









-
Low temperature stress profoundly impairs multiple aspects of the plant life cycle, spanning from seed germination and vegetative growth to photosynthesis, flowering, and fruit setting. Consequently, it leads to a significant reduction in crop yield, quality and other important commercial traits. Moreover, cold stress constrains the geographic distribution of plant species. In the context of tomato, exposure to low temperature stress disrupts cell membrane integrity, leading to electrolyte leakage, compromised photosynthetic efficiency, and reduced carbohydrate accumulation[1].
The CBF-COR signaling pathway plays a pivotal role in plant response to cold stress. This pathway is initiated by the rapid induction of the transcription factor CBF/DREB1 (C-repeat binding factor / dehydration-responsive element binding factor 1) in response to low temperatures. CBFs specifically bind to the CRT/DRE cis-acting element in the promoter regions of cold-responsive genes (CORs), thereby activating COR expression to orchestrate plant growth and development under cold stress[1]. Furthermore, the CBF-COR pathway is subject to modulation by various phytohormones. Abscisic acid (ABA) stands out as one of the most important phytohormones in plant stress responses. The production of the ABA precursor xanthoxin relies on the catalytic activity of NCED (9-cis-epoxycarotenoid dioxygenase), with its activity determining the rate of ABA biosynthesis[2,3]. The gene expression of NCED1 and ABA content can be highly induced to enhance the plant cold tolerance through crosstalk with other phytohormones, such as strigolactones and brassinosteroids[4,5]. In tomatoes, an upstream player of NCED is the tetratricopeptide repeat protein SlREC2 (REDUCED CHLOROPLAST COVERAGE 2), which interacts with SlBCH1b (β-RING CAROTENE HYDROXYLASE 1b) to induce ABA biosynthesis, CBF-COR pathway, and photoprotection activity. This interaction ultimately results in improved cold tolerance in tomatoes[6,7].
Transcriptome analysis has revealed that approximately 10% of ABA-responsive genes in Arabidopsis are up-regulated in response to cold stress[8]. Some COR gene promoters contain ABA-responsive cis-elements (ABRE) targeted by ABRE-binding proteins/factors (AREBs/ABFs)[9]. For instance, ectopic expression of the seed-specific ABRE-binding protein ABI3 triggered the expression of specific COR genes in vegetative tissues, thus enhancing the Arabidopsis freezing tolerance[10]. ABA Overly-sensitive 5 (ABO5) and ABO6 are involved in the ABA signaling pathway in Arabidopsis. ABO5 encodes a pentapeptide repeat protein required for cis-splicing of mitochondrial nad2 intron 3[11], while ABO6, a DEXH-box RNA helicase, mediates the interaction between ABA and auxin signaling[12]. Loss-of-function mutants of ABO5 and ABO6, resulting from primary defects in RNA splicing or editing of mRNAs encoding complex I proteins, show hyposensitivity to cold stress and ABA treatment[13].
SlMETS1, coding a methionine synthase in tomato, has been reported to catalyze the synthesis of methionine (Met) in the folate-dependent single-carbon metabolism, thereby bridging the folate pathway and plant immunity[14,15]. S-adenosylmethionine (SAM), produced from Met by S-adenosylmethionine synthetase (SAMS), serves as a universal methyl donor for DNA, RNA, histone, and protein methylation via methyltransferases[16] SAM is also a precursor for polyamines and ethylene synthesis[17]. Overexpressing SAMS1 was reported to promote the synthesis and oxidation of polyamines, leading to improved H2O2-induced antioxidant protection and enhanced tolerance of transgenic plants to low temperature stress[14]. These findings suggest a positive influence of SAM on the cold resistance in plants.
This study investigates the involvement of METS1, ABO5 and ABO6 in response to low temperature stress in tomato, focusing on their dependence on ABA biosynthesis and signaling pathways. The investigation was conducted on wild-type tomato plants and two ABA biosynthesis-deficient mutants, notabilis (not) and sitiens (sit), which were subjected to cold acclimation or cold stress with normal temperature as the control. We analyzed the correlations between gene expression levels and low temperature tolerance. Furthermore, we validated the involvement of SlMETS1 in the early response of plants to low temperature stress through virus-induced gene silencing (VIGS), and found that the early response is enhanced by SlNCED.
