










-
Soil salinization and alkalization represent a major abiotic stress that significantly constrains global plant growth and agricultural productivity. Current estimates indicate that approximately 1.13 billion hectares of agricultural land are affected by salt and alkali stresses, accounting for over 20% of the world's total cultivated area[1]. These stresses degrade soil health by elevating pH, impairing water retention, and reducing nutrient availability, thereby posing a substantial threat to sustainable agriculture. Tea plant, a vital economic crop in China, is particularly susceptible to salt and alkali stress due to its preference for acidic soils. The ongoing expansion of tea cultivation into marginal lands with high salinity and alkalinity has led to significant reductions in yield and quality, resulting in considerable economic losses. Therefore, deciphering the molecular basis of salt and alkali tolerance in tea plant is fundamental for guiding breeding strategies aimed at enhancing stress resistance.
Long non-coding RNAs (lncRNAs) are defined as RNA molecules exceeding 200 nucleotides in length that do not code for proteins. Historically, due to their low expression levels and limited sequence conservation compared with mRNAs, lncRNAs were often dismissed as 'transcriptional noise', and their biological significance was overlooked[2]. However, recent studies have identified numerous functional lncRNAs that play critical regulatory roles in plant growth, development, and stress responses. For instance, in tomato, lncRNA16397 interacts with the SlGRX gene to confer resistance to late blight. Overexpression of lncRNA16397 was shown to alleviate membrane damage and enhance pathogen resistance by upregulating SlGRX expression and reducing ROS accumulation[3]. Similarly, lncRNA33732, activated by the transcription factor WRKY1, enhances resistance by inducing RBOH expression and promoting H2O2 accumulation during the early defense response[4]. LncRNAs can also function by interacting with microRNAs (miRNAs). In resistant tomato varieties, lncRNA39026 sequesters miR168a, leading to the upregulation of its target gene SlAGO1, and enhancing late blight resistance[5]. Beyond disease resistance, lncRNAs like DRIR are involved in abiotic stress responses of Arabidopsis thaliana. DRIR expression is strongly induced by drought and salt stress, and its overexpression enhances tolerance to these stresses[6]. In cotton, lncRNA973 acts as a positive regulator of salt tolerance, as evidenced by increased sensitivity in knockout lines and enhanced tolerance in overexpression lines[7].
Numerous studies have demonstrated that lncRNAs can directly modulate transcription in plants, and this regulatory process occurs primarily within the nucleus. LncRNAs can serve as cofactors, modulating transcription factor activity, or directly binding to DNA to activate gene expression[8]. For example, the Arabidopsis lncRNA ELF18-induced long non-coding RNA 1 (ELENA1) promotes the expression of the pathogen-related gene PR1, enhancing disease resistance[9]. Conversely, the lncRNA SVALKA, which is transcribed antisense to CBF1, represses CBF1 transcription and thereby modulates cold tolerance[10].
Although lncRNA research in tea plant has advanced in recent years, studies have mainly focused on traits such as aroma formation and disease resistance. For instance, 3,556 lncRNAs were associated with aroma formation pathways in black tea[11], evolutionarily conserved OPR trans-lncRNA pairs were implicated in jasmonic acid (JA)-mediated disease resistance[12], and Cslnc170 was shown to positively regulate tea plant resistance to anthracnose by modulating the expression of its potential target gene CsLOX4[13]. Guo et al.[14] identified lncRNA81246, an lncRNA in tea plant, which modulates disease resistance by interfering with the miR164d-CsNAC1 module. Recently, Jiang et al.[15] demonstrated that the lncRNA Cslnc256 enhances tea plant resistance to Colletotrichum camelliae by acting as an endogenous target mimic of CsmiR395, thereby relieving the miRNA-mediated suppression of sulfate metabolism-related genes. However, the roles of lncRNAs in response to salt and alkali stresses remain largely unexplored in tea plant. To date, no lncRNA involved in salt or alkali tolerance has been functionally characterized, representing a significant knowledge gap.
The Jasmonate-ZIM domain (JAZ) proteins are key regulatory factors in the JA signaling pathway, characterized by two conserved domains: ZIM and Jas. The ZIM domain, containing a TIFY motif, mediates interactions with co-repressors like NINJA and TPL, facilitating JAZ degradation and interaction with transcription factors such as MYC2[16,17]. JAZ proteins integrate signals from upstream components (COI1, RGLG3/4, etc.), and regulate downstream targets, including bHLH transcription factors such as MYC2/3/4 and MYB, to coordinate responses to abiotic stresses[16,18]. For example, in cotton, GhJAZs interact with GhMYC2/GhMYC2a to regulate salt tolerance[19].
In tea plant, 13 JAZ genes (CsJAZs) have been identified, distributed across 11 subfamilies[20]. Transcriptional regulatory network prediction and expression pattern analysis revealed that these genes play important roles in growth, development, abiotic stress response, and hormone regulation in tea plant. Promoter analysis revealed an abundance of hormone-responsive elements, particularly for GA, SA, ABA, and JA, in most CsJAZ genes. Consistent with this, hormone treatments induced the expression of multiple CsJAZs; for instance, CsJAZ1, 2, 3, 5, 6, 11, and 12 were upregulated by ETH, while subsets were responsive to GA and ABA[20]. Functionally, alternative splicing of CsJAZ1 negatively regulates flavan-3-ol biosynthesis[21], and under heat stress, CsJAZ6 is activated by CsHSFA1b/2 to interact with CsEGL3 and CsTTG1, reducing catechin accumulation[22]. Nevertheless, the specific functions of CsJAZ proteins under alkali stress are poorly understood.
Our previous transcriptome analysis of tea plant tender leaves under salt and alkali stresses identified CsJAZ2 as a stress-responsive gene that responds to both salt and alkali stress[23]. Coexpression network prediction also revealed a differentially expressed lncRNA, designated lnc87821 based on its gene ID, potentially cis-interacting with CsJAZ2[24]. This lncRNA represents the first candidate for functional characterization of a lncRNA involved in abiotic stress tolerance in tea plant, forming a novel regulatory module essential for salt and alkali stress responses. To investigate the roles of CsJAZ2 and lnc87821 in response to salt and alkali stresses, this study conducted molecular identification, expression profiling, functional validation, and interaction analysis of these two genes. By characterizing the lnc87821-CsJAZ2 module, this study addresses the current gap in knowledge regarding lncRNA-mediated abiotic stress regulation in tea plant. We propose a functional mechanism model for the lnc87821-CsJAZ2 module in responding to salt and alkali stresses, offering new insights for the molecular breeding of stress-resistant tea plant.
-
One-year-old cutting seedlings of the tea plant cultivar 'Shaancha 1' with uniform growth were selected as experimental materials. The seedlings were pre-cultivated hydroponically in a nutrient solution (elemental composition: N 2,850 µmol·L−1, K 1,000 µmol·L−1, Ca 750 µmol·L−1, P 100 µmol·L−1, S 2,130 µmol·L−1, Mg 1,040 µmol·L−1, Al 400 µmol·L−1, Fe 6 µmol·L−1, B 9 µmol·L−1, Mn 18 µmol·L−1, Zn 1.5 µmol·L−1, Cu 0.4 µmol·L−1, Mo 0.5 µmol·L−1, pH 5.5) for one month under controlled environmental conditions (temperature: 25 ± 3 °C; relative humidity: 60%–70%; photoperiod: 12-h light/12-h dark). For stress treatments, the nutrient solution was supplemented with either 200 mmol·L−1 NaCl to impose salt stress or 150 mmol·L−1 NaHCO3 to impose alkali stress. Tender leaf samples were collected at 0, 1, 3, 6, 12, 24, and 48 h post-treatment, then snap-frozen in liquid nitrogen and stored at −80 °C for subsequent nucleic acid extraction. Additionally, leaf, stem, root, and flower tissues were sampled to analyze the specific expressions of genes in different tissues. Three independent biological replicates were maintained for each treatment and time point.
RNA extraction, gene cloning, and sequence alignment
-
Total RNA was extracted from tender leaves using the RNA extraction kit for polysaccharide and polyphenol plant (DP441, Tiangen, Beijing, China) according to the instructions. First-strand cDNA for mRNA analysis was synthesized with the HiScript II 1st Strand cDNA Synthesis Kit (Vazyme, Nanjing, China), while cDNA for lncRNA analysis was generated using the lnRcute lncRNA cDNA First Strand Synthesis Kit (KR202, Tiangen, Beijing, China). Gene-specific primers were designed based on the sequences of CsJAZ2 and lnc87821 (Supplementary File 1). The target sequences were amplified by PCR using the high-fidelity PrimeSTAR DNA Polymerase (R010A, Takara, Dalian, China), with the first-strand cDNA serving as the template. The obtained PCR products were inserted into the pMD19-T vector and then introduced into E. coli DH5α competent cells. Positive clones were selected on LB solid medium supplemented with 100 mg·L−1 ampicillin. Plasmid sequencing was performed by Xi'an Tsingke Biology Co., Ltd. The obtained sequences were aligned and analyzed using DNAMAN 7.0 software, and phylogenetic analysis was conducted using MEGA7 software.
qRT-PCR analysis
-
First-strand cDNA was synthesized from total RNA according to the manufacturer's instructions for the HiScript II Q Select RT Reagent Kit (Vazyme, Nanjing, China). qRT-PCR was subsequently performed using the ChamQ SYBR Color qPCR Master Mix (Vazyme, Nanjing, China) on a QuantStudio®5 Real-Time PCR System (Thermo Fisher Scientific, USA). For the quantification of lncRNA expression, the procedure followed the protocol provided with the lnRcute lncRNA Fluorescence Quantitative Detection Kit (SYBR Green) (FP402, Tiangen, Beijing, China). The Csβ-actin gene and the CsU6 gene were employed as reference genes for cytoplasmic and nuclear RNAs, respectively. Gene-specific primers designed for qPCR are provided in Supplementary File 1. Relative gene expression was determined by the 2−ΔΔCᴛ method according to Livak & Schmittgen[25].
