









-
The normal growth of plants is the premise of survival and propagation, enabling crops to mature in time and obtain the expected yield. Light serves as the source of energy and developmental switch throughout the plant life cycle, thereby dramatically determining plant growth and architecture[1]. Plants alter their morphological and growth features responding to light signals of varying intensity and quality for their survival and optimal growth[1,2]. Ultraviolet (UV) radiation occupies a minor fraction of sunlight, yet it imparts many positive and negative effects on plant growth[2]. The increasingly enhanced UV radiation in solar radiation due to global warming would potentially induce DNA damage[3]. DNA damage affects cellular functionality and disturbs the normal growth of the whole organism[4]. Plants are constantly exposed to endogenous cellular processes and exogenous environmental events, which can impair genomic integrity through DNA damage[5]. Effective DNA damage detection mechanisms that enable the cells to quickly activate repair mechanisms are crucial for maintaining genomic integrity[6].
All organisms possess the mechanisms for repairing DNA damage[7]. Because of the sessile lifestyle, plants have developed many ways to protect themselves from UV radiation in sunlight. For example, some UV absorbing pigments are synthesized in plant leaves; various DNA repair mechanisms are expressed to repair UV induced damage. The repair of UV-damaged DNA is mainly performed by two pathways, light-dependent repair and light-independent (dark) repair[8]. Light-dependent repair utilises photoreactivation, which is primarily a form of repair for lower organisms and as organisms evolve. Light-independent repair is known as nucleotide excision repair (NER)[9], which does not directly reverse the DNA damage but replaces DNA damage with undamaged nucleotides[10,11]. In Arabidopsis, RAD23, DDB1A, CSA, and XPB are involved in this pathway, and DDB1A is recognized as a key gene essential for the DNA repair process[12−15].
Damage DNA Binding Protein 1A (DDB1A) is involved in the damage recognition stage of the NER pathway. DDB1 was first identified in patients with human xeroderma pigmentosum who are hypersensitive to UV-B radiation because of defects in DNA repair[16−18]. Subsequently, the homologous gene of DDB1 was found in plants such as Arabidopsis, tomato, and rice. Arabidopsis has two highly related DDB1 proteins, named DDB1A and DDB1B. A null mutant (ddb1a) of DDB1A was constructed in Arabidopsis and this mutant showed some different phenotypes from the wild-type plants; for example, the ddb1a mutant had significantly longer hypocotyls, more lateral roots, and fewer rosette leaves and the shoot height was significantly reduced compared to that in the wild-type[13]. Bernhardt et al.[13] revealed that DDB1A is critical for embryo development and affects whole plant development. Two high-pigment-1 (hp-1) and high-pigment-1w (hp-1w) mutants were obtained in tomato with the phenotype displaying higher anthocyanin levels, shorter hypocotyls, and enhanced fruit pigmentation compared to their semi-isogenic wild-type counterparts[19]. Lieberman et al.[20] found that the higher anthocyanin level phenotypes in hp-1 and hp-1w were controlled by the same gene, DDB1. Ishibashi et al.[21] cloned DDB1 in rice, with resulting characterization indicating that DDB1 is mainly expressed in proliferative tissue, with increased expression after UV radiation. These results indicated that DDB1 expression is related to cell proliferation and might primarily be required for DNA repair during DNA replication. Further, DDB1A performs not exactly the same function in different species.
Plant mutants exhibiting growth retardation-related phenotypes such as diminished growth and sensitive responses to UV radiation are conductive to characterize the function of genes linking growth to light-dependent DNA repair[2,22−25]. In this study, we screened two allelic mutants with retarded growth, grm1 and grm2, from EMS-mutagenized populations of Chinese cabbage wild-type 'FT'. These mutants exhibited stunted vegetative growth with smaller leafy-heads, and their mutated trait stably inherited. Their causal gene was mapped to the BraA09g024830.3C (BrDDB1A) on A09 chromosome, which encodes the putative Damaged DNA Binding Protein 1A in Chinese cabbage. The mutants exhibited normal growth and morphology identical with the wild-type in a low UV-radiation environment. Our results demonstrated that BrDDB1A is required for maintaining regular plant growth, which provide evidence for characterizing the functional gene in DNA repair associated with Chinese cabbage growth.
-
The grm1 mutant was obtained by constructing a mutant library as previously described by Huang et al.[26]. Isolated 'FT' microspores from the Chinese cabbage doubled haploid (DH) line were treated with 0.16% EMS for 15 min[27]. grm2 with the same phenotype as grm1 was selected from the mutant library constructed with 0.8% EMS-treated germinated 'FT' Chinese cabbage seeds. The stable inheritance of grm1 and grm2 mutants was exhibited after multiple generations.
When the third true leaf appeared in grm1 and grm2 mutants, the leaf length, leaf width, and plant width of five plants were measured every 3 d for a total of 22 d. The fresh weight and dry weight of three plants were measured every 6 d for 45 d. At the heading stage, the head length, head width, and head weight of five plants were measured.
UV sensitivity assays
-
To investigate whether the growth of two grm mutants would be sensitive to UV, we simultaneously sowed 100 grm1, 100 grm2 and 100 wild-type 'FT' under the same environmental conditions, and randomly selected 50 plants of each at the seedling stage (3rd leaf stage, 18 d after seeding) as the control and the experimental group, respectively. The control group (50 grm1, 50 grm2 and 50 wild-type 'FT') was grown in the greenhouse under natural conditions (day length of 16 h/8 h, 22–25 °C/15–18 °C day/night regime). The experimental group (50 grm1, 50 grm2 and 50 wild-type 'FT') was grown in an artificial climate chamber for plant growth, in which photoperiod and ambient temperature were in accordance with those of the control group. The growth of the two plant groups was observed from the seedling stage (18 d after seeding) to the rosette stage (40 d after seeding). The average maximum UV intensity perpendicular to the plants surfaces under the two conditions was 1.89 mW/m2 and 0.01 mW/m2, as measured with a UV meter (Tenmars-TM213).
Scanning electron microscopy (SEM) observation
-
The third true leaf of grm1 and grm2 mutants were observed by SEM (Hitachi, Japan) as previously described[28].
