










-
Fruit trees are important agricultural resources because of their diversity and adaptability. Due to the growing market for fruits, improving the breeding of high-quality fruit tree varieties and breeding efficiency has become a major focus of research. Traditional fruit tree breeding techniques have some difficulties because of their long juvenile period, high heterozygosity, and incompatibility. PTC is an essential asexual propagation method that can increase propagation coefficients, shortening seedling periods, and ensure the stable inheritance of desirable traits.
The process of PTC consists of continuous dedifferentiation (callus formation) and redifferentiation (plant regeneration) stages[1,2]. Studies have demonstrated that growth regulators in plant are crucial in vitro culture. The callus from Dendrocalamus sinicus hypocotyls was induced on Murashige-Skoog (MS) medium containing 2.10 mg·L−1 6-benzylaminopurine (6-BA) and 3.10 mg·L−1 2,4-dichlorophenoxyacetic acid (2,4-D), and the induction rate was 88.87%[3]. Cnidium officinale root explants showed a maximum callus induction rate of 75% on MS medium containing 2.3 mol·L−1 2,4-D and 2.2 mol·L−1 benzyladenine (BA)[4]. In Salvia tebesana shoot apical meristem explants, the callus induction rate could reach 100% when using 1-naphthylacetic acid (NAA) (0.5 and 1.0 mg·L−1) or in combination with 6-benzylaminopurine (BAP) (0.5 mg·L−1) and 2,4-D (0.5, 1.0, and 1.5 mg·L−1) in conjunction with BAP (0.5 mg·L−1)[5]. PGR are therefore essential for the development of callus.
And epigenetic variations often occur in PTC[6]. A crucial epigenetic modification mode is DNA methylation[7−9], it is also affected during callus induction[10,11]. The relative expression level of DNA methyltransferase gene DNA METHYLTRANSFERASE 1 (MET1), DNA METHYLTRANSFERASE CHROMOMETHYLASE 3 (CMT3) and DOMAIN REARRANGED METHYLASE 2 (DRM2) and REPRESSOR OF SILENCING 1 (ROS1) has also changed during the callus formation[11−13]. The most popular PGR used to produce callus in varied plants are auxin and cytokinin[14−16]. The induction effect was affected by the concentration and combination of PGR, plant species and explant[17]. Previous research discovered that under the induction of 2,4-D, the level of DNA methylation in wheat mature embryos decreased and then increased[10]. The medium containing 1.0 mg·L−1 6-BA and 0.1 mg·L−1 2,4-D, whereas DNA methylation level and the relative expression level of DNA methyltransferase-related genes (MET1, CMT3) were lowest had the highest rate of callus induction (91.0%)[11]. Nevertheless, there are few reports on how growth regulators affect DNA methylation during the formation of pear callus.
Auxin/cytokinin is essential for callus formation, and the transcription level of PGR related genes changes with the formation of callus[18,19]. Some previous studies have found that AHK 2 and AHK 4 encode cytokinin receptors[20−23]; ARR 5 and APRR 5 can be induced by cytokinin[24]; CKX 7 catalyzed cytokinin oxidation[25]. The production of indole-3-acetic acid (IAA) -amino acid conjugates is catalyzed by auxin responsive genes GH 3.1 and GH 3.6, it provides a defense mechanism against excess auxin in plants[26]. Auxin transport genes Pin-like 3, Pin-likes 5, and Pin-likes 6 participate in auxin homeostasis by regulating auxin metabolism[27]. Therefore, in this study we selected these related genes for relative expression verification.
The MSAP and qRT-PCR technologies were applied in this study to examine changes of DNA methylation and gene expressions. Also the gene expression of auxin and cytokinin in pear (Pyrus ussuriensis Maxim) callus induced by TDZ and IBA was investigated. At the same time, the effects of TDZ and IBA on DNA methylation and auxin and cytokinin related gene expression in callus induction of pear leaf were discussed. Our findings shed new light on the plant dedifferentiation DNA methylation mechanism.
