








-
Pinellia ternata (Thunb.) Breit. is a perennial herbaceous plant in the Araceae family. It is found in the wild in most of China, as well as in Japan and South Korea. P. ternata is known in China as 'Banxia', and the first records of P. ternata can be traced back to the 'Shen Nong's Herbal Classic' of the Han Dynasty[1]. In the field of traditional Chinese medicine, the dried Banxia tuber often has anti-depressant, wound-healing, anti-coughing and anti-vomiting functions, and also has anti-fungal, anti-inflammatory, antioxidant, sedative-hypnotic, insecticidal activities[2].
The Pinellia genus ranges in the number of chromosomes and has abundant germplasm resources. Chromosomal haplotypes 2−16 have been reported, and the basic chromosome number (x) varies greatly[3]. The Pinellia genus is generally thought to have a common basic chromosome number of X = 13[3], but not in integer multiples, so there is a possibility of other chromosome cardinalities. Pinellia pedatisecta and Pinellia yaoluopingensis are two species in which diploidy has been identified, and P. yaoluopingensis differs in taxonomic classification from P. ternata only in the presence or absence of beaded buds, making its classification somewhat controversial. The formation of P. ternata bulbils is involved in asexual reproduction, which means that P. ternata may be an asexual organism that evolved through polyploid changes resulting in descending dysploidy. Thus, the X = 13 chromosome set is theorized to be more primitive than the X = 7−9 variation[3,4]. There is unresolved controversy around the classification of P. ternata, and the disagreement between its classification in the Flora Republicae Popularis Sinicae (FRPS) and Flora of China (FOC) is an obstacle to the study of P. ternata. The species classification and germplasm resources are unclear and thus cannot be correlated with drug efficacy, which directly affects the dosage of medicine and its curative effect.
Reviews published in recent years have mainly reviewed pharmacological activities, processing to reduce toxicity, genetic cultivation, etc. Mao & He[5] gave a detailed review of the germplasm resources, genetic diversity, and active ingredients of P. ternata and suggested that molecular identification or primary active ingredients should be used as quality markers to distinguish the panel from counterfeit products. Bai et al.[6] has described the main components and medicinal mechanisms of P. ternata mainly from chemical composition, pharmacology and toxicity reviews. Pharmacological and molecular targets of P. ternata can be analyzed using network pharmacology. Peng et al.[7] was more specific about the treatment of pineal and the mechanism by which the toxicity was reduced. There are many reviews of P. ternata which have been published in recent years, and a number of new ideas and research projects have been proposed. But there is a gap in the field of molecular genetic breeding of P. ternata. Therefore, this is the main focus in this paper.
Meanwhile, bacterial disease and rising temperatures lead to declines in the production of P. ternata. P. ternata is harvested in the summer and autumn, and it is exposed to temperatures above 30 °C, which leads to rapid withering of the P. ternata plant[3,8,9]. High temperature induces a higher expression of heat shock proteins in plants, and a lower expression of other proteins, affecting growth and development. Bacterial diseases are the most important diseases destroying the growth of P. ternata. With the gradual rise in exports, there is an increased number of artificial P. ternata plantations. However, soft rot and other bacterial diseases, which are harmful to plants in hot weather, have significantly reduced the production of P. ternata[10]. Diseases and improper cultivation management will directly affect crop yield and quality, resulting in extreme economic losses.
Some studies on the resources of P. ternata have been reported, but few focused on its germplasm resources, production, cultivation, and functional genes, and the taxonomy of the plant remains a subject of debate. This study, therefore, focuses on these topics and reviews the recent literature on P. ternata to provide direction for future development and practical application of P. ternata in germplasm resources and breeding.
-
P. ternata normally has 2−5 leaves, but occasionally only 1. The petiole is 15−20 cm long, the base is sheathed, and there are 3-5 mm diameter bead buds in or above the sheath or at the base of the leaf blade (apex of the petiole). Beady buds germinate on the parent plant or after falling into the ground. Seedling leaves are ovate-cordate to halberd-shaped, entirely simple, and measure 2−3 by 2−2.5 cm; the leaves of older plants are 3-split, lobed green, pale-backed, oblong-elliptic or lanceolate, acute at both ends and measure 3−10 by 1−3 cm; the lateral lobes are slightly shorter; entire or with inconspicuous shallow undulating concrete, lateral veins 8−10 pairs, fine, reticulated, dense, set veins 2 circles. Inflorescence petioles are 25−30 (−35) cm long, longer than petioles. The tuber globose is 1−2 cm in diameter, with fibrous roots[1]. P. ternata grows below 2500 meters above sea level and is commonly found on grassy slopes, wastelands, corn fields, field edges, or in sparse forests[11] (Fig. 1).