-
Wild-type tomato (Solanum lycopersicum L. 'Moneymaker', Strain ID TOMJPF00002) was ordered from TOMATOMA (Tomato Mutants Archive,
https://tomatoma.nbrp.jp/ ). The ABA defective mutants notabilis (not) and sitiens (sit), with deletion in the gene SlNCED1 (Solyc07g056570) and SlAAO (Solyc01g009230) respectively, were donated by Prof. Dr. Liang Sun in China Agricultural University. Tomato seeds were soaked in water at 55 °C for 15 min and subsequently in distilled water for 12 h before being placed on a wet filter paper in a petri dish for germination in darkness at 28 °C for 48 h. Germinated seeds were then transferred to a substrate mixture consisting of peat, vermiculite, and perlite (2:1:1, v/v/v) in a 50-well tray. The growth conditions comprised a 16-8 h photoperiod, temperatures of 25 °C/20 °C (day/night), and a photosynthetic photon flux density (PPFD) of 600 μmol·m−2·sec−1. Upon reaching the four-leaf stage, seedlings were subjected to either cold acclimation at 12 °C for 1 d followed by exposure to 4 °C or direct cold stress treatment at 4 °C. All other growth conditions remained consistent except for the specified temperature adjustments.Virus-induced gene silencing (VIGS)
-
Tomato virus-induced gene silencing was performed as per Liu et al.[18]. Monoclonal colonies carrying the target construct of TRV1, TRV2 or TRV2-METS1 were preincubated at 28 °C and 200 rpm overnight in 2 mL of LB liquid medium containing appropriate antibiotics. The bacterial solution was mixed with fresh LB liquid medium at a ratio of 1:200 the next day and incubated until reaching an OD600 of 0.8−1.5. After centrifugation at 4,000 rpm for 8 min, bacteria pellets were resuspended in infiltration media (10 mM MgCl2, 10 mM MES, 200 mM Acetosyringone) to OD600 = 1.0. The TRV1 suspension was mixed separately with either TRV2 or TRV2-METS1 at 1:1 (v/v), and incubated at room temperature for 2−3 h.
Two-week-old tomato seedlings were infiltrated with the prepared suspension using a 1 mL needle-free syringe. Infiltration was performed on the abaxial side of two cotyledons and the first leaf of each plant. The infiltrated plants were kept in darkness for 2 d (16 h/8 h, 22 °C/18 °C, day/night, ) before being returned to the previous growth conditions until reaching the four-leaf stage. Leaves of individual plant were collected for RNA extraction and silencing efficiency evaluation. The tomato seedlings with higher than 50% silencing efficiency were used for experiments.
Quantitative analysis of total chlorophyll content
-
The total chlorophyll content was determined as previously described[19]. Briefly, the third leaf of the plants were sampled, excluding midribs. Ten leaf discs with a diameter of 8 mm were collected from three individual plants for each sample. Samples were incubated in 10 mL of 95% ethanol in the dark for 24 h to extract chlorophyll. Absorbance was measured at 652 nm, and the total chlorophyll concentration (C, mg·L−1) was calculated using the formula: Total chlorophyll content (C, mg·L−1) = (1,000 × OD652)/34.5. Each biological replicate included three technical replicates.
Measurement of Relative Electrolyte Permeability (REP)
-
REP was assessed using an established method from Liu et al.[19]. The second leaf from cold-treated tomatoes were immersed in 20 mL of distilled water and shaken at 200 rpm for 2 h before measurement (S1). The same procedure was followed for the control that was distilled water without tomato leaves (S01). Both the test and control samples were boiled for 15 min and cooled to room temperature, then measured again (S2 and S02, respectively). REP was calculated using the formula:
REP (%) = (S1 − S01) / (S2 − S02) × 100
Measurement of malondialdehyde (MDA) content
-
One gram of plant material was ground in 2 ml 10% TCA (Trichloroacetic acid) extraction buffer. The resulting mixture was then transferred to a 10 mL centrifuge tube and brought to a final volume of 10 mL with TCA buffer. Supernatant was collected after centrifuge at 4,000 rpm for 10 min, from which 2 ml was mixed with equal volume of 0.6% TBA (Thiobarbituric acid). The mixture was boiled in a water bath for 15 min, followed by centrifuging at 4,000 rpm for 10 min. Absorbance was measured at 450 nm, 532 nm, and 600 nm. MDA concentration (μmol/L) was determined as follows:
MDA concentration (μmol/L) = (6.45 × (OD532 − OD600) − 0.56 × OD450 ) × reaction liquid volume / extract volume (mL) × 1000.
MDA content (μmol/g) = (MDA concentration × total extract (L)) / sample fresh weight (g).
RNA extraction and RT-qPCR
-
RNA was extracted from the entire true leaves of cold-treated or the control plants. RNA extraction, Reverse transcription reaction, and RT-qPCR were conducted following the procedures previously described[20]. Tomato leaves were collected from three to five individual plants per sample with each plant serving as a biological replicate. Each biological replicate was subjected to at least three technical replicates for RT-qPCR analysis.
