









-
Amomi Fructus, known as 'Sharen' in Chinese, is an important traditional medicine that is widely used throughout southeast and east Asia, and which has been used for medicinal purposes for over 1,300 years[1−3]. In China, Amomi Fructus is used in traditional Chinese medicine (TCM) as a prescribed treatment for various gastrointestinal diseases[4−7]. It is also a popular condiment to food in many Asian countries[8]. According to the Chinese Pharmacopoeia, Amomi Fructus originates from the ripe fruits of Amomum villosum, A. xanthioides, and A. longiligulare[9]. However, the Japanese Pharmacopoeia lists only A. xanthioides as the source of Amomi Fructus.
Of the three Amomum species, A. villosum, which originates from Yangchun City in Guangdong Province of China, has traditionally been regarded as having superior medicinal quality[10−13]. Consequently, A. villosum fetches a substantially higher price than the other two Amomum species. Nonetheless, many recent studies indicate that A. xanthioides and A. longiligulare have biological functions that can promote the treatment of spleen and stomach stagnation, and other digestive disorders[14−17]. Moreover, A. villosum and A. xanthioides share many similar physical characteristics; the two species can be distinguished by neither chemical methods, microscopy, nor DNA barcoding. Instead, experts must examine the color of the fresh fruit and root sheath to tell the two species apart. In contrast, A. longiligulare is morphologically and chemically distinct from A. villosum and A. xanthioides, and is also distinguishable by DNA barcoding.
Chloroplasts (cp) are the major organelles responsible for photosynthetic and biosynthetic activities in plant cells. The cp genome also provides a particularly useful model for understanding the evolution and systematics of plants, as well as for comparative genetic studies owing to its highly conserved, simple structure and genetic content[18,19]. In studies on phylogenetics, DNA barcoding, population biology as well as transcriptomics, genetic information from cp genomes has been used to elucidate the relationships between different species, their common structures, and unique features[20−24]. With the rapid development of gene sequencing technology in recent years, increasing numbers of cp sequences have quickly been obtained[25−29]. At present, the cp genomes of over 1,500 plant species have been entered into the National Center for Biotechnology Information (NCBI) database.
In this study, we determine the complete cp genome sequences of A. villosum, A. xanthioides and A. longiligulare. To advance understanding of the relationships among the three Amomum medicinal species, we undertake a comparative analysis to clarify details of their physical characteristics and chloroplast genome features. We also construct a phylogenetic tree for Amomum species that is based on 53 protein-coding genes found in 16 closely related species and two outgroup species. Overall, our results provide molecular information that is valuable for guiding the conservation of the three Amomum medicinal species and for improving understanding of their phylogenetic relationships.
-
Fresh plants of A. xanthioides, A. villosum and A. longiligulare were collected from Jinghong City in Yunnan Province, Yangchun City in Guangdong Province and Haikou City in Hainan Province (China), respectively. Total genomic DNA was extracted from samples weighing approximately 100 mg using the DNeasy Plant Mini-Kit (Qiagen Gmbh, Hilden, Germany) according to the manufacturer’s instructions. Purified genomic DNA was quantified using a TBS-380 fluorometer (Turner BioSystems Inc., Sunnyvale, CA, USA). Genomic DNA was fragmented into 400–500 bp using a Covaris M220 Focusd-ultrasonicator (Covaris, Woburn, MA, USA). Library preparation was conducted using the TruSeq DNA Sample Prep Kit. At least 5 μg of genomic DNA was used for each strain when constructing the sequencing library for Illumina sequencing. Paired-end libraries with an insert size of ~300 bp were constructed according to the manufacturer’s instructions (Bioscientific, AIR™ Paired-End DNA Sequencing Kit). Subsequently, the 100 bp at each end was sequenced using Illumina Hiseq4000.
