









-
MADS-box genes regulate diverse aspects of growth and development in flowering plants, including photoperiod response and flowering time control[1], flower meristem determination and floral organ identification[2], pollen fertility regulation[3], seed and fruit development[4], and endocarp development and vegetative development regulation[5]. Typically, MADS-box transcription factors are involved in specifying the identity of floral organs, including sepals, petals, stamens, and carpels[6], the genetic formation control of which was deciphered with a genetic model (ABC model) was recently advanced to the ABCDE model[7]. Briefly, class A genes including APETALA1 (AP1) in Arabidopsis thaliana[8], are needed for the formation of sepals. Represented by AtAPETALA3 (AtAP3), AtPISTILLATAb(AtPI), class B plus class A genes determine the development of petals[9]. AtAGAMOUS (AtAG), a representative class C gene, functions alone to control the formation of carpels, Class B plus class C genes are required for the identification of stamens[10]. Class D genes, such as AtSEEDSTICK (AtSTK), individually regulate the specification of ovules[11], while class E genes represented by AtSEPALLATA (AtSEP) genes such as AtSEP1, AtSEP2, AtSEP3, and AtSEP4, are necessary for establishing the identity of petals, stamens, and carpels[12].
Different lineages from B-class MADS box genes can give rise to distinct phenotypes. AtAP3 and AtPI function as B-class MADS box genes in Arabidopsis[13,14], DEFICIENS (DEF) and GLOBOSA (GLO) constitute B class MADS box genes function activity in A. majus[15,16]. Mutations in either one of these genes leads to the transformation of petals into sepals and that of stamens into carpels[13−18]. The AP3 lineage gives rise to two AP3-like lineages due to another major duplication at the base of the core eudicots. Two AP3-like lineages contain euAP3 that contains AP3 itself, and TM6, which lacks a representative in A. thaliana[19,20]. Silencing Tomato AP3 (TAP3) in the tomato ovary using a specific promoter resulted in male sterility and parthenocarpy[21]. In consideration of the differences between siliques in Arabidopsis and pome fruit derived from both ovary and hypanthium tissues in apple (Malus domestica), silencing of the MdPI gene confers parthenocarpy and results in the transformation of petals into sepals and of stamens into pistils, respectively[22,23]. In the parthenocarpic fruit (pat) mutant of tomato, autonomous development of ovary is associated with the altered expression of class B MADS box genes, including stamenless (sl), pistillate (pi) and SlDEFICIENS (DEF)[24]. Different from the phenotype of mutation in TAP3, RNA interference (RNAi)-induced reduction in TM6 expression results in different phenotypes with homeotic defects primarily in stamens and normal carpel tissue in appearance[25]. Tomato male sterility may be a consequence of reduced expression of SlTM6[26]. Overexpression of TM6 can partially restore the tap3 second whorl phenotype, but TAP3 and TM6 showed different expression patterns and distinct functions in flower development[25]. Different lineages from class B MADS-box genes have complex biological functions during flower development, these orthologues in pear remain largely unrevealed.
Ectopic expression is sufficient for the identification of classic B function. Ectopic expression of AtAP3 results in the conversion of carpels into stamens, and fails to form a functional gynoecium, resulting in a female sterile flower[13]. Flowers of plants ectopically expressing AtPI exhibit different phenotypes, with partial replacement of sepals by petals[9]. Plants overexpressing AtAP3 and AtPI together produce flowers containing two outer whorls of petals and inner whorls of stamens and the early flowering and leaf curling[9]. Overexpression of MdPI results in the complete conversion of sepals into petals and in the production of distinctly flattened fruit as a consequence of restricted cell expansion[27]. Despite that, overexpression of TM6 in wild-type background leads to the production of normal floral identification[25], given that TM6 has more prominent expression in the stamens and carpels, the function of TM6 and its roles in fruit development require further understanding.
The down-regulation of TAP3 leads to parthenocarpic fruit set in tomato[21] and the members of B class MADS-box were down-regulated during fruit set[28]. Exogenous application of gibberellic acid 4+7 (GA4+7) reduced the expression of PbTM6a and PbTM6b in parthenocarpic pear ovary[29]. It compelled us to figure out the function of PbTM6 on the fruit set process in pear. Silencing TM6 in Medicago truncatula[30] or in tomato[25] or mutating FaTM6 in in the octoploid Strawberry leads to defects in the anthers and low viability of pollen grains[31]. Because of the long breeding cycle and arduous genetic transformation of pear, investigation of the function of candidate genes in this plant species is difficult. Research related to PbTM6 genes has not been reported in pear to date.
In this study, we explored the potential role of PbTM6 downstream GA4+7 induced in pear, by generating transgenic tomato lines overexpressing PbTM6a. Specifically, we examined the function of PbTM6a in fertility, and preliminarily explored the regulatory mechanism underlying its role in male sterility. Our results have shed light on the function of PbTM6a in pear, and broaden the function of class B MADS box genes.
-
Sixteen-year-old 'Dangshansu' pear trees grafted onto Pyrus betulifolia Bge rootstocks were used as materials for sampling flowers, which were collected from pear experimental base of Northwest A&F University located in MeiXian County, Shaanxi Province, China (34.28° N, 108.22° E; 562 m). Samples of all treatments were immediately frozen in liquid nitrogen after removal, and stored at −80 °C.
Tomato Ailsa Craig (Solanum lycopersicum L.), received from Xiangqiang Zhan, from Northwest A&F University, Yangling, China, was used for genetic transformation. Tomato planting conditions: 25 °C during the day, 18 °C at night, and photoperiod 18/6 h.
Phylogenetic analysis and sequence alignment
-
The MADS-box proteins from ABCDE model were retrieved from NCBI database (
www.ncbi.nlm.nih.gov ) and a phylogenetic tree was constructed by the software MEGA X with the Maximum Likelihood method and bootstrap analysis. Boot-strap values were calculated from 1,000 replicate analyses. The constructive methods were as per our previous study[32]. The amino acid sequence alignment of AtAP3, SlTM6, PbTM6a and PbTM6b was carried out through DNAMAN version 9.The protein accessions used were as follows: AtAG (Arabidopsis thaliana, P17839.2); AtAGL11 (Arabidopsis thaliana, Q38836.1); AtAP1 (Arabidopsis thaliana, CAA78909.1); AtAP3 (Arabidopsis thaliana, AAD51902.1); AtCAL (Arabidopsis thaliana, BAH30315.1); AtAGL8 (Arabidopsis thaliana, Q38876.1); AtFUL (Arabidopsis thaliana, OAO94650.1); AtPI (Arabidopsis thaliana, P48007.1); AtSEP1 (Arabidopsis thaliana, OAO95853.1); AtSEP2 (Arabidopsis thaliana, OAP07944.1); AtSHP1 (Arabidopsis thaliana, OAP06129.1); AtSHP2 (Arabidopsis thaliana, NP_565986.1); MdAG = MdMADS15 (Malus domestica, NP_001315863.1)[33]; MdAGL1 = MdMADS14 (Malus domestica, XP_008342376.1)[33]; MdAGL11 = MdMADS10 (Malus domestica, NP_001280931.1)[23]; MdAP1 (Malus domestica, ACD69426.1); MdCAL = MdMADS2 (Malus domestica, XP_008393256.1)[34]; MdDEF = MdMADS23 (Malus domestica, XP_028962429.1; HM122607); MdSEP1 = MdMADS8 (Malus domestica, NP_001280893.1)[35]; MdPI (Malus domestica, CAC28022.1); PbTM6a (Pyrus bretschneideri, XP_009346295.1); PbTM6b (Pyrus bretschneideri, XP_009378222.1); SlAG = TAG1 (Solanum lycopersicum, NP_001266181.1)[36]; SlAGL11 = TAGL1 (Solanum lycopersicum, XP_004241906.1)[37]; SlDEF=TAP3 (Solanum lycopersicum, NP_001234077.2)[25]; SlFUL (Solanum lycopersicum, NP_001294867.1); TAP3 (Solanum lycopersicum, ABG73412.1)[25]; SlPI = TPI = SlFBP1 (Solanum lycopersicum, NP_001234075.2; ABG73411.1; XP_004245202.1)[25]; SlMADS6 = TM6 = SlTDR6 (Solanum lycopersicum, QHB49937.1; ABG48621.1; NP_001311309.1)[25]; SlSEP1 = TM29 (Solanum lycopersicum, NP_001233911.1)[38]; SlTAGL1 (Solanum lycopersicum, NP_001300859.1); SlAGL6 (Solanum lycopersicum, NP_001348459.1); VvAGL11 = VvMADS5 = VvAG3 = (Vitis vinifera, QSX80212.1; A0A217EJJ0.1); VvAP3 = VvAP3a = VvPMADS1 = VvDEF (Vitis vinifera, E0CPH4.1; RVW50380.1)[39]; VvMADS2 (Vitis vinifera, XP_019080194.1); VvMADS1 = VvAGAMOUS/SHATTERPROOF (Vitis vinifera, NP_001268105.1)[40]; VvAG (VvAGAMOUS) = AG1 or AG2 (Vitis vinifera, NP_001268097.1)[41]; AnDEFA (Antirrhinum majus, P23706.1); AnGLO (Antirrhinum majus, Q03378.1).
