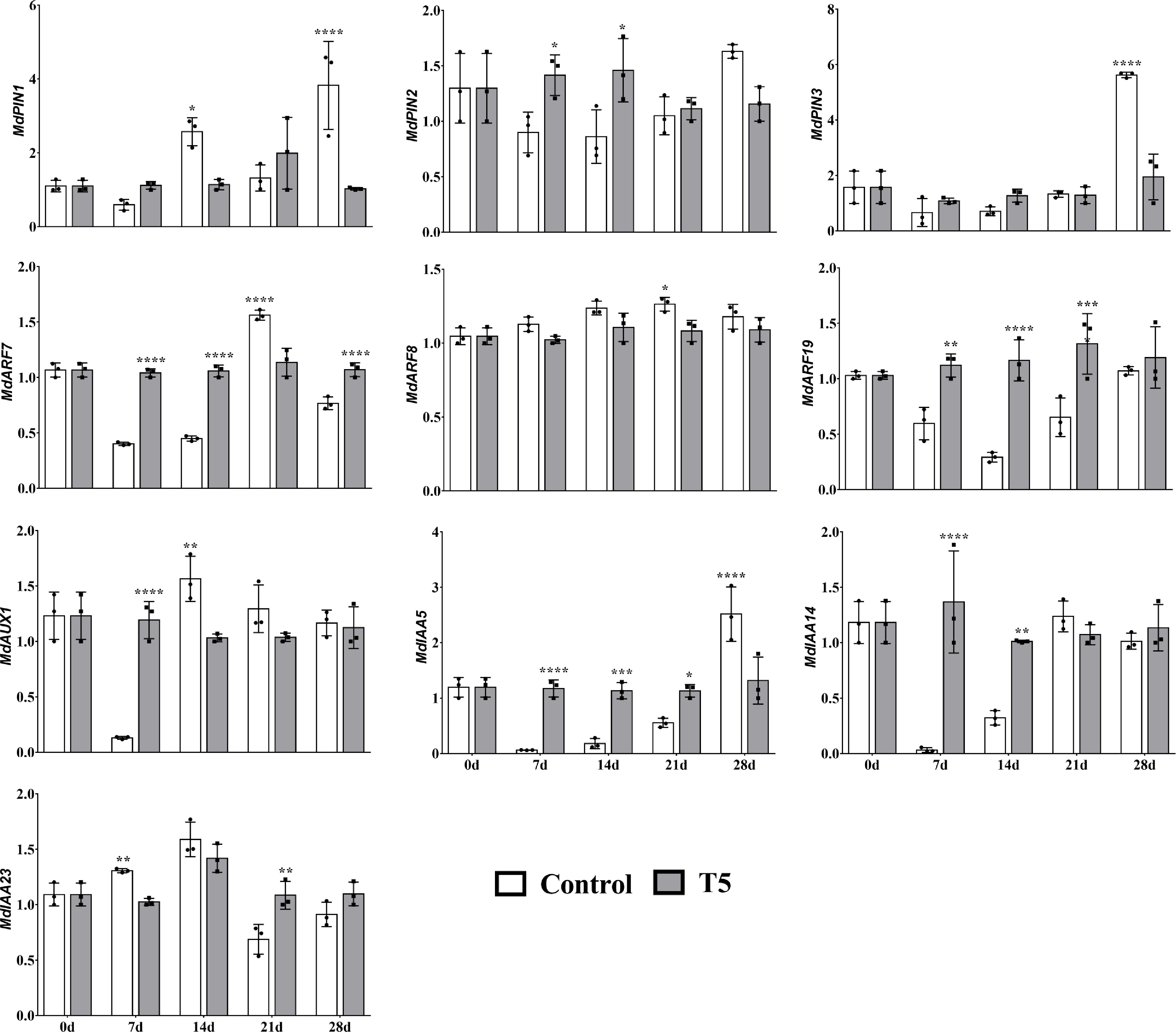
Figure 5. Effect of nitrate treatment (control and T5, 72 mM L-1) on the relative expression of IAA-related genes over this study's time course (0 d, 7 d, 14 d, 21 d, and 28 d) during LR development. Error bars refer to the average value ± SD from three biological replicates. Asterisks indicate significant differences at * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Effect of nitrate treatment on the expression of root development- and cell cycle-related genes
-
The transcript abundances of root development- and cell cycle-related genes related to LR formation and development were assessed. The relative expressions of MdWOX11, MdLBD16, and MdSHR were higher at all time points; similarly, the expressions of MdGATA1 and MdSCR1 were higher at all time points (except at 28 d) in T5 nursery plants than in the control nursery plants (Fig. 6). MdARRO1 was expressed higher at 14 d, 21 d, and 28 d in the control group than in the T5 nursery plants. The expression of cell cycle-related genes, including MdCYCD1;1 and MdCYCP4;1, seemed higher at 7 d, 14 d, and 21 d in T5 nursery plants than in the control nursery plants; however, MdCYCD3;1 was higher at 7 d in T5 nursery plants than in the control nursery plants.
