









-
Pansy (Viola × wittrockiana Gams.), an important biennial plant in the Violaceae, is widely grown as a commercial flower for its white, yellow, orange, red, blue, and purple flowers in winter and spring. In some cultivars, anthocyanins accumulate at the base of the petals to generate variegated patches or blotches[1]. For example, purple blotches on the white flowers of 'Mont Blanc' mainly contain cyanidin-p-coumarylglycoside, and purple blotches on the yellow flowers of 'Rhinegold' contain mainly delphinidin-3:5-p-coumaryl glucorhamnoside[2]. In our previous research, we showed that blotch formation was caused primarily by accumulation of cyanidin and delphinidin.
Anthocyanins are major pigments from the flavonoid family that are responsible for flower color, pollinator attraction, and UV protection[3,4], and the pathways of flavonoid and anthocyanin biosynthesis are well understood in most model plants[5]. They involve a variety of biosynthesis genes, such as phenylalanine ammonia-lyase (PAL), chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), flavonoid 3′-hydroxylase (F3′H), flavonoid 3′,5′-hydroxylase (F3′5′H), dihydroflavonol 4-reductase (DFR), and anthocyanidin synthase (ANS)[6]. Almost all anthocyanidins undergo several modifications, including glycosylation, acylation, and methylation, which are catalyzed by glycosyltransferases (GTs), acyltransferases (ATs), and methyl transferases (MTs), respectively. Together, these genes synergistically regulate the biosynthesis of anthocyanins[7].
The transcriptional regulation of flavonoid and anthocyanin biosynthesis genes by transcription factor (TF) complexes of R2R3-MYB, bHLH, and WD40 family members has been widely demonstrated[5]. For example, AtMYB11, AtMYB12, and AtMYB111 of Arabidopsis thaliana target similar genes in the flavonoid biosynthesis pathway, including CHS, CHI, F3H, and flavonol synthase 1 (FLS1)[8]. Likewise, basic region/leucine zipper (bZIP) TFs have equally pivotal and diverse roles in the regulation of anthocyanin biosynthesis. An example is AtHY5, which promotes the expression of AtMYB12 and the accumulation of anthocyanins under light conditions in Arabidopsis[8].
Anthocyanin biosynthesis is typically regulated by specific phytohormones. In previous studies, jasmonic acid[9], cytokinins[10], ethylene[11], brassinosteroids[12], and abscisic acid[13] have been shown to promote anthocyanin accumulation by activating the transcription of anthocyanin biosynthesis genes, usually via TFs. In Lycium, for example, ABA stimulated transcription of the MYB-bHLH-WD40 (MBW) TF complex to upregulate expression of genes in the flavonoid biosynthetic pathway, thereby promoting anthocyanin production and fruit coloration[14]. In apple treated with ABA, the MdbZIP44 TF enhanced the ability of MdMYB1 to bind to the promoters of the anthocyanin biosynthesis genes MdDFR and MdUFGT[15].
Tyrosine is an aromatic α-amino acid that contains a phenolic hydroxyl group[16]. It is also a precursor for the shikimate pathway, which feeds into the anthocyanin pathway when tyrosine ammonia lyase (TAL) is active[17]. In recent years, tyrosine has been shown to upregulate anthocyanin-positive TFs such as PAP1, PAP2, and EGL3 and induce anthocyanin biosynthesis in Arabidopsis thaliana[18]. Recently, we found that tyrosine decarboxylase (TYDC), which catalyzes tyrosine to tyramine, was significantly upregulated in blotched areas relative to non-blotched areas of pansy petals, suggesting that tyrosine may play an important role in anthocyanin biosynthesis. In addition, our research indicated that tyramine might contribute to flower color and blotch formation as a co-pigment of anthocyanins in pansy[19]. Based on these findings, we speculated that tyrosine might also effect anthocyanin biosynthesis in pansy petals. However, more details are needed to fully understand how tyrosine affects anthocyanin biosynthesis in pansy.
In the present study, we performed virus-induced gene silencing (VIGS) to block the expression of VwTYDC and thus increase tyrosine content in order to determine whether this would promote anthocyanidin accumulation in pansy. Anthocyanin-free areas of pansy petals were treated with exogenous tyrosine or water and were then used for transcriptome sequencing, metabolite analysis, gene expression verification, and anthocyanin measurements. The results revealed a possible mechanism by which tyrosine contributes to the formation of pansy petal blotches and provide new insights into the production of anthocyanins in plants.
-
Viola × wittrockiana 'Mengdie' (Fig. 1) plants were used as materials and were planted at the horticultural farm of Hainan University (20.03N, 110.19E), in Haikou, Hainan Province, China. The plants were cultivated in 18-cm pots from winter to early spring (November 2019 through March 2020). During the planting period, soil moisture remained above 60%, the average daily temperature was about 20 °C, and the daylength was about 11 h. When the flowers were in full bloom, they were used for VIGS assays and ABA treatments.

Figure 1.
Phenotypes of VwTYDC-silenced Viola × wittrockiana. (a) Blank control; (b) flower phenotype after infection with pTRV2-VwCHS; (c) flower phenotype after infection with pTRV2-VwANS; (d) flower phenotype after infection with pTRV2-VwTYDC.
Virus-induced gene silencing
-
The pTRV1 and pTRV2 vectors were used to perform the VIGS assay on blooming pansy flowers. Partial sequences of VwANS (300 bp), VwCHS (270 bp) and VwTYDC (300 bp) were cloned into the pTRV2 vector, with VwCHS and VwANS serving as positive controls. The primers are shown in Supplemental Table S1. Agrobacterium strain GV3101 was transformed with the plasmids, pTRV1, pTRV2, pTRV2-VwANS, pTRV2-VwCHS and pTRV2-VwTYDC, then cultured in liquid YEP medium containing 100 mg/L kanamycin and 20 mg/L rifampicin. All samples were shaken at 28 °C at 250 rpm until the OD600 value reached 1.0. The bacterial cultures were then centrifuged, and the supernatants were removed. The residues were resuspended in MMA solution (10 mM MgCl2·6H2O, 10 mM 2-[N-morpholino] ethanesulfonic acid [MES], and 148 uM acetosyringone) and the OD600 value was readjusted to 1.0. Each MMA solution containing pTRV2-VwCHS, pTRV2-VwTYDC, pTRV2-VwANS, or pTRV2 was mixed with MMA solution containing pTRV1 (V/V, 1:1) and incubated in the dark for 2–3 h. The mixed solutions were then injected into the epidermis at the top of the pansy stem, and any phenotypic changes in the petals were observed and photographed over subsequent days.
Tyrosine and ABA treatment of non-blotched areas on pansy petals
-
Petals of blooming pansy flowers were treated with 3 mM tyrosine (CAS:60-18-4, Sigma, USA). A syringe tip was used to pierce the upper epidermis of non-blotched areas, and tyrosine solution was smeared onto the punctured regions. Plants in the control group were treated with ddH2O. After 24 h, the treated tissues were collected for anthocyanin analysis and total RNA extraction. All samples were immediately frozen in liquid nitrogen and stored at −80 °C. Treatment with 50 μM ABA (S-ABA, CAS:14375-45-2, Phyto Technology Laboratories, USA) was similar to the tyrosine treatment, and all treatments had three biological replicates.
