








-
MCM1, AGAMOUS, DEFICIENS and SRF (MADS)-box transcription factors (TFs) are well-known for their regulatory roles in plant growth and development processes[1]. MADS-box proteins have four typical motifs from the N-terminal to C-terminal, including M, I, K, and C. The M motif contains 58–60 amino acids and is highly conserved in all MADS-box proteins, conferring the functions of protein dimerization and nuclear location[2]. The I domain contains almost 30 amino acids and is less conserved, conferring the functions of DNA-specific integration and dimerization[3]. The K domain contains almost 70 amino acids and has the functions of isomerization and protein-protein interaction[4]. The C domain is versatile in sequence length and contains hydrophobic amino acids, has the function of transcriptional activation, and contains more complex protein compounds[5].
MADS-box TFs implement their regulatory roles by interacting with homologous or heterogenous proteins[1]. A MADS-box transcription factor RIN binds with 241 proteins to play key regulatory roles in tomato[6]. MuMADS1 (also MaMADS36) is an AGAMOUS (AG)-like MADS-box gene previously isolated from banana fruits, whose expression is greatly induced by exogenous ethylene and closely related to banana fruit ripening at the transcription level[7]. Recently, we found that two MADS-box transcription factors MaMADS36 and MaMADS55 coordinately regulated banana fruit starch degradation mediated by MaGWD1 and MaMADS36 play a pivotal role in regulating banana fruit ripening[8, 9].
The Ovate Family Proteins (OFPs) are plant-specific and shape regulators by controlling subcellular localization of protein complexes in Arabidopsis[10, 11], and many crops including tomato[12], pepper[13] and cotton[14]. We first isolated an ovate family protein named MaOFP1 from banana fruits and demonstrated that this protein interacted with the K-domain of MuMADS1 to regulate fruit ripening and quality formation[15, 16]. However, the protein expression characteristics of MuMADS1 and MaOFP1 in banana fruits and their fine interaction mechanism remain unclear.
In the present study, we found that MuMADS1 and MaOFP1 proteins differentially expressed during banana fruit ripening process and the K-domain of MuMADS1 interacted with the ovate-domain of MaOFP1. This result sheds light on further understanding the regulatory mechanisms of MADS-box and OFP transcription factors and provided target genes for oriented biological breeding.
-
Differentially developing Baxi (BX, AAA genotype) and Fenjiao (FJ, ABB genotype) fruits including 0 day after flower (DAF), 20 DAF, and 80 DAF (0 day postharvest, DPH) were obtained from the Institute of Tropical Bioscience and Biotechnology banana plantation. The healthy middle fruit combs were selected and separated into single fruit fingers. After surface-sterilization of the fruit fingers using 0.1% sodium hypochlorite for 10 min, they were air-dried and then stored at 22 °C and 80% relative humidity. The 8 DPH and 14 DPH fruits for BX and 3 DPH and 6 DPH fruits for FJ were obtained according to the ethylene production. The selected materials were frozen in liquid nitrogen and stored at −80 °C before extraction of protein at different stages of ripening.
Prokaryotic protein expression
-
MuMADS1 and MaOFP1 were cloned into pCZN1 vector and transformed into E. coli Arctic Express. The recombinants were identified by sequencing. The recombinants were cultured overnight and diluted 1:100 with fresh Luria–Bertani (LB) medium supplemented with kanamycin (50 mg·L−1) at 37 °C. Isopropyl-b-d-thiogalactopyranoside (IPTG; 0.4μM) was used as an additive for 3 h to induce the protein expression. The E. coli cells were collected, broken, and purified. The target proteins were separated using 12% SDS–PAGE and stained with Coomassie blue.
