










-
Cacao (Theobroma cacao L.) is a tropical species native to the Amazon basin of South America[1]. Owing to the main ingredient of chocolate, cocoa has a high economic value and is widely cultivated throughout the world's tropical regions[2−4]. The livelihoods of 40-50 million people worldwide depend on the production of cocoa beans, which amounted to 5.7 million tons of production from cultivated areas of 12.3 million hectares in 2020[5]. The antioxidant, cardiovascular protective and anti-inflammatory properties of cocoa beans are due to their abundance in theobromine, flavonoids, and other vital components[6−8]. Cocoa is also used in beverages, desserts, cosmetics and other industries[1,9,10].
Given the late-acting or ovarian form of self-incompatibility, cross-pollination operates at higher fruit set rates and generates plenty of genetic diversity for the natural cacao germplasm[4,11]. Generally, cocoa has been subdivided into three traditional groups: Criollo, Forastero, and Trinitario, based on morphological traits and geographical origins[4,12]. The Trinitario group has been recognized as the hybrids of Criollo and Forastero[13,14]. It should be noted that Motomayor et al.[13] proposed a new classification system for cacao germplasm using microsatellite marker technology, which included nine new groups and the Criollo group. There are large differences in trait phenotypes between cocoa groups[13,15].
Unfermented cocoa beans show a high bitterness and astringency taste, hard for consumers to accept[16,17]. Products from roasted and fermented cocoa beans can obtain the distinctive cocoa flavor[18,19]. Some investigations have proven that the content of theobromine, caffeine, and polyphenols are decreased to different degrees after the fermentation process[20−22]. The most crucial aspect for the consumer market is the aroma composition of cocoa bean, which also affects the bean quality[23]. Numerous volatile organic compounds, including volatile (acids, alcohol, ketone, etc.) and non-volatile organic compounds (amino acids, sugar, etc.), which are generated during the fermentation, drying, and roasting of the beans, contribute to the aroma of cocoa[19,24].
As a generalization, the cocoa groups mentioned above are classified commercially into fine flavor cocoa (Criollo and Trinitario) and bulk cocoa (Forastero) based on their aroma texture. A significant amount of the world's production of cocoa is in bulk, compared to 12% of the fine flavor cocoa, it lacks floral, fruity, and other premium aromas[25]. Of course, high-quality aroma is only an important basis in classification, and it is also related to many factors, such as planting and processing methods[25]. The demand for fine flavor cocoa on the commercial market is constantly increasing, which encourages the urgent expansion of new sources of fine flavor cocoa[14,25,26].
The compositions of the volatile compounds released by cocoa beans of various genetic groups have already been the subject of previous studies[27−29]. For the cocoa beans from Hainan Province, China, high levels of furans, hydrocarbons, alcohols and esters were found in unfermented Trinitario beans. Very high levels of hydrocarbons, alcohol and acids were characterized in Criollo unfermented beans. However, Forastero has no distinctive performance in terms of volatile compounds[28]. As for the fermented bean samples from Ecuador, Criollo generated volatiles of floral, fruity and woody aromas, while Forastero generated a large number of floral and sweet volatiles[29]. Criollo could be well identified in terms of aromatic composition, which distinguishes it from the other two groups[27].
In contrast to cultivation, fermentation, or roasting processes, the quality and aroma of fermented and unfermented beans in the Criollo, Forastero and Trinitario groups from the same region have not yet been investigated. Therefore, the present study aims to assess the quality phenotype and volatile characteristics variations of fermented and unfermented Criollo, Forastero and Trinitario cocoa beans. The investigation results will be used to evaluate volatile polymorphisms in different groups of cocoa beans in China, thus providing aromatic information as the basis for future breeding or biotechnological applications.
-
In the present study, nine cacao lines representing the three genetic groups, Criollo, Forastero, and Trinitario, were cultivated in the Cacao Germplasm Repository of the Spice and Beverage Research Institute, CATAS, Hainan Province, China. All the germplasm was introduced from southeast Asia and west Africa, and studies were conducted to select germplasm with prominent phenotypes from natural hybrid offspring. It should be noted that the selected Forastero lines were introduced from Africa and they have the characteristics of the Amelonado variety. Additionally, the three Criollo lines were introduced from Indonesia and showed typical characteristic traits. During the major ripening period (February to April) of 2022, 15 to 20 healthy cacao pods were harvested from each line respectively, then transported to the laboratory, where they were dealt with individually as per standard methods in a timely manner. The pods were opened and the beans were taken out. Beans of three pods were washed with clean water and dried at 45 °C in a drying oven (Blast high-temperature oven, Shanghai Heheng Instrument Equipment Co., Ltd., China) to a moisture content of 5%−7% for the unfermented treatment beans. The remaining beans were placed in a jar and fermented, temperature-controlled in an incubator. The fermentation process was as follows: 48 h at 35 °C, 48 h at 40 °C, 48 h at 45 °C, and finally 24 h at 50 °C. During this whole process, the beans were mixed well after every 24 h, and excess fermentation broth was removed. Finally, the fermented beans were also dried in a 45 °C oven to a moisture content of 5%−7%, turned every 2 h for the first 12 h, and then every 8 h to ensure even heating and easier drying.
During sampling, the beans were ground in a rotating blade mill (JC-FW200, Qingdao Juchuang Jiaheng Analytical Instrument Co., Ltd., China) and sieved through a pore size of 0.425 mm. The powder obtained was finally stored at −20 °C until sample preparation. The unfermented samples of three groups were designated as C01, C02, C03 (Criollo), F01, F02, F03 (Forastero), and T01, T02, T03 (Trinitario), and the fermented samples of three groups were correspondingly designated as C11, C12, C13 (Criollo), F11, F12, F13 (Forastero), and T11, T12, T13 (Trinitario). The details of each line are listed in Supplemental Table S1.
Determination of cocoa butter
-
Cocoa butter content was analyzed through ultrasound-assisted extraction using a previously validated method of petroleum ether extraction[30]. Approximately 1.50 g of each sample was extracted with 15.00 mL of petroleum ether using an ultrasonic bath at a fixed power of 250 W (FRQ-1006HT, Hangzhou Frante Ultrasonic Technology Co., Ltd., China) for 10 min and centrifuged for 10 min at 8,000 rpm. The extraction process was repeated three times for each sample. Then, the obtained precipitates were vacuum-dried, and weighed after drying. The content of cocoa butter is calculated by dividing the quantity difference by the sample quality.
Theobromine and caffeine determination
-
In the quantification of theobromine and caffeine levels, each sample was analyzed using a previously used liquid chromatography method[31]. Firstly, 0.50 g of cocoa powder was weighed, then operated as follows: adding 90.00 mL of distilled water, extracting by ultrasonic (room temperature, 30 min, 250 W), diluting to 100.00 mL, and finally filtering with the pore size of 0.22 μm filter (13 mm × 0.22 μm, nylon, ANPEL Laboratory Technologies (Shanghai) Inc. China). The samples were analyzed by a semi-preparative HPLC Agilent 1260 Series LC system (Agilent Technologies, Palo Alto, CA, USA) equipped with an auto sampler, a diode array detector (DAD) set to 275 nm. The compounds were separated on a Zorbax column (XDB-C18, 250 × 4.6 mm × 1.8 μm) at 40 °C. The flow system consisted of acetic acid to methanol and water in a ratio of 1:20:79, with an injection volume of 10.00 μL and a flow rate of 1.00 mL/min.
Polyphenol determination
-
The Folin-Ciocalteu method, with some changes, was employed to determine the total content of polyphenols[32]. Approximately 0.20 g of each sample was extracted with 10.00 mL of a 70% methanol solution using a water bath for 10 min at 70 °C and centrifuged for 10 min at 8,000 rpm. The supernatant was filtered through a filter membrane with a 0.22 μm filter, and then the obtained supernatant was diluted 100 times with distilled water. Subsequently, 1.00 mL of the diluted extract was transferred to a 15.00 mL centrifuge tube and reacted with 5.00 mL of 10% Folin-Ciocalteu solution for 5 min, followed by the addition of 4.00 mL of 7.5% Na2CO3 solution for 60 min. Finally, the absorbance of the solution at 765 nm was measured in comparison with the standard curve of prepared gallic acid solutions using a UV-Vis spectrophotometer (UV2310 II, Shanghai, China). All tests were carried out in triplicate. Polyphenol content was expressed as gallic acid equivalents (GAE).