-
The test tube pear (Pyrus ussuriensis Maxim) buds were cultured using the MS medium as the basic medium and 1.0 mg·L−1 of 6-benzylaminopurine (6-BA), 0.2 mg·L−1 of 3-indolebutyric acid (IBA), 3% (w/v) sucrose, and 0.6% (w/v) agar[28]. After bringing the pH to 5.8, it was autoclaved for 20 min at 121 °C. The culture temperature was 25 °C, and 16 h was set as the photoperiod. To induce callus, the leaves of pear in vitro cultured for one month were applied as explants. From the test-tube plantlets, only fully developed leaves (those closest to the stem) were selected. Two leaves from each of the 115 plants from each treatment were selected, for a total of 30 leaves from each treatment that were measured. As a technical repeat, the 30 leaves were separated into three groups, each with 10 leaves. The callus induction medium was NN69 medium with different concentration combinations of TDZ (1.0, 3.0, and 5.0 mg·L−1) and IBA (2.0, 4.0, and 6.0 mg·L−1), 30 g·L−1 of sucrose, and 7 g·L−1 of agar. Leaves of pear similar in size and from the same growth period were used. The two ends of the leaves were cut and five wounds were made in the middle of the leaves perpendicular to the main pulse. The explants were then laid flat in callus induction medium and cultured in darkness for 2 weeks. Following 2 weeks of induction, callus was collected for further analysis.
DNA extraction
-
Using a Plant Genomic DNA Kit (Vazyme, Nanjing, China), the genomic DNA of the pear (Pyrus ussuriensis Maxim) was isolated. 1% agarose gels were used to assess DNA quality. −20 °C was used to keep DNA samples.
Reactions of digestion and ligation
-
DNA samples were digested using EcoR I + Hpa II and EcoR I + Msp I. Hpa II
and Msp I recognize and digest the identical site (5′-CCGG-3′), but their sensitivities differ to DNA methylation. Hpa II can recognize and digest non-methylation sites and single-chain methylation sites but cannot digest double-chain methylation sites; that is, it is unable to enzymatically digest sites that contain mCCGG, CmCGG, and mCmCGG. Msp I can recognize and digest non-methylation sites and double-stranded medial cytosine methylation sites and cannot enzymatically digest single-stranded lateral cytosine methylation sites; that is, it cannot enzymatically digest UmCCGG and mCmCGG sites. As a result, several polymorphic PCR amplification bands can represent various DNA methylation states in MSAP analysis. Pre-amplification and selective amplification
-
The fragment length polymorphism program was used to guide two amplification processes[29]. In Supplemental Table S1, the primer combinations needed for amplification are displayed. Supplementary Table S2 displays the primer combinations' base sequences. Pre-amplified PCR reactions had a total volume of 20 µL and contained 2 µL of ligation products and 0.5 µL of E and H preamplification primers (e.g., Supplemental Table S2), Taq polymerase 1 U (Shenggong), 4 µmol·L−1 of dNTPs (Shenggong), and 2 µL of 10× PCR buffer (Shenggong). 26 cycles of 30 s each at 94 °C, 1 min at 56 °C, and 1 min at 72 °C were used to amplify the DNA fragments. The pre-amplified product was then diluted with ddH2O for selective amplification. 0.5 µL of E and H selective amplification primers were added into the diluted product (e.g., Supplemental Table S2), Taq polymerase 1 U, 4 µmol·L−1 of dNTPs, and 2 µL of 10× PCR buffer supplemented with ddH2O to 20 µL. The PCR conditions used were 13 cycles at 94 °C for 30 s, 65 °C for 30 s (reduced by 0.7 °C per cycle), 72 °C for 1 min, and 23 cycles at 94 °C for 30 s, 56 °C for 30 s, and 72 °C for 1 min.
Polyacrylamide gel electrophoresis and data analysis
-
The products of PCR were buffered and denatured for 10 min at 94 °C. Vertical electrophoresis analysis was performed with 6 % denaturing polyacrylamide gel. After silver staining, only the clear and repeatable bands were further converted to binary character matrices, where '+' denotes the presence of bands and '−' denotes no bands. The four types of identified bands are Type I (+, +), Type II (+, −), Type III (−, +), and Type IV (−, −) (e.g., Supplemental Table S3). Type I, no methylation in both EcoR I/Hpa II and EcoR I/Msp I digested; Type II, methylation appeared only in EcoR I/Hpa II degested; Type III, methylation appeared in EcoR I/Msp I digested; Type IV, methylation appeared in EcoR I/Hpa II and EcoR I/Msp I digested. The calculation formula of DNA methylation rate is as follows: Total amplified products = I + II + III; Total methylated products = II + III + IV; Per-methylated ratio (%) = [(III +IV)/(I + II + III + IV)] × 100; Hemi-methylated ratio (%) = [II/(I + II + III + IV)] × 100.