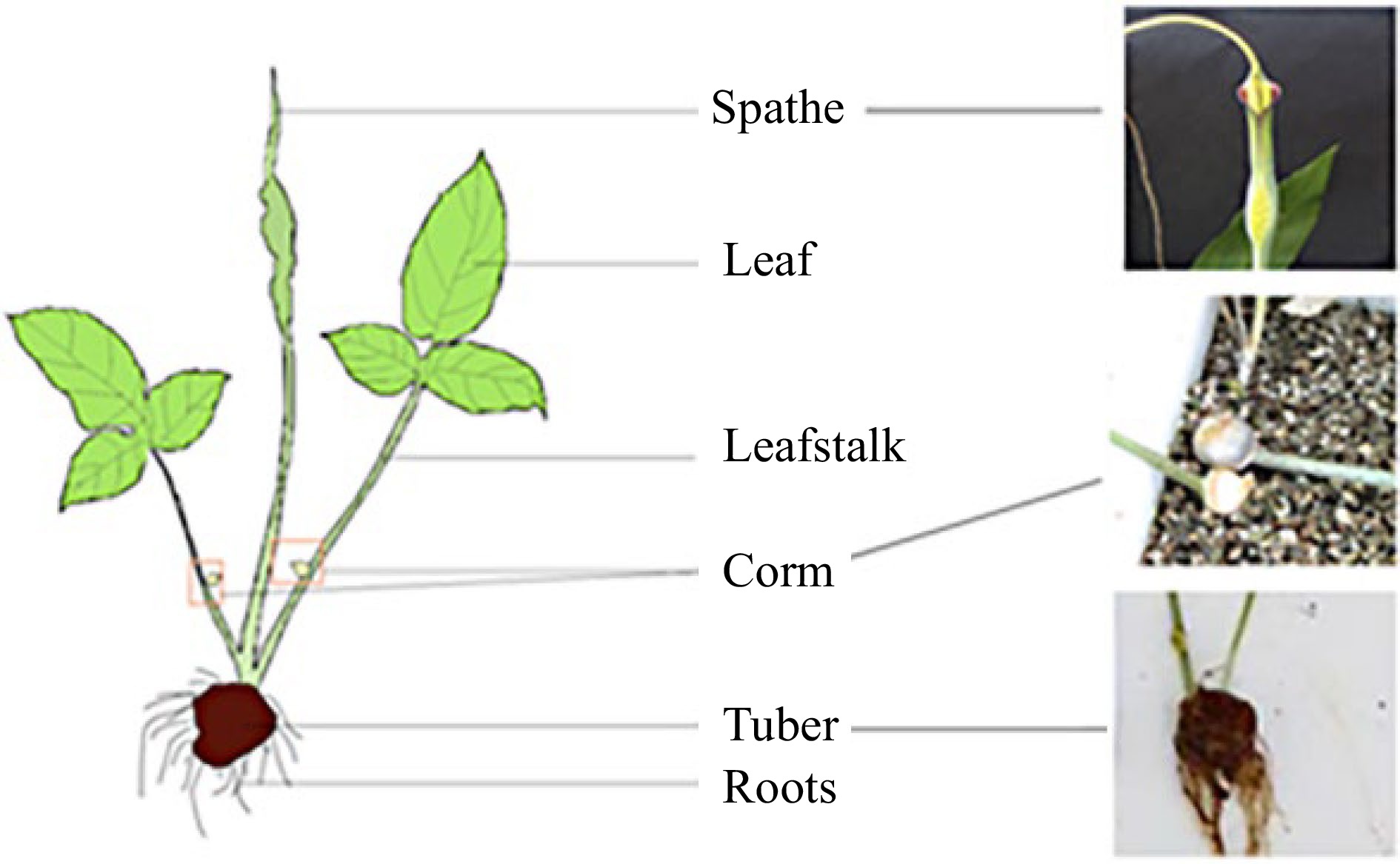
Figure 1.
The biological morphology and organs of P. ternata. The Spathe, corms and tubers are shown in the sketch (left) and the photo (right).
The Pinellia genus is rich in germplasm resources and widely distributed, with five species (Pinellia peltate, Pinellia pedatisecta, Pinellia cordata, Pinellia integrifolia, Pinellia ternata) occurring in China according to FRPS statistics, and nine species (Pinellia peltata, Pinellia pedatisecta, Pinellia cordata, Pinellia integrifolia, Pinellia ternata, Pinellia polyphylla, Pinellia fujianensis, Pinellia yaoluopingensis, Pinellia tripartita) described by 2010 version of FOC. P. peltata, P. pedatisecta, P.cordata, and P. integrifolia are classified clearly (Table 1), but P. ternata is a polyploid heterozygous species which makes its classification controversial[1,12]. For example, P. yaoluopingensis was considered a diploid Pinellia, but the 2010 FOC lists it as a species of the Pinellia genus alone. Mingwang et al.[3] showed that P. yaoluopingensis lacked the bead bud structure of asexual reproduction. In terms of the degree of evolution, it was concluded that P. yaoluopingensis may be the product of aneuploid evolution from the sexual reproduction to the asexual reproduction stage which produced bead buds[3,13], thus retaining its taxonomic status as a species. However, several studies report that the polyploid heterozygous species of P. ternata contains a diploid genome. The number of haploid chromosomes is not unique, including X = 13 and X = 9, two common haploid chromosomes numbers. In addition, the P. ternata leaf shape also changed during the planting process, and the species could not be completely separated by traditional leaf shape classification[3].
Table 1. Species comparison of Pinellia genus between Flora Republicae Popularis Sinicae (FRPS) and Flora of China (FOC).
Latin name of FRPS Latin name of FOC Taxonomic character Place of origin Pinellia peltata Pinellia peltata Leaf blade entire, Leaf blade peltate, ovate or oblong. Fujian, Zhejiang Pinellia pedatisecta Pinellia pedatisecta Leaf blade always pedate, leaflets 6−11; bulbils absent. Sichuan Pinellia cordata Pinellia cordata Tuber globose; leaf blade sagittate-oblong, cordate-ovate, base deeply cordate; bulbils present at base of petiole and at base of leaf blade Anhui, Fujian, Guangdong, Guangxi, Guizhou, Hubei, Hunan, Jiangxi, Zhejiang. Pinellia integrifolia Pinellia integrifolia Leaf blade ovate or oblong, base obtuse or shallowly cordate, 5–19 × 1.5–6 cm Chongqing, Hubei, Sichuan Pinellia ternata Pinellia ternata Leaf blade trisect, sometimes pedate with only 5 leaflets, Bulbils present at petiole below middle, or both at proximal part of petiole and at base of leaf blade Widely distributed in China, excluding inner Mongolia, Qinghai, Xinjiang, and Tibet (Japan, Korea; Europe and North America) Pinellia polyphylla Leaf blade deltoid-ovate or broadly ovate, base deeply cordate, 6–33 × 4–22 cm Sichuan Pinellia fujianensis Rhizome cylindric; leaf blade broadly sagittate; bulbils at base of petiole Fujian Pinellia yaoluopingensis Leaf blade trisect, sometimes pedate with only 5 leaflets, leaflets oblong or lanceolate. Petiole lacking bulbils, bulbils emerging only from tuber; lateral leaflets usually bifid Anhui, Jiangsu Pinellia tripartita Leaf blade only deeply 3-partite, anterior lobe broadly ovate or ovate-oblong, sessile; bulbils absent. Hong Kong (Japan) The classification of species plays an active role in the study of genetic reproduction. A clearer demarcation between species will be the key to studying the genetic evolution and clinical use of P. ternata. Molecular techniques offer considerable improvements and overcome many of the significant challenges of phenotypic testing for a better understanding of Pinellia genus diversity.