Determination of volatile compounds
-
Volatile compounds were performed using a previously reported Head-Space Solid Phase Micro-Extraction (HS-SPME) and Gas Chromatography-Mass Spectrometry (GC-MS) method[33]. Approximately 2.00 g of each sample was placed in a 10.00 mL head-space vial. The sample was incubated at 60 °C for 10 min, then the fiber (50/30 μm DVB/CAR/PMDS, Agilent, USA) was exposed for 30 min at the same temperature. Subsequently, the volatile compounds were analyzed by GC-MS (7890B-5977B, Agilent, USA), equipped with a DB-WAX column (30 m × 0.25 mm × 0.25 μm). The operating conditions were programmed as follows: 40 °C of initial oven temperature for 4 min, from 40 to 96 °C at 3 °C/min, from 96 to 150 °C at 1.5 °C/min, from 150 to 210 °C at 10 °C/min. High purity helium gas was used as a carrier gas at a flow rate of 1.00 mL/min. The injector was operated in split less mode at 230 °C with an inlet and quadruple temperature of 250 °C and a scan range of 30 to 450 m/z in full scan mode. The identification of volatile compounds was conducted by comparing the mass spectra of the compounds in the samples with the NIST 17 Library. An alkane series (C7-C30) was a run under the same GC-MS conditions to obtain linear retention indices (LRI) for non-isothermal Kovats. Aroma descriptors for the compounds were identified by the online databases VCF 16.9 (
www.vcf-online.nl ), and the Good Scents Company (www.thegoodscentscompany.com/indeX.html ).Statistical analysis
-
A one-way ANOVA was employed to test the variation of quality traits among the different cacao groups (p < 0.05) by the Duncan method. Venn diagrams were performed on BioLadder platform (
www.bioladder.cn ). Origin 2021 is used for PCA analysis and drawing bar charts. Box plot, clustering heat maps, and correlation heat maps were conducted using R package version 4.0.4 for Windows. The clustering method was hierarchical clustering. -
The cocoa butter content of unfermented beans were higher than those of fermented beans in the three cacao groups[20], with a range of 37.98%−44.00% (Table 1). Statistical analysis indicated that Trinitario showed a significant change(p < 0.05) in cocoa butter content after fermentation, with a reduction of 5.70%. It is well known that cocoa butter plays a crucial role in the production of chocolate, and variations in its content may have a direct impact on the taste buds and physical characteristics of cocoa products[16,34].
Table 1. Quality traits of unfermented and fermented beans for the three cacao groups.
Germplasm Cocoa butter
(%)Theobromine
(mg/g)Caffeine
(mg/g)Polyphenol
(mg GAE/g)F0 42.37 ± 1.94ab 13.83 ± 0.50a 1.92 ± 0.83a 63.54 ± 4.69a C0 42.12 ± 1.46ab 14.49 ± 0.84a 1.24 ± 0.11a 64.40 ± 2.96a T0 44.00 ± 2.24a 13.56 ± 1.67a 2.69 ± 1.10a 66.21 ± 1.92a F1 38.76 ± 2.58b 12.01 ± 1.24a 1.73 ± 0.75a 53.64 ± 6.53ab C1 37.98 ± 1.93b 11.68 ± 1.53a 1.18 ± 0.09a 45.35 ± 12.44b T1 38.30 ± 1.36b 12.49 ± 1.18a 2.04 ± 0.58a 46.41 ± 5.49b The value for each group is the mean ± SD of the triplicate material in the group. Mean values assigned with a common letter within the same column are not significantly different according to Duncan's multiple range tests at the 5% level. Theobromine and caffeine content
-
The maximum contents of theobromine and caffeine were found in Forastero (13.83 mg/g) and Trinitario (2.69 mg/g) in unfermented beans. Measurements of theobromine and caffeine were consistent with previous research, resulting in lower levels after fermentation due to their diversion and diffusion during the fermentation process[22,35,36]. As the fermented beans with the lowest theobromine and caffeine concentrations (11.68 and 1.18 mg/g, respectively), Criollo is less bitter than Forastero and Trinitario[37].
Total polyphenol content
-
In the three groups of cocoa beans, the total polyphenol content ranged from 45.35 to 66.21 mg GAE/g (GAE, gallic acid equivalent)[6]. The astringent taste of cocoa beans is mainly caused by polyphenols, but fermentation reduces their content to mitigate this taste[21,38]. The total polyphenol content of Criollo and Trinitario was statistically significantly decreased after fermentation (p < 0.05). The fermented beans of Criollo showed the smallest total polyphenol content and are therefore less astringent[18,39].
We discuss this based on the average values of three germplasm within the same group. The average data for all quality traits showed a decrease in the level of quality after fermentation. But it has to be said that after fermentation, the caffeine content of F01 and the polyphenol content of T01 have both increased (Supplemental Table S2). The fermentation conditions of cocoa beans is difficult to control in the natural environment, and susceptible to environmental influences[40], and the quality of fermented beans fluctuates widely[39,41]. The fermentation treatment in this study was performed under strict control conditions in the room, avoiding the interference of environmental factors. Therefore, the changes or differences in different cacao groups are mainly caused by genetic background[38]. However, it is undeniable that these quality traits are also influenced to a large extent by environmental, fermentation, and drying factors[18,22,36,38,42].