To identify variations in methylation during callus induction, non-methylated and methylated fragments were categorized in accordance with MSAP fragments. Site alterations in methylation status came in four different types: type A, maintaining methylation; type B and C, demethylation; type D, remethylation, in which type B, C, and D are polymorphic methylation bands. Type A is to maintain methylation type, including A1, A2, A3; Type B including B1, B2, B3, B4, B5, B6, B7, B8, is demethylated; Type C includes C1, C2, C3, C4 and C5, is also demethylated; D type includes D1, D2, D3, D4, D5 and D6, which are remethylation type (Supplemental Fig. S1). Type B, type C and type D are polymorphic methylation bands. The percentages of methylation patterns were defined as A(%) = 100 × A / (A + B + C + D); B(%) = 100 × B / (A + B + C + D); C(%) = 100 × C / (A + B + C + D); D(%)= 100 × D / (A + B + C + D).
Quantitative Real-time PCR Analysis
-
Using an EASY Spin Plant RNA Kit (TianGen, Beijing, China), total RNA from callus was obtained, and A NanoDrop 2000C equipment (Thermo Fisher Scientific, Waltham, MA, USA) was used to assess the purity of the RNA. M-MLV reverse transcriptase was used to synthesize cDNA from 1 g of total RNA (Vazyme, Nanjing, China). For qRT-PCR amplification, the following conditions were used: 5 minutes at 95 °C, 45 cycles at 60 °C, 15 seconds at 95 °C, and 30 seconds at 72 °C. The FastStart Essential DNA Green Master Kit was used in the standard mode with the Roche 480 real-time PCR system (Basil, Switzerland). The total reaction system was 20 µL including 2 µL cDNA in 10-fold dilution. The Pyrus actin gene was used as a reference to normalize the qRT-PCR data, and the method via 2−ΔΔCᴛ was used to calculate the relative gene expression level[30]. Supplemental Table S4 lists the primers used.
-
Twenty pairs of primers used amplified a total of 2,534 clearly recognizable bands, including 838 methylation sites, accounting for 33.07% of all amplified bands. When TDZ was 3.0 mg·L−1 and IBA was 4.0 mg·L−1, the induction rate was 54% (Fig. 1 and Table 1), and the methylation rate of callus was 27.96% (Table 2). The callus induction rate was 24.00% and 11.33%, respectively, when TDZ was 3.0 mg·L−1 and the IBA concentration was either 2.0 or 6.0 mg·L−1 (Fig. 1 and Table 1), and the methylation rate was 29.91% and 26.37%, respectively (Table 2). Callus induction rates were as low as 14.67% and 4.67%, respectively when IBA concentration was 4.0 mg·L−1 and TDZ concentration was either 1.0 mg·L−1 or 5.0 mg·L−1 (Fig. 1 and Table 1), and the callus methylation rate was 32.92% and 31.15%, respectively (Table 2). As the IBA concentration increase, the methylation level decreased. As the TDZ concentration increased, the methylation level first fell and subsequently rose. After the medium was supplemented with 3.0 mg·L−1 TDZ and 4.0 mg·L−1 IBA, the callus' methylation rate was 27.96%.
