








-
Absidia, a genus belonging to the Cunninghamellaceae family, encompasses mesophilic organisms that thrive under moderate temperature conditions. These fungi exhibit optimal growth within a temperature range of 25 to 34 °C[1−7]. Previously, Absidia also included thermophilic species and mycoparasites[4,5,7]. However, advancements in molecular tools have brought more stability to the classification of Absidia sensu lato[4,5,7]. Phylogenetic analyses indicated that these genera did not belong to Absidia, but resided in Lichtheimiaceae instead. Hence, species of Absidia sensu lato were segregated into three distinct genera, Absidia sensu stricto, Lichtheimia and Lentamyces, based on phylogeny, physiology and morphology, which includes features such as the zygospores and their appendages[4,5,7]. Thermophilic species have since been reclassified under Lichtheimia, while mycoparasites now belong to Lentamyces[4,5,7].
Absidia species are usually isolated as saprobes in soil, but also on dung, and other organic debris[8−12]. The genus is ubiquitous with a broad distribution. They are found in temperate, tropical and subtropical countries such as Brazil, China, Thailand and South Korea[8−13]. Absidia species usually produce sporangiophores that are erect, arising singly or in whorls, with subsporangial septa (one or more). Sporangiophores are usually produced in whorls and bear a terminal columellate and apophysate pyriform sporangium. The columellae usually have apical projections distinct from other genera within the Cunninghamellaceae, and zygospores have finger-like appendages, usually produced on equal suspensors[9,11,13].
The taxon Absidia has experienced a rapid influx of new species in the last few years. Various novel taxa have been identified in Brazil, China, Thailand and South Korea[8−13]. Species are usually delineated using the ITS and LSU genetic markers. Some studies also include protein-coding genes such as actin (ACT) and translation elongation factor (EF-1α), which increases the reliability of the phylogenies[9,10]. However, it is well known, that obtaining the ITS rDNA sequence data and protein coding genes in this genus is extremely difficult and often cloning is required to obtain good quality DNA sequences[6,9].
In an attempt to explore the diversity of zygosporic fungi in northern Thailand, soil samples were collected from Nan Province. During the sampling process, an Absidia strain was isolated. We characterized this new isolate based on molecular phylogenetic analyses, and morphophysiological characteristics. The results revealed that the isolated strains differed significantly from known Absidia species. Consequently, we introduce this newly isolated strain as a novel species within Absidia, accompanied by a taxonomic diagnosis and photoplates. By characterizing and introducing this strain as a new species, we expand the taxonomic knowledge of the genus, and broaden our understanding of the evolutionary and ecological dynamics within this group of fungi. Furthermore, the identification of new species adds to the overall knowledge of fungal diversity, ultimately contributing to broader scientific research.
-
Soil samples were collected from Nan province, Thailand in January 2020. During this time, the average temperature in Thailand ranges from 24–32 °C. Organic debris were manually removed from the surface of the soil prior to sampling. Sterile shovels and spoons were used to dig the surface layer (around 1–5 cm) and collect the soil. The samples were transferred to zip lock bags and kept under ice until it was possible to store it at 4 °C.
The dilution plating method was used to isolate the fungus[14]. The sample was diluted to a ratio of 1:5 and 1:10 with sterile distilled water. The mixture was then shaken for 2 h at 25 °C. Subsequently, 100 µL of the supernatant was transferred to fresh media supplemented with chloramphenicol. The media used for inoculation were malt extract agar (MEA) (Himedia™), PDA and yeast malt extract (YMA) (yeast extract: 3 g; malt extract: 3 g; peptone: 5 g; glucose: 10 g; agar: 15 g; distilled water: 1 L). A flame sterilized glass spreader was used to spread the supernatant on the media. Once completed, the agar plates were wrapped in parafilm and kept at 20 °C. The inoculated plates were checked daily for fungal growth. Once growth (3 d post inoculation) was observed, fungal tips were transferred to fresh agar plates to acquire axenic cultures.