Genetic analysis
-
To investigate the genetic inheritance of grm1 and grm2 mutants, F1, F2, and BC1 generations were constructed by crossing 'FT' plants with grm1 and grm2 mutants. The segregation ratio of F2 and BC1 generations was analyzed using the Chi-squared (
$\chi^2 $ $\chi^2_{0.05,1} $ Mapping population construction and BSR-Seq analysis of grm1
-
The grm1 mutant was crossed with '15A110', a DH line of Chinese cabbage. In the F2 mapping population (grm1 crossed with '15A110'), two extreme mixed pools, a mutant pool (LM-pool), and a normal pool (WT-pool) were constructed for BSR-Seq. The total RNA was extracted using an RNApure Total RNA Kit (Aidlab, Beijing, China). Pass filter data were obtained on an Illumina HiSeq 2500 platform by GENEWIZ (Suzhou, China). Clean reads were obtained using Cutadapt software (version 1.9.1) to filter low quality data and remove contamination and joint sequences. Clean reads were compared with the reference genome for analysis by Hisat software (v2.0.14)[29]. By comparing the results of each sample with the reference genome, mpileup processing was carried out using SAMtools (v0.1.18) software to obtain the possible single-nucleotide variant (SNV) results of each sample. SNV results were annotated using ANNOVAR (v2013.02.11) software. Euclidean distance (ED) was used to estimate the genetic distance between SNVs and the target trait. ED was calculated as follows:
$ ED=\sqrt{(\mathrm{A}\mathrm{F}\mathrm{ }-\mathrm{ }\mathrm{A}\mathrm{S})2\mathrm{ }+\mathrm{ }(\mathrm{C}\mathrm{F}\mathrm{ }-\mathrm{ }\mathrm{C}\mathrm{S})2+\mathrm{ }(\mathrm{G}\mathrm{F}\mathrm{ }-\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{G}\mathrm{S})2+\mathrm{ }(\mathrm{T}\mathrm{F}\mathrm{ }-\mathrm{ }\mathrm{T}\mathrm{S})2} $ A, C, G, and T represented the corresponding bases in the F-pool and S-pool. For example, AF indicates the frequency of the 'A' base in the F-pool[30]. To eliminate background noise, the ED value of each different SNV site was processed to the power of 5 (ED5)[31]. All ED5 values were sorted and the SNV sites corresponding to the top 1% of ED5 values were screened. Specific chromosome regions were further located according to the distribution of differential SNV sites.
DNA extraction and linkage analysis
-
The cetyltrimethyl ammonium bromide (CTAB) method was used to extract the DNA of two parents (grm1 and '15A110') and 3,022 F2 recessive homozygous plants, according to the method of Murray & Thompson[32] with minor modifications. According to the BSR-Seq result, the simple sequence repeats (SSRs) and insertion/deletion (InDel) markers were designed by Primer Premier5 software and synthesized in GENEWIZ (Suzhou, China). The polymorphic markers were screened for linkage analysis. PCR was used to amplify the DNA, and polyacrylamide gel electrophoresis was used to examine the PCR products as previously described by Huang et al.[26].
Whole-genome re-sequencing
-
For whole-genome re-sequencing, grm1 mutant and 'FT' DNA was extracted using a DNA Secure Plant Kit (Tiangen, Beijing). Two libraries with inserted fragments of 400 bp were constructed and paired-end sequencing was performed on the grm1 mutant and 'FT' libraries using an IlluminaHiSeq sequencing platform and next-generation sequencing technology. Clean reads were obtained by filtering raw reads according to the following requirements: (1) removal of joint contamination using Adapter Removal (version 2)[33]; (2) the sliding window method was used to filter the low-quality reads; (3) removal of reads ≤ 50 bp. BWA (0.7.12-r1039)[34] software was used to compare the clean reads to the reference genome (Brassica_rapaV3.0) using the mem program according to default parameters. GATK software[35] was used for single nucleotide polymorphism (SNP) and InDel detection. ANNOVAR[36] software was used to annotate SNP and InDel loci.
Candidate gene analysis
-
According to the gene sequence information, primers were designed to amplify full-length coding sequences (CDs) and promoter sequences of candidate genes from grm1, grm2, and 'FT' samples. The amplified products were purified using a Gel Extraction Kit (Omega, USA), and then, the purified products were ligated into the pGEM -T Easy Vector (Promega, USA) and finally transformed into TOP10 competent cells (CWBIO, China). The recombinant plasmids were sequenced in GENEWIZ (Suzhou, China). The sequence analysis was performed using DNAMAN software (v6.0.3.99).
RT-qPCR analysis
-
Brgrm1 expression was compared between grm1 and 'FT' samples. Different organs (root, stem, leaf, flower, bud, pod) and leaves from different periods (the cotyledon, the first true leaf, the third true leaf, the sixth true leaf, the rosette leaf, the head leaf) served as templates for RT-qPCR. Total RNA was extracted using an RNApure Total RNA Kit (Aidlab, Beijing, China) and reverse-transcribed into cDNA using FastQuant RT Super Mix (Tiangen, China). cDNA was used as a template with primers for RT-qPCR analysis. Primer sequences are shown in Supplemental Table S1. RT-qPCR products treated with 2× UltraSYBR mixture (Low Rox) (CWBIO, China) were evaluated using the QuantStudio 6 Flex Real-Time PCR System (Applied Biosystems, USA). Three biological replicates were used for each test. Expression levels were calculated using the 2−ΔΔCᴛ method[37] with Actin serving as an internal control.
Bioinformatics analysis
-
The SMART software (
http://smart.embl-heidelberg.de/ ) was used to predict the conserved domains of the candidate gene. The tertiary structure of the protein encoded by the candidate gene was predicted using Phyre2 software (www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index ). Subcellular localization of the protein was predicted via Plant-mPLoc (www.csbio.sjtu.edu.cn/bioinf/plant-multi/# ). The predicted nucleus localization of BrDDB1A was analyzed using INSP[38] (Identification Nucleus Signal Peptide) prediction software at:www.csbio.sjtu.edu.cn/bioinf/INSP/ .Statistical analysis
-
The error values were calculated to measure the accuracy of the data; to reduce error values, three biological replicates were used. Significant differences were assessed using the IBM SPSS Statistics software (v22.0.0.0).
-
grm1 and grm2 mutants both displayed growth retardation with a smaller leafy head compared to the 'FT' phenotype (Fig. 1 and Table 1). The growth curve showed that the growth rate of grm1 and grm2 mutants was slower than 'FT' growth; the difference in leaf length, leaf width, and plant width was significant at 9–15 d (Supplemental Fig. S1a−c). The fresh and dry weights of grm1 and grm2 mutants were dramatically lower than those of 'FT' plants (Supplemental Fig. S1d, S1e). SEM results showed that the number of cells in grm1 and grm2 mutants was higher than those in 'FT' plants at the same location and period. Correspondingly, cell size in grm1 and grm2 mutants was smaller than that in 'FT' plants (Fig. 2).