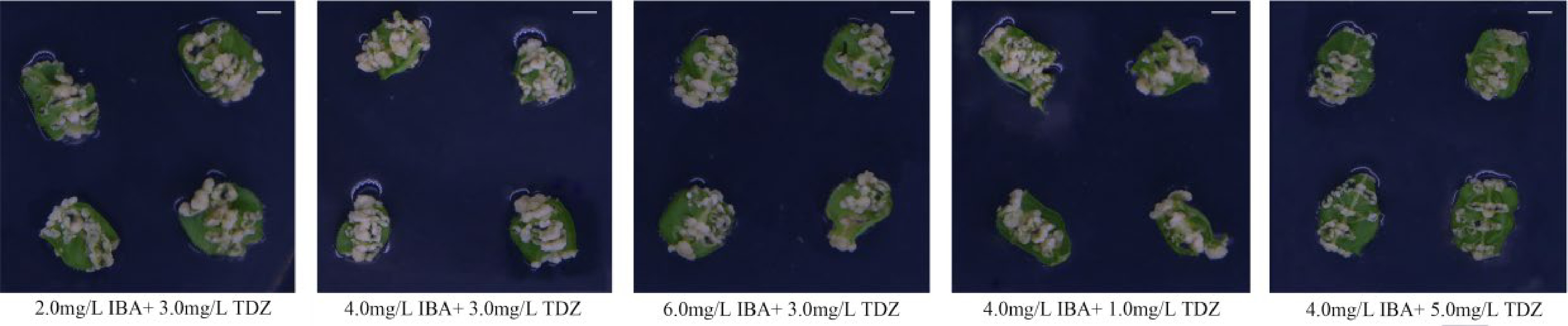
Figure 1.
Comparison of pear callus induced by different TDZ and IBA concentrations. Scale bars = 0.5 cm.
Table 1. Effects of TDZ and IBA on the differentiation of pear leaf explants.
PGR Combination No. of total
explantsNo. of
callusInduction
rate (%)2.0 mg·L−1 IBA 3.0 mg·L−1 TDZ 50 13 24.00 ± 3.06b 50 14 50 9 4.0 mg·L−1 IBA 50 26 54.00 ± 2.00a 50 26 50 29 6.0 mg·L−1 IBA 50 10 11.33 ± 4.67cd 50 5 50 2 1.0 mg·L−1 TDZ 4.0 mg·L−1 IBA 50 7 14.67 ± 2.91c 50 5 50 10 3.0 mg·L−1 TDZ 50 26 54.00 ± 2.00a 50 26 50 29 5.0 mg·L−1 TDZ 50 2 4.67 ± 1.76d 50 4 50 1 Samples were collected 2 weeks after induction in the dark. Only callus with sizes greater than or equal to 2 mm × 2 mm × 2 mm were counted. All analysis were repeated three times and the mean ± S.E. was calculated. Callus induction rate = number of callus in existence/total number of explants × 100%. Columns with different letters are significantly difference at P < 0.05 by Duncan’s multiple range test. Table 2. Comparison of DNA methylation level in pear callus induced by different TDZ and IBA concentrations.
Plant growth regulator TDZ 3.0 mg·L−1 IBA 4.0 mg·L−1 IBA (mg·L−1) TDZ (mg·L−1) MSAP band type 2.0 4.0 6.0 1.0 3.0 5.0 I 396 407 416 379 407 389 II 53 37 31 48 37 49 III 63 72 64 72 72 53 IV 53 49 49 66 49 74 Total amplified bands 512 516 516 499 516 491 Total methylated bands 169 158 144 186 158 176 Methylated ratio (%) 29.91 27.96 26.37 32.92 27.96 31.15 Permethylated ratio (%) 20.53 21.42 20.00 24.42 21.42 22.48 Hemi-methylated ratio (%) 9.38 6.55 6.37 8.50 6.55 8.67 Analysis of DNA methylation patterns in pear callus induced by TDZ and IBA
-
Methylation was mainly maintained in each treatment, both methylation and demethylation frequently happened simultaneously. Three percent of fragments were demethylated when the TDZ concentration was between 1.0 mg·L−1 and 3.0 mg·L−1, and 2.36% of fragments were demethylated when TDZ was between 3.0 mg·L−1 and 5.0 mg·L−1 (Table 3). 5.88% of fragments were demethylated when the IBA concentration ranged from 2.0 mg·L−1 to 4.0 mg·L−1, and 1.68% of fragments were demethylated when IBA concentration changed from 4.0 to 6.0 mg·L−1 (Table 3).
Table 3. MSAP pattern of pear callus induced by different TDZ and IBA concentrations.