Subcellular localization analysis
-
The subcellular localization of CsJAZ2 was investigated by fusing its coding sequence (CDS) to the N-terminus of GFP in the pCAMBIA2300-GFP vector (35S::GFP) without the stop codon, resulting in the recombinant plasmid 35S::CsJAZ2::GFP. Specific primers used for plasmid construction are provided in Supplementary File 1. A. thaliana (Col-0) protoplasts were isolated from 4-week-old fully expanded rosette leaves. Leaf strips (0.5–1 mm) were digested in an enzyme solution containing 1.5% (w/v) cellulase R10 and 0.4% (w/v) macerozyme R10 in mannitol-based buffer for 3–4 h in the dark, with gentle shaking. The released protoplasts were filtered, washed, and resuspended in MMg solution (0.4 mol·L−1 mannitol, 15 mmol·L−1 MgCl2, 4 mmol·L−1 MES, pH 5.7). For transfection, 20 µg recombinant plasmid was gently mixed with 200 µL of protoplast suspension and 220 µL of PEG-calcium transfection solution (40% PEG 4000, 0.2 mol·L−1 mannitol, 0.1 mol·L−1 CaCl2). Transfected protoplasts were transferred to a 6-well plate and incubated in the dark at 25 °C for 12–18 h. GFP fluorescence was observed using a laser scanning confocal microscope (TCS SP8 SR, Leica, Germany).
The subcellular localization of lnc87821 was determined using a nuclear-cytoplasmic fractionation assay according to Li et al.[26], with all steps performed using RNase-free tips and tubes. Fresh root tips from hydroponically grown tea plant were finely minced and subjected to nuclear and cytoplasmic separation using a commercial extraction kit (BB-361123, Bestbio, Nanjing, China), supplemented with protease inhibitors. Tissue homogenates were filtered through Miracloth (475855, Merck, Germany). RNA was extracted separately from nuclear and cytoplasmic fractions, followed by lncRNA-specific reverse transcription and qRT-PCR analysis to determine the localization of lnc87821.
Identification of transcriptional activation activity of CsJAZ2
-
For transcriptional activation analysis, the coding sequence of CsJAZ2 (stop codon removed) was cloned into the pGBKT7 vector to create a fusion with the DNA-binding domain (CsJAZ2-BD). Following amplification with primers from Supplementary File 1, the fragment was inserted into the vector. The resulting plasmid, alongside empty BD (negative control) and P53-BD + T-AD (positive control) plasmids, was transformed into yeast Y2H Gold cells with a commercial kit (SK2400, Coolab, Beijing, China). Following transformation, yeast cells were plated on SD/-Trp dropout medium and incubated at 30 °C for 3 d. PCR-confirmed positive colonies were then streaked onto SD/-Trp-His-Ade plates supplemented with X-α-gal for functional testing. Plates were incubated under the same conditions, and growth status as well as color development was recorded after 3 d.
Construction of Arabidopsis lines overexpressing CsJAZ2
-
The Arabidopsis lines overexpressing CsJAZ2 were generated via Agrobacterium tumefaciens (strain GV3101)-mediated floral dip transformation. Wild-type (WT) Arabidopsis (Col-0) plants were grown until flowering. Prior to transformation, plants were well-watered to maintain high tissue turgor. An overnight culture of Agrobacterium harboring the 35S::CsJAZ2::GFP construct was inoculated into 20 mL of LB liquid medium supplemented with 50 mg·L−1 kanamycin and 25 mg·L−1 rifampicin, and cultured at 28 °C with constant shaking (180 r·min−1), until the OD600 reached 1.0. The bacterial cells were collected by centrifugation at 4,000 r·min−1 for 3 min, and were resuspended in infiltration medium (1/2 × MS salts, 5% sucrose, 0.05% Silwet-77) to a final OD600 of 0.8. The Arabidopsis inflorescences were subjected to a 10–15 s dip in the bacterial suspension. Post-transformation, plants were kept in darkness and high humidity for 12 h, after which they were moved to a growth chamber under normal light conditions. The floral dip procedure was repeated twice weekly for three weeks. Mature T1 seeds were harvested, air-dried, and surface-sterilized before sowing on 1/2 × MS medium with 50 mg·L-1 kanamycin to select transgenic seedlings. Resistant T1 plants were transferred to soil to set seed, and T2 seeds were subsequently harvested from individual lines. The T2 progeny were screened by germination on 1/2 × MS medium containing 25 mg·L−1 kanamycin. After two weeks, lines showing a segregation ratio of approximately 3:1 (resistant : sensitive) were identified, and kanamycin-resistant (green) seedlings were transplanted to soil. T3 seeds from these plants were screened again on antibiotic medium, and resistant plants were transferred to soil for seed production. To confirm transgene expression, RNA was extracted from leaf tissues of T3 plants and subjected to qRT-PCR analysis. Homozygous overexpression lines showing high CsJAZ2 transcript levels were selected for subsequent experiments.
Detection of hydrogen peroxide (H2O2) and superoxide anion (O2−) content in Arabidopsis leaves
-
The accumulation of H2O2 and O2− in treated Arabidopsis leaves was assessed using both quantitative and histochemical methods. For quantitative analysis, the contents of H2O2 and O2− were measured using commercial assay kits (H2O2: BC3595, Solarbio, Beijing, China; O2−: BC1290, Solarbio, Beijing, China) according to the manufacturers' protocols. Each treatment included four independent biological replicates. For histochemical detection, leaf samples were immersed in either a 1 mg·mL−1 3,3'-diaminobenzidine (DAB) solution to visualize H2O2, or a 0.5 mg·mL−1 nitroblue tetrazolium (NBT) solution to detect O2−. The samples were incubated in the solutions overnight, in the dark. Subsequently, the leaves were decolorized by boiling in 80% ethanol for at least 10 min to remove chlorophyll. The staining intensity, indicative of H2O2 or O2− accumulation, was then observed and qualitatively detected.
Treatments and transcriptome sequencing analysis of A. thaliana
-
WT Arabidopsis and a homozygous transgenic line overexpressing CsJAZ2 (line #10-6) was exposed to 200 mmol·L−1 NaCl (salt stress), or 100 mmol·L−1 NaHCO3 (alkali stress). Leaves from the same positions were collected after 48 h of treatment, then snap-frozen in liquid nitrogen and stored at –80 °C until further analysis. Transcriptome sequencing and the construction of the corresponding cDNA library was conducted by Beijing Berry Genomics Biotechnology Co., Ltd. Raw sequencing data was processed, and differentially expressed genes (DEGs) were analyzed as previously described[23], using the Arabidopsis TAIR10 genome assembly (
www.arabidopsis.org/download/list?dir=Genes%2FTAIR10_genome_release ) as a reference genome. The expression levels of genes were quantified using FPKM values with the StringTie software, differential expression analysis was performed using the DESeq R package (1.16.1), and the resulting p-values were adjusted using Benjamini & Hochberg’s approach for controlling the false discovery rate. Genes with an adjusted value of p < 0.01, and an absolute value of log2 (fold change) > 1 found by DESeq were considered as differentially expressed.Silencing of CsJAZ2 and lnc87821 in tea plant
-
The silencing of CsJAZ2 and lnc87821 in tea plant was performed using antisense oligonucleotides (AsODNs), following the methodology established by Zhang et al.[27]. AsODNs targeting CsJAZ2 and lnc87821 were designed using the online Soligo software (
http://sfold.wadsworth.org/cgi-bin/index.pl ). For the efficiency of silencing, three AsODNs with the binding energy less than –10 kcal·mol−1 and located at relatively distant positions within the sequence were selected from filtered output for antisense oligos for each target, along with corresponding sense oligonucleotides (sODNs) serving as controls (Supplementary File 1). The AsODNs and sODNs were synthesized by Xi'an Tsingke Biotechnology Co., Ltd. Before treatment, the three selected AsODNs (or sODNs for controls) were pooled and diluted to a final concentration of 20 μmol·L−1. Uniformly grown new shoots (one bud with two leaves) of tea plant were excised and immediately immersed in the oligonucleotide solution. Leaf samples were collected at 6, 12, 24, and 48 h after treatment, with four biological replicates per treatment and time point.Coding potential prediction of lnc87821 and GUS verification
-
The coding potential of lnc87821 was evaluated using a combined bioinformatic and experimental approach. Computational analysis was performed with the Coding Potential Calculator (CPC;
http://cpc2.cbi.pku.edu.cn ), where negative prediction scores indicate non-coding potential, and a positive score suggests protein-coding capability. Additionally, prediction of open reading frames (ORFs) within the lnc87821 sequence was performed utilizing the NCBI ORF Finder tool (www.ncbi.nlm.nih.gov/orffinder ).To experimentally validate the coding capacity, a β-glucuronidase (GUS) reporter assay was conducted following the method of Zhang et al.[28]. Specific primers (Supplementary File 1) were designed to clone the predicted ORF region of lnc87821, as well as a mutated version (mORF), into the pCAMBIA1300-GUS vector, generating the constructs 35S::ORF::GUS, and 35S::mORF::GUS. These recombinant plasmids, along with the empty 35S::GUS vector (negative control), were transformed into Agrobacterium GV3101. The bacterial suspensions were infiltrated into leaves of tobacco plants, with three independent plants infiltrated per construct. After 48 h, GUS staining was performed using a commercial kit (SL7160, Coolab, Beijing, China) following the manufacturer's instructions.