-
The role of the phytohormone ABA in cold stress response has been recognized in tomatoes. In this study, we conducted a time-course experiment to explore the phenotypic differences between ABA deficient mutants (not and sit) and the wild type in response to cold acclimation (1 d at 12 °C followed by 4 °C treatment, hereafter referred to as 12 °C − 4 °C) and direct 4 °C cold treatment. Plants exposed to cold stress displayed symptoms of water loss and wilting compared to those under normal temperature. Notably, not mutants showed those symptoms after 6 h under cold acclimation, while sit mutants exhibited them after 12 h (Fig. 1a, b; Supplemental Fig. S1a, b). However, similar symptoms occurred in both the not and sit mutants after 6 h of direct 4 °C cold treatment (Fig. 1c). The severity of wilting escalated over time under both low temperature stress conditions.

Figure 1.
Phenotypes of WT and ABA biosynthesis mutants in response to cold stress. The representative phenotypes of tomato WT, not, and sit mutants under (a) normal growth condition, (b) cold acclimation and (c) cold stress conditions for 0, 2, 6, 12, 24, and 48 h. WT, not, and sit seedlings, reaching the four-leaf stage, were moved to 4 °C for cold treatment (4 °C). Cold acclimation was performed by transferring seedlings to 12 °C for 1 d before moving to 4 °C (12 °C − 4 °C). Seedlings grown at normal growth condition (25 °C) were used as the control. Scale bars indicate 5 cm.
To understand the physiological basis of the observed symptoms, we measured the total chlorophyll content, REP and MDA content in WT, not, and sit plants after cold acclimation and direct 4 °C cold treatments. The total chlorophyll content in not and sit remained unchanged after cold acclimation (12 °C − 4 °C) but was higher than that in the WT (Fig. 2a, c). A similar trend was observed for sit mutant during direct 4 °C cold stress (Fig. 2b). Conversely, not mutant displayed a gradual chlorophyll increase over time and a significant rise after 24 h of direct 4 °C cold treatment (Fig. 2b). REP of not mutants exhibited the highest levels under both cold conditions, followed by WT (Fig. 2d−f). The MDA content in not and sit showed no obvious change compared to the WT under the cold acclimation (12 °C − 4 °C) condition (Fig. 2g). Under 4 °C stress condition, the MDA content in not displayed a gradual increase over time and a sharp rise after 24 h, while sit mutants had a sudden increase at 6 h (Fig. 2b). However, the not showed continuous increase in the MDA content after 12 h under 25 °C normal conditions (Fig. 2i). At the 12-h time point, both the total chlorophyll content and REP of ABA deficient mutants were significantly higher than those of WT during cold treatments (Fig. 2j−l).
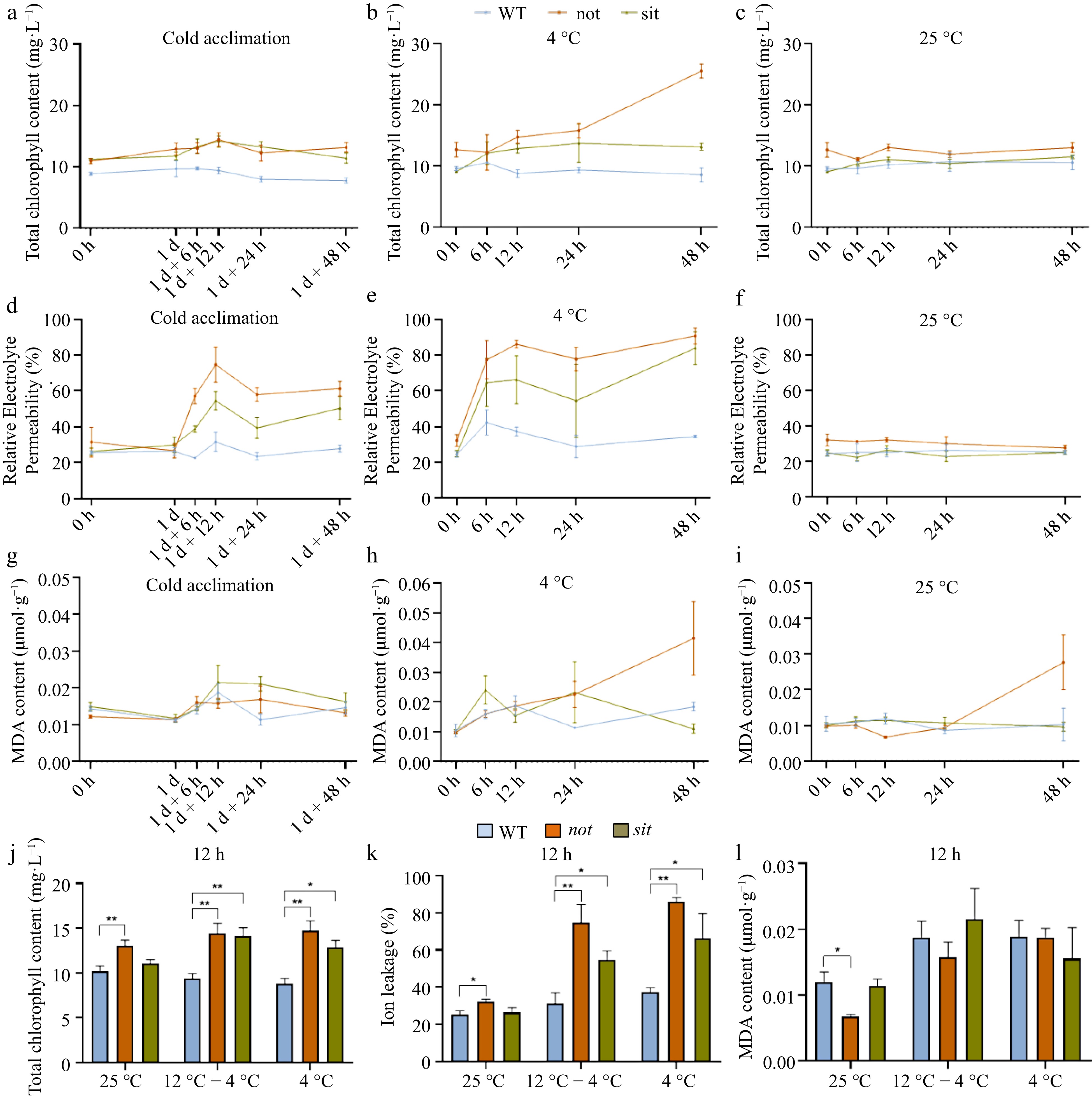
Figure 2.