Tissue-specific expression analysis
-
PbTM6a and PbTM6b specific quantitative primers were designed by their reference sequence on the NCBI website (
www.ncbi.nlm.nih.gov ) (Supplemental Table S1). The flowers were separated into different floral organs including sepals, hypanthium, petals, filament, anther, style, and ovary of two days before anthesis, anthesis and four days after anthesis and then were used for RNA extraction and subsequently tissue-specific expression analysis. Total RNA extraction and Real-time fluorescent Quantitative Polymerase Chain Reaction (qRT-PCR) was performed as previously described[42]. The pear actin-β gene was used as an internal standard for expression analysis (Supplemental Table S1).Subcellular localization
-
The CDS of PbTM6a after the removal of termination codons was cloned into the pCAMBIA2300 vector fusing with the Green Fluorescent Protein (GFP) reporter under the Cauliflower mosaic virus (CaMV) 35S promoter using primers with adaptors (Supplemental Table S2). Agrobacterium mediated transient transformation was used for subcellular localization. Agrobacterium containing recombinant plasmid was revitalized twice and suspended with cell resuspension solution (10 μmol·L−1 MES, 10 μmol·L−1 MgCl2, 200 μmol·L−1 AS, pH = 5.6), then was transformed into tobacco (Nicotiana benthamiana) leaves by injection and the empty vector expressing GFP was used as a positive control. After culture in the dark for 1 d and in the light for 2 d, the back of the tobacco leaf was made into freehand section for GFP observation. The GFP signal was observed with a fluorescence microscope (BX63, OLYMPUS, Japan). Activate and adjust the concentration of the bacterial solution till OD600 = 0.6, then inject the bacterial solution from the back of the tobacco leaf using a syringe.
Agrobacterium-mediated tomato genetic transformation
-
The full-length PbTM6a (LOC103938026) coding sequence (CDs) was isolated from 'Dangshansu' cDNA using primers with adaptors (Supplemental Table S2), and then cloned into the pBI121 vector to generate a recombinant plasmid using a Clone-Express One Step Cloning kit (Vazyme, Nanjing, China), then transformed into Agrobacterium tumefaciens strain LBA4404 by heat-shock method. Agrobacterium revitalized and suspensions prepared were manufactured following the methods as reported in our lab[43]. Tomato genetic transformation was carried out as previously reported[43].
Paraffin sectioning
-
Paraffin sectioning experiments were carried out as previously reported[29]. Samples collected were immediately fixed in formaldehyde–acetic acid–alcohol fixative and stored at 4 °C. The samples were dehydrated in ethanol/xylene and embedded in paraffin, sectioned into 8-µm-thick slices, dried, and stained with safranin and fast green. A microscopic imaging system (BX51 + PD72 + IX71, Olympus, Japan) was used to observe the anatomical images.
Pollen vigor measurement
-
TTC (2,3,5-triphenyl tetrazolium chloride) solution was performed to determine pollen grain viability[44]. Fresh pollen of wild-type and transgenic tomato were evenly sprinkled on the glass slide, respectively. Dripped with a drop of TTC dye solution, and then covered with a cover glass. Prepared glass slides were observed under an optical microscope after incubating at 37 °C for 15−20 min. The reaction of TTC with succinate dehydrogenase in the mitochondria of living cells yielded red formazan, which are used to indicate cell viability. Living pollen were stained in light red and deeper red. Percentage of pollen viability was calculated by the stained pollen number including light red and deeper red pollen, divided by the total pollen number. Percentage of vigorous pollen was calculated by the deeper stained pollen number divided by the total pollen number.
Pollen germination test in vitro
-
Pollen germination in vitro medium was prepared as follows: 12 g sucrose and 0.03 g Ca (NO3)2·4H2O, 0.02 g MgSO4·7H2O, 0.01g KNO3 and 0.005 g H3BO3 were added to 100 ml ddH2O. Agar powder was added to make the final concentration of 0.1% (m/v) after adjusting the pH to 6.5. Next, the medium was heated until the agar completely melted, and then cooled to room temperature. A drop of unset culture medium was added onto the glass slide as the pollen germination bed. The pollen was evenly sprinkled on the culture bed, cultivated for 1 h under dark conditions at 28 °C. A drop of Sigma-aldrich (0.1%) was added onto the glass slide treated above, then covered with a cover glass. After staining for 5 min, the pollen germination situation was assessed under a fluorescent microscope in vitro.
Reciprocal crosses experiment
-
Reciprocal crosses experiment was carried out as previously reported[45]. For the female parent, the stamens were removed two days before flowering to prevent self-pollination. Before and after pollination, the inflorescence was bagged for the prevention of natural hybridization. For the male parent, blooming flowers with bright yellow petals, stamens golden, un-scattered pollen were chosen to take anthers and made into pollen. A pencil's rubber-head was used as a pollinator.
Scanning electron microscope observation of pollen
-
Pollens of PbTM6a-OE and wild type from the day before flowering was collected and glued to an aluminum sample holder with conductive glue. Scanning electron microscope (S-3400N) was applied for the visualization of pollen morphology. Three visual fields of scanning electron microscope were used for the statistics of normal pollen grain.
Hormone content determination
-
The determination of hormone content was performed as previously described[46]. Briefly, tomato stamen samples of wild-type and PbTM6a-OE were collected 2 d before anthesis with at least three replicates per treatment. 0.5−1 g of the sample was added into a 2 mL centrifuge tube, and the weight of each sample was recorded for analysis, 1 mL ethyl acetate was used as the extract and added into centrifuge tube, subsequently with vortex and shaken for 10 min, centrifuged at 12,000 rpm for 10 min, and the supernatant was transferred into a new 2 mL tube. The liquid collected was evaporated to dryness with a nitrogen blower, and then dissolved with 200 μL methanol (50%) and filtered through a 0.22-µm filter membrane before testing. The hormones levels were determined by ultra-performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS) (AB SCIEX Triple TOF 5600+, Darmstadt, IN, USA). UPLC-MS/MS was performed using an ACQUITY UPLC HSS T3 (1.8 mm, Waters, USA) column (2.1 mm × 100 mm). The specific parameters were as in our previous study[46].
-
To confirm which class-B MADS box genes the two PbTM6 genes were orthologues of, phylogenetic analysis of classical MADS box genes were performed. The results showed that both PbTM6 belong to the B-class MADS box genes and were more closely related to TM6 than to TAP3 and AtAP3 (Fig. 1).