-
P. ternata was first recorded in Shaanxi Province (China) during the Wei-Jin period in Shen Nong's Herbal Classic as a traditional medicine slightly used in each prescription[14]. P. ternata plants were later found in the Shandong, Anhui, Jiangsu, and Zhejiang provinces[12]. During the Tang Dynasty, P. ternata was grown in various regions of China, but it was produced at the highest quality in Henan, Anhui, and Jiangsu provinces[15] (Table 2). During the Song, Ming, and Qing dynasties, P. ternata with the highest medicinal value was produced in Shandong Province. In the Republican period, P. ternata from Hubei province was regarded as a genuine medicinal material[16−18]. In China, the origin of P. ternata developed from west to east and then from east to west, and now it is distributed in most areas of the country, except Xinjiang, Tibet, Qinghai, and Inner Mongolia. And some countries in East Asia such as Korea and Japan also have P. ternata production[1]. Changes in the main origin of P. ternata have occurred throughout history, which may be related to climate change. The climate affects the quality, causing changes in the main production area.
Table 2. Record of the main planting area of P. ternata.
Dynasty High quality growing area Reference Wei-Jin period Shaanxi Province Supplementary Records of Famous Physicians Wei-Jin period Shaanxi Province, Shandong Province, Anhui Province, Zhejiang Province, Jiangsu Province,
Shanghai MunicipalityCollective Notes to the Canon of Materia medica Tang dynasty Henan Province, Anhui Province, Jiangsu Province Qianjin Yi Fang Song dynasty Shandong Province Bencao Tujing (Illustrated Classics of Materia Medica) Ming dynasty Shandong Province Qing dynasty Anhui Province, Shandong Province Materia Medica in Qing Dynasty The Republican period Hubei Province Drug production identification -
Species genetic diversity plays an important role in adaptability to natural environments and is closely related to the evolution and genetic variation of population structure[19]. The study of species genetic diversity is instructive for screening the high-quality germplasm resources of P. ternata and its polyploid complex species, as well as the evolutionary relationships among species. To study the genetic diversity of P. ternata, a variety of gene identification methods were used to reveal the geographical distribution of P. ternata in an attempt to predict the evolutionary principles and direction of germplasm resources. Various molecular markers such as SRAP (Sequence-related amplified polymorphism), TRAP (Target region amplified polymorphism), ISSR (Inter-simple sequence repeat), ITS (Internal transcribed spacer), and cpDNA (Chloroplast DNA) have been used to investigate the relationships, germplasm resources, taxonomy, origin, and evolution of Pinellia[20,21]. Genetic diversity analysis of panel germplasm resources based on ISSR markers showed that geographic factors were more important than leaf shape classification[22]. A comprehensive analysis of the Jaccard similarity coefficient of each population of P. ternata showed high similarity among populations with different leaf types, with a lower similarity between different regions, suggesting that the germplasm resource groups of P. ternata, especially the wild type of P. ternata, should be mainly divided into regions and supplemented by phenotypes[23]. RAPD (Random amplifed polymorphic DNA) analyses of different populations have shown that the germplasm resources of P. ternata are highly mixed in both natural and cultivated populations[24]. Some plant individuals share the same phenotype, but genotypes vary according to the composition of genetic material. RAPD and RFLP (Restriction fragment length polymorphism) analyses of P. ternata materials from China and Korea have revealed that genetic diversity within a species can be found in specific regions, and the geographic origin of the tubers of P. ternata can be easily identified[25]. Pan et al. studied the genetic diversity and pedigree structure of wild Pinellia chinensis, revealing the causes of its geographical distribution pattern and speculating on the potential refuge of the species during the ice age. The ITS sequences of Pinellia nuclear genes from 205 individuals were analyzed by PCR (Polymerase chain reaction) amplification and sequencing[26], revealing that Pinellia may have several glaciated sanctuaries in China, which are located in southwest, central, and east China. This is consistent with the hypotheses that Pinellia originated in the middle and lower reaches of the Yangtze River[27] and that Pinellia cell types increased from the southwest to the northeast[28]. Based on the chloroplast genome, an adaptive evolution analysis of the three species of Pinellia revealed that they form a monophyletic evolutionary clade, among which P. pedatisecta diverged first, indicating that P. pedatisecta is not a basal group of Pinellia, while P. peltata may be[29]. This differs from our prior understanding and could help to further understand the genetic diversity of the panel and better screen for high-quality germplasm resources.
Mutual introduction in different regions is the norm in the production, but it is necessary to consider the relationship between germplasm resources and geographical environment. Introduction cannot be blindly introduced and high-quality resources need to be selected according to local conditions, but there is no unified standard in the industry, it is necessary to formulate industrial standards to promote the development of P. ternata production.
