









-
In angiosperms or flowering plants, 'Florogenesis' is the transitioning process from vegetative tissue to reproductive organs or flowers in a plant's apical meristem[1]. This transition is governed by flowering genes in which expression is influenced by environmental factors such as vernalization, photoperiod, gibberellins, an autonomous pathway[2]. In Arabidopsis, several genes have been discovered which are involved in flowering and act as floral integrators. These flowering genes include FT, SOC1, CO, VRN1, PPD, FCA, FLD, and FLK[3,4]. These floral integrator genes specifically upregulate flowering by promoting transition from vegetative to flowering or repressing floral repressor genes. Repressor genes act as repressors of floral integrator genes and upregulate the expression of repressors, including genes such as FLC, FRI, FLX, VRN2, and SVP[3, 5−6].
FLC and FLC-like are floral repressors found in many dicotyledon plants, such as Malus[7], Rosa[8], Coffea[9] and Brassica[10]. The FLC gene is regulated by temperature changes throughout the year, both in annuals and perennials. In summer, FLC expression is upregulated through FRIGIDA (FRI) by binding the FLC promoter through the DNA-binding protein SUPPRESSOR OF FRIGIDA4 SUF4[11]. In addition, FRI expression is upregulated by FLX; both SUF4 and FLX are in the FRI-specific pathway[12]. In winter, FLC is down-regulated through a process of vernalization as prolonged exposure of low temperature in winter in the meristem gradually reduces the expression of FLC[13]. In addition to the vernalization pathway, the autonomous pathway reduces the expression of FLC both in the meristem and leaves[13]. Gradual reduction of FLC allows FLOWERING LOCUS T (FT) to be expressed in the leaves and transported through the phloem to reach to the meristematic tissue and stimulate the MADS box genes, thereby inducing flowering in Arabidopsis[14].
In wheat and barley, the flowering pathway is regulated by photoperiod, vernalization and the circadian clock[15]. Vernalization gene-2 (VRN2) is a dominant repressor or inhibitor of flowering in winter wheat (Triticum aestivum; a monocot grass) that is down-regulated by vernalization (a cold period; winter)[15]. Subsequently, the floral integrator leads to flowering in winter wheat while spring wheat doesn't require vernalization due to a non-functional VRN2 gene. However, vernalization is a facultative stimulus for earlier flowering in spring wheat[16]. In contrast, maize (Zea mays) and rice (Oryza sativa) rely on plant age to build up sufficient energy requirements in order to transition to flowering through epigenetic action of miR172[17]. In monocot geophytes (defined as herbaceous perennial plants with underground storage organs, e.g. bulbs, corms, tubers, etc., that promote winter survival), such as Gladiolus, Lilium, Tulipa, Narcissus and Crocus, the flowering pathway is poorly understood. Factors of plant growth influencing flowering in commercial geophytes for commercial production are well known[1, 18]. A clearly delineated genetic pathway for monocots has yet to be explored and characterized for many floricultural crops, such as gladiolus. In contrast, the Arabidopsis model is readily applicable to temperate dicotyledon plants[19] but may be only partially useful for monocots. Only a few flowering genes have been discovered in monocot ornamental geophytes[1], such as FT-like in Allium cepa[20,21], FT in Narcissus[22], NLF in Narcissus[23], LFY in Allium sativum[24,25] and in Lilium[26]. Recently, many flowering genes have been discovered in Lilium ×formolongi (FT, CO-like, AP2, GA1, SOC1) and/or proposed for L. formosanum (VER1, VER2)[27,28]. The discovery of flowering genes in geophytes serves as a valuable resource to model flowering pathway(s) therein.
Geophytes such as Gladiolus, Lilium, Tulipa, Narcissus and Crocus are floricultural crops with ornamental value wherein flowering is essential to maintain the marketing value for these crops. Gladiolus ×hybridus Rodigas, commonly known as gladiolus(-i), is commercially cultivated as a cut flower and as garden or landscape plants. Gladioli are geophytic plants with underground modified stem structures known as corms, producing cormels as a means of vegetative propagation[29]. Flower initiation and development are crucial steps for its success as a cut flower. Therefore, understanding the flowering pathway is vital for genetics and breeding to improve the floral market value.
Gladiolus has a genome size of 1,100 Mbp, although it is unclear whether this is for haploid or diploid and the species is unknown[30]. The genome weight for gladiolus was recently measured in G. communis as 0.67−0.68 pg for monoploid G.s. (1Cx, pg) and in G. italicus at 0.61 pg for monoploid G.s. (1Cx, pg)[31]. The limited knowledge of the gladiolus genome is also reflected in the lack of identified gladiolus flowering genes, since none have been discovered except for the gibberellin receptor gene GID1a[31]. The relationship of GID1a with flowering has not been established. In Arabidopsis, gibberellin binds to the gibberellin receptor forming the GID1 complex that binds to DELLA, causing its degradation, thereby enabling SOC1 and LFY to be upregulated, leading to flowering[32−33].
Understanding the flowering pathway and gene expression is important for efficient selective breeding of gladiolus for rapid generation cycling (RGC) or early flowering types that flower in < 1 year from seed. An important flowering gene is FLC, a major flowering repressor found in Arabidopsis and many dicot species. FLC plays a vital role in the control of flower initiation[13]. Gladiolus (Iridaceae) has both summer and winter flowering species. It had been hypothesized that there was no FLC gene in monocot species until the FLC homologue was discovered in some cereal crops, such as Triticum aestivum[34], Hordeum vulgare[35] and Brachypodium distachyon[36]. These studies did not discover FRI genes, which upregulate FLC expression in Arabidopsis thaliana[11]. A hypothesis to test would be that some monocots do not possess the flowering repressor FLC gene and rely on alternative gene(s) to acts as a miR172 repressor as in Zea and Oryza[17]. Additionally, it remains unknown whether there is a FLC-dependent pathway in all other monocots.
In Arabidopsis, FLC is located between two flanking genes, UFC (located 4.7 Kb upstream of FLC) and DOWNSTREAM OF FLOWERING LOCUS C (DFC; found 6.9 Kb downstream of FLC)[37]. The UFC gene expression is repressed by vernalization and independent of FLC repression due to vernalization[37]. Thus, both FLC and UFC are repressed by vernalization, yet are not dependent on each other for expression; the suppression is through chromatin modification in an epigenetic manner[37]. The VRN1 gene is expressed with vernalization and acts as a floral integrator whereas UFC is repressed and required by VRN1 expression dependently[38]. The potential role for UFC in flowering has yet to be discovered and it may not involve flowering at all since vernalization only represses UFC in seeds while DFC is repressed by vernalization of the plant[38]. Insertion of the NPTII gene between the UFC and FLC region confirmed NPTII response to cold as the whole cluster region of FLC responded to cold[37]. In the UFC protein of A. thaliana, a conserved domain DUF966, has 92 amino acids, although its function is still unknown[39]. This lack of knowledge in DUF966 function creates a challenge to identify the function of UFC protein. A recent study showed the role of UFC in A. thaliana, with the genes SOK2 (with a role in embryogenesis, root initiation, growth and branching of the primary and lateral roots)[39] and DUF966 (with a defense-stress response)[40, 41].
The FLX gene encodes a putative leucine zipper domain which is required for FRI-mediated activation of FLC in Arabidopsis[42] (Fig. 1). Up-regulating FLC occurs in winter annual Arabidopsis[33], while late flowering phenotypes exhibit strong expression of FLX which indicates its role in the suppression of flowering[42]. Several genes have been discovered in the FLX gene family, e.g. FLX-LIKE1 (FLL1), FLX-LIKE2 (FLL2), FLX-LIKE3 (FLL3), FLX-LIKE4 (FLL4)[11,33]. FLX and FLL4 are the most crucial genes in flowering time control in Arabidopsis[33].