DNA extraction, promoter cloning, and analysis
-
Genomic DNA of tea leaves was extracted using the Polysaccharide and Polyphenol Plant DNA Extraction Kit (DP360, Tiangen, Beijing, China) according to the manufacturer's protocol. Specific primers (Supplementary File 1) were designed to amplify an approximately 1,900 bp genomic region upstream of the CsJAZ2 coding sequence, corresponding to its putative promoter. The amplified fragment was confirmed by sequencing and then analyzed for cis-regulatory elements with the PlantCARE online database (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html ). The identified elements were visualized using TBtools software (v7.0).Cluster analysis of gene expression
-
Based on the transcriptome datasets of tea plant under salt and alkali stresses from our previous study[23], gene expression levels were represented by FPKM (Fragments Per Kilobase of transcript per Million mapped reads) values. Expression patterns under each stress condition were independently clustered using TBtools software (v7.0). In the resulting heatmaps, genes with similar expression profiles are grouped together, reflecting coordinated transcriptional responses.
Statistical analysis
-
All data was analyzed by one-way ANOVA. Where significant effects were detected, mean separation was performed using Duncan's multiple range test or Fisher’s protected LSD test (p ≤ 0.05), implemented in SPSS Statistics (version 20). Data in figures are presented as means ± standard deviation (SD).
-
CsJAZ2 and lnc87821 were cloned from the leaves of the tea plant cultivar 'Shaancha 1'. The CDS sequence of CsJAZ2 was 1,092 bp in length, while lnc87821 had a sequence of 947 bp (Supplementary Files 2, 3). Sequence alignment revealed that CsJAZ2 contains TIFY, CCT, and GATA domains. Among these, the TIFY domain is characteristic of JAZ family proteins, whereas the GATA domain is typically associated with GATA-type transcription factors. Phylogenetic analysis of CsJAZ2 with JAZ homologs from Arabidopsis and tea plant indicated that CsJAZ2 is most closely related to CsJAZ5 (Supplementary File 4).
Transcriptome data from our previous study demonstrated that CsJAZ2 expression was upregulated under both salt and alkali stress conditions (Fig. 1a). To further investigate the stress-responsive expression patterns of CsJAZ2 and lnc87821, their transcript levels were analyzed during short-term salt and alkali treatments. Under salt stress, the expression of CsJAZ2 increased significantly, peaking at 24 h at approximately 3.5-fold of the control level. Under alkali stress, CsJAZ2 expression also increased gradually, reaching about 2-fold of the control at 48 h. In contrast, lnc87821 expression was downregulated under both stress conditions (Fig. 1b). These results indicate that CsJAZ2 is induced by salt and alkali stress, whereas lnc87821 is suppressed. Furthermore, the expression of lnc87821 and CsJAZ2 exhibited a significant negative correlation under both salt stress (r = –0.8287) and alkali stress (r = –0.7901), suggesting a potential negative regulatory relationship between them.
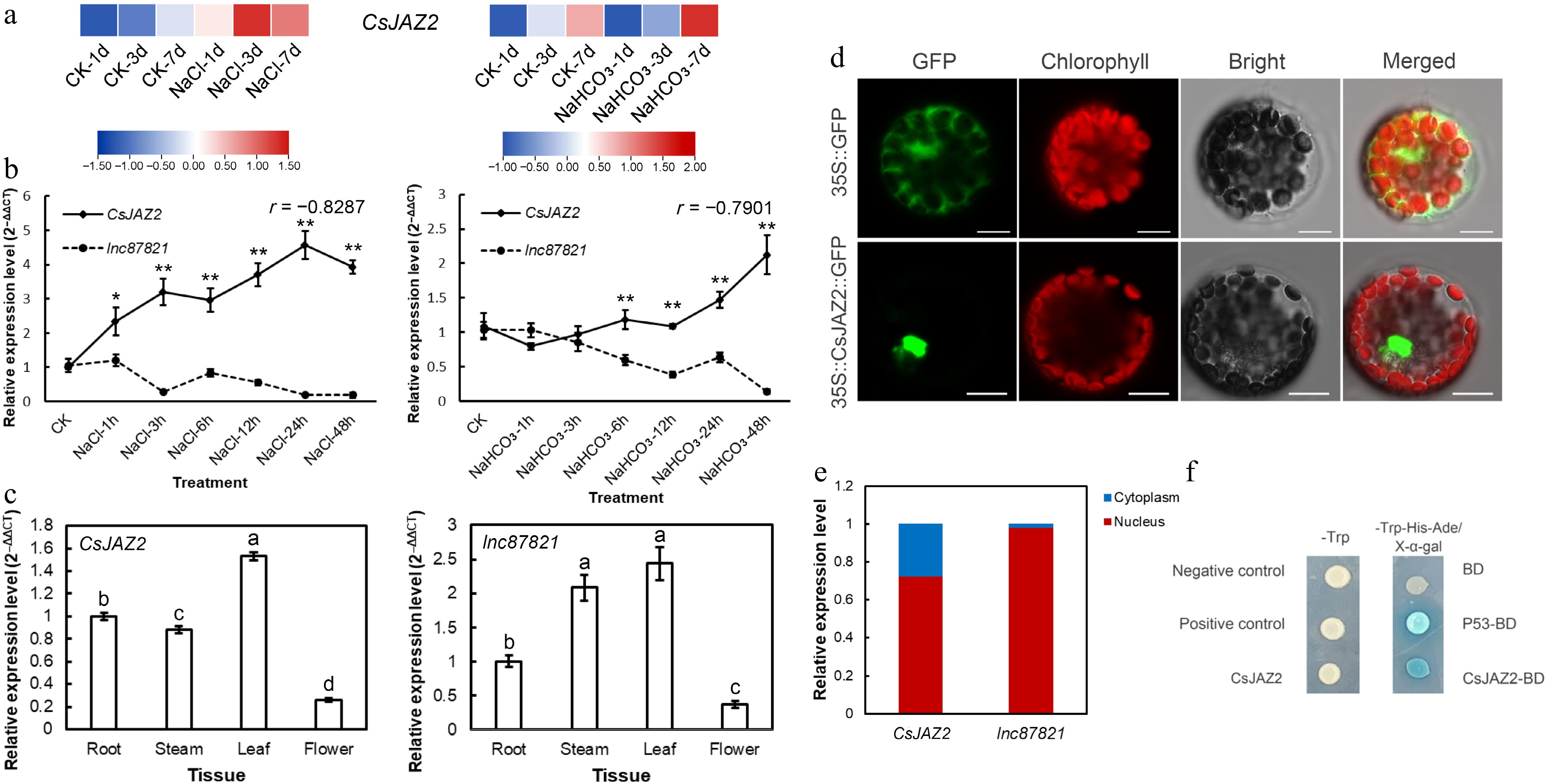
Figure 1.
Expression analysis and subcellular localization of CsJAZ2 and lnc87821. (a) Expression levels of CsJAZ2 under salt and alkali stress. (b) Expression of lnc87821 and CsJAZ2 were significantly negatively correlated under salt and alkali stress. Data is the mean ± SD. * Indicates significant difference between salt and alkali stress, * p ≤ 0.05, ** p ≤ 0.01. (c) Expression pattern analysis of CsJAZ2 and lnc87821 in different tissues of tea plant. Different letters indicate significant difference between groups, p ≤ 0.05. (d) Subcellular localization reveals the nuclear localization of CsJAZ2, scale bar = 10 μm. (e) Nuclear-cytoplasmic fractionation reveals the nuclear localization of lnc87821. (f) Yeast one-hybrid assay verified the transcriptional activation activity of CsJAZ2.
To examine the tissue-specific expression profiles of CsJAZ2 and lnc87821, their expression levels were determined in leaves, stems, roots, and flowers. Both genes showed the highest expression in leaves and the lowest in flowers (Fig. 1c).
Both CsJAZ2 and lnc87821 are located in the nucleus
-
The subcellular localization of CsJAZ2 was assessed by transiently expressing a 35S::CsJAZ2::GFP fusion construct in Arabidopsis protoplasts. The resulting GFP fluorescence was observed with laser scanning confocal microscopy. As shown in Fig. 1d, the green fluorescence of CsJAZ2-GFP was predominantly observed in the nucleus, distinct from the red auto-fluorescence of chloroplasts, indicating nuclear localization of CsJAZ2.
Since lncRNAs are non-coding, the localization of lnc87821 was assessed by nuclear-cytoplasmic fractionation followed by qRT-PCR. As shown in Fig. 1e, lnc87821 was predominantly detected in the nuclear fraction, accounting for 98.08% of its total expression across nuclear and cytoplasmic compartments. By contrast, gene CsJAZ2 showed a distinct distribution pattern. These results indicate that lnc87821 is primarily localized in the nucleus, suggesting its potential function in nuclear regulatory processes.