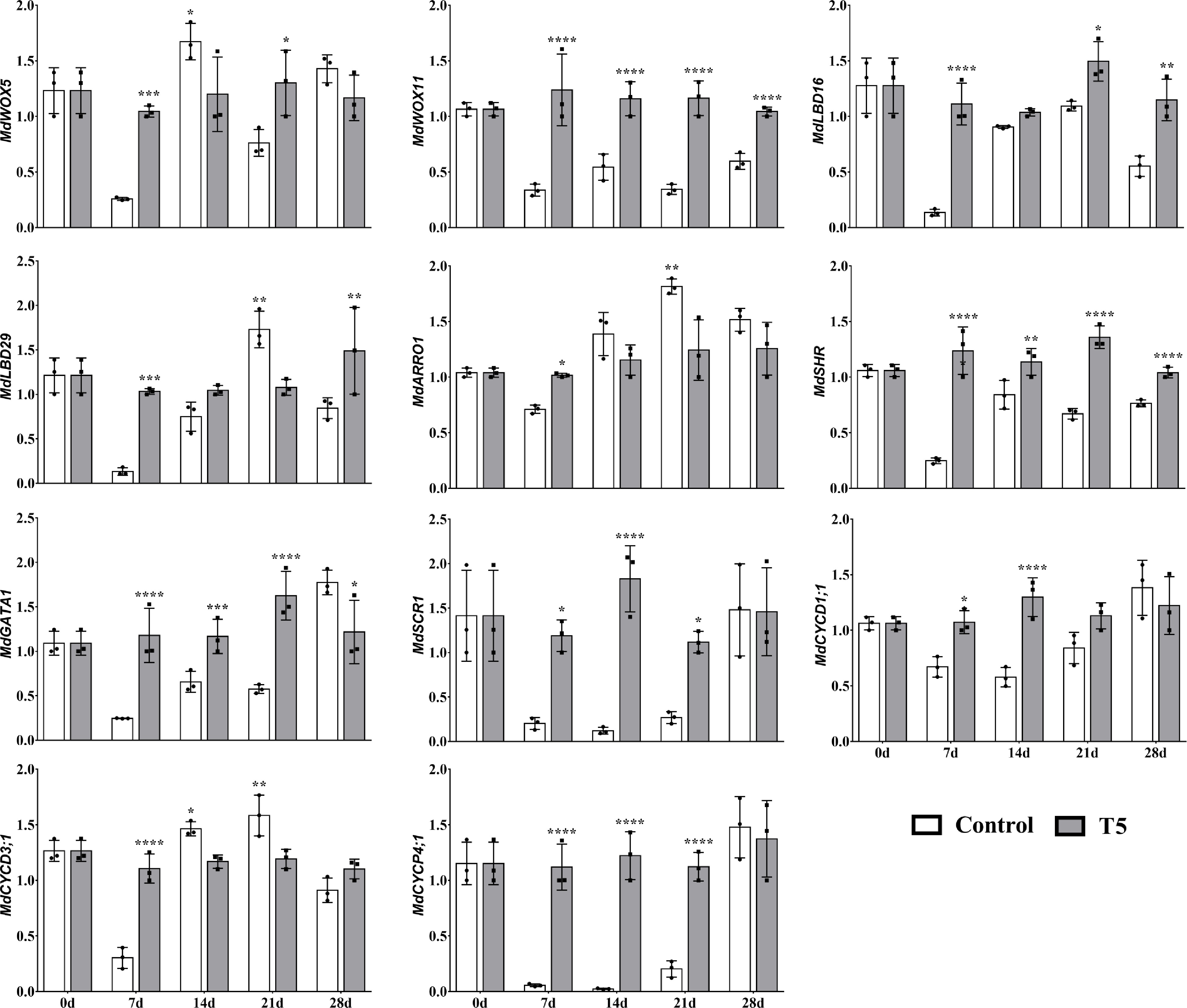
Figure 6. Effect of nitrate treatment (control and T5, 72 mM L-1) on the relative expression of root development- and cell cycle-related genes over the course of the study (0 d, 7 d, 14 d, 21 d, and 28 d) during LR development. Error bars refer to the average value ± SD from three biological replicates. Asterisks indicate significant differences at * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
-
In apple, LR development is separated into three phases: the initiation of LR, the development of LR primordia, and the emergence of LR[34]. Previous studies established that the effect of nitrate on root growth is not exerted by the direct perception of external nitrate, but largely depends on the magnitude of nitrate absorbed by the plant[35−37]. The effect of nitrate on LR development differs between the different concentrations of nitrate. Several nitrate treatments showed that T5 treatment more significantly affects LR growth than the other treatments and control (Fig. 1). From the results, we can determine that T5 was sufficient for LR formation and development in B9 (Fig. 1). In addition, the effect of external nitrate availability on the elongation of LRs is prominently restricted by the increase in the meristematic activity of the mature LR tips[35−37]. Our results showed that T5 had a more significant effect on LR formation and development than the control (Fig. 2). Likewise, the size of the root surface area is closely tied with the area available for root absorption, a vital signal for root activity. Root diameter is also an important signal for root activity, with fine roots being indicated to have a crucial role in the absorption of nutrients and minerals. In this study LRs were divided into four groups based on their available diameters: < 0.5 mm, 0.5−2 mm, 2−4.5 mm, and > 4.5 mm. From this perspective, the categories < 0.5 mm and 0.5−2 mm appeared enriched in all measured parameters in all treatments compared with the control nursery plants ( Table 1).