-
Genetic breeding contributes to significant improvement in yield and product quality by enhancing tolerance to biological and abiotic stresses. P. ternata is a polyploid complex population with complex haplotypes and multiple chromosome numbers. P. ternata is agreed to be a population with multiple haplotypes 2n = 2x, 4x, 6x, 8x, 9x, and 10x with X = 13, among which the octoploid panel with 2n = 8x has the highest proportion in each population and is the most widely distributed P. ternata population[3,4] The increased vigor of polyploids has made them a goal of many plant breeders over the last century, with increasingly improved plant varieties obtained by inducing polyploids and/or exploiting natural polyploids in a variety of ways[30]. He et al.[31] and Jia et al.[32] reported the successive induction and identification of the 16-ploidy of P. ternata. Colchicine was used to induce the polyploid transformation of P. ternata based on the original 8-ploidy, and a P. ternata polyploid showed better resistance and increased yield. Pathway enrichment analysis of differentially expressed single genes (DEGs) by Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) showed that genes involved in the 'starch and sucrose metabolism', 'purine metabolism', and 'photosynthesis' pathways were significantly induced in the P. ternata polyploid. Six transcription factors (MYB, WRKY, bHLH, lateral organ boundary domain (LBD), homologous domain zip (HD-ZIP), and ethylene response factor (ERF)) may play an important role in phenotypic and medicinal composition differences between 8-ploid and 16-ploid materials of P. ternata and may improve the resistance of 16-ploid strains to biotic and abiotic stresses[33].
Unlike other weeds and normal crop plants, P. ternata has a very low reproductive rate under natural conditions. It mainly relies on its bulbs to reproduce asexually, with a mature plant growing only 7-8 bulbs per year. The rapid propagation technology of P. ternata has been gradually developed and further promotes the differentiation of leaves in vitro. The cultivation of large tuber plants favors the development of large and numerous beaded buds[34]. Xie et al.[35] used tissue culture to study the virus-free technique on the leaves of P. ternata with an 80% virus-free rate, finding that the NAA (α-naphthoacetic acid) project improved the differentiation efficiency of tissue[36]. Xu et al.[37] selected an MS medium as the most suitable medium for P. ternata growth when screening basic media, and studied the hormone ratio of explant disinfection treatment and plant regeneration. Wang et al.[38] reported that low concentrations of 2,4-dichlorophenoxyacetic acid (2,4-D), indole-3-acetic acid (IAA), and NAA were suitable for microtuber induction, while multiple concentrations of kinetin (KT) and zeatin (ZT) had no significant effect on petiole microtuber induction. When different concentrations of auxin or cytokinin were added to the MS medium, the petioles of the explants were mostly polar. Building on previous experiments on in vitro culture of P. ternata, Jie et al.[39] performed suspension cell culture on P. ternata and identified the succinic acid component. There was no significant difference in succinic acid composition relative to cultured P. ternata, but toxicity was lower in the cell suspension. Liu et al.[40] found the alkaloid content in cells cultured in vitro was much higher than in cultivated plants.
Heterosis refers to the phenomenon in which offspring of a cross between two genetically distinct parents outperform their parents in terms of growth potential, viability, fecundity, resistance to stress, yield, and quality[41]. Conventional breeding has produced many hybrids of medicinal plants, such as peony, mint, and rehmannia[42−44]. For the polyploid hybrid species P. ternata, the chromosomal ploidy of the parents should be taken into account in crossbreeding. The ploidy of P. ternata is complex, with two reproductive states: asexual reproduction and sexual reproduction, with sexual reproduction dominated by asexual reproduction. It is necessary to take into account whether the breeding material is capable of sexual reproduction when conducting cross-breeding for P. ternata. Sun et al.[45] pointed out that a possible cause of hybridization failure is difficulty in chromosome pairing due to inconsistency in the number of parental chromosomes, which indicates incompatibility between the two parties. Wang et al.[46] suggested that parental affinity, which may be related to the diversity of chromosome multiplicities of the P. ternata, is the main factor affecting the seed set rate for hybridization. Luo et al.[47] crossbred material from different populations of P. ternata for evaluation and measured the organic acid and guanosine content of the F1 generation. Maternal parentage has a greater influence than paternal parentage on the accumulation of organic acids and guanosine in the F1 generation and the accumulation of organic acids and guanosine in the tuber of P. ternata has a somewhat antagonistic effect[47]. The purpose of crossbreeding was to improve agronomic traits or stress resistance, but the present study only focuses on the effective constituents, and failed to show the advantages of cross breeding. Meanwhile, the evolution of P. ternata tends to reproduce asexually, to increase the reproductive efficiency and promote the adaptation to the environment. This adaptability is closely related to drug efficacy and yield as well as processing. However, it contradicts crossbreeding and is a key issue to be considered in subsequent breeding work.
-
Many functional genes are crucial to the growth of P. ternata (Fig. 2). Transcriptome analysis of P. ternata bulb development confirmed that GAPDH is the best reference gene in the development stage and a variety of tissues of P. ternata, laying the foundation for analyzing the mechanism of bulb formation[48]. Heat stress is an important environmental factor restricting the growth of P. ternata. Spermidine (SPD) and melatonin (MLT) are processed to enhance the heat tolerance of P. ternata under heat stress conditions, likely by regulating heat shock transcription factors with different regulatory modes. These findings suggest that MLT and SPD participate in the short duration of HSR, which is an important clue for the study of improving the heat tolerance of P. ternata[49]. Tian et al.[50] cloned the P. ternata heat shock protein gene PtsHSP17.2 in transgenic tobacco and demonstrated that PtsHSP17.2 is more highly expressed in transgenic plants with heat stress induction. This study enhances the current understanding of thermal adaptation and further strengthens the study of P. ternata's genetic reproduction. Meanwhile, stearic acid desaturase genes (PtSAD) have been confirmed to play an important role in heat stress. High temperature induces the expression of PtSAD, but other stresses (drought, cold, and salt stress) do not. Overexpression of PtSAD increases the fraction of unsaturated fatty acids, and negative feedback regulation reduces the thermal resistance of P. ternata[51]. Bacteria causes many serious diseases that have seriously affected the yield of P. ternata in recent years. Soft rot is a disease caused by P. carotovorum subsp. Carotovorum (Pcc). P. ternata infection, which appears as a water-stained scab, is highly transmissible and leads to tuber rot and decreased yield[52]. pY11T-3-1, as Ps. aeruginosa, has a wide antagonistic spectrum and host selectivity and has an inhibitory effect on Pcc[53]. Biological control is also a new trend against plant diseases in recent years, which also plays an important role in the production of Pinellia. The P. ternata (Thunb.) Breit T2 plus line system began to show symptoms at 20 h after infection with Pcc. During infection, genes related to growth and development were downregulated and genes related to disease resistance were reprogrammed. These results provided the initial inspiration and molecular mechanism for studying the resistance mechanism of soft rot on P. ternata[54].