Volatile compounds
-
Similarly, the mean of the three materials in the group were represented as the corresponding cacao group. Forty-eight volatile compounds were detected in unfermented beans and 79 volatile compounds were detected in fermented cocoa beans from the three groups (Table 2 & Fig. 1c). A total of 81 volatile compounds were detected, and the same classes of compounds were found in other experiments, including 19 alcohols, 21 esters, eight acids, eight aldehydes, seven ketones, four hydrocarbons, three pyrazines, two furans, four lactones and other types of volatile components (two phenols, one pyridine, one sulfide, and one pyrrole) collectively[26,37].
Table 2. Volatile compounds before and after fermentation of three groups cocoa beans.
Code Compound LRI Relative content (%) Aroma description F0 C0 T0 F1 C1 T1 Alcohols A1 2-Methyl-3-buten-2-ol 1002 1.98 ± 0.58 0.65 ± 0.65 1.01 ± 0.53 0.03 ± 0.03 0.08 ± 0.08 0.01 ± 0.01 Herbal, earth, oily A2 Isobutanol 1068 0.46 ± 0.27 0.28 ± 0.26 0.47 ± 0.31 0.07 ± 0.03 0.22 ± 0.14 0.10 ± 0.07 Apple, cocoa, wine A3 2-Pentanol 1108 46.33 ± 13.39 29.22 ± 7.88 38.31 ± 24.80 NA NA NA Green, fusel oil A4 3-Methyl-1-butanol 1202 3.37 ± 1.45 3.85 ± 3.29 5.05 ± 0.69 0.32 ± 0.22 1.44 ± 1.17 0.72 ± 0.08 Banana, fruity, fusel oil A5 1-Pentanol 1247 0.17 ± 0.17 0.12 ± 0.12 0.06 ± 0.04 NA 0.03 ± 0.03 NA Fruity, green A6 2-Hexanol 1311 0.50 ± 0.50 0.26 ± 0.01 0.43 ± 0.14 NA NA NA Wine, fruity, fatty A7 2-Heptanol 1322 1.04 ± 1.72 6.31 ± 3.44 6.80 ± 1.27 0.06 ± 0.05 0.51 ± 0.57 0.13 ± 0.07 Fruity, citrus A8 1-Hexanol 1342 0.18 ± 0.18 0.40 ± 0.05 0.23 ± 0.04 NA 0.04 ± 0.04 NA Green, herbal, fruity A9 3-Ethoxy-1-propanol 1374 NA NA NA 0.06 ± 0.06 0.13 ± 0.08 0.06 ± 0.02 Fruity A10 2-Octanol 1421 NA NA NA 0.08 ± 0.05 0.06 ± 0.02 NA Spicy, green, wood A11 2-Nonanol 1519 NA 0.27 ± 0.10 0.83 ± 0.83 0.19 ± 0.08 1.01 ± 0.21 0.27 ± 0.20 Fruity, green A12 2,3-Butanediol 1536 1.03 ± 0.91 1.55 ± 0.62 1.19 ± 0.55 13.24 ± 7.34 10.22 ± 4.77 12.89 ± 2.84 Creamy, fruity A13 Linalool 1544 0.49 ± 0.49 0.30 ± 0.30 0.35 ± 0.16 0.27 ± 0.16 0.16 ± 0.14 0.08 ± 0.02 Rose, floral, green A14 1-Octanol 1555 NA 0.14 ± 0.14 0.07 ± 0.07 0.03 ± 0.03 0.08 ± 0.08 0.05 ± 0.01 Waxy, green A15 1,3-Butanediol 1574 3.15 ± 1.97 3.12 ± 1.55 4.23 ± 3.31 4.74 ± 1.67 4.04 ± 0.93 4.74 ± 0.87 A16 Propylene glycol 1587 NA 0.13 ± 0.13 0.08 ± 0.06 0.07 ± 0.07 0.11 ± 0.04 0.07 ± 0.03 A17 1-Phenylethanol 1798 0.38 ± 0.38 NA 0.17 ± 0.17 0.09 ± 0.05 0.20 ± 0.15 0.04 ± 0.00 Floral, honey, rose A18 Benzyl alcohol 1857 NA NA NA 0.02 ± 0.00 0.12 ± 0.08 0.03 ± 0.02 Floral, rose A19 2-Phenylethanol 1890 0.63 ± 0.21 1.19 ± 0.19 1.42 ± 0.75 6.17 ± 5.01 18.88 ± 13.74 8.10 ± 6.24 Floral, honey, rose Esters E1 Ethyl acetate 903 10.09 ± 4.66 13.03 ± 4.23 9.70 ± 6.52 2.34 ± 2.08 8.94 ± 6.12 3.90 ± 2.51 Fruity, sweet, grape E2 Isobutyl acetate 973 NA NA NA 0.12 ± 0.01 0.10 ± 0.03 0.12 ± 0.01 Apple, floral, herbal E3 Ethyl butanoate 994 0.57 ± 0.57 0.74 ± 0.18 0.15 ± 0.15 NA 0.12 ± 0.12 NA Fruity, green, apple E4 2-Pentyl acetate 1039 1.53 ± 0.32 7.32 ± 3.01 1.92 ± 0.84 0.03 ± 0.01 0.05 ± 0.05 0.05 ± 0.01 Herbal, fruity, green E5 Isopentyl acetate 1104 NA NA NA 0.72 ± 0.44 1.01 ± 0.02 1.34 ± 0.50 Fruity, sweet, banana E6 Ethyl hexanoate 1225 0.27 ± 0.27 0.26 ± 0.16 0.22 ± 0.13 0.13 ± 0.10 0.21 ± 0.13 0.12 ± 0.14 Fruity, green, banana E7 2-Heptyl acetate 1257 0.19 ± 0.19 1.95 ± 0.58 0.27 ± 0.27 0.04 ± 0.04 0.17 ± 0.17 0.13 ± 0.13 Fenugreek, fruity E8 Ethyl heptanoate 1329 NA NA 0.07 ± 0.07 0.03 ± 0.01 0.04 ± 0.02 0.02 ± 0.02 Fruit, wine E9 Acetoin acetate 1377 NA NA NA 0.32 ± 0.15 0.19 ± 0.11 0.31 ± 0.14 Sweet, creamy E10 Ethyl octanoate 1429 NA NA NA 0.09 ± 0.03 0.22 ± 0.12 0.09 ± 0.08 Fruity, apricot, brandy E11 2-Hydroxyethyl acetate 1627 NA NA NA 0.19 ± 0.04 NA 0.19 ± 0.04 E12 Ethyl decanoate 1630 NA NA NA 0.01 ± 0.01 0.02 ± 0.01 NA Grape, brandy, grape E13 Diethyl succinate 1667 NA NA 0.22 ± 0.22 NA 0.24 ± 0.05 NA Fruity, floral E14 1-Phenylethyl acetate 1684 NA 0.11 ± 0.11 0.08 ± 0.08 0.02 ± 0.00 0.02 ± 0.00 0.02 ± 0.02 Green, fruity E15 Ethyl phenylacetate 1766 NA 0.11 ± 0.11 NA 0.08 ± 0.02 0.15 ± 0.06 0.12 ± 0.05 Floral, rose, honey E16 2-Phenylethyl acetate 1793 NA NA NA 2.58 ± 2.01 2.94 ± 2.08 3.14 ± 0.50 Floral, rose, honey E17 Isobutyl benzoate 1816 NA NA 0.44 ± 0.17 0.04 ± 0.01 0.04 ± 0.02 0.07 ± 0.01 Sweet, fruity E18 Ethyl laurate 1834 NA NA NA 0.02 ± 0.02 0.02 ± 0.00 0.01 ± 0.01 Sweety, floral E19 Ethyl cinnamate 2097 NA NA NA 0.02 ± 0.02 0.03 ± 0.02 0.02 ± 0.02 Balsamic, fruity E20 Eugenyl acetate 2239 NA NA NA 0.02 ± 0.02 0.17 ± 0.17 0.25 ± 0.25 Woody, clove, floral E21 Ethyl palmitate 2246 NA NA NA 0.12 ± 0.01 0.13 ± 0.07 0.12 ± 0.02 Waxy Acids Ac1 Acetic acid 1443 6.30 ± 3.52 9.15 ± 0.82 6.22 ± 4.56 52.56 ± 9.01 35.42 ± 20.58 52.57 ± 8.27 Sour, vinegar, pungent Ac2 Propanoic acid 1531 NA NA NA 0.06 ± 0.03 0.04 ± 0.04 0.04 ± 0.01 