Anthocyanin detection
-
The anthocyanin metabolome of petals treated with tyrosine or ddH2O was analyzed using a UPLC-ESI-MS/MS system (UPLC, ExionLCTM AD, Sciex; MS, Applied Biosystems 6500 Triple Quadrupole, Sciex). The frozen petals were ground into powder (30 Hz, 1.5 min), and 50 mg of powder was weighed and extracted in 0.5 mL methanol/water/hydrochloric acid (500:500:1, V/V/V). The extract was then vortexed for 5 min, ultrasonicated for 5 min, and centrifuged at 4 °C and 13,000 rpm for 3 min. The residue was re-extracted by repeating the above steps again under the same conditions. The supernatants were collected and filtered through a 0.22-μm membrane filter (Anpel) before LC-MS/MS analysis with a Waters ACQUITY BEH C18 column (1.7 µm, 2.1 mm × 100 mm) and a solvent system of water (0.1% formic acid): methanol (0.1% formic acid). The gradient program was 95:5 V/V at 0 min, 50:50 V/V at 6 min, 5:95 V/V at 12 min, hold for 2 min, 95:5 V/V at 14 min; and hold for 2 min. The flow rate was 0.35 mL/min, the temperature 40 °C, and the injection volume 2 μL. The ESI source operation parameters were as follows: ion source, ESI+; source temperature, 550 °C; ion spray voltage (IS), 5,500 V; curtain gas (CUR), 35 psi. Anthocyanin contents were detected by MetWare based on the AB Sciex QTRAP 6500 LC-MS/MS platform.
ABA analysis
-
ABA contents were analyzed using an ESI-HPLC-MS/MS system (HPLC, Agilent 1290; MS/MS, Applied Biosystems 6500 Quadrupole Trap). Frozen petals were ground into powder (30 Hz, 1.5 min), and 1.5 g of powder was weighed and extracted in 0.4 mL isopropanol/water/hydrochloric acid. Then, 8 μL of 1 μg/mL internal standard solution was added, and the mixture was shaken at low temperature for 30 min. Dichloromethane was added, the extract was shaken at low temperature for 30 min and centrifuged at 4 °C and 13,000 rpm for 5 min, and the supernatants were removed. The residual organic phase was dried with nitrogen and reconstituted in methanol. The extract was centrifuged at 4 °C and 13,000 r/min for 10 min. The supernatants were filtered through a 0.22-μm filter membrane and measured by HPLC-MS/MS using a Poroshell 120 SB-C18 column (2.7 μm, 2.1 mm × 150 mm) and a solvent system of (A) methanol (0.1% formic acid) and (B) water (0.1% formic acid). The gradient program was 0–1 min, 20% A; 1–9 min, 20%–80% A; 9–10 min, 80% A; 10–11 min, decrease to 20% A; 11–15 min, 20% A. The injection volume was 2 μL. The ESI source operation parameters were as follows: ion source, ESI+; source temperature, 400 °C; IS, 4,500 V; CUR, 15 psi. Analyst software (Sciex) was used to analyze the ABA data.
RNA extraction, cDNA library construction, and mRNA sequencing
-
RNA was extracted from the tyrosine- and water-treated areas of pansy petals using a modified TRIzol method. Three biological replicates were used for the analysis. High-quality RNA was used for mRNA sequencing: mRNA was enriched with Oligo(dT) beads, and the enriched mRNA was then broken into short fragments using fragmentation buffer and reverse transcribed into cDNA using random primers. Second-strand cDNA was synthesized using DNA polymerase I, RNase H, dNTP, and buffer. The cDNA fragments were purified with the QiaQuick PCR extraction kit, end repaired, poly(A) tailed, and ligated to Illumina sequencing adapters. The ligation products were selected by agarose gel electrophoresis, PCR amplified, and sequenced using the Illumina HiSeq 4000 platform by Gene Denovo Biotechnology Co. (Guangzhou, China).
mRNA transcriptome data analysis
-
Sequencing reads were filtered by removing adaptor-contaminated reads, reads with > 10% unknown nucleotides (Ns), and reads with > 40% low-quality (Q-value ≤ 20) bases. The high-quality clean reads were mapped to rRNA to identify residual rRNA reads, and such reads were removed. The remaining high-quality clean reads were mapped to the reference transcriptome using Bowtie 2 with default parameters[20], and the mapping ratio was calculated. Trinity was used to assemble the clean sequencing data to obtain unigenes[21].
The gene abundances were calculated and normalized to RPKM (Reads Per Kilobase per Million reads)[22], and the expression levels of unigenes in the two treatments were calculated. Differentially expressed genes (DEGs) between the tyrosine and ddH2O treatments were identified and filtered using edgeR (
www.bioconductor.org/packages/release/bioc/-html/edgeR.html ) based on FDR < 0.05 and |log2FC| > 1. The DEGs were input to Blast2GO software and in-house perl scripts for gene ontology (GO) term analysis, and KEGG pathways were assigned to the assembled genes using the KEGG Automatic Annotation Server (KAAS,www.genome.jp/kegg/ ) in order to determine their biological functions, metabolic pathways, and signal transduction pathways. To identify TFs, the assembled transcriptomes were searched against the Plant Transcription Factor Database (http://planttfdb.cbi.pku.edu.cn ) using hmmsearch v3.0 (http://hmmer.org ).The protein functional annotations of unigenes were obtained by performing blastx (
www.ncbi.nlm.nih.gov/BLAST ) against the NR, Swissprot, KEGG, and COG/KOG annotation databases (E-value < 10−5). Functions of unigenes that did not receive protein annotations were predicted with ESTScan[23]. After GO annotation of unigenes with Blast2GO software[24], WEGO was used to calculate functional classification statistics for all unigenes with GO annotations[25].qRT-PCR analyses of mRNA
-
The RNA samples obtained for transcriptome sequencing were also used for quantitative reverse transcription-polymerase chain reaction (qRT-PCR). Reverse transcription was performed for single-stranded DNA synthesis using the PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa, Shanghai, China) according to the manufacturer's protocol. qRT-PCR was performed with three technical replicates and three biological replicates. The 18S gene was used as an internal control for normalization. qRT-PCR analysis was performed using Luna Universal qPCR Master Mix (New England Biolabs, Ipswich, MA, USA) according to the manufacturer's instructions with denaturation at 95 °C for 60 s and 40 cycles of amplification (95 °C for 15 s, 60 °C for 30 s). Expression levels of the target genes relative to the internal control were calculated using the 2−ΔΔCᴛ method[26]. The gene-specific primers are shown in Supplemental Table S2.
Transcription factor phylogenetic analyses
-
Protein sequences of the MYB and bZIP TFs from Arabidopsis were obtained from NCBI. The gene sequences of pansy MYBs and bZIPs were obtained from the transcriptome, and their conserved domains were analyzed using SMART software in Genomic mode with default parameters. Phylogenetic trees of the two TF families were constructed using MEGA 5.02 with default parameters.
Statistical analysis
-
ABA content experiments and qRT-PCR analyses were performed in triplicate. Univariate values were analyzed using ANOVA, and mean values were compared using Duncan's new multiple range test (P < 0.05) with SAS software (SAS Institute Inc., Cary, NC, USA).
-
VIGS was performed to verify the function of VwTYDC. As shown in Fig. 1, there was no change in flower phenotype after injection of the blank control pTRV2 (Fig. 1a), but the blotches disappeared or became smaller when plants were injected with the positive controls pTRV2-VwANS and pTRV2-VwCHS (Fig. 1b, c). In the pTRV2-VwTYDC treatment, the flower color changed to pink (Fig. 1d), indicating pigment accumulation in petals when VwTYDC was silenced.
Phenotypic changes and anthocyanin accumulation of pansy petals treated with exogenous tyrosine
-
To determine how tyrosine effects anthocyanin accumulation in pansy petals, non-blotched petal areas were treated with exogenous tyrosine. Petal phenotypes were observed after 24 h of treatment with water or tyrosine: there were no obvious changes in the non-blotched areas following water treatment (Fig. 2a), but cyanic stripes appeared in the non-blotched areas of pansy petals following treatment with tyrosine (Fig. 2b).