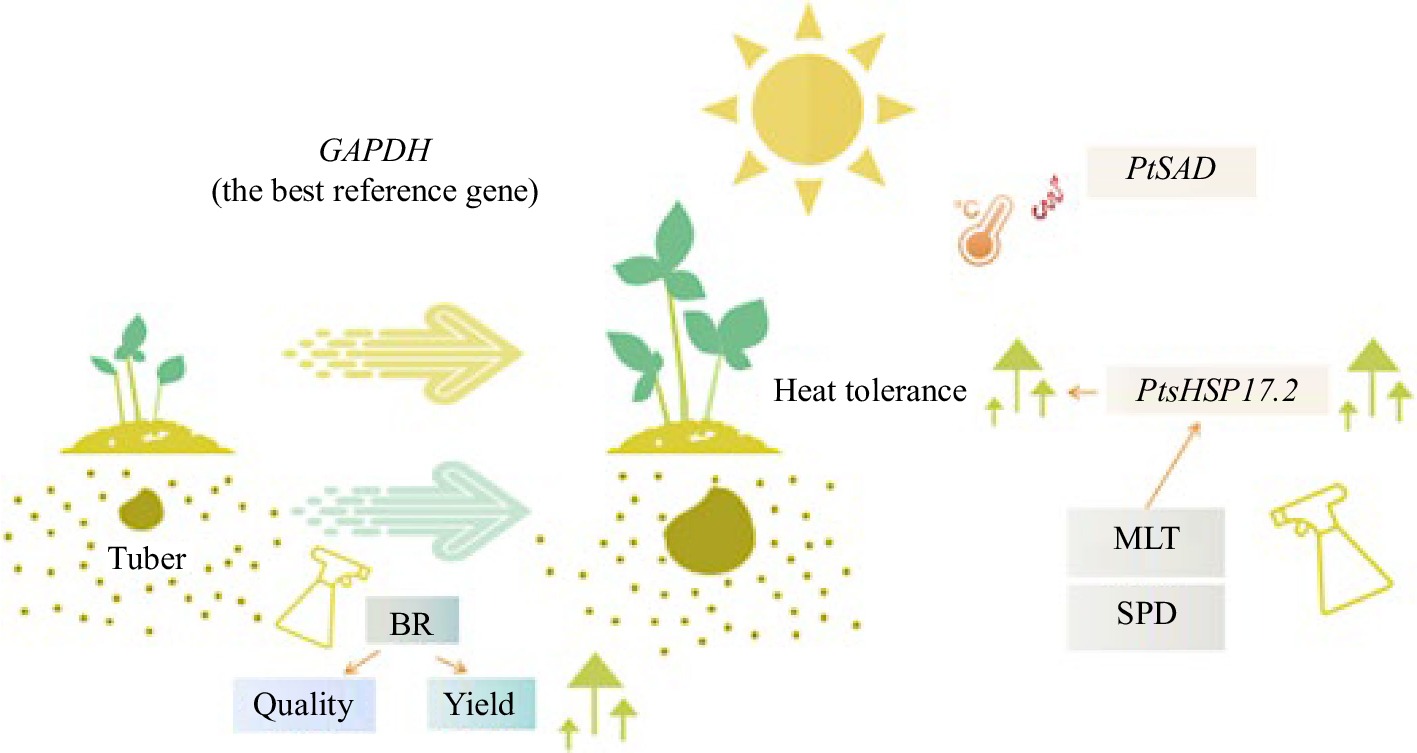
Figure 2.
Studies on functional genes in P. ternata production.
A hormone, Brassinolide (BR), is involved in many important physiological and biochemical regulatory processes, such as plant gene expression, reproductive development, and photosynthesis to regulate plant growth[55,56]. A series of recent experimental studies tested the effect of BR on P. ternata. Guo et al.[57] conducted BR treatment on two varieties of P. ternata, finding that BR improves the quality and yield of P. ternata. The subsequent BR treatment of P. ternata showed that the catabolism of starch to maltodextrin and maltose in the tuber decreased with lower expression of beta-amylase (BAM) and isoamylase (ISA), and the catabolism of cellulose to D-glucose increased with higher expression of endoglucanase (edg) and beta-glucosidase (BGL) genes. The photoprotective mechanism of P. ternata improved and the content of abscisic acid (ABA) decreased, further revealing the effect of brassinolide (BR) on the formation mechanism of the P. ternata tuber[58]. In addition, the optimal harvest date of plants treated with BR was evaluated using two indexes of yield and quality change, revealing 75 days after treatment with 0.10, 0.50, and 1.00 mg/L BR to be the optimal harvest point[59]. In recent years, there have been a number of studies on the in vivo regulation and expression of genes induced by exogenous hormones in order to improve abiotic stress in plants. Pinellia are prone to die above ground and hibernate below ground in hot and dry conditions. This pattern of growth has serious implications for P. ternata cultivation. How to breed and improve excellent P. ternata varieties, and how to improve their agronomic properties and disease resistance are also the focus of current research.
In recent years, the research and application direction of P. ternata is inclined to the anti-insect application of lectin, which is effective and widely used. However, there are few gene mining and functional gene verification of P. ternata itself. Therefore, the follow-up research should focus on the molecular mechanism of P. ternata itself, and the use of gene editing or overexpression to verify the function of functional genes is the focus of future research.