Western blot
-
Equaling banana fruit protein from different ripening stages were separated using SDS–PAGE and lectrotransferred to a PVDF membrane (Millipore Corporation, Bedford, MA, USA) at 100 V for 60 min. The membrane was soaked in 5% non-fat milk in a TTBS solution [0.2 M TRIS-HCl (pH 7.6), 1.37 M NaCl, 0.1% Tween-20] for 1 h at 37 °C. The proteins were incubated with the MuMADS1 and MaOFP1 polyclonal antibodies in 5% non-fat milk in a TTBS solution for 3 h and four 5 min rinses at 37 °C. This membrane was then immersed with a horseradish peroxidase-conjugated goat antirabbit antibody (Zoonbio biotechnology,
www.zoonbio.com/ ) for 1 h at 37 °C, and subjected to four 5 min rinses in a TTBS solution. The blot was performed with a SuperECL Plus kit (Applygen, Beijing, China), and the signal was exposed with X-ray film.Pull down assay
-
Aimed at the GST pull-down identification, full-length cDNA of MuMADS1 was cloned into the pGEX-4T-1 vector, whereas that of MaOFP1 was cloned into the pET-32a vector. All recombinant proteins were subjected to perform GST pull-down as described by Hu et al.[17]. Approximately 100 µg of GST and MuMADS1 fusion protein was immobilized in 50 µL of glutathione agarose and equilibrated. Approximately 100 µg of His-MaOFP1 fusion protein was added to the immobilized GST-MuMADS1 and GST after three washes with PBST. The two fusion proteins were incubated overnight at 4 °C under gentle rotation. The bound proteins were eluted with elution buffer (10 mM glutathione in PBS, pH 8.0) and analyzed by immunoblotting.
Yeast two hybrid (Y2H) assay
-
Y2H assay was performed using a MATCHMAKERTM GAL4 Y2H 3 according to the instructions (Clontech,
www.clontech.com/ ). The K domain fragment of MuMADS1 was cloned into the pGBKT7 bait vector and then immunodetected. Meanwhile, the ovate domain and non-ovate domain fragments of MaOFP1 were amplified and cloned into the pGADT7 to fuse with the activation domain (AD) of GAL4, respectively. Self-activation was detected on selective synthetic dropout medium plates (SD/-Trp, SD/-Trp/-His, SD/-Trp/-His+X-α-gal). Then, combinations were simultaneously transformed into the yeast strain AH109. The transformants containing plasmids pGADT7 and pGBKT7 were used as a negative control. The interactions were evaluated by growth on selective medium (SD/-Ade/-His/-Leu/-Trp+x-α-gal).Bimolecular Fluorescence Complementation (BiFC) assay
-
The K domain of MuMADS1 cDNA fragment was fused into pXY106 containing the N-terminal fragment of the yellow fluorescent protein (nYFP), and an ovate domain fragment of MaOFP1 was fused into pXY104 containing the C-terminal fragment of the YFP (cYFP). The Agrobacteria GV3101 cells containing two plasmids, pXY106-K domain and pXY104-ovate domain, were co-transformed the three week old tobacco (Nicotiana benthamiana) leaves by transiently infiltrating according to the method of Schweiger & Schwenkert[18]. After infiltration, plants were cultivated at 22 °C with 16 h photoperiod for at least 2 d before detection. The fluorescent photographs of YFP were shotted with a Zeiss Axioskop 2 Plus fluorescence microscope (Axio Scope, Germany). All assay results were repeated at least three times.
-
Compared with the MaACTIN, MuMADS1 was differentially expressed during BX and FJ fruits development and ripening process. With the progress of BX fruit development, the expression level gradually increased and reached the peak at 0 DPH, and then gradually decreased. MuMADS1 was highly expressed at 0 DAF and then gradually decreased and reached the lowest point at 6 DPH during FJ fruit development and ripening process (Fig. 1). These results indicated that MuMADS1 played an important role in fruit development and early fruit ripening stages. The expression level of MaOFP1 during BX and FJ fruits development and ripening process was significantly lower than MuMADS1. Moreover, the expression level of MaOFP1 in FJ was higher than in BX (Fig. 1), which suggested that MaOFP1 plays more important role in FJ than in BX. This result was inconsistent with our previous report that the OFP family possesses functions that are more vital in BX than in FJ[19], which is probably because of the difference between mRNA and protein level. However, this result was consistent with our previous study that the expression characteristics of MuMADS1 and MaOFP1 in transcription level in BX was antagonistic indicating their complicated regulatory mechanism in banana fruit ripening and quality formation[15].

Figure 1.
Differential expression of MuMADS1 and MaOFP1 proteins at banana fruit developing and ripening stages. (a) Phenotype of banana fruit developing and ripening stages. A1−A5 represent 0 DAF, 20 DAF, 80 DAF (0 DPH), 8 DPH and 14 DPH of BX, respectively. (b) Protein expression. B1−B5 represent 0 DAF, 20 DAF, 80 DAF (0 DPH), 3 DPH and 6 DPH of FJ, respectively.