The CsJAZ2 protein has transcriptional activation activity
-
To investigate whether CsJAZ2 functions as a transcriptional activator, a yeast one-hybrid assay was performed using the Y2H Gold strain. The empty BD vector was used as the negative control, while the P53-BD + T-AD pair was used as the positive control. On a SD/–Trp–His–Ade medium supplemented with X-α-gal, yeast cells transformed with the empty BD vector failed to grow and showed no blue coloration. In contrast, both the positive control and yeast expressing the CsJAZ2-BD fusion grew normally and developed a distinct blue color (Fig. 1f), demonstrating that CsJAZ2 possesses transcriptional activation activity.
Overexpression of CsJAZ2 enhances salt and alkali stress tolerance in A. thaliana
-
To investigate the functional role of CsJAZ2 in salt and alkali stress responses, two independent Arabidopsis overexpression lines (CsJAZ2-OE#1-7, and #10-6) were generated. To assess the physiological impact of CsJAZ2 overexpression, soil-grown transgenic and WT seedlings were irrigated with 100 mmol·L−1 NaHCO3 or 200 mmol·L−1 NaCl for 7 d. Under alkali stress, both CsJAZ2-OE lines displayed better growth than WT, which showed pronounced growth inhibition (Fig. 2a). Although no visible phenotypic differences were observed under salt stress, histochemical staining and quantitative assays revealed lower accumulation of H2O2 and O2− in the leaves of overexpression lines compared with WT under salt stress, and reduced O2− under alkali stress (Fig. 2b, c, f, g). These results suggest that CsJAZ2 mitigates ROS accumulation under both salt and alkali stress, thereby enhancing the antioxidative capacity of Arabidopsis.
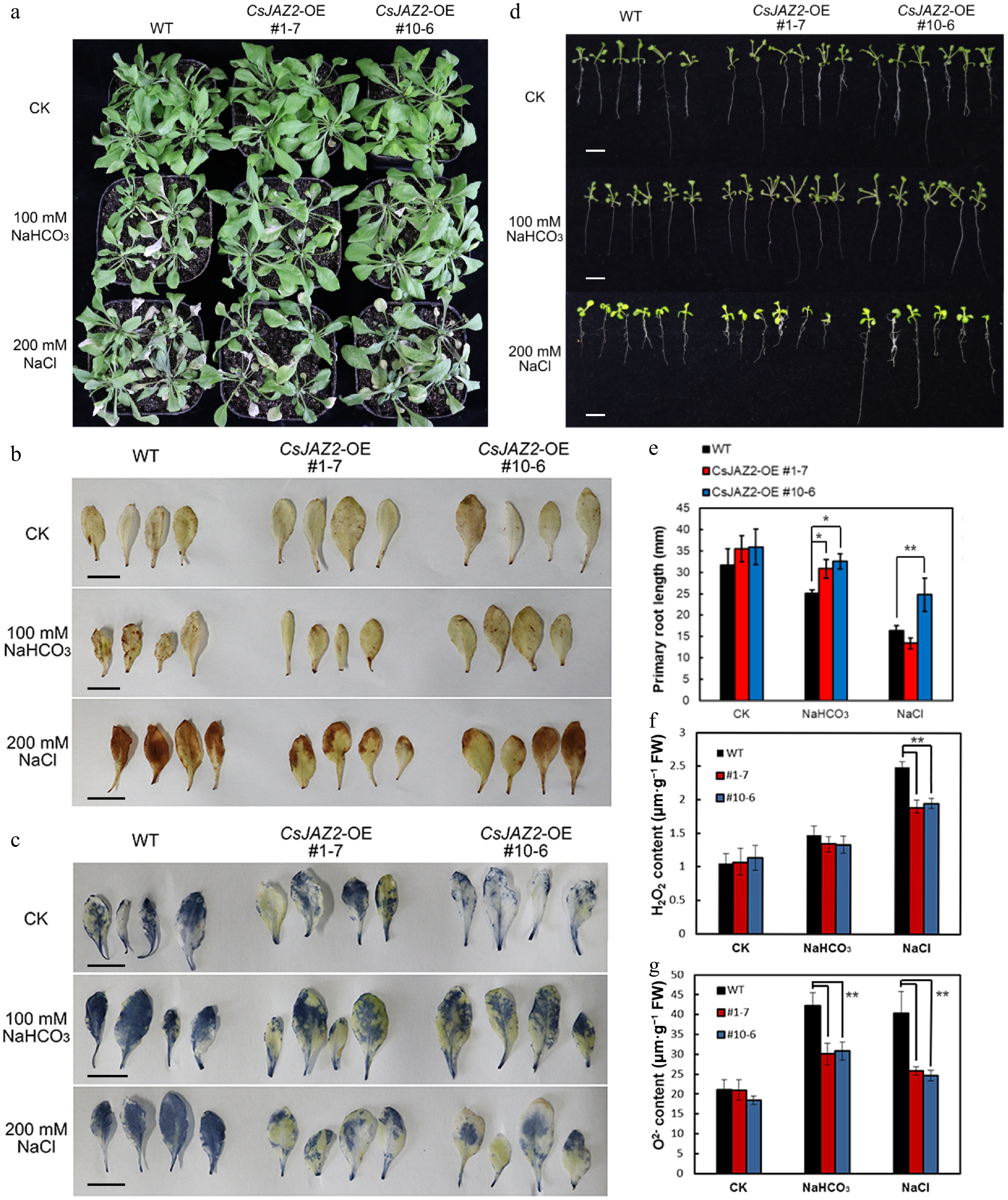
Figure 2.
Analysis of the resistance of CsJAZ2-OE Arabidopsis under salt and alkali stresses. (a) The growth condition of CsJAZ2-OE Arabidopsis under salt and alkali stresses. (b) DAB staining of H2O2 in leaves of CsJAZ2-OE Arabidopsis under salt and alkali stresses, scale bar = 1 cm. (c) NBT staining of O2− in leaves of CsJAZ2-OE Arabidopsis under salt and alkali stresses, scale bar = 1 cm. (d) The root growth of CsJAZ2-OE Arabidopsis under salt and alkali stresses, scale bar = 1 cm. (e) Statistics of root length. (f) Statistics of H2O2 content in leaves. (g) Statistics of O2− content in leaves. Data is the mean ± SD, * represents significant difference (p ≤ 0.05), ** represents extremely significant difference (p ≤ 0.01).
In addition, germination of WT and transgenic lines was assessed separately on 1/2 MS medium supplemented with either 100 mmol·L−1 NaHCO3 or 200 mmol·L−1 NaCl. Root growth was evaluated after 10 d. Under alkali stress, both overexpression lines exhibited longer roots compared with WT (Fig. 2d, e). Under salt stress, line #10-6 showed a significant increase in root length relative to the control, indicating that CsJAZ2 enhances root tolerance to both salt and alkali stresses.
Transcriptome analysis of CsJAZ2-overexpressing A. thaliana
-
To elucidate the molecular mechanism by which CsJAZ2 confers salt and alkali stress tolerance in Arabidopsis, transcriptome sequencing was performed on WT and CsJAZ2-OE#10-6 plants under control (CK), salt stress (NaCl), and alkali stress (NaHCO3) conditions. Differential gene expression analysis identified 768, 1,640, and 1,051 DEGs under CK, NaCl, and NaHCO3 treatments, respectively (Fig. 3a). Venn diagram analysis revealed 46 common DEGs across all three treatments, and 245 DEGs shared between salt and alkali stress conditions (Fig. 3b).
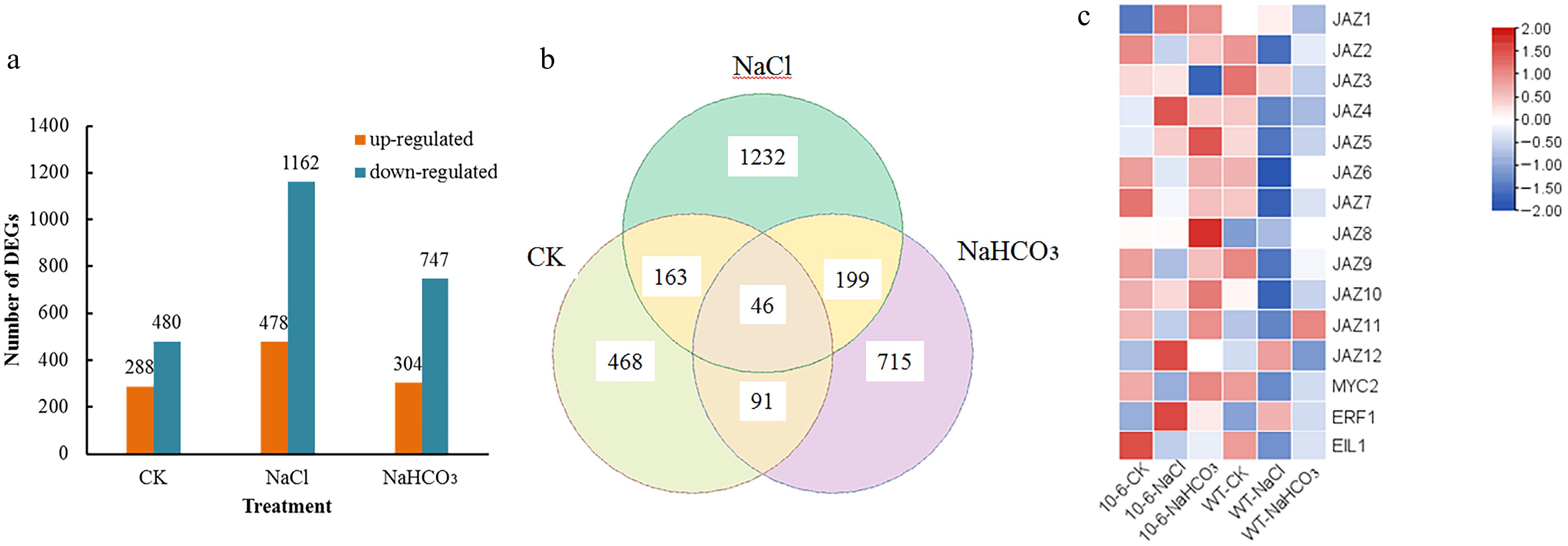
Figure 3.