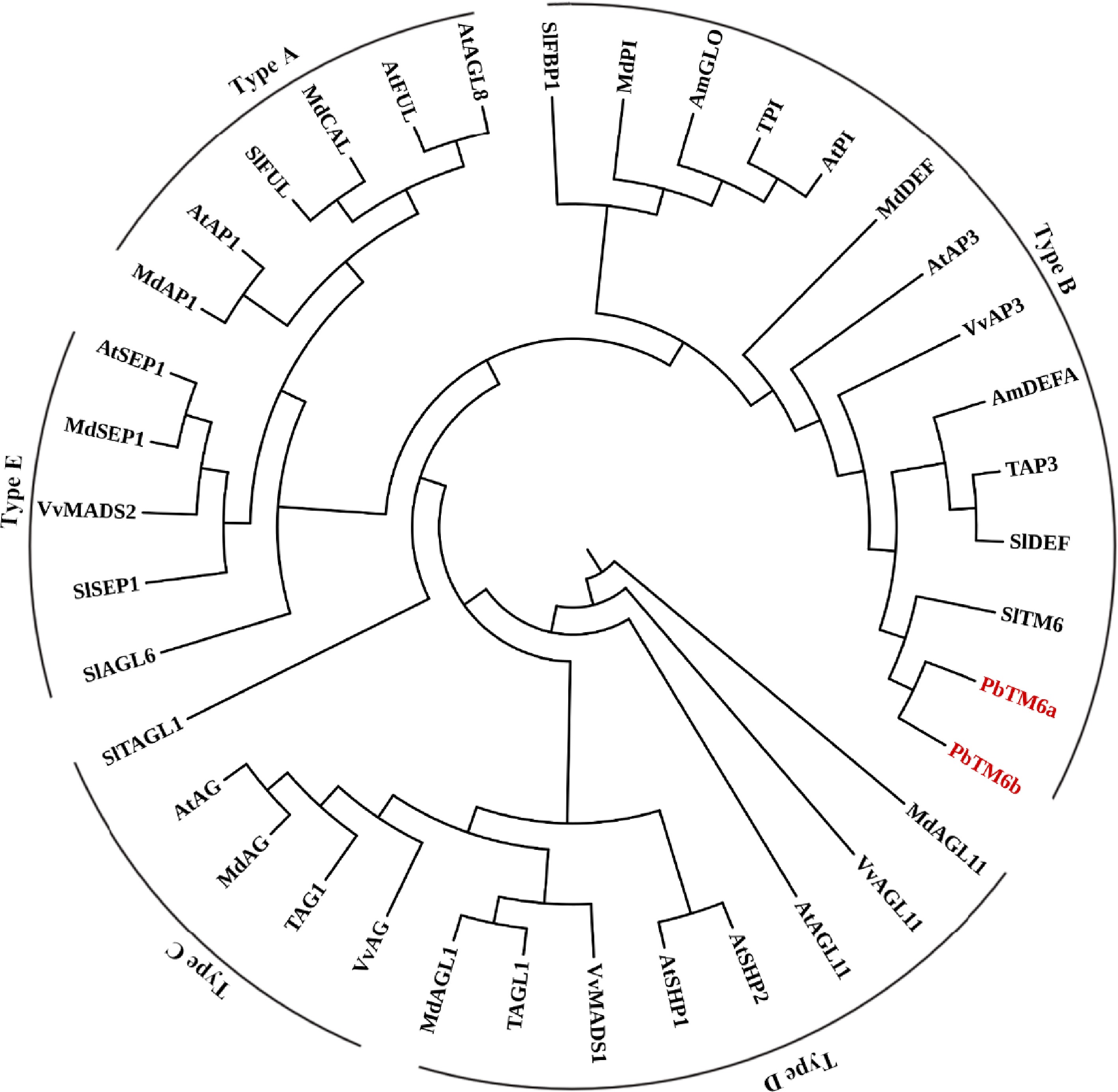
Figure 1.
Phylogenetic analysis of PbTM6 and other MADS box proteins involved in the classic ABCDE model.
Alignment of the amino acid sequences of PbTM6a, PbTM6b, AtAP3 and TM6 showed that all of these proteins possess a highly conserved MADS domain (1-60 aa) at the N-terminus, and the amino acid sequences of PbTM6a and PbTM6b were highly similar (Fig. 2a). Expression levels of PbTM6a and PbTM6b in the flower organs of pear were analyzed by qRT-PCR at developmental stages (2 d before anthesis, at anthesis and 4 d after anthesis) (Supplemental Fig. S1). The results showed that PbTM6a and PbTM6b shared similar expression patterns. The expression levels of PbTM6a and PbTM6b were high in anthers, filaments and petals, which suggested that these genes play important roles in the development of male reproductive organs and petal development (Fig. 2b & c). The expression of PbTM6a and PbTM6b was down-regulated in pear fruitlets by treatments of GA4+7 (Supplemental Fig. S2). The down-regulation of PbTM6a and PbTM6b might participate in the pear fruit set process. Subcellular localization analysis via the heterologous expression of a PbTM6a-GFP fusion in Nicotiana benthamiana leaf epidermal cells showed a fluorescence signal exclusively located in the cell nucleus (Supplemental Fig. S2b), indicating that PbTM6a is a nuclear-localized protein. In summary, our results showed that PbTM6a and PbTM6b correlate with B-class MADS-box genes, and highly expressed in male reproductive organs and petals.
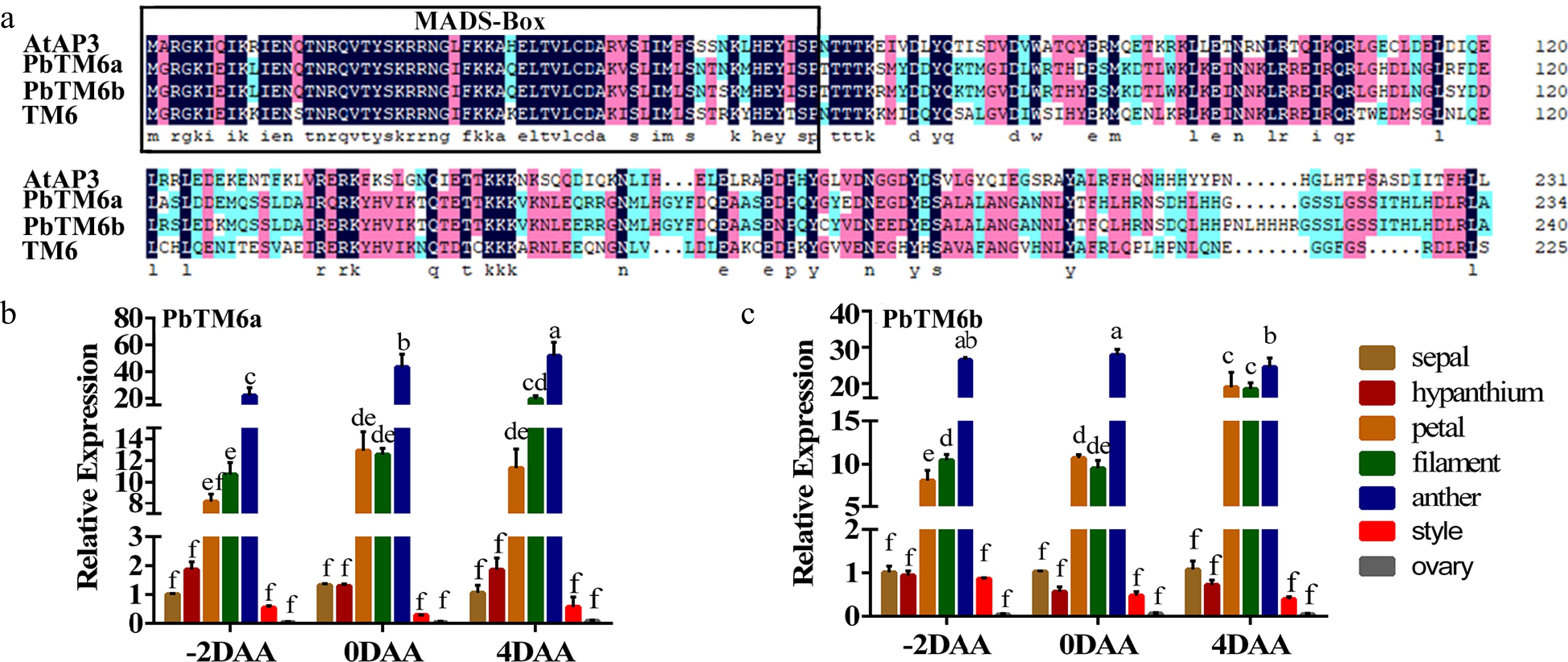
Figure 2.