Plant endogenous hormones are crucial for regulating LRs as they can respond to varying environmental cues, such as drought and the physiological role of silicon in alleviating drought stress[38−40] through a signaling network within the plant, and are effective for cell-fate-determination and cell-specialization. On the other hand, the regulating mechanisms and the contribution of each hormone and their complex signaling networks during LR formation are inadequate. Auxin seems essential for inducing LRs in Malus hupehensis apple seedlings[41]. Hormones are found to be indirectly involved in LR regulation by interacting with auxin[41]. In this regard, the obtained results illustrated that nitrate treatment increased IAA levels (Fig. 3). This upturn in IAA content appears to be important for establishing the induction of LR formation and development in B9 apple nursery plants and may possibly represent a positive physiological response in the roots[42,43]. The content of ABA was higher in the control nursery plants and showed a general decline, and then slightly increased at 28 d, in both groups (Fig. 3). These outcomes are consistent with the previous reports that ABA negatively controls LR formation[24]. However, the ZR level was increased at the induction stage in the T5 nursery plants, which may be triggered by the existence of cellular programming needed for cell division; these results are in line with an earlier report[44]. JA inhibits AR formation[45]; JA concentration increased in the control nursery plants from 0 d to 21 d and decreased at 28 d (Fig. 3). Altogether, the results of the hormonal content analysis indicated that the formation and development of LRs were affected by complex hormonal regulations. In contrast, the underlying mechanism associated with their interactions requires further study.