Acidic, pungent, cheesy Ac3 Isobutyric acid 1561 NA NA NA 0.28 ± 0.17 0.30 ± 0.22 0.20 ± 0.04 Acidic, buttery, cheese Ac4 Butanoic acid 1622 NA NA NA 0.03 ± 0.01 0.03 ± 0.01 0.03 ± 0.03 Acetic, cheese, butter Ac5 2-Methylbutanoic acid 1662 NA NA NA 2.94 ± 2.94 0.28 ± 0.28 NA Pungent, cheese Ac6 Isovaleric acid 1664 0.30 ± 0.30 0.36 ± 0.07 0.17 ± 0.08 0.68 ± 0.42 0.68 ± 0.49 0.58 ± 0.14 Sour, stinky Ac7 Hexanoic acid 1839 NA NA NA 0.03 ± 0.01 0.05 ± 0.03 0.03 ± 0.00 Sour, fatty Ac8 Octanoic acid 2056 NA NA NA 0.05 ± 0.01 0.05 ± 0.02 0.04 ± 0.01 Acid, cheese Aldehydes Ald1 Hexanal 1046 0.24 ± 0.24 0.54 ± 0.54 0.37 ± 0.37 NA 0.01 ± 0.01 NA Grass, fresh, fruity Ald2 Octanal 1279 NA 0.12 ± 0.12 0.10 ± 0.05 0.02 ± 0.01 0.02 ± 0.01 NA Citrus, fatty Ald3 Nonanal 1387 1.19 ± 0.78 0.99 ± 0.67 0.77 ± 0.47 0.15 ± 0.02 0.14 ± 0.07 0.13 ± 0.07 Citrus, fatty Ald4 2-Furaldehyde 1448 0.58 ± 0.58 0.54 ± 0.54 NA NA 0.15 ± 0.15 0.14 ± 0.14 Baked bread, almond Ald5 Decanal 1480 NA NA NA 0.07 ± 0.02 0.04 ± 0.02 0.27 ± 0.01 Orange peel, floral Ald6 Benzaldehyde 1503 0.13 ± 0.13 NA NA 0.10 ± 0.06 0.35 ± 0.27 0.17 ± 0.09 Almond, cherry Ald7 Phenylacetaldehyde 1619 NA NA NA 0.05 ± 0.02 0.08 ± 0.03 0.04 ± 0.03 Honey, nutty Ald8 2-Phenyl-2-butenal 1899 NA NA NA NA 0.09 ± 0.06 0.03 ± 0.03 Cocoa, honey, roast Ketones K1 2-Pentanone 946 11.10 ± 1.17 9.06 ± 0.52 10.71 ± 5.75 0.26 ± 0.25 0.13 ± 0.09 0.12 ± 0.02 Fruity K2 2-Heptanone 1166 0.57 ± 0.71 2.64 ± 1.20 4.01 ± 3.35 0.10 ± 0.10 0.24 ± 0.22 0.08 ± 0.07 Fruity, green K3 Acetoin 1271 1.02 ± 0.26 0.84 ± 0.21 0.69 ± 0.45 3.67 ± 2.59 1.63 ± 0.74 2.09 ± 0.39 Buttery, creamy K4 Hydroxyacetone 1285 NA 0.13 ± 0.13 NA 0.02 ± 0.00 0.03 ± 0.02 0.02 ± 0.01 Caramellic, buttery K5 2-Hydroxy-3-pentanone 1348 NA NA NA 0.19 ± 0.19 0.04 ± 0.02 0.02 ± 0.02 Truffle, earthy, nutty K6 2-Nonanone 1383 NA 0.70 ± 0.06 0.26 ± 0.26 0.05 ± 0.05 1.73 ± 1.46 0.03 ± 0.03 Green, herbal K7 Acetophenone 1625 3.20 ± 3.20 0.17 ± 0.17 0.72 ± 0.43 0.35 ± 0.10 0.65 ± 0.37 NA Floral, almond Hydrocarbons H1 β-Myrcene 1148 0.98 ± 0.98 NA 0.66 ± 0.41 0.22 ± 0.16 0.13 ± 0.10 0.04 ± 0.01 Balsam, spicy H2 Styrene 1242 NA NA NA 0.03 ± 0.01 0.02 ± 0.00 0.02 ± 0.02 Balsam, plastic H3 Alloocimene 1366 NA NA NA 0.08 ± 0.08 0.02 ± 0.02 NA Floral, herbal H4 α-Humulene 1643 0.38 ± 0.10 0.29 ± 0.29 0.48 ± 0.26 0.08 ± 0.03 0.08 ± 0.02 0.05 ± 0.01 Woody Pyrazines P1 2,3-Dimethylpyrazine 1338 0.16 ± 0.12 0.16 ± 0.11 0.11 ± 0.03 0.25 ± 0.20 3.31 ± 3.86 0.27 ± 0.03 Caramel, roasted, nutty P2 Trimethylpyrazine 1397 NA NA NA 0.18 ± 0.16 0.06 ± 0.03 0.18 ± 0.02 Nutty, cocoa, baked potato P3 Tetramethylpyrazine 1464 NA NA NA 4.43 ± 6.06 0.51 ± 0.36 4.27 ± 1.19 Chocolate, coffee, cocoa Furans F1 trans-Furan linalool oxide 1461 0.66 ± 0.66 NA 0.21 ± 0.05 0.05 ± 0.03 0.07 ± 0.04 0.05 ± 0.01 Floral, citrus F2 Furaneol 2017 NA NA NA 0.02 ± 0.02 0.05 ± 0.05 0.05 ± 0.05 Caramel, sweet, strawberry Lactone L1 γ-Valerolactone 1583 0.15 ± 0.15 NA NA 0.04 ± 0.03 NA 0.09 ± 0.09 Herbal, sweet L2 γ-Butyrolactone 1599 0.57 ± 0.32 0.56 ± 0.47 0.60 ± 0.76 0.30 ± 0.16 0.50 ± 0.18 0.42 ± 0.36 Creamy, caramel L3 Pantolactone 2001 NA NA NA 0.08 ± 0.03 0.12 ± 0.06 0.10 ± 0.05 Cotton candy, burnt L4 5-Acetyldihydro-2(3H)-furanone 2024 NA NA NA 0.02 ± 0.00 0.04 ± 0.02 0.06 ± 0.01 Wine Others O1 Methionol 1704 NA 0.11 ± 0.11 NA 0.06 ± 0.06 0.43 ± 0.39 0.12 ± 0.17 Onion, soup O2 trans-Pyran linalool oxide 1756 NA NA NA 0.03 ± 0.01 0.04 ± 0.01 0.03 ± 0.01 Floral, sweet O3 2-Acetylpyrrole 1946 NA NA NA 0.04 ± 0.01 0.03 ± 0.02 0.07 ± 0.02 Bready, cocoa, hazelnut O4 Eugenol 2152 0.12 ± 0.12 0.39 ± 0.39 0.17 ± 0.17 0.03 ± 0.03 0.18 ± 0.18 0.04 ± 0.02 Clove, spicy O5 Elemicin 2207 NA 1.93 ± 1.93 NA 0.04 ± 0.03 0.13 ± 0.16 0.20 ± 0.20 Spicy, floral The value for each group is the mean ± SD of the triplicate material in the group. 'NA' indicates that the substance has not been detected. 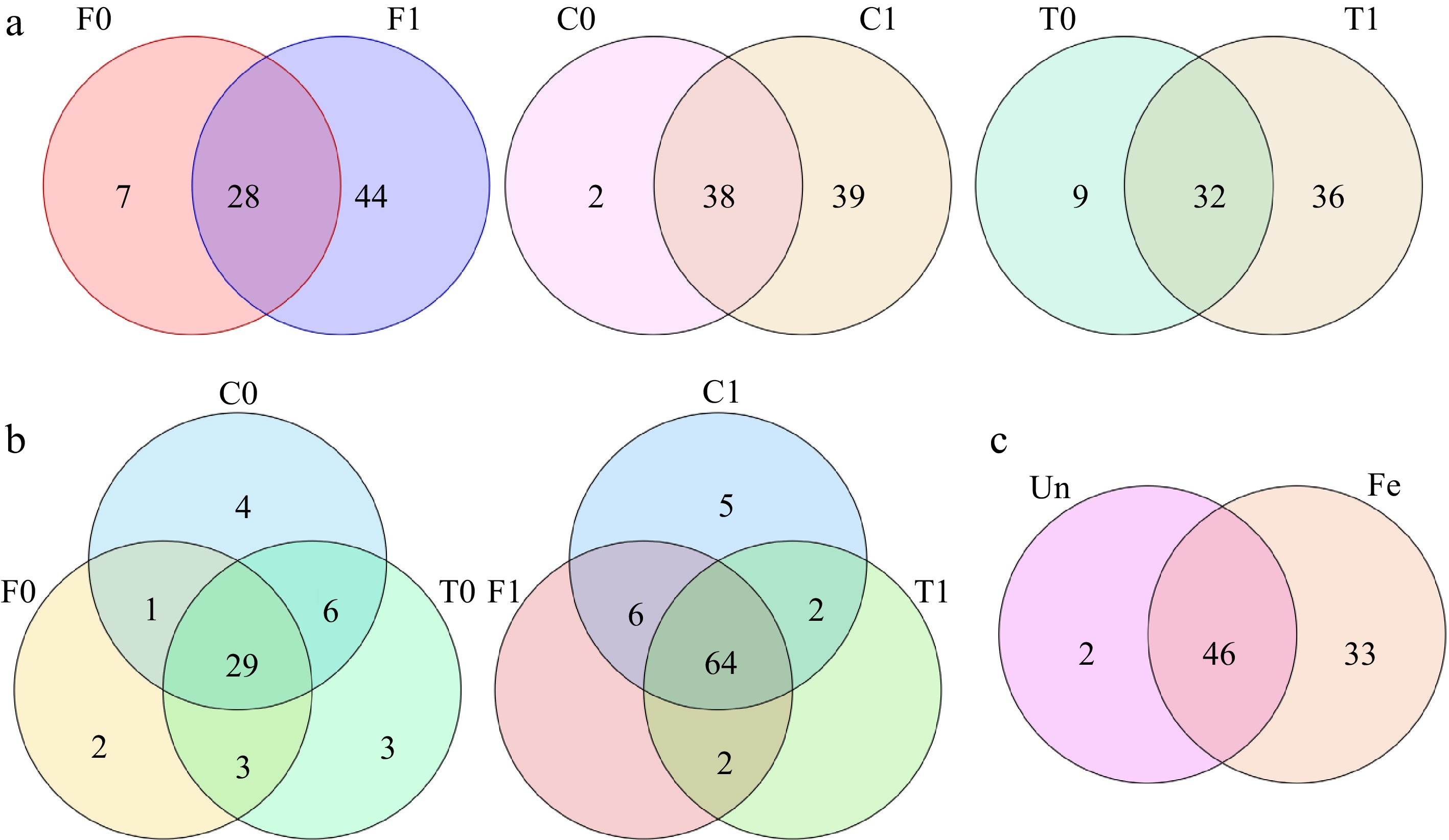
Figure 1.
Venn diagram of volatile compounds. (a) Comparison of the number of volatile compounds in unfermented and fermented beans within the same group. (b) Comparison of the number of volatile compounds between unfermented and fermented beans in the three groups. (c) Number of volatile compounds shared by unfermented and fermented beans in the three groups (Un: unfermented beans, Fe: fermented beans).
Among the unfermented beans, Trinitario had the most volatile compounds (41 compounds), followed by Criollo (40 compounds) and Forastero (35 compounds) contained the lowest compounds (Fig. 2b). On average of each group, the Trinitario group exhibited the highest volatile compounds, followed by the Criollo group. The Forastero group showed the lowest volatile compounds (Fig. 3). The Criollo, Forastero and Trinitario groups exhibited four, two and three specialized volatile compounds respectively (Fig. 1b). Diversity of the volatile component number and average number within groups were converged with different cacao genotypes, in agreement with the results of Qin et al.[28]. Trinitario contained the highest total volatility, approximately twice that of Forastero (Fig. 2a).