Figure 1.
Phenotypic characterization of the wild-type 'FT' and grm1 and grm2 mutants. (a) The wild-type 'FT' at seedling stage. (b) grm1 at seedling stage. (c) grm2 at seedling stage. (d) The phenotype of 'FT' (left), grm1 (middle), and grm2 (right) at rosette stage. (e) The phenotype of grm2 (left), F1 (middle, grm2 × grm1), and grm1 (right) at rosette stage. (f) Leafy heads of the grm1 (left) and wild-type 'FT' (right). (g) Leafy heads of the grm2 (left) and wild-type 'FT' (right).
Table 1. Identification of agronomic characters in grm1, grm2 and wild-type 'FT' at the heading stage
Characteristics 'FT' grm1 grm2 Mean of head weight (kg) 0.41 ± 0.05 0.10 ± 0.00* 0.16 ± 0.03* Mean of head length (cm) 12.73 ± 0.68 11.14 ± 0.11* 11.71 ± 0.32* Mean of head width (cm) 9.22 ± 0.82 5.88 ± 0.07* 6.35 ± 0.29* Mean of head length/head
width ratio1.38 ± 0.09 1.89 ± 0.01* 1.84 ± 0.05* 
Figure 2.
Observation of the leaf epidermal cells by scanning electron microscopy (SEM). (a) 'FT' at 200 μm, (b) grm1 at 200 μm, (c) grm2 at 200 μm, (d) 'FT' at 100 μm, (e) grm1 at 100 μm, (f) grm2 at 100 μm.
Allelism of grm1 and grm2
-
The grm1 and grm2 were identified from EMS-mutagenized populations of wild-type 'FT' isolated-cultured microspores and seeds, respectively, but they both exhibited very similar growth retardation phenotype. Therefore, to confirm whether their mutant phenotype might be caused by the same gene, we conducted cross pollination between grm1 and grm2 for testing their allelism. It was found that the phenotype of all 50 plants in the hybrid F1 generation from grm1 plants crossed with grm2 plants was consistent with the mutant phenotype (Fig. 1e, f). This result indicated that grm1 and grm2 were caused by allelic mutation.
Inheritance of mutant trait in grm1 and grm2
-
The genetic inheritance of the grm1 and grm2 mutants was calculated. The segregation ratio of grm1 F2 and BC1 generations are listed in Table 2 and the segregation ratios of grm2, F2, and BC1 generations are listed in Table 3. For grm1 mutants, all F1 generation plants showed the 'FT' phenotype. Of a total of 553 F2 individuals, 130 plants displayed the mutant phenotype, whereas 423 plants displayed the 'FT' phenotype; the F2 segregation ratio was approximately 3:1. Of a total of 106 BC1 plants, 45 and 61 plants displayed mutant and 'FT' phenotypes, respectively; the BC1 segregation ratio was approximately 1:1. Overall, these results indicated that the grm1 mutant trait was controlled by a single nuclear recessive gene (Brgrm1). The segregation ratios of grm2 F2 and BC1 generations were also 3:1 and 1:1, respectively. The grm2 mutant trait was thus also controlled by a single nuclear recessive gene (Brgrm2).
Table 2. Genetic analysis of the grm1 mutant.
Generation Total 'FT' grm1 Segregation ratio χ2 P1 ('FT') 50 50 0 P2 (grm1) 50 0 50 F1 (P1 × P2) 50 50 0 F1 (P2 × P1) 50 50 0 BC1 (F1 × 'FT') 90 90 0 BC1 (F1×grm1) 106 61 45 1.356:1 2.424 F2 553 423 130 3.254:1 0.699 Table 3. Genetic analysis of the grm2 mutant.
Generation Total 'FT' grm2 Segregation ratio χ2 P1 ('FT') 50 50 0 P2 (grm2) 50 01 50 1 F1 (P1 × P2) 50 50 0 F1 (P2 × P1) 50 50 0 BC1 (F1 × 'FT') 100 100 0 BC1 (F1 × grm2) 150 78 72 1.083:1 0.24 F2 300 231 69 3.348:1 0.682 Primary mapping of Brgrm1 to chromosome A09 by BSR-Seq
-
The grm1 F2 population was used to map the target region by BSR-Seq. Approximately 40,000,000 clean reads were obtained from the LM-pool and WT-pool. The ED5 value of SNV differences was plotted in the reference genome. The distribution of ED5 values on the chromosomes is shown in Fig. 3. Brgrm1 was mapped to chromosome A09. Using the top 1% of ED5 values as a threshold, five intervals were obtained on chromosome A09 (Supplemental Table S2). SSR markers were designed, two parents (grm1 and '15A110') were used to screen the linkage marker, and 300 F2 individuals were used to verify the target region (Supplemental Fig. S2). Finally, Brgrm1 was mapped between the SSR markers ZMD-9 and ZMD-44 on chromosome A09; the genetic distances were 1.52 cM and 1.79 cM, respectively (Fig. 4a).