Cold tolerance evaluation of tomato WT, not, and sit in response to low temperature. WT, not, and sit seedlings, reaching the four-leaf stage, were moved to 4 °C for cold treatment (4 °C). Cold acclimation was performed by transferring seedlings to 12 °C for 1 d before moving to 4 °C (12 °C − 4 °C). Seedlings grown under normal growth conditions (25 °C) were used as the control. For cold tolerance evaluation, time-course experiments were conducted to measure the total chlorophyll content (a)−(c), Relative Electrolyte Permeability (REP) (d)−(f), and MDA content (g)−(i) in WT, not, and sit mutants after low temperature treatment for 0, 6, 12, 24, and 48 h. Bar graphs represent (j) total chlorophyll content, (k) REP, and (l) MDA content in WT, not, and sit mutants at the 12-h from the same time-course experiments as (a)−(i). Values are means of minimum three replicates, with each containing 9 to 10 seedlings. Date are expressed as mean ± standard deviation, and one-way ANOVA followed by Student's t-test was used for significant difference analysis. * p < 0.05, ** p < 0.01, *** p < 0.001.
Overall, these results demonstrate that ABA synthesis deficient mutants were more sensitive to cold acclimation and cold stress than the WT, with the sit mutant exhibiting higher sensitivity to direct 4 °C cold stress than the not mutant (Fig. 2a, b, d, e, h). In addition, these findings support the influence of ABA biosynthesis on plant responses to low temperature stress.
SlNCED may influence SlMETS1 induction in the early cold stress response
-
Grafting is a valuable technique for enhancing the cold tolerance of tomato plants. Through an analysis of various mobile mRNA databases across different vegetable crops, we identified METS1 as a pivotal mobile signal in response to cold stress[20]. Nevertheless, its precise role in cold stress remains largely unexplored, particularly concerning its interactions with the ABA signaling pathway. Therefore, we examined the relative expression of SlMETS1, two well-characterized cold responsive genes (CBF1 and COR47-like), and two genes identified in tomatoes that are involved in ABA signaling (SlABO5 and SlABO6) in WT, not and sit mutants under cold stress. To eliminate the interference from expression variations at 25 °C (Supplemental Fig. S2a−c), we normalized the expression levels under low temperature stress using the room temperature expression as the denominator.
The relative expression of SlMETS1 was slightly down-regulated in WT and sit mutants after 2 h of cold acclimation and 4 °C cold treatment, and then up-regulated, peaking at 6 h before decreasing over time (Fig. 3a, c). In contrast, SlMETS1 showed upregulation in not mutants as early as 2-h time point, suggesting a potential influence of SlNCED on SlMETS1 gene expression during cold stress (Fig. 3b). CBF1 and COR47-like are two cold stress-related genes. SlCBF1 and SlCOR47-like expression significantly increased under cold stress in all genotypes (Fig. 3d−i), consistent with the typical response of plants to low temperature. There were no significant differences in the expression patterns of SlABO5, SlABO6 and SlMETS1 in WT. The trends of expression dynamics of SlABO5 and SlABO6 were similar in not and sit mutants, except for the highest expression occurring at 6 h under cold acclimation and 24 h under direct 4 °C cold treatment. In sit mutants, SlABO5 and SlABO6 expression peaked at 6 h under both conditions, with SlABO6 showing a slight rebound at 24 h under cold acclimation conditiona (Fig. 3j−o).