Transcriptome analysis of CsJAZ2-OE Arabidopsis. (a) Statistics of up- and downregulated DEGs under different treatments. (b) Venn diagram of DEGs under different treatments. (c) Expression heatmap of Arabidopsis JAZ and its downstream genes.
Expression analysis of the JAZ family and downstream regulatory genes showed that their transcript levels were generally higher in CsJAZ2-overexpressing plants than in WT under both salt and alkali stress (Fig. 3c), suggesting that CsJAZ2 may enhance stress tolerance through coordinated regulation of related genes in the JAZ signaling network.
Furthermore, among the stress-responsive DEGs, we identified numerous genes involved in ROS scavenging, such as POD, GST, MDHAR and SOD. The expression patterns of their homologs in tea plant under corresponding stress conditions were also analyzed (Supplementary File 5). Notably, under salt stress, several antioxidant genes such as TEA007569 (POD) and TEA010026 (GRX) were upregulated in CsJAZ2-OE plants but downregulated in tea plant. Conversely, TEA015341 (GST) was downregulated in tea plant but upregulated in transgenic Arabidopsis. A similar regulatory pattern was observed under alkali stress, where two tea plant genes TEA006224 (POD) and TEA011287 (GST) were upregulated in OE plants, while TEA015341 (GST) and four homologs were downregulated. These results indicate that CsJAZ2 modulates the expression of antioxidant-related genes in Arabidopsis, potentially enhancing ROS scavenging capacity and contributing to improved salt and alkali stress tolerance.
AsODN-mediated silencing of CsJAZ2 in tea shoots
-
To functionally characterize CsJAZ2 in tea plant, we performed antisense oligonucleotide (AsODN)-mediated gene silencing in tea shoots. As shown in Fig. 4a, the expression level of CsJAZ2 was significantly reduced at 6 h after AsODN treatment compared with the sense oligonucleotide (sODN) control. Although transcript levels gradually recovered from 12 to 48 h, they remained significantly lower than those in the control, confirming effective suppression of CsJAZ2. Consistent with this, expression of three downstream genes CsMYC2, CsERF1, and CsEIL1 were also downregulated, indicating that silencing CsJAZ2 affects its regulatory network.
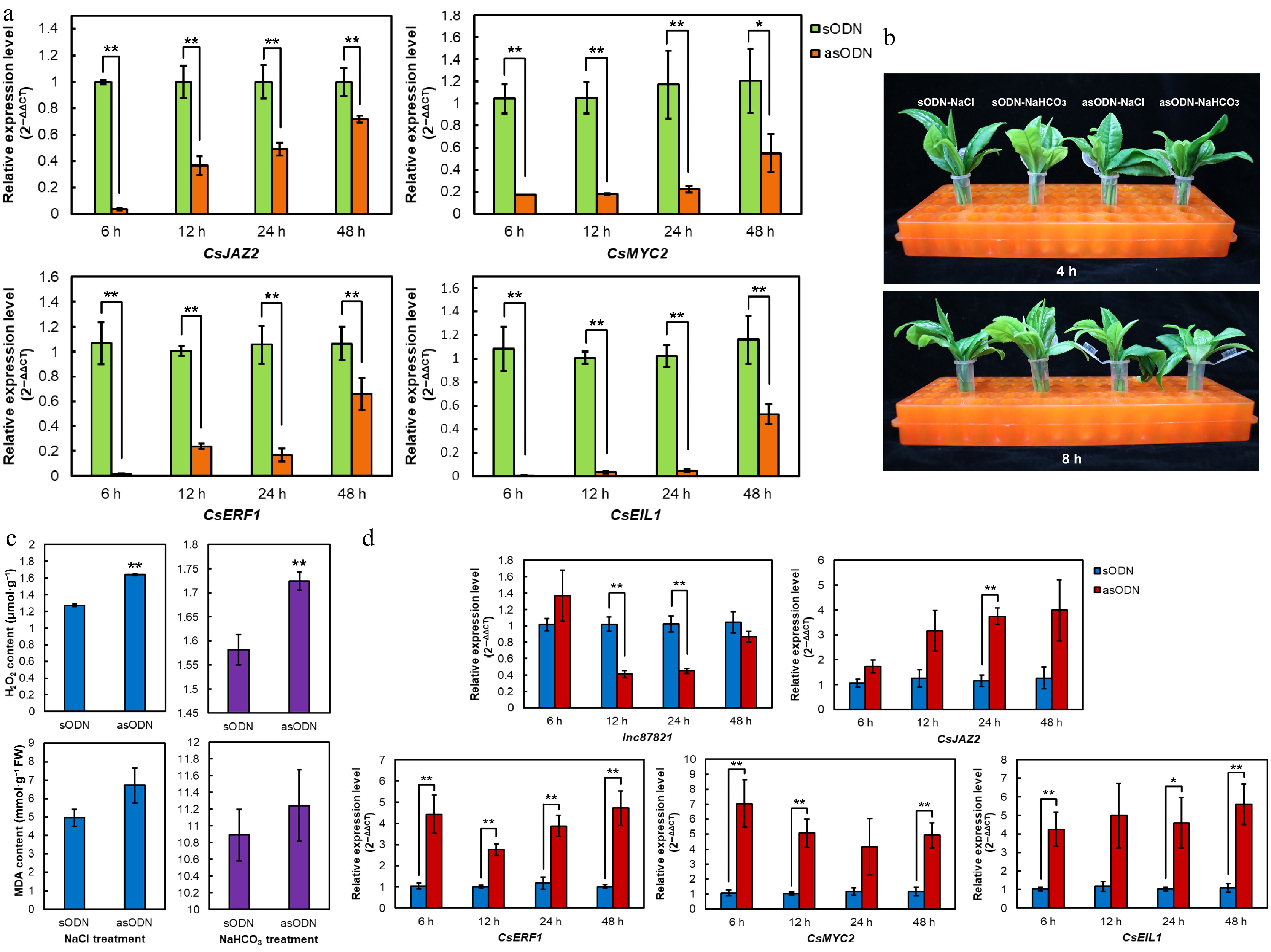
Figure 4.
AsODN-based suppression of CsJAZ2 and lnc87821 in tea shoots. (a) Effects of CsJAZ2 silencing on the expressions of downstream genes. (b) 6 h-silencing of CsJAZ2 in tea shoots treated with salt and alkali stresses. (c) Determination of H2O2 and MDA contents after salt and alkali treatments. (d) Effects of AsODN inhibition of lnc87821 on expressions of CsJAZ2 and its downstream genes. Data is the mean ± SD, * represents significant difference (p ≤ 0.05), ** represents extremely significant difference (p ≤ 0.01).
Based on the observed peak silencing efficiency of CsJAZ2 at 6 h post-AsODN treatment, the silenced tea shoots were subsequently exposed to salt (NaCl) and alkali (NaHCO3) stress for 8 h. Physiological analysis revealed that AsODN-treated samples displayed marginally enhanced leaf wilting under both stress conditions compared with the control (Fig. 4b). Quantitative assays showed significantly higher H2O2 accumulation in silenced shoots, whereas MDA content remained statistically unchanged (Fig. 4c). These results indicate that CsJAZ2 functions as a negative regulator of H2O2 accumulation and contributes to enhanced stress tolerance in tea plant under salt and alkali stress.
Lnc87821 negatively regulates CsJAZ2 expression in tea shoots
-
To further investigate the regulatory relationship between lnc87821 and CsJAZ2, we performed AsODN-mediated silencing of lnc87821 in tea shoots. As shown in Fig. 4d, compared with the control, expression levels of lnc87821 were significantly reduced at 12 and 24 h after AsODN treatment, confirming effective suppressions. Concurrently, expression of CsJAZ2 was upregulated at 24 h, and its downstream targets CsMYC2, CsERF1, and CsEIL1 also showed consistent upregulation throughout the treatment period. These results demonstrate that lnc87821 acts as a negative regulator of CsJAZ2 and its downstream signaling pathway.
Coding potential prediction and GUS verification of lnc87821
-
Recent studies suggest that some lncRNAs may harbor short open reading frames (sORFs) with peptide-coding potential[8]. To evaluate whether lnc87821 encodes a functional peptide, we analyzed its coding potential using the Coding Potential Calculator (CPC), and identified putative ORFs through the NCBI ORF Finder. The coding gene CsJAZ2 was included as a positive control. The CPC analysis yielded a coding potential score of –1.13943 for lnc87821, supporting its classification as a non-coding RNA. Nevertheless, a short ORF potentially encoding 42 amino acids was identified within its sequence (Fig. 5a).