Assembly and annotation of the chloroplast genome
-
The chloroplast genomes were assembled by MITObim (v1.9.1)[30]. Gaps in the assembled sequences were filled using GapCloser[31]. The final circular step was checked manually. The final assembly generated a circular genome sequence with no gaps. Annotation of the chloroplast genome was performed using DOGMA (
www.dogma.ccbb.utexas.edu/ )[32]. tRNAs were identified using tRNAscan-SE (v1.23,http://lowelab.ucsc.edu/tRNAscan-SE ) and rRNAs were determined using RNAmmer (v1.2,www.cbs.dtu.dk/services/RNAmmer/ )[33,34]. Identification of predicted coding sequences (CDS) was performed using Glimmer version 3.02 (http://cbcb.umd.edu/software/glimmer )[35]. All CDS with less than 300 base pairs were discarded. The cp genomes were illustrated using the software OrganellarGenome DRAW (http://ogdraw.mpimp-golm.mpg.de )[36].Sequence analyses
-
To investigate the distribution of condon usage, relative synonymous condon usage values (RSCU) were detected using the program CondonW (v1.4.2, available from:
http://downloads.fyxm.net/CodonW-76666.html )[37]. The REPuter program was used to identify repeats including forward, palindrome, reverse and complement sequences with a length ≥ 30 bp and sequences for which ≥ 90% were identified in chloroplast genomes[38]. The SSRs (Simple sequence repeats) were detected using MISA software using the thresholds: 10, 5, 4, 3, 3, and 3 repeat units for mono-, di-, tri-, tetra-, penta- and hexa-nucleotide SSRs, respectively. mVISTA software was used to compare the complete cp genomes of A. xanthioides, A. villosuman and A. longiligulare[39].Phylogenetic analysis
-
A total of 24 complete chloroplast genomes were downloaded from the NCBI, with the genomes of Kingia australis and Mauritia flexuosa serving as outgroups. The 53 protein-coding gene sequences commonly present in 27 species, including the three species in this study, were aligned using Clustal W2[40]. Subsequently, RAxML-HPC 2.7.6.3 and PAUP in XSEDE at CIPRES Science Gateway (
www.phylo.org/ ) were employed to construct the ML (Maximum likelihood) tree with 1,000 bootstrap replicates[41−43]. -
All three Amomum chloroplast genomes displayed typical quadripartite structures, which had similar lengths and base compositions in their respective genomes and corresponding regions (Supplemental Table S1). The largest genome was that of A. villosum (162,355 bp), while those of A. xanthioides and A. longiligulare were 161,889 bp and 161,990 bp, respectively. The total GC content of the A. villosum genome (35.1%) was slightly lower than that of the other two species (both 35.2%). In A. villosum , a higher GC content was found in the inverted repeats (IRs) region than in the large single-copy (LSC) and small single-copy (SSC) regions, a pattern that was also observed in the other two Amomum species as well as in the cp genomes of other species reported in the literature (Fig. 1, Supplemental Figs S1 & S2). The GenBank accession numbers of A. villosum, A. longiligulare, and A. xanthioides are MH165483, MH165484, and MH165485, respectively.
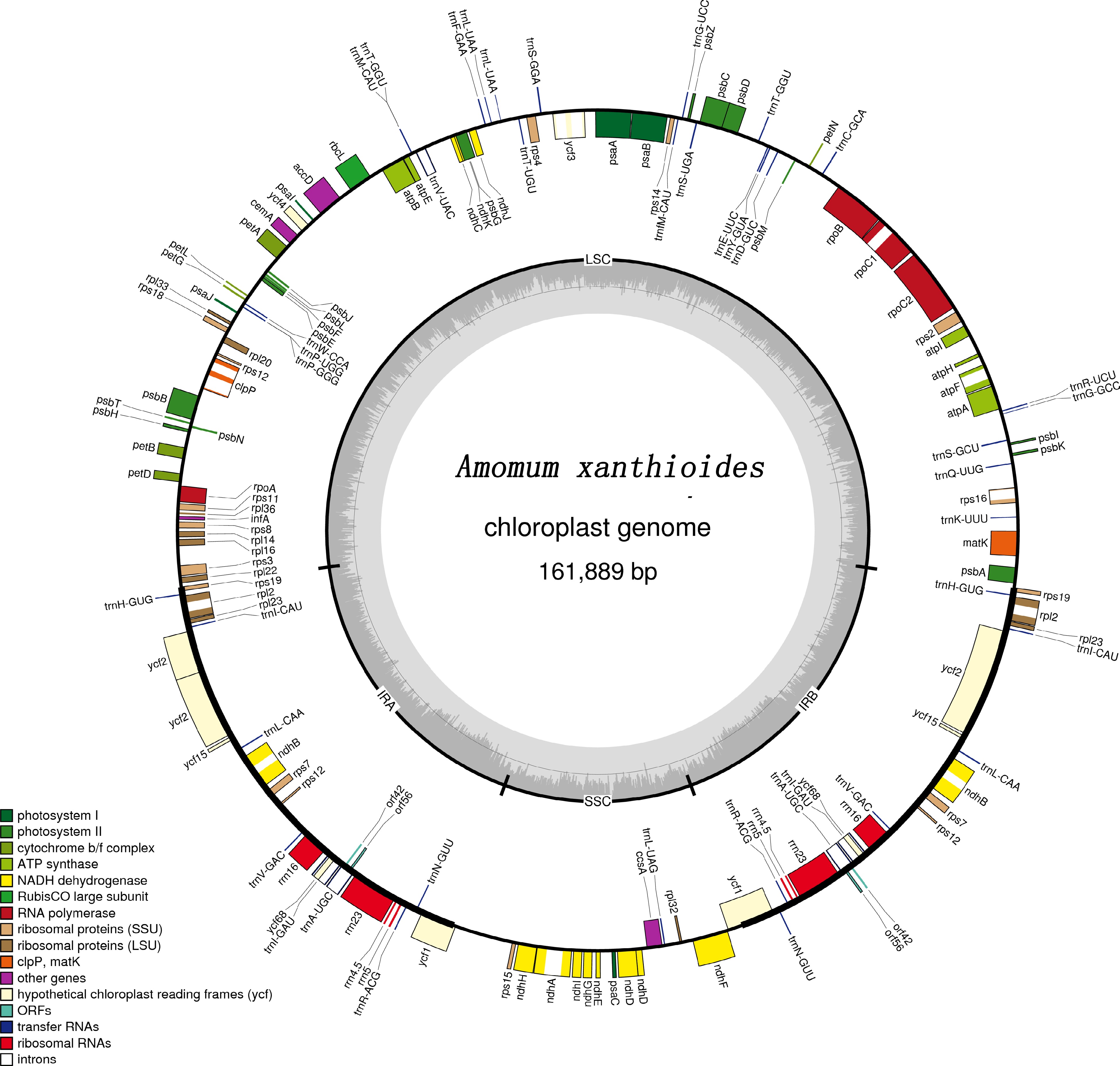
Figure 1.