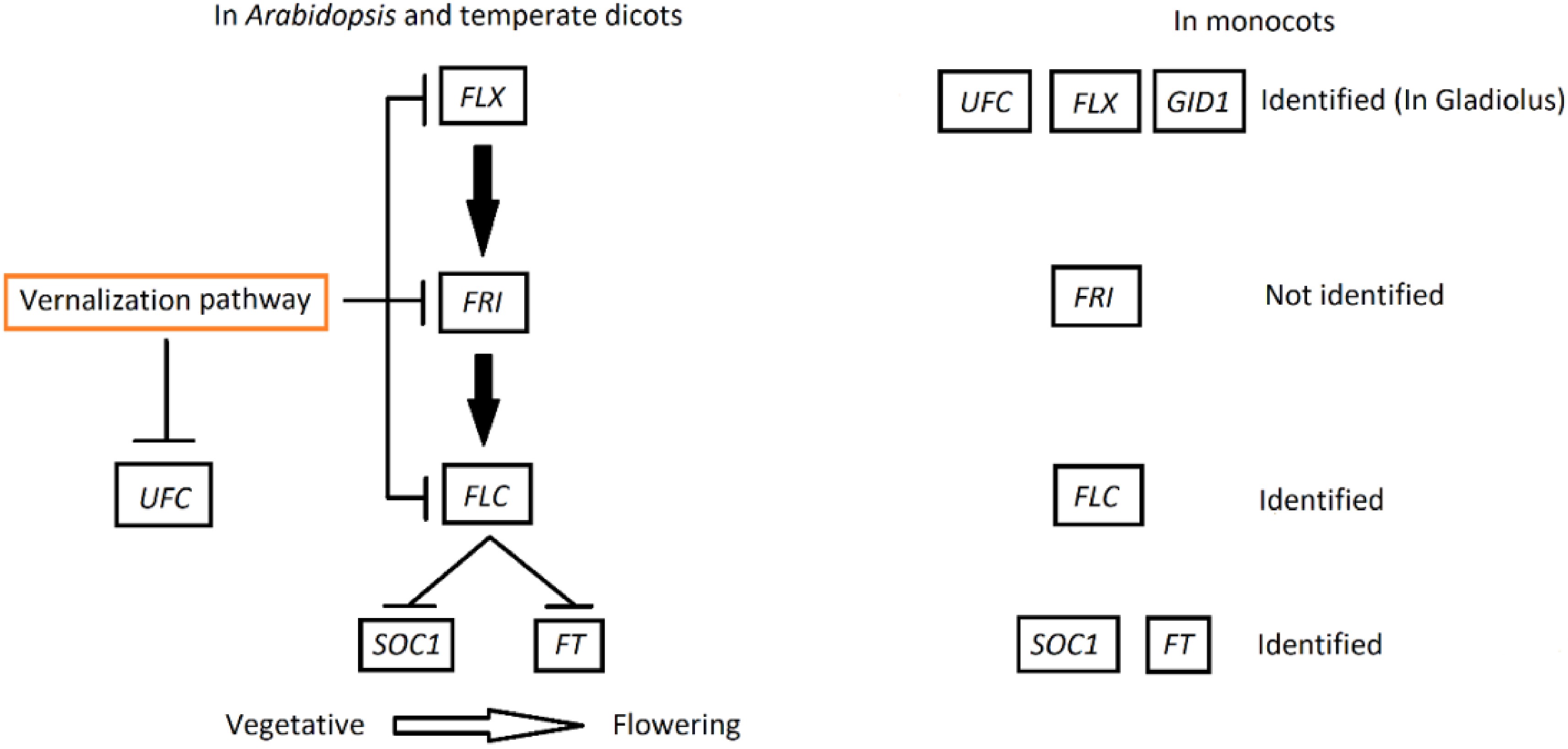
Figure 1.
Model represents portion of flowering pathway regarding the role of the FLX gene in flowering along with the UFC gene in Arabidopsis and temperate dicots. FLX upregulates FRI which also upregulates the expression of the FLC protein and suppresses flowering by repressing the expression of the floral integrator SOC1 and FT. The vernalization pathway downregulates the expression of FLX, FRI and FLC genes, allowing the floral integrators to initiate flowering in vegetative state of dicots, while the vernalization pathway also downregulates the UFC gene[37]. In monocots, FLX, FLC, GID1, SOC1 and FT have been identified[22,35,51], while UFC and FLX have just been identified in gladiolus (in the current study). GIBBERELLIN-INSENSITIVE DWARF1 (GID1) regulates GA synthesis. However, FRI was not identified either by lacking the presence of these repressor genes or monocots relying on other options of the flowering pathway genes.
In order to test whether FLC is present in gladiolus, the adjacent gene (UFC) will also be probed, along with FLX, which is part of the FLC-dependent mechanism. Therefore, the objective of this study is to identify whether UFC and FLX genes occur in the genetically diverse gladiolus germplasm of the University of Minnesota Gladiolus Breeding Program. The null hypotheses tested are: Ho1 = There is no difference among gladiolus genotypes in the existence of the UFC gene; Ho2 = There is no difference among gladiolus genotypes in the presence of FLX.
-
Rapid Generation Cycling (RGC) in gladiolus is the ability of flowering in the first year from seed as an annualized perennial. Eight such RGC genotypes were included in this study[43−45], along with six breeding genotypes and three commercial cultivars ('Vista', 'Glamini®' and 'Carolina Primrose'; Table 1). Classically, seed-propagated gladiolus have 3−5 years as juvenile, non-flowering (vegetative) seedlings before a phase change into flowering (reproductive) adults[44]. The University of Minnesota Gladiolus breeding program developed gladiolus genotypes with a reduced juvenility period and phase change to flowering in less than one year from seed[43−45].
Table 1. Gladiolus genotypes used in this study, their codes and whether they are Rapid Generation Cycling (RGC): + is for RGC genotypes and – for Non-RGC genotypes. All gladiolus genotypes were tested for the presence of UFC, FLX and FRI genes.