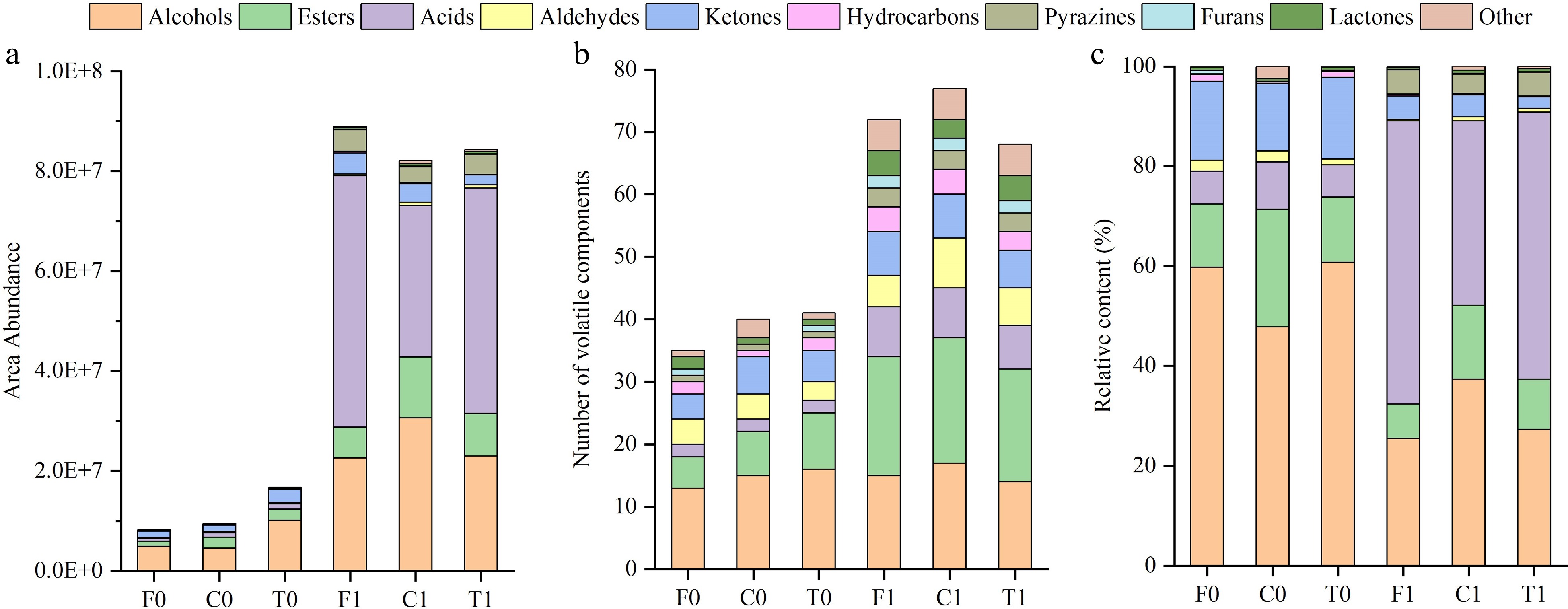
Figure 2.
Volatile compound buildup map. (a) Total area abundance of volatile components. (b) Number of volatile components. (c) Relative content of volatile components.
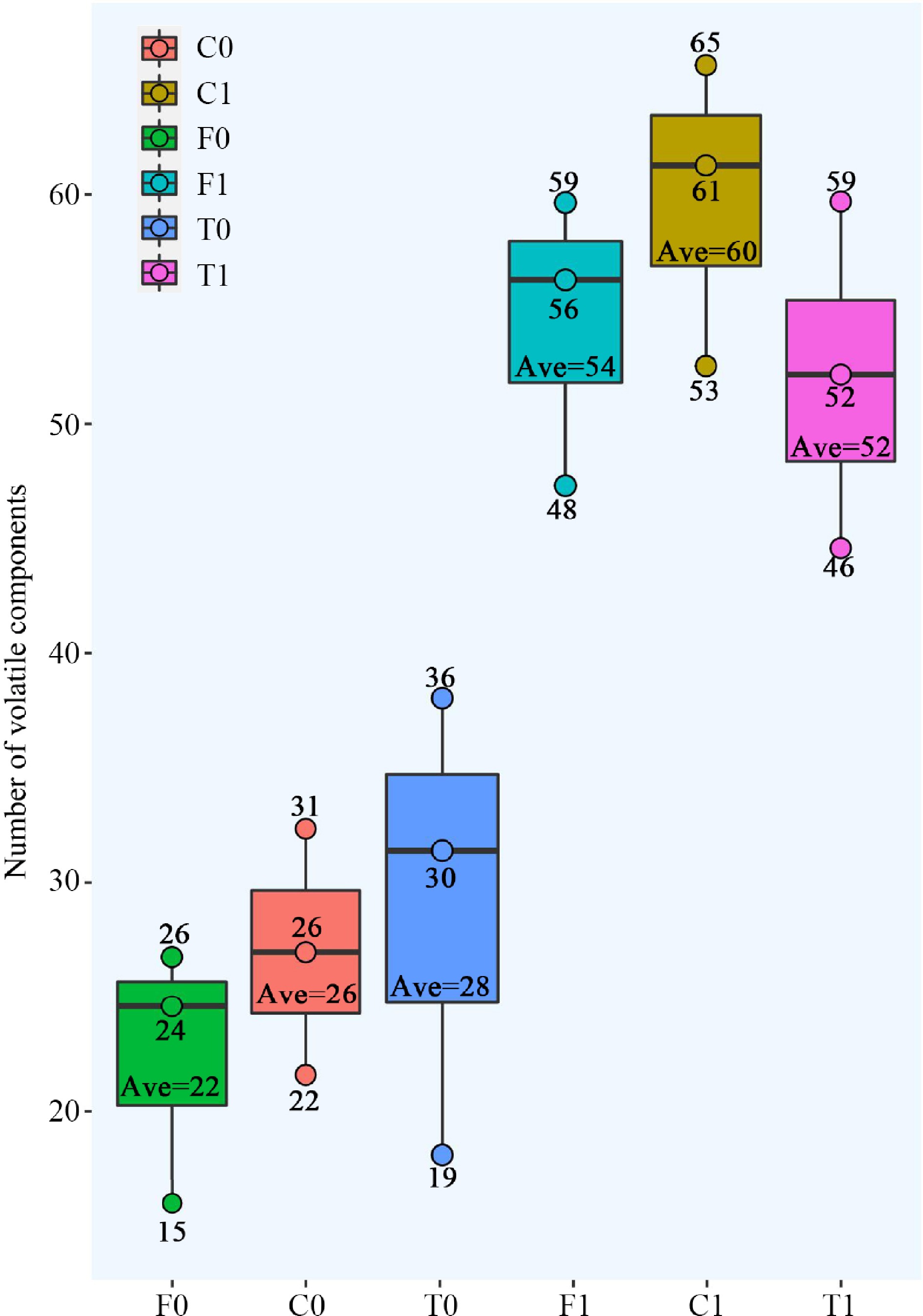
Figure 3.
Number of volatile compounds within the three cacao groups. The three dots on the box represent the number of volatile compounds for the three replicates; 'Ave' represents the average (rounded).
As for the fermented beans, Criollo had the highest number of volatile compounds (77 compounds), followed by Forastero (72 compounds) and Trinitario (68 compounds). On average of each group, the Criollo group exhibited the highest volatile compounds, followed by the Forastero group. The Trinitario group showed the lowest volatile compounds (Fig. 3). Only the Criollo group exhibited five specialized volatile compounds (1-pentanol, 1-hexanol, ethyl butanoate, diethyl succinate, and hexanal) compared to the other two groups (Fig. 1b). These compounds have good aromas (fruity, green, etc.) and may be responsible for the good quality of Criollo.
In both unfermented and fermented beans, there were significant differences in the number of volatile components among the three cacao groups (Fig. 1c). After the fermentation process, the total amount of volatile compounds in cocoa beans increased by 5 to 10-fold (Fig. 2a) and the number of volatile compounds (Fig. 2b) significantly increased. The results revealed that the fermentation process is a crucial step in enhancing the flavor of cocoa beans[24].