(a) Amino acid sequence alignment of AtAP3, PbTM6a, PbTM6b, and TM6. MADS-box conserved domains are outlined in black. Expression of (b) PbTM6a and (c) PbTM6b in different floral organs of pear at different developmental stages. −2 DAA, 2 d before anthesis; 0 DAA, anthesis; 4 DAA, 4 d after anthesis. Data represents mean (± standard deviation (SD). Significant differences (P < 0.05) among treatments are determined by one-way analysis of variance (ANOVA), indicated with different lowercase letters.
Heterologous expression of PbTM6a in tomato reduces seed number and size
-
Since the expression patterns of PbTM6a and PbTM6b in floral organs and the amino acid sequences of the encoded proteins were highly similar, PbTM6a had higher abundance and more significantly decreased expression in parthenocarpic ovaries than PbTM6b[29], PbTM6a was chosen for further analysis.
To explore the potential function of PbTM6a in floral organ identity and fruit development, three PbTM6a overexpression (PbTM6a-OE) lines were generated in tomato (Supplemental Fig. S3a). It appeared similar between the wild type and PbTM6a-OE lines during the vegetative growth phase. Plant height and growth as leaf size, shape and color of wild-type and PbTM6a-OE plants were nearly uniform (Supplemental Fig. S3b). Overexpression of PbTM6a seemed to not affect the formation of floral organs compared with the wild type in appearance (Supplemental Fig. S3c). Moreover, flower and fruits of the wild type and PbTM6a-OE lines appeared similar during fruit development (Fig. 3a). The number and size of seeds were significantly reduced in the mature fruits of PbTM6a-OE lines compared with those of the wild type (Fig. 3b), although the mature fruits of wild-type plant showed traces of retarded seed formation. The statistics of seed number per fruit and seed weight proved the reduced number and size of seeds produced by PbTM6a-OE lines (Fig. 3c & d). Thus, we showed that the overexpression of PbTM6a in tomato reduces seed size and number.
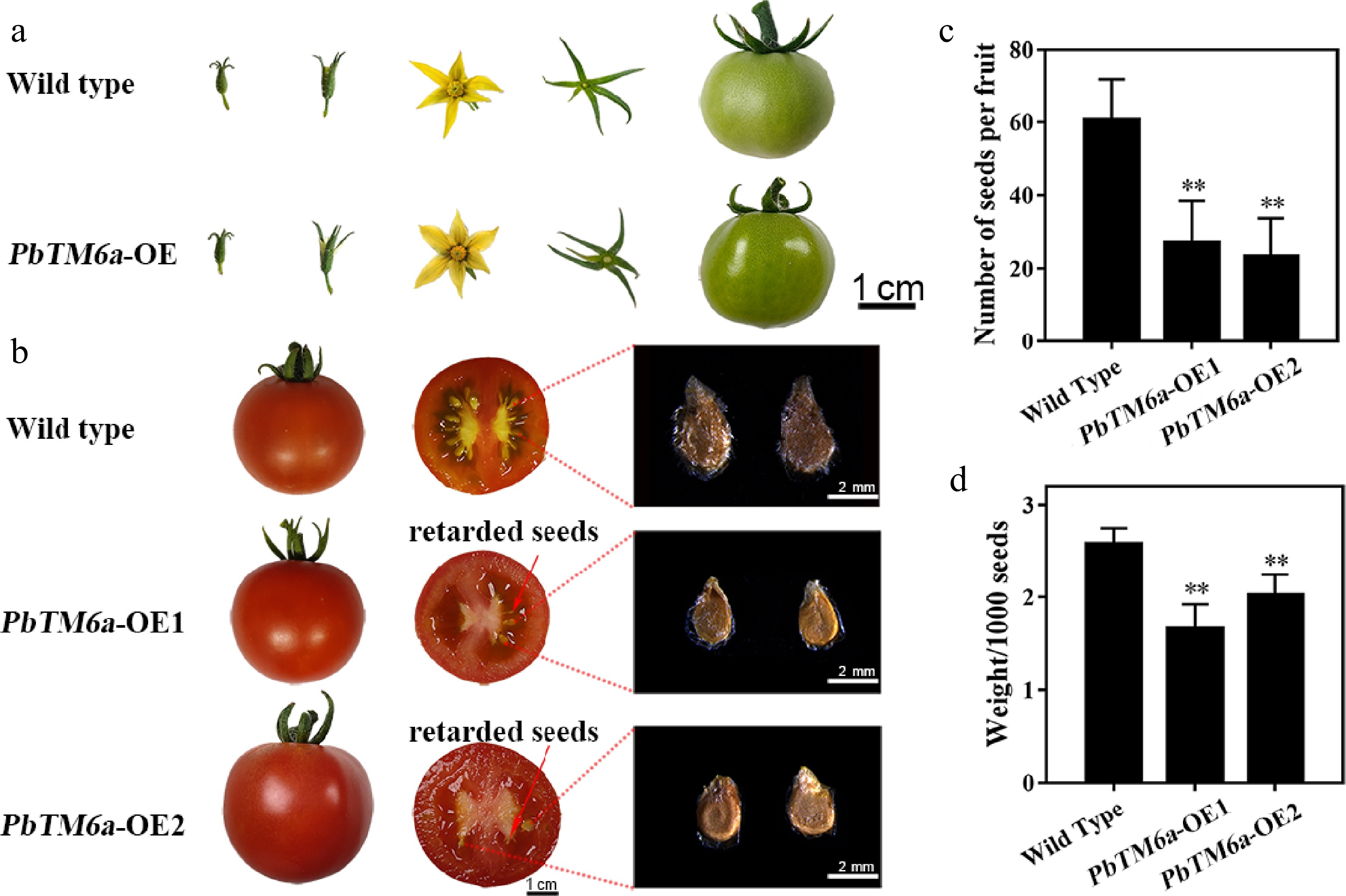
Figure 3.
Phenotypic analysis of wild type and transgenic tomato plants. (a) Comparison of the growth and development of tomato fruits and the structure of floral organs between the wild type and PbTM6a overexpression (PbTM6a-OE) lines. (b) Heterologous overexpression of PbTM6a decreases the number and size of tomato seeds. (c) Number of wild-type tomato plants and PbTM6a-OE lines. (d) Thousand seed weight of wild-type plants and PbTM6a-OE lines. Data represents mean (± standard deviation (SD). Significant differences (P < 0.05) among treatments are determined by one-way analysis of variance (ANOVA), indicated with different lowercase letters.
Transgenic tomatoes with overexpression of PbTM6a produced anthers and ovaries with subtle changes potentially contributed to reduction in seeds size and number compared to wild type
-
To explore the cause of the reduction in seed size and number in transgenic tomato, stamens at the late stage of anther development were selected for preparing and analyzing paraffin sections (Fig. 4a). Both transgenic and wild type lines could produce anthers with similar structure and pollen grains (Fig. 4b−g). In addition, the connective tissues between homolateral pollen sacs were broken in wild type, but the stomium bound to the connective tissue appears to be more strongly formed in PbTM6a-OE anthers, which would influence anther dehiscence.