Interaction of MuMADS1 and MaOFP1
-
The interaction of MuMADS1 and MaOFP1 has previously been investigated by Y2H[15]. To further identify whether MuMADS1 really interacted with the MaOFP1 protein, an in vitro GST pull-down assay was conducted with MuMADS1-GST and MaOFP1-His proteins that were expressed and purified from E. coli BL21. The result showed that a GST-tagged MuMADS1, but not GST alone, interacts with the His-tagged MaOFP1 protein (Fig. 2). Once again this result proved the interaction of MuMADS1 with MaOPF1, which deepened the interaction theory of MADS-box with non MADS-box transcription factors such as BELL1[20], MuUBA[21], SEUSS[22] and NF-YB[23].
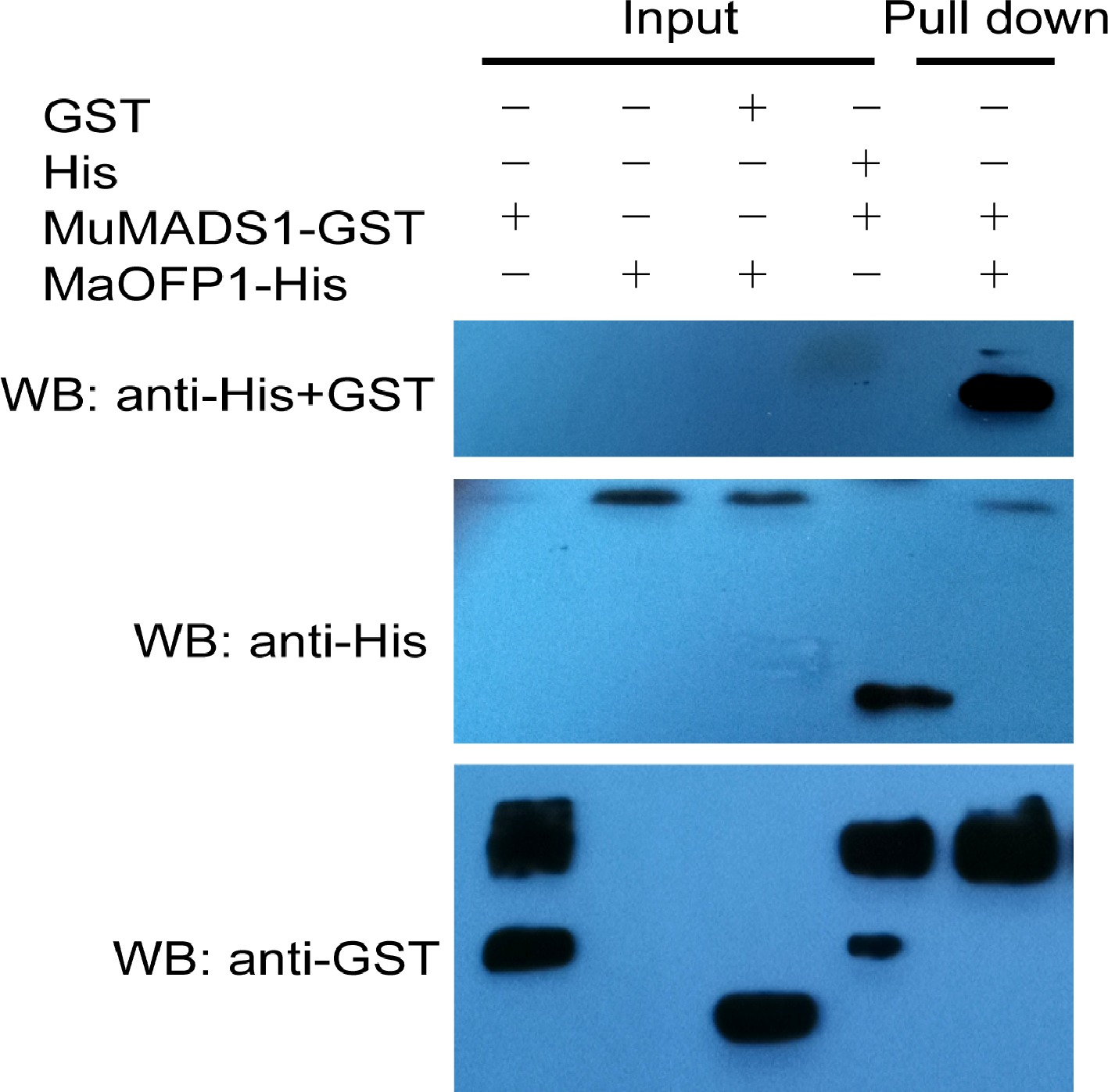
Figure 2.
