








-
Cucumber (Cucumis sativus L.) is an important vegetable crop in Cucurbitaceae and is widely planted worldwide. According to the statistics of the world Food and Agriculture Organization (FAO), the production area of cucumber in China was 1.27 million hectares with a total output of 73.36 million tons in 2020 (
https://www.fao.org/faostat ). Cucumber fruits are an indispensable part of human's dietary structure, which are consumed immature either fresh or pickled[1−3]. In the Asian market, cucumber has been adapted to certain morphological traits such as skin color, firmness, crispness, taste quality among others. Among them, the fruit peel color or skin color of fresh market cucumber is a master trait affecting consumer acceptance and marketability. For example, in Northern China markets, dark green fruit peel is more popular than light green peel fruit, which is also an important breeding target in cucumber.In terms of its diversity, the color of immature cucumber fruit displays a wide range of fruit peel colors. Currently, the peel colors that have been studied and reported include dark green, light green, yellow green, and white[4−7] (Fig. 1). In recent times, many genetic studies have focused on the peel color characteristics of cucumber fruit. Previous studies concerning cucumber fruit skin color mainly focused on inheritance and gene primary mapping. Six major genes such as w/CsAPRR2, D/CsDull, B/CsMYB60, lgp/CsARC5, lgf/CsYcf54, ygp/CsMYB36 have been cloned that control the color of fruit peel. And these genes were highly expressed in ovary and pericarp. Although most of the genes such as CsARC5, CsYcf54, CsAPRR2, CsMYB36 and CsMYB60 regulate metabolism pathway of chlorophyll and flavonoids to affect the peel color, there are few studies on the molecular regulation mechanism of cucumber pericarp color formation. However, there are few systematic reviews on the research progress of genetic rule and mechanism of cucumber peel color recently. Here, from careful analysis of recent literature, we summarized the research progress on genetic regulation and molecular mechanism of cucumber peel color, hoping to provide the clues to understanding the metabolic and functional roles of pigments biosynthesis pathways for other cucurbit crop breeding.

Figure 1.
Representative immature fruit peel colors in commonly consumed cucumber varieties.
-
The epidermis of cucumber fruit, formed by growth and development after ovary wall differentiation, is referred to as the pericarp[2]. The pericarp consists of exocarp, mesocarp and endocarp. In cucumber fruit, it is generally considered that the pericarp refers to the exocarp. Since there are different requirements for peel color in domestic and overseas markets, so it is of great practical significance to improve peel color according to different market needs. Like other vegetable crops, immature cucumber fruit have different peel colors ranging from light green to white[8−13].
Generally, the colors of the peel mainly depend on the content and formation of pigments[7]. Because the peel contains many secondary metabolites, such as pigments, tannins, aromatic compounds, and so on, it can have a wide range of colors[14,15]. In most plants, the content of chlorophyll, carotenoids and flavonoids have become the main factors affecting the color of plants. Currently, in the research of cucumber, information has been reported regarding the fruits of candidate genes such w/CsAPRR2, lgp/CsARC5, lgf/CsYcf54, ygp/CsMYB36, which affects the skin color by regulating chlorophyll, and B/CsMYB60, which regulates the emergence of orange peel by flavonoids[4−7]. Xishuangbanna cucumber and orange-yellow watermelon have a high β-carotene content, which gives their flesh an orange or yellow color[16]. Recent research has also revealed that a vast number of transcription factors control fruit ripening, which in turn affects the accumulation of carotenoids, chlorophyll, and flavonoids. For example, the GOLDEN2-LIKE(GLK) transcription factor regulating chlorophyll levels can affect pepper[17], tomato[18] and Arabidopsis thaliana[19]. Manoharan et al. determined that CYC-B can regulate carotenoid content to produce orange tomatoes[20]. SiMYB12 also plays a vital role in regulating flavonoids and leads to the production of pink tomatoes[21, 22]. Gu et al. found that rice seed coat color and dormancy genes are controlled by pleiotropic genes, which regulate the color of the peel by regulating the formation of flavonoids[23]. In addition, the sikkim cucumber (Cucumis sativus var. Sikkimensis) is featured with some morphological traits like brown fruit with fine and heavy netting. The major locus B, H, and Rs that regulate the shape and color of the pericarp have so far been identified. Periderm tissue formation in the sikkim cucumber is coordinated by spatiotemporal interplays between cutin, suberin and lignin pathways[24, 25].