Morphological characters were observed using a compound microscope (Nikon Eclipse Ni) and images of fungal structures were captured using a Nikon DS-RI2 digital camera. The fungus was preserved in 15% glycerol and water. The ex-type culture was deposited in the Mae Fah Luang University Culture Collection (MFLUCC) and an inactive dried culture (on MEA and 2.5% glycerol) was deposited as the holotype in Mae Fah Luang University (MFLU) Herbarium, Chiang Rai, Thailand. The new taxon was registered in Index Fungorum (2023).
DNA extraction and PCR amplification
-
Mature fungal cultures (grown for 3–5 d in MEA at 25 °C) were used for genomic DNA extraction. The total genomic DNA was extracted using the G-spin™ Total DNA Extraction Kit (Intron Biotechnology, South Korea) following the manufacturer’s instructions. The partial fragments of ITS and LSU were amplified using polymerase chain reaction (PCR) using the primers ITS4/5 and LR0R/LR7, respectively[15,16]. The PCR conditions for both ITS and LSU were as follows, initial heat treatment for 5 min at 94 °C, 30 cycles with a denaturation step at 94 °C for 30 s, annealing at 52 °C for 45 s and an elongation step of 90 s at 72 °C and a final elongation period of 7 min at 72 °C.
The PCR products were purified using gel purification and subsequently with MEGAquick spin plus fragment DNA purification kit (Intron Biotechnology, South Korea). Sequencing was performed using an Applied Biosystems 3130XLDNA analyzer (Bionics, South Korea).
Phylogenetic analyses
-
The raw chromatograms were viewed using BioEdit to check the quality of the sequences and to remove ambiguous bases at both ends. Each sequence was subjected to a Blast search in GenBank to find the closest taxa and check for chimera and/or contamination. The forward and reverse reads were merged using SeqMan. The taxon sampling aimed to cover the genetic diversity of the genus. DNA sequence data were extracted from GenBank to build the dataset for phylogenetic analyses (Table 1). Individual ITS and LSU matrices were built and aligned using MAFFT on the online platform (
https://mafft.cbrc.jp/alignment/server/ )[17]. The alignment matrix was then trimmed to remove ambiguous bases using TrimAl 1.2[18].Table 1. Data used for phylogenetic analysis in this study and their corresponding GenBank accession numbers. Type species are denoted by T. Sequences derived in this study are shown in bold.