Figure 2.
Effect of tyrosine treatment on non-blotched areas of Viola × wittrockiana petals. (a) ddH2O treatment; (b) tyrosine treatment. The treated areas are marked by a red box.
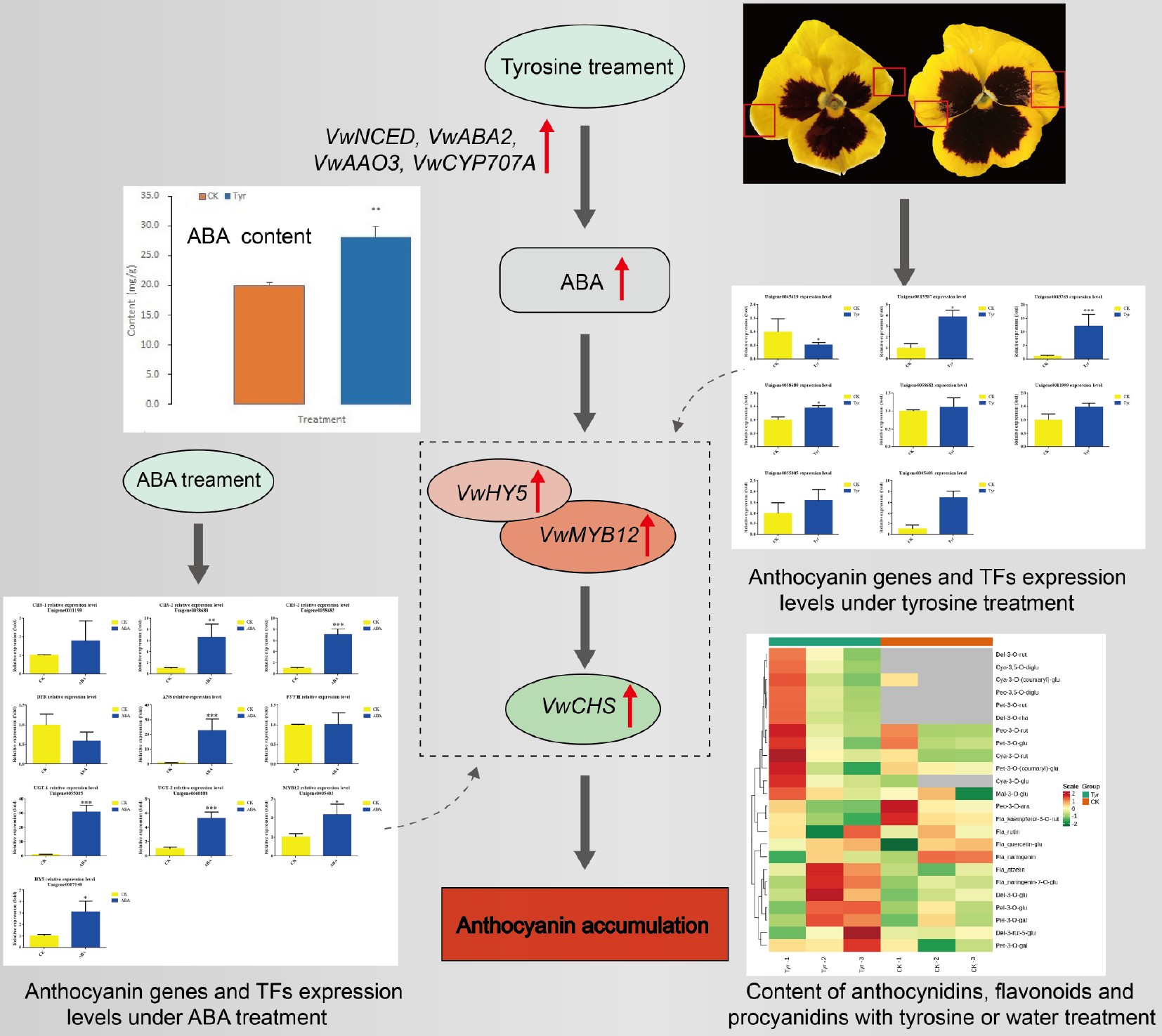
ESI-HPLC-MS/MS was used to scan for 108 anthocyanidins, flavonoids, and procyanidins in the petal areas that received tyrosine or water treatment, and 24 compounds were detected in at least one of the treatments (Supplemental Table S3). The LC-MS/MS data have been uploaded to MetaboLights under number MTBLS3419. The contents of all 24 compounds in different samples were analyzed by hierarchical clustering analysis (Figs 3 & 4). The contents of two metabolites, cyanidin-3-O-glucoside and cyanidin-3-O-rutinoside, differed based on the threshold Fold Change ≥ 2 or Fold Change ≤ 0.5 (Fig. 3). However, if compounds with an expression level of N/A in the CK treatment were included, there were nine differentially abundant anthocyanidins, including cyanidins, delphinidins, petunidins, peonidins and pelargonidins. Among these, cyanidin-3-O-rutinoside showed the highest content of 5.098 ng/g. Delphinidin-3-O-rutinoside, delphinidin-3-O-rhamnoside, cyanidin-3,5-O-diglucoside, petunidin-3-O-rutinoside, peonidin-3,5-O-diglucoside, peonidin-3-O-galactoside, and pelargonidin-3-O-(coumaryl)-glucoside were detected in the tyrosine treatments but not in the water treatments (Fig. 4).

Figure 3.
Violin plot of anthocyanins in petals of Viola × wittrockiana treated with tyrosine or ddH2O. The box in the middle represents the interquartile range. The black horizontal line in the middle is the median, and the outer shape represents the distribution of the data.
