









-
Cells are the basic unit of plant organization and function[1]. The cell cycle regulates plant cell division, differentiation and expansion, which ultimately affects plant growth, development and reproduction[2]. The cell cycle is a process that a viable cell undergoes from the end of the last division to the end of the next division, which includes the mitotic cycle and the endoreduplication cycle[3]. Cell cycle progression is one of the basic characteristics of biological activities, which is closely related to the development of many higher plants. Thus, the study on the regulation of cell cycle progression has become one of the hotspots of molecular biology in recent years.
In eukaryotes, the cell cycle is a complex regulatory process affected by multiple factors, in which cyclin-dependent kinases (CDKs) play key roles in regulating cell cycle. CDK binds to cyclin partners to activate CDK kinase activity by forming a Cyclin-CDK complex, which triggers G1/S phase to G2/M phase transition and controls cell cycle progression. Studies have shown that the activity of CDKs can be regulated by some proteins, among which Kip-related protein (KRP) as CDK inhibitors can bind to the CDKs and affect their activities[4]. KRP proteins can affect cell cycle process and regulate plant growth and development[5]. The C-terminus of the KRP proteins usually contain a conserved functional region composed of about 30 amino acids, which is necessary for ICK/KRPs to bind to the CDK-CYC complex[6]. However, the diversity of the N-terminal sequence of the KRP proteins leads to the low similarity of KRP protein sequences in plants. The cyclin-dependent kinase inhibitor KRPs can finely regulate the activity of CDK and negatively modulate the cell cycle process.
Plant growth and development is determined by the coordination of cell division and cell expansion, which depends on the precise regulation of cell mitosis cycle and the nuclear replication cycle. Overexpressing the KRP gene in plants exhibited a number of similar phenotypes, including smaller plant biomass, serrated leaves, and reduced cell numbers[7]. In Arabidopsis, seven KRP genes were identified. AtKRP4 and AtKRP5 were mainly expressed in dividing cells, while AtKRP1 and AtKRP2 were highly expressed in differentiated cells, and AtKRP3, AtKRP6, and AtKRP7 were expressed in both cells[5]. Interestingly, when root-knot nematodes infected Arabidopsis roots, AtKRP1 and AtKRP2 genes can inhibit the mitosis of giant cells by increasing their expression level, which prevented the proliferation of adjacent cells to hinder the root-knot nematode development[8]. The AtKRP1 gene driven by the GL2 promoter in Arabidopsis not only resulted in a significant decrease in the DNA content of epidermal hair cells, but also caused programmed death of epidermal hair cells[9]. Overexpression of AtKRP3 in Arabidopsis can increase the DNA ploidy level of shoot apical meristem and leaf cells, which changed cell arrangement characteristics and reduced cell volume[10]. In tomato, overexpression of cyclin-dependent kinase inhibitors (SlKRP) in mesocarp cells can relieve the link between intracellular replication and cell growth[11]. These reports suggest that the biological function of KRPs is to inhibit cell division.
The precise regulation of the cell cycle is also critical for plants to respond to environmental stress and changes[12]. For instance, protein kinase WEE1 affects cell numbers by inhibiting CDK1 to stop cells undergoing mitosis[13]. WEE1 protein is an important indicator of DNA replication and a damage checkpoint. When plant DNA is damaged, WEE1 is induced to express and inhibit the cell cycle to cope with abiotic stresses[14]. Also, the drought stress inhibits the enzymatic activity of CDK in Arabidopsis roots, causing the root tip meristem to stop dividing to adapt to the external environment[15]. Remarkably, few studies have been reported on how KRP responds to environmental stresses.
Tomato is an important economic horticultural plant that is popular with consumers worldwide. However, during the growth and development of tomato, its yield and quality are severely impacted by a variety of abiotic stresses, such as drought, salinity, and chilling[16]. The study of plant resistance-related genes has important guiding significance for the production of high-quality tomatoes in abnormal environmental conditions. Although some advances in the understanding of the action of KRP have been reported in Arabidopsis thaliana, genome-wide information of SlKRP family members in tomato has not been executed. Therefore, exploring the biological function and molecular mechanism of the cell cycle important regulatory gene SlKRP in abiotic stress provides a theoretical basis for cultivating tomato varieties with strong stress resistance. In recent reports, plant KRP proteins bind to CDK protein to inhibit their activity, thereby inhibiting cell division and affecting plant growth and development[17]. Although the SlKRP genes have been identified in tomato, characteristics of the tomato SlKRP family genes have not been systemically studied. Here, we identified six SlKRP genes by genome-wide analysis, and performed bioinformatics analyses to analyze phylogenetic construction, chromosome distribution, gene structure, protein interaction network, co-expression analysis, and gene duplication. Then, the expression profiles in response to abiotic stress and in various tissues of tomato plant were characterized via qRT-PCR. Furthermore, silencing of SlKRP5 increased the sensitivity to drought stress. Therefore, this work provides a theoretical basis for further functional studies of the SlKRP family and provides potential targets for future tomato improvement.
-
To identify the SlKRP genes in the tomato genome, the KRP protein sequences in Arabidopsis were obtained from the TAIR database (
https://www.Arabidopsis.org/ ). All Arabidopsis AtKRP genes were reported previously[5]. The conserved sequence of the CDI domain (pfam02234) was used to identify the SlKRP family genes. The genome database of tomato (SGN,https://solgenomics.net/ ) and the National Center for Biotechnology Information (NCBI,https://www.ncbi.nlm.nih.gov/ ) was used to search for the SlKRP genes based on the CDI domain through BLAST. Further, we used HMMER software to search the SlKRP genes of tomato. A total of six SlKRP genes were identified in tomato, namely the Solyc02g090680, Solyc09g061280, Solyc12g098310, Solyc09g091780, Solyc03g044480, and Solyc01g108610. According to the existing reports, Solyc02g090680, Solyc09g061280, Solyc12g098310, and Solyc09g091780 were named as SlKRP1, SlKRP2, SlKRP3 and SlKRP4, respectively[11]. As Solyc01g108610 has high homology with AtKRP6, Solyc01g108610 was named as SlKRP6. Solyc03g044480 was named as SlKRP5.ExPASy (https://web.expasy.org/protparam ) was used to calculate their physio-chemical characteristics (molecular weight, and isoelectric point). The secondary structure of SlKRP proteins was predicted using Novopro (www.novopro.cn/tools ). We predicted the phosphorylation potential for SlKRPs via the NetPhos-3.1 website (https://services.healthtech.dtu.dk/service.php?NetPhos-3.1 ).Analysis of chromosomal location, gene structure and gene duplication
-
According to the chromosome location information of SlKRP genes provided by the SGN website, MG2C (
http://mg2c.iask.in/mg2c_v2.1/ ) was performed to draw the schematic representation of SlKRP genes chromosomal positioning. The gene structures of SlKRPs were illustrated using the Gene Structure Display Server (GSDS,http://gsds.cbi.pku.edu.cn/ ). The Multiple Collinearity Scan toolkit (MCScanX) with default parameters was used to examine the gene duplication events of SlKRP genes[18].Phylogenic analyses
-
Sequence alignment of KRP proteins from tomato, tobacco, Arabidopsis, capsicum and eggplant was carried out with the ClustalW program and the integrated tool MEGAX (
www.megasoftware.net ). Then, a phylogenic tree was built with the neighbor-joining (NJ) algorithm, wherein the bootstrap replicate value was set as 1000.Identification of cis-elements of SlKRP genes
-
The 3-Kb promoters of six SlKRP genes were extracted from the SGN database and the cis-regulatory elements were analyzed using the Plant CARE website (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ). The identified CAREs visualized using the Toolkit for Biologists integrating various biological data handling tools (TBtools)[19].Plant materials and methods of stress treatment
-
Tomato seedings were cultured in a cave containing vermiculite: peat: perlite (1:3:1 v/v/v). The seedlings were grown in a glass greenhouse, 14 h at 28 °C/10 h at 18 °C (day/night) photocycle, and the relative humidity was 75%. Uniform seedlings at the three-leaf stage were selected and transferred to plastic pots (10 cm × 10 cm × 10 cm, one plant per pot) for the stress treatment. Tomato seedlings were treated with 150 mmol·L−1 NaCl for stress treatment. Tomato seedlings were transferred to dry substrate for drought stress treatment. Tomato seedlings were placed in 4 °C incubators to simulate low temperature stress. Both treatment group and control group were treated for 12 h. Leaves were also immediately frozen in liquid nitrogen and stored at −80 °C. The six individual tomato seedlings were treated for each experiment.