Species name Voucher no. GenBank accession number ITS LSU Absidia abundans CGMCC.3.16255T NR_182590 ON074683 Absidia aguabelensis URM 8213T MW763074 MW762874 Absidia alpina CGMCC 3.16104 OL678133 Absidia ampullacea CGMCC 3.16054 MZ354138 MZ350132 Absidia anomala CBS 125.68T NR_103626 NG_058562 Absidia bonitoensis URM 7889T MN977786 MN977805 Absidia brunnea CGMCC.3.16055 MZ354139 MZ350133 Absidia caatinguensis URM7156T NR_154704 NG_058582 Absidia californica CBS 126.68T − NG_056998 Absidia caerulea NRRLA9483 − − Absidia caerulea CBS 104.08 JN205811 MH866107 Absidia cornuta URM 6100T NR_172976 MN625255 Absidia cuneospora CBS 102.59 JN205819 JN206579 Absidia cylindrospora var. cylindrospora CBS 100.08 JN205822 JN206588 Absidia cylindrospora var. nigra CBS 127.68T − NG_058560 Absidia cylindrospora var. rhizomorpha CBS 153.63T − NG_058563 Absidia edaphica MFLU 20-0415 − MT393986 Absidia edaphica MFLU 20-0416T MT396372 MT393987 Absidia fusca CBS 102.35T NR_103625 NG_058552 Absidia glauca CBS 101.08T NR_111658 MH866105 Absidia heterospora SHTH021 JN942683 JN982936 Absidia heterospora CBS 101.29T − NG_058564 Absidia jindoensis CNUFC-PTI1-2 MF926623 MF926617 Absidia jindoensis CNUFC-PTI1-1T MF926622 MF926616 Absidia koreana EML-IFS45-2 KR030063 KR030057 Absidia koreana EML-IFS45-1T KR030062 KR030056 Absidia macrospora FSU4746 AY944882 − Absidia macrospora CBS 697.68T − NG_058549 Absidia ovalispora HMAS 249158 MW264133 MW264074 Absidia ovalispora CGMCC 3.16018T MW264071 MW264130 Absidia panacisoli SYPF 7183 MF522181 MF522180 Absidia panacisoli CBS 140959T NR_159563 NG_063948 Absidia pseudocylindrospora EML-FSDY6-2 KU923817 KU923814 Absidia pseudocylindrospora CBS 100.62T NR_145276 NG_058561 Absidia psychrophilia FSU4745 AY944874 EU736306 Absidia repens FSU 4726 EU484288 − Absidia repens NRRL1336 − AF113448 Absidia repens CBS 115583T NR_103624 NG_058551 Absidia soli MFLU 20-0413 MT396371 MT393985 Absidia soli MFLU 20-0414T MT396373 MT393988 Absidia spinosa FSU551 AY944887 EU736307 Absidia spinosa FSU552 AY944888 EU736308 Absidia spinosa var. biappendiculata CBS 187.64 − MH870040 Absidia stercoraria