-
P. ternata tends to grow in a warm, moist, shaded environment and can be planted in forests, among fruit trees, or with other crops. Strong light leads to the phenomenon of lodging: the leaves wither, the buds fall, and then new plants grow and develop[60]. According to statistics from the General Administration of Customs (General Administration of Customs of the People's Republic of China.
www.customs.gov.cn ), P. ternata exports reached 169.581145 million yuan in 2022, ranking among China's top ten export medicinal materials exports in 2020[61]. As a traditional Chinese medicinal substance as not a source of food plant, and it is relevant for the treatment of Covid-19. Its export price has been rising rapidly since 2019, up 56.55% (Fig. 3), but the annual export volume is gradually decreasing. Japan was the largest exporter of P. ternata in 2022, accounting for 51% of the total annual exports, followed by Korea with 23% and the Taiwan region of China with 15% (Fig. 4). P. ternata is exported to Asia, North America, and even some countries and regions in Australia. Since the Covid-19 global outbreak, the demand for P. ternata at home and abroad is increasing, wild resources are scarce, and disease outbreak has been frequenting in artificial planting. Therefore, it is urgent to improve the quality and productivity to maintain the sustainable development of P. ternata industry. The export quality standard is very important in the export trade. At the same time, it is necessary to develop export processed products and deep-processed products to support the development of related industries. Industry and development of P. ternata should be closely related to genetic breeding. The development of the cultivation cannot be separated from the collection, exploitation and utilization of resources. The adaptability of the P. ternata should be coordinated with the local climate, and the yield and quality should also be taken into account, which are inseparable from the work of genetic breeding.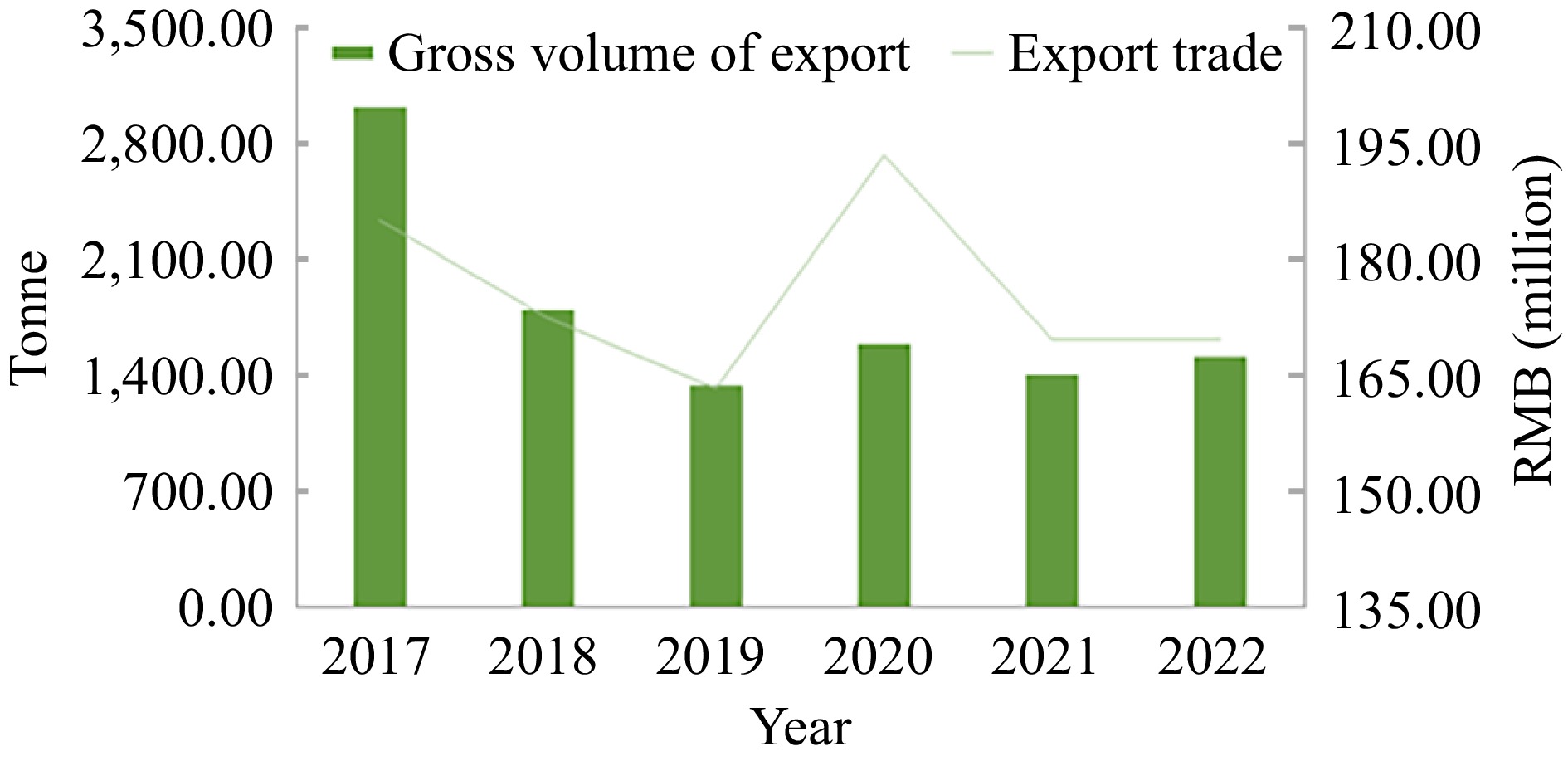
Figure 3.
The volume of P. ternata exports and total export trade over the past six years.