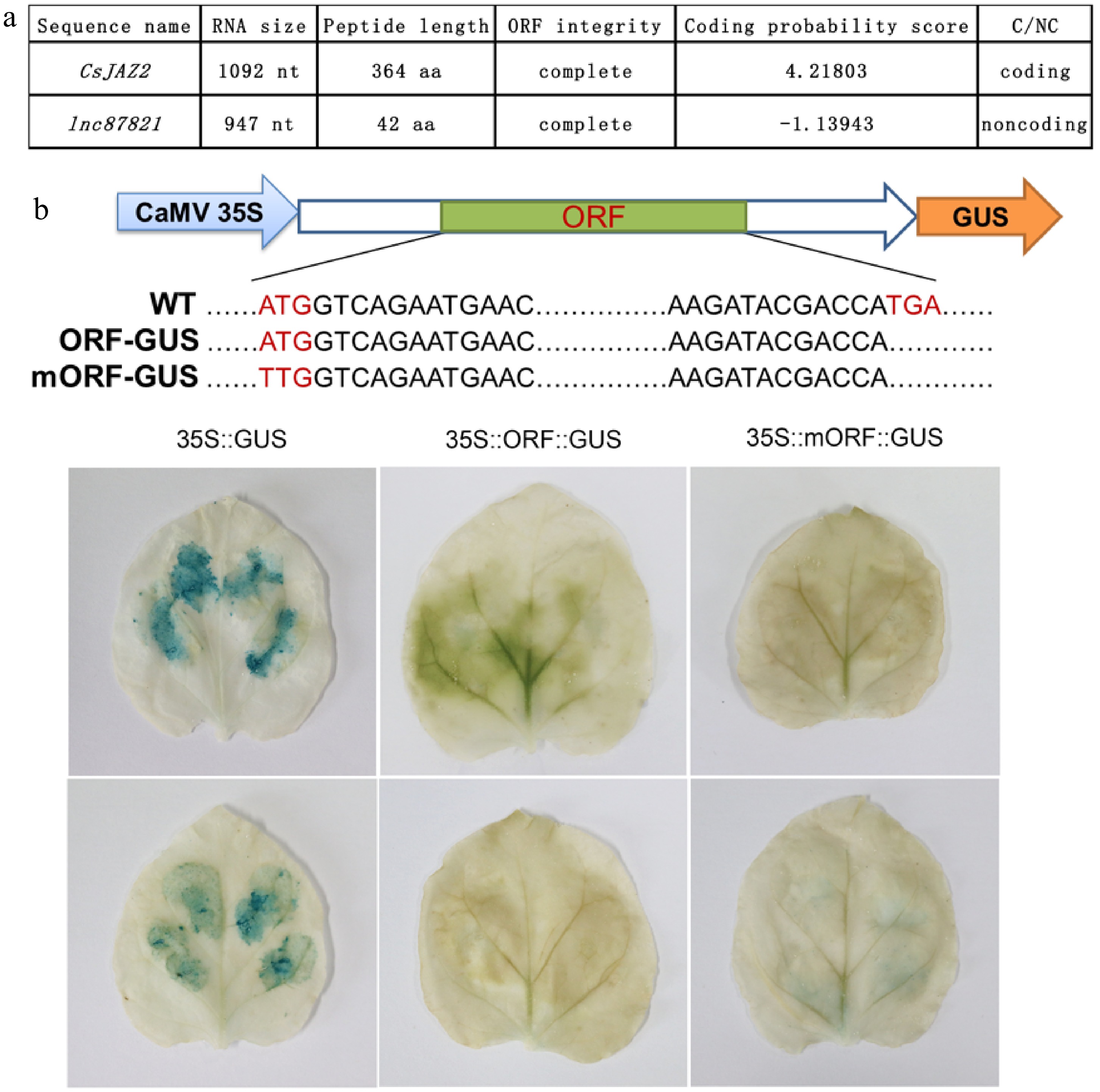
Figure 5.
Prediction and GUS verification of coding potential of lnc87821. (a) Coding potential prediction of lnc87821. (b) GUS verification of coding capability of lnc87821.
To experimentally test whether this sORF is translated, the putative ORF and a mutated version with a disrupted start codon (mORF) were cloned into a GUS reporter vector under the control of the 35S promoter. As shown in Fig. 5b, strong GUS activity was observed in tobacco leaves expressing the positive control (35S::GUS), whereas no detectable signal was observed in leaves expressing 35S::ORF::GUS or 35S::mORF::GUS. These results confirm that lnc87821 does not encode a translated peptide, and likely functions primarily as a non-coding RNA.
Prediction of the regulatory mechanism of lnc87821 on CsJAZ2 under salt stress and alkali stress
-
To investigate the upstream regulatory mechanism of CsJAZ2, a 1,900 bp promoter region upstream of the coding sequence was cloned and sequenced (Supplementary File 2). Cis-regulatory element analysis was performed using the PlantCARE database. As shown in Fig. 6a and Supplementary File 6, the promoter sequence contains multiple putative regulatory elements, including light-responsive elements, hormone-responsive motifs, and binding sites for MYB and MYC transcription factors. These findings suggest that the expression of CsJAZ2 is potentially regulated by light conditions, hormonal signals, and MYB/MYC transcription factors.
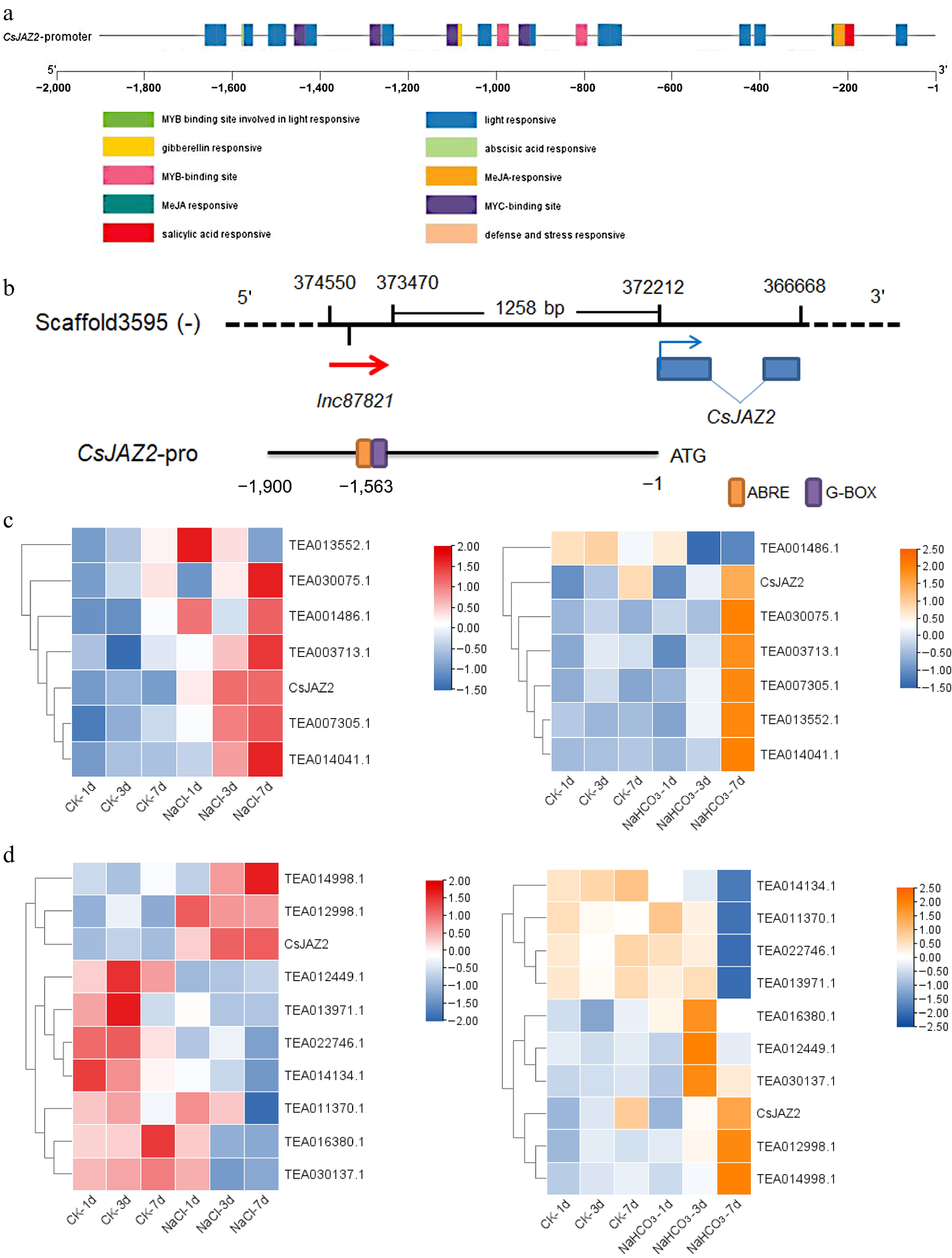
Figure 6.
Prediction of the regulatory mechanism of lnc87821 on CsJAZ2 under salt and alkali stresses. (a) Analysis of cis-acting elements of CsJAZ2 promoter. (b) Position relation of lnc87821, CsJAZ2, and its promoter. (c) Expression clustering analysis of differentially expressed bZIPs and CsJAZ2 under salt and alkali stresses. (d) Expression clustering analysis of differentially expressed bHLHs and CsJAZ2 under salt and alkali stresses.