EML-DG8-2 KU168829 KT921999 Absidia stercoraria EML-DG8-1T KU168828 KT921998 Absidia zygospora RSPG 214 KC478527 - Absidia zygospora ANG28 DQ914420 - Absidia zygospora MFLUCC 23–0061T OR104965 OR104992 Absidia chinensis CGMCC.3.16056 MZ354140 MZ350134 Absidia cinerea CGMCC.3.16062 MZ354146 MZ350140 Absidia digitula CGMCC 3.16058 MZ354142 MZ350136 Absidia globospora CGMCC.3.16031 MW671537 MW671544 Absidia healeyae UoMAU1 MT436028 MT436027 Absidia jiangxiensis CGMCC 3.16105 OL678134 - Absidia lobata CGMCC 3.16256 ON074690 ON074679 Absidia medulla CGMCC 3.16034 MW671542 MW671549 Absidia montepascoalis CNUFC HT19001T MW473494 MW561560 Absidia multispora URM 8210 MN953780 MN953782 Absidia oblongispora CGMCC 3.16061 MZ354145 MZ350139 Absidia pararepens CCF 6351 MT193670 MT192307 Absidia pernambucoensis URM 7219 MN635568 MN635569 Absidia purpurea CGMCC 3.16106 OL678135 - Absidia radiata CGMCC 3.16257 ON074698 ON074684 Absidia saloaensis URM 8209T MN953781 MN953783 Absidia saloaensis DXL2020 MN953781 MN953783 Absidia sichuanensis CGMCC 3.16258T NR_182589 ON074688 Absidia sympodialis CGMCC 3.16064 MZ354148 MZ350142 Absidia terrestris FMR 15024 LT795004 LT795593 Absidia terrestris FMR 14989T LT795003 LT795005 Absidia turgida CGMCC.3.16032 MW671540 MW671547 Absidia varians CGMCC.3.16065 MZ354149 MZ350143 Absidia virescens CGMCC.3.16066 MZ354150 MZ350144 Absidia xinjiangensis CGMCC.3.16107 OL678136 Absidia yunnanensis CGMCC 3.16259T NR_182591 NG_149054 Absidia zonata CGMCC.3.16033 MW671541 MW671548 Chlamydoabsidia padenii NRRL 2977T − AF113453 Chlamydoabsidia padenii CBS 172.67T NR_153872 JN206586 Cunninghamella bainieri FSU319 − EU736313 Cunninghamella homothallica CBS 168.53 MH857147 NG_058833 Cunninghamella phaeospora CBS 692.68 AF254934 NG_058812 Halteromyces radiatus NRRL6197 − AF157192 Halteromyces radiatus CBS 162.75 NR_145293 NG_057938 Maximum likelihood (ML) phylograms were inferred using RAxML-NG 1.1.0 and IQ-tree in the online CIPRES Portal (