Current studies have also shown that the color of cucumber peel changes due to differences in pigment content, mainly chlorophyll and flavonoids. For example, Liu et al. found that the chlorophyll content and the number of chloroplasts in the white peel material Q24 were significantly reduced compared with the fruit green peel material Q30, and eventually resulted in a white peel color[6]. Zhou et al. found that the alteration of the ARC5 gene on chromosome 7 affected the development of chloroplasts, which in turn led to bright green peels[5]. The study of Lun et al. showed that the mutation of CsYcf54 gene on chromosome 6 affects the synthesis of chlorophyll and leads to the decrease of chlorophyll in the cucumber fruit peel showing the light green phenotype[4]. Song et al. located a cucumber peel irregular stripe gene IST, which may participate in chlorophyll synthesis and affect the irregularity of peel stripes[26]. In recent research, Chen et al. found the total content of these flavonoid metabolites in L14 old peel was 95 times higher than that of L19 old peel and 35 times higher than that of L14 young peel of cucumber. Additionally, key transcription factors that belong to R2R3-MYB, bHLH51 and WRKY23 might be the major drivers of transcriptional changes in the peel between L14 and L19[27−31]. These results suggest that genes which regulate fruit skin color mainly affect the content of pigments such as chlorophyll and flavonoids by participating in chloroplast development, resulting in different fruit skin colors (Fig. 2).
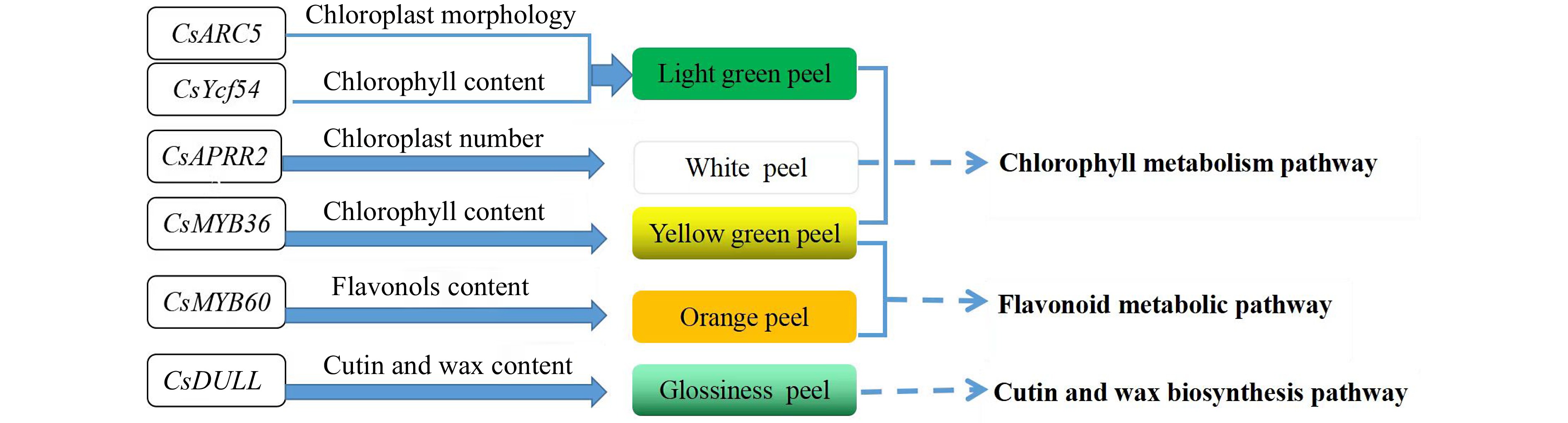
Figure 2.
Regulation network of skin color genes of immature cucumber.