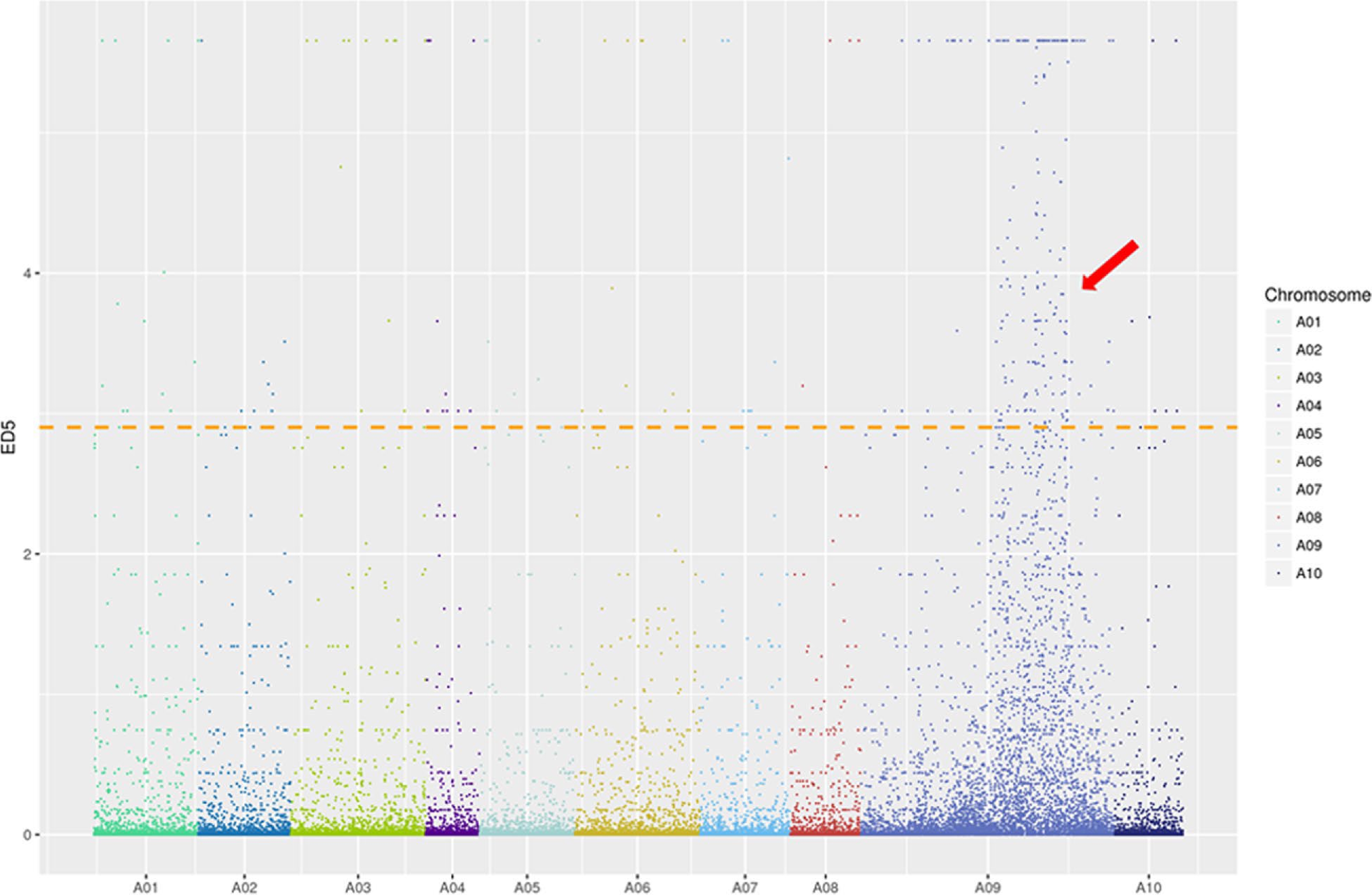
Figure 3.
ED5 (Euclidean distance to the power of 5) distribution of filtered single nucleotide polymorphisms (SNPs) on chromosomes based on bulked segregant RNA-seq (BSR-Seq). Note: X-axis represents the chromosome in Brassica rapa and Y-axis represents the ED5 values of filtered SNPs. The horizontal line is the correlation threshold of the top 1%.
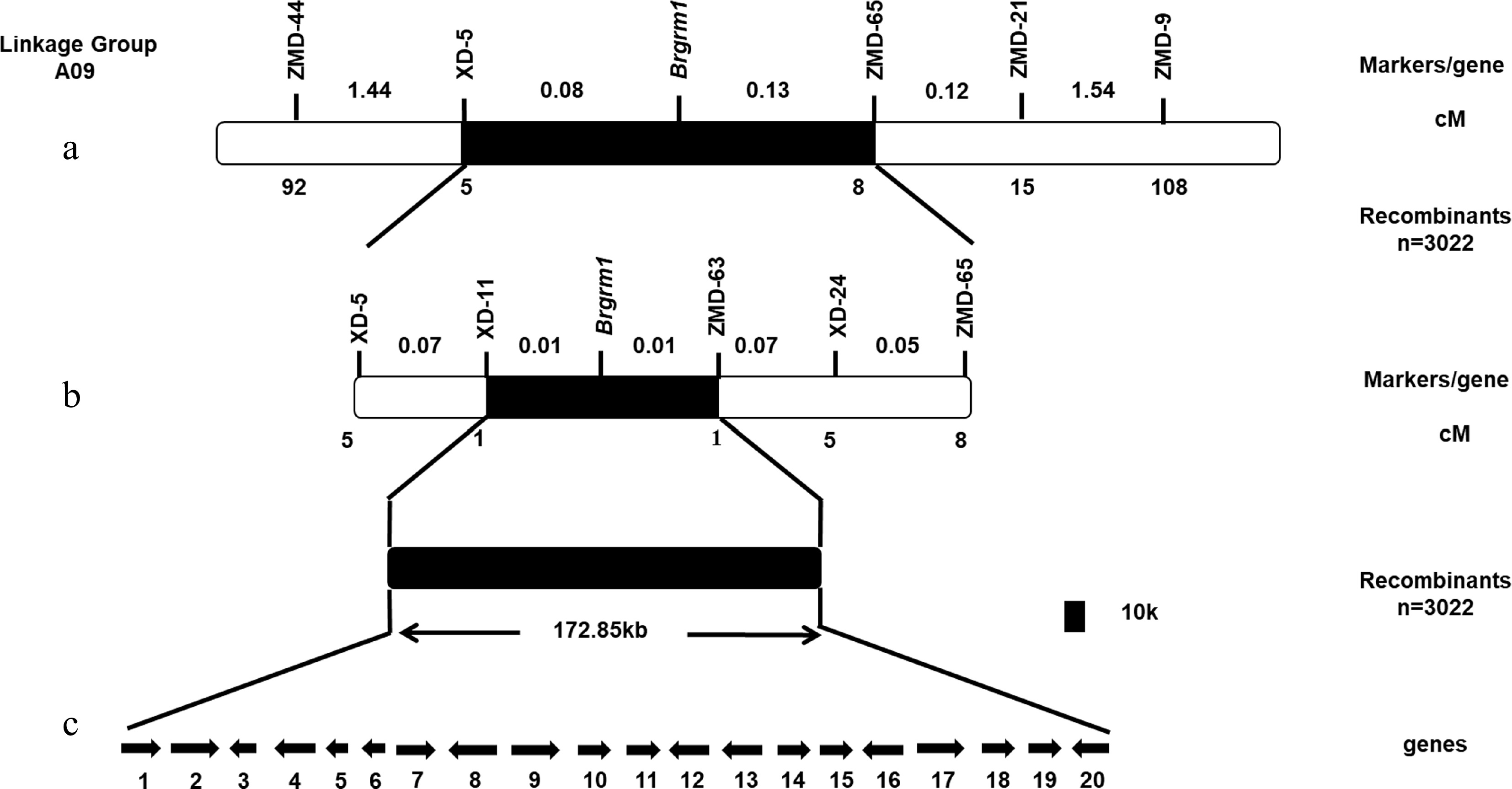
Figure 4.