www.phylo.org/portal2 ) andhttp://iqtree.cibiv.univie.ac.at/ respectively with bootstrap support obtained from 1,000 pseudo replicates[19,20]. Bayesian inference (BI) analysis was also performed on the online CIPRES Portal (www.phylo.org/portal2 ) using MrBayes on XSEDE 3.2.7a[21]. The Bayesian tree was built by running four simultaneous chains of 2 × 106 generations and a sampling frequency of 100. The burn-in phase was estimated using Tracer software. The first 1,000 trees represented the burn-in and was herein discarded. Convergence was declared when the average standard deviation of split frequencies reached 0.01 or below. The substitution models of molecular evolution were estimated for each genetic marker using jModelTest2 on XSEDE in the CIPRES Portal. The best fit model for both the ITS and LSU was GTR+G+I under the Akaike information criterion. -
The ITS and LSU concatenated dataset comprised 82 sequences including three outgroup taxa. The final trimmed alignment consisted of 1,401 sites: ITS: 449, LSU: 952. The final matrix contained 790 distinct alignment patterns and the likelihood of the best scoring ML tree was −21,271.48146. The topologies obtained from the ML and BI analyses were congruent, and similar to previous studies. In both phylogenetic analyses our isolate grouped with two unclassified strains namely Absidia sp. RSPG 214 and soil fungal sp. ANG28 with maximum support. This clade clustered separately from other known, and validly described Absidia species with a statistical support of 91/100/1 (ML/ML/PP). Together, they are sister to the clade formed by A. jindoensis and A. jiangxiensis.
The genetic distance in the trimmed ITS alignment of the new species and its sister taxa was computed. The genetic distance between our new species and A. jindoensis ranges from 11.5%−13.5%, and 24%−34.5% to A. jiangxiensis. This provides additional concrete evidence that validates the new species.
Absidia zygospora Hurdeal VG & Gentekaki E, sp. nov. Fig. 1, 2
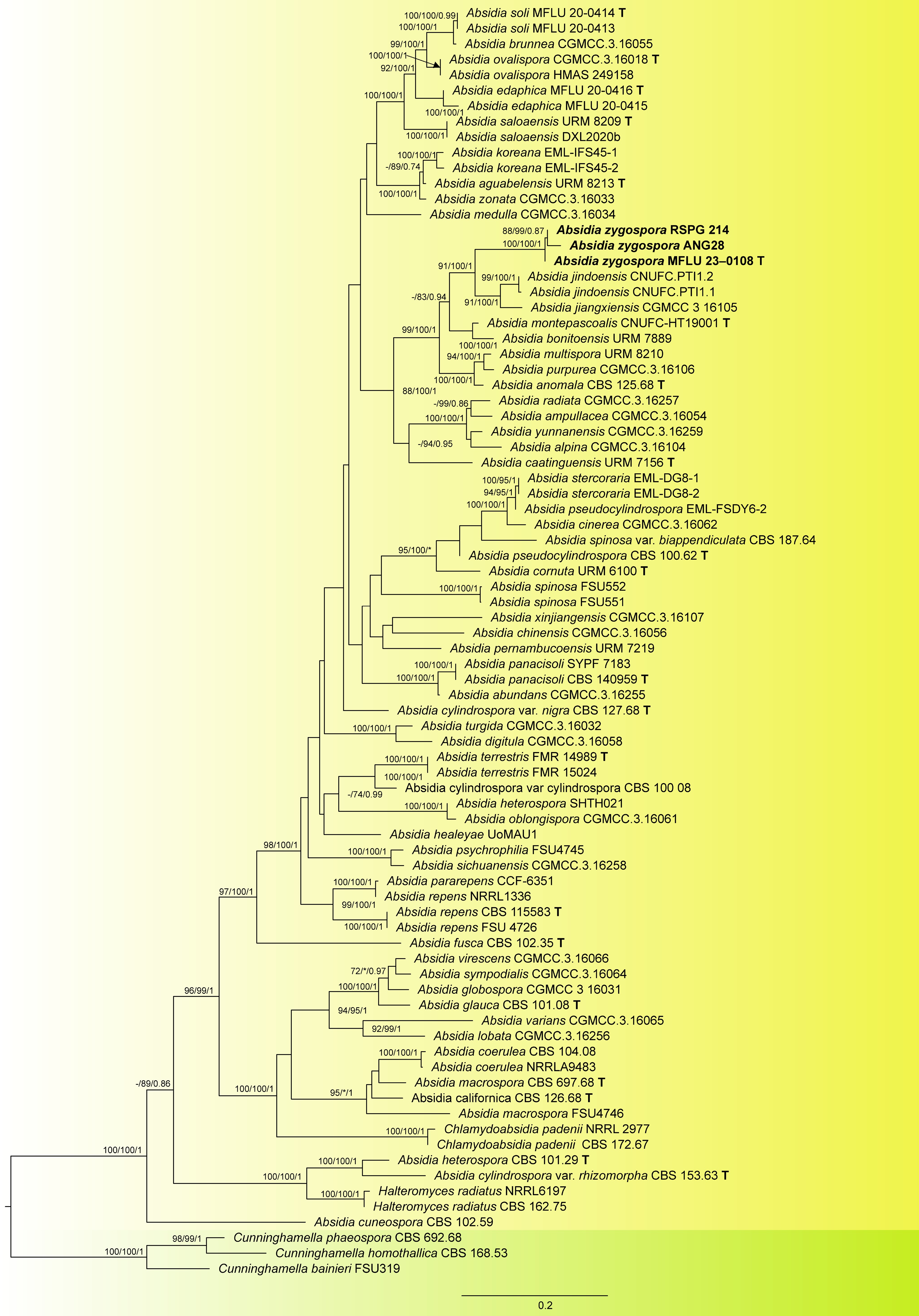
Figure 1.
Maximum likelihood phylogram inferred from 82 taxa and 1401 characters based on ITS, and LSU matrix using GTR+G+I model. ML bootstrap supports (≥ 70%) and Bayesian posterior probability (≥ 0.70) are indicated above the branches or near the nodes as ML/ML/PP. Tree is rooted using Cunninghamella homothallica (CBS 168.53), C. phaeospora (CBS 692.68), and C. bainieri (FSU319). Strains of the new species are in bold and the type species in the dataset are indicated using T. (−) represent bootstrap support lower than 70%.