Genotype Code RGC Gladiolus ×hybridus 21213 1 + Gladiolus × hybridus 2220 2 + Gladiolus ×hybridus 2231 3 + Gladiolus ×hybridus 2337 4 + Gladiolus ×hybridus 35314 5 + Gladiolus ×hybridus 3923 6 + Gladiolus ×hybridus 3931 7 + Gladiolus × hybridus 74210 8 + Gladiolus ×hybridus 7736 9 + Gladiolus ×hybridus 28236 10 − Gladiolus ×hybridus 15531 11 − Gladiolus ×hybridus 20732 12 − Gladiolus ×hybridus 60314 13 − Gladiolus dalenii 'arolina Primrose' 14 − Gladiolus ×hybridus 'Beatrice' 15 − Gladiolus ×hybridus 'Glamini'® 16 − Gladiolus ×hybridus 'Vista' 17 − The designed probe for the UFC gene in Gladiolus (RAPiD genomics® LLC; Gainesville, FL, USA;
https://rapid-genomics.com ) resulted in a total of 433 sequences; 161/433 sequences had read hits of the UFC gene with various percentages of coverage. Of these, 34 sequences were chosen, based on the largest length with two sequences per genotype due to the presence of two alleles per gene. These sequences represent the genomic variability of UFC in gladiolus. The sequences were analyzed for gene prediction using the HMM-based gene structure prediction of FGENESH using A. thaliana (Generic) as the specific gene-finding parameter since the gene prediction is optimized for A. thaliana. Results confirmed the presence of UFC exons of a protein and coding sequence. The UFC coding sequences of each gladiolus genotype were analyzed in Genoeious® by pair-alignment with its genomic sequence to determine each exon. After the pair-alignment, the coding sequence was translated and aligned in a multi-alignment process using MUSCLE alignment in the neighbor joining clustering method and CLUSTALW sequencing scheme with UFC proteins of other monocot and dicot species: Ananas comosus, Musa acuminata, Elaeis guineensis, Asparagus officinalis, Arabidopsis thaliana and Glycine max. The UFC gene in Gladiolus ×hybridus was assigned to GhUFC as a label while genotype 14, G. dalenii 'Carolina Primrose', is assigned to GdUFC.There are two alleles of the UFC gene found in gladiolus, designated as A and B. Thus, the genes are designated as GhUFC-A and GhUFC-B. The median number of amino acids of the GhUFC-A protein is 420 amino acids across all gladiolus genotypes, with some genotypes having less than 420 amino acids. One genotype has 451 amino acids which could be due to an insertion, while GdUFC-A also has 420 amino acids. The second allele, the GhUFC-B protein, has a range of 375 to 410 amino acids. GdUFC-B has 286 amino acids with an incomplete protein, missing many amino acids and a stop codon (Figs 2 & 3). Gladiolus genotypes 1 (G. ×hybridus 21213; Table 1) and 15 (G. ×hybridus 'Beatrice'; Table 1) were selected for pair-alignment with the species of comparison (Table 2). Similarities of identity protein sequences and the percentage of GhUFC-A and GhUFC-B occur at range of ~30% to 57% across all species (Table 2). The intron-exon organization of GhUFC-A in G. ×hybridus 'Beatrice' is similar to Elaeis guineensis and Asparagus officinalis in term of exon splicing (Fig. 4) while GhUFC-B in G. ×hybridus 21213 (genotype 1; Table 1) has some similarity with Ananas comosus exons splicing. The remaining genotypes fall into these two configurations of the exon; the configuration shows the location of the conserved domain for UFC gene DUF966, which is found in Arabidopsis and other selected species of comparison. The DUF966 domain has the 92 and 93 amino acids of Ananas comosus. The multi alignment of the UFC protein conserved domain is conserved across species, although it is polymorphic (Fig. 5). With the high identity matching in Gladiolus, genotypes 1 and 15 exhibit a range of ~65% to ~86% across all investigated species for the DUF966 domain of the UFC protein (Table 3). The GhUFC-A allele has a high identity across gladiolus genotypes (Fig. 6), with the polymorphic exception of G. ×hybridus 2231 (genotype 3; Table 1) and G. ×hybridus 3931 (genotype 7). GhUFC-B is also conserved and identical in sequence with G. ×hybridus 20732 (genotype 12), due to missing amino acids.
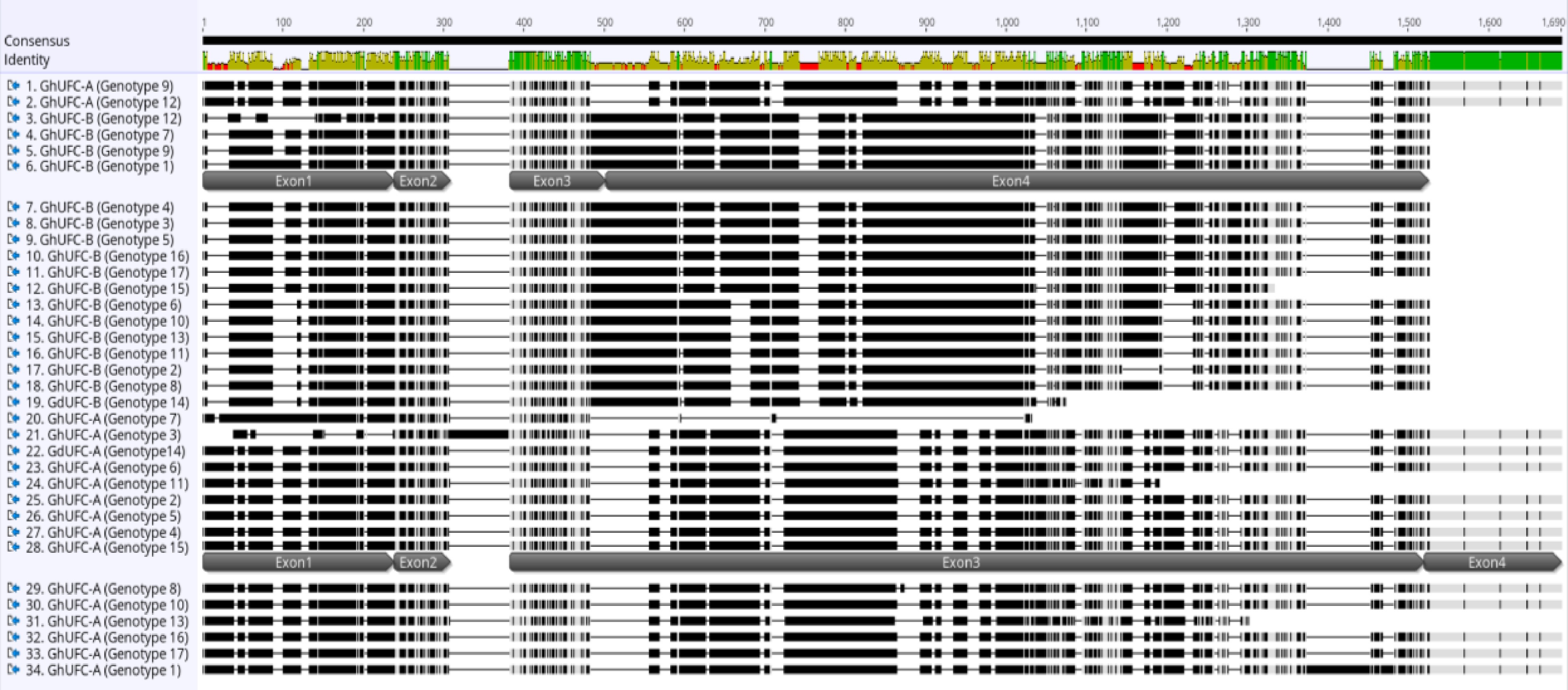
Figure 2.
Multi-alignment of UFC coding sequence in G. ×hybridus (GhUFC) and Gladiolus dalenii (GdUFC), the alignment is for the 17 genotypes, each genotype has 2 alleles, allele A and allele B: GhUFC-A, GdUFC-A, GhUFC-B, GdUFC-B. Both alleles has 4 exons but allele A size is larger in coding sequence than allele B. The alignment shows insertion and missing coding sequences in some genotypes. The multi-alignment is done in MUSCLE pair-alignment using neighbor joining cluster method and CLUSTALW sequencing scheme (Geneious®).