Construction of genetic and physical maps of Brgrm1 and analysis of candidate genes. (a) Preliminary mapping of Brgrm1. (b) Fine mapping of Brgrm1. (c) Candidate gene analysis in target region. Arrows indicate the direction of gene transcription.
Fine mapping of Brgrm1
-
To map Brgrm1, more SSR and InDel markers were designed between the ZMD-9 and ZMD-44 markers. Of these, six polymorphic markers, XD-5, XD-11, ZMD-63, XD-24, ZMD-65, and ZMD-21, were screened (Supplemental Table S1). A total of 3022 F2 individuals were used to locate Brgrm1. The label of recombinants indicated that XD-5 and XD-11 were located on one side, and ZMD-63, XD-24, ZMD-65, and ZMD-21 were located on the other side of Brgrm1. Finally, Brgrm1 was mapped between the XD-11 and ZMD-63 markers with a physical distance of 172.85 kb. Both XD-11 and ZMD-63 markers had one recombinant, and the genetic distance was 0.01 cM (Fig. 4b).
Identification of the mutant gene by whole-genome re-sequencing
-
Twenty genes were contained in the target region (Fig. 4c and Supplemental Table S3). Whole-genome re-sequencing was used to predict the candidate gene. In total, 67,518,344 and 93,975,642 clean reads were obtained from the grm1 mutant and 'FT', respectively. Comparisons of the results between grm1 and 'FT' provided 1,075,244 SNPs and 389,539 InDels from the grm1 mutant and 1,363,821 SNPs and 368,451 InDels from 'FT' samples. The criteria for mutation screening were 1) the mutations occurred in the target region and 2) the mutations were homozygous and non-synonymous. The results suggested that one InDel was screened in the target region of the grm1 mutant. A base deletion (A) occurred in the 17th exon of BraA09g024830.3C. Gene annotation indicated that BraA09g024830.3C encoded the Damaged DNA Binding Protein 1A, which plays a role in damaged DNA repair. Therefore, BraA09g024830.3C was regarded as the candidate gene.
Cloning and co-segregation analysis of the candidate gene
-
The primers (Supplemental Table S1) were designed to clone the full length (B-1~B-9) and CDs (CD-1~CD-5) of BraA09g024830.3C from grm1 mutant and 'FT'. Cloning and sequencing results showed that the base (A) deletion in the 17th exon (Fig. 5a) caused a frameshift mutation, ultimately resulting in the termination of amino acid coding (Fig. 5b). Three primers (Pro-1~Pro-3) (Supplemental Table S1) were designed to clone the promoter sequence of BraA09g024830.3C from the grm1 mutant and 'FT'. The result showed that the promoter sequence of BraA09g024830.3C was not different between grm1 and 'FT'. To further validate the mutation locus, two recombinants of the two closest markers were cloned. The result showed that the two recombinants had the same base deletion compared to the sequence of the grm1 mutant in the 17th exon (Fig. 5c). Therefore, this InDel was co-segregated with the growth retardation trait. These results showed that BraA09g024830.3C was the strongest candidate gene.
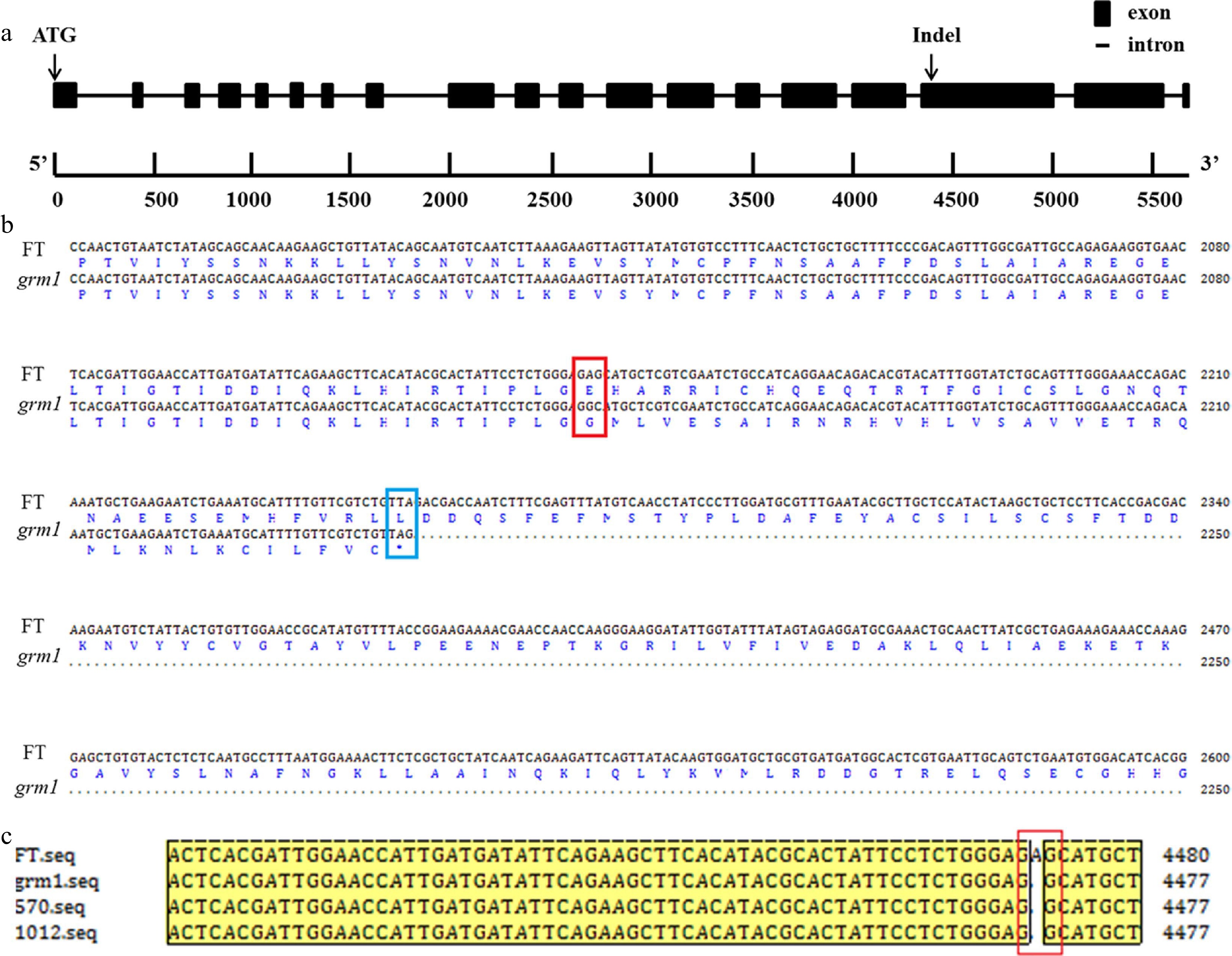
Figure 5.