Type IBA (mg·L−1) No.of band Percentage TDZ (mg·L−1) No. of band Percentage 2.0 4.0 6.0 1.0 3.0 5.0 M H M H M H M H M H M H A1 + + + + + + 364 422 (88.66%) + + + + + + 351 418 (89.51%) A2 + − + − + − 45 + − + − + − 47 A3 − + − + − + 13 − + − + − + 20 B1 − − + + + + 6 28 (5.88%) − − + + + + 0 14 (3.00%) B2 − − + − + − 5 − − + − + − 0 B3 − − + − − − 0 − − + − − − 0 B4 − − − + − + 2 − − − + − + 1 B5 + − − + − + 0 + − − + − + 0 B6 − − − + + + 1 − − − + + + 0 B7 − − + − + + 7 − − + − + + 1 B8 + − + + + + 7 + − + + + + 12 C1 − − − − + + 1 8 (1.68%) − − − − + + 2 11 (2.36%) C2 − − − − − + 3 − − − − − + 5 C3 + − + − + + 1 + − + − + + 1 C4 + − + − − + 0 + − + − − + 1 C5 − − − − + − 3 − − − − + − 2 D1 + + + + − + 5 18 (3.78%) + + + + − + 13 24 (5.14%) D2 + + + + − − 1 + + + + − − 1 D3 + + − − − − 7 + + − − − − 1 D4 + − + − − − 3 + − + − − − 7 D5 + − − − − − 1 + − − − − − 2 D6 − + − + − − 1 − + − + − − 0 Total 476 467 The NCBI pear (Pyrus bretschneideri Rehd) genome (
www.ncbi.nlm.nih.gov/genome/12793 ) was chosen to contain three highly homologous DNA methylation maintenance-related genes (PbMET1, PbCMT3, PbDRM2) and demethylation-related genes (PbROS1), and the relative expression level of methylation and demethylation-related genes in this process were compared by qRT-PCR. This was done in accordance with the protein sequence of DNA methylation-related enzyme According to the findings, when IBA 4.0 mg·L−1, PbMET1, PbCMT3, and PbDRM2 reduced first and subsequently increased when TDZ concentration increased. In callus generated by 1.0 mg·L−1 TDZ, where the methylation level was 32.92%, the expression of genes associated to methylation was highest (Fig. 2a and Table 2). In callus generated by 3.0 mg·L−1 TDZ, the expression of methylation-related genes was the lowest and the level of methylation was the lowest at 27.96% (Fig. 2a and Table 2). The relative expression of demethylation-related genes PbROS1 showed an opposite trend to the changes of DNA methylation-related genes (Fig. 1a). The highest gene expression and lowest methylation levels were seen when the TDZ concentration was 3.0 mg·L−1 (27.96%, Table 2). When IBA concentration changed, the genes PbMET1, PbCMT3, PbDRM2 and PbROS1 showed similar changes (Fig. 2b). The callus generated by 2.0 mg·L−1 IBA had the highest methylation level and the highest expression of genes related to methylation, both of which were 29.91% (Table 2). In the callus generated by 4.0 mg·L−1 IBA, the amount of methylation was low (27.36%), and the expression of genes related to methylation was at its lowest (Table 2). When IBA was 4.0 mg·L−1, PbROS1 showed the highest gene expression and the lowest methylation level (27.96%, Fig. 2b and Table 2).
Figure 2.
Expression analysis of methylation related genes in pear callus induced by different TDZ and IBA concentrations.
Analysis of expression of auxin and cytokinin-related genes in pear callus induced by TDZ and IBA
-
During callus formation, we studied the expression of genes associated to auxin and cytokinin during callus development. Significant alterations in the expression of associated genes were brought about by the treatment of leaves with various amounts of TDZ and IBA (Fig. 3). On the seventh day following treatment, PbGH 3.1-1, PbPin-likes 3 and PbCKX 7 expression was considerably lower in IBA 4.0 mg·L−1 and TDZ 3.0 mg·L−1 treatment group, but PbAHK 4 expression was significantly greater in the IBA 4.0 mg·L−1 and TDZ 1.0 mg·L−1 treatment group and decrease significantly in comparison to the other treatments (Fig. 3a). PbGH 3.1-1 and PbPin-likes 3 expression was lower significantly than that of IBA 6.0 mg·L−1 and TDZ 3.0 mg·L−1 treatments, but significantly up-regulated than that of other treatments seven days after treatment, whereas PbCKX 7 and PbAHK 4 expression was significantly lower than that of other treatments (Fig. 3b).