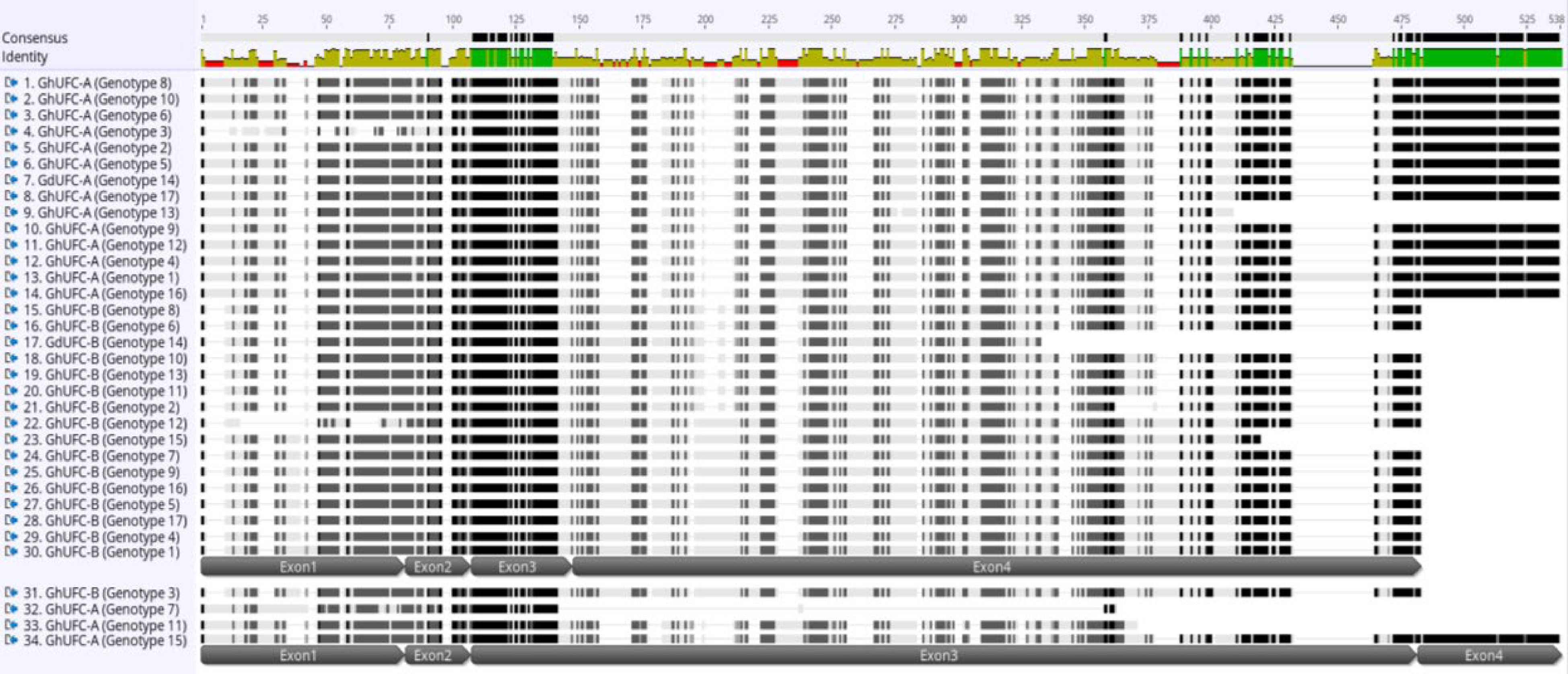
Figure 3.
Multi-alignment of UFC amino acid sequence in Gladiolus ×hybridus (GhUFC) and G. dalenii (GdUFC), the alignment is for the 17 genotypes, each genotype has 2 alleles, allele A and allele B: GhUFC-A, GdUFC-A, GhUFC-B, GdUFC-B. Both alleles has 4 exons but allele A size is larger in amino acid sequence than allele B. The alignment shows insertion and missing amino acid sequences in some genotypes. The alignment identify conserved amino acid sequences (green color). The multi-alignment is done in MUSCLE pair-alignment using neighbor joining cluster method and CLUSTALW sequencing scheme (Geneious®).
Table 2. The identity of amino acid sequences and number (%) of two UFC proteins (GhUFC-A, GhUFC-B) in two Gladiolus (genotypes 1 and 15) in relation to other species (Gene locus/ID) through pair alignment, using MUSCLE alignment for the neighbor joining clustering method and the CLUSTALW sequencing scheme (Geneious®).
Species (Gene locus/ID) Gladiolus ×hybridus genotype Genotype 1 Genotype 15 GhUFC-A (%) GhUFC-B (%) GhUFC-A (%) GhUFC-B (%) Ananas comosus (Aco009327) 189 (33.51) 249 (53.21) 191 (34.35) 231 (52.98) Musa acuminata (GSMUA_Achr5T28540_001) 251 (54.09) 170 (39.35) 250 (57.74) 148 (31.36) Elaeis guineensis (p5.00_sc00099_p0095) 254 (52.05) 172 (40.86) 250 (51.23) 155 (39.85) Asparagus officinalis (evm.model.AsparagusV1_08.3493) 213 (46.61) 148 (35.58) 213 (50.0) 135 (29.87) Arabidopsis thaliana (AT5G10150) 137 (28.54) 129 (29.79) 138 (31.72) 117 (29.32) Glycine max (Glyma.11G193000.1) 181 (33.64) 226 (52.19) 188 (35.67) 207 (52.01) 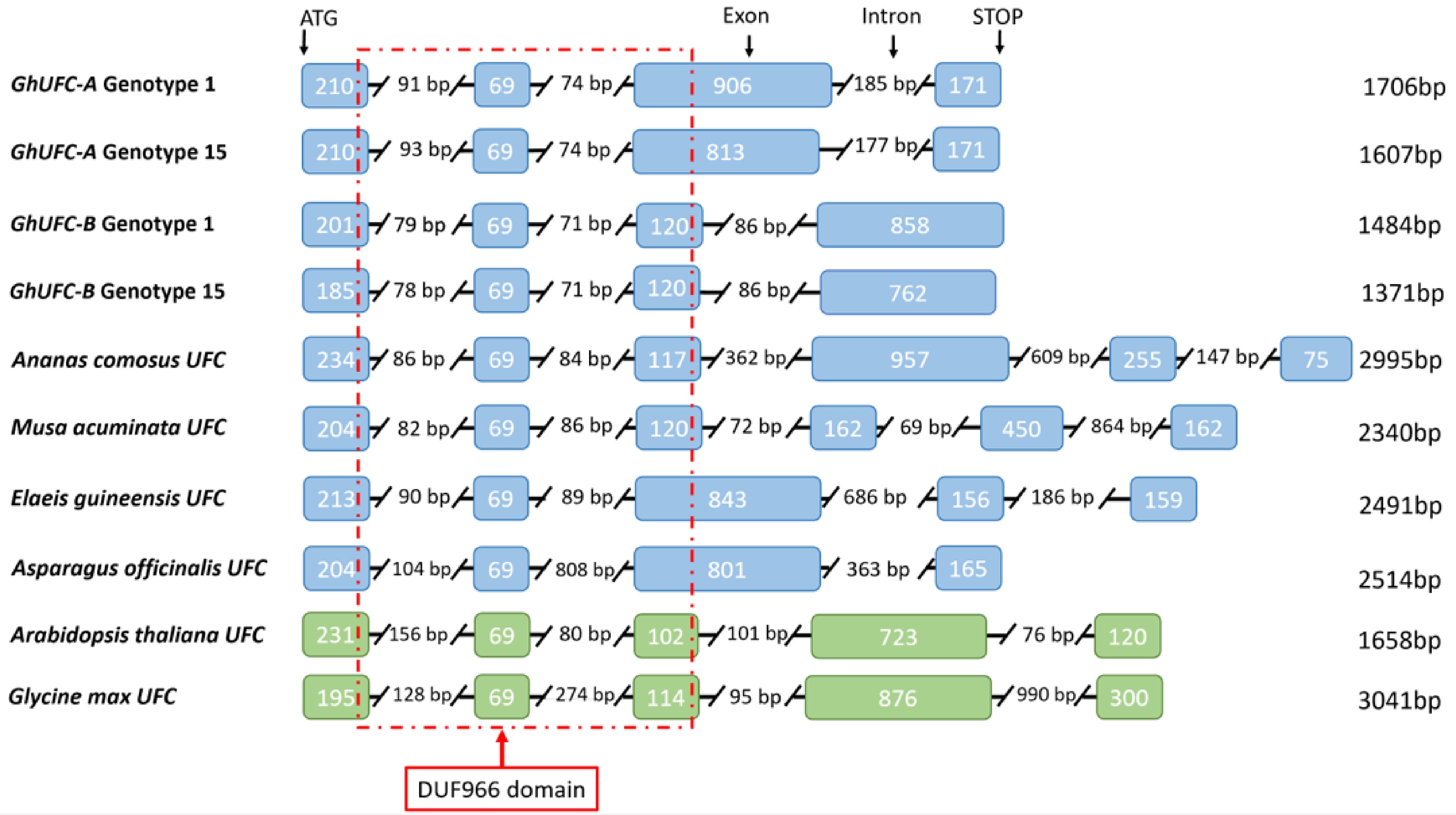
Figure 4.