Gene map of the complete chloroplast genome of A. xanthioides. Genes located on the inside and outside of the circle are transcribed clockwise and counterclockwise, respectively. The inner circle indicates the GC and AT content.
A total of 139, 138, and 139 genes were encoded from the chloroplast genomes of A. xanthioides, A. villosum, and A. longiligulare, respectively. The three Amomum species also had different tRNA amounts: a total of 39, 38, and 39 tRNAs were identified in A. xanthioides, A. villosum, and A. longiligulare, respectively. The three species were also found to share another 92 protein-coding genes and eight rRNAs. Duplicates were found in ten protein-coding genes, seven tRNAs, and all rRNAs genes. After the duplicates were removed, there were 118 unique protein-coding genes, 117 unique tRNAs, and 118 unique rRNAs genes (Table 1). Although the tRNA trnI-CAU was present in A. longiligulare and A. xanthioides, it was absent from A. villosum. Among these genes, nine protein-coding (atpF, ndhA, ndhB, rpoC1, rps16, rpl2, rpl16, ycf15, ycf68) and four tRNA (trnA-UGC, trnI-GAU, trnL-UAA, trnV-UAC) genes contained one intron, and three genes (rps12, ycf3, clpP) contained two introns (Table 2).
Table 1. Gene contents of three Amomum chloroplast genomes.
No. Group of genes Gene names Amount 1 Photosystem I psaA, psaB, psaC, psaI, psaJ 5 2 Photosystem II psbA, psbB, psbC, psbD, psbE, psbF, psbG, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ 16 3 Cytochrome b/f complex petA, petB, petD, petG, petL, petN 6 4 ATP synthase atpA, atpB, atpE, atpF*, atpH, atpI 6 5 NADH dehydrogenase ndhA*, ndhB*(×2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK 12(1) 6 RubisCO large subunit rbcL 1 7 RNA polymerase rpoA, rpoB, rpoC1*, rpoC2 4 8 Ribosomal proteins (SSU) rps2, rps3, rps4, rps7(×2), rps8, rps11, rps12**(×2), rps14, rps15, rps16*, rps18, rps19(×2) 15(3) 9 Ribosomal proteins (LSU) rpl2*(×2), rpl14, rpl16*, rpl20, rpl22, rpl23(×2), rpl32, rpl33, rpl36 11(2) 10 Proteins of unknown function ycf1(×2), ycf2(×2), ycf3**, ycf4, ycf15*(×2), ycf68*(×2) 10(4) 11 Other genes accD, clpP**, matK, ccsA, cemA, infA 6 12 Transfer RNAs# 39/38/39 tRNAs 39/38/39(7) 13 Ribosomal RNAs rrn4.5(×2), rrn5(×2), rrn16(×2), rrn23(×2) 8(4) * Gene with one intron; ** gene with two introns; (×2) indicates the number of the repeat unit is 2. # indicates different amount among A. xanthioides, A. villosum and A. longiligulare. Table 2. Gene with introns in three Amomum chloroplast genomes and the length of exons and introns.
Gene Location Exon I (bp) Intron I (bp) Exon II (bp) Intron II (bp) Exon III (bp) atpF# LSC 378 770/771/771 207 − − clpP# LSC 252 627/626/629 237 908/909/909 72 ndhA# SSC 567 1,056/1,067/1,054 513 − − ndhB IR 849 603 783 − − rpl16# LSC 8 1,045/1,042/1,038 420 − − rpl2 IR 426 658 396 − − rpoC1# LSC 1605 751/752/739 438 − − rps12 LSC 156 − 159 156 159 rps16# LSC 234 703/707/721 46 − − trnA-UGC IR 38 803 35 − − trnI-GAU# IR 35/42/35 937 42/35/42 − − trnL-UAA# LSC 35 563/534/534 50 − − trnV-UAC# LSC 37 606/606/603 38 − − ycf3# LSC 138 773/773/774 249 710/711/710 135 # indicates different amount among A. xanthioides, A. villosum and A. longiligulare. IR contraction and expansion
-
The LSC/IRa/SSC/IRb junctions of the three Amomum cp genomes were compared, and expansions and contractions in IR boundary regions were also observed (Fig. 2). The ycf1 gene was located at the SSC/ IRa boundary in A. villosum and A. longiligulare, but located entirely within the IRa region in A. xanthioide. In addition, the SSC/IRb border extended into the ycf1 pseudogene in all three Amomum species. Overall, while the IR/SC junctions of all three Amomum species were similar, A. xanthioides displayed differences at the ycf1 gene in comparison with the other two species.

Figure 2.
Comparison of the borders of the LSC, SSC, and IR regions in the genomes of the three Amomum species. Ψ indicates pseudogenes. The number above the gene feature indicates the distance between the ends of genes and border sites. Figure is not drawn to scale.