In vitro GST pull-down assays of the interaction between MuMADS1 and MaOFP1. Anti-His and GST immunoblot shows the amount of MaOFP1-His bound by the indicated MuMADS1-GST protein.
The K-domain of MuMADS1 interacted with the ovate-domain of MaOFP1
-
Previous investigation demonstrated that the K-domain of MuMADS1 interacts with MaOFP1[15]. However, the fine interaction mechanism of K-domain of MuMADS1 with MaOFP1 remains unclear. To further investigate the fine interaction mechanism of the MuMADS1 K-domain with MaOFP1, a Y2H assay was performed. The K-domain of MuMADS1 was cloned into pGADT7, and the ovate domain and nonovate domain were cloned into pGBKT7 (Fig. 3a). The result indicated that the K-domain of MuMADS1 could interact with the ovate domain of MaOFP1 (Fig. 3b).
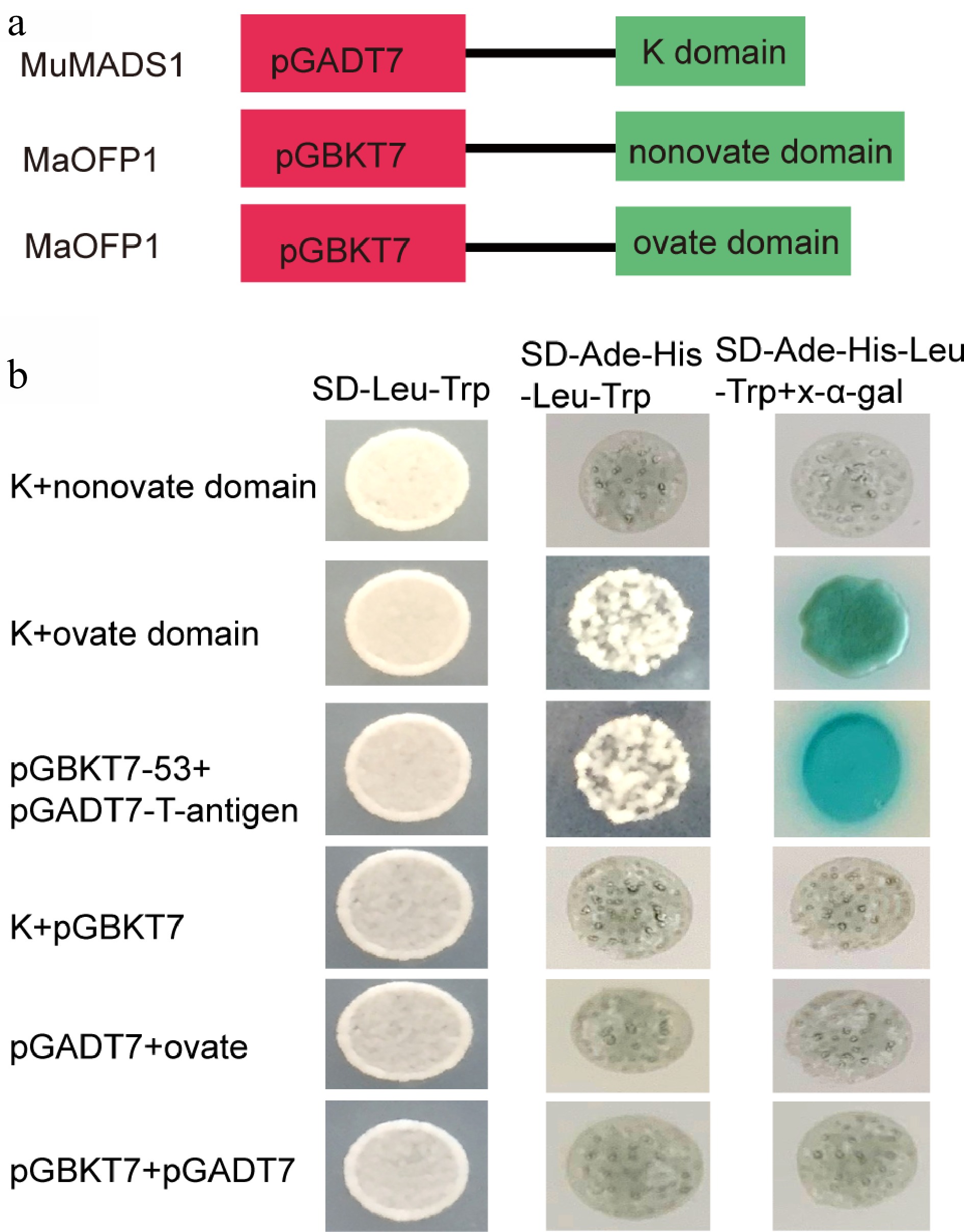
Figure 3.