Intron-exon configuration of the UFC genes in Gladiolus ×hybridus of genotypes 1 and 15 in relation to several species. Monocot species are highlighted in blue: Ananas comosus, Musa acuminata, Elaeis guineensis and Asparagus officinalis. Dicot species highlighted in green for Arabidopsis thaliana and Glycine max. Sequences were aligned based on first exon sequences. Total length of the gene’s coding region is listed on the right of each respected species. The red line represents the conserved domain DUF966 in relation to the location of the domain with exon configuration.

Figure 5.
Alignment of the globular region containing DUF966 domain of UFC proteins from Gladiolus ×hybridus of genotypes 1 and 15, Ananas comosus, Musa acuminata, Elaeis guineensis, Asparagus officinalis, Arabidopsis thaliana and Glycine max. Green coloration shows identical amino acid sequence; yellow color highlights the polymorphisms while red color shows the cytosine amino acid. The conserved domain DUF966 is 92 amino acids.
Table 3. Identity of amino acid sequences, number (%) of UFC proteins in the conserved domain DUF966 in Gladiolus genotypes 1 and 15 in relation to other species (Gene locus/ID) through pair alignment.
Species (Gene locus/ID) Gladiolus × hybridus genotype Genotype 1 Genotype 15 GhUFC-A (%) GhUFC-B (%) GhUFC-A (%) GhUFC-B (%) Ananas comosus (Aco009327) 70 (76.09) 79 (84.95) 79 (84.95) 79 (84.95) Musa acuminata (GSMUA_Achr5T28540_001) 78 (84.78) 71 (78.02) 78 (84.78) 71 (78.02) Elaeis guineensis (p5.00_sc00099_p0095) 79 (85.87) 72 (79.12) 79 (85.87) 72 (79.12) Asparagus officinalis (evm.model.AsparagusV1_08.3493) 75 (82.42) 70 (76.09) 75 (82.42) 70 (76.09) Arabidopsis thaliana (AT5G10150) 61 (66.30) 61 (64.89) 62 (67.39) 61 (64.89) Glycine max (Glyma.11G193000.1) 70 (76.92) 75 (82.42) 70 (76.92) 75 (81.52) 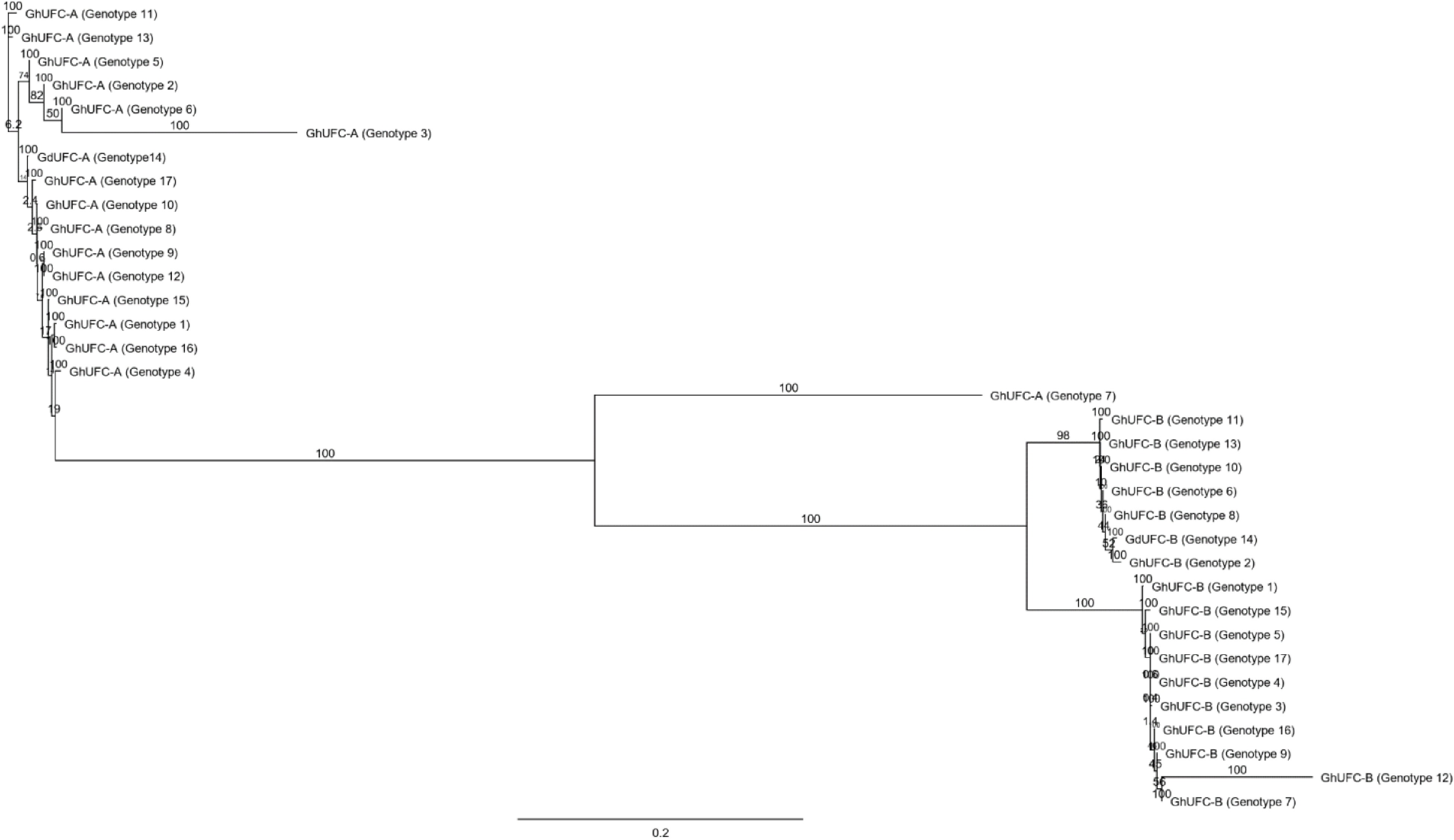
Figure 6.
The phylogenetic tree of all UFC genotypes in Gladiolus; genetic distances were computed using the Tamura-Nei method and are in the units of the number of base substitutions per site. The tree build using the Neighbor-Joining method and the bootstrap test was performed for each tree (500 replicates) and the tree format is organized and ordered with a scale bar of 0.2 (Geneious®).
The FLX gene was identified in Gladiolus in 12/17 genotypes; 11 G. ×hybridus genotypes have GhFLX whereas GdFLX is identified in genotype 14, G. dalenii 'Carolina Primrose' (Table 4). The range of amino acid proteins are from 146 to 254 amino acids, missing the stop codon. Three genotype sequences in G. ×hybridus 2231, 3923, and 'Glamini'® (genotypes 3, 6 and 16, respectively; Table 1) have the longest amino acid chain. FLX is present in many species; in Arabidopsis it belongs to the FLX gene family, FLX, FLOWERING LOCUS C EXPRESSOR-LIKE 1 (FLL1), (FLL2), (FLL3) and (FLL4)[33]. Based on the pair-alignment, GhFLX and GdFLX match FLL1 with as high as 50% amino acid identity (Table 4). The similarities of sequences in identity of gladiolus genotypes range from ~26% to ~65% across all investigated species of the entire FLX protein; the highest identity is the Ananas comosus match with ~65%. The multi-alignment for all FLX indicates that the tested gladiolus genotypes with the longest amino acid sequences lack exons (Fig. 7). A pair-alignment test with Ananas comosus – FLX reveals that GhFLX G. ×hybridus 'Glamini'® (genotype 16; Table 1) lacks two exons and a stop codon (Fig. 8).
Table 4. Number (%) of amino acid sequences of GhFLX protein in Gladiolus genotypes 3 and 6 (genotype 16 is identical to genotype 6) in relation to the other species through pair alignment; similarities of sequences in identity of gladiolus genotypes ranged from ~26% to ~65% across all investigated species of the whole FLX protein; alignment is done in MUSCLE, using the neighbor joining clustering method and the CLUSTALW sequencing scheme (Geneious®).