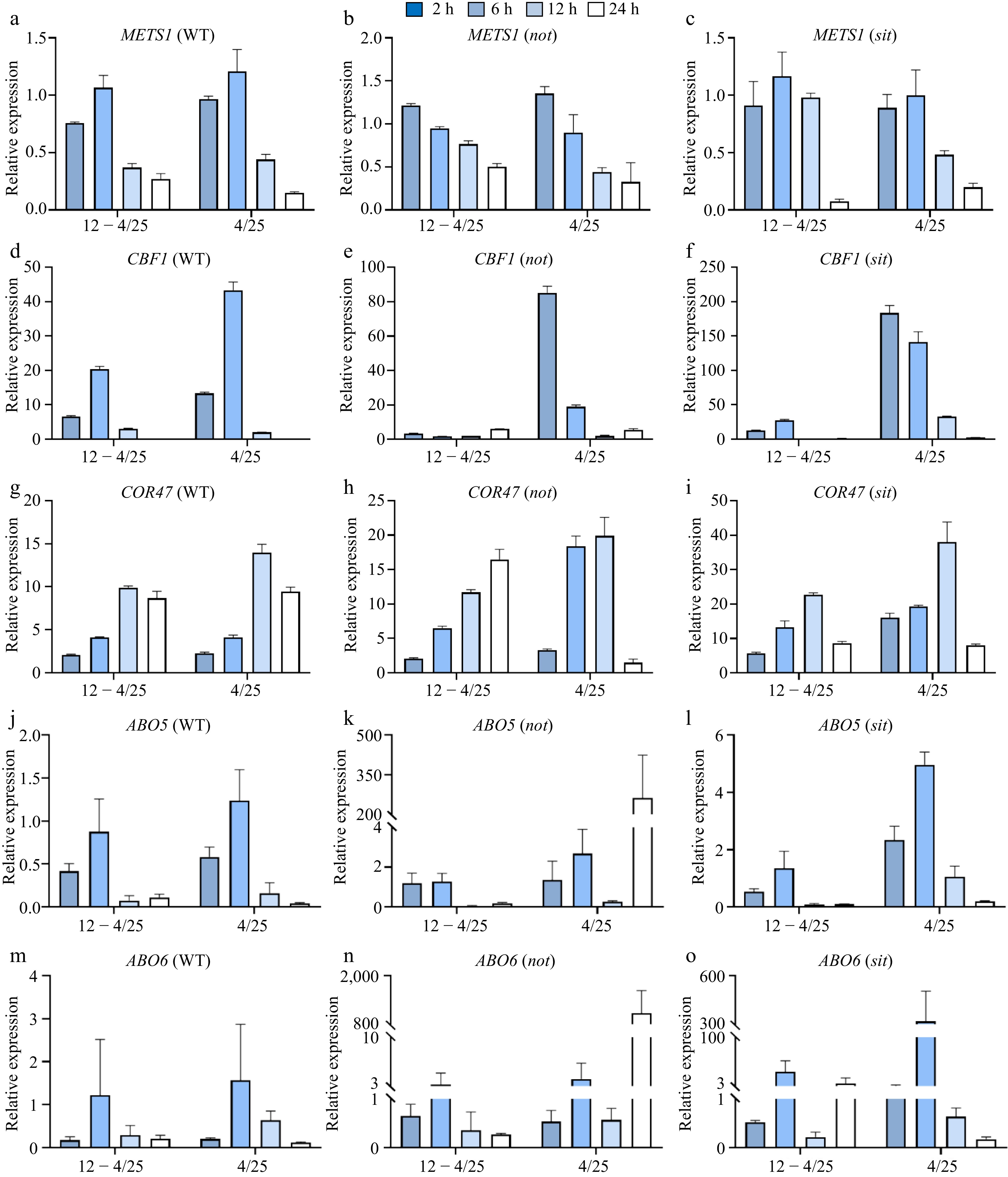
Figure 3.
Expression dynamics of SlMETS1, CBF1, COR47, ABO5 and ABO6 under cold stress. Cold treatments of plants and time-course experiments were the same as described in Fig. 2. Gene relative expression levels were determined by RT-qPCR in tomato WT, not, and sit. The expression levels under cold acclimation and 4 °C cold stress were normalized to room temperature (12 − 4/25, 4/25), and SlEfα was used as internal reference. Relative expression of (a)−(c) SlMETS1, (d)−(f) CBF1, (g)−(i) COR47, (j)−(l) ABO5, and (m)−(o) ABO6, respectively. The experiments were repeated at least three times with similar results.