Expression analysis for SlKRP genes
-
For analysis of SlKRP genes expression patterns under different conditions, including various abiotic stresses, growth and development, the total RNA of plant samples was isolated with an OminiPlant RNAkit (Cwbio, Beijing, China). Then, complementary DNAs were synthesized using an HiScript III RT SuperMix reverse transcriptase kit (Vazyme, Cat. #R323-01) according to the manufacturer's instruction. qRT-PCR was used to determine the gene transcription using 96-well blocks with the ABI QuantStudio 3 (Applied Biosystems, USA). The following qRT-PCR program was used: the template denaturation at 95 °C for 3 min; followed by amplification for 40 cycles with a melting temperature of 95 °C for 10 s and an annealing temperature of 68 °C for 15 s. The comparative 2−ΔΔCᴛ method was used to calculate the relative expression levels of target genes, and the β-actin gene (Soly11g008430) was used as an internal control. The primers for RT-PCR are listed in Supplemental Table S1.
We analyzed the expression patterns of SlKRP genes under shade, sun, ABA, IAA and ACC treatments using the published RNA-seq datasets[20]. TBtools was used to draw heatmaps.
Protein subcellular localization
-
To investigate the subcellular localizations of the SlKRP proteins, the coding sequence of SlKRP without the stop codon was amplified by PCR and then cloned into the expression vector pGWB405 with GFP under the control of the CaMV35S promoter by homologous recombination. The fusion constructs were transformed into tobacco leaves as described previously[21]. The nuclei were detected by DAPI staining. GFP fluorescence were detected at 48 h following transfection using laser confocal microscope (LSM; Carl Zeiss, Thornwood, NY, USA). The primers for subcellular localization are listed in Supplemental Table S1.
Virus-induced gene silencing (VIGS)
-
A particular fragment of SlKRP5 was designed and amplified by PCR using specific primers (Supplemental Table S1). The fragment from SlKRP5 was inserted into the tobacco rattle virus RNA2 (TRV2) vector to construct recombinant vector TRV2:SlKRP5, and were introduced into Agrobacterium tumefaciens GV3101. A. tumefaciens cells containing TRV1 were mixed with TRV2:SlKRP5, pTRV2:00 (negative control), or pTRV2-SlPDS (positive control) vectors at a volume ratio of 1 : 1. The plants at two-leaf stage were infiltrated with inoculant of Agrobacterium suspensions (OD600 = 0.5). When leaves of pTRV2:SlPDS plants emerged photobleached phenotype, we performed qRT-PCR assay to determine the SlKRP5 expression in TRV2:SlKRP5 plants and calculate silencing efficiency. VIGS assays were conducted as previously described[22].
Drought stress treatment
-
SlKRP5-silenced (TRV2:SlKRP5) and control (TRV2:00) plants were grown in soil and treated with drought stress. For imitation of drought stress, roots of plants were watered with 150 ml of 25% (w/v) PEG6000 solution every day and the whole treatment lasted for 3 d in total. Normally growing plants were used as controls. Drought stress treatment were conducted as previously described[23]. And then, the physiological indicators were measured. Each treatment contained six plants.
Measurement of antioxidant enzyme activity
-
The content and activities of malondialdehyde (MAD), superoxide dismutase (SOD), and peroxidase (POD) were determined at different wavelengths using enzyme-labeled instrument according to a protocol described previously[22]. The production of hydrogen peroxide (H2O2) and superoxide anion (
${\text O_2^-} $ -
In this study, based on the amino acid sequence of the Pfam CDI domain, six SlKRP gene members were obtained via SGN, NCBI, and other public databases in the tomato genome. We further analyzed the SlKRP proteins, such as the chromosome location, the amino acid length, the molecular weight, and the theoretical isoelectric points (Table 1). We found that SlKRP genes were unevenly distributed on chromosomes 1, 2, 3, 9 and 12 (Fig. 1a). The amino acids (aa) length of SlKRP proteins ranged from 188 aa (SlKRP2) to 232 aa (SlKRP3). In addition, the molecular weights and theoretical isoelectric points of these proteins ranged from 2,1489.04 Da (SlKRP2) to 2,6152.29 Da (SlKRP3) and from 4.14 (SlKRP5) to 9.67 (SlKRP3), respectively (Table 1). Gene family is generated by either tandem repeats or large-scale fragment repeats during the evolutionary process[24]. Segmental gene duplication revealed that SlKRP1 is highly similar to SlKRP3, SlKRP5 and SlKRP6, indicating that they undergo intrachromosomal or interchromosomal fragment replication (Fig. 1b).
Table 1. Physico-chemical characteristics for the SlKRP family genes.
KRP member Gene ID Chr. no. Strand CDS length (bp) Protein length (aa) Molecular weight/Da PI SlKRP1 Solyc02g090680.2.1 2 + 639 212 24,011.99 6.06 SlKRP2 Solyc09g061280.2.1 9 − 567 188 21,489.04 9.45 SlKRP3 Solyc12g098310.1.1 12 + 699 232 26,152.29 9.67 SlKRP4 Solyc09g091780.2.1 9 − 633 210 23,844.81 9.22 SlKRP5 Solyc03g044480.2.1 3 − 612 203 24,207.86 4.14 SlKRP6 Solyc01g108610.2.1 1 + 657 218 24,102.7 5.46 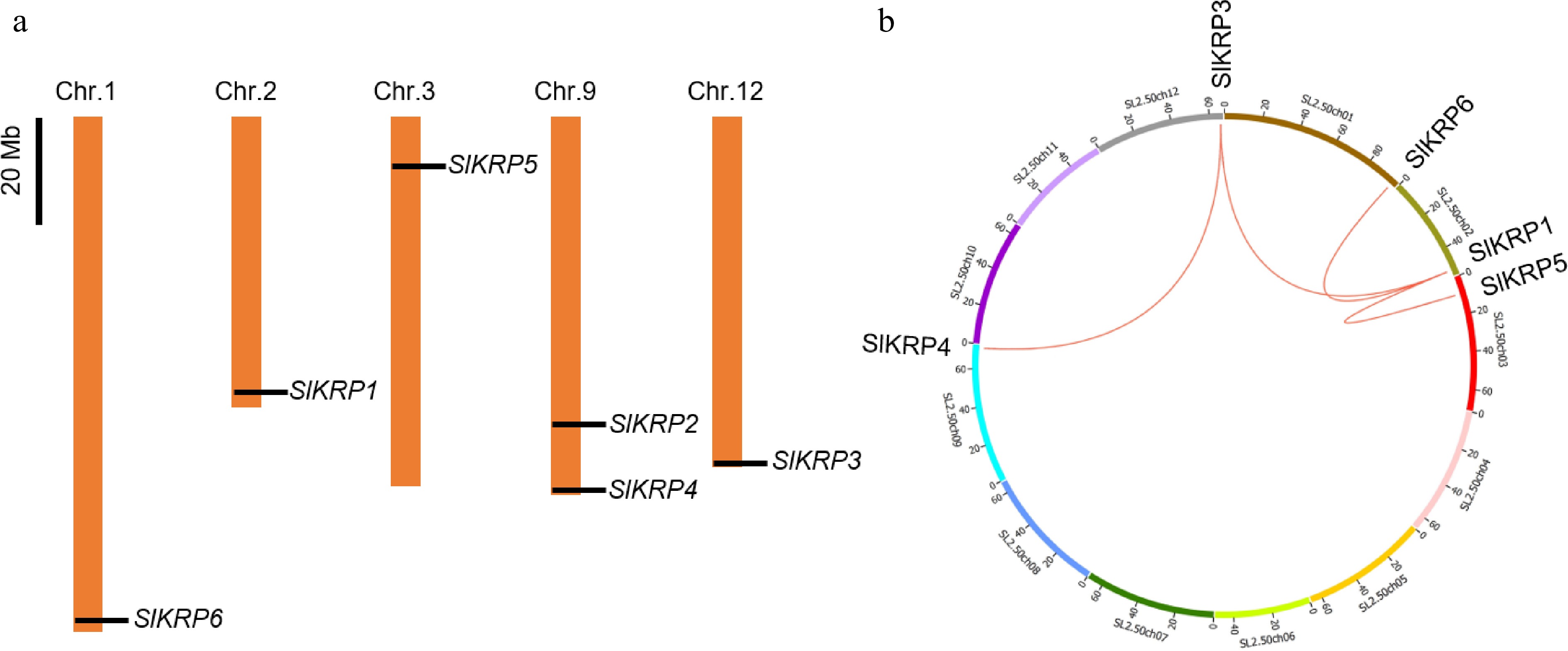
Figure 1.
(a) Distribution and (b) duplication of SlKRP genes on tomato chromosomes. Chromosome numbers are indicated at the top of each bar.