Species (Accession no.) Gladiolus × hybridus genotype GhFLX Genotype 3 (%) GhFLX Genotype 6 and 16 (%) Ananas comosus (XP_020095672.1) 180 (64.98%) 177 (64.60%) Musa acuminata (XP_009420070.1) 122 (48.03%) 122 (48.03%) Elaeis guineensis (XP_010922618.1) 132 (50.00%) 131 (49.62%) Arabidopsis thaliana – FLX (NP_001154541.1) 82 (30.48%) 82 (30.48%) Arabidopsis thaliana – FLL1 (NP_566492.1) 135 (50.00%) 135 (50.00%) Arabidopsis thaliana – FLL2 (NP_001320766.1) 96 (36.09%) 94 (35.34%) Arabidopsis thaliana – FLL3 (NP_564678.1) 104 (34.67%) 104 (34.67%) Arabidopsis thaliana – FLL4 (NP_001119474.1) 67 (26.38%) 68 (26.77%) Glycine max (Glyma.15g269300) 156 (57.14%) 155 (56.78%) 
Figure 7.
Multi-alignment of FLX amino acid sequence in Gladiolus ×hybridus (GhFLX) with other species; Arabidopsis thaliana, Ananas comosus, Elaeis guineensis, Musa acuminata and Glycine max. The alignment is for the three gladiolus genotypes (3, 6 and 16) each genotype has 3 exons. The alignment shows missing amino acid sequences in gladiolus genotypes 3, 6 and 16 as amino acid sequences does not have a stop codon. The alignment identifies conserved amino acid sequences (green color). Note: Arabidopsis thaliana – FLL4 is a functional protein which has two exons only (Lee and Amasino, 2013). The multi-alignment is done in MUSCLE pair-alignment using neighbor joining cluster method and CLUSTALW sequencing scheme (Geneious®).
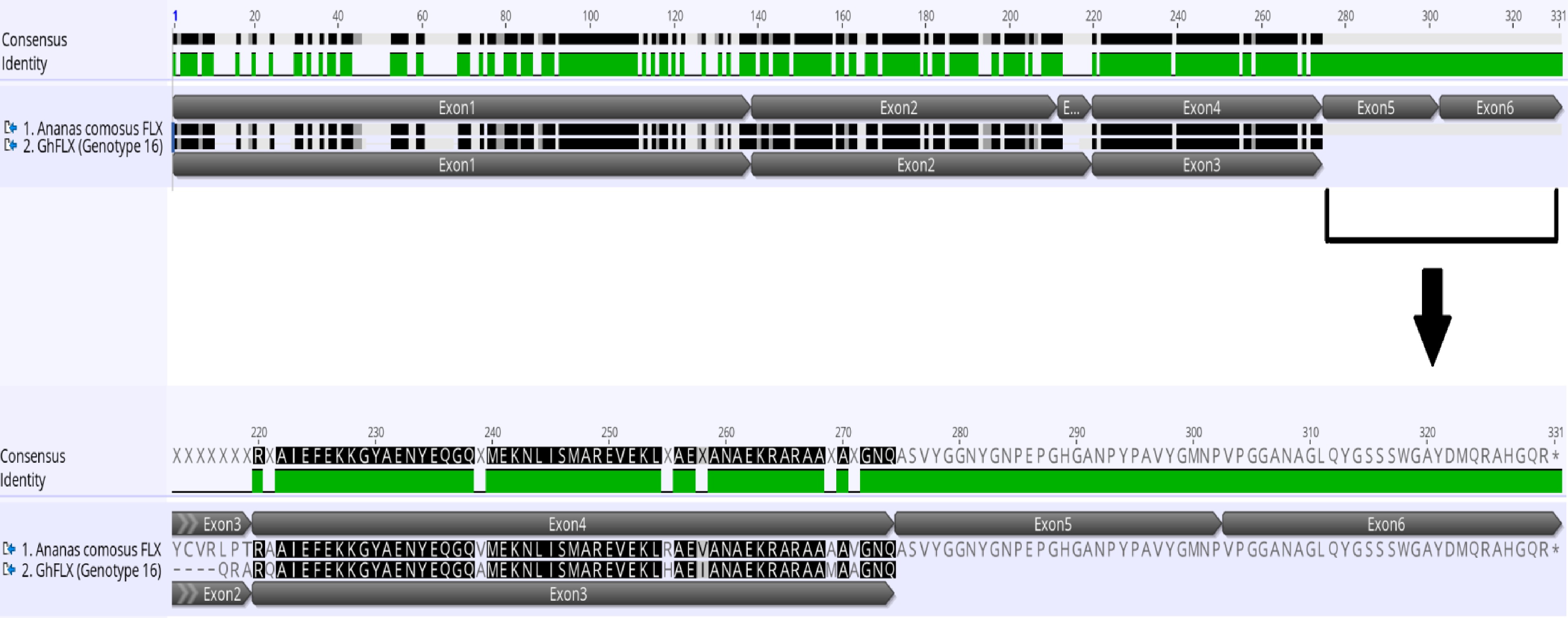
Figure 8.
Pair alignment of FLX protein in Ananas comosus and Gladiolus ×hybridus genotype 16 showing the complete FLX protein in Ananas comosus with five exons while incomplete FLX protein in Gladiolus ×hybridus (GhFLX) which lacks the remaining two exons and stop codon. The alignment is done in MUSCLE pair-alignment using neighbor joining cluster method and CLUSTALW sequencing scheme (Geneious®).
-
The presence of a putative UFC gene in gladiolus is confirmed with two alleles, GhUFC-A and GhUFC-B. It is highly possible that allelic number is due to tetraploidy in cultivated gladioli (2n = 4x = 60)[46−48]; ploidy levels of G. dalenii have not been reported[46]. The cultivated G. ×hybridus are interspecific hybrids[49]. Therefore, the presence of different alleles would be expected in the diverse array of genotypes included in this study: 13 are from University of Minnesota gladiolus breeding program (interspecific hybrids) and four are commercial gladiolus cultivars with unknown ancestry and relatedness (Table 1). The GhUFC-A gene has ~50% identity with Musa acuminata, Elaeis guineensis and Asparagus officinalis (Table 2). The splicing of Elaeis guineensis and Asparagus officinalis exons is similar to GhUFC-A (Fig. 4). GhUFC-B gene splicing in the first 4 exons is similar to Ananas comosus, Arabidopsis thaliana and Glycine max UFC gene splicing (Fig. 4). These divergences in splicing of the UFC gene support UFC presence in gladiolus with two alleles[46]. Further tests should be done to identify whether or not UFC is also present in diploid gladiolus species, such as G. murielae, G. tristis and G. carneus[50].
The UFC gene is responsive to vernalization by lowering expression alongside FLC and DFC in Arabidopsis thaliana, as all these genes are in the cluster of vernalization stimulus region[37]. FLC is a floral repressor, the overexpression of FLC results in a delay in flowering[51], while overexpression of UFC does not result in altering the flowering time[37]. Thus, UFC is adjacent to FLC, both are repressed by vernalization, yet UFC does not show any influence on flowering time. This was observed herein since the genotypes in this study include both RGC-1, which are early flowering gladiolus able to reach flowering in the first year from seed and the classical later-flowering gladiolus which requires 3-5+ years to flower from seed. The multi-alignment of UFC protein in RGC-1 genotypes does not show any difference from non-RGC-1 genotypes. Thus, the UFC gene most likely isn't involved in flowering, at least directly, which was proven in a UFC study in A. thaliana[38]. The main differences represent the differences between alleles of UFC-A and UFC-B, regardless of the gladiolus genotypes tested herein (Fig. 6).