-
With the deepening of genetic research on skin color traits of cucumber commercial fruits, genes controlling skin color traits have been gradually excavated and located. At present, the discovered molecular markers include polymorphic markers ME9EM1-309 and ME9EM14-425, which are 6cM and 8.3cM distant from the green gene of cucumber peel, respectively[32]. Molecular markers E43M61 and E34M59, SSR23517 and SSR23141 linked to the w gene of cucumber white skin trait[33]. The markers SNP2145087 and SNP3286953 were related to lgp[6]. In addition, there are molecular markers SSR10 and SSR37 associated with the skin color uniform(u) gene[34]. What’s exciting are genes related to light green skin, white skin, yellow-green skin and stripe of young fruit have been mainly cloned (Table 1).
Table 1. Genes that control peel color in cucumber were identified in recent years.
Pericarp color character Regulatory gene (ID) Function Reference NCBI reference sequence Yellow green CsMYB36 (Csa2G352940) Interacts with genes related to chlorophyll synthesis Hao et al. 2018[7] XP_004143035.1 White CsAPRR2/W (Csa3G904140) Chlorophyll content and chloroplast number decreased Tang et al. 2018[35] XP_011652667.1 Light green CsYcf54 (Csa6G133820) An important component of cyclase in chlorophyll synthesis Lun et al. 2016[4] XP_004139926.1 Bright green CsARC5 (Csa7G051430) Chloroplast division Zhou et al. 2015[5] XP_004136854.1 Stripe CsIST (Csa1G005490) The number and volume of chloroplasts decreased Song et al. 2020[26] XP_004138283.1 Glossiness CsDULL (Csa5G577350) The regulation of cutin and wax
biosynthesis/transportationZhai et al. 2022[3] NP_001295848.1 Although progress has been made in determination of major gene-controlled cucumber immature fruit peel color, several issues appeared during those processes. For instance, the genetic distance between the molecular markers developed by the genetic population constructed with different parents and the target gene was inconsistent[35]. Even if the same experimental materials were used, due to the difference in the size of the genetic population, the obtained markers and the localization interval of the target genes are different[36, 37], or using the same materials but different mapping methods to obtain the immature cucumber peel color candidate genes which are located not even on the same chromosome[4, 5]. These results indicate that the genetic localization and gene mining of commercial cucumber skin color traits may be restricted by many factors such as population size, marker selection, genetic background and environmental conditions. Therefore, although some genes that regulate the skin color of cucumber fruits have been preliminarily mapped by previous studies (Table 1), due to the relative complexed color traits, the location interval of many important genes is still large, and the target genes are still difficult to be defined, for example, u, which further affects the in-depth research on the function and regulation mechanism of skin color genes in commercial fruits.
Since the 1930s, researchers have noted the diversity of cucumber skin color and the genetic characteristics of those phenotype. Nevertheless, there were still two viewpoints on the genetic rules of skin color genes of commercial cucumber fruits: the one is favored quality traits controlled by a single gene, and the other is favored quantitative traits controlled by major effect genes and modification genes. The possible reasons for the disunity of the above research conclusions are as follows: (1) the investigation results most rely on simple visual results, which tends to lead to the poor accuracy of the standard classification of skin color difference (Fig. 1); (2) In recent years, with the help of a chromatic meter and other means to the determination of pigment content after peeling and cutting fruit skins, it is easy to add pulp to the determination results meaning that pigment content cannot fully represent the color of fruit skins[10−12]; (3) The genetic background and investigation environment, fruit development stages of test materials of separated research groups are different, which may lead to inconsistent conclusions. Although the results differ due to the different genetic background, testing method and growing environment, the skin color character of commercial cucumber fruit should be a relatively simple quantitative genetic character. At present, it has been established physiologically that the commercial color of cucumber is caused by the difference of pigment composition, content and chloroplast number[38]. Some candidate genes controlling fruit color have been successfully mined from the genetic level, but functional verification of gene editing has not been completed (Table 1).
