








-
While Nepal has achieved strides in national development, rural areas still face challenges in achieving sustainable agriculture and food security, particularly for women farmers. National statistics show progress in areas like income and poverty reduction[1−3], however, a significant portion of the rural population (28%) grapples with multidimensional poverty[3]. Slow growth in the agriculture industry and the rural economy are important factors that contribute to rural poverty. Farmers in Nepal have been engaged in farming, rearing cattle, and other small-scale productive endeavors for hundreds of years. Farming and livestock-keeping used to be integrated as well as keeping trees on the farmland or in the backyard as a source of fodder, wood for fuel, and litter compost. Such an agroforestry system are standard historical practice in many communities in Nepal[4]. These practices align with the principles of circular agriculture, which emphasizes minimizing waste and maximizing resource use within closed-loop systems. For the vast majority of economically engaged smallholders and the poorest households, it serves as their primary activity. The majority of the population relies on agriculture (70% of rural livelihood) as their primary source of income, accounting for two-thirds of the workforce, however, contributing only 21.3% of Nepal's GDP[1,5].
The demands on agricultural production in Nepal have increased due to the country's rapid population growth and urbanization. Even though Nepal's agricultural output has increased over the past ten years, it is still below its potential when compared to that of its neighbors. Small farms, challenging terrain, limited access to farmer-friendly and affordable agricultural technologies, lack of seeds and fertilizers, loss of habit which is the traditional method of storing seed stock, lack of all-weather road connectivity between urban and rural areas, heavy reliance on seasonal rainfall, constrained agricultural markets, and subsistence farming have long been cited as the main obstacles keeping the nation in a state of low agricultural productivity. The small amount of private sector involvement in the agriculture industry is another contributing factor. Climatic change, labor migration, and changes in marketing patterns (increase import) are additional factors that hinder agriculture production[6].
Increasing labor migration significantly decreases agricultural activities and production in the country[7,8]. The labor migration that mostly involves men pursuing work in foreign countries have only increased the burden on women to perform additional domestic caretaker tasks, exacerbating already substantial workloads[9]. There are important factors that lead to a decline in agricultural production, particularly among women farmers. The lack of access to information on market prices and trends can make it difficult for farmers to make informed decisions about what crops to grow and when to sell them, while a lack of access to markets can limit their opportunities to sell their products and earn a fair price[10,11]. This can lead to a lack of motivation to produce more, particularly if it is difficult to earn a sustainable income from farming. Additionally, the challenges of agroforestry, including the need for long-term planning and investment, can make it a less attractive option for farmers who are struggling to make ends meet. All of these factors can contribute to a decline in agricultural production, particularly among women farmers who may face additional barriers such as limited access to information, resources, and decision-making power[5,12].
Decline in the production of agricultural goods means that there is a reduction in the amount of food and other agricultural products that are being produced. When there is a high demand for food and a decline in agricultural production, it can lead to a decrease in the availability of food for people to consume[13]. This, in turn, can lead to the need to spend a high amount of a country’s economy to import basic foodstuffs. Another serious consequence is food insecurity, which is a condition where people do not have access to enough food to meet their basic nutritional needs. Food insecurity can have serious consequences for individuals, households, and communities. It can lead to malnutrition, which can have negative effects on physical and cognitive development and can increase the risk of illness and disease[13−16]. Although overall agriculture production is increasing at the national level, trends over the last decade show that many parts of Nepal still face chronic to minimal food insecurity[17].
Food insecurity can also lead to economic hardship, as people may have to spend a larger proportion of their income on food or may have to forego other basic needs to purchase food. Food insecurity is a multidimensional problem that not only impacts health and nutrition but also has economic and social consequences[14,18]. Decline in agriculture production and food insecurity can also have negative social and political implications. It can lead to social unrest, political instability, and even conflict, as people compete for scarce resources[19]. Therefore, it is important to ensure that agriculture production is sustainable and resilient and that efforts are made to address the underlying causes of the decline in production to prevent food insecurity. Sustainable agriculture is essential for addressing the problem of food insecurity and ensuring food security for future generations[20,21]. Furthermore, the decline in agricultural production, lack of market for products, and limited access to information on market trends and demands has resulted in farmers losing interest in agriculture in general[22]. To that end, the aim of the study is to examine farming practices, assess the marketability of agricultural products, and propose interventions for promoting sustainable agriculture and enhancing market access for smallholder farmers in Devdaha Municipality. Specifically, this study aims to: (1) Analyze the current farming practices and challenges faced by farmers in the region. (2) Evaluate the marketability of agricultural products, prioritizing high-potential, profitable, and marketable crops. (3) Recommend interventions and strategies to enhance sustainable agriculture, improve market access, and reduce dependency on imported food items.