Interaction analysis of the K-domain of MuMADS1 and MaOFP1 by Y2H. (a) Schematic diagram of vector construction for Y2H. (b) Y2H assay.
The ovate domain of MaOFP1 was designated as the Domain of Unknown Function 623 (DUF623)[12]. The known functions of OFPs are shape regulators, also respond to plant hormones and responses to stress[24]. Previous studies have demonstrated that MuMADS1 plays important roles in banana fruit softening and sugar accumulation[9, 16]. The interaction of K-domain of MuMADS1 with the ovate domain of MaOFP1 suggests the novel roles of OFP in fruit ripening and quality formation, which greatly enriched the function of DUF623.
Identification of the K-domain of MuMADS1 interacted with the ovate-domain of MaOFP1
-
The fine interaction the K-domain of MuMADS1 interacted with the ovate-domain of MaOFP1 was further demonstrated by BiFC assay. The K-domain of MuMADS1 was cloned into pXY106 of nYFP, and the ovate domain and nonovate domain were cloned into pXY104 of cYFP (Fig. 4a). The result indicated that the K-domain of MuMADS1 could interact with the ovate domain of MaOFP1 to produce yellow fluorescence (Fig. 4b), further building on our previous report[15]. This result demonstrated that MADS-box TFs could interact not only with MADS-box proteins but also with non-MADS family proteins, which is highly consistent with Masiero et al.[23] and Liu et al.[21].
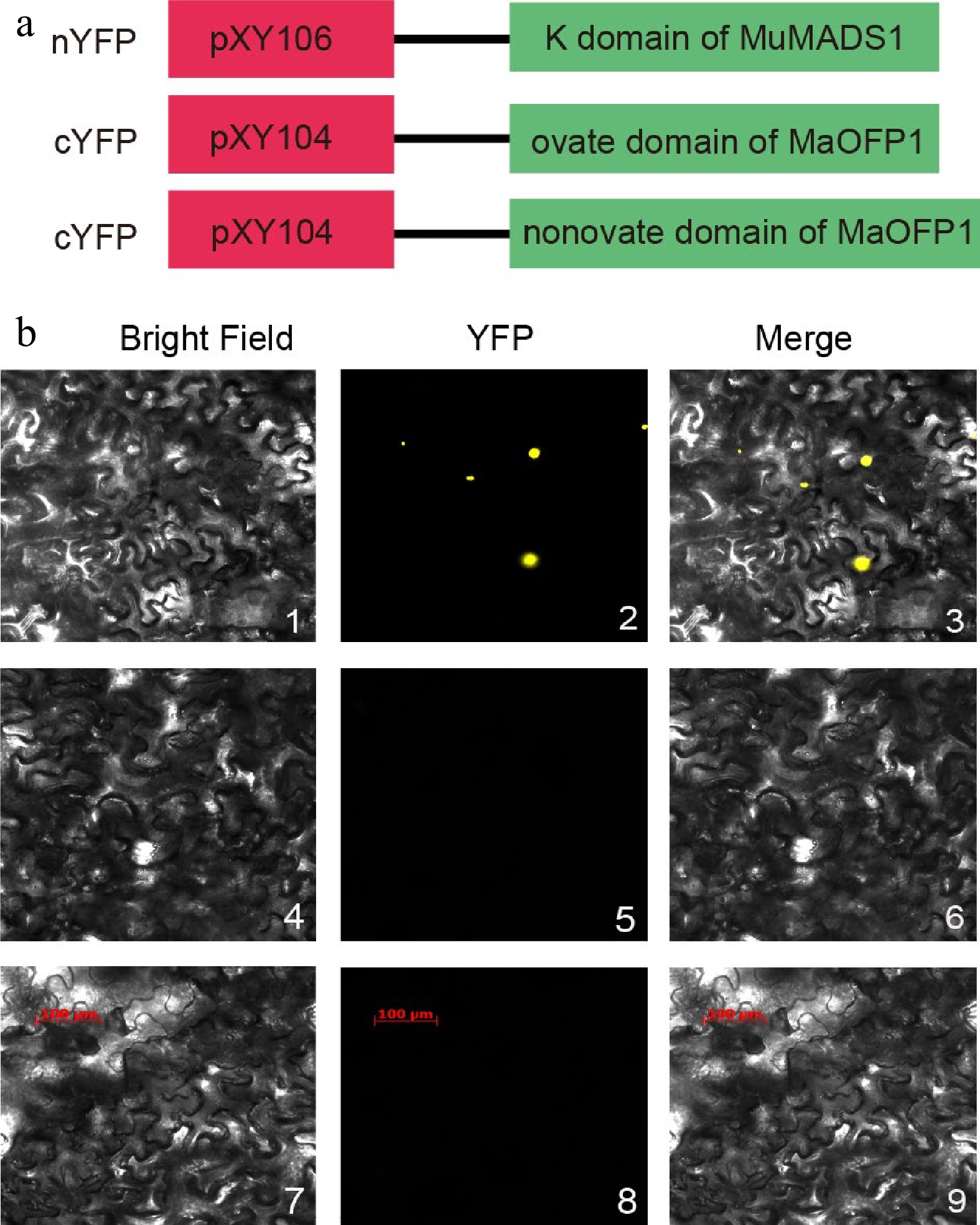
Figure 4.
Interaction identification of the K-domain of MuMADS1 and ovate-domain of MaOFP1 by BiFC. (a) Schematic diagram of vector construction for BiFC. (b) BiFC assay. 1, 2, 3, K-pXY104 + ovate-pXY106; 4, 5, 6, K-pXY104 + nonovate-pXY106; 7, 8, 9, K-pXY104 + pXY106.
-
The expression of MuMADS1 protein was higher than that of MaOFP1 during BX and FJ fruits development and ripening process. The K-domain of MuMADS1 interacted with the ovate-domain of MaOFP1 to implement their regulatory roles.
The work was sponsored by the National Natural Science Foundation of China (NNSFC, No. 31872161 and 32172269), and China Agriculture Research System of MOF and MARA (No. CARS-31-03).
-