-
Although skin color of immature fruit is an important external quality characteristic of cucumber, the mechanism of their formation is still unclear. Current progress on fruit color study shed light for researchers. In one report, the white pericarp is due to the early termination of the translation of the candidate gene APRR/w by 1 base insertion, which leads to the decrease of chlorophyll content and chloroplast number in pericarp, making the appearance of white pericarp[6]. Different from white pericarp formation, the possible mechanism for the bright green appearance of pericarp is that changes in amino acid translation in the exon region of ARC5 gene affect chloroplast development[5]. The light green pericarp phenotype was caused by a non-synonymous mutation in the coding region of CsYcf54 gene, which affected the synthesis of chlorophyll and led to the decrease of chlorophyll in cucumber fruit skin and leaves[4]. Similarly, CsMYB36 were reported recently may regulate yellow green coloration by interacting with Casparian strip (CsCASP1) and pigment synthesis protein (CsMYC2) involved in peel color development in cucumber[7].
According to the above reports, it is not difficult to find that those genes may lead to chloroplast development or participate in the key steps of chlorophyll synthesis. However, the deeper principle or mechanisms are still lacking. In fact, chlorophyll accumulation in fruits are affected by a variety of factors, such as environmental conditions such as light, temperature and endogenous clues such as chloroplast structural integrity, plant hormones, transcription factors (TFs) expression and so on. Fruit peel color is a quantitative character, and its gene expression is affected by the environment[7]. Therefore, to better understand the CsAPRR2, CsARC5, CsYcf54, CsIST, CsMYB36 gene function will provide an insight into mechanisms explained from a classic chlorophyll and chloroplast development perspective.
Latest results indicate that other cucumber fruit characteristics can have an impact on skin color[39, 40]. For example, the wax layer of cucumber fruit is distributed on the surface of the outer fruit, similar to a layer of white powder, and will cover the original skin color of cucumber to some extent, resulting in visual differences in cucumber skin color. According to recent research, CsDULL, a gene that controls peel gloss, influences the accumulation of peel cutin and wax by regulating two potential target genes, CsGPAT4 and CsLTPG1[3]. Abnormal expression of CsWAX2 in transgenic cucumbers specifically affected both very long chain (VLC) alkanes and cutin biosynthesis. The higher expression of CsWIN1 and wax biosynthesis genes was reflected in the glossier appearance of pericarp, possibly the result of higher wax ester content and higher integration of small trichomes in the pericarp[41, 42]. As a result, it would be a novel approach to studying the mechanism of fruit color to pay attention to changes in visual differences in peel color caused by other factors.
-
Iron-sulfur cluster proteins (Fe-S) are necessary for chloroplast synthesis, chloroplast biogenesis and its key biological functions require sufficient iron-sulfur cluster assembly. Fe-S clusters are important repair groups in organisms. The biosynthesis of Fe-S clusters has been extensively studied in bacteria and yeast[43]. In contrast, much remains to be discovered about the biogenesis of Fe-S clusters in higher plants. It has been reported that the Fe-S cluster is a chemically simple but versatile protein repair group that can be used for photosynthesis, respiration and nitrogen fixation[44, 45]. In chloroplasts, Fe-S clusters play a key role in photosynthesis. Since Cysteine (cys) is determined to be the source of Fe-S formation, NifS-like proteins with cys de-sulfurase activity are likely to participate in this process. NifS is a cysteine de-sulfurase enzyme that converts cysteine into alanine and sulfide and is used to assemble sulfur into Fe-S clusters[46]. In the mitochondrial respiratory complex and the chloroplast photosynthetic process, ferrithioprotein is essential for electron transfer. Additionally, the iron-sulfur repair group is a cofactor for sulfite and nitrite reductase and is believed to involve in the assimilation of sulfur and nitrogen in the chloroplast, respectively[47, 48]. As a matter of fact, potatoes are deficient in sulfur, leaf veins and leaves will turn yellow on a large scale, and the growth rate will be slow[49].
Despite the physiological significance of numerous sulfur-containing compounds, it is still unknown how sulfur atoms are transported and specifically delivered to receptors in various pathways. Higher plants belong to the sulfurtransferases (STRs) family, which has an average of 20 members. The primary sequences, protein domain organization, and subcellular localization of these members vary substantially. The roles of STR5 and STR17a in arsenate tolerance, STR13/CNX5 in Moco biosynthesis and cytoplasmic tRNA thiomodification, and STR1 in cysteine degradation and H2S production are just a few of the functions of STR members. Surprisingly, more research needs to be carried out on the precise function of other STRs, especially those found in chloroplasts[50].