-
This study was carried out in Devdaha Municipality, which is located in Rupandehi District, Lumbini Province of Nepal. The Municipality has a total of 12 wards (Fig. 1), which are distributed across 136.95 km2 of geographical area and extended from the Terai flatland (101 m) to the Churia range (1,126 m). The Municipality is intersected by rain-fed streams such as Rohini, Bhaluhi, and Ghodaha. According to the 2021 census conducted by the Central Bureau of Statistics (CBS), Devdaha Municipality has 17,300 households with a total population of 72,457 that include 34,033 (47%) males and 38,424 (53%) females. Population aged above 10 years are 60,545, of that 32,856 are economically active and 15,096 (45.9%) of those economically active, are engaged in agriculture and forestry activities. Out of that 14,658 are skilled and elementary workers in the agriculture and forestry occupation. The male population engaged in agriculture and forestry (41.04%) is much less than the female population (58.96%). The census data indicates the agriculture and forestry sector holds significant potential for growth and development in Devdaha Municipality. About 27.22% of Devdah municipality is occupied by agricultural land (Fig. 1), but agriculture occupies the least part of the economy.
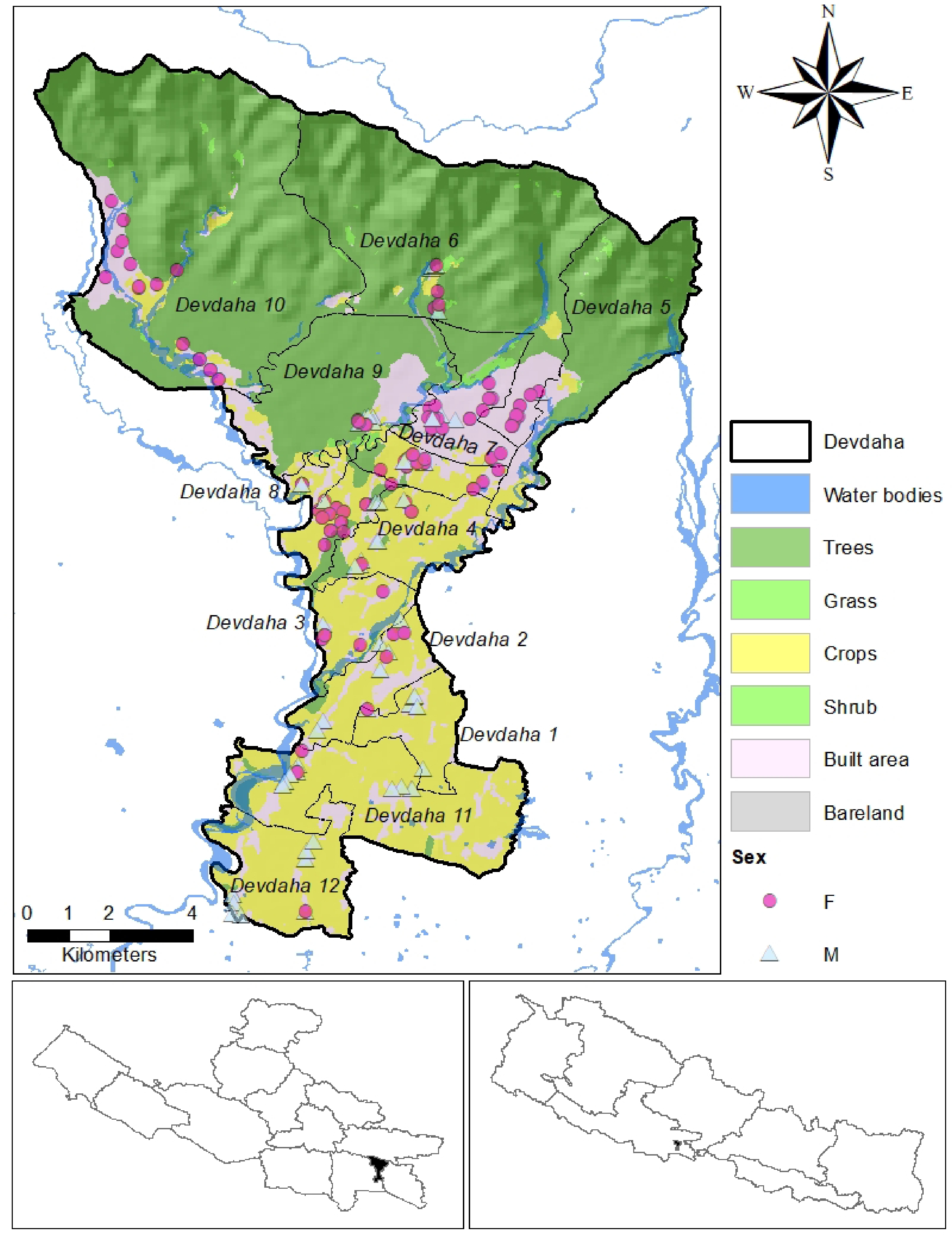
Figure 1.
Map of the study area showing different landuse.
Women have a high degree of involvement in farming activities, ranging from cropland preparation to harvesting products, livestock raising, and post-harvesting activities. The high involvement