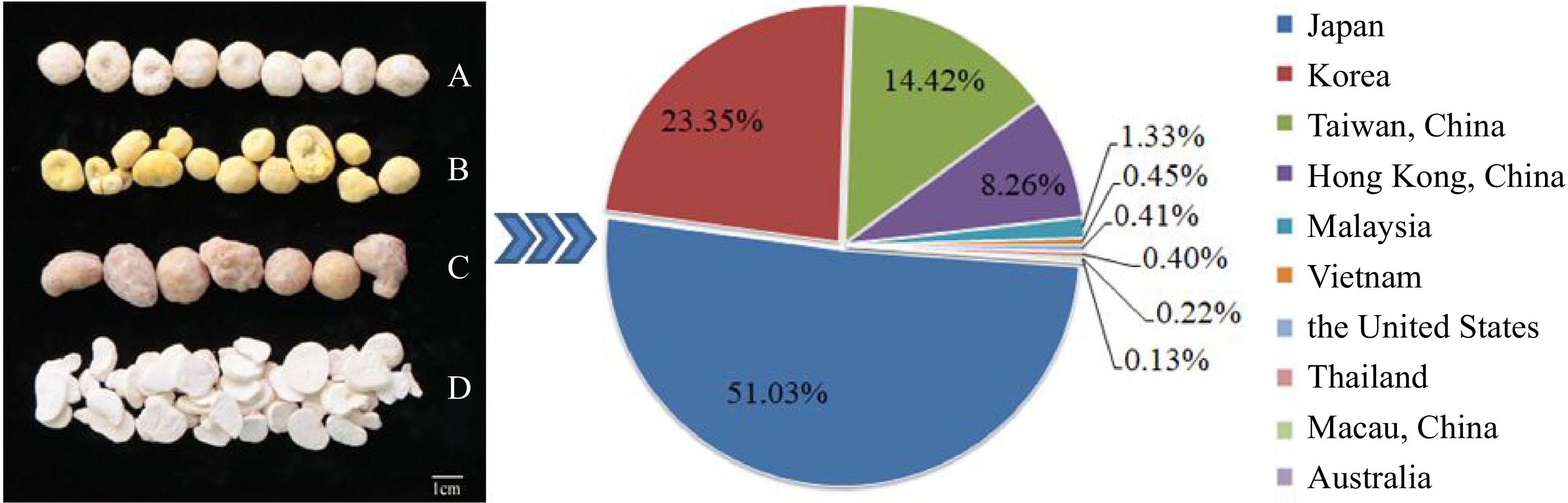
Figure 4.
The proportion of P. ternata (left) exports of major countries and regions in 2022 (right). (A) Pinelliae Rhizoma; (B) Pinelliae Rhizoma Praeparatum (Fa-Banxia); (C) Pinelliae Rhizoma Praeparatum Cum Zingibere et Alumine (Jiang-Banxia); (D) Pinelliae Rhizoma Praeparatum Cum Alumine (Qing-Banxia).
-
P. ternata is a plant commonly used in traditional Chinese medicine for over 2,000 years to relieve phlegm and cough. This review synthesizes and reviews in detail the research on genetic breeding, genetic diversity, production and planting, and functional gene research of P. ternata (Fig. 5).
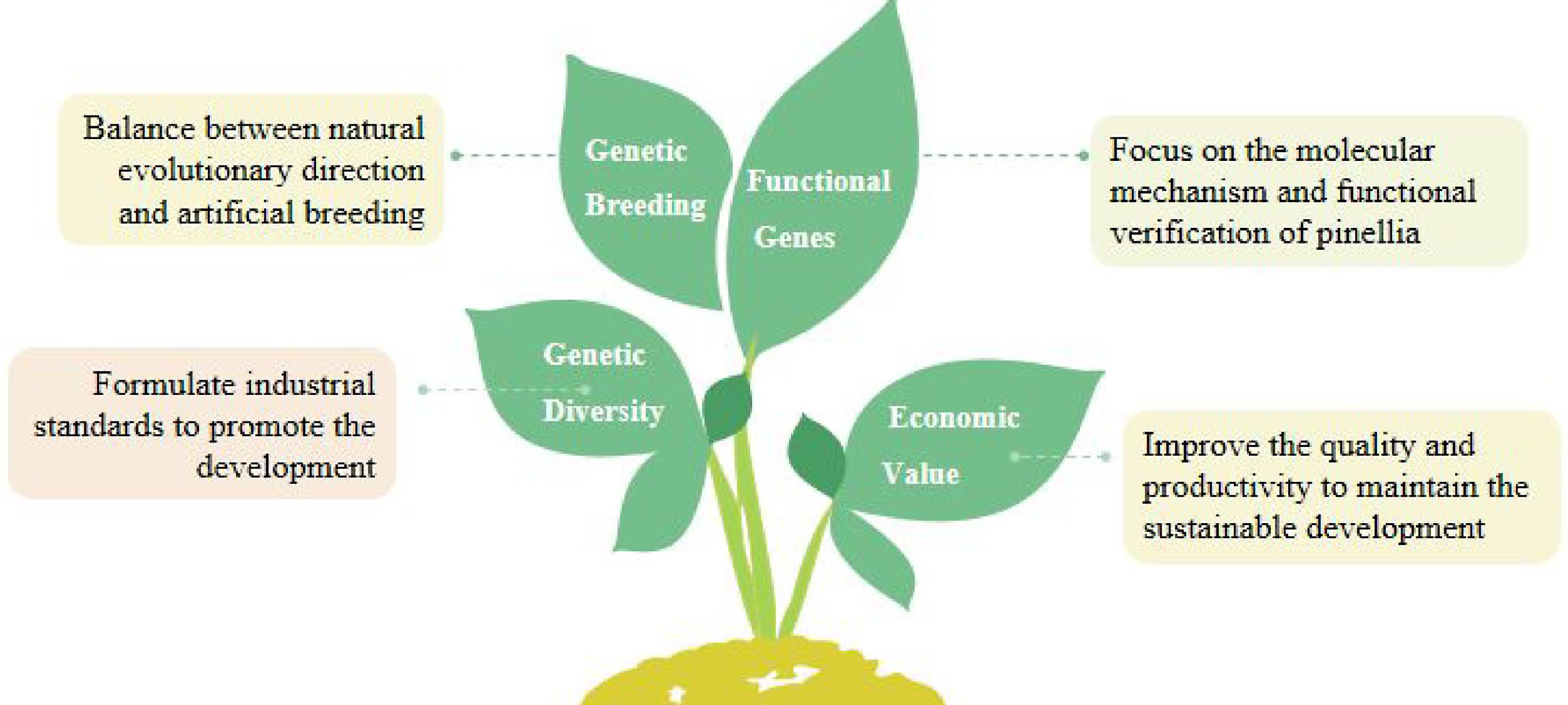
Figure 5.