Condon usage
-
There were 53963, 54118, and 53996 codons in the chloroplast genomes of A. xanthioides, A. villosum, and A. longiligulare, respectively. The codon usage frequency and RSCU were analyzed. Leucine and tryptophan were the most and least encoded universal amino acids, respectively. Except for methionine and tryptophan, most amino acids are encoded by multiple, synonymous codons (Fig. 3). Additionally, in all three Amomom species, codon usage was biased towards codons that terminated in A or U; this has similarly been observed in other species[19, 23, 27].
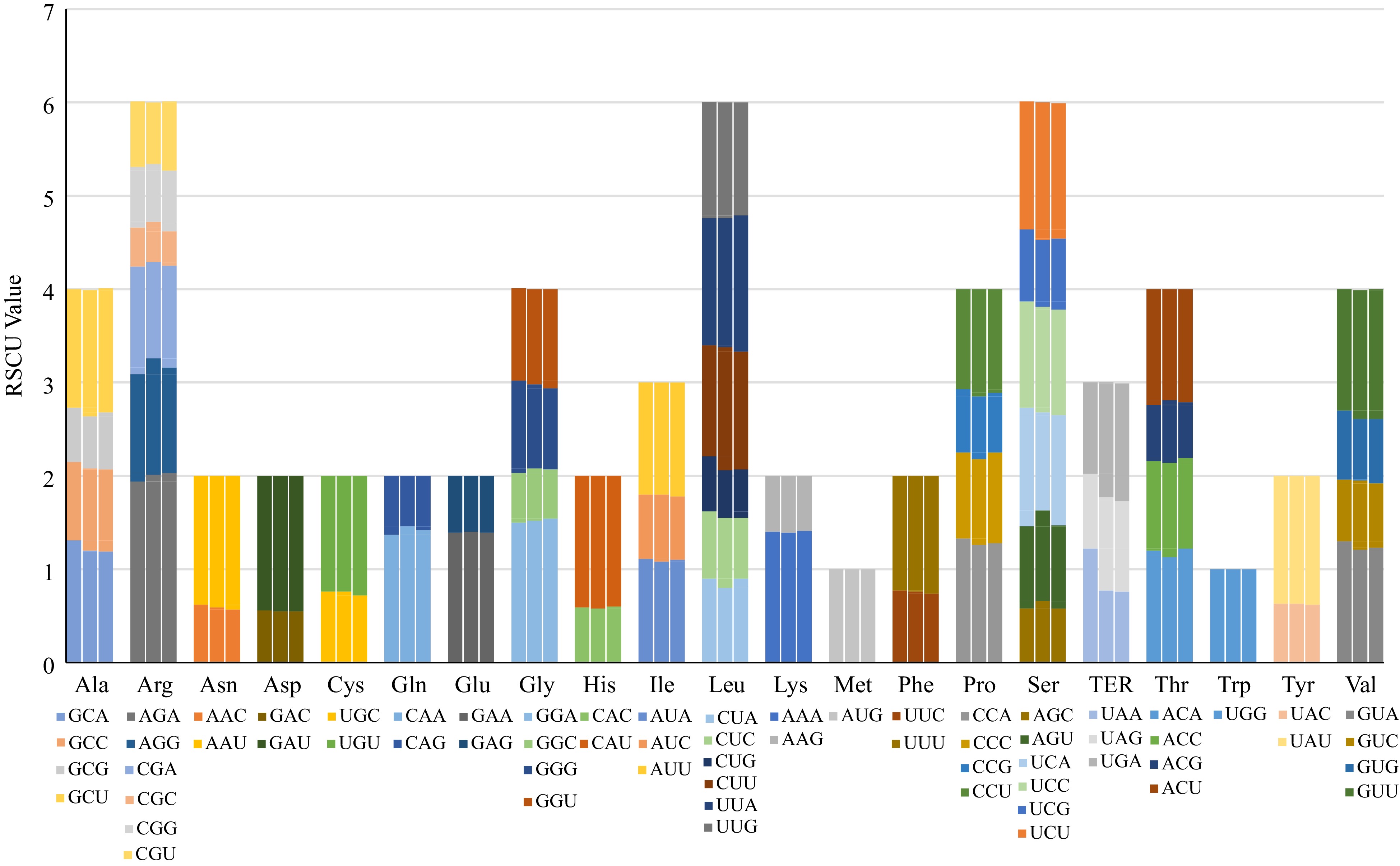
Figure 3.
Codon content of 20 amino acids and stop codons in all protein-coding genes of the chloroplast genomes. The histogram (as read from left to right) shows the different numbers of codons found in A. xanthioides, A. villosum and A. longiligulare.
Repeat structure and analysis of simple sequence repeats
-
An analysis of repeat sequences using REPuter detected no complement repeats in all three Amomum chloroplast genomes. Figure 4 shows the results of the analysis for repeat structures in the three Amomum species. We found that while the chloroplast genome of A. xanthioides had the fewest palindromic repeats (7) and forward repeats (14), it had the most reverse repeats (13) among the three Amomum species. In comparison, the chloroplast genome of A. villosum had the greatest number of palindromic repeats (28) and forward repeats (18), and no reverse repeats. The chloroplast genome of A. longiligulare had 21 palindromic repeats, 20 forward repeats, and one reverse repeat. Most of the repeated units were different among the three Amomum species; that is, any repeated sequences were molecularly unique to each species.