Structures and motifs analysis of SlKRP genes in tomato
-
We further analyzed the exon–intron structures and motif compositions of the SlKRP genes in the tomato genome (Fig. 2). The lengths of SlKRP genes were from 1,437 bp to 3,001 bp. The SlKRP1, SlKRP5 and SlKRP6 genes contained four exons and three introns, while The SlKRP2, SlKRP3 and SlKRP4 genes contained three exons and two introns. Also, conserved domain of SlKRP proteins was cyclin-dependent kinase inhibitor (CDI) and existed in C-terminal region of SlKRP proteins (Supplemental Fig. S1). We speculated that SlKRP genes had evolutionary diversity according to gene structure and gene length. Further, we identified the 20 different motifs (Motifs 1–20) from the SlKRP genes in the tomato genome using the MEME program (Table 2). The amino acid lengths of these conserved motifs ranged from 6 to 44 aa, of which motif 1 was CDI domain. The motif 1 was identified in six SlKRP proteins. We found that the SlKRP genes that classified in the same category had similar conserved domains.
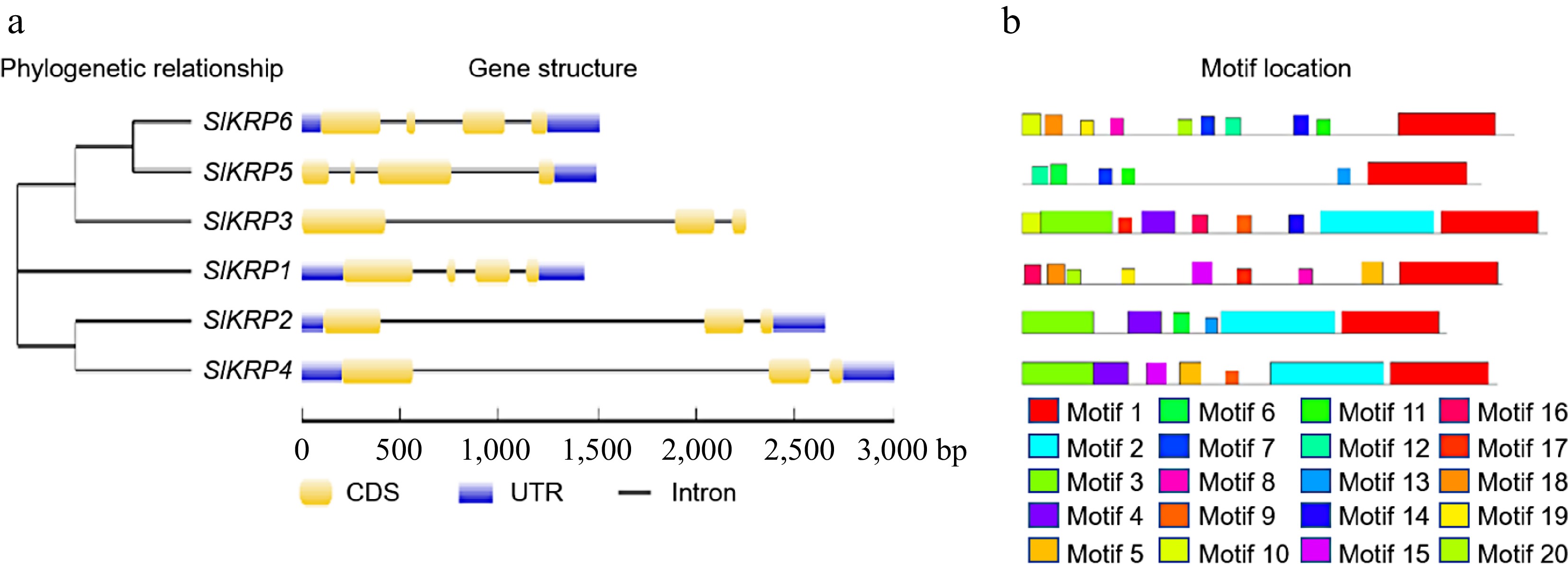
Figure 2.
Gene structures and conserved domains of SlKRP genes. (a) Exon-intron structure of the SlKRP genes. (b) Distributions of conserved motifs in SlKRP proteins. The 1−20 motifs in SlKRP proteins were identified by the MEME program (
http://meme-suite.org/ ), which were displayed by different colored boxes. Motif 1 is cyclin-dependent kinase inhibitor domain.Table 2. Details of conserved motifs in tomato KRP proteins.
Motif Length (aa) Best possible match 1 43 IPTEAELEEFFTAAEKRQQKRFIEKYNFDFVKDEPLEGRYEWV 2 44 NLLEFEGRKRTTRESTPCSLIRDPDNIPTPGSSTRRTNANEANGRVPNSI 3 32 MGKYJRKTGKVLDVSPLGVRTRAKTLALKRLQ 4 15 GGCYLQLRSRRLEKP 5 9 PKPQIPKVC 6 7 CDNYHPV 7 6 CCSSCY 8 6 CAMSYS 9 6 MEGQKW 10 8 MGEFLKKC 11 6 PDEKCG 12 7 MMKKKRK 13 6 DEILFP 14 7 NFKPIDN 15 9 QGNGVPCEP 16 7 RRKHKCK 17 6 SGGGDG 18 8 INGEMKIM 19 6 KRDGDL 20 6 VAEVAI Phylogenetic analysis of KRPs
-
To study evolutionary patterns of KRP genes in the plants, we obtained KRP homologous of tobacco, Arabidopsis, capsicum and eggplant by sequence alignment in NCBI databases (Supplemental Table S2). Then, an unrooted phylogenetic tree with KRP protein sequences from five species was performed via MEGA software (Fig. 3). We found that the six SlKRP proteins can be classified into three classes, including Class I (SlKRP1 and SlKRP5), Class II (SlKRP2, SlKRP3 and SlKRP4), and Class III (SlKRP6). The phylogenetic analysis indicated that a closer orthologous relationship of SlKRP proteins in each clade was observed between tomato and eggplant, perhaps indicating the closest relationship between the tomato and eggplant.
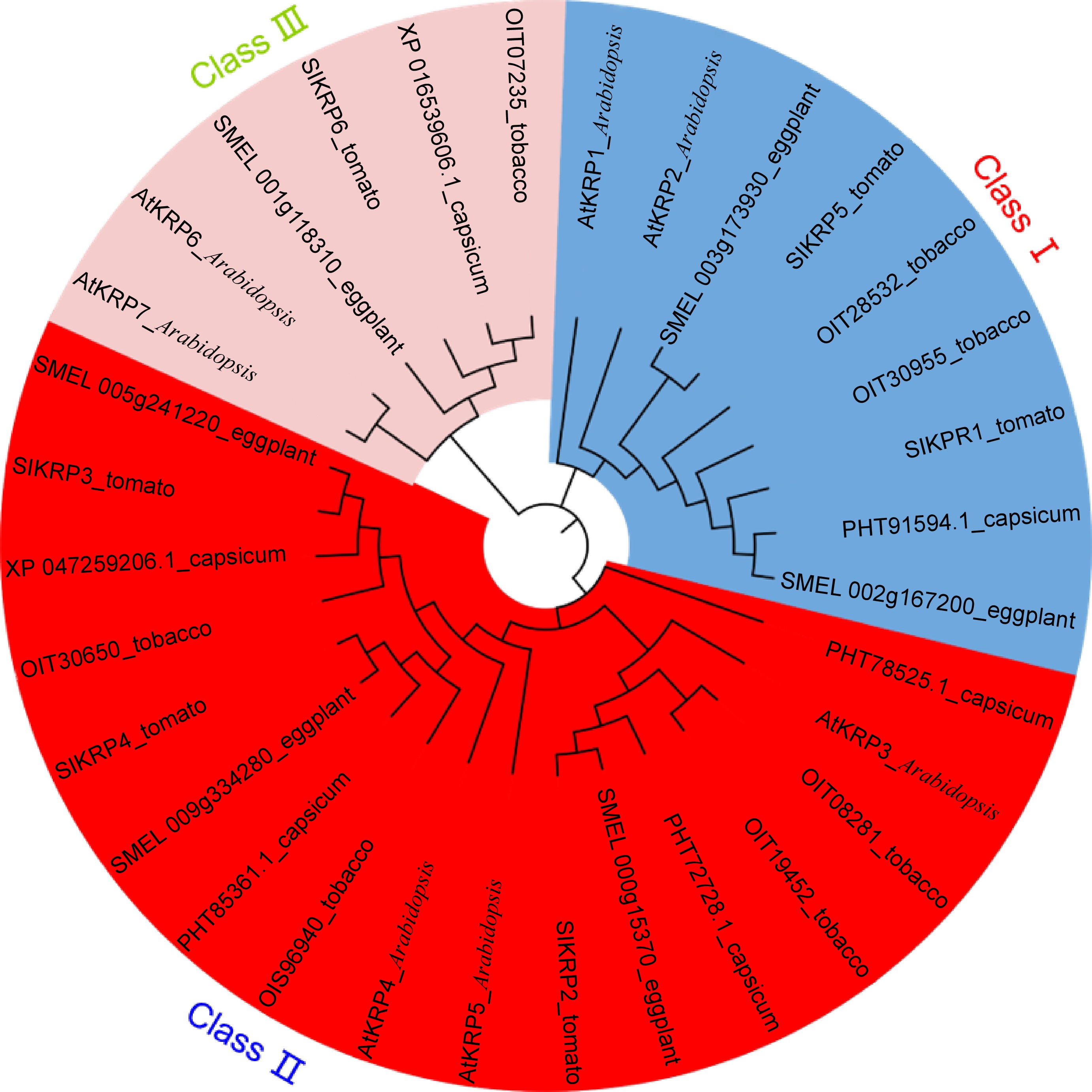
Figure 3.