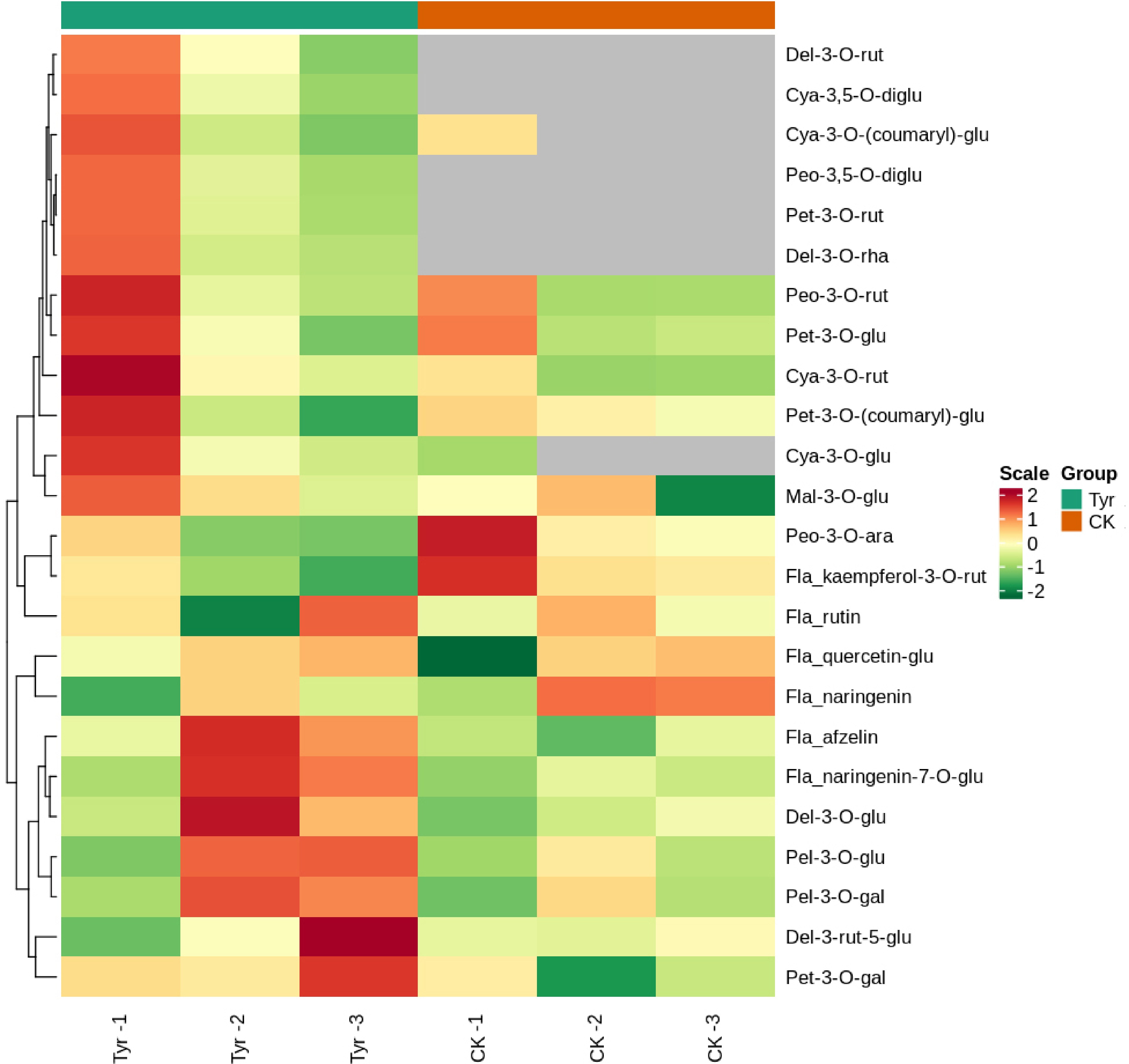
Figure 4.
Heatmap of the contents of the 24 compounds detected in different Viola × wittrockiana samples analyzed by hierarchical clustering analysis. The horizontal axis shows the sample information, and the vertical axis shows the metabolite information. The tree on the left of the figure shows the metabolite clustering, and the scale is the metabolite content after standardization. A deeper red color indicates a higher content, and gray indicates that the compound was not detected. Groups indicate the different treatments. CK: ddH2O treatment; Tyr: tyrosine treatment.
RNA sequencing, gene functional annotation, and classification
-
Three cDNA libraries were constructed from petals treated with tyrosine, and 57,665,984, 41,950,452 and 49,379,404 high-quality reads were obtained. Three more cDNA libraries were constructed from the control samples, and 52,086,074, 46,811,096, and 48,288,596 high-quality reads were obtained. The final transcriptome assembly contained 90,732 genes with an average length of 735 nt and an N50 of 1,172 nt. The sequencing raw data have been deposited into the NCBI Sequence Read Archive (SRA) under accession number PRJNA754504.
To assign putative functions to the assembled genes, their sequences were searched against public databases; 50,829 genes were annotated by the NR database, and 36,481 genes were annotated by the KOG database. GO annotations (16,976 genes) and KEGG pathway annotations (15,175 genes) were also obtained to gain more insight into the putative gene functions. In the GO analysis, the terms metabolic process, catalytic activity, cellular process, binding, and single-organism process were the top five annotations with the largest number of genes (Supplemental Fig. S1). In the KEGG pathway analysis, the pathways with the greatest unigene enrichment were metabolic pathways (5790, 38.15%), followed by biosynthesis of secondary metabolites (3089, 20.86%), and biosynthesis of antibiotics (1622, 10.69%).
Identification of DEGs related to anthocyanin biosynthesis
-
A total of 19,438 DEGs were detected between water- and tyrosine-treated areas of pansy petals: 6401 downregulated and 13,037 upregulated (Supplemental Fig. S2). Among these, 4525 were mapped to 131 KEGG pathways. There were nine DEGs related to anthocyanin biosynthesis in the tyrosine-treated areas, and all but unigene0045619 were upregulated (Table 1). VwHCT (unigene0045619, unigene0015507), VwC3′H (unigene0083763, unigene0029000), and VwCHS (unigene0058680, unigene0058682, unigene0011199) were involved in the flavonoid biosynthesis pathway (ko00941), whereas VwUGT75C1 (unigene0060888, unigene0055085) was involved in the anthocyanin biosynthesis pathway (ko00942).
Table 1. Putative anthocyanin structural genes that were differentially expressed in response to tyrosine treatment in Viola × wittrockiana petals.
Gene ID Annotation RPKM
(Tyr treatment)RPKM
(H2O treatment)log2 (T/CK) FDR Up/Down unigene
0045619VwHCT 0.54 1.48 −1.45 0.05 Down unigene
0015507VwHCT 2.31 0.56 2.26 5.35e-12 Up unigene
0083763VwC3’H 3.60 0.34 3.42 1.18e-28 Up unigene
0029000VwC3’H 1.80 0.02 6.86 8.38e-15 Up unigene
0058680VwCHS 213.61 100.30 1.09 4.65e-35 Up unigene
0058682VwCHS 87.86 42.41 1.05 1.18e-41 Up unigene
0011199VwCHS 10.51 3.40 1.63 6.24e-33 Up unigene
0060888VwUGT75C1 13.30 5.71 1.22 2.95e-21 Up unigene
0055085VwUGT75C1 177.86 66.99 1.41 3.19e-105 Up VwHCT, shikimate O-hydroxycinnamoyltransferase; VwC3'H, coumaroylquinate 3'-monooxygenase; VwCHS, chalcone synthase; VwUGT75C1, anthocyanidin 3-O-glucoside 5-O-glucosyltransferase. Analysis of differentially expressed MYB transcription factors
-
There were 13 differentially expressed MYB TFs: eight upregulated and five downregulated (Supplemental Table S4). We constructed a phylogenetic tree that included the MYB TFs of Arabidopsis thaliana to provide insight into the potential functions of these differentially expressed MYB TFs (Supplemental Fig. S3). We found that unigene0005403 was highly homologous to the Arabidopsis TFs AT5G49330.1, AT2G47460.1, and AT3G62610.1, which encode AtMYB111, AtMYB12, and AtMYB11. A blastx search of the transcriptome data indicated that this unigene was also highly similar to an MYB12-like TF gene from Cicer arietinum (E-value < 10−5), and we therefore speculated that it was likely to be an MYB12-like gene in pansy.
qPCR of key anthocyanin biosynthesis and transcription factor genes
-
The relative expression levels of 10 DEGs (nine biosynthesis genes and one TF gene) related to the anthocyanin biosynthesis pathway were verified in tyrosine- and ddH2O-treated petals by qPCR. The expression levels of these DEGs were again promoted by tyrosine, with the exception of unigene0045619 (HCT-1-like) (Fig. 5), consistent with the transcriptome sequencing results (Table 1).