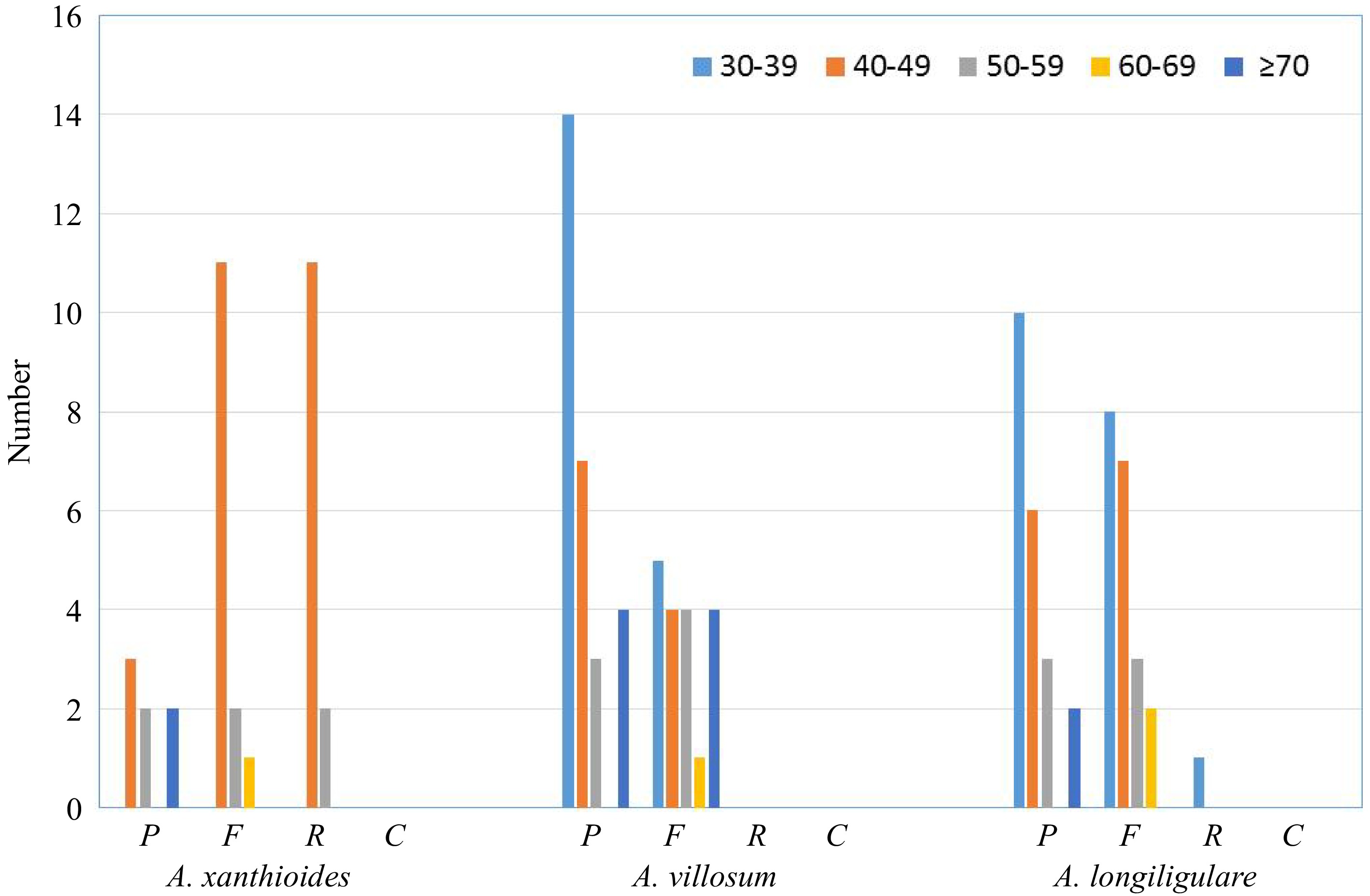
Figure 4.
Repeat sequences in three Amomum chloroplast genomes. REPuter was used to identify repeat sequences with length ≥ 30 bp and sequences for which ≥ 90% were identified in chloroplast genomes. Different letters are used to indicate forward repeats (F), palindromic repeats (P), reverse repeats (R), and complementary repeats (C). Repeats of different lengths are indicated in different colors.
A total of 94 SSRs were detected in the chloroplast genome of A. xanthioides (Table 3). Rich mononucleotide repeats were identified. Specifically, there were 57, 15, 3, 13, 2 and 4 mono-, di-, tri-, tetra-, penta- and hexa-nucleotide repeats, respectively. In line with the results of previous studies on many other plants species[44,45], high contents of AT were found in the SSRs of A. xanthioides. Only 17 SSRs were composed of G and C, while 77 SSRs were composed of A and T. The chloroplast genomes of the other two Amomum species shared similar types and amounts of SSRs. The chloroplast genome of A. xanthioides had two and five types of SSRs that presented in different amounts from the chloroplast genomes of A. villosum and A. longiligulare, respectively.
Table 3. SSR types and amounts in three Amomum chloroplast genomes.