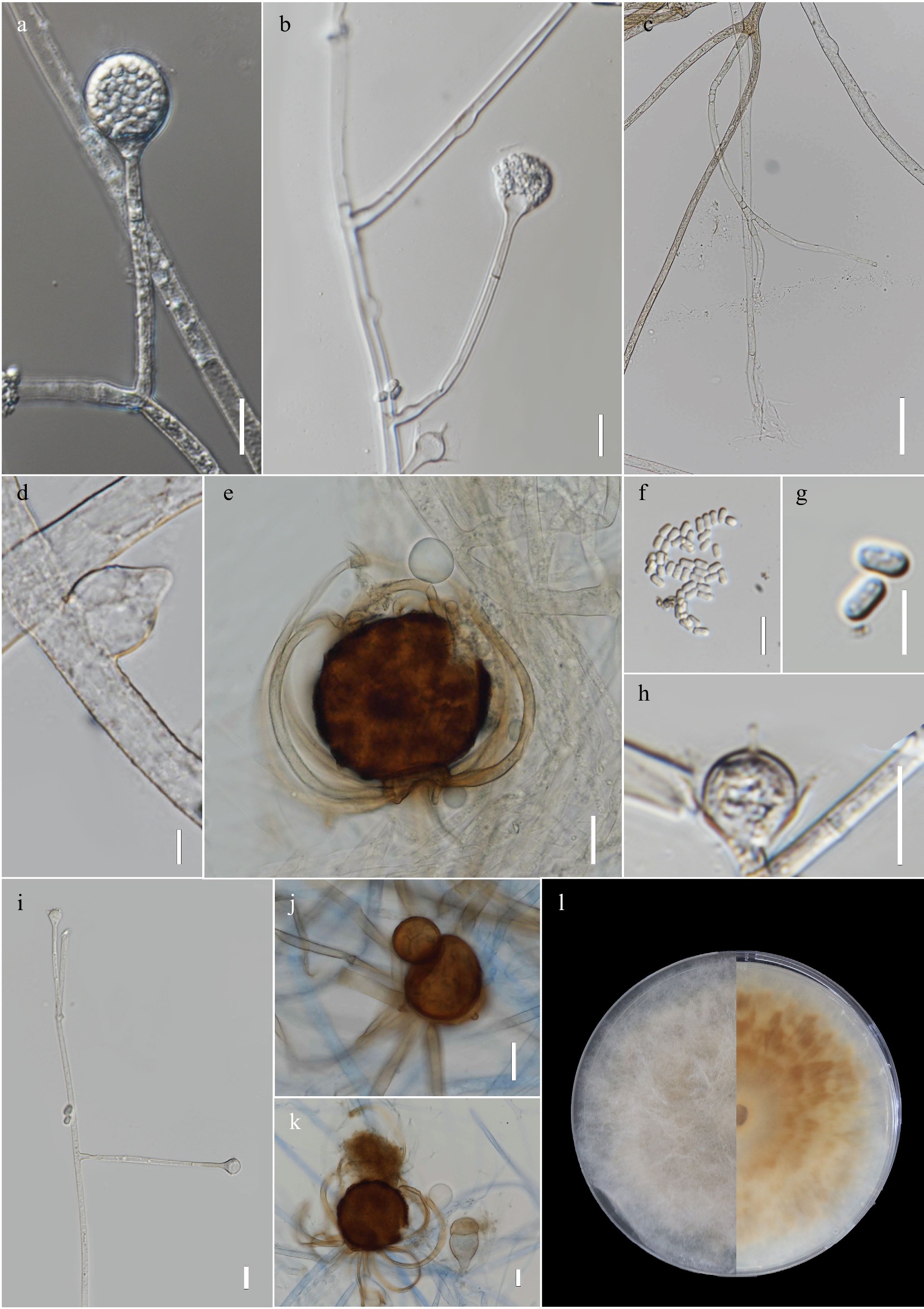
Figure 2.
Absidia zygospora MFLUCC 23–0061 (ex-type). (a) Developing sporangium. (b) Simple sporangiophore with sporangium, and subsporangial septation and columella with apical projection. (c) Rhizoids, (d) Swollen hyphae. (e), (j)–(k) Zygospores with unequal suspensors and finger-like appendages. (f)–(g) Sporangiospores. (h) Columella with apical projection. (i) Simple branching of sporangiophores. (l) Front and obverse images of culture in PDA. Scale bars: a–d, f, h, i = 10 µm, e, j–k = 20 µm, g = 5 µm.