To understand the underlying mechanism of how nitrate treatment promotes LR growth, we measured the expression levels of nitrate transporters and assimilation-related genes. The NRT genes play various biological roles during nitrate uptake and distribution inside the plant[4]. The identification of two different gene families: NRT1 and NRT2, encodes low- and high-affinity nitrate transporters, respectively. NRT1.1 represses LRs at low nitrate concentrations by its auxin transport activity that lowers LR auxin accumulation[46,47]. The expression of MdNRT1.1 was upregulated in control nursery plants; thus, we can propose that fewer roots (in response to control) are related to the higher expression of MdNRT1.1 and lower IAA level compared with T5 nursery plants (Figs. 3 & 4). Expression of HATS-type gene, NRT2.1, is inhibited by high nitrate concentrations and activated by low nitrate concentration. Additionally, the transcript of MdNRT2.1 was observed higher in response to control nursery plants during the course of this study (except at 28 d); conversely, the expression showed a first increasing trend at 7 d, 14 d, and 21 d, and decreased expression levels were detected towards 28 d in the control group (Fig. 4). MdATG18a is functionally involved in nitrate uptake, transport, and assimilation by the upregulation of high-affinity nitrate transporters (MdNRT2.1 and MdNRT2.5) and nitrate reductase (MdNIA2)[48]. In this study, we observed that MdATG18a might positively control nitrate transport and assimilation during LR growth by affecting the expression of MdNRT2.5 and MdNIA2, whose expression levels were higher at all time points (except MdNRT2.5 at 28 d), but did not affect MdNRT2.1 (Fig. 4). NIR and NR are involved in the process of reducing nitrate to ammonium in tight regulation. NR is prompted by nitrate and its expression depends on external nitrate[49]. From our results, MdNR expression was upregulated in T5 nursery plants compared with the control at 7 d, 14 d, and 21 d (Fig. 4), which suggested that MdNR might play a key role in the regulation of LRs in B9 apple rootstock. Further work is needed to understand the roles of nitrate-related genes during LR growth.
Regarding auxin signaling pathways, the degradation of AUX/IAA promoted ARF expressions, which in succession stimulated the transcription of the auxin-responsive gene[26]. Several genetic studies have revealed the involvement of ARFs in the processes of plant growth. In Arabidopsis, ARF7 and ARF19 genes exhibited particular dynamic gene expression patterns in the development of seedlings, rooting, and embryogenesis[50,51]. The arf7 and arf19 single mutants reduced ARs and LRs, but double mutants triggered significantly fewer roots[50,51]. Agreement was found with this study, where MdARF7 and MdARF19 expression was higher at all time points (except MdARF7 at 21 d) in T5 nursery plants than in the control nursery plants (Fig. 5), so they are considered to be critical players in the growth of apple LRs. PIN-FORMED (PIN) auxin polar transport maintains the auxin gradient in root tips, triggering root growth[52]. Some mutants influence homeostasis, which in turn affects adventitious rooting. The expression of the rice mutant (OsPIN1) was involved in polar auxin transport and affected the process of AR emergence and tillering. They established that the auxin level and distribution are imperative in different organs[53]. Additionally, the member of the PIN family regulates the auxin efflux system and behaves as a promoter for auxin efflux in the plasma membrane, which is crucial for root growth. Conversely, not every pin mutation affects root growth, and some pin mutations show strong defects in root formation[54]. Unsurprisingly, this study indicated that the transcript levels of MdPIN1, MdPIN2, and MdPIN3 were upregulated at most time points, which shows that these genes have a primary role in LR regulation (Fig. 5).