Figure 3.
Analysis of expression of auxin and cytokinin-related genes in pear callus induced by different TDZ and IBA concentrations.
-
Studies into plant developmental processes[31], gene functions[32], micropropagation[33], and the creation of transgenic plants with properties[34] have all made substantial use of PTC. Much research has revealed that epigenetic alterations take place during the process of PTC[35−40]. DNA methylation, genomic imprinting, nucleolar dominance, maternal impact, gene silencing, transposon activation, and RNA editing have received the majority of epigenetics research attention[41]. One of the most significant epigenetic changes is DNA methylation. In plant genomes, 5-methylcytosine (5mC) is one of the most widely used epigenetic markers in plant genomes[8].
Previous studies have shown that PGR in plants affect callus induction during PTC[4,5,41]. In this study, we examined the dedifferentiation process in tissue culture because this process is important for regeneration, genetic transformation, and the establishment of a transient verification system. The differentiation direction of explants depends on cytokinin and auxin, which is advantageous to callus growth. In PTC, the most common growth regulators are two categories: auxins (NAA, IBA, 2,4-D and so on) and cytokinins (TDZ, 6-BA, BAP, and so on). The greatest callus induction rate was 86% in peanut (Arachis hypogaea L.) embryos grown on MS medium suppled with 5.5 mg·L−1 BAP and 1.5 mg·L−1 NAA[42]. When the kinetin (KT) was 4.65 µmol·L−1 and IBA was 19.60 µmol·L−1 to induce callus, the rate of induction was 97.75%[43]. In this work, we found that TDZ and IBA could affect callus induction. Callus development was greatest and the induction rate was highest (54%) with 3.0 mg·L−1 TDZ and 4.0 mg·L−1 IBA. Previous studies have demonstrated that PGR have a favorable impact on the development of callus in mangosteen stem explants[44]. The highest rate of callus induction was 75% when root explants generated callus on MS medium containing 2.3 µmol·L−1 of 2,4-D and 2.2 µmol·L−1 of BA[4]. On MS medium supplied with varied dosages of 2,4-D, NAA, and BAP, explants generated callus at a rate of up to 100%[5].
In addition, several studies have shown that PGR can cause changes in DNA methylation level during PTC[11,45−47]. Treatment of barley callus with 0.5 μmol·L−1 brassinolide (BR) led to a slight change in the DNA methylation level compared with the control group[47]. DNA methylation in vitro pitaya shoots was sensitive to low concentrations of gibberellin (GA), but the sensitivity of DNA methylation to high concentrations of GA was reduced[45]. Only a few research have looked at how TDZ and IBA affect DNA methylation during plant callus induction. In this process there have also been limited findings on methylation and demethylation. We found that different concentrations of TDZ and IBA during pear callus induction led to differences in DNA methylation. And methylation and demethylation related genes were also different significantly among treatments. When TDZ was 3.0 mg·L−1 and IBA was 4.0 mg·L−1, the level of methylation was low (27.96%) and the callus induction rate was maximum (54%). Meanwhile, the methylation-related genes had the highest relative expression, while that of demethylation-related genes was lowest. On the medium supplied with 0.1 mg·L−1 2,4-D and 1.0 mg·L−1 6-BA, Brassica napus had the lowest level of methylation and the maximum rate of callus induction (91.0%), demonstrating that hormone function and callus differentiation were connected to methylation level[11]. The induction rate and the level of DNA methylation caused by 6-BA and 2, 4-D were compared, and it was discovered that there were substantial variations in both of these parameters[46]. These conclusions are supported by our experimental data.
The relative expression level of DNA methyltransferase and demethylase genes during callus development triggered by various doses of PGR has not yet been extensively studied. The relative expression level of genes involved with DNA methyltransferase (MET1, CMT3) in Brassica napus callus induced by varied doses of PGR were significantly different[11]. At the same time, many studies have shown that MET1 and DRM2 methyltransferases affect the formation of Arabidopsis callus[48 ]. According to a study by Chen et al., DNA methyltransferase genes (CMT3, DRM2) and DNA demethylase genes (ROS1) may control DNA methylation in the Dimocarpus longan[49]. We discovered that the relative levels of expression of DNA methyltransferase genes (PbMET1, PbCMT3, PbDRM2) and DNA demethylase genes (PbROS1) were significantly different in callus induced by different PGR. This was similar with the findings of previous research[11,48,49].