Index Fungorum number: IF 900230
Etymology: named after its ability to produce sexual spores
Holotype: MFLU 23–0108
Asexual morph on MEA at 25 °C: Sporangiophores unbranched (mostly) or in whorls (2−4), initially hyaline, turning brown as the culture matures, up to 2.5 μm wide. Subsporangial septum present at the base of the sporangium (
$\overline{\text{x}} $ $\overline{\text{x}} $ $\overline{\text{x}} $ $\overline{\text{x}} $ Sexual morph on MEA at 25 °C after 30 d: Homothallic. Zygosporangia globose to subglobose, 50–77.5 × 59.5–76.5 μm (
$\overline{\text{x}} $ Culture characteristics – Colony grows faster in MEA than PDA. Within 3 d, the colony attains a diameter of 35 mm at 25 °C in PDA, while in MEA, it reaches 45 mm. At 15 and 20 °C, colony growth rate is similar from day 1 to day 3, in both MEA and PDA. The same is observed at temperatures 25 and 30 °C. Day-old cultures in MEA, at 25 °C, are white with a slight yellowish to pale brown tint around the inoculation plug. At this stage, only mycelial growth is observed, no reproductive structures, such as the sporangia, are produced. On day 2, the colony has a pale brown color and the outer most part of the colony is white (perimeter). Few sporangia on single sporangiophores are observed. On day 3, a pale brown colony with a white outer most layer persists. Production of sporangia and sporangiophores remain minimal, with most sporangia still immature (no colored sporangia). Most of the sporangiophores and sporangia are formed near the inoculation plug, hence in the older part of the culture. Whorls of sporangiophores which are typical of Absidia are not observed and if observed only whorls of 2 are seen. At the same temperature (25 °C), growth is quite slow and sporangiophore and sporangia formation is rare to none in PDA. However, by day 3, zygosporangia with finger-like appendages can be observed. More zygosporangia appeared to be produced on PDA than in MEA, in which only a few are observed. The fungus can grow at 15, 20, 25, 30 °C, with the optimal range being 25–30 °C. Growth was not observed at 10 and 37 °C.
Material examined: Thailand, Nan province, isolated from soil, on January 2020, collected and isolated by Vedprakash G. Hurdeal, ex-type culture, MFLUCC 23–0061; holotype: MFLU 23–0108.
Notes: In the phylogenetic analyses A. zygospora grouped as sister to the clade formed by A. jindoensis and A. jiangxiensis with high statistical support. Genetic distances between the new species and its sister taxa also provided further evidence for the introduction of the new taxon. Physiological data show that the new species has a lower growth rate than A. jiangxiensis. A more significant difference in rate of growth was seen between A. zygospora and A. jindoensis (45 mm vs 90 mm at 25 °C in MEA after 3 d). The new species produces smaller sporangia, and columella than sister taxa. The zygospores produced by A. zygospora are within the range of the other species, and are produced on significantly unequal suspensors. Given that equal suspensors comprise a defining characteristic of Absidia an amendment in the genus description needs to be undertaken. The same phenomenon is seen in A. jiangxiensis. The production of the appendages is usually from the larger suspensor (also seen in A. jiangxiensis).
-
Ecologically, Absidia species contribute to organic matter decomposition which is vital for recycling nutrients and have other functions such as in soil structure and aggregation. In this study, a new Absidia species, isolated from soil, is introduced based on morphological data, colony growth and phylogenetic analyses. The new species has smaller sporangia, columella and spores compared to the sister taxa (clade comprising of Absidia jindoensis CNUFC-PTI1-2, CNUFC-PTI1-1 and Absidia jiangxiensis CGMCC.3.16105). Inferred phylogeny indicates that our isolate is new with statistical support obtained by maximum likelihood (IQ-Tree and RAxML) analyses and Bayesian inference. Herein, phylogeny is inferred based on ITS and LSU genetic markers which are the most available sequences for this genus. ITS is generally considered the barcode for fungi. For Absidia, ITS can generally be used to differentiate species. However, cloning is often required to obtain the good quality ITS sequence data which renders the introduction and discovery of a new taxon difficult. As more taxa are introduced, the topology of the Absidia tree changes. This can be seen from the phylogenies of various taxonomic studies of this genus, highlighting the importance of taxon sampling[8−13]. Previously, spore shape was proposed as a taxonomically informative character of Absidia[7]. Our new species produces cylindrical spores and groups with Absidia species that produce similar spores. Hence, even with the establishment of more taxa, spore shape categorization is informative for initially identifying in which clade a species might be place.