Alcohols
-
The relative contents of alcohols was about 50% in unfermented beans and 20%−40% in fermented beans (Fig. 2), where large amounts were essential due to the production of floral, fruity and green aromas[28,40,43]. 2-pentanol was the most abundant substance in the volatile compounds of unfermented beans (Table 2 & Fig. 4). Interestingly, 2-pentanol was not detected in the fermented beans, and the same was true for 2-hexanol (Fig. 1c). While the final profiles of fermented cocoa beans were characterized by a higher content of 2,3-butanediol, 1,3-butanediol and 2-phenylethanol.
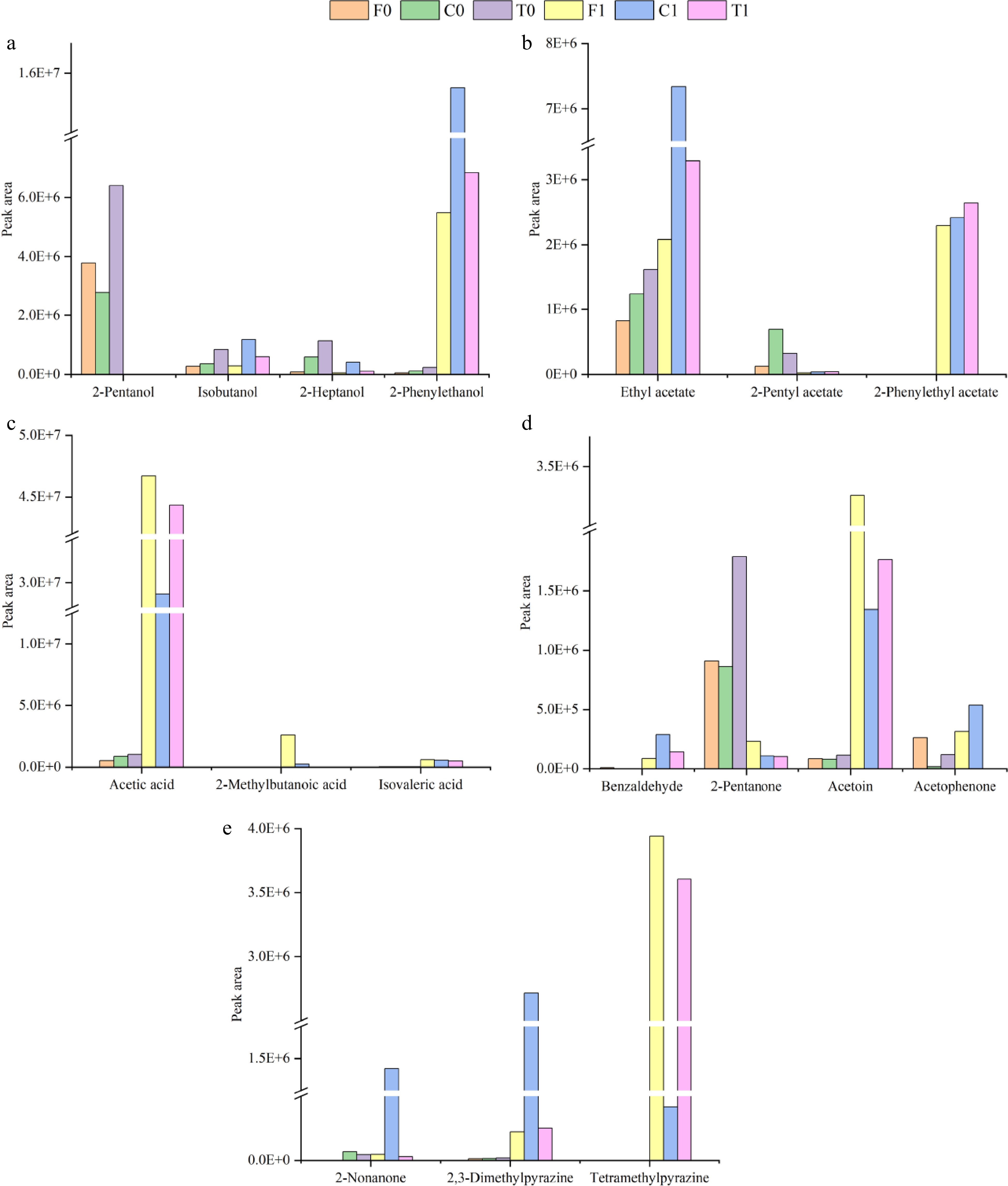
Figure 4.
Peak areas of major volatile compounds.
Isoamyl alcohol, 2-heptanol and 2-phenylethanol are considered important volatile compounds because they produce desirable flavors (fruity, floral)[43,44]. In particular, 2-phenylethanol has been identified as an important compound in fine flavor cocoa[45]. However, the 2-heptanol content was decreased after fermentation (Fig. 4a). The final profile of the unfermented cocoa beans was characterized by the highest content of isoamyl alcohol, 2-heptanol, and 2-phenylethanol in Trinitario, followed by Criollo and Forastero. The characteristics of the fermented cocoa beans were clearly different, with Criollo containing the highest content of the three alcohol compounds mentioned above, in particular producing about twice as much 2-phenylethanol as the other two groups, which gives it a more fruity and floral aroma, followed by Trinitario (Fig. 4a).
Esters
-
The esters are mainly derived from the reaction between organic acids and alcohols during anaerobic fermentation, providing the fruity flavor of cocoa beans[46]. A total of 10 esters were detected before fermentation, which was increased to 21 kinds after fermentation (Fig. 2b). The highest levels of esters were found in the unfermented cocoa beans of Criollo and, interestingly, in the fermented beans as well (Fig. 2c), confirming that esters were the characteristic volatile compounds of Criollo[47]. Ethyl acetate was the most abundant ester in both unfermented and fermented beans, with the amount in fermented beans increasing by a factor of two to four in comparison.
The fruity and green aromas are stronger in Criollo fermented beans because of the highest content of ethyl acetate, which is also one of the main volatile components of cocoa[43]. Directly related to the fine flavor of cocoa is 2-phenylethyl acetate[40], which appeared only after fermentation and released notes of rose and honey, with Trinitario fermented beans showing the highest level and Forastero fermented beans showing the lowest level, although the differences were not significant (Fig. 4b). 2-pentyl acetate, the main volatile compound in cocoa pulp[48], was also present at high levels in the volatile composition of unfermented beans but its level was reduced in fermented beans, likely due to its conversion to other substances during fermentation (Fig. 4b). The excellent flavor of fermented beans is largely due to acetate compounds produced by acetic acid bacteria (AAB)[49], and both the quantity and content of these compounds increased after fermentation.
Acids
-
Through the fermentation process, acids were advanced from two components (acetic acid and isovaleric acid) to eight components (Table 2). The content of acetic acid and isovaleric acid was observed 30 to 90 times higher and 20 times boost in fermented beans compared to unfermented beans. Acetic acid was the highest component of volatile compounds in fermented beans and was also highly odour-active[43,50], with relative levels as high as 35%−52% (Table 2). Acetic acid permeates into the inside of the cocoa beans and forms aromatic compounds (esters and higher alcohols, etc.), influencing the flavor profile of fermented beans[51]. Furthermore, the roasting process adversely affects the concentration of acetic acid, resulting from its high volatility[50,52]. In the current study, acetic acid content was significantly lower in Criollo group fermented beans than those in Trinitario and Forastero groups, supporting the viewpoint of Santander et al.[39]. Generally, the pulp of Criollo group fresh cocoa beans is thinner than that of Trinitario and Forastero groups, in which carbohydrates are consumed and transformed into acetic acid, carbon dioxide, and flavor compounds under the action of yeasts and AAB. 2-methylbutanoic acid also showed a higher content in Forastero fermented beans (Fig. 4c). It is best to avoid producing isovaleric acid in large quantities because it has an unpleasant flavor (souring) and is produced during the fermentation and drying processes[46].
Aldehydes
-
Aldehydes further contribute to the development of desirable flavors and influence the formation of pyrazines[35]. The components of aldehyde in the final volatile profile were different between fermented and unfermented treatments. Eight aldehydes were detected in fermented cocoa beans, which is twice the number of aldehydes in unfermented cocoa beans, and all aldehydes can be found in Criollo fermented beans (Table 2). The aldehyde content of fermented cocoa beans increased about threefold, with phenylethylaldehyde being detected after fermentation and being the main flavour marker of chocolate, imparting floral and other flavours[47]. Benzaldehyde endows roasted almond flavor, which showed the highest content in Criollo and the lowest content in Forastero (Fig. 4d), and it is one of the volatiles that strongly enriched the characteristic aroma of fermented cocoa beans[44].