Existing evidence suggests that lncRNAs can modulate the expression of adjacent genes by interfering with transcription factor binding to promoter regions[8]. Analysis of the CsJAZ2 promoter revealed that the transcriptional region of lnc87821 overlaps with ABRE and G-box cis-acting elements (Fig. 6b), indicating a potential role for lnc87821 in regulating the activity of these motifs. Given that ABRE and G-box elements are known binding sites for bZIP and bHLH transcription factors, respectively[29,30], we further examined the coexpression patterns of CsJAZ2 with bZIP and bHLH family genes in tea plant transcriptomes under salt and alkali stress. Cluster analysis identified several transcription factors, including bZIP transcription factor TEA007305.1, TEA003713.1, TEA014041.1, and bHLH transcription factor TEA012998.1 and TEA014998.1, that exhibit strong expression correlation with CsJAZ2 (Fig. 6c, d), suggesting that these factors may be involved in the lnc87821-CsJAZ2 regulatory pathway under salt and alkali stresses.
-
The JAZ protein family plays a critical role in the JA signaling pathway and is widely involved in plant stress responses, growth, and development[31]. In tea plant, 13 JAZ family genes have been identified, and they are known to participate in abiotic stress responses, hormone interactions, and developmental processes[20]. Although previous studies have demonstrated that JAZ genes regulate flavonoid biosynthesis and metabolism in tea plant[21,22], their specific roles in abiotic stress tolerance remained poorly understood. In this study, we observed that the expression of CsJAZ2 was induced under both salt and alkali stress conditions. To further investigate its function, we cloned and analyzed CsJAZ2, revealing that it contains a conserved GATA domain, classifying it as a member of the GATA transcription factor family. Transcriptional activation assays in yeast confirmed that CsJAZ2 possesses transcriptional activation activity. Moreover, subcellular localization analysis determined the nuclear localization of CsJAZ2, consistent with its identity as a transcription factor.
Tea plant response to salt and alkali stresses involves complex regulatory mechanisms. Previous studies from our group have demonstrated that these stresses adversely affect tea plant growth, photosynthetic performance, and induce oxidative stress in leaves[23], ultimately compromising tea quality and yield. Evidence from other species indicates that JAZ family members can modulate stress tolerance; for instance, OsJAZ9 functions as a transcription factor enhancing salt resistance in rice[32], while GsJAZ2 improves alkali stress tolerance in soybean[33]. The functional roles of CsJAZ2 to salt and alkali stress tolerance was assessed using both gain-of-function (heterologous expression in A. thaliana), and loss-of-function (AsODN-mediated silencing in tea plant) strategies. Transgenic Arabidopsis lines overexpressing CsJAZ2 exhibited enhanced tolerance to both salt and alkali stress, accompanied by improved ROS scavenging capacity. Transcriptome analysis revealed upregulation of antioxidant-related genes in these transgenic lines under salt and alkali stresses. Conversely, silencing of CsJAZ2 in tea shoots led to downregulation of its downstream targets, increased ROS accumulation and enhanced membrane lipid peroxidation. Collectively, these results demonstrate that CsJAZ2 enhances salt and alkali stress tolerance by modulating downstream antioxidant pathways, thereby alleviating oxidative damage in tea plant.
LncRNAs are increasingly recognized as key regulators in diverse plant physiological processes through multiple molecular mechanisms[34,35]. A well-documented mode of lncRNA action involves the cis-regulation of genomically adjacent genes. For instance, in tomato, lncRNA33732 enhances resistance to late blight by activating the expression of the neighboring RBOH gene, thereby promoting ROS scavenging[4]. Similarly, in peach, lncRNA5 upregulates its adjacent gene PpTCP18 to suppress branch formation[36]. Additional mechanisms include coexpression patterns between lncRNAs and nearby genes[3], as well as promoter-derived lncRNAs that modulate the transcription of downstream genes by forming triplex structures with their promoters[37]. Despite these advances, the functional roles of lncRNAs in tea plant remain largely unexplored, with no prior reports experimentally validating lncRNA-mediated regulation of downstream targets. In this study, we identified a stress-responsive lncRNA, lnc87821, located 1,258 bp upstream of CsJAZ2 and partially overlapping its promoter region, through comparative analysis of salt stress-induced lncRNAs and the tea plant reference genome. This genomic arrangement suggested a potential cis-regulatory role for lnc87821 in negatively modulating CsJAZ2 expression, thereby influencing salt and alkali stress responses. Consistent with this hypothesis, expression analysis under salt and alkali stress revealed a significant negative correlation between lnc87821 and CsJAZ2 transcript levels, further supporting the proposed regulatory relationship.
As the majority of lncRNAs lack protein-coding capacity, they typically exert regulatory functions at the transcriptional level through nucleic acid-based mechanisms, predominantly within the nucleus. In this study, nuclear-cytoplasmic fractionation coupled with qRT-PCR confirmed the predominant nuclear localization of lnc87821. Both lnc87821 and CsJAZ2 exhibited the highest expression levels in tea leaves, suggesting their potential functional significance in this tissue. Despite emerging evidence that certain lncRNAs can encode functional small polypeptides, computational assessment using the CPC and ORF predictions indicated that although lnc87821 contains a small ORF, it lacks coding potential. This conclusion was further substantiated by GUS reporter assays, which demonstrated no detectable translation activity from the predicted ORF. Collectively, these results confirm that lnc87821 functions primarily as a non-coding RNA.
Loss-of-function approaches are widely employed to characterize the biological roles of lncRNAs in plants. For example, Li et al.[26] demonstrated that CRISPR/Cas9-mediated knockout of lncRNA1459 in tomato regulates fruit ripening, while Zhang et al.[28] used virus-induced gene silencing (VIGS) to suppress lncRNA354 in cotton, revealing its involvement in salt stress response via the miR160b-ARF module. In tea plant, AsODN-mediated silencing has become an established method for functional genetic studies. Applying this approach, we successfully suppressed lnc87821 expression and observed consequent upregulation of CsJAZ2 and its downstream targets, confirming lnc87821 as a negative regulator of CsJAZ2.
The negative regulatory relationship between lnc87821 and CsJAZ2 bears interesting comparisons with other plant lncRNA systems. In tomato, lncRNA16397 enhances pathogen resistance by upregulating its target SlGRX[3], representing a positive regulatory mode distinct from the negative regulation observed for lnc87821. Similarly, lncRNA33732 activates its neighboring RBOH gene[4], whereas our findings reveal lnc87821 as a repressor of CsJAZ2. This contrast highlights the diversity of lncRNA-mediated regulatory mechanisms in plants, with some lncRNAs acting as transcriptional activators and others as repressors. The negative regulation exerted by lnc87821 is more reminiscent of lncRNA SVALKA in Arabidopsis, which represses CBF1 transcription during cold stress[10], suggesting that repressive lncRNAs may represent a common regulatory strategy in abiotic stress responses.
Promoter analysis revealed that the transcriptional region of lnc87821 overlaps with two key cis-acting elements in the CsJAZ2 promoter: ABRE and G-box, which are known binding sites for bZIP and bHLH transcription factors, respectively[28,29]. This genomic arrangement suggests a potential mechanism by which lnc87821 might interfere with transcription factor access to these elements. Coexpression analysis further identified several transcription factors (CsbZIP43, CsbZIP13, CsbHLH137, and CsbHLH117) exhibiting expression patterns closely correlated with CsJAZ2 under salt and alkali stresses, making them candidate regulators of CsJAZ2. We hypothesize that lnc87821 transcription through this region may physically occlude transcription factor binding to the ABRE and G-box elements, a mechanism analogous to transcriptional interference reported in other eukaryotic systems. Alternatively, the lnc87821 transcript itself might compete with these transcription factors for promoter occupancy. To test these possibilities, future experiments should include a dual-luciferase reporter assay to determine whether lnc87821 directly binds to the CsJAZ2 promoter, and chromatin immunoprecipitation (ChIP) assays to examine whether lnc87821 overexpression or silencing alters the recruitment of bZIP/bHLH transcription factors to the ABRE/G-box elements. These follow-up studies would provide direct mechanistic evidence for the proposed regulatory model.
Promoter analysis revealed that the transcriptional region of lnc87821 overlaps with two key cis-acting elements in the CsJAZ2 promoter: ABRE and G-box, which are known binding sites for bZIP and bHLH transcription factors, respectively[29,30]. Coexpression analysis further identified several transcription factors (CsbZIP43, CsbZIP13, CsbHLH137, and CsbHLH117) exhibiting expression patterns closely correlated with CsJAZ2 under salt and alkali stresses. These findings suggest that lnc87821 may modulate CsJAZ2 expression by interfering with the binding of bZIP/bHLH transcription factors to its promoter, though this proposed mechanism requires further experimental validation.
Based on the experimental validation of our hypothesis, a negative regulatory model for lnc87821-CsJAZ2 module is proposed, elucidating its role in tea plant tolerance to salt and alkali stress. This model provides mechanistic insight into a previously uncharacterized lncRNA-transcription factor interaction in tea plant, and expands our understanding of lncRNA diversity in stress responses. The hypothetical mechanism is delineated as follows: under non-stress conditions, elevated expression of lnc87821 likely interferes with the binding of bZIP/bHLH transcription factors to the CsJAZ2 promoter, thereby suppressing CsJAZ2 transcription; under salt and alkali stresses, the downregulation of lnc87821 alleviates this repression, permitting bZIP/bHLH factors to activate CsJAZ2 expression. The subsequent increase in CsJAZ2 levels enhances the transcription of downstream target genes, reinforcing ROS scavenging capacity and ultimately improving tea plant tolerance to salt and alkali stresses. This model provides mechanistic insight into how a stress-responsive lncRNA interfaces with the JA signaling pathway, uncovering a previously unknown regulatory mechanism for the antioxidant defenses under abiotic stress in a perennial woody crop.