In this study, the genetic distance in the ITS between A. zygospora MFLUCC T20-0309 and A. jindoensis ranges from 11.5%−13.5 %, and 24%−34.5 % to A. jiangxiensis, which seems in this case to meets the criterion for the establishment of a new species[22,23].
Our isolate forms a clade with the unclassified strains Absidia sp. RSPG 214 and soil fungal sp. ANG28, which are therefore referred to as A. zygospora. This supplements evidence to the introduction of the new species by providing additional information in terms of rDNA sequences and increasing taxon sampling. With several strains isolated from various places of the globe, important ecological information can be deducted in terms of the distribution. Interestingly, Absidia sp. RSPG 214 was also found in Thailand, but in Surat Thani, a southern province providing clues on the distribution of this species in the country. Meanwhile, ANG28 was isolated from soil in the United States.
Currently, few reports on the sexual morph of Absidia species has been published. In this study, we provide a description based on both the sexual and asexual stages of A. zygospora. The sexual stage is typical of the genus with finger-like appendages, but with unequal suspensors. The presence of unequal suspensors is in conflict with the generic description whereby, the genus is said to produce zygospore strictly on equal suspensors. Hence, the generic description of Absidia is amended to include zygospores produced on both equal and unequal suspensors. This finding underscores our limited knowledge of this genus and emphasizes the necessity for further taxonomic investigations, including the discovery of more species. Moreover, accurate taxonomic identification is essential for comprehending the biodiversity and distribution patterns of Absidia species, including the newly isolated species from the Nan Province in Thailand.
Vedprakash G. Hurdeal thanks Mae Fah Luang University and Mushroom Research Foundation for the Ph.D. scholarship and their support in the research on basal fungi. He acknowledges the Thesis or Dissertation writing grant (Oh7702(6)/0156) and research publication grant of Mae Fah Luang University. E. B. Gareth Jones thanks the King Saud University, Kingdom of Saudi Arabia for the award of a Distinguished Scientist Fellowship (DSFP). The authors thank Shaun Pennycook for his help in the nomenclature of the species.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hurdeal VG, Jones EBG, Gentekaki E. 2023. Absidia zygospora (Mucoromycetes), a new species from Nan Province, Thailand. Studies in Fungi 8:15 doi: 10.48130/SIF-2023-0015 

Absidia zygospora (Mucoromycetes), a new species from Nan Province, Thailand
- Received: 29 March 2023
- Accepted: 19 June 2023
- Published online: 26 September 2023
Abstract: Absidia is one of the most commonly isolated fungi among Cunninghamellaceae. The genus comprises saprobes isolated from soil, dung and other organic debris such as leaf litter. During a survey aimed at exploring the diversity of basal lineages of soil fungi, samples were collected from Nan province, Thailand. This led to the collection of a new Absidia isolate from soil. Characterization of the new isolate was based on morphological characters, colony growth and DNA sequence data. Phylogenetic analyses indicate that the new isolate comprises a lineage distinct from other described species. Morphological characterization showed that the isolate has smaller sporangia and columellae than its sister taxa. Furthermore, physiological data and genetic distance analysis supported the establishment of the new taxon. Hence, in this study, a new species of Absidia (A. zygospora) is introduced based on morphology, phylogeny and physiology.
-
Key words:
- 1 new species /
- Mucorales /
- Soil /
- Terrestrial habitat /
- Zygosporic fungi