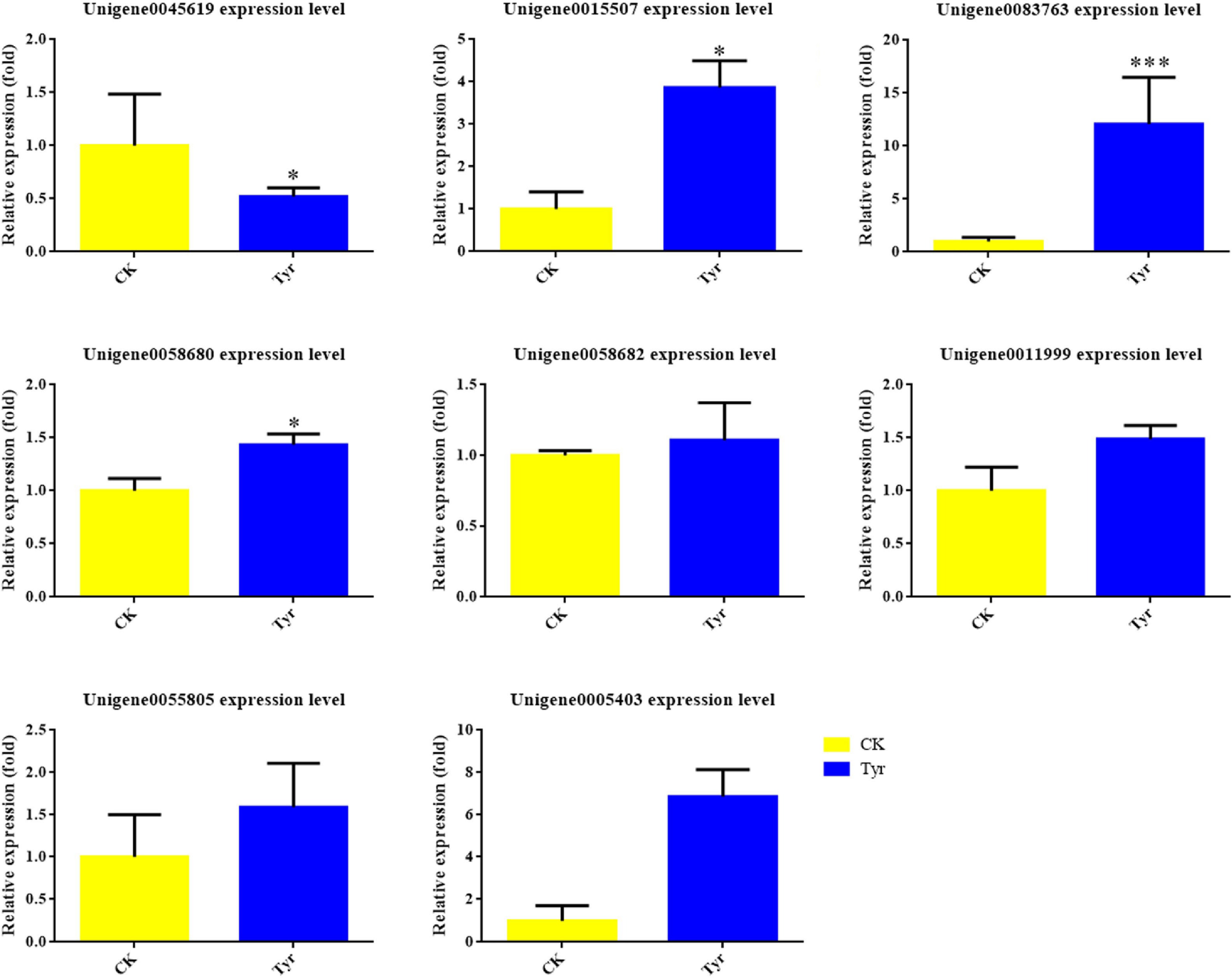
Figure 5.
The relative expression levels of anthocyanin biosynthesis and transcription factor genes in Viola × wittrockiana petals that received tyrosine (Tyr) or ddH2O (CK) treatment. * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001.
Tyrosine upregulates ABA biosynthesis-related genes and promotes ABA production
-
The transcriptome data revealed that expression levels of some key genes in the ABA biosynthesis pathway were significantly higher in the tyrosine-treated areas than in the H2O-treated areas based on FDR < 0.05 and |log2FC| > 1 (Table 2). This result suggested that the ABA content might be higher in the tyrosine-treated areas. We therefore analyzed ABA content and confirmed that it was significantly higher in the tyrosine-treated areas than in the H2O-treated areas (Fig. 6).
Table 2. Differentially expressed genes related to ABA biosynthesis in Viola × wittrockiana.
Gene ID Annotation RPKM
(Tyr treatment)RPKM
(H2O treatment)log2
(T/CK)FDR Up/Down unigene
0004917VwNCED 2.20 1.03 1.10 0.00 Up unigene
0068071VwNCED 2.41 0.56 2.10 7.18e-6 Up unigene
0068072VwNCED 1.16 0.35 1.72 0.00 Up unigene
0082413VwABA2 0.33 0.01 4.51 0.00 Up unigene
0073280VwAAO3 8.81 4.28 1.04 2.46e-31 Up unigene
0017204VwCYP707A 18.30 0.81 4.50 7.13e-212 Up unigene
0087548VwHY5 0.63 0 9.29 0.01 Up VwNCED, 9-cis-epoxycarotenoid dioxygenase; VwABA2, xanthoxin dehydrogenase 2; VwAAO3, abscisic-aldehyde oxidase; VwCYP707A, (+)-abscisic acid 8'-hydroxylase. 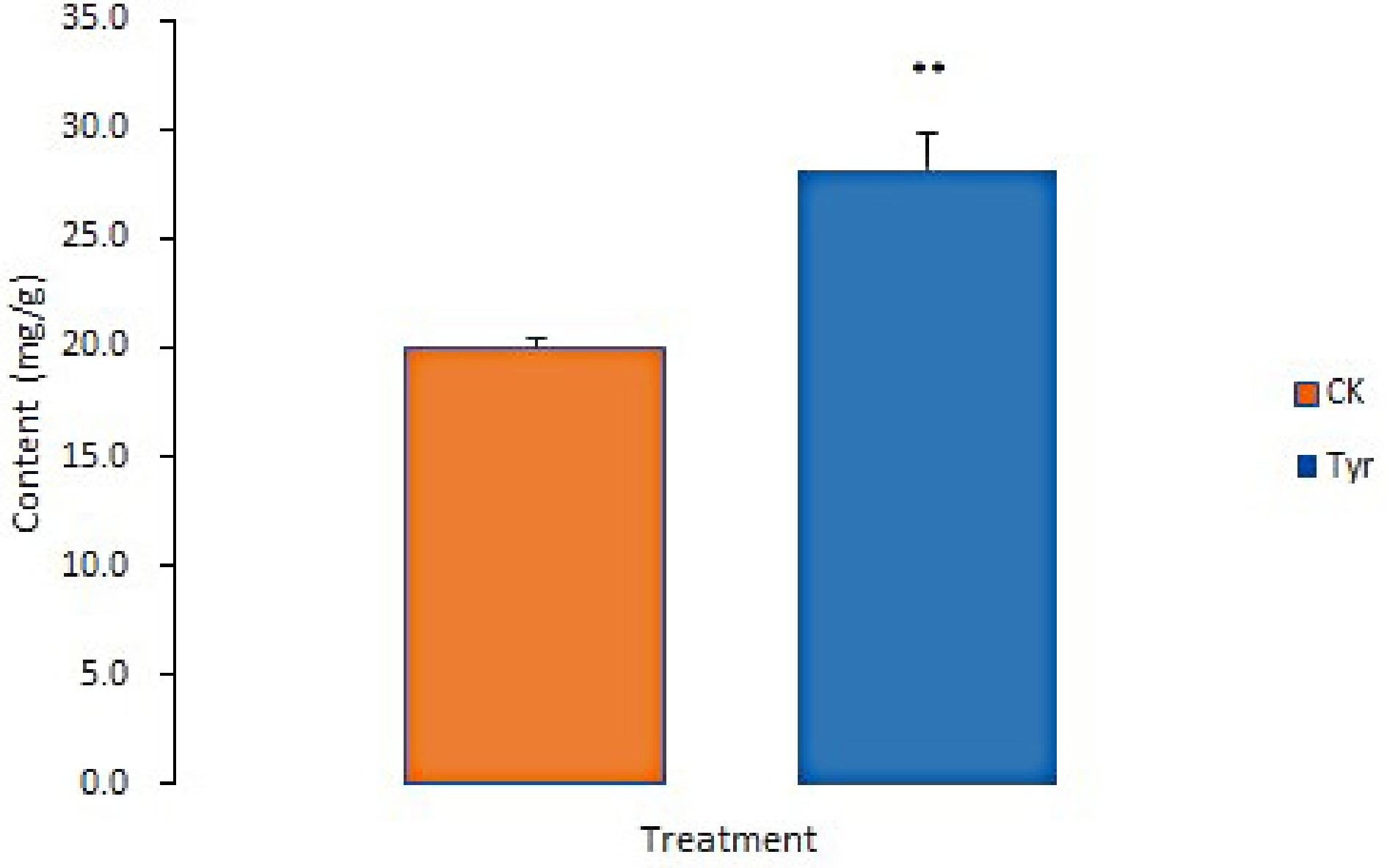
Figure 6.