-
In this study, CsJAZ2 and its genomically adjacent lncRNA, lnc87821, both implicated in tea plant responses to salt and alkali stress, were successfully cloned. lnc87821 was found to be downregulated under salt and alkali stresses, and was identified as a negative regulator of CsJAZ2. The tolerance to salt and alkali stress was enhanced by CsJAZ2 through the improvement of ROS-scavenging capacity, which was achieved by the modulation of downstream target genes and antioxidant-related components (Fig. 7). Furthermore, it is speculated that the expression of CsJAZ2 may be regulated by lnc87821 through transcriptional interference with the binding of upstream transcription factors to the cis-elements in its promoter region. The lnc87821-CsJAZ2 module offers potential applications in tea plant breeding: CsJAZ2 serves as a candidate marker for marker-assisted selection, while targeted downregulation of lnc87821 could strategically enhance stress tolerance. These findings provide novel insights for molecular breeding aimed at enhancing salt and alkali tolerance in tea plant. However, the precise regulatory mechanism of the lnc87821-CsJAZ2 module and the functional roles of its downstream targets remain to be fully elucidated. It should also be noted that a transient AsODN-mediated suppression approach was utilized for gene silencing in this study, as stable transgenic tea plant lines were not obtained. The evidence for the proposed lncRNA-mRNA interaction could be strengthened by future validation using stable genetic transformation systems. Future validation using stable transformation systems, combined with CRISPR/Cas9-mediated genome editing, will be essential for fully elucidating lncRNA-mRNA regulatory networks and advancing the precision breeding of stress-resistant tea cultivars.
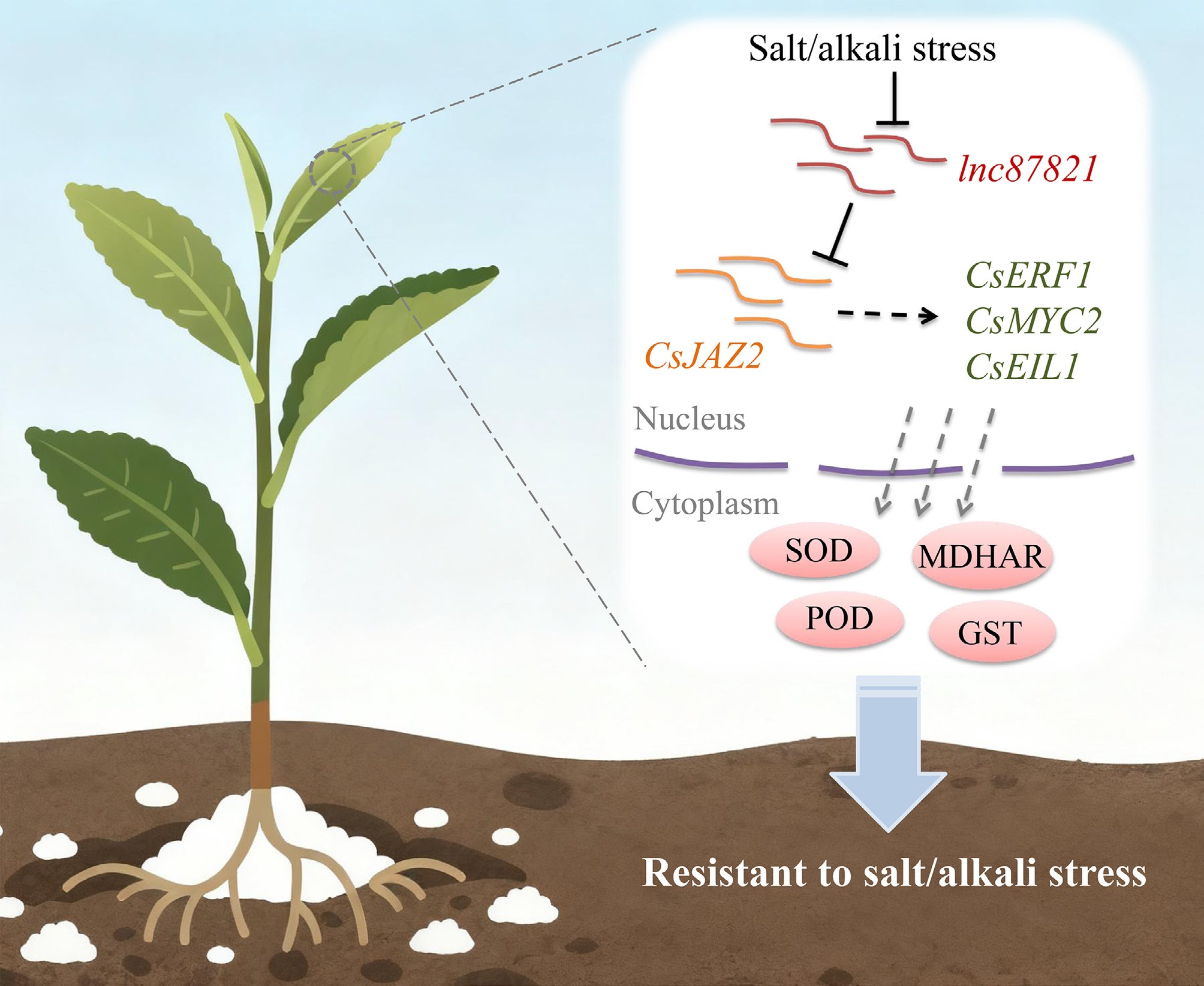
Figure 7.
A predicted model for lnc87821-CsJAZ2 participating in the salt and alkali stress responses in tea plant.
-
The authors confirm their contributions to the paper as follows: study conception and design: Yu Y, Wang W, Zan L, Wan S; experiments performed: Wan S, Zhang Y, Xiao Y; data analysis: Wan S, Zhang Y, Liu L; draft manuscript preparation: Wan S, Yan F, Xu Y. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author upon reasonable request.
-
This work was supported by Shaanxi University of Technology Doctoral Talent Launch Project (SLGRCQD031), the 'Co-construction of City and School' scientific research project of State Key Laboratory of Qinba Biological Resources and Ecological Environment (SXC-2112), the Key R&D Program Project of Shaanxi Province (2024QY2-GJHX-18), the earmarked fund for Modern Agro-industry Technology Research System (CARS-19) and Shaanxi Provincial Department of Education Special Scientific Research Program for Local Service (25JC033).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/bpr-0026-0008.
- Supplementary File 1 Primer sequences used in the experiments.
- Supplementary File 2 Sequences of genes and promoters.
- Supplementary File 3 Cloning of CsJAZ2 and lnc87821.
- Supplementary File 4 Domain and phylogenetic tree analysis of CsJAZ2 protein.
- Supplementary File 5 Differentially expressed genes related to active oxygen scavenging under salt and alkali stresses.
- Supplementary File 6 Cis-acting elements of CsJAZ2 promoter.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wan S, Zhang Y, Xiao Y, Liu L, Wang W, et al. 2026. Long non-coding RNA lnc87821 negatively regulates CsJAZ2 to modulate salt and alkali stress responses in Camellia sinensis. Beverage Plant Research 6: e023 doi: 10.48130/bpr-0026-0008 

Long non-coding RNA lnc87821 negatively regulates CsJAZ2 to modulate salt and alkali stress responses in Camellia sinensis
- Received: 13 January 2026
- Revised: 26 February 2026
- Accepted: 03 March 2026
- Published online: 25 June 2026
Abstract: Tea plant (Camellia sinensis [L.] O. Kuntze), a major economic crop in China, is an acidophilic species with stringent requirements for its growth environment. Soil salinization is increasingly threatening the sustainable cultivation of tea plant. However, the lncRNA-mediated regulatory mechanisms underlying salt and alkali stress tolerance remain largely elusive. Based on our preliminary research, a long non-coding RNA (lncRNA) in tea plant, lnc87821, was predicted to potentially cis-regulate its downstream target gene CsJAZ2, and respond to salt stress. Therefore, this study aimed to experimentally validate the lnc87821-CsJAZ2 regulatory relationship. By integrating gene cloning, expression analysis, coding potential prediction, GUS reporter assays, and gene silencing in tea shoots, we elucidated the negative regulatory role of lnc87821 on CsJAZ2, and confirmed its functional relevance under salt and alkali stresses. Under these conditions, the expression of lnc87821 and CsJAZ2 exhibited significant negative correlations. Silencing lnc87821 in tea shoots significantly upregulated the expression of CsJAZ2 and downstream antioxidant-related genes. Furthermore, CsJAZ2 enhanced the scavenging capacity of reactive oxygen species (ROS) under salt and alkali stresses. Furthermore, bioinformatic promoter analysis of CsJAZ2 suggested that lnc87821 might modulate CsJAZ2 transcription by interfering with the binding of transcription factors to the G-box and ABRE cis-acting elements. Our results demonstrate that lnc87821 functions as a negative regulator in tea plant tolerance to salt and alkali stresses by modulating CsJAZ2. This study provides novel insights into the stress adaptation mechanisms of tea plant and identifies candidate genes for molecular breeding for stress resistance.
-
Key words:
- Camellia sinensis /
- Salt stress /
- Alkali stress /
- lnc87821 /
- CsJAZ2