Ketones
-
Ketones such as 2-pentanone and 2-heptanone, which were natural compounds present in the unfermented beans, decreased throughout fermentation and resulted in a lower content level in the fermented beans. Following fermentation, beans displayed an increased concentration of several key ketones, particularly acetoin, which is known as a technological indicator of cocoa processing and a key component of the buttery and creamy flavors in chocolate[46,50]. Additionally, 2-nonanone was detected in much higher concentrations in the Criollo group fermented beans than the Trinitario and Forastero groups (Fig. 4e), imparting a more fruity flavor[44]. Acetophenone produces floral aromas that contribute to the flavor and quality of the cocoa[35,45], and it is interesting to note that this compound was not detected in Trinitario fermented beans (Fig. 4d).
Pyrazines
-
Pyrazines are mainly generated during the Maillard reaction and Strecker degradation of the roasting process and also form in a small amount during the drying process, imparting roasted incense and nutty notes[35,40,46]. The final profile of fermented beans was characterized by three pyrazine components of 2,3-dimethylpyrazine, trimethylpyrazine, and tetramethylpyrazine. The three group's cocoa beans presented an approximate percentage of pyrazines. Trimethylpyrazine and tetramethylpyrazine are considered important and were mainly produced after fermentation[53], with tetramethylpyrazine being present in large quantities in fermented beans of the Trinitario and Forastero groups, while 2,3-dimethylpyrazine was predominant in Criollo fermented beans (Fig. 4e). However, pyrazine is not thought to be responsible for determining the superior aroma of Criollo[23,54].
Furans, lactones and other compounds
-
Two furans, recognized as trans-furan linalool oxide and furaneol, were found in the present study, of which furaneol was specifically presented in fermented beans. Four types of lactones have been discovered, and interestingly, 5-acetyldihydro-2(3H)-furanone has a wine aroma. Some other volatile compounds, including one pyran, one sulfide, one pyrrole and two phenolic compounds, were detected in the present study and showed differences between fermented and unfermented beans. We detected two phenolic compounds (eugenol and elemicin) that released floral flavors instead of smoky flavors, despite the fact that phenolic compounds are frequently believed to be connected with smoky flavors in cocoa (Table 2)[40,43]. 2-acetylpyrrole is unique to fermented beans and provides the flavor of roasted cocoa[43].
Principal Component Analysis (PCA)
-
Principal component analysis (PCA) is a routinely employed statistical analysis method and has been successfully applied to analytical results, both for individual compounds and component combinations[37]. In order to evaluate the possibility of differentiating the samples taking into account their genotype, this statistical tool was applied. In both unfermented and fermented beans, the two first principal components (PCs) were sufficient to explain more than 99.5% of the variation in the original data (Fig. 5).
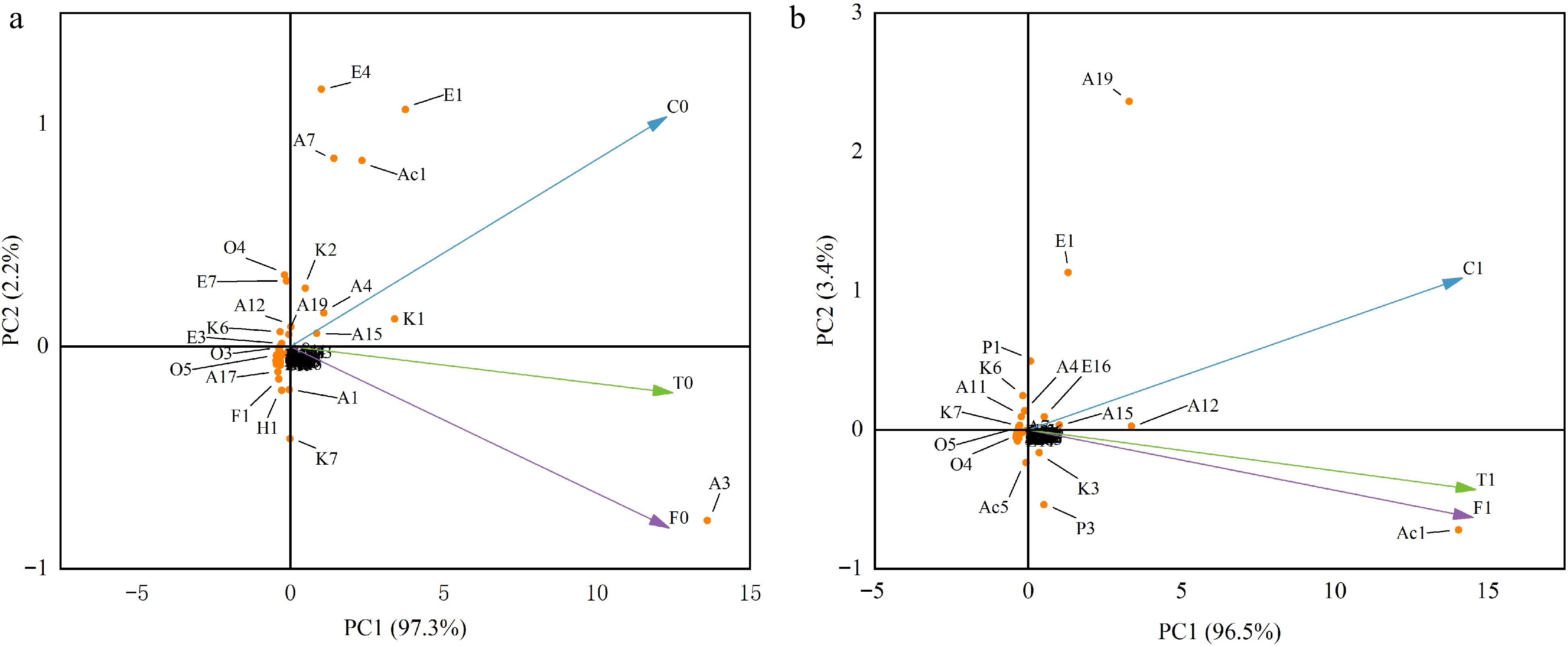
Figure 5.
PCA analysis of unfermented and fermented beans in three cacao groups. (a) unfermented beans. (b) Fermented beans. The code in the figure corresponds to the corresponding compound.
As illustrated in Fig. 5a, the unfermented beans of Criollo, which situated in the positive region of PC2, were clearly differentiated from those of the Trinitario and Forastero groups. Criollo beans combine high contents of 2-heptanol, ethyl acetate, 2-Pentyl acetate and acetic acid, giving it high scores on PC2[27]. Conversely, the high content of 2-pentanol caused the Forastero and Trinitario groups to be located in the negative regions of PC2. Additionally, Trinitario, derived from the hybrid of Criollo and Forastero, was also located in the position of PC2 between the two groups. The positional relationship of the three groups of fermented beans in the principal component analysis was similar to that of unfermented beans (Fig. 5b). 2-phenylethanol (floral), ethyl acetate (fruity), 2-nonanol (fruity, green), and 2,3-dimethylpyrazine (roasted, nutty) were closely related to Criollo and were endowed with floral, fruity, green, roasted, and nutty aroma characteristics. 2,3-butanediol (creamy, fruity), acetic acid (sour), 2-methylbutanoic acid (pungent, cheese), acetoin (buttery, creamy), and tetramethylpyrazine (roasted, nutty) were the main components strongly correlated with the Forastero group, imparting sour, buttery, fruity, toasty and nutty sensory notes. Trinitario is located between them, but leans more towards Forastero.
Cluster heat map and correlation analysis
-
The peak areas of the volatile compounds for each cacao line are shown in Fig. 6, with log10 conversions performed for each parameter. Significant variations were detected among the three cacao groups. For unfermented treatment, Criollo and Trinitario were classified into one cluster, while Forastero and Trinitario were grouped together in fermented treatment (Fig. 6). After the fermentation process, volatile compounds and content were significantly changed, which was also presented in the different lines of the group, indicating that the volatile phenotype was influenced by the complex genetic background of the cacao lines[4,40,53,55].