Phylogenetic trees showing KRP genes from tomato, tobacco, Arabidopsis, capsicum and eggplant. The KRP proteins were classified into three subfamilies and distinguished by different colors.
The expression pattern of SlKRP genes
-
To further understand the biological functions of SlKRP genes, we performed qRT-PCR to analyze the expression of six SlKRP genes in different tomato tissues, including root, stem, leaf, flower, and fruits of different developmental stages. qRT-PCR analysis showed that the six SlKRP genes had obvious tissue-specific expression (Fig. 4a). Among them, the expression of SlKRP5 and SlKRP6 was relatively higher in leaves. SlKRP1, SlKRP3, and SlKRP4 exhibited relatively higher expression in anthesis, indicating that the three genes may be involved in cell division of tomato fruit. The expression abundance of SlKRP2 gene gradually increased with fruit development, indicating that it may be involved in fruit ripening and quality formation. Moreover, we also performed transcriptomic datasets published by SGN website to analyze the differential expression of the SlKRP genes in fruits of the wild variety 'LA1589' and the cultivar 'Heinz 1706', suggesting that SlKRPs may be involved in the formation of tomato fruit size (Supplemental Figs S2 & S3). Taken together, these results suggested that the six SlKRP genes had functional divergence in regulating tomato growth and development.
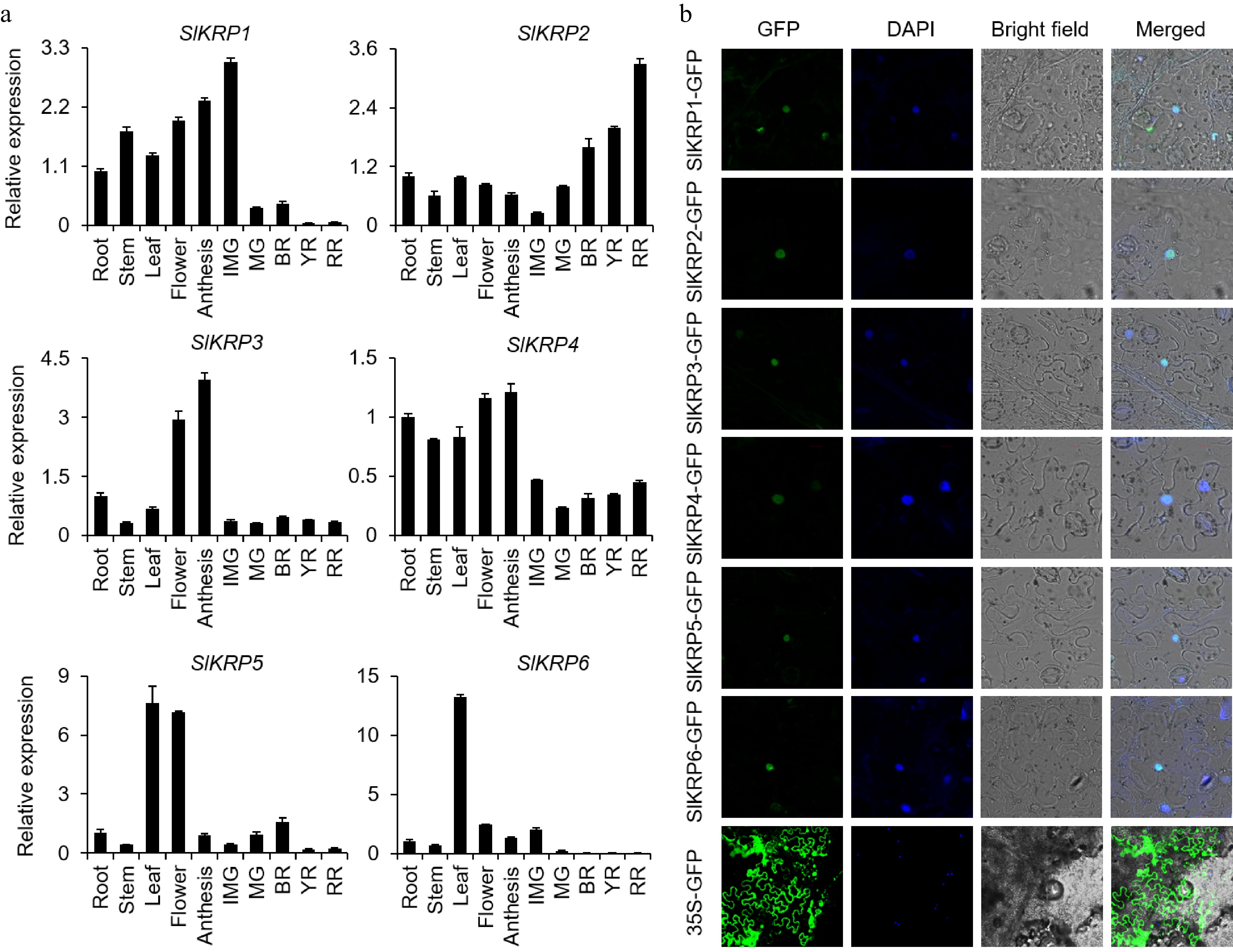
Figure 4.
Expression characterization of SlKRP genes. (a) Expression profiles of SlKRP genes in different tissues, including root, stem, leaf, flower, anthesis, immature green fruit (IMG), mature green fruit (MG), breaker fruit (BR), yellow fruit (YR), and red ripe fruit (RR). The expression levels of SlKRP genes were calculated using the CT method. Data shown are means ± SD (n = 3). The β-actin gene was used as an internal control. (b) Subcellular localization of SlKRP-GFP (green fluorescent protein) fusion proteins in tobacco leaf epidermal cells. The nuclei were determined by DAPI staining.
To investigate the subcellular localization of SlKRP proteins, we constructed SlKRP-GFP fusion proteins driven by the CaMV35S promoter, which were transiently expressed in 5-week-old tobacco leaves. Confocal microscope observation showed the SlKRP-GFP fluorescence signal overlapped with that of DAPI, a nuclear localization marker[25]. Therefore, these KRP proteins were localized in the nucleus (Fig. 4b).
Cis-regulatory elements in the promoters of SlKRP genes
-
Gene transcription is regulated by cis-regulatory elements (CAREs) in the promoter sequence[26]. To further explore the function of SlKRP genes, 131 CAREs were identified in the 3-Kb promoters of SlKRP genes, which were categorized into ten responsive groups, containing ABA-, MeJA-, light-, low temperature-, GA-, defense and stress-, SA-, Auxin-, drought-responsive element,and cell cycle regulation. It is worth noting that there was CARE involved in cell cycle regulation in the SlKRP5 promoter, indicating that its potential function may be cell cycle regulation (Fig. 5). These CAREs indicated that SlKRPs may play an important role in response to abiotic and biotic stress.