Gene structure and amino acid sequence alignment. (a) Gene structure of BraA09g024830.3C with an insertion/deletion (InDel). (b) Alignment of nucleotide sequence and amino acid sequence of BraA09g024830.3C from 'FT' and grm1 mutant. The site created by the non-synonymous single nucleotide polymorphism (SNP) is shown by a red empty box and the coding termination is shown by a blue empty box. (c) Alignment of BraA09g024830.3C with nucleotide sequences in the two F2 recombinants, the grm1 mutant, and 'FT'. Note: the two F2 recombinants were from the most closely linked markers, XD-11 and ZMD-63.
Cloning of BraA09g024830.3C in the grm2 mutant
-
As grm1 and grm2 were triggered by the allelic mutations, the promoter sequence and full-length sequence and CDs of BraA09g024830.3C were also cloned from the grm2 mutant. The result showed that the promoter sequence of BraA09g024830.3C was not different between grm2 and 'FT'. The resulting full-length clone indicated that a base substitution (G to A) occurred in the first intron of grm2 (Fig. 6a, b). The cloning and sequencing of CDs showed that the first intron with 263 bp was retained in grm2 and resulted in a frameshift mutation, ultimately leading to a termination amino acid codon (TAA) (Fig. 6c). These results showed that BraA09g024830.3C was the causal gene, which accounts for the phenotype of growth retardation in Chinese cabbage, and it was named BrDDB1A.

Figure 6.
Gene structure and nucleotide sequence alignment. (a) Gene structure of BrDDB1A in grm2 with a single nucleotide polymorphism (SNP). (b) Nucleotide sequence alignment of BrDDB1A.3C in 'FT' and grm2 mutant. c 263-bp intron retention occurred in the first intron of BrDDB1A in the grm2 mutant.
Expression analysis of BrDDB1A in the grm1 and grm2 mutant
-
RT-qPCR detection of the BrDDB1A expression levels in grm1, grm2, and 'FT' showed that BrDDB1A expression from different organs (root, stem, leaf, flower, bud, pod) was dramatically lower in grm1 than in 'FT' (Fig. 7a), and BrDDB1A expression in grm1 and grm2 leaves from different periods (the cotyledon, the first true leaf, the third true leaf, the sixth true leaf, the rosette leaf, the head leaf) was significantly lower than that in 'FT' (Fig. 7b, c).
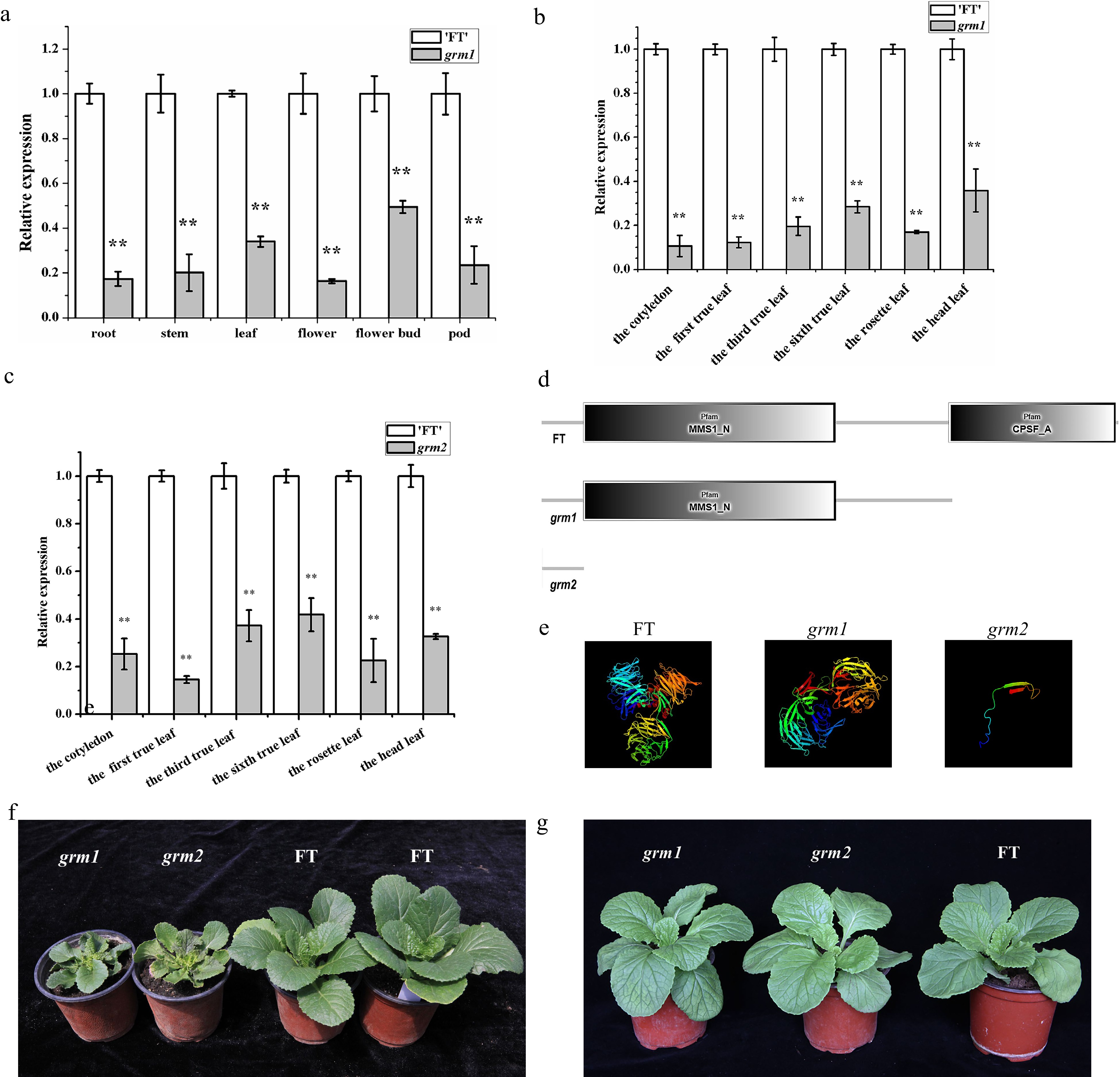
Figure 7.