ABA contents in Viola × wittrockiana from different treatment groups. ** p ≤ 0.01. CK, ddH2O treatment. Tyr, tyrosine treatment.
We also analyzed differentially expressed bZIP TFs of pansy by building a phylogenetic tree with their sequences and those of all bZIP TFs from Arabidopsis (Supplemental Fig. S4). The upregulated unigene0087548 was highly homologous to the Arabidopsis TFs AT3G17609.2 and AT3G17609.3, which encode homologs of AtHY5, an important bZIP TF that binds to the promoter of the bZIP TF gene ABA insensitive 5 (ABI5) to activate ABA biosynthesis[27].
Exogenous ABA upregulated the expression of anthocyanin biosynthesis genes
-
To determine whether ABA could activate anthocyanin-related genes in Viola × wittrockiana, we treated non-blotched areas of pansy petals with ABA and used ddH2O as a control treatment. The treated petal areas were then collected for qRT-PCR analysis. As shown in Fig. 7, ABA treatment upregulated the expression levels of multiple anthocyanin biosynthesis genes, including VwCHS, VwANS, and VwUGT, as well as the TF genes VwMYB12 and VwHY5.
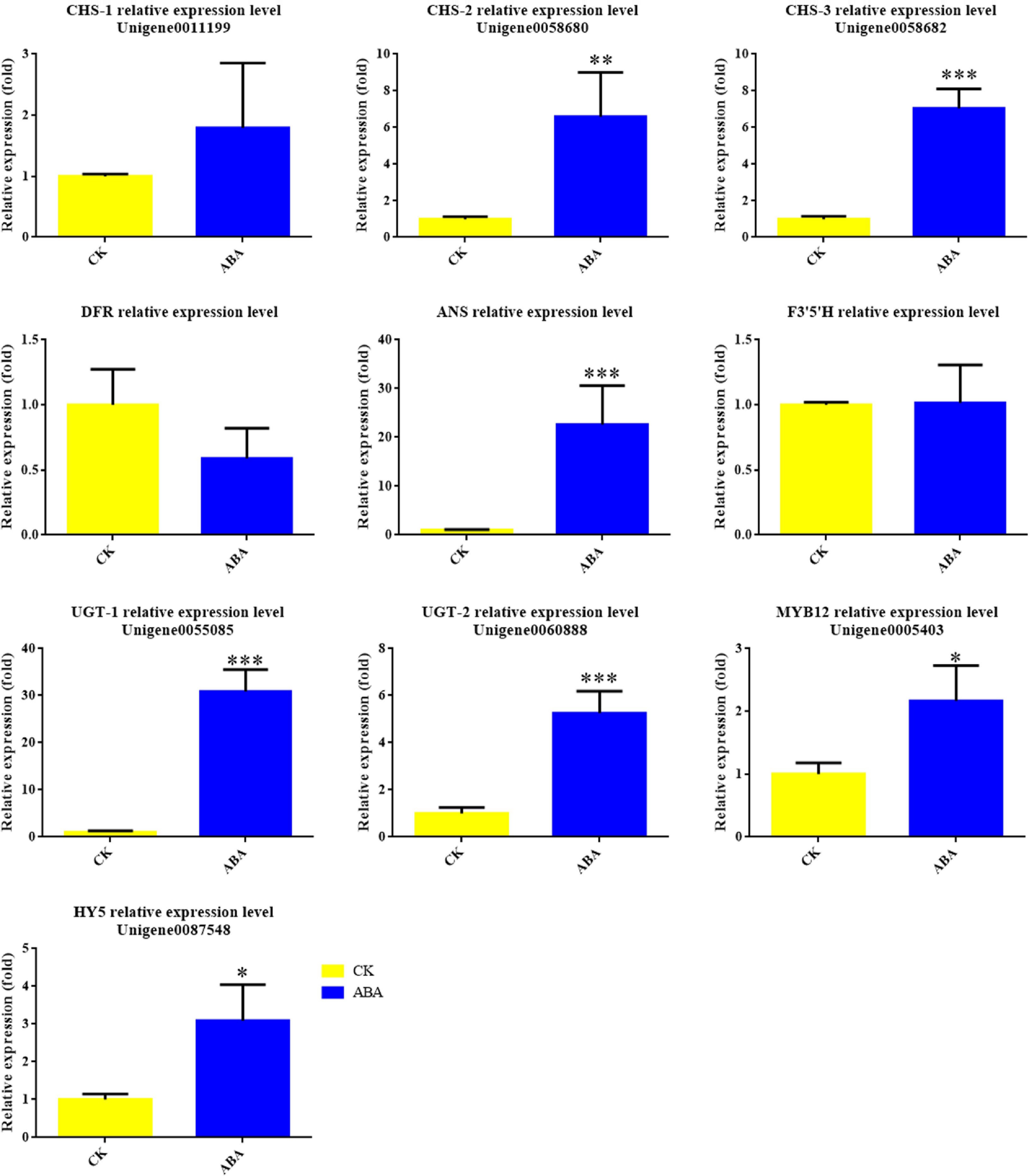
Figure 7.
Expression levels of anthocyanin biosynthesis genes and transcription factors in Viola × wittrockiana with and without ABA treatment. * p ≤ 0.05, ** p ≤ 0.01, ***p ≤ 0.001.
-
Tyrosine has recently been shown to promote anthocyanin biosynthesis, and the molecular mechanism by which it induces anthocyanin biosynthesis in Arabidopsis has been investigated. Exogenous tyrosine is now known to induce anthocyanin biosynthesis and accumulation by upregulating anthocyanin biosynthesis-related genes, including DFR, LODX, and UGT[18]. Tyrosine has also been found to promote the biosynthesis of flavonoids, the substrates of anthocyanin biosynthesis[28]. These results suggest that tyrosine may serve as an efficient regulatory metabolite for anthocyanin biosynthesis.