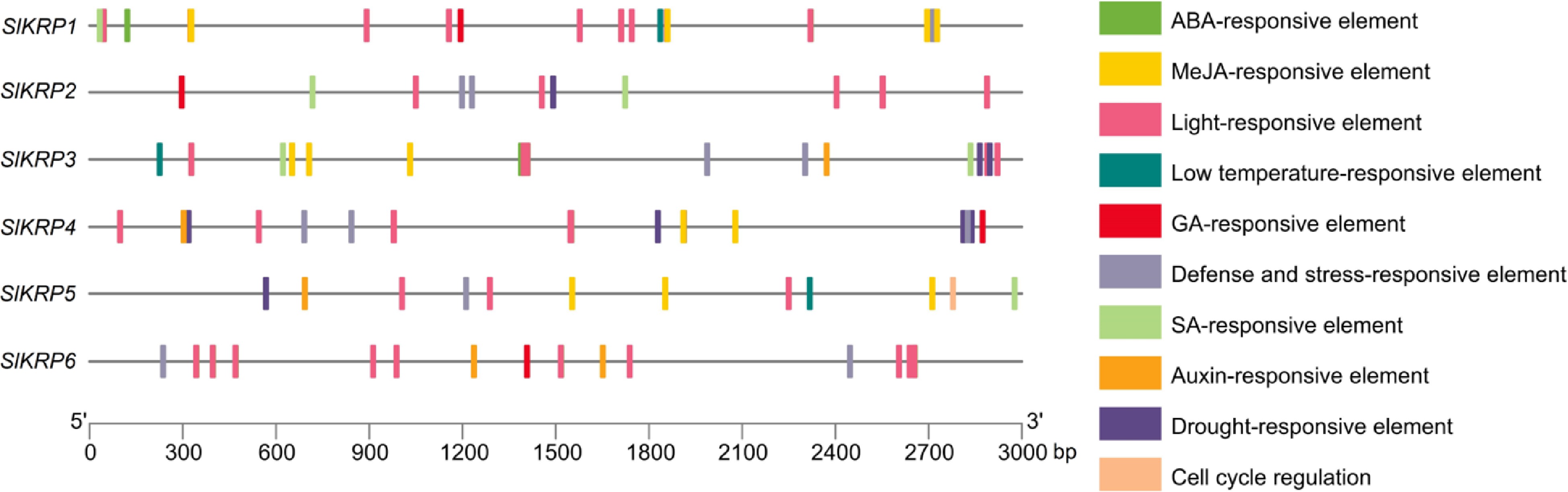
Figure 5.
Cis-regulatory elements in the promoter regions of SlKRP genes.
Expression patterns of SlKRP genes in response to abiotic stress
-
To analyze the SlKRP genes response to three abiotic challenges, such as salt, drought, and low temperature, we analyzed the expression profiles of the SlKRP genes under stress conditions. qRT-PCR analysis showed that expression abundance of SlKRP1, SlKRP2, SlKRP3, SlKRP4, and SlKRP5 genes were significantly suppressed after salt treatment (Fig. 6a). Under drought treatment, the expression level of SlKRP3 was increased by 2.6 times compared with that in tomato seedlings under normal conditions, and the expression levels of SlKRP5 and SlKRP6 decreased to 30.8% and 21.8% of the control, respectively (Fig. 6b). The expression of five SlKRPs was significantly different in response to low temperature, of which 4 SlKRP genes were significantly downregulated compared with normal temperature (Fig. 6c). Low temperature significantly increased expression level of SlKRP2 by 8.6-fold compared with the control. We further explored the expressional responses of SlKRPs to light treatment using the published transcriptomic datasets, and found that shade treatment can up-regulate the expression of SlKRP1, SlKRP3, and SlKRP6. Whereas, the expression of SlKRP2, SlKRP4, and SlKRP5 genes showed an opposite pattern to that under sunlight treatment (Fig. 6d).
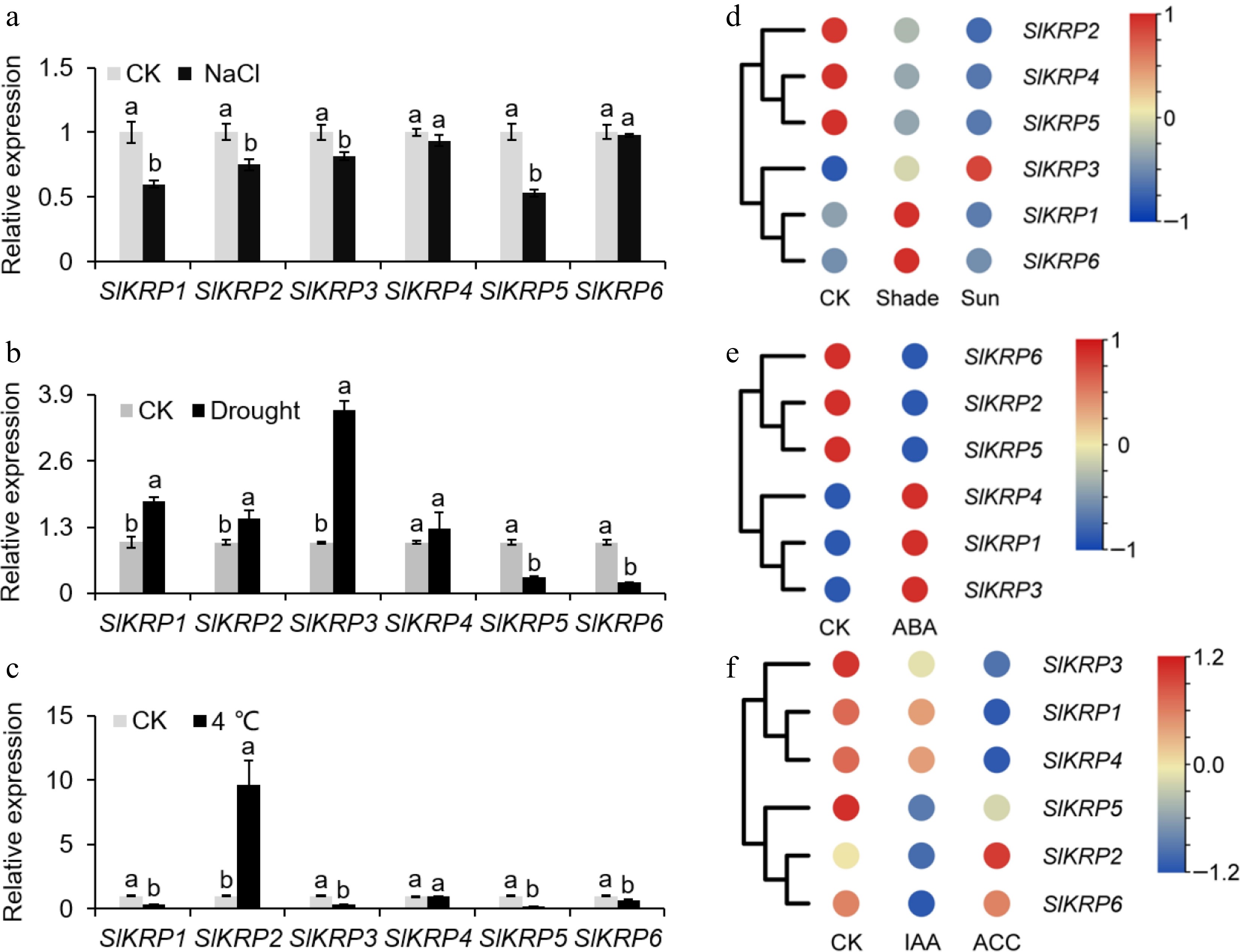
Figure 6.
Expression profiles of SlKRP genes under various environmental stressors and phytohormone treatment. Expression profiles of SlKRP genes in tomato plants after (a) salt , (b) drought, and (c) low temperature treatments. The β-actin gene was used as an internal control. The data are presented as the means ± SDs (n = 3). The different letters indicate statistically significant differences at a 5% level of significance according to Tukey's pairwise comparison tests. A heatmap displaying SlKRP genes expression under (d) light, (e) ABA, IAA and (f) ACC treatments. Red and blue represented up-regulated and down-regulated gene expression after treatment, respectively.
In addition to environmental stress, plant hormones are the key factors in regulating the whole process from seed germination to fruit formation, which reveals the regulation mechanism of crop agronomic trait formation[27]. Therefore, we analyzed the expression profiles of the SlKRP genes in response to phytohormone treatments with ABA, IAA, and ACC, showing that the six SlKRP genes differentially expressed. Under ABA treatment, SlKRP1, SlKRP3, and SlKRP4 were upregulated, and other SlKRPs were significantly down-regulated (Fig. 6e). Among the six SlKRP genes, the expression of SlKRP2, SlKRP3, SlKRP5, and SlKRP6 was down-regulated after IAA application compared with CK. Under ACC treatment, the expression of four SlKRPs, including SlKRP1, SlKRP3, SlKRP4, and SlKRP5 genes exhibited differential compared with the normal condition, and SlKRP6 were less sensitive to treatment (Fig. 6f). In short, our analysis suggested that SlKRP genes played regulatory roles in response to phytohormone and environmental stress in tomato plants.