The solution of P. ternata is proposed for breeding and production at present.
P. ternata, as a medicinal material, is often mixed with plants of the same genus. However, P. pedatisecta Schott and P. ternata are the only species of the Pinellia genus that are used as medicines and listed in Chinese Pharmacopoeia[2]. P. ternata tubers are often indistinguishable when mixed with tubers from the same genus confusing both doctors and patients. In the chemical separation of P. pedatisecta Schott, a monomer compound, triglochinic acid, can be used to effectively distinguish tubers mixed with others from the same genus[62]. In addition, the ITS2 sequence can be used to effectively and stably identify P. ternata and its counterfeits[63]. However, there are a variety of plants under the taxonomy of Pinellia and the classification is inconsistent. Some plants have not been reported, so there is an error in identification which brings serious problems for drug identification. More recent studies suggest that more existing germplasm resources should be collected for research and discussion[64]. The lack of germplasm resources seriously restricts the process and development of P. ternata research.
Breeding is the key means of solving the problem of production. Molecular breeding has become the best choice to solve the problem of species breeding. The genome is the key to revealing the origin, evolution, and genetic breeding of a species. Unfortunately, P. ternata often appears as a polyploid heterozygote, especially 7-ploid and 8-ploid, making it complex and difficult to study. Researchers can look to solutions to similar problems in different species which could be applied in P. ternata to construct the genome. In a similar situation involving the peanut genome, multiple resources were collected to establish the peanut reference genome for resequencing, providing new insights into the genetic evolution of the plant[65]. For crucian carp with multiple ploidies, the separation of characters through traditional hybridization can reveal the origin of the species[66,67]. As for P. ternata, traditional breeding methods, such as self-cross, backcross, or cross may reveal the evolutionary process and direction of P. ternata and provide ideas for its genetic breeding. At the same time, the use of modern molecular genetics may be able to explain the path of P. ternata evolution, such as gene editing or overexpression could be used to identify gene function, and molecular marker could be designed to screen target resources.
In addition, more research should be done to relieve plant disease by molecular biological application. The demand for P. ternata is increasing gradually, and the problems of production and quality need to be solved. Firstly, varieties with high yields and resistance to diseases should be selected and cultivated by breeding. Secondly, natural active pesticides, such as antagonists, natural active molecules, etc., should be developed based on existing research to prevent the breeding of diseases combined with cultivation management. Future research should also focus on the discovery of disease-resistant and insect-resistant resources, the exploitation of functional genes under environmental stresses, and the utilization of functional genes and molecular biological techniques to benefit P. ternata production and processing.
-
Conceptualization: Liu D; performed experiment: Zhang J, Luo M, Xu J and Wang M; supervision: Liu D, Miao Y; writing preparation: Zhang J, Luo M; funding acquisition: Liu D, Luo M. All authors have read and agreed to the published version of the manuscript.
Supported by the Funds for Key Program for Traditional Chinese Medicine of Hubei University of Chinese Medicine (Grant No. 2022ZZXZ001), the Postdoctoral innovation research position in Hubei Province, the special fund for the construction of modern agricultural industrial technology system (grant no. CARS-21), the third batch of industrial technology system projects in Hubei Province (grant no. HBHZD-ZB-2020-005), and key project at central government level (Grant No. 2060303).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Jingyi Zhang, Ming Luo
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang J, Luo M, Miao Y, Xu R, Wang M, et al. 2023. Germplasm resources, genetic diversity, functional genes, genetic breeding, and prospects of Pinellia ternata (Thunb.) Breit: a review. Medicinal Plant Biology 2:13 doi: 10.48130/MPB-2023-0013 

Germplasm resources, genetic diversity, functional genes, genetic breeding, and prospects of Pinellia ternata (Thunb.) Breit: a review
- Received: 31 March 2023
- Accepted: 19 August 2023
- Published online: 07 October 2023
Abstract: Pinellia ternata (Thunb.) Breit. is a traditional Chinese medicinal plant that has been widely used in China, Japan and Korea to relieve cough, vomiting, and inflammation. This review summarizes the recent research focus on germplasm resources, genetic diversity, functional genes, genetic breeding and prospects of P. ternata. The impact of germplasm resources and biogeography are the key factors of the effects of traditional Chinese medicinal materials, avoiding the medical negligence caused by using the confused medicine. Genetic diversity and genetic breeding are the basis of germplasm improvement. A virus-free technique of tissue culture is used to rapid propagation of P. ternata, promoting the production of seedlings without season restrictions. Functional gene research is the theoretical basis and target of germplasm improvement. Many genes, such as PtsHSP17.2 and PtSAD have been confirmed to play an important role in heat stress, guiding the selection and breeding of heat-resistant and drought-resistant P. ternata resources. Still, some problems exist in the production of P. ternata, presenting a challenge in breeding and cultivation. We summarize previous studies here and propose directions for further study to advance the research in the production of P. ternata.
-
Key words:
- Pinellia ternata /
- Germplasm resources /
- Genetic diversity /
- Functional genes /
- Genetic breeding