In the present research, VIGS silencing of VwCHS and VwANS produced obvious fading of petal blotches, thus confirming that the VIGS protocol worked well in pansy. By contrast, VIGS silencing of VwTYDC induced pigment accumulation in pansy petals (Fig. 1), demonstrating that an increase in tyrosine promoted pigment biosynthesis in pansy. Moreover, the non-blotched parts of the petals showed some cyanic spots or stripes after tyrosine treatment (Fig. 2). Flowers were also significantly smaller after VIGS treatment. This phenomenon may reflect the ability of the VIGS technique to silence relevant developmental genes, thereby affecting organ development; alternatively, wounding caused by the silencing treatment may have affected flower development (Fig. 2)[29,30]. Anthocyanin metabolome analysis demonstrated that contents of cyanidin-3-O-glucoside and cyanidin-3-O-rutinoside were significantly enhanced by tyrosine treatment. Furthermore, delphinidin-3-O-rutinoside, delphinidin-3-O-rhamnoside, cyanidin-3,5-O-diglucoside, petunidin-3-O-rutinoside, peonidin-3,5-O-diglucoside, peonidin-3-O-galactoside, and pelargonidin-3-O-(coumaryl)-glucoside were detected in the tyrosine treatment but not the water control (Figs 3 & 4). Consistent with these findings, the transcriptome results indicated that VwHCT, VwC3′H, VwCHS, and VwUGT were upregulated in the tyrosine-treated areas. All these results suggest that tyrosine may promote anthocyanin accumulation by upregulating a number of anthocyanin biosynthesis-related genes.
Twenty-four different anthocyanins and related compounds were detected in the different samples (Fig. 4), among which almost all of the anthocyanins had been reported previously in Viola[31]. Although the Tyr-1 data differed somewhat from those of Tyr-2 and Tyr-3, probably owing to difficulties in accurately sampling specific petal areas, cyanidin-3-O-rutinoside was the major enriched anthocyanin with the highest average content (5.098 ng/g) in tyrosine-treated areas, similar to the main anthocyanidins previously reported in the blotched areas of pansy petals[1].
Tyrosine promotes anthocyanin biosynthesis via ABA synthesis in pansy
-
Previous studies have reported that endogenous or exogenous factors can induce anthocyanin accumulation through ABA. In Lycium, for example, developmental cues transcriptionally activated LbNCED1 and thereby enhanced accumulation of ABA. ABA then stimulated transcription of the MYB-bHLH-WD40 TF complex, which in turn upregulated the expression of structural genes in the flavonoid biosynthetic pathway, ultimately promoting anthocyanin production and fruit coloration[32]. In bilberry (Vaccinium myrtillus), red light induced high expression of specific genes of anthocyanin biosynthesis and ABA signal perception and metabolism, including 9-cis-epoxycarotenoid dioxygenase (NCED), the ABA receptor pyrabactin resistance-like (PYL), and an abscisic acid 8'-hydroxylase gene (CYP707A) that functions in ABA catabolism[33]. ABA has been reported to promote anthocyanin accumulation by upregulating the expression of CHS, ANS, or UGT[34]. In our research, exogenous tyrosine application significantly upregulated the expression of several ABA biosynthesis genes, including VwNCED, VwABA2, VwAAO3, and VwCYP707A (Table 2). At the same time, the concentration of ABA was significantly higher in the tyrosine-treated areas of pansy petals (Fig. 6). Moreover, ABA treatment also significantly upregulated the expression of anthocyanin biosynthesis genes, including VwCHS, VwANS, and VwUGT, similar to the effect of tyrosine treatment on gene expression (Table 1). These results support a model in which tyrosine promotes anthocyanin biosynthesis via ABA accumulation in pansy petals.
Transcription factors may be important mediators of tyrosine-promoted anthocyanin biosynthesis
-
Anthocyanin biosynthesis genes are transcriptionally regulated by the MYB-bHLH-WD40 complex[35]. In Arabidopsis, exogenous tyrosine upregulates the expression of MBW complex genes including PAP1, PAP2, GL3, EGL3, and TTG1[18]. In this study, we found eight significantly upregulated MYB-like unigenes, among which unigene0005403 had high homology to MYB12 from Cicer arietinum. Phylogenetic analysis with Arabidopsis MYBs also supported the hypothesis that unigene0005403 may have similar functions to AtMYB12 (Supplemental Fig. S3), which upregulates early anthocyanin biosynthesis genes such as CHS, CHI, and F3H[36]. These results suggest that unigene0005403 may also promote anthocyanin accumulation by upregulating anthocyanin biosynthesis genes. According to the transcriptome and qPCR data for VwCHS, VwCHI, and VwF3H, only VwCHS was significantly upregulated in response to tyrosine, and we thus speculate that unigene0005403 may act mainly on VwCHS in pansy.
bZIP TFs may also regulate anthocyanin biosynthesis by interacting synergistically with the MYBs[37]. In Arabidopsis, AtHY5 is a bZIP gene that can activate the expression of MYB12/PFG1 and MYB75/PAP1[38]. HY5 is also known to activate the expression of MYB12 and MYB111[39], which are involved in the regulation of flavonol synthase[40]. Moreover, HY5 can respond to ABA by specifically binding to ABI5 chromatin[41]. In this research, unigene0087548, annotated as an HY5-like gene, and unigene0005403, annotated as an MYB12 homolog, were both significantly upregulated in ABA-treated pansy petals (Fig. 7), suggesting that HY5 may have responded to ABA treatment and interacted synergistically with VwMYB12 to promote anthocyanin biosynthesis in pansy.
A proposed model of the promotion of anthocyanin biosynthesis by tyrosine in pansy
-
On the basis of our experimental results, we propose a working model of the promotion of anthocyanin biosynthesis by tyrosine treatment in pansy. Tyrosine treatment activates ABA production by upregulating VwNCED, VwABA2, VwAAO3, and VwCYP707A. Then the bZIP TF VwHY5 responds to ABA accumulation and activates VwMYB12 to upregulate VwCHS expression and then induce anthocyanin production in non-blotched areas of pansy petals (Fig. 8).
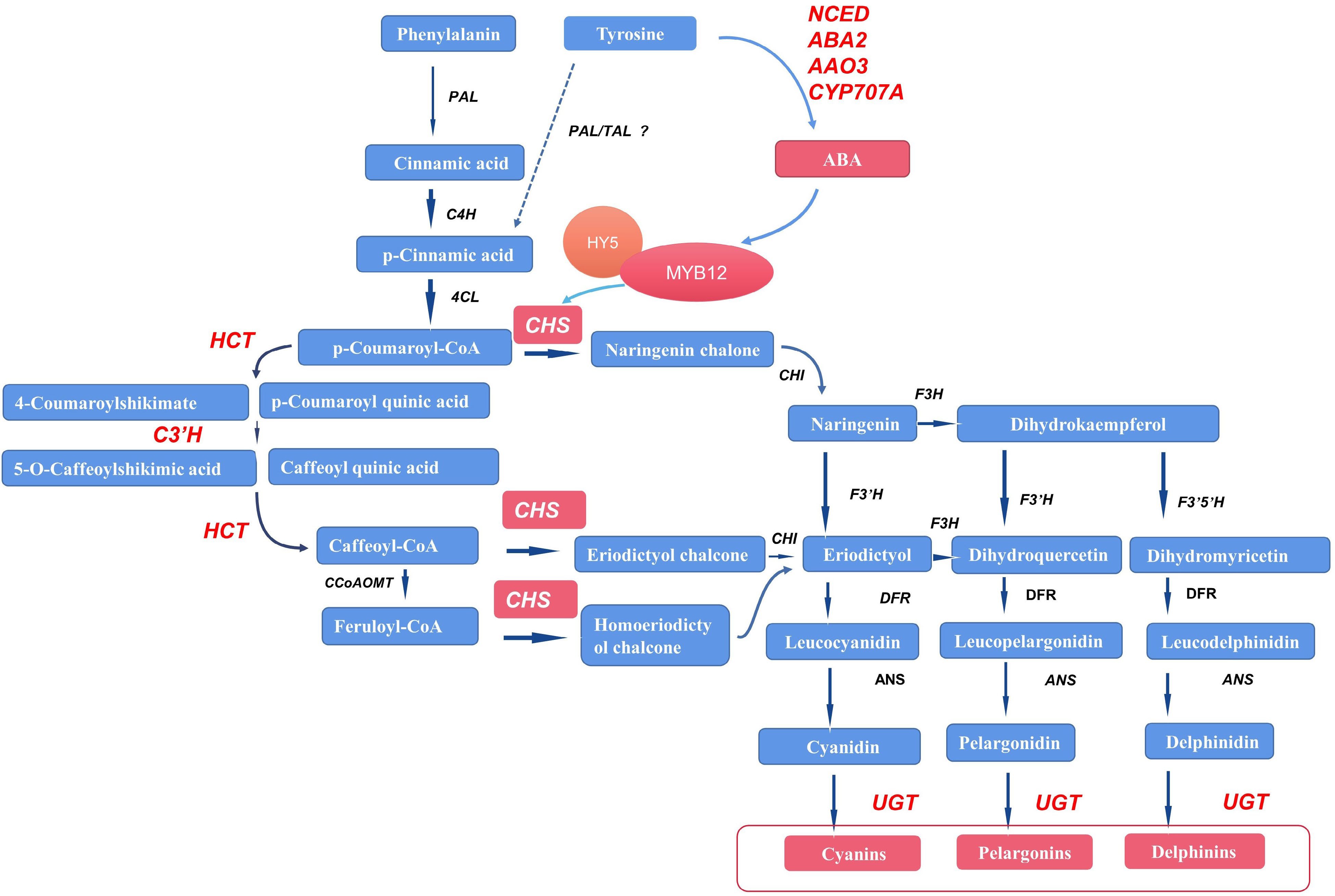
Figure 8.