According to earlier research, the rhodanese protein-encoding gene CsYP which regulates peel yellowing, was located by BSA sequencing and molecular markers on the 3,577 (dark green) and 3,578 (light green) cucumber types in our group[51]. The rhodanese (Rhod), which is abundantly found in nature, was initially identified in bovine serum protein in 1933[52]. By directing sulfur atoms to make or repair iron and sulfur clusters in proteins, it can aid in reconstructing or restoring the structure of such proteins[53]. It is widely known that the Fe-S cluster is crucial for the development of the chloroplast structure. Rhodanese-domain proteins are extensively distributed in plants and other creatures, although little is known about most of their biological functions. CRLT, a thiocyanate enzyme required for chloroplast translation, was recently discovered in Chlamydomonas[54]. TROL (thylakoid rhodanese-like protein), a thylakoid membrane nuclear encoding component required for efficient linear electron flow(LEF) in vascular plants, was also discovered in related Arabidopsis thaliana studies[55].
-
Among the existing Cucurbitaceae with more than 800 species known worldwide, cucumber has the smallest genome and the most studied gene function so far[19]. Cucumber research most focused on the biological basis of cucumber fruit development. Cucumber varieties grown and used around the world have rich skin color characteristics. These variation characters facilitate research into the biological basis of fruit color. Cucumber fruit skin color development is a complex biological process regulated by genotypes, transcription factors, endogenous/exogenous hormones, and enzymes. This paper discussed the physiological characteristics, gene mining, and regulatory mechanism of cucumber peel color.
As a new model plant for fruit development, cucumber still has a certain gap with traditional model plants such as Arabidopsis and rice in gene function research. Research reveals that while there have been several investigations into the genetic basis of cucumber peel color features, the majority of them have focused on placement rather than molecular regulation or particular functional analysis. Network analysis of the metabolome and transcriptome has been applied as a powerful tool to investigate the novel genes and pathways in plants, so it is also effective for exploring the novel pathway involved in the formation of yellow peel in cucumber[56, 57]. According to current research, most research on cucumber pericarp color traits focuses on the young pericarp, with little focus on the mature pericarp color traits, particularly the molecular genetic effect. To investigate the genetic mechanism and gene effect of cucumber mature pericarp color is the foundation for future molecular genetic improvement, breeding, and utilization of cucumber pericarp color. Therefore, the next research direction should focus on the exploration of regulation mechanism, further supplement and improve the genetic law of cucumber peel color and provide a technical basis for creating new germplasm to meet market demand.
This research was supported by the Hainan Provincial Joint project of Sanya Yazhou Bay Science and Technology City (320LH042); Sanya Institute of China Agricultural University (SYND-2021-18; SYND-2022-20); and the Construction of Beijing Science and Technology Innovation and Service Capacity in Top Subjects (CEFF-PXM2019_014207_000032), Chinese Universities Scientific Fund 2022TC106. The authors are grateful to members of Ren's lab for technical assistance and discussions.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Cui Y, Li S, Dong Y, Wu H, Gao Y, et al. 2023. Genetic regulation and molecular mechanism of immature cucumber peel color: A review. Vegetable Research 3:9 doi: 10.48130/VR-2023-0009 

Genetic regulation and molecular mechanism of immature cucumber peel color: A review
- Received: 09 September 2022
- Accepted: 11 January 2023
- Published online: 09 March 2023
Abstract: Peel color is an essential exterior fruit quality attribute determining the commodity sale of fresh market cucumber. The color phenotype of cucumber peel are traits which are not only controlled by genetics but also affected by the environment. However, understanding the molecular mechanism of skin color formation in immature cucumbers is still in its infancy. In addition, the latest systematic review on research are also lacking. In this review, we first analyzed the physiological factors that influence pericarp color change. The research progress of cucumber peel color was then reviewed at the genetic and molecular levels, and the existing problems in the research were also mentioned. Meanwhile, this paper presents new insights on fruit skin color in order to provide a reference for the fruit development of cucumber and other Cucurbitaceae crops.
-
Key words:
- Cucumber /
- Gene /
- Molecular mechanism /
- Peel color