SHORT ROOT (SHR) is also knotty with the formation of the LR stem cell niche and AR's meristem[55], where SHR is directly targeted by SCARECROW (SCR) and plays a key role in LR development and maintains the stem cell niche in LRs[55]. In addition, SCR also regulates the radial organization of the root and is required cell autonomously for distal specification of the QC, which in turn regulates the stem cell fate of the immediately surrounding cells. This study revealed that MdSCR1 and MdSHR regulated LR growth in B9 (Fig. 6). In Arabidopsis, auxin promotes the expression of WOX11/12 and its protein binds to the WOX5/7 promoter to trigger their transcripts[31]. WOX11 also stimulates the transcript abundances of MdLBD16 and LBD29[56]. Moreover, LBD29 and WOX5 are also involved in maintaining the renewal of stem cells by regulating cell cycle-related genes[57,58]. Because of the above-mentioned reports, we suggest that endogenous auxin accumulation promotes the MdWOX11 expression, which activates the MdLBD16 and MdLBD29 expression. Combined expressions of these genes promoted LR development by the upregulation of cell cycle-related genes (MdCYCD1;1 and MdCYCP4;1) in T5 nursery plants (Fig. 6).
-
This study showed that nitrate application promoted LR formation and development in B9 nursery plants by elevating the endogenous concentrations of IAA and GA, and by reducing the ABA, JA, and ZR content at most time points. Furthermore, nitrate treatment upregulated the expression of nitrate-related genes as well as auxin biosynthesis and transduction-related genes, which led to an increase in auxin levels at all time points. Subsequently, the expression levels of MdSHR, MdGATA1, and MdSCR1 were prompted, leading to an increase in LR development. The current study shows the involvement of nitrate in the growth of LRs by an IAA-mediated pathway, where higher auxin concentrations elevated MdWOX11 expression, which controls the MdLBD16 and MdLBD29 expressions, with the combined effects of these genes stimulating LR development by inducing cell cycle-related genes. This study suggests that the involvement of endogenous hormones, specific genes, and pathways in nitrate metabolism mediates LR formation and development. The particular mechanism related to crosstalk between nitrate and other hormones needs further research.
-
Morphological homogenous and healthy B9 apple rootstock nursery plants were used as test materials and were obtained from Shaanxi QingMei Biotechnology Co., Ltd. B9 nursery plants were grown on 1/2 Hoagland solution (formulation is shown in Supplemental Table S1) for 15 d to recover from the transplant shock. The solution was replaced weekly. The hydroponic culture details are presented in Mao et al.[32]. In the preliminary experiment, 108 nursery plants were cultured hydroponically for five weeks and treated with five different nitrate treatments, namely T1, 2.25 mM L−1; T2, 9 mM L−1; T3, 18 mM L−1; T4, 36 mM L−1; and T5, 72 mM L−1, without nitrate served as a control (details are shown in Supplemental Table S2). After identifying the suitable amount of nitrate, a total of 162 fresh B9 nursery plants were cultured, of which half were treated with nitrate (T5), and the remaining served as a control to explore the mechanism by which nitrate increases LR growth.
Measurement of root morphological parameters and endogenous hormones
-
Root morphological parameters, including root length (cm), surface area (cm2), volume (cm3), and others, were measured using a scanner, and the images were analyzed by WinRHIZO, Regent instrument (Inc., Quebec, Canada) as explained by Mao et al.[32]. Plant height (cm), stem diameter (mm), and root numbers were calculated as described by Mao et al.[32]. Samples were collected at 0 d, 7 d, 14 d, 21 d and 28 d, with three biological replicates for each treatment at each time point. The harvested samples were immediately dipped in liquid nitrogen and stored at −80 °C until further use. Approximately 0.6 g (200 mg per replicate) of fresh weight of LR was harvested from the control and T5 treated nursery plants at 0 d, 7 d, 14 d, 21 d and 28 d to measure hormone levels. The extraction and purification of gibberellic acid 3 (GA3), abscisic acid (ABA), brassinosteroids (BR), jasmonic acid (JA), zeatin riboside (ZR), gibberellic acid (GA), and indole-3-acetic acid (IAA) content were performed according to our previously published method[33]. Three biological replicates were carried out for each treatment at each time point.