During the winter cold period, vernalization suppress both FLC and UFC expression[37], which allows floral gene integrators to promote flowering. The hypothesis would be that, after vernalization and flowering, UFC protein involvement is in embryogenesis and root initiation such that growth and branching occur in the spring season (since it doesn't occur in the winter season). This could explain how FLC and UFC are both negatively responsive to vernalization stimuli in the cluster genes area, while upstream of UFC is not responsive to vernalization[37].
The identification of FLX in gladiolus raises the question whether gladiolus follows the Arabidopsis dicot model of the flowering pathway. In the winter annual, A. thaliana, flowering is promoted after vernalization, which suppresses the floral suppressor FLC that is upregulated by FRI through activation of FRI complex of (FRI, FRL1, FRL2, FES1 and SUF4) proteins in addition to FLX protein. FLX was proven to provide transcriptional activity for the FRI complex[11]. A loss of function of FLX in A. thaliana resulted in early flowering phenotypes[42], which indicates the clear role of FLX in flowering. The role of FLX in gladiolus has not been tested, particularly in RGC-1 genotypes and pedigrees. Thus, FLX upregulation of FRI in gladiolus would be a rational approach. However, FRI was not detected in gladiolus, using the primer design of A. thaliana FRI (At4g00650) because the FRI gene has not been previously detected in any monocotyledon species. Thus, the primer used from A. thaliana did not detect FRI in any of the 17 gladiolus genotypes[45]. Our results with gladiolus provide additional data in support of these previous results for FRI in monocots. In addition, VRN2, the repressor of flowering in cereals and A. thaliana was not detected in gladiolus, using the primer design of Triticum monococum, T. durum and Hordeum vulgare (Aljaser[45]; Supplemental Table S1, Supplemental Fig. S1 & S2). This is not conclusive evidence as the genetic similarities between A. thaliana and Gladiolus are low, given that GhUFC-A is ~32% and GhFLX is 50% identical to A. thaliana genes. Therefore, there could be an FRI gene in gladiolus but this would require better primer design to locate the gene because the presence of GhFLX might indicate in the presence of other flowering repressor genes as FRI protein upregulates FLX in A. thaliana and is part of the flowering pathway[11]. In addition, Musa acuminata, Elaeis guineensis and Ananas comosus are all monocots and tropical species which have FLX and SUF4 genes as part of the FRI complex[51]. This indicates the presence of some of the FRI complex components while a lack of identification of FRI gene itself creates divergent possibilities: a) either there are FRI and FLC genes in these species or b) a lack of these genes and the presence of SUF4 and FLX genes have other unknown flowering pathway purposes. Since GhFLX and GdFLX have similarities to FLL1, reaching up to 50% identity in amino acids, FLX gene is part of the gene family, FLL1-FLL4[11,33]. While the role of FLL1 in flowering pathway is not proven, FLX and FLL4 are the most crucial genes in control of flowering time in Arabidopsis[33]. Additionally, the relationship between UFC and FLX indicates that a mutation in FLX influences UFC expression, e.g., the flx mutant in A. thaliana[42].
The next step in this research would be to identify the UFC gene in diploid gladiolus species to determine if the allele is similar to GhUFC-A, GhUFC-B or a third allele. Use of diploids would simplify the study to determine the function of UFC protein in gladiolus by silencing and knocking out the gene. Locating the physical location of the UFC gene in Gladiolus will help in testing if there are other UFC genes in gladiolus as part of a UFC gene family, since the first discovered UFC gene (At5g10150) in A. thaliana is located in the cluster genes UFC, FLC and DFC on chromosome 5[37]. UFC is also designated SOK2 and the other UFC genes are grouped in SOK gene family such as SOK1 (At1g05577), SOK3 (At2g28150), SOK4 (At3g46110) and SOK5 (At5g59790)[39].
Identifying the FLX gene in Eurasian species of Gladiolus, particularly G. italicus, G. imbricatus and G. communis, would be informative since these winter-hardy, perennial species grow in temperate habitats that require vernalization to break corm dormancy in the winter season[29,52,53]. Conversely, identifying FLX in subtropical gladiolus species, such as G. crassifolius, G. laxiflorus and G. atropurpureus[54], would allow comparison of FLX among these different habitats to further support the influence in FLX in the gladiolus flowering pathway. Furthermore, the use of transgene silencing of FLX in gladiolus would determine whether or not FLX influences the production of a rapid flowering phenotype gladiolus (RGC-1), as was reported in the loss of flx function in A. thaliana[42]. In conclusion, the discovery of UFC and FLX genes in gladiolus provides insight into understanding flowering and vernalization responses in ornamental, monocot geophytes.
-
Rapid generation cycling is a powerful tool that can be implemented to reduce the juvenility period in perennial crops such as gladioli and are being applied in the breeding program crop ideotype. Although it is possible to annualize a perennial crop through genetically modifying the flowering pathway by overexpression a positive flowering regulator or inserting blocker of flowering suppressor, such biotechnological methods require regulatory approval. Therefore, conventional breeding methods for early flowering are widely accepted and do not require regulation for cultivar release.
The search for FLC and its regulatory genes in gladiolus is a step to uncover the flowering pathway in geophytes. To uncover if FLC is present in Gladiolus, we searched for linked genes with FLC. In Arabidopsis, FLC is adjacent to two genes, UFC and DFC, both of which are downregulated by vernalization. The discovery of UFC in gladiolus, as well FLX (which upregulates FRI), is crucial to establish the flowering pathway. These may be early indicators of the presence of FLC homologue in gladiolus. Discovery of both genes are important to understand the flowering mechanism and genes in the flowering pathway to aid in breeding and selection of early flowering gladioli from seed or corms.
-
The 17 gladiolus genotypes used in this study (Table 1) were chosen to represent a range of diversity within cultivated gladioli (Gladiolus ×hybridus, G. dalenii) which includes nine genotypes of Rapid Generation Cycling-1 (RGC-1; ones that flower in < 1 year from seed; [43−45]) and eight genotypes Non-RGC genotypes (that require 2 to 5 years to flower from seed). Fourteen of these genotypes are interspecific parents and hybrids created by the University of Minnesota Gladiolus Breeding Program, while three additional genotypes are commercial cultivars. Originally, 'Carolina Primrose' was introduced as G. primulinus. All gladiolus pedigrees used in this experiment are published[43−45] and commercial cultivars 'Beatrice' (an open-pollinated seedlings of unknown origin, occurring in a private garden, Brookfield, Vermont, in 2003). 'Beatrice' was selected for its winter hardiness, surviving in USDA Z3). 'Glamini'® a series of shorter in height than tall summer gladiolus which blooms early and has a range of flowering colors[55]. One genotype 'Carolina Primrose' is an heirloom gladiolus, bred in 1908, with yellow flowers, collected at an old homesite in North Carolina and derived from G. dalenii (Table 1)[56].
Greenhouse environment
-
Mature gladiolus corms (competent to flower) were planted into 1,679.776 cm2 square, deep pots (Belden Plastics, St. Paul. MN, USA) in week 23 (2017) and grown for 18 weeks. Containers were filled with SS#8-F2-RSi potting soil, 'SunGrow' (Sun Gro Horticulture, Agawam. MA, USA). The corms were grown in a long day photoperiod (0800 – 1600 HR supplied by 400-W high-pressure sodium lamps + 2200 to 0200 h night interruption, > 150 µmol m−2 sec−1) at a minimum setpoint of 18 oC (day/night), 70%–80% relative humidity, with irrigation accomplished using constant liquid feed (CLF) of 125 ppm N from water-soluble 20N–4.4P–16.6K (Scotts, Marysville, OH, USA) and deionized water on weekends. Standard fungicide drenches and insecticides were applied either monthly or as needed, respectively.