SSR type Repeat unit Amount Ratio (%) A. xanthioides A. villosum A. longiligulare A. xanthioides A. villosum A. longiligulare Mono A/T 57 62 58 100 100 100 Di GA/TC 2 2 2 13.3 10.5 11.8 AT/AT 13 17 15 86.7 88.2 86.7 Tri GCT/AGC 1 1 1 33.3 25.0 33.3 TTC/GAA 1 1 1 33.3 25.0 33.3 GGA/TCC 1 1 1 33.3 25.0 33.3 ATA/TAT 0 1 0 0.0 25.0 0.0 Tetra TTAT/ATAA 2 2 2 15.4 18.2 18.2 AAAG/CTTT 1 1 1 7.7 9.1 9.1 ATTT/AAAT 2 2 1 15.4 18.2 9.1 CTAA/TTAG 1 1 1 7.7 9.1 9.1 AGAA/TTCT 2 2 2 15.4 18.2 18.2 CGTA/TACG 1 1 1 7.7 9.1 9.1 CATA/TATG 1 1 0 7.7 9.1 0.0 GAAT/ATTC 1 1 1 7.7 9.1 9.1 TATT/AATA 1 1 1 7.7 9.1 9.1 AATT/AATT 1 1 1 7.7 9.1 9.1 Penta TTAAA/TTTAA 1 1 1 50.0 50.0 50.0 AATCA/TGATT 1 1 1 50.0 50.0 50.0 Hexa TGATAG/CGATCA 1 1 1 25.0 25.0 50.0 GAAGAG/CTCTTC 1 1 0 25.0 25.0 0.0 TCCTCT/AGAGGA 1 1 0 25.0 25.0 0.0 TCACTA/TAGTGA 1 1 1 25.0 25.0 50.0 Interspecific comparisons of sequence identity among the chloroplast genomes of the three Amomum species were conducted using mVISTA (Fig. 5). We found that the chloroplast genomes of all three Amomum species were highly conserved, and that their IR regions were less divergent than their LSC and SSC regions. However, any non-coding regions appeared to be more variable globally than the coding regions. Highly divergent regions were located at inter-gene spacers, including matK-trnK-UUU, rps16-trnQ-UUG, atpF-atpH, atpH-atpl and psbM-trnT-GGU, with ycf1 being the most divergent coding region. These highly divergent regions could be used to develop potential markers for future phylogenetic analyses and molecular identification of Amomum species.
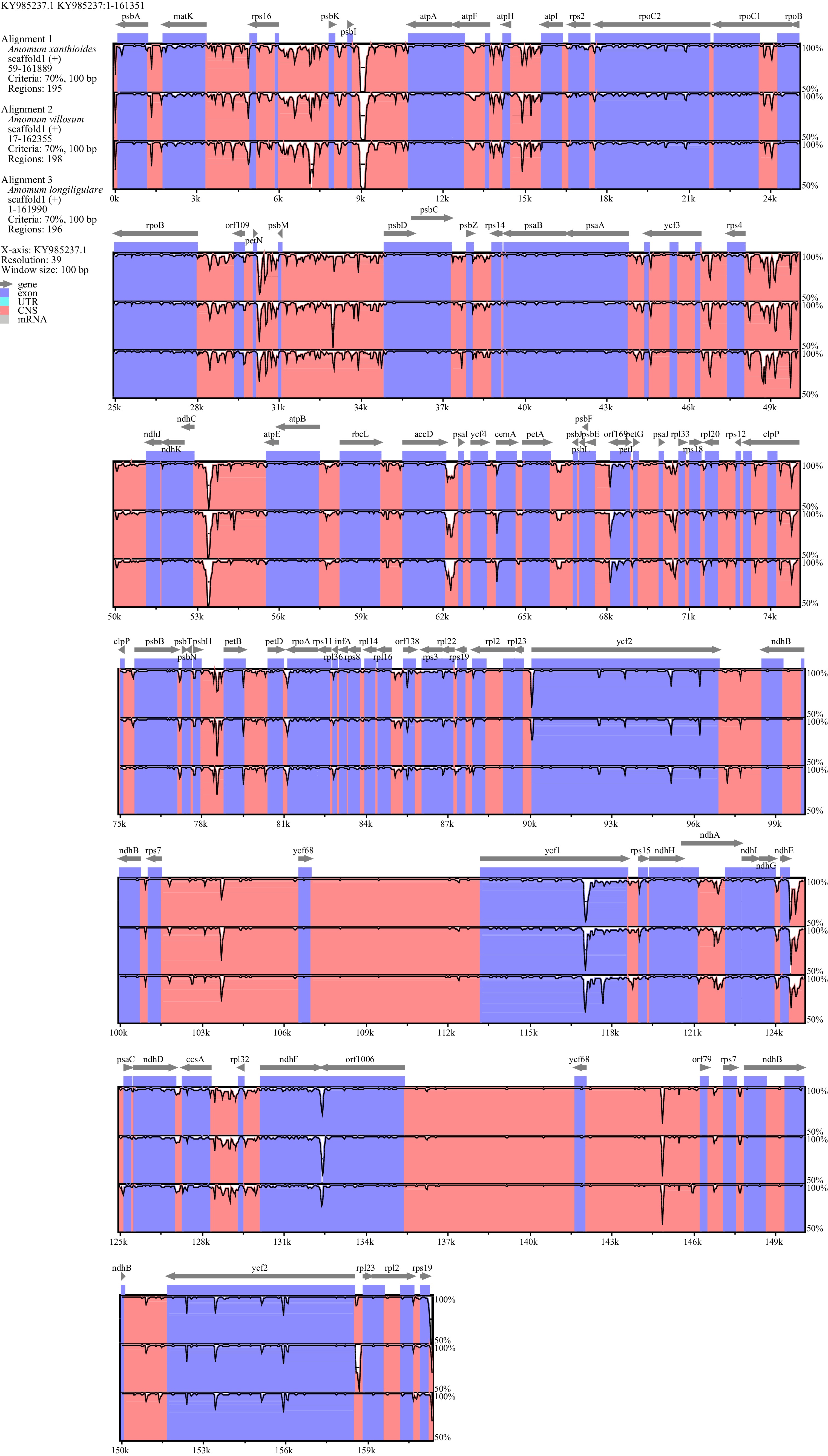
Figure 5.