Juhua Liu is the Editorial Board member of journal Fruit Research. She was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board members and her research group.
-
# These authors contributed equally: Yunke Zheng, Maoni Fu
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng Y, Fu M, Wang J, Miao H, Jia C, et al. 2023. Protein expression and fine interaction mechanism of banana MuMADS1 and MaOFP1. Fruit Research 3:8 doi: 10.48130/FruRes-2023-0008 

Protein expression and fine interaction mechanism of banana MuMADS1 and MaOFP1
- Received: 08 December 2022
- Accepted: 23 February 2023
- Published online: 03 April 2023
Abstract: MuMADS1 is an AGAMOUS (AG)-like MADS-box gene previously isolated from banana fruits. Previous investigation demonstrated that the K-domain of MuMADS1 interacts with MaOFP1. However, the expression characteristics of MuMADS1 and MaOFP1 proteins in banana fruits and their fine interaction mechanism need to be further investigated. In the present study, the expression characteristics of MuMADS1 and MaOFP1 proteins during banana fruits development and ripening process was investigated by Western blot. The interaction of MuMADS1 and MaOFP1 was demonstrated again by Pull down. The fine interaction mechanism of K-domain of MuMADS1 interacting with the ovate-domain and non-ovate-domain of MaOFP1 was further investigated by yeast two hybrid (Y2H) and bimolecular fluorescence complementation (BiFC) assay. The result indicated that MuMADS1 was differentially expressed during Baxi (BX) and Fenjiao (FJ) fruits development and ripening process. With the progress of banana fruit development, the expression level gradually increased and reached the peak at 0 DPH, and then gradually decreased. The expression level of MaOFP1 during BX and FJ fruits development and ripening process was significantly lower than MuMADS1. Moreover, the expression level of MaOFP1 in FJ was higher than in BX. The K-domain of MuMADS1 interacted with the ovate-domain of MaOFP1. These results laid the foundation for deeply understanding the role of MuMADS1 and MaOFP1 in banana fruits and provide gene resources for banana fruit ripening regulation.
-
Key words:
- Banana (Musa acuminata) /
- MuMADS1 /
- MaOFP1 /
- Protein expression /
- Interaction