Relationship between SlKRP family genes and cell cycle related genes
-
The ICK/KRP family genes are major regulators of cyclin-dependent kinase activities in several plants, which regulate endoreplication and cell division in plants[28]. Gene co-expression is a technical method to show the interaction between genes based on their expression data[29]. To further understand the relationship within tomato SlKRP, SlCDK and cyclin gene expression, we performed co-expression analysis using transcriptome data of these genes from tomato fruits at nine developmental stages[30]. The expression abundance of SlKRP1 was positively correlated with the most SlCDK and cyclin genes transcription, while the expression level of SlKRP1 was highly negatively correlated with the that of SlCDKD1. The expression levels of SlKRP3, SlKRP5, and SlKRP6 showed positive correlation with that of some SlCDK and cyclin genes. The expression level of SlKRP2 was highly negatively correlated with the expression of most CDK and cyclin genes. In short, these results reveal that these genes may interact with each other to regulate the cell cycle (Fig. 7).
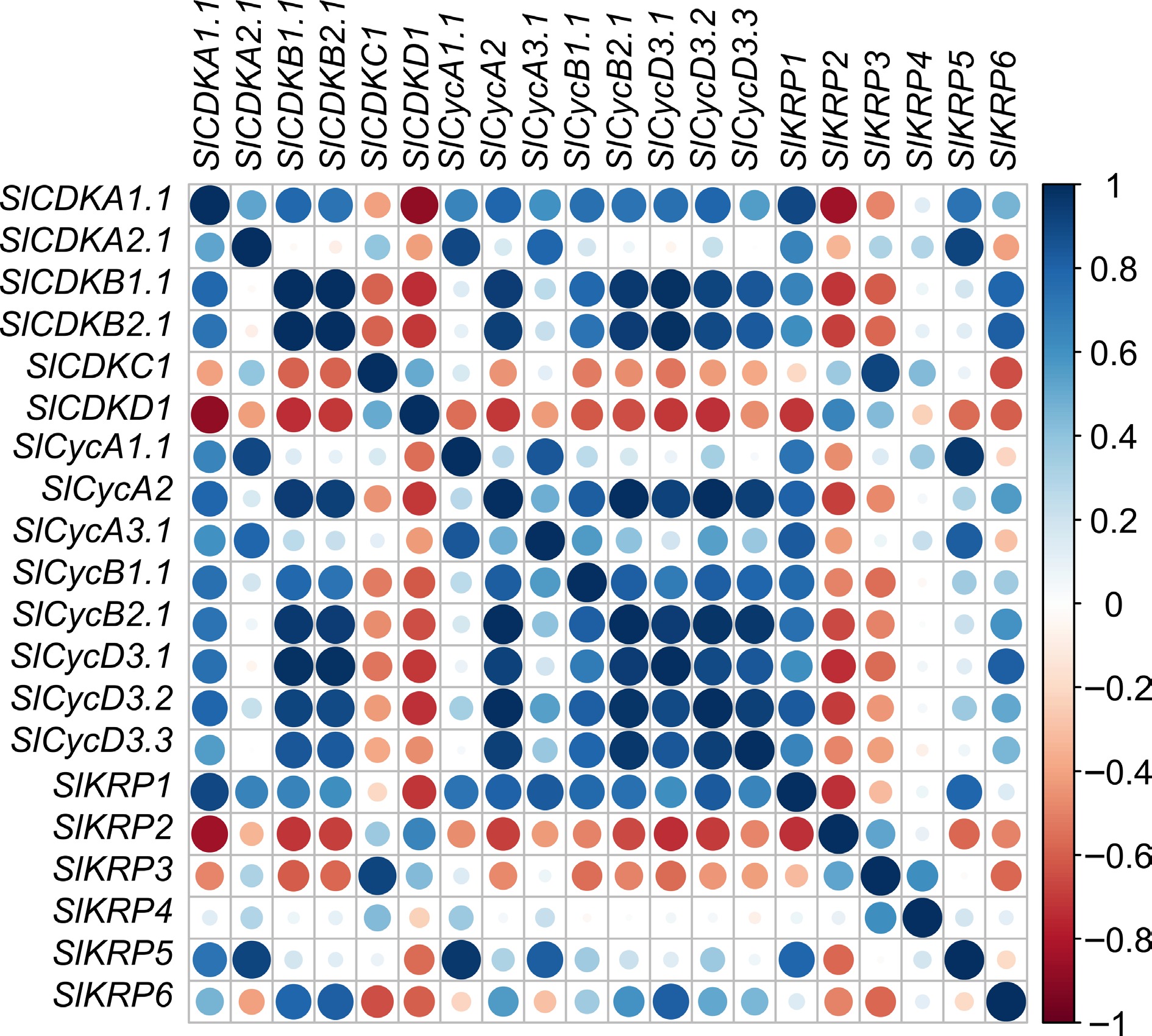
Figure 7.
Co-expression analysis of SlKRP, SlCDK and cyclin genes in tomato. The color of the circles indicates the correlation coefficient value of gene co-expression in the R environment.
Furthermore, to gain insight into the possible biological functions of SlKRPs, we searched proteins that may interact with SlKRPs via the STRING database (Fig. 8). Interestingly, many proteins involved in cell cycle regulation were predicted to be associated with SlKRP proteins, showing that SlKRPs played essential roles in regulation of the cell cycle. Among the proteins, cyclins interacted with CDK proteins to control cell cycle progression[31]. WEE1 protein kinase is a member of the serine/threonine protein kinase family, which mainly inhibits the activity of CDC protein to regulate cell mitosis[32]. However, we did not find proteins associated with SlKRP5. Therefore, these results indicate that SlKRP proteins interacted with cell cycle-related proteins to regulate cell cycle progression.
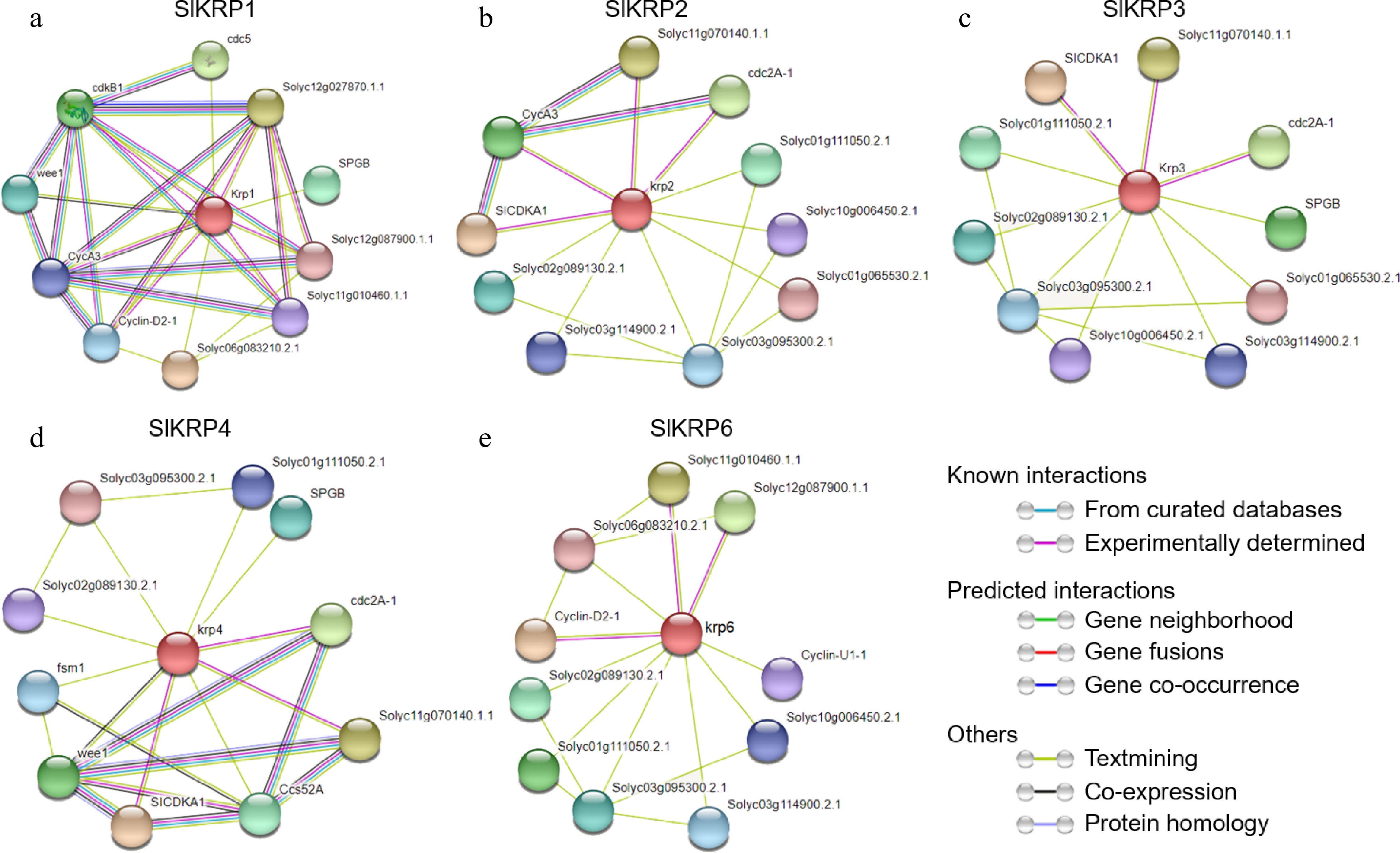
Figure 8.