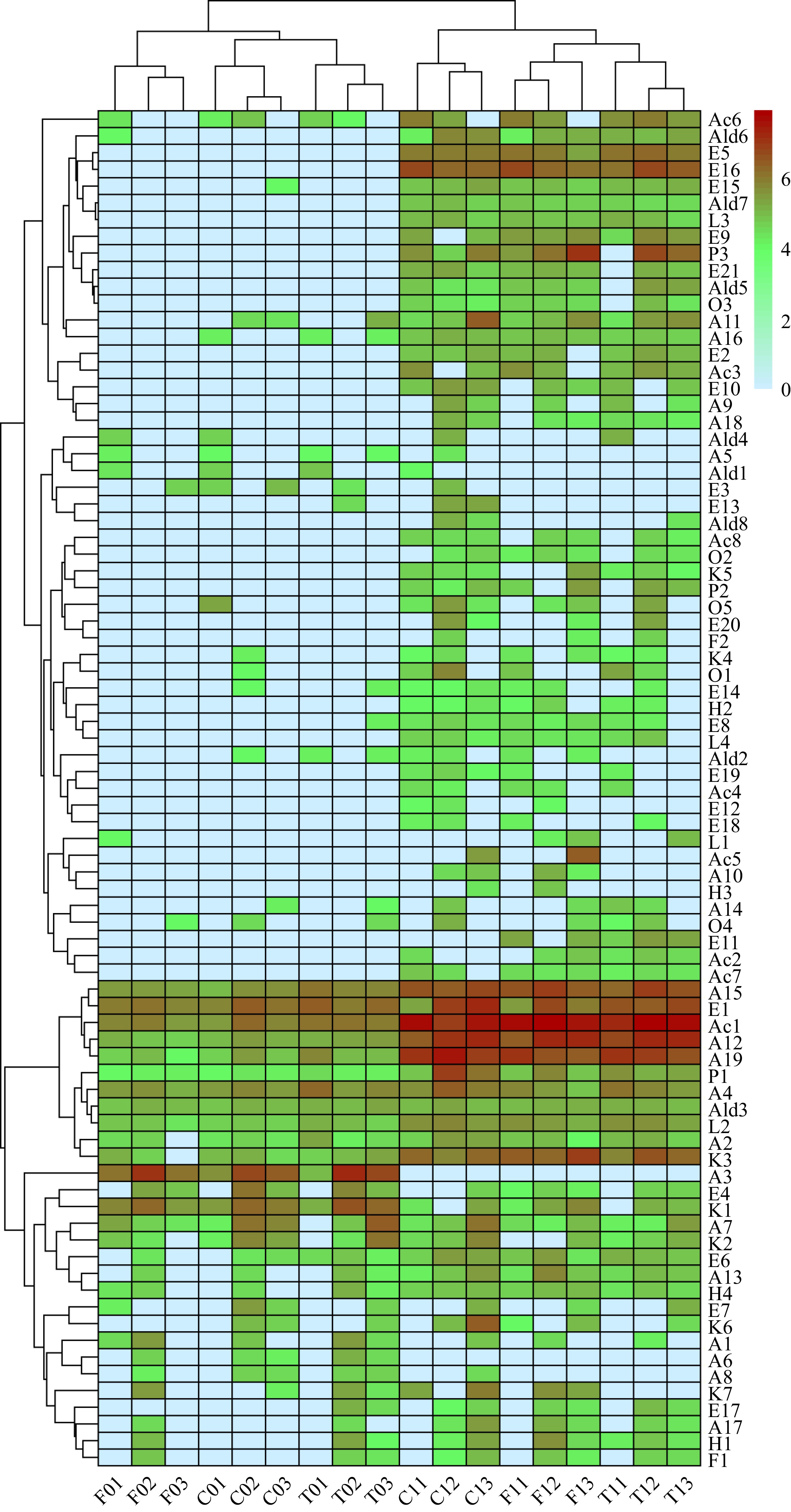
Figure 6.
Volatile compounds cluster heat map. The code in the figure corresponds to the corresponding compound.
The majority of quality traits were positively correlated with volatile components in unfermented beans (Supplemental Fig. S1a), revealing that substance accumulation was conducted with volatile components in synchronization. During the fermentation process, aldehydes inhibit the production of pyrazines[35], a significant negative correlation was determined between the two components (Supplemental Fig. S1b). Comprehensively, the phenotypic value of all tested quality traits was reduced in different magnitudes, contrarily, the content of volatile components was increased significantly, resulting in a negative correlation between quality traits and volatile components in fermented beans (Supplemental Fig. S1b).
-
The fermentation process has a significant impact on the test quality traits of unfermented and fermented Criollo, Forastero and Trinitario beans in Hainan Province, China. The phenotypic value of polyphenols, theobromine, caffeine and cocoa butter content is decreased to different degrees, of which polyphenols are shows to decrease by the largest decrease. For the unfermented beans, the Trinitario group exhibits the highest total content of volatile components, which include alcohols and esters, 2-pentanone, 2-heptanone, 2-heptanol and 1,3-butanedio. The Criollo group contains a higher content of ester components, such as 2-pentyl acetate and 2-heptyl acetate. Fermented beans of the Criollo group are transformed to a flavor profile with high levels of alcohols and esters, including 2-phenylethanol, isoamyl alcohol, ethyl acetate, and high levels of 2-nonanone, 2,3-dimethylpyrazine. Trinitario and Forastero groups contain large amounts of acidic compounds and tetramethylpyrazine, and ketone compounds, 2-methylbutanoic acid and acetoin are also enriched in the Forastero group of fermented beans. Nevertheless, the optimum fermentation time of cacao beans differ significantly, depending on varieties[4,22]. Therefore, in order to produce fermented cocoa beans with good flavor, different fermentation times should be set according to different varieties in production. The present results provide a baseline for a better understanding of the effect of the fermentation process for the three cacao groups cultivated in the Hainan region, principally according to the quality trait identification and profiles of volatile compounds, aiming to improve excellent cacao varieties with fine flavor.
-
The authors confirm contribution to the paper as follows: study conception and design: Yang D, Wu B, Wu G, Li F; data collection: Qin X, Zhao X, Zhu Z; analysis and interpretation of results: Yan L, Zhang F; draft manuscript preparation: Yang D, Wu B. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This research was funded by the Hainan Province's Key Research and Development Project of China (No. ZDYF2021XDNY123) and Central Public-interest Scientific Institution Basal Research Fund (No.1630142022003).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Dewei Yang, Baoduo Wu
- Supplemental Table S1 Agronomic traits and pictures of each material.
- Supplemental Table S2 Quality characteristics of fermented and unfermented beans for all cocoa materials.
- Supplemental Fig. S1 Heat map of quality traits and volatile compound correlations. (a) unfermented beans. (b) fermented beans. The black diagonal separates them. The pie charts in the boxes indicate the strength of the correlations, red is positive, blue is negative, and "×" represents significance (p < 0.05).
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang D, Wu B, Qin X, Zhao X, Zhu Z, et al. 2024. Quality differences and profiling of volatile components between fermented and unfermented cocoa seeds (Theobroma cacao L.) of Criollo, Forastero and Trinitario in China. Beverage Plant Research 4: e010 doi: 10.48130/bpr-0024-0002 

Quality differences and profiling of volatile components between fermented and unfermented cocoa seeds (Theobroma cacao L.) of Criollo, Forastero and Trinitario in China
- Received: 18 October 2023
- Revised: 22 December 2023
- Accepted: 25 December 2023
- Published online: 01 March 2024
Abstract: Fermentation plays a crucial role and contributes to the quality of cocoa beans, the basis of chocolate products. The aim of this study was to evaluate the levels of chemical and aromatic compounds in the cocoa beans of Criollo, Forastero and Trinitario in China. Besides fermentation, unfermented beans of the three genetic groups were also analyzed. The results indicated that genotype involvement played an important role, contributing to cocoa volatile profiling and quality traits. The three cocoa groups' mass indicators all demonstrated some degree of decline after fermentation. After HS-SPME-GC-MS testing, 48 compounds were found in raw beans and 79 compounds in fermented beans. Trinitario contained the most volatile compounds with the highest levels of alcohol, while Criollo contained the highest ester content in unfermented beans. Alcohols and esters were characteristic components of Criollo fermented beans with 37.33% and 14.81%, respectively. The content of acetic acid was as high as 50% in the fermented beans of Forastero and Trinitario and only 35.42% in Criollo. The feature compounds of Criollo fermented beans showed an aromatic profile related to flower flavor, fruit flavor, green flavor, baking flavor and nut flavor. While the characteristic volatile compounds were mainly associated with descriptors of sour, buttery, fruity, roasted and nutty notes for Forastero and Trinitario fermented beans. The results of this study revealed the characteristics of three genetic groups of cocoa beans that will ultimately contribute to the breeding of excellent aroma varieties and cocoa production sustainability in Hainan Province, China.
-
Key words:
- Cocoa /
- Volatile components /
- Quality traits /
- Different cocoa groups