METS1 reduces damage severity during early cold stress stage
-
To investigate the role of SlMETS1 in low temperature tolerance, SlMETS1 was silenced using VIGS (Supplemental Fig. S3a). Both the control and the SlMETS-silenced plants (hereafter referred to as TRV2-METS1) were subjected to 4 °C low temperature treatment before physiological phenotypes observation and leakage of plasmo membrane measurement. Different from the control plants, TRV2-METS1 plants wilted after 8 h of cold treatment but gradually recovered (Fig. 4a). The changes of REP were similar between TRV2-METS1 and the control plants at both room and 4 °C temperature (Fig. 4b, c), except for a significant increase occurred in TRV2-METS1 after being treated for 8 h and decreasing to the level of the control after 24 h (Fig. 4b, c). The relative expression levels of SlCBF1 and SlCOR47-like significantly increased in response to cold stress in TRV2-METS1 comparing with the control (Supplemental Fig. S3b). Therefore, SlMETS1 inhibition increased the sensitivity of plants to low temperature stress in the early stage, suggesting a positive role of METS1 in mitigating the damage severity caused by cold stress during this stage.
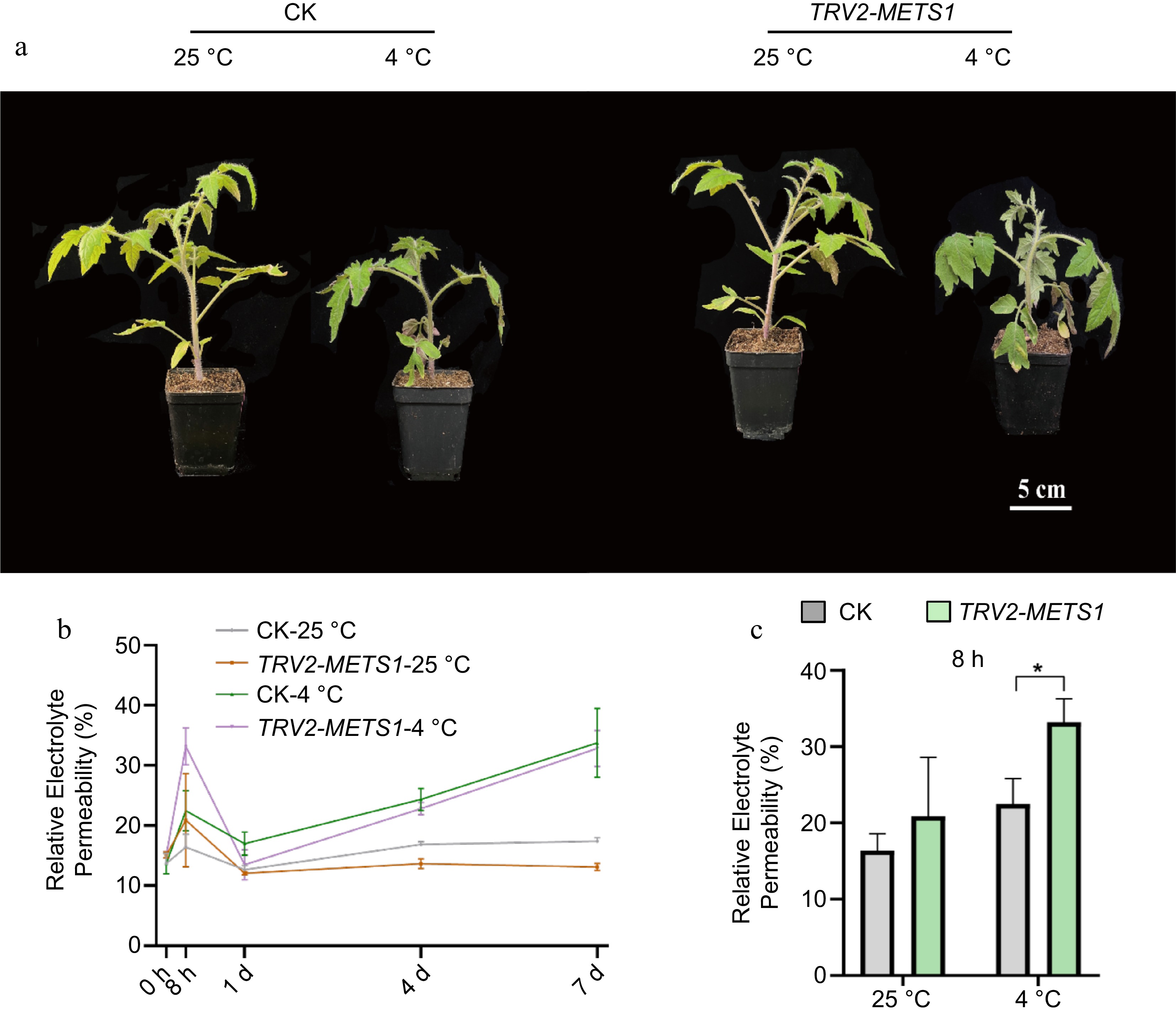
Figure 4.