Sequence alignment in the chloroplast genomes of three Amomum species using mVISTA software. The top line shows the genes in order (with the direction of transcription indicated by an arrow). A cut-off of 70% identity was used for the plots. The Y-axis indicates the value of percent identity and ranges from 50% to 100%.
Phylogenetic analyses
-
Due to the limited availability of extensive biological samples per species in previous phylogenetic studies on chloroplast genomes of Amomi Fructus[46−49], we addressed this issue by integrating all sequences in our study. We have arranged a comprehensive set of 12 chloroplast genomes, including A. villous, A. longiligulare, and A. xanthioides, alongside 13 from the Zingiberales clade. For phylogenetic analysis, we utilized two Arecales species as out-groups. We performed multiple sequence alignment to construct the maximum parsimony (MP) and maximum likelihood (ML) phylogenetic tree based on 53 common protein-coding genes (Fig. 6). The maximum likelihood (ML) phylogenetic analysis yielded robust results, with A. longiligulare clustered into a monophyletic clade separately from A. villosum and A. xanthioides, and branches of A. villosum and A. xanthioides formed a distinct clade. Furthermore, the nested relationship observed among all samples of A. villosum and A. xanthioides. The results suggested that the common protein-coding genes can be used as a basis for identifying A. longiligulare, but cannot be used as a basis for identifying A. villosum and A. xanthioides. The phylogenetic positions of the three Amomum species as determined in this study align with previous results obtained with DNA barcoding[50, 51]. Our findings therefore provide an empirical resource to aid the identification of Amomum species as well as species of other genera using the cp genome.
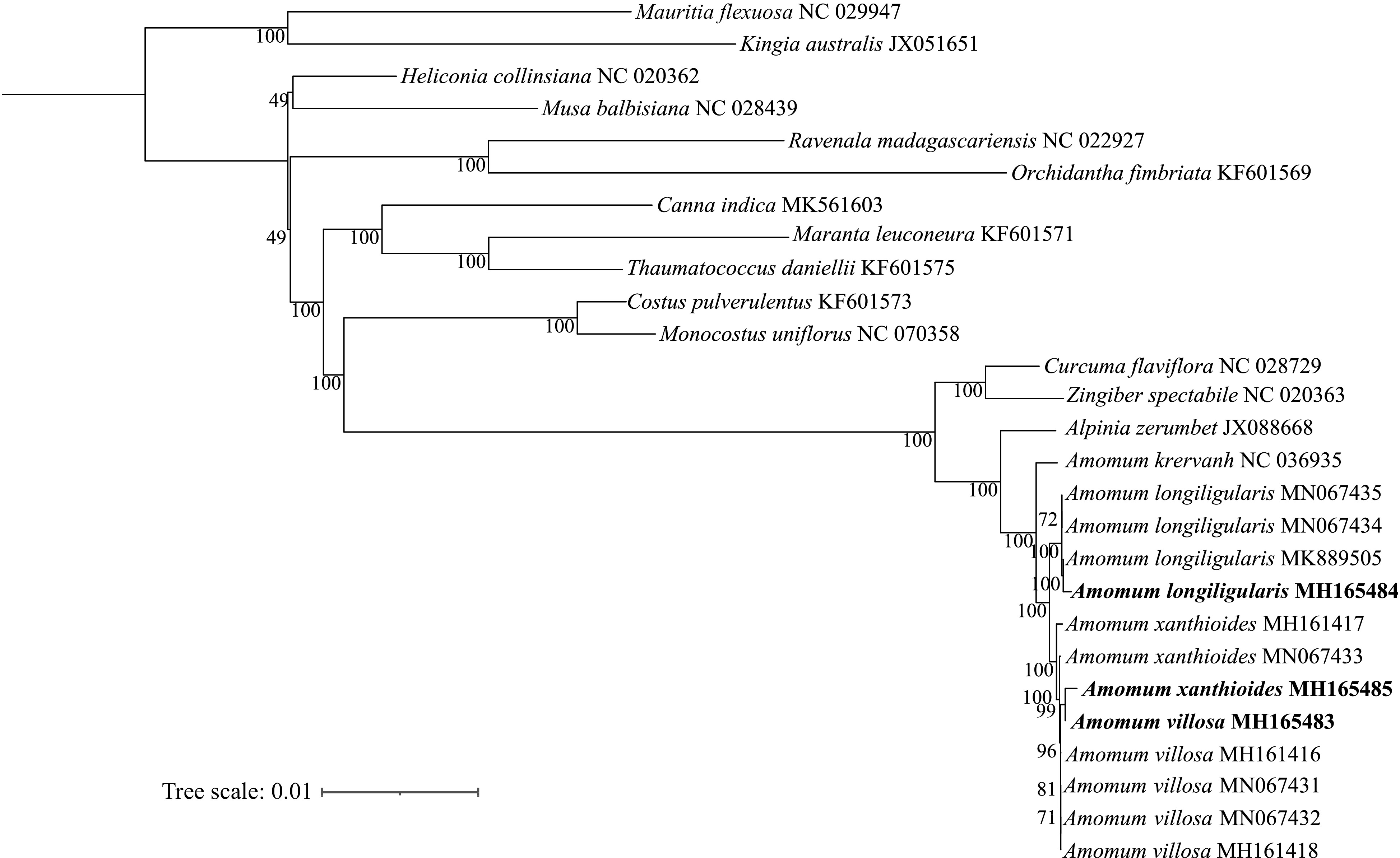
Figure 6.