RNA extraction, cDNA synthesis, and RT-qPCR analysis
-
Total RNA was extracted using Plant RNA Purification Reagent in line with supplier's protocol (Invitrogen), and the genomic DNA was removed using rDNase I RNase free (Takara). The RNA quality was checked by 2100 Bioanalyser (Agilent Technologies, Santa Clara, USA) and subsequently confirmed by ND-2000 (NanoDrop Technologies). The transcript abundances of nitrate-, IAA-, root development- and cell cycle-related genes were analyzed using RT-qPCR and gene primers were designed using Primer 6.0 (Supplemental Table S3). The RT-qPCR assay was conducted as described in our previous work[59]; for normalization, an apple ACTIN gene was used[4,33,60]. Three biological and three technical replicates were made. The selected gene's relative expression levels were measured using the 2−ΔΔCᴛ method[61].
Statistical analysis
-
Graphs were plotted and statistically evaluated using GraphPad Prism version 7.00 for Windows (One-way ANOVA followed by Dunnett's multiple comparison test (Fig. 1) and Two-way ANOVA followed by Sidak's multiple comparison test (Figs. 2−6) GraphPad Software, San Diego California USA ( https://www.graphpad.com). p < 0.05 was considered statistically significant. The results are presented as mean ± SEM. ns = p > 0.05, * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
- This work was financially supported by the National Key Research and Development Program of China (2018YFD1000101, 2019YFD1000803), Shaanxi Apple Industry Science and Technology Project (2020zdzx03-01-04), Tang Scholar by Cyrus Tang Foundation (C200022002), The China Apple Research System (CARS-27), Tang Scholar by Cyrus Tang Foundation and Northwest A&F University.
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 1/2 Hoagland nutrient solution formula.
- Supplemental Table S2 1/2 Hoagland nutrient solution contains 2.25 mM L−1 of KNO3. The formulation of treatments is as follows.
- Supplemental Table S3 Sequence of primers used for expression analysis, F for the former primer, R for the reverse primer.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Tahir MM, Zhang X, Shah K, Hayat F, Li S, et al. 2021. Nitrate application affects root morphology by altering hormonal status and gene expression patterns in B9 apple rootstock nursery plants. Fruit Research 1: 14 doi: 10.48130/FruRes-2021-0014

Nitrate application affects root morphology by altering hormonal status and gene expression patterns in B9 apple rootstock nursery plants
- Received: 03 August 2021
- Accepted: 26 November 2021
- Published online: 14 December 2021
Abstract: Lateral roots (LRs) are critical for absorbing water, minerals, and nutrients. Nitrate is a vital signaling molecule for regulating LR growth. However, understanding how nitrate interacts with endogenous hormones and genes to coordinate LR development is a bottleneck. In this study, B9 apple rootstock nursery plants were cultured hydroponically and treated with different nitrate treatments (Control; T1, 2.25 mM L−1; T2, 9 mM L−1; T3, 18 mM L−1; T4, 36 mM L−1; and T5, 72 mM L−1), where T5 outperformed the other treatments. Nursery plants were subsequently treated with control and T5 to explore the underlying physiological and molecular mechanisms by which nitrate promoted LR growth. Plant height, stem diameter, root morphological parameters, endogenous hormones, and gene expression levels were measured. Results showed that T5 promoted LR growth by increasing the endogenous contents of indole-3-acetic acid (IAA) and gibberellic acid (GA), and by decreasing the contents of abscisic acid (ABA), jasmonic acid (JA), and zeatin riboside (ZR) at most time points. Furthermore, the relative expression levels of nitrate transporter genes were upregulated in the control group, and nitrate assimilation genes were upregulated in T5 nursery plants at various time points. In addition, the transcript abundances of auxin-related genes were higher in T5 nursery plants, which increased auxin contents. MdSHR, MdGATA1, and MdSCR1 expressions were induced, hence increasing LR growth. In contrast, higher auxin content raised MdWOX11 expression, which enhanced the MdLBD16 and MdLBD29 expression levels, thus prompting the transcripts of cell cycle-related genes: MdCYCD1;1 and MdCYCP4;1. Overall, the changes in hormonal content and gene expression levels followed superior LR growth by T5.
-
Key words:
- Apple rootstock /
- Nursery plants /
- Nitrate /
- Lateral roots (LRs) /
- Formation and development /
- Hormones /
- Gene expression