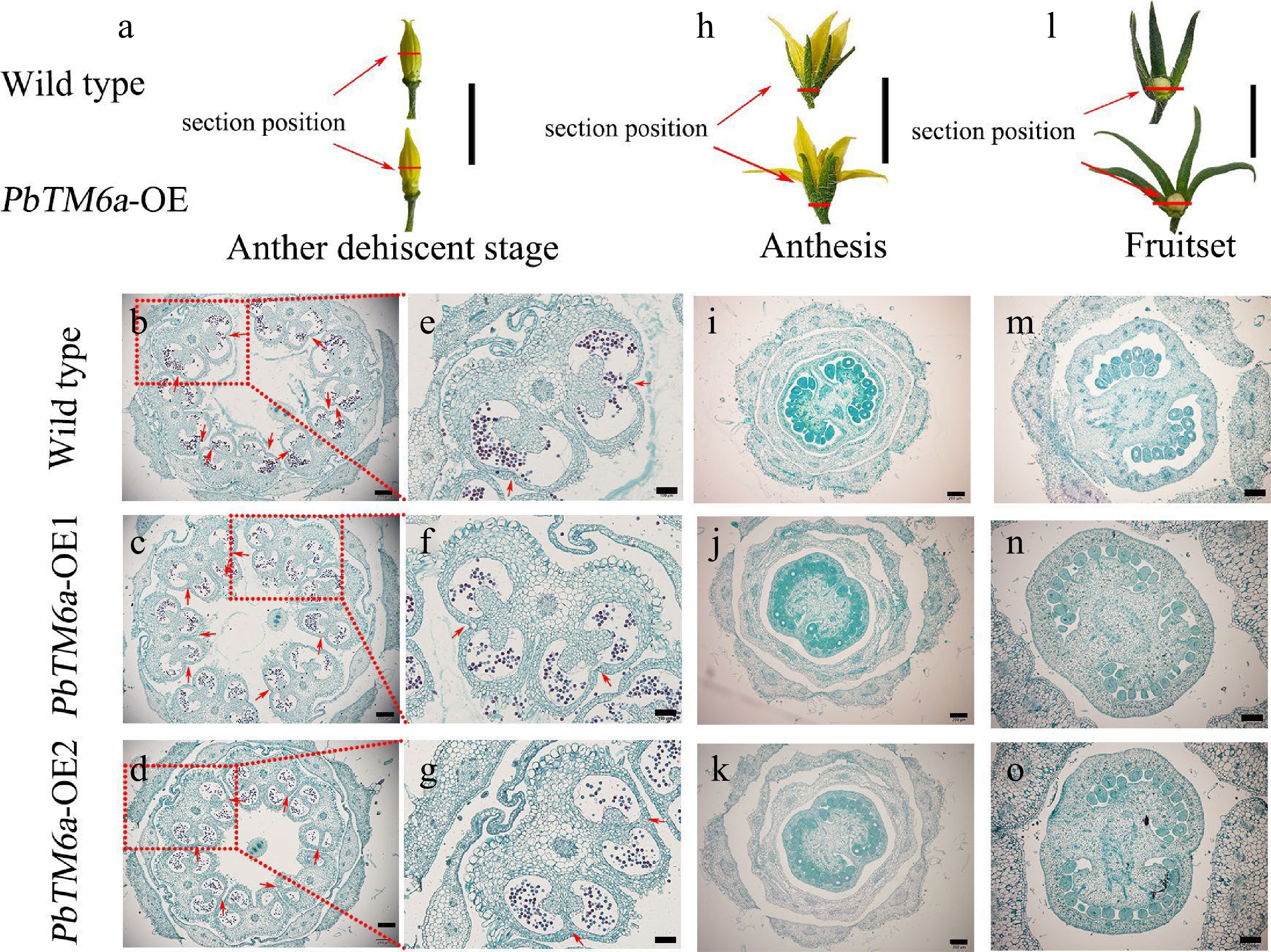
Figure 4.
Morphological and histological observations of floral organs collected from wild type plants and PbTM6a-OE lines. Red lines indicate the section position. (a) Stamens of wild type plants and PbTM6a-OE lines after sepals removed at the anther dehiscence stage. (b)−(d) Histological analysis of anthers of the wild type plants and PbTM6a-OE lines. (e)−(g) Further analysis of anthers shown in (b)−(d). Arrows indicate the position of stomium. (h) Morphology of wild type and PbTM6a-OE flowers at anthesis. (i)−(k) Histological analysis of the cross-section of flowers shown in (h). (l) Analysis of the small tomato fruit of wild-type plants and PbTM6a-OE lines after fruit set. (m)−( o) Histological analysis of the cross-section of ovary shown in (l). Scale bars: 2 cm in (a), (h),( l); 200 µm in (b), (c), (d), (i), (j), (k), (m), (n), (o); 100 µm in (e), (f), (g).
To confirm whether the reduction in seed number was related to defects in the ovule, the ovary was sampled at anthesis and fruit setting for histological observation. Results showed that the number of maternal ovules seemed similar between PbTM6a-OE lines and the wild type (Fig. 4h−k). The total number of ovules in transgenic tomato plants was similar to that in wild type plants even after successful pollination and fertilization (Fig. 4m−o). Moreover, the placenta in transgenic ovaries was larger than that in wild type and occupied the most space of ovaries which might lead to the arrest of ovules development (Fig. 4h−o). These results indicate that the strongly formed stomium and inflated placenta in transgenic tomatoes might contribute to the reduction in seeds size and number.
Reduced vigor and germination of PbTM6a-OE pollen interfere with successful fertilization
-
Because tomato is a self-pollinating crop, defects in pollen germination were speculated as the cause of poor pollination and fertilization. To determine pollen vigor, the pollen of just-opened tomato flowers were selected and used for TTC staining and pollen germination in vitro. Pollen with cell viability was stained red. Pollen with no cell viability was not stained. Most of the pollen of wild type plants was stained deep pink with TTC, while most pollen of PbTM6a-OE lines stained light pink or did not stain (Fig. 5a−c). The results of pollen germination experiments indicated that the majority of wild-type pollens were capable of germination, whereas only a few transgenic pollens could germinate (Fig. 5d−f). The percentage of fertile pollen was approximately 95% in wild-type plants, which was significantly higher than that in PbTM6a-OE1 (8%) and PbTM6a-OE2 (11%) lines (Fig. 5g). Similarly, the percentage of vigorous pollen in wild-type plants (81%) was higher than that in PbTM6a-OE lines (Fig. 5h). The statistics of germinated pollen supported the results of the germination experiment (Fig. 5i). Overall, our results demonstrated that tomato plants overexpressing PbTM6a produced pollen with weak vigor and reduced germination.
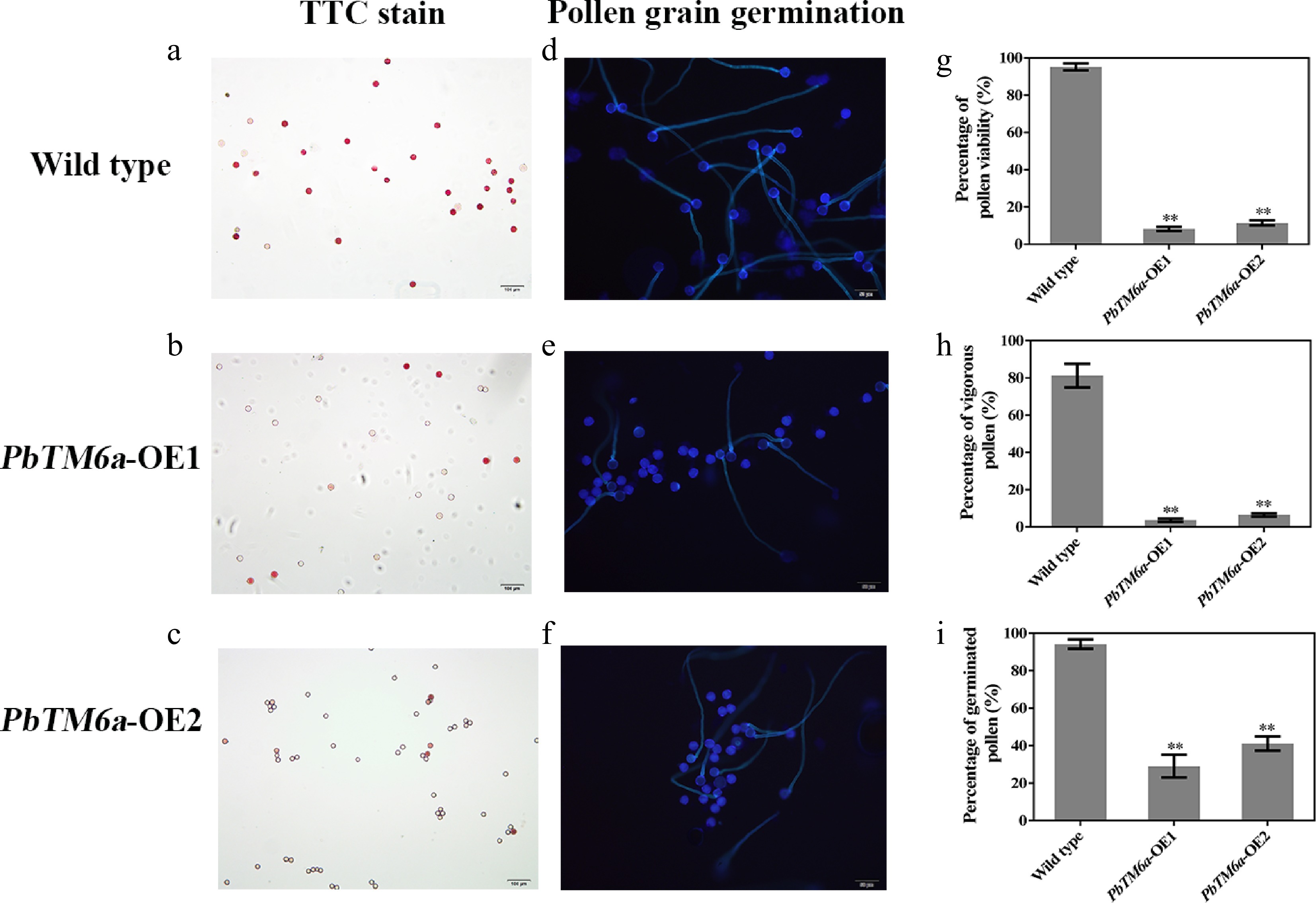
Figure 5.