In order to comprehend how auxin and cytokinin-related genes are expressed during induction of pear callus. This study analyzed the expression of auxin response genes (PbGH 3.1-1, PbGH 3.1-1, PbGH 3.6), transport genes (PbPin-likes 3, PbPin-likes 5, PbPin-likes 6), cytokinin response genes (PbARR 6, PbAPRR 5, PbAHK 2, PbAHK 4) and cytokinin degradation genes (PbCKX 7, PbCKX 7-like) during callus formation. IBA and TDZ have an effect on auxin biosynthesis. When using various IBA concentrations, the auxin synthesis gene is considerably up-regulated as the concentration increases. It is worth noting that under the treatment of different dosages of TDZ, the auxin synthesis essential genes PbGH 3.1-1, PbGH 3.1-2 and PbGH 3.6 were significantly up-regulated on the 14 days following with TDZ 3.0 mg·L−1 and IBA 4.0 mg·L−1. PbPin-likes 6 was also considerably down-regulated in terms of transport, demonstrating that auxin is required to participate in the later stage of callus formation. The study of teaplant also confirmed that auxin played a vital role in the formation of callus[50]. The synthesis of cytokinins was more significantly affected by TDZ. Under the treatment of TDZ 3.0 mg·L−1 and IBA 4.0 mg·L−1, PbARR 5 and PbAHK 4 were significantly up-regulated at 14 days after treatment, indicating that the synthesis and metabolism of cytokinins were mainly affected by TDZ concentration and played an vital role in callus formation[51].
In summary, different combinations of TDZ and IBA concentrations have an effect on the pear callus induction and DNA methylation. Methylation and demethylation occurred simultaneously during pear callus induction. TDZ and IBA affected callus formation through transcriptional regulation. Our study sheds new light on the DNA methylation process of plant callus induction.
The authors acknowledge funding provided by the National Key Research and Development Program of China (2018YFD1000100) and the China Agricultural Research System (CARS-29-07).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primer combinations used for MSAP analysis.
- Supplemental Table S2 Adapters and primers used for MSAP analysis.
- Supplemental Table S3 Cytosine methylation patterns and methylation status.
- Supplemental Table S4 Primers for qRT-PCR analysis.
- Supplemental Figure S1 Amplification images of DNA methylation patterns in callus induced by different concentrations of TDZ and IBA.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou F, Zhou S, Cui S, Liu J, Yang Y, et al. 2023. Effects of TDZ and IBA on DNA methylation during callus induction in pear (Pyrus ussuriensis Maxim). Fruit Research 3:7 doi: 10.48130/FruRes-2023-0007 

Effects of TDZ and IBA on DNA methylation during callus induction in pear (Pyrus ussuriensis Maxim)
- Received: 06 October 2022
- Accepted: 20 February 2023
- Published online: 20 March 2023
Abstract: One of the essential epigenetic processes in higher eukaryotes, DNA methylation is essential for maintaining genome integrity and influencing gene expression. There is limited research on how plant growth regulators (PGR) affect DNA methylation during the pear callus formation process. In this study, using methylation-sensitive amplification polymorphism (MSAP), DNA methylation in pear callus caused by various quantities of thidiazuron (TDZ) and indole-3-butyric acid (IBA) was compared. The concentrations of TDZ and IBA could affect the callus induction rate and methylation level. It was discovered that induction rate and methylation were negatively correlated. The rate of pear callus induction was highest (54%) while the medium suppled with 3.0 mg·L−1 TDZ and 4.0 mg·L−1 IBA, although the matching DNA methylation level was lowest (27.96%). Additionally, there were significant difference in the level of genes' expression between different treatments and both methylation and demethylation are regulated by these genes. As for genes related to auxin and cytokinin, gene expression analysis revealed that their levels of expression after different concentrations of TDZ combined with IBA present in differences significantly. Our findings shed light on DNA methylation mechanisms of plant tissue culture (PTC) dedifferentiation.
-
Key words:
- Pear /
- TDZ /
- IBA /
- Callus induction /
- DNA methylation /
- MSAP /
- Epigenetics.