SlMETS1 inhibition increases sensitivity to early 4 °C cold stress in tomato. SlMETS1 suppression allele (TRV2-SlMETS1) was obtained by VIGS via TRV2-SlMETS1 construct, while the control (CK) was transformed with TRV2 construct without SlMETS1 insertion. (a) Representative phenotype of plants TRV2-SlMETS1 and the CK at 25 and 4 °C for 8 h. (b) Relative Electrolyte Permeability (REP) of TRV2-SlMETS1 and the CK plants exposed to 4 or 25 °C for 0 h, 8 h, 1 d, 4 d, and 7 d. (c) REP of TRV2-SlMETS1 and the CK plants exposed to 4 or 25 °C for 8 h. Represented values were obtained from minimum three replicates, with each containing nine seedlings. REP difference among groups were detected by t-test. *, p < 0.05, **, p < 0.01, n = 9.
-
The pivotal role of ABA in the regulation of cold tolerance has been firmly established. ABA activates leucine zipper (bZIP) transcription regulators that bind to the ABA-responsive element on the COR gene promoter to induce their expression[9,21], thus playing a crucial role in plant's response to low-temperature stress. This study revealed a positive role of tomato methionine synthase SlMETS1 in response to low temperature stress, potentially through ABA and ethylene synthesis pathways. SAMS is vital in catalyzing the formation of S-adenosylmethionine (SAM), a precursor for polyamines and ethylene synthesis. Increased SAM levels promote the synthesis and oxidation of polyamines, thereby enhancing plant cold tolerance[14]. Interestingly, our findings revealed a different expression pattern of SlMETS1 in the not compared to the wild type. In addition, SlMETS1 suppression increased the sensitivity of tomato to low temperature stress during the early stage.
In this study, SlMETS1 was found to be expressed similarly in sit mutant and WT after 2 h of cold acclimation and cold treatment, while it was notably up-regulated in not mutant at the same time point. The not mutant is characterized by the deletion of the SlNCED gene, which encodes a key enzyme in ABA biosynthesis in higher plants. The expression differences suggest a possible interplay between SlMETS1 and SlNCED. SlNCED is a 9-cis-epoxycarotenoid dioxygenase that contributes to tomato fruit ripening[22], salt tolerance[23], drought tolerance[24], pollen ripening[25], pigmentation and carotenoid composition during ripening[26]. The SlNCED family in tomato consists of three members: SlNCED1 (Solyc07g056570), SlNCED2 (Solyc08g016720), SlNCED6 (Solyc05g053530). NCED2 is associated with JA-mediated ABA biosynthesis in tomato plants, thus functioning in response to cold stress. SlNCED2 can be induced by exogenous MeJA and cold temperature[27]. In addition, SlNCED1 also can be regulated by the brassinosteroids (BRs) biosynthesis gene DWF and the key BR signaling gene BZR1 in response to cold stress[4]. SlHY5 can regulate the expression of SlNCED6 and promote the accumulation of ABA. Silencing of SlNCED significantly weakened tomato cold resistance[28].
Our findings demonstrate a rapid induction of SlMETS1 expression under cold stress for both the wild type and the not mutant, indicating SlMETS1 expression as an early response to low temperature stress in tomato. This was further supported by the increased sensitivity of SlMETS1-silenced plants to cold treatment after 8 h. The SlNCED knockout preceded the expression peak of SlMETS1 under cold stress, suggesting that SlNCED inhibition accelerated the response of SlMETS1 to cold stress. Hence, SlMETS1 may regulate cold stress response via interplaying with the ABA biosynthesis pathways. However, the precise molecular regulation mechanisms and potential interactions between SlMETS1 and SlNCED require further investigation.
Additionally, the SAM cycle, in which SlMETS1 is involved by catalyzing the synthesis of SAM biosynthetic precursor Met, is also regulated by ABA[14,15]. SAMS, a key enzyme that catalyzes the formation of SAM, can be regulated by various phytohormones, including ethylene (ETH), gibberellin (GA3), α-naphthylacetic acid (NAA), 6-benzylaminopurine (BAP), indole-3-butyric acid (IBA) and ABA[17,29−30]. It is plausible that ABA modulates METS1 expression and SAM cycle to help plants better cope with stresses.