Assessment of pollen vigor via the viability evaluated by TTC staining and germination in vitro. (a)−(c) Visualization of pollen viability of PbTM6a-OE lines and wild-type plants in vitro by TTC staining. (d)−(f) Evaluation of PbTM6a-OE and wild-type pollen germination in vitro under a fluorescence microscope. Statistics of the percentage of (g) pollen viability, (h) vigorous pollen and (i) pollen germination rate (i) in PbTM6a-OE lines and wild-type plants. Data represents mean (± standard deviation (SD). Significant differences (P < 0.05) among treatments as determined by one-way analysis of variance (ANOVA), are indicated with different lowercase letters.
To determine whether the reduced seed number and size of PbTM6a-OE line were maternally inherited, we carried out reciprocal crosses. Emasculated wild-type tomato plants pollinated with PbTM6a-OE pollen produced lower seed number per fruit because of inadequate fertilization; however the seed number was partly restored in transgenic tomato pollinated with wild-type pollen (Fig. 6a). The statistics of seed number coincide with our observation (Fig. 6b, Supplemental Table S3). Therefore, low-vigor pollen produced by PbTM6a-OE lines appear to be the dominant factor responsible for unsuccessful fertilization, which in turn led to the production of fruits with less seeds.

Figure 6.
Comparison of seed formation and seed number among the progeny of reciprocal crosses and self-pollinations. (a) Seed formation and (b) seed number per fruit produced by PbTM6a-OE (♀) × wild Type (♂) and wild Type (♀) × PbTM6a-OE (♂) reciprocal crosses and wild-type and PbTM6a-OE self-pollinationas. Data represents mean (± standard deviation (SD). Significant differences (P < 0.05) among treatments are determined by one-way analysis of variance (ANOVA), indicated with different lowercase letters.
Low viability of pollen was caused by defective pollen grain development
-
To determine the cause of the low viability of pollen, we observed pollen morphology by scanning electron microscopy (S-3400N). The pollen grain displayed a smooth and plump surface in wild-type (Fig. 7a), while transgenic pollen showed a sunken surface which was plump in appearance (Fig. 7b, c). Statistics showed that PbTM6a overexpressing tomato lines produced much less normal pollen grains compared to that of wild-type (Fig. 7d). Collectively, these findings led us to conclude that PbTM6a overexpression in tomato led to defective pollen grains with low vitality and germinating capacity, causing poor pollination and fertilization, which contributed to the formation of fewer seeds per fruit compared with the wild type.
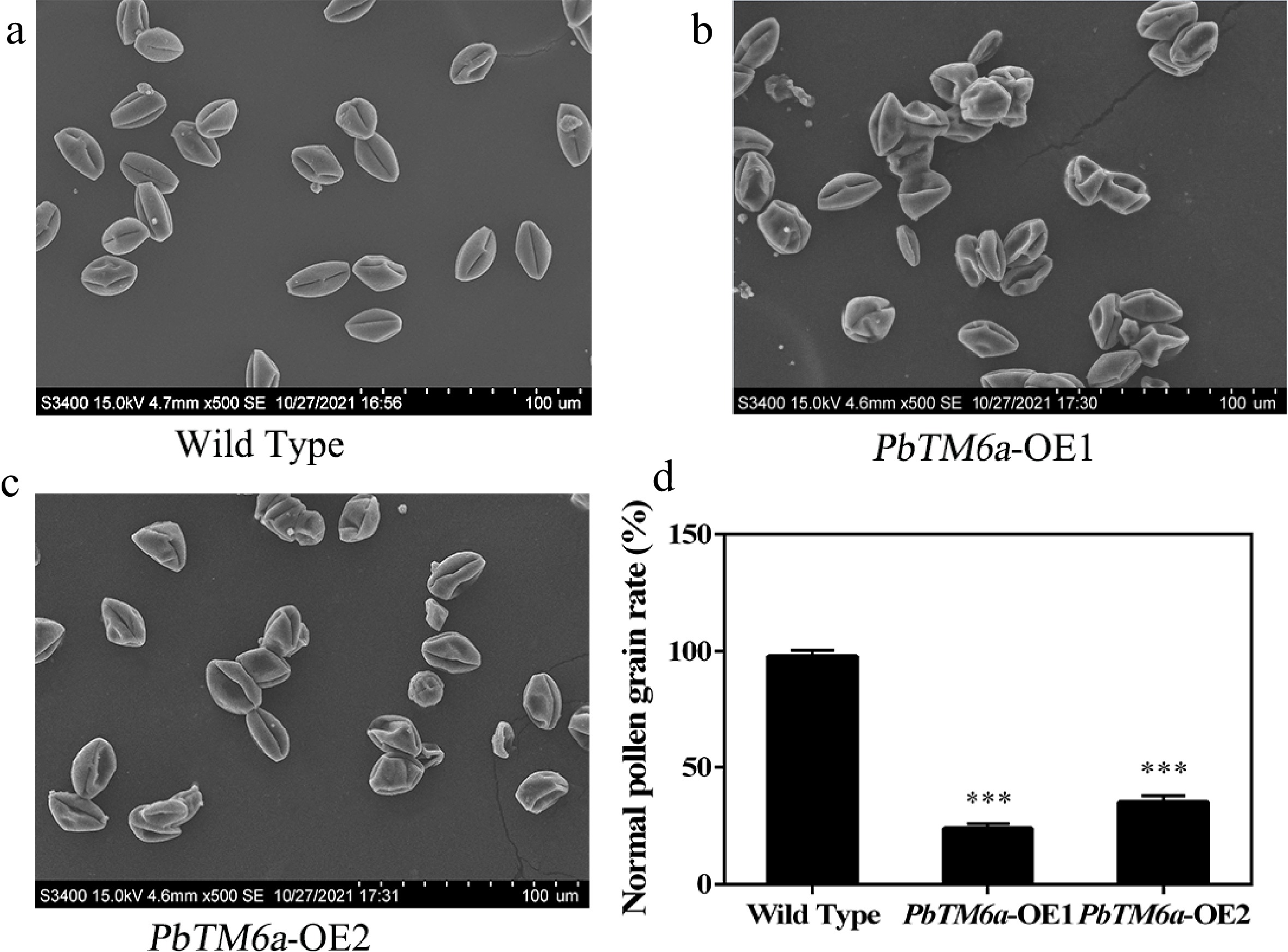
Figure 7.