The expression and protein analyses of BrDDB1A. (a) The expression pattern of BrDDB1A from different organs of 'FT' and grm1. (b) The expression pattern of BrDDB1A at different developmental periods from 'FT' and grm1 leaves. (c) The expression pattern of BrDDB1A at different developmental periods from 'FT' and grm2 leaves. Asterisks indicate significant differences among grm1, grm2 and 'FT' (t test, P < 0.05). (d) The analyses of conserved domains of BrDDB1A in 'FT', grm1, and grm2. (e) Protein tertiary structure of BrDDB1A in 'FT', grm1, and grm2. (f) The wild-type 'FT', grm1 and grm2 plants were cultured in the greenhouse with 1.89 mW/cm2 UV radiation. (g) The wild-type 'FT', grm1 and grm2 plants were cultured indoors with 0.01 mW/cm2 UV radiation.
Bioinformatics analysis of BrDDB1A
-
The SMART software showed that the conserved domain of BrDDB1A was different in the two mutants from that in 'FT' (Fig. 7d). BrDDB1A had two conserved domains, MMS1_N and CPSF_A in 'FT'; in contrast, it had only one conserved domain, MMS1_N, in grm1 and had no conserved domains in grm2. Additionally, the Phyre2 results showed that the protein tertiary structure was changed in the two mutants compared with that in 'FT' (Fig. 7e). BrDDB1A in 'FT' contained three β-propeller (βPa–βPc) subdomains: an independent β-propeller (BP) domain (BPB) and a clam-shaped double-propeller fold (BPA-BPC). Conversely, in grm1, the base deletion led to the disappearance of the BPA domain from BrDDB1A and destroyed the structure of the clam-shaped double-propeller fold. Additionally, in grm2, the intron retention resulted in the disappearance of both the BPB and BPA-BPC domains. Importantly, different variations of the same gene in the two mutants led to changes in the BrDDB1A tertiary protein structure. Meanwhile, the subcellular localization of BrDDB1A was predicted via Plant-mPLoc, it was found that BrDDB1A might encode a nucleus-localized protein (Supplemental Fig. S3).
Mutation of BrDDB1A induced UV-sensitive growth
-
It has been reported that overexpression of Arabidopsis DDB1A enhances UV tolerance[11]. To investigate whether the dysfunction of the BrDDB1A in the two grm mutants makes them sensitive to UV, we observed two groups of 50 mutants and 50 wild-type plants that grew in different growth environments. When the mutant and wild-type plants were grown in the greenhouse, the mutant plants exhibited the growth retardation phenotype (Fig. 7f). By contrast, when the mutant and wild-type plants were grown indoors, the mutant plants showed wild-type traits, and there were no significant differences between wild-type and mutants in terms of phenotypes (Fig. 7g).
-
Screening mutants by EMS mutagenesis has been widely applied to numerous plants including Arabidopsis, rice, wheat, and Chinese cabbage[39−42]. EMS mutagenesis is advantageous because of its high mutation efficiency and diversity of single base mutations and because it results in saturation mutagenesis. In this study, two allelic mutants (grm1 and grm2) were identified from the EMS-mutagenized populations of Chinese cabbage 'FT', whose growth retardation were triggered by the outation of BraA09g024830.3C (BrDDB1A). EMS is an alkylating agent that largely accomplishes the mutagenesis process as follows: first, the oxygen of the guanine (G) is alkylated. Then, G is paired with thymine (T) during DNA replication. Finally, a base substitution occurs (G:C to A:T). This single base mutation is the main form of EMS mutagenesis. Herein, the grm1 mutant was obtained by EMS treatment of isolated microspores and its mutated phenotype was caused by the termination of amino acid coding resulted from the deletion of a base (A) in the 17th exon of BrDDB1A. However, the base deletion/insertion mutants did not conform to those of typical EMS mutagenesis. A previous study reported a naturally occurring mutant during microspore culture; Chen et al.[43] obtained a natural stigma exsertion mutant in the isolated microspore culture of ornamental kale. Therefore, we speculated that the grm1 mutant might be a natural mutant produced during the isolated microspore culture process. In the grm2 mutant, we found that a base substitution (G-A) occurred in the last base of the first intron, which conformed to typical EMS mutagenesis and resulted in an intron retention event. We speculated that this might have been due to the mutation of the acceptor site of the 3′-end in the alternative splicing event from AG to AA, which destroyed the GT-AG rule of the splicing site.
Allelism test can be used to verify whether the mutations are allelic, that is, the causal gene is the same while the mutated sites occurring in the causal gene might be distinct. In our study, the grm1 and grm2 were obtained from EMS-mutagenized populations of wild-type 'FT' isolated-cultured microspores and seeds, respectively. Considering that their growth retardation phenotypes are highly similar to each other, and they possess similar genetic characteristics, we carried out the allelism test between the mutants. The method of the allelism test is crossing two mutants to construct an F1 generation and observing the phenotype of the F1 generation. If the phenotype of the F1 generation is a mutant phenotype, the mutant genes of the two mutants are allelic; if the F1 generation recovers to a wild-type phenotype, the two mutant genes are complementary and belong to non-allelic genes. Multani et al.[44] obtained a brachytic2 mutant (br2) in maize. The cloned br2 gene was found to encode a protein that is probably involved in auxin polar transport. Pilu et al.[45] isolated a new brachytic2 mutant (br2-23). The two mutants, br2 and br2-23, were crossed in the allelism test and the F1 generation phenotype was that of the mutant phenotype; these results indicated that the two mutations of br2 and br2-23 are allelic. Our allelism test of the grm mutants (grm1 and grm2) showed that their mutations belonged to allelic mutation. Compared with WT, they displayed consistent mutant phenotypes of stunted growth. The causal gene BrDDB1A was identified in grm1 through BSR-Seq combined with whole-genome re-sequencing, and verified by the allele mutations existed in grm1 and grm2. Thus, we suggested that the mutation of BrDDB1A triggered the growth retardation in grm1 and grm2 mutants of Chinese cabbage.