DNA extraction and probe design
-
Newly expanded gladiolus leaves were harvested, placed in an ice box and sent to RAPiD Genomics® LLC (Gainesville, FL, USA;
http://rapid-genomics.com/home/ ) for DNA extraction, probe design, sequencing and computable analysis. Probe designs for the UFC gene were based on banana, Musa acuminata subsp. malaccensis accession XM_009383889, from the GenBank Nucleotide Core[57] and oil palm, Elaeis guineensis accession XM_010920607.2[58]. Probe design for FLX gene were based on oil palm, Elaeis guineensis accession XM_010924316.2[59] and date palm, Phoenix dactylifera accession XM_008801571.2[60]. The designed probe for UFC able to capture the locus in Musa acuminata and Elaeis guineensis by capturing the 2x coverage of the UFC exons in Musa acuminata and Elaeis guineensis, while the FLX probe captures the locus in Elaeis guineensis and Phoenix dactylifera. Probes are amplified in short reads of UFC and FLX genes in gladiolus. The reads are sequenced through the Illumina dye sequencing technique, the raw data is demultiplexed using Illuminas BCLtofastq and then assembled using MaSuRCA® software[61], creating full assembly sequence scaffolds. Afterwards, read mapping using the reference genome and blasting to filter all assembled sequences for hits to the sequences, provided the probes design (UFC and FLX). Then, count read numbers for each assembled sequence passed the filtering, accruing the final sequences for genetic analysis. Gene sequences are currently being deposited into GenBank.Genetic analysis
-
The sequence data for the UFC and FLX genes used in this study were found in the genetic sequence database under the following accession/ID numbers: Ananas comosus (Aco009327) UFC gene is from the Pineapple Genomics Database[62]; Musa acuminata (GSMUA_Achr5T28540_001) UFC from the Banana Genome Hub[63]; Elaeis guineensis (p5.00_sc00099_p0095) UFC from the Malaysian Oil Palm Genome Programme[64]; Asparagus officinalis (evm.model.AsparagusV1_08.3493) UFC from the Asparagus Genome Project[65]; Arabidopsis thaliana (At5g10150) UFC from The Arabidopsis Information Resource (TAIR)[66]; Glycine max (Glyma.11G193000.1) UFC from the SoyBase[67]. The FLX protein was from the GenBank Nucleotide Core with accession numbers as follows: Ananas comosus (XP_020095672.1)[68], Musa acuminata (XP_009420070.1)[69], Elaeis guineensis (XP_010922618.1)[70], Arabidopsis thaliana – FLX (NP_001154541.1)[71], Arabidopsis thaliana – FLL1 (NP_566492.1)[72], Arabidopsis thaliana – FLL2 (NP_001320766.1)[73], Arabidopsis thaliana – FLL3 (NP_564678.1)[74], Arabidopsis thaliana – FLL4 (NP_001119474.1)[75] and Glycine max (Glyma.15g269300) FLX from the SoyBase[67].
Generated sequences were analyzed for gene prediction using the HMM-based gene structure prediction of FGENESH with Arabidopsis thaliana (Generic) as the specific gene-finding parameter. The predicted genes for UFC and FLX were analyzed in multi-alignment using Geneious© software (Biomatters Ltd, Auckland, NZ). The UFC protein sequences of gladiolus were analyzed for conserved domains using the Protein Homology/analogy Recognition Engine V 2.0 (Phyre2) browser[76]. Then the alignment of conserved domain was formed to compare the matching and differences in each amino acid in the sequences of gladiolus and the other comparison species. A phylogenetic tree of all UFC genotypes of Gladiolus was formed by computing the distances using the Tamura-Nei method and were in the units of the number of base substitutions per site. The tree building used the Neighbor-Joining method and a bootstrap test was performed for each tree (500 replicates).
Funding for this research was supported by grants from the Minnesota Agricultural Experiment Station (MAES21-0045) and the Minnesota Gladiolus Society which funded sequencing and greenhouse charges. The Kuwaiti Government funded a Graduate (Ph.D.) Scholarship for Jaser Aljaser to conduct this research as part of his Ph.D. dissertation.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 The primer design for VRN2 gene in the tested cereal and gladiolus samples.
- Supplemental Fig. S1 Gel electrophoresis image of PCR VRN2 gene bands of 1500-2000 bp at 52oC annealing temperature. Rapid Generation Genotype (RGC) does not require vernalization to flower. Gladiolus palustris is wild Eurasian species which require vernalization to flower, as well as all the cereal species require vernalization to flower. Both gladiolus samples do not show presenceVRN2gene fragment amplification, although the primer design was based on the consensus sequence of VRN2 in Triticum monococum, Triticum durum, Hordeum vulgare and Aegilops tauschii. The failure to amplify VRN2 gene in gladiolus genotypes could indicate that gladiolus may have VRN2 gene homolog, but the gene sequence varies greatly to cereals.
- Supplemental Fig. S2 Gel electrophoresis image PCR VRN2 gene band of 1500-2000 bp at 51℃ annealing temperature. Arranged from left wells to right; 1. High Ranger Plus 100 bp DNA ladder, 2.Triticum monococum PI 272561 (control), 3.Gladiolus × hybridus RGC genotype 2231, 4.Gladiolus × hybridus RGC genotype 1531151, 5.Gladiolus × hybridus RGC genotype 16883, 6.Gladiolus murielae, 7.Gladiolus cardinalis, 8.Gladiolus flanganii, 9.Gladiolus papilio, 10.Freesia alba, 11.Iris sibirica, 12.Hippeastrum sp . 13.Lilium longiflorum ‘Nelly White’ , 14.Tulipa sp. ‘Queen of Night’, 15.Crocosmia × crocosmiiflora ‘Mistral', 16.Crocus sativus, 17.Cypella coelestis, 18.Allium cepa ‘Flat of Italy’, 19.Hyacinth orientalis ‘Purple voice’ 20.Tecophilaea cyanocrocus. Thus, the designed primer was able to amplify VRN2 band in Triticum monococum while no band was amplified in any of geophytic genotypes, the designed primer was based on the consensus sequence of VRN2 in Triticum monococum, Triticum durum, Hordeum vulgare and Aegilops tauschii.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Aljaser JA, Anderson NO, Noyszewski A. 2022. Discovery of UPSTREAM OF FLOWERING LOCUS C (UFC) and FLOWERING LOCUS C EXPRESSOR (FLX) in Gladiolus ×hybridus, G. dalenii. Ornamental Plant Research 2:13 doi: 10.48130/OPR-2022-0013 

Discovery of UPSTREAM OF FLOWERING LOCUS C (UFC) and FLOWERING LOCUS C EXPRESSOR (FLX) in Gladiolus ×hybridus, G. dalenii
- Received: 20 April 2022
- Accepted: 17 August 2022
- Published online: 29 August 2022
Abstract: The lack of identified flowering genes in ornamental geophytic crops, such as Gladiolus, is critical to further genetic research. The UPSTREAM OF FLOWERING LOCUS C (UFC) gene is adjacent to FLOWERING LOCUS C (FLC) which is a floral repressor; FLC EXPRESSOR (FLX) upregulates FRIGIDA which upregulates FLC expression. The purpose of this research was to determine whether two flowering genes exist in Gladiolus ×hybridus and G. dalenii: UFC and FLX. Seventeen early flowering and commercial cultivars possess the UFC gene with four exons in two allelic forms. The sequenced UFC gene, when translated into its amino acid sequence and set in pair-alignment to other species, has < 57% in amino acid identity to Musa acuminata. The FLX gene in gladiolus has 3/5 (60%) exons in common with Ananas comosus; pair-alignment of the exons has ~65% identity of FLX to A. comosus. The UFC protein consists of a conserved domain, DUF966, which is higher in identity (86%) and pair-alignment with Elaeis guineensis. The two newly-discovered genes in gladiolus, UFC and FLX, provide insight into the flowering mechanism, flowering pathway genes, and vernalization response.
-
Key words:
- Gladiolus xhybridus /
- Gladiolus dalenii /
- flowering genes /
- Rapid generation cycling