Analysis of the morphology of wild-type and PbTM6a-OE pollen by scanning electron microscopy. (a) Overview of plump pollen grains of wild type. (b) Overview of plump pollen grains of PbTM6a-OE1. (c) Overview of plump pollen grains of PbTM6a-OE2. (d) Statistics of the percentage of pollen grain with normal surface in wild-type and PbTM6a-OE lines. Data represents mean (± standard deviation (SD) and asterisks represents significant differences (P < 0.01) as determined by one-way ANOVA.
Levels of JA, ABA, IAA, and GA3 were reduced in transgenic stamens
-
Plant hormones were tightly related to pollen development. Therefore, we assessed plant hormone levels in transgenic tomato stamens overexpressing PbTM6a (Fig. 8). Remarkably, the content of JA and ABA were reduced in the stamens of PbTM6a-OE lines, and the levels of IAA and GA3 were significantly decreased. Thus, we concluded that overexpression of PbTM6a in tomato led to the suppression of endogenous JA, ABA, IAA and GA3 levels in stamens, which may be related to male sterility.
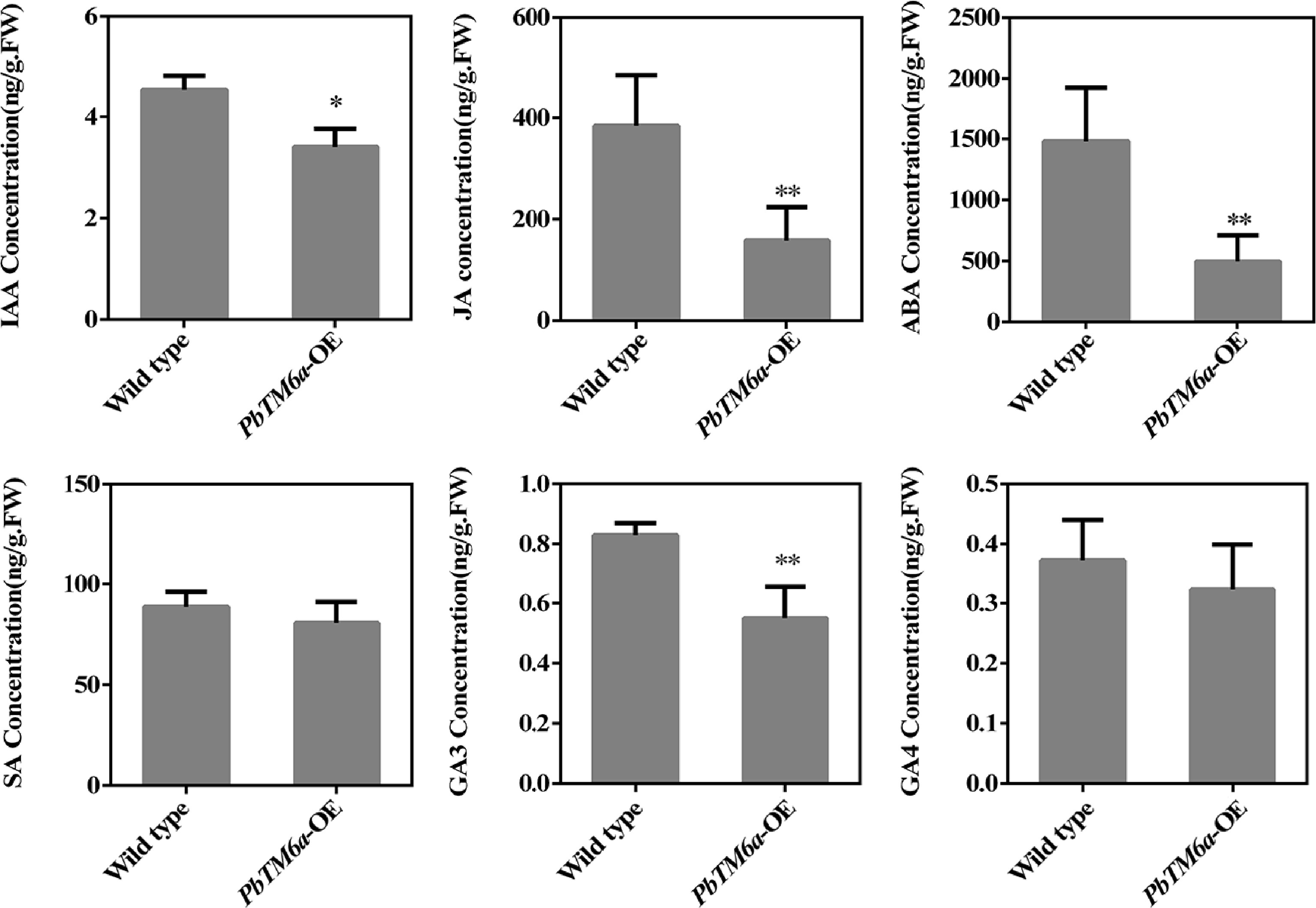
Figure 8.
Levels of IAA, JA, SA, ABA, SA, GA3, and GA4 in wild-type and PbTM6a-OE stamens. Data represents mean (± standard deviation (SD). Asterisks represents significant differences (P < 0.05) as determined by one-way ANOVA.
-
Although class-B MADS box genes, especially for PI and AP3, exhibit a highly conserved function in the formation of petals and stamens[22,25,27], mutations in these genes have been reported to cause distinct new phenotypes including parthenocarpy, male sterility[25,26,27]. Orthologous to AP3, TM6, plays a key role in anther development in strawberry[31] and tomato[26]. However, the role of TM6 gene in pollen and seed development remains unknown, and research related to TM6 genes has not yet been reported in pear. Here we identified PbTM6a and PbTM6b genes from the transcriptome of parthenocarpic ovaries induced by GA4+7, and their expression was suppressed (Supplemental Fig. S1). Here, we reported the response of PbTM6 genes to GA4+7 in pear.
Based on phylogenetic analysis of typical members of the ABCDE model[7], PbTM6a and PbTM6b were classified into class B of the MADS-box gene family (Fig. 1). Like other MADS-box proteins, which contain a highly conserved DNA-binding MADS-box domain of 56–60 amino acids at the N-terminus[3]. The PbTM6a and PbTM6b proteins were also found to possess the conserved DNA-binding MADS-box domain (Fig. 2a). In view of the parthenocarpic capacity of MdPI[22,27], repressed expression of PbTM6a and PbTM6b induced by the GA4+7 treatment impelled us to determine the involvement of PbTM6 in floral organ identity. Our results showing higher expression of PbTM6 in anther, filament and petal were consistent with previous reports, according to which class-B MADS-box genes are highly expressed in stamen and petal[9,25]. The PbTM6a and PbTM6b genes are homoeologues which arose through whole genome duplication. PbTM6a had higher abundance and more significantly decreased expression in parthenocarpic ovaries than PbTM6b[29], PbTM6a was chosen for further analysis.
Because pear has a long breeding cycle and is difficult to transform, tomato was chosen as the heterologous expression system for exploring the role of PbTM6a in flowers and fruit development. Mutant in TM6 mediated by CRISPR/Cas9 system in the octoploid cultivated strawberry leads to distinct petals with modest defects in overall size and color and anthers with a severe reduction in pollen content and viability[31]. However, overexpression of PbTM6a in tomato did not lead to the conversion of petals and stamens and did not affect the vegetative development of transgenic plants compared with the wild-type (Fig. 3a & Supplemental Fig. S3). Thus, we speculated that PbTM6a has distinctive features compared with classical B-class MADS-box genes such as AP3 and PI.