A previous study showed that DDB1A is involved in the initial damage recognition stage of the NER pathway in response to UV radiation[17,46]. UV stress is considered an important factor affecting the plant growth rate, because the intensity of UV radiation reaching the earth's surface is constantly increasing[47]. UV radiation induces the formation of lesions that can obstruct replication and transcriptional processes and might alter chromatin structure[6,8]. For example, UV radiation can cause a series of morphological changes in rice, such as plant height decreases, leaf thickening, cotyledon curling, stem elongation, leaf expansion, and root ratio (UV radiation plant/normal plant) decreases[48,49]. Increased UV radiation inhibits cotton growth and results in dwarfism[50]. DDB1A recognizes DNA damage and initiates the NER processes[6]. DDB1A might also function to alter chromatin structure and recruit the NER-factor to DNA damage sites[51]. In Arabidopsis, the overexpression of DDB1A increases UV-resistance, and the loss of functional mutant in DDB1A results in UV sensitivity[11]. Likewise, the phenotype of the grm1 and grm2 mutants was consistent with the morphological changes caused by UV radiation in rice[49]. In our study, we observed that the expression levels of BrDDB1A in grm1 and grm2 mutants were significantly lower than that in wild-type plants and there was no difference in the promoter sequence between WT and the two allelic mutants. Previous studies in Arabidopsis (DDB1A), rice (OsUV-DDB1) and Aspergillus nidulans (DdbA) have shown that the mRNA levels of these homologic genes all increased after UV exposure, of which the transcript levels might correlate with the UV-induced DNA repair rate[11,21,52]. Therefore, it was conjectured that the lower mRNA level in grm mutants was affected by the failure of dysfunctional BrDDB1A to respond to UV radiation, while the normal BrDDB1A in WT could trigger the function in UV-induced DNA repair, in turn resulting in the higher mRNA level. We also found an interesting phenomenon that when mutant plants were grown indoors under low UV radiation conditions, the mutants exhibited normal growth resembling the wild-type. Therefore, we speculated that the phenotype of grm1 and grm2 might be caused by UV radiation damage. Moreover, mutations could have led to changes in the structure of BrDDB1A, making it unable to accurately identify the UV radiation damage site; thus, the NER repair pathway would not be correctly initiated, ultimately resulting in the growth retardation phenotype in Chinese cabbage. This hypothesis requires further investigation.
In summary, we identified BrDDB1A as the target gene for the growth retardation in grm1 and grm2 mutants. BrDDB1A might be associated with maintaining normal plant growth under UV-stress in Chinese cabbage. This is the first report indicating that BrDDB1A functions in maintaining regular plant growth of Chinese cabbage. Our study provides insight for further investigation of the regulatory mechanism of BrDDB1A with respect to the growth and development of Chinese cabbage.
The research was funded by the National Natural Science Foundation of China (Grant No. 31730082). We would like to thank Editage (
www.editage.cn ) for English language editing.-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Meidi Zhang, Jiaqi Zou
- Supplemental Table S1 Primer sequences of two mutants.
- Supplemental Table S2 Target region of Brgrm1 by BSR-Seq .
- Supplemental Table S3 Candidate genes within the located region.
- Supplemental Fig. S1 Growth curve of grm1, grm2 and ‘FT’ at seedling stage. (A) leaf length, (B) leaf width, (C) plant width, (D) fresh weight, (E) dry weight.
- Supplemental Fig. S2 Screening of molecular markers tightly linked to Brgrm1. (A) Screening of polymorphic SSR markers between the two parents. (B) Recombinants with ZMD-44 in the mapping population. (C) Recombinants with ZMD-9 in the mapping population. P1: mutant grm1. P2: DH line ‘15A110’. Asterisks indicate recombinants.
- Supplemental Fig. S3 The predicted localization of BrDDB1A was in the nucleus.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang M, Zou J, Huang S, Fu W, Gao Y, et al. 2022. BrDDB1A maintains regular growth of Chinese cabbage via UV tolerance. Vegetable Research 2:17 doi: 10.48130/VR-2022-0017 

BrDDB1A maintains regular growth of Chinese cabbage via UV tolerance
- Received: 07 September 2022
- Accepted: 24 October 2022
- Published online: 29 November 2022
Abstract: Normal plant growth and development ensures the timely maturity and expected yield of crops. The repair mechanism of genome damage under adverse circumstances is essential for maintaining regular plant growth. Herein, two allelic growth retardation mutants of Chinese cabbage, grm1 and grm2, were obtained from EMS-mutagenized populations of wild-type 'FT' isolated-cultured microspores and seeds, respectively. Both mutants stably inherited and exhibited stunted growth with smaller leafy-heads. Genetic analysis and allelism test manifested that the mutated trait was triggered by a same single recessive nuclear gene. Via BSR-Seq, Brgrm1 was mapped to a target region including 20 genes on chromosome A09. Whole-genome re-sequencing revealed that BraA09g024830.3C in grm1 had a single base (A) deletion in the 17th exon, leading to a termination codon (TAG). Genotyping showed that the mutated phenotype co-segregated with the InDel in recombinants of the closest linkage markers. In addition, cloning of BraA09g024830.3C in grm2 found that a base substitution (G-A) occurred in the last base of the 1st intron causing an additional 263-bp retention in coding sequences, which in turn led to a termination codon (TAA). BraA09g024830.3C (BrDDB1A) is a homolog of Arabidopsis thaliana Damaged DNA Binding Protein 1A (DDB1A), a key gene of UV tolerance involved in DNA damage repair. The two mutants exhibited normal plant growth identical with wild-type under an extremely low UV radiation. Our results demonstrated that BrDDB1A contributes to maintaining regular plant growth in Chinese cabbage, which provide insights into elucidating the molecular mechanism underlying Chinese cabbage growth and development.
-
Key words:
- BrDDB1A /
- EMS /
- Plant growth /
- Chinese cabbage /
- BSR-Seq /
- Allelic mutation