Possible pathways by which tyrosine treatment may promote anthocyanin biosynthesis in Viola × wittrockiana. The red color represents upregulation of genes and metabolites. The dashed arrow represents the possible pathway. PAL/TAL indicates that PAL may function like TAL in pansy, although this remains to be verified. PAL: phenylalanine ammonia-lyase; TAL: tyrosine ammonia-lyase; C4H: cinnamate 4-hydroxylase; 4CL: 4-coumarate-CoA ligase; HCT: shikimate O-hydroxycinnamoyltransferase; C3′H: 5-O-(4-coumaroyl)-D-quinate 3'-monooxygenase; CHS: chalcone synthase; F3H: naringenin 3-dioxygenase; F3′H: flavonoid 3'-hydroxylase; F3′5′H: flavonoid 3',5'-hydroxylase; DFR: flavanone 4-reductase; ANS: anthocyanidin synthase; UGT: UDP glucuronosyltransferase, falconoid 3-O-glycosyltransferase; CCoAOMT: caffeoyl-CoA O-methyltransferase; VwNCED: 9-cis-epoxycarotenoid dioxygenase; VwABA2: xanthoxin dehydrogenase; VwAAO3: abscisic-aldehyde oxidase; VwCYP707A: (+)-abscisic acid 8'-hydroxylase.
Another possible pathway is that exogenous tyrosine promotes the production of p-coumaric acid, which then leads to increased anthocyanin content. Tyrosine is known to give rise to p-coumaric acid through the catalysis of tyrosine ammonia-lyase (TAL); p-coumaric acid is then acted upon by 4-coumarate-CoA ligase (4CL) to form p-coumaroyl CoA, the main precursor of the anthocyanin biosynthesis pathway. When the content of tyrosine increases, p-coumaric acid production may be upregulated, thus promoting anthocyanin biosynthesis. However, no unigenes in the Viola × wittrockiana transcriptome were annotated as TAL in the present study. We speculate that phenylalanine ammonia-lyase of pansy (VwPAL) may have the same function as TAL, as PAL has been shown to function like TAL in some other plants[42]. However, more experiments are needed to verify this hypothesis.
This research was supported by the National Natural Science Foundation of China (Grant No. 32160719, 32060365 and 31760590), the Natural Science Foundation of Guizhou Province (No. ZK [2022] 095), and the Cultivation Research Program of Guizhou University (No. [2018] 5781).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/TP-2022-0009)
-
Received 7 September 2022; Accepted 2 November 2022; Published online 29 November 2022
-
Tyrosine promoting the anthocyanin biosynthesis in pansy.
Tyrosine decarboxylase gene of pansy (VwTYDC) expressed differently in blotches and non-blotched areas of petals.
Tyrosine induced ABA synthesis and promoted anthocyanin biosynthesis in pansy petals.
-
# These authors contributed equally: Zheng Cui, Jia Gu
- Supplementary Table S1 Primers used for the construction of pTRV2 vectors for VIGS.
- Supplementary Table S2 Primer used for qPCR.
- Supplementary Table S3 Contents of anthocynidins, flavonoids and procyanidin in tyrosine or water treatment as determined by ESI-HPLC-MS/MS
- Supplementary Table S4 Differentially expressed MYB transcription factors.
- Supplementary Figure S1 Gene ontology annotation of the Viola × wittrockiana transcriptome data.
-
Supplementary Figure S2
Volcano plot of differentially expressed genes (DEGs, FDR<0.05 and="">1) between the 3 mM Tyrosine and ddH2O treatments. Red, green, and black dots represent up-regulated DEGs, down-regulated DEGs, and non DEGs.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Cui Z, Gu J, Li J, Zhao A, Fu Y, et al. 2022. Tyrosine promotes anthocyanin biosynthesis in pansy (Viola × wittrockiana) by inducing ABA synthesis. Tropical Plants 1:9 doi: 10.48130/TP-2022-0009 

Tyrosine promotes anthocyanin biosynthesis in pansy (Viola × wittrockiana) by inducing ABA synthesis
- Received: 07 September 2022
- Accepted: 02 November 2022
- Published online: 29 November 2022
Abstract: Viola × wittrockiana (pansy) is an important ornamental plant, particularly during winter and spring. In previous studies, we found that the tyrosine decarboxylase gene of pansy (VwTYDC) was expressed differently in blotched and non-blotched areas of pansy petals, suggesting that tyrosine may have a role in anthocyanin biosynthesis. In this study, we found that virus-induced gene silencing of VwTYDC caused an accumulation of pink pigmentation in pansy petals. Likewise, exogenous tyrosine treatment (TYRT) induced the formation of black stripes in non-blotched petal areas. Metabolome analysis indicated that the contents of two anthocyanins, cyanidin-3-O-glucoside and cyanidin-3-O-rutinoside, increased significantly in the TYRT areas. RT-qPCR results revealed that the anthocyanin-related genes VwHCT, VwC3′H, VwCHS, and VwUGT were upregulated in the same areas. Transcriptome analysis revealed that four genes involved in the abscisic acid (ABA) biosynthesis pathway (VwNCED, VwABA2, VwAAO3, and VwCYP707A) were significantly upregulated in the same TYRT areas. ABA content was measured by ESI-HPLC-MS/MS, and ABA content was significantly higher in TYRT areas than in control areas. In addition, when exogenous ABA was spread onto non-blotched petal areas, anthocyanin biosynthesis genes were upregulated just as with tyrosine. Thus, transcriptome and metabolite analyses revealed a possible novel regulatory network for anthocyanin biosynthesis in which tyrosine induces ABA synthesis and ABA then promotes anthocyanin biosynthesis in pansy petals.
-
Key words:
- Tyrosine /
- Transcriptome /
- Anthocaynins metabolome /
- Abscisic acid /
- Viola × wittrockiana