Phylogenetic tree constructed using the maximum likelihood (ML) method based on 53 protein-coding genes from 16 species in the Zingiberales clade. Numbers at nodes indicate the values of bootstrap support.
-
Here, we constructed the complete chloroplast genomes of A. xanthioides, A. villosum, and A. longiligulare and performed detailed comparisons of the genomes of the three Amomum species. We found that although the cp genomes of the three Amomum species were relatively conserved, they differed in their gene contents as well as patterns of IR contraction and expansion. We investigated repeat sequences and SSRs with the aim of facilitating the development of polymorphic microsatellites and new molecular markers. Our phylogenetic analyses revealed A. xanthioides and A. villosum clustered together, which further supports previous observations of the two species’ high similarity in terms of morphological characteristics, chemical components, and DNA barcodes, suggesting that the evidence does not support considering A. xanthioides as an independent species. In the Chinese Pharmacopoeia, for example, A. xanthioides is considered a variety of A. villosum. Therefore, identification of A. villosum and A. xanthioides using common protein-coding genes of chloroplast genomes is not recommended.
Overall, our findings are useful to evolutionary, phylogenic, population and barcoding studies. The specific results should also inform efforts to genetically engineer and conserve Amomum species, which are important medicinal resources.
-
The authors confirm contribution to the paper as follows: study conception and design: Su C; data collection: Wang H, Wang S, Zhang W, Wang B; analysis and interpretation of results: Su C, Zhao J, Meng X; draft manuscript preparation: Su C, Zhao J, Wang S. All authors reviewed the results and approved the final version of the manuscript.
-
The complete chloroplast genome sequences of A. villosum, A. longiligulare, and A. xanthioides was submitted to GenBank (
www.ncbi.nlm.nih.gov ) (accession numbers: MH165483, MH165484, and MH165485). This work was supported by the National Key R&D Program of China from the Ministry of Science and Technology of China (Grant No. 2021YFE0100900). We thank Chao Xiong and TopEdit (
www.topeditsci.com ) for their linguistic assistance during the preparation of this manuscript.-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Gene map of the complete chloroplast genome of A. villosum. Genes located on the inside and outside of the circle are transcribed clockwise and counterclockwise, respectively. The inner circle indicates the GC and AT content.
- Supplemental Fig. S2 Gene map of the complete chloroplast genome of A. longiligulare. Genes located on the inside and outside of the circle are transcribed clockwise and counterclockwise, respectively. The inner circle indicates the GC and AT content.
- Supplemental Table S1 Base composition of three Amomum chloroplast genomes.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Su C, Wang H, Wang S, Zhao J, Meng X, et al. 2023. Analysis of whole chloroplast genomes from three medicinal Amomum species. Medicinal Plant Biology 2:14 doi: 10.48130/MPB-2023-0014 

Analysis of whole chloroplast genomes from three medicinal Amomum species
- Received: 09 May 2023
- Accepted: 08 September 2023
- Published online: 11 October 2023
Abstract: Amomum xanthioides, A. villosum and A. longiligulare are three medicinally important herbs that are widely cultivated in southeast Asia. The ripe fruits of all three species are not only used worldwide in treatments for gastrointestinal diseases, but are also popular condiments in cooked food. In this study, we determined and analyzed the complete chloroplast (cp) genome sequences of these three Amomum species. The sizes of the cp genomes of A. xanthioides, A. villosum and A. longiligulare were 161,889 bp, 162,355 bp, and 161,990 bp, respectively. The cp genomes of all three species displayed a typical quadripartite structure. The cp genomes of A. xanthioides, A. villosum, and A. longiligulare encoded a total of 139, 138, and 139 genes, respectively, of which 118, 117, and 118 genes were single-copy genes. These included 92 protein-coding genes, eight rRNAs, as well as 39, 38, and 39 tRNAs, respectively. A comparative analysis revealed that the cp genomes of the three Amomum species had similar characteristics and patterns of organization. However, they also varied in terms of gene content, the expansion of inverted repeats, codon usage, repeat sequences, and simple sequence repeats. A phylogenetic analysis strongly supported a sister relationship between A. xanthioides and A. villosum. Overall, the results advance understanding of the relationships among the three medicinally valuable Amomum species, and provide basic molecular information to aid conservation efforts as well as research in phylogenetics and systematics.
-
Key words:
- Amomum /
- Medicinal plants /
- Chloroplast genomes /
- Phylogeny