The relative expression changes of SlMETS1 during cold stress in wild type and sit mutants were comparable. In addition, two ABA-sensitive proteins, ABO5 and ABO6, showed similar expression trends in the wild type and sit mutants under cold acclimation and 4 °C cold stress. However, the expression of SlABO5 and SlABO6 in not mutants differed from that in WT and sit mutants. These findings suggest that the response of SlABO5 and SlABO6 to cold stress is influenced by SlNCED, confirming the role of SlNCED as the rate-limiting enzyme of ABA biosynthesis. Notably, the increased expression of SlABO5 and SlABO6 in not mutants after 24 hours may be associated with mitochondrial metabolism, as the knockout of ABO6 in Arabidopsis affects plant response to ABA via accumulating higher levels of ROS in mitochondria[12].
In addition, grafting is widely used to improve the cold tolerance of horticultural crops (Solanaceae, Cucurbitaceae etc.) in production. We previously identified 18 orthologous genes that were consistently present across multiple databases containing predicted or identified long-distance phloem-delivered mRNAs in A. thaliana, grape, cucumber heterografted with watermelon, and N. benthamiana heterograft systems[31−34]. These 18 mRNAs, including METS1, were mainly enriched in biosynthesis of amino acids, nitrogen metabolism, and methionine metabolism pathways[20]. By conducting a blast search against the tomato genome, we identified the expression of 18 mobile mRNAs in both shoot and root of tomato during cold acclimation and cold stress treatments (Supplemental Fig. S4). Notably, SlMETS1 exhibited a transient up-regulation in the relative expression level in shoot after 3 h of cold treatment, indicating SlMETS1 mRNA positively involved in early cold response in tomatoes. Thus, we proposed that, as a potential systemic mRNA signal, SlMETS1 may be involved in the regulation of cold tolerance in shoot tissues.
-
The accession numbers of all genes used in this paper were obtained from the Sol Genomics Network (Solanum_lycopersicum SL4.0
https://solgenomics.net/ ). They were listed in Supplemental Table S1. -
The authors confirm contribution to the paper as follows: study conception and design: Gao L, Zhang W; performed the experiments: Liu Z, Chen Y; draft manuscript preparation: Liu Z, Zhang W; data collection: Liu Z, Chen Y and Wang T. All authors reviewed the final version of this manuscript and granted approval for its publication.
-
All data generated during this study are included within the article or its supplementary files.
-
We declare that these experiments comply with the ethical standards and legislation in China, and all commercial cultivars were procured in accordance with national and international guidelines.
We thank Ikann-editorial team for their language editing assistance. This work was co-funded by The 2115 Talent Development Program of China Agricultural University to W. Zhang and the Earmarked Fund for China Agriculture Research System (CAS-23) to Lihong Gao.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Zixi Liu, Ying Chen
- Supplemental Fig. S1 Genetic background identification by RT-PCR and RT-qPCR.
- Supplemental Fig. S2 RT-qPCR reveals expression changes of SlMETS1, CBF1, COR47 in WT, not, and sit under normal condition.
- Supplemental Fig. S3 SlMETS1 gene silencing efficiency identification and CBF-COR gene expression analysis in SlMETS1 silenced plants.
- Supplemental Fig. S4 Relative expression levels of 18 mRNAs in tomatoes subjected to cold and cold acclimation.
- Supplemental Table S1 Oligonucleotides used in the study.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Z, Chen Y, Xiang Q, Wang T, Gao L, et al. 2023. Tomato SlMETS1 positively regulates cold stress tolerance by involving ABA biosynthesis. Vegetable Research 3:28 doi: 10.48130/VR-2023-0028 

Tomato SlMETS1 positively regulates cold stress tolerance by involving ABA biosynthesis
- Received: 22 August 2023
- Accepted: 21 September 2023
- Published online: 24 October 2023
Abstract: Low temperature stress has severe detrimental effects on plant growth and crop yield. While the role of phytohormone ABA in regulating cold stress tolerance has been recognized, its underlying regulation mechanisms remain poorly understood. Here, we investigate the involvements of SlMETS1 (Methionine Synthase 1) and SlABO5/6 (ABA Overly-Sensitive 5/6) in response to low temperature stress, and their reliance on the abscisic acid (ABA) pathways using wild-type tomato (WT) and ABA defective mutants not (notabilis) and sit (sitiens). Our findings reveal that cold stress influences the expression of SlMETS1, SlABO5, and SlABO6. The sit mutant exhibits similar expression tendency to the WT. However, the not mutant shows distinctive expression patterns of these three genes. In fact, they respond more rapidly and earlier to cold stress in not compared to the WT and sit. SlMETS1 inhibition by virus-induced gene silencing (VIGS) leads to cold stress sensitivity, resembling ABA defective mutants. These discoveries suggest an association of SlMETS1, SlABO5, and SlABO6 with SlNCED in the ABA biosynthesis and signaling pathways during cold stress, providing new molecular evidence for the relevance between methionine synthesis, ABA signaling, cold tolerance and shoot-root communication in plants.
-
Key words:
- ABA /
- SlMETS1 /
- Cold tolerance /
- Shoot-root communication