Surprisingly, the number of seeds per fruit was significantly reduced in PbTM6a-OE lines compared with the wild type (Fig. 3b−d). Through histological observations of floral organs, PbTM6a-OE lines have more strongly formed stomium bound to the connective tissue in the pollen sac and squeezed ovules in a narrow ovary chamber, which seems to contribute to the reduction in seed number and size (Fig. 4). Further reciprocal crosses and the determination of pollen vigor and germination rate demonstrated that the low vigor pollen produced by PbTM6a-OE lines leading to unsuccessful fertilization is the dominant cause for the production of fruits with fewer and smaller seeds (Figs 5−7). Meanwhile the effect of the development of transgenic ovary and ovule itself on the seed formation cannot be excluded in this study. Given that the PbTM6a and PbTM6b have much lower expression in the ovary compared to petals and anthers (Fig. 2b, c), the development of the ovary and ovule seems not to be the dominant factor affecting seed number and size. Consistent with the speculation, FaTM6 is not expressed in carpels, carpels of mutants in FaTM6 shows normal development in octoploid strawberry[31].
Interestingly, SlTM6 has been characterized as a candidate gene for the tomato male sterile-1526 locus, where the promoter and first four exons of the TM6 gene were absent[26]. The RNAi of SlTM6 resulted in flowers with homeotic defects primarily in stamens[25]. It is possible that the heterogenous overexpression of PbTM6a leads to post-transcriptional regulation of other endogenous MADS-box genes in tomato. Mutation in FaTM6 leads to smaller and darker anthers with clear defects in the epidermal cell layer which produce less pollen grain with aberrant and collapsed structure and impaired viability[31]. Here ectopic expression of PbTM6a can produce abnormal pollen with reduced viability and low germination and sunken surface (Figs 5 & 7). Sophisticated regulation of TM6 expression is required for pollen development. It demonstrated that TM6 has a conserved role in the anther and pollen formation.
The integrated regulatory network of JA and other plant hormones participates in male organ development[47]. Decreased levels of JA, IAA, ABA and GA3 were detected in the stamens of PbTM6a overexpression lines (Fig. 8). In Arabidopsis, the deficiency of JA is tightly associated with male organ development, mutations in genes encoding JA biosynthetic enzymes result in failure of filament elongation, delayed anther dehiscence, and unviable pollen[48]. Decreased pollen vigor and germination in PbTM6a-OE lines may be a consequence of the decreased JA level. It has been reported that the exogenous application on undehisced anthers can remove the block in pollen release[49]. The specific mechanism between PbTM6a and genes related to regulating JA content needs to be further demonstrated.
Auxin and its signaling play a key role in male gametogenesis. Two auxin biosynthesis genes, YUC2 and YUC6, exhibit high expression in anther procambium, endothecium, tapetum, tetrads and microspores, and the yuc2yuc6 double mutant is unable to form viable pollen grains[50]. Microsporocyte- or microspore-specific, but not tapetum-specific, overexpression of YUC2 gene in the yuc2yuc6 double mutant is sufficient to rescue abortive pollen formation[51]. Similar to the male-sterile phenotype of the JA mutants, the arf6-2 arf8-3 double mutant produces unviable pollens[49]. Male sterility observed in our study could also be a consequence of insufficient IAA content. The production of auxin in anthers is necessary for the advancement of pollen from the microspore stage to the bicellular stage in both Arabidopsis and tomato[51]. Auxin seems to act upstream of JA in the development of pollen grains.
A gradual elevation in ABA and IAA levels accompanies the formation of microsporocytes and microspores in the male-fertile lines of petunia (Petunia hybrida L.)[52]. Ultrahigh levels of ABA and IAA arise in reproductive cells of male-sterile lines. The addition of ABA enhances the viability and reduces the death of barley Hordeum vulgare L., which suggests that ABA acts as a potential signal of male sterility[53]. Although the involvement of ABA in male sterility lacks direct evidence, accumulating indirect evidence shows that ABA interacts with the sugar signaling pathway[54]. Carbohydrates are necessary for sustaining pollen development[55]. In tomato, silencing of Lycopersicum Invertase 5 (LIN5), which encodes a cell wall invertase, decreases seed number per fruit and reduces pollen germination and viability[56]. The role of ABA in male sterility control needs to be further studied in the future.
In addition, the promotion of bioactive GAs to pollen viability has been determined widely. Bioactive GAs is involved in pollen exine formation and the programmed cell death of tapetal cells in Arabidopsis, rice and tomato[57]. Exogenous GA3 application promotes pollen viability and number under cold conditions in almond (Amygdalus communis L.)[58]. Although the effects of GA4+7, when applied to flowers, on pollen germination in pear were not assessed in this study, the expression of PbTM6a was repressed by the application of GA4+7 in pear, confirming that PbTM6a acts downstream of GA4+7 in pear. The low pollen viability production mediated by PbTM6a maybe takes in the process of the regulation of GAs to pollen development. Overexpression of PbTM6a in tomato showed that low GA level may be the cause of the weak vigor of pollen (Fig. 8).
-
In summary, heterologous overexpression of PbTM6a is insufficient for the classical function of class B MADS-box genes in the identification of floral organs, but instead of reduced plant fertility. The specific mitotic progression and the molecular mechanism underlying the role of PbTM6a in pollen development should be elucidated in the future. The results of this study provide insight into the conserved role of MADS-box genes in controlling male fertility, and enhance our understanding of the mechanism underlying the function of PbTM6 in pear.
This study was supported by the earmarked fund for CARS-28 (China Agriculture Research System).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Haiqi Zhang, Wei Han
- Supplemental Table S1 List of qRT-PCR primers.
- Supplemental Table S2 List of primers cloning PbDEFs.
- Supplemental Fig. S1 Expression levels of PbTM6a and PbTM6a in control and GA4+7-treated ovaries. DAA, Days After Anthesis. Data represents mean (± standard deviation (SD). Asterisk represents significant differences (P< 0.05) as determined by one-way ANOVA.
- Supplemental Fig. S2 Floral organs and PbTM6a subcellular localization. (a) Different floral organs of pear. (b) Subcellular localization of PbTM6a.
- Supplemental Fig. S3 Detection and observations of reproductive and vegetative growth of PbTM6a-OE lines (a) DNA amplify of PbTM6a in transgenic tomato (b) Observations of vegetative growth of wild type and transgenic tomato. (c) Observations of flowers of wild type and transgenic tomato.
- Supplemental Table S3 Seeds numbers of tomato with overexpressing PbTM6a gene in mutual pollination experiment.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang H, Han W, Linghu T, Zhao Z, Wang A, et al. 2023. Overexpression of a pear B-class MADS-box gene in tomato causes male sterility. Fruit Research 3:1 doi: 10.48130/FruRes-2023-0001 

Overexpression of a pear B-class MADS-box gene in tomato causes male sterility
- Received: 29 August 2022
- Accepted: 25 November 2022
- Published online: 17 January 2023
Abstract: B-class MADS-box genes are sufficient for the specification of petals and stamens; however, the role of Tomato MADS-box protein 6 (TM6) in seed formation in pear remains largely unknown. In this study, PbTM6a and PbTM6b, characterized as negative regulators of the response to GA4+7, were identified as classic B-class MADS-box genes. Additionally, both of the genes encoding proteins carried the highly conserved MADS-box domain, and showed high expression levels in anther, petal and filament of 'Dangshansu'. Overexpression of PbTM6a in tomato reduced the number of seeds per fruit. Analysis of the anatomical structure of floral organs revealed that the reduction in seed number in transgenic fruits might be attributed to an obstacle of pollen release due to strongly formed stomium and limited ovary space of ovule development. Moreover, triphenyl tetrazolium chloride (TTC) staining and in vitro germination tests of pollen grains indicated that PbTM6a overexpression reduced pollen viability and germination rates. Reciprocal crosses showed that the reduction in seed number in transgenic fruits was dominantly caused by the decreased fertility of pollen grains. Subsequent scanning electron microscopy showed that sterile pollen grains were caused by abnormal pollen grains. Additionally, the reduced levels of jasmonic acid (JA), abscisic acid (ABA), indole-3-acetic acid (IAA) and gibberellin A3 (GA3) in transgenic stamens contributed to the development of sterile pollen. Collectively, our results reveal the role of PbTM6a genes in controlling male infertility and broaden our understanding of the mechanism underlying the function of B-class MADS-box genes in pear.
-
Key words:
- PbTM6 /
- MADS-Box /
- Male fertility /
- Pear