The predicted protein interaction network of the SlKRP proteins in tomato using string database.
Inhibited expression of SlKRP5 reduces drought tolerance in tomato
-
Drought treatment significantly affected SlKRP5 expression in tomato, suggesting that SlKRP5 potentially is involved in drought stress responses. To further elucidate the importance of SlKRP5 in basal drought tolerance, we performed virus-induced gene silencing (VIGS) assay to suppress SlKRP5 expression in tomato. SlPDS-silenced plants revealed a photo-bleached phenotype after 15 d of infiltration (Fig. 9a). qRT-PCR results showed that was SlKRP5 expression in the TRV2:SlKRP5 plants was significantly lower than that in control (TRV2:00) plants (Fig. 9b), illustrating that SlKRP5 was successfully silenced. We use the 25% PEG600 to treat the control and TRV2:SlKRP5 plants to simulate drought stress. Most of the leaves of the TRV2:SlKRP5 plants became withered significantly after drought stress (25% w/v PEG6000) for 3 d, nevertheless those of the control plants displayed mild curling (Fig. 9c). Under drought stress, DAB and NBT staining results showed that TRV2:SlKRP5 plants had higher accumulations of O2− and H2O2 than the control plants, reflected the greater degree of membrane damage of TRV2:SlKRP5 plants (Fig. 9d & e). Furthermore, we measured the activities of the main ROS-scavenging enzymes, including SOD and POD and found that their activities were remarkably decreased in the SlKRP5-silenced plants after drought stress (Fig. 9g & h). Collectively, these results indicate that SlKRP5 is critical for tomato resistance against drought.
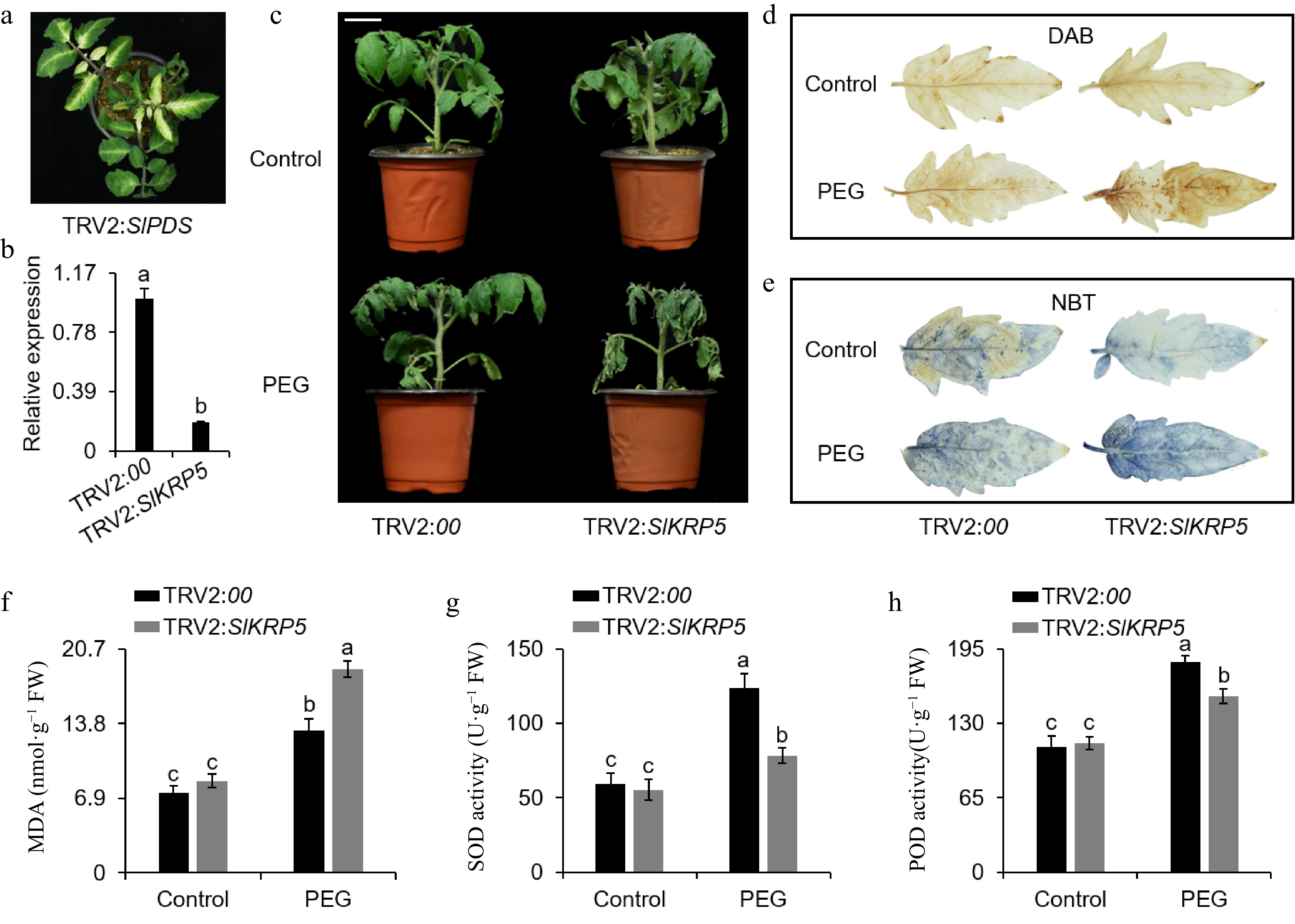
Figure 9.
Knockdown of SlKRP5 reduces drought stress tolerance in tomato. (a) Phenotype of SlPDS-silenced tomato plant via VIGS. (b) Expression level of SlKRP5 in SlKRP5-silenced and control plants. (c) Phenotype of SlKRP5-silenced and control plants treated with PEG6000 (25% w/v). Scale bar, 3 cm. DAB staining for (d) H2O2 and (e) NBT staining for superoxide in control and SlKRP5-silenced tomato leaves. (f) MDA content, (g) SOD activity and (h) POD activity in SlKRP5-silenced and control plants. The data are presented as the means ± SDs (n = 3). The different letters indicate statistically significant differences at a 5% level of significance according to Tukey's pairwise comparison tests.
-
KRP proteins are key regulators of CDK-Cyclin complex activities in endoreduplicating cells, which were identified in several plants, such as A. thaliana, O. sativa, and Z. mays[33]. However, systemic study on the roles of SlKRP genes in tomato was limited. Herein, we used the HMMER model and BLASTP to identify SlKRP family protein sequences through comparative analyses based on the Arabidopsis ICK/KRP sequences (Table 1). In maize, all of the inhibitors of cyclin-dependent kinase proteins mainly localized to the nucleus[34]. Consistent with previous reports, subcellular localization indicated that the six KRP proteins were located in the nucleus in this study. Meanwhile, these SlKRP proteins have similar secondary structures (Supplemental Fig. S4). These results provide essential information for the further functional analysis of KRPs in the plants.
The C-terminal region of KRPs genes in plants is considered as a conserved functional motif, which is involved in interaction with CDK proteins and inhibits their activity[35]. The interaction region of KRPs with D-type cyclins and A-type CDKs is located at the C-terminus of protein[15]. In vitro enzymatic activity assay demonstrates that KRP protein can inhibit CDK activity in plant[5]. However, the tomato ICK/KRP protein SlKRP1, which lacked the conserved C-terminal region, can still interact with SlCYCD3[36]. Similarly, we performed multiple sequence alignment analysis of tomato SlKRP family and found that the C-terminal region of SlKRP proteins contain a conserved domain (Supplemental Fig. S1). On the other hand, SlKRP proteins had multiple phosphorylation sites (Supplemental Fig. S5), which indicated that SlKRP proteins may be phosphorylated by other kinases such as CDK In conclusion, the SlKRP protein structures conferred functional diversity in regulating plant growth. In this study, we identified six SlKRP genes on the five chromosomes of the tomato genome and performed phylogenetic analysis to reveal that the six SlKRP genes were classified into three classes, which is consistent with reports on Arabidopsis thaliana (Fig. 3). However, a total of seven AtKRP genes were identified in Arabidopsis[5]. The interchromosomal segment duplication and tandem duplication in plants are important driving forces for the evolution of genome and genetic systems[37]. The gene duplication analysis displayed that the SlKRP gene family is mainly characterized by interchromosomal segment duplication, suggesting that interchromosomal segmental duplication is the main expansion mechanism of these SlKRP genes in tomato (Fig. 1b). Exon-intron structure of gene is essential in the study of gene family evolution[38]. More meaningfully, we also found that the structure of interchromosomal segmental duplication genes is highly similar. These results indicated that structural and expression differences in the SlKRP gene family may confer the functional diversity in regulation of plant growth and development and tolerance to abiotic stresses. Fine-tuning of KRP expression abundance was demonstrated to be a key characteristic of cell cycle control. The expression patterns of SlKRP1-SlKRP4 in vegetative organs (roots, leaves, flowers) and developing fruits of the cherry tomato cultivar WVa106 has been reported[11]. The results showed that SlKRP4 was mostly expressed in the early stage of fruit development, while SlKRP1 was generally highly expressed in tomato vegetative organs. These findings are consistent with our results on expression patterns of SlKRPs measured in the processing tomato cultivar Heinz1706. However, our data suggested that SlKRP3 was lowly expressed in the late stage of tomato fruit development, which was different from previous reports. The difference in SlKRPs expression patterns may be due to various tomato cultivars. Here, we used qRT-PCR analysis from different tissues (root, stem, leaf, flower, and fruit) to explore the expression of the SlKRP gene family, showing that SlKRPs displayed the unique expression profiles in different tissues of tomato (Fig. 4). It is worth noting that cell division and expansion occurred in early fruit development and directly influenced the size and shape of fruit[39]. SlKRP1 had high expression mostly in young fruit development, which implied that SlKRP1 may be involved in cell division in young fruit and determine the tomato fruit weight. In addition, the cyclin-dependent kinase inhibitor KRP can inhibit the mitotic CDKA;1 kinase complex to regulate the mitosis-to-endocycle transition, which impacted the leaf size in Arabidopsis[40]. Indeed, SlKRP5 and SlKRP6 showed relatively high expression in the leaves than other organs, suggesting that these two genes may synergistically regulate tomato leaf development. The expression abundance of SlKRP2 gradually increased with the development of tomato fruits, demonstrating that SlKRP2 may affect the fruit ripening. Plant growth and development are influenced by various environmental factors and phytohormones, such as light, temperature, abscisic acid, which can regulate several cell cycle gene transcriptions to affect plant growth[6,41]. For instance, ABA treatment can significantly induce the expression abundance of AtKRP1 gene in Arabidopsis[15]. In this study, we found that most SlKRP genes had diverse cis-elements on their promoters, such as ABA-responsive, Light-responsive, and Low temperature-responsive boxes (Fig. 5). Additionally, the cell cycle regulation element existed in the promoter of the SlKRP5 gene. Interesting, the transcripts of SlKRP1, SlKRP5, and SlKRP6 were significantly suppressed by almost all tested abiotic stressors, especially the expression of SlKRP5 decreased to 53.1%, 30.9% and 17.8% of that in normal conditions after salt, drought and low temperature treatments. Also, the transcripts of SlKRP family genes were not obviously affected after the treatment of exogenous ABA, IAA and ACC (Fig. 6). These data indicated that SlKRPs might play a vital role in stress resistance.
Drought stress is a major factor affecting plant growth and development, which limits agricultural production. The water loss can disrupt the ion balance, increase oxidation-reduction potential, produce reactive oxygen species, and even destroy macromolecules in plants. Drought causes the oxidation reaction in plant cell membrane lipids to produce malondialdehyde. Plants can quickly sense drought stress and then regulate expression of stress-related genes, which finally resist drought through self-regulation. Abiotic stress can cause damage to plant DNA, which in turn causes cell cycle retention[42]. The interaction between CDK and KRP maintains cell cycle process to alleviate damage of abiotic stress to plants. The expression of some KRP genes was induced by abiotic stress or ABA, indicating that KRP genes may play an important role in drought resistance[43]. Some cell cycle-related genes improve plant drought tolerance by regulating stomatal development. CAREs in promoter and abiotic stress induced expression analysis showed that SlKRP5 has a regulatory role in abiotic stress. In this study, we performed the VIGS experiment to obtain SlKRP5-silenced plants. Under drought treatment, MDA content was significantly increased and antioxidant enzyme activity was decreased in SlKRP5-silenced plants. Therefore, SlKRP5 is a novel target gene in genetic engineering to enhance drought tolerance in tomato.
-
In this study, we identified six SlKRP genes which are located on tomato five chromosomes and classified into three main classes by phylogenetic analysis. Analyses on gene structure, protein motifs, subcellular localization and protein interaction network of SlKRP genes revealed their evolutionally conserved function. Tissue-specific expression patterns of SlKRP genes indicated specific roles of each SlKRP gene in tomato fruit development, fruit ripening, and leaf size. Both cis-element analyses in promoter regions of SlKRP genes and their expressions in various treatments suggested that SlKRP genes may play important roles in the phytohormone signaling and response to abiotic stresses. Silencing of SlKRP5 via VIGS assay reduces drought tolerance in tomato. Our studies lay foundation for the analysis of SlKRP function in tomato plant development and stress responses.
This work was supported by National Natural Science Foundation of China (32202497), Taishan Scholar Foundation of Shandong Province (tsqn201812034), China Postdoctoral Science Foundation (2022M711967), and National Natural Science Foundation of China (31872951).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Genzhong Liu, Zhangfeng Guan, Mingxuan Ma
- Supplemental Fig. S1 Multiple sequence alignment of the SlKRP proteins in tomato.
- Supplemental Fig. S2 Expression of the SlKRP genes in immature green fruits from 'Heinz 1706' and 'LA1589'.
- Supplemental Fig. S3 Expression of SlKRP genes in different tissues of tomato fruits at four developmental stages.
- Supplemental Fig. S4 Prediction of SlKRP protein’s secondary structure via Novopro website. Spiral and arrow represented α-helix and β-pleated sheet, respectively.
- Supplemental Fig. S5 Phosphorylation site prediction of tomato SlKRP proteins via the NetPhos-3.1 website.
- Supplemental Table S1 Primers used in this study.
- Supplemental Table S2 The identified amino acid sequences of KRP in tomato, tobacco, Arabidopsis, capsicum and eggplant.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu G, Guan Z, Ma M, Wang H, Liu X, et al. 2023. Genome-wide identification and molecular characterization of SlKRP family members in tomato and their expression profiles in response to abiotic stress. Vegetable Research 3:27 doi: 10.48130/VR-2023-0027 

Genome-wide identification and molecular characterization of SlKRP family members in tomato and their expression profiles in response to abiotic stress
- Received: 27 February 2023
- Accepted: 13 July 2023
- Published online: 10 October 2023
An Author Correction to this article was published on 31 January 2024,
http://doi.org/10.48130/vegres-0024-0006.
Abstract: The cell cycle has an essential role in the regulation of plant growth, development, and stress responses, which is controlled by the complex of cyclin / cyclin-dependent kinases (CDKs). Kip-related proteins (KRPs) as CDK inhibitors are involved in the precise regulation of cell cycle progression. However, the comprehensive identification of SlKRP family genes in tomato has not been achieved. Here, a total of six SlKRP proteins were identified from the tomato genome and divided into three classes via phylogenetic analysis. Chromosomal localization analysis revealed the interchromosomal segment duplication among SlKRP genes. Analyses on gene structures and conserved motifs indicated that the SlKRP genes were evolutionarily conserved. The subcellular localization analysis showed all SlKRP proteins were located in the nuclei. Six SlKRPs had distinct expression in different tissues. Their expressions were affected by the plant hormones (ABA, IAA, and ethylene) and various abiotic stresses (salt, drought, and low temperature), which were correlated with different cis-acting regulatory elements (CAREs) in the 3-Kb promoter regions of these genes. In addition, co-expression relationship and protein interaction network analysis proved that SlKRP proteins may interact with CDKs and cyclins. To further explore the function of SlKRPs in tomato, VIGS assay was performed to obtain SlKRP5-silenced plants and demonstrated that silencing of SlKRP5 increased the sensitivity to drought stress. These findings provide references for the further functional analysis of KRPs in the future.
-
Key words:
- Cell cycle /
- CDK inhibitors /
- KRP /
- Comprehensive identification /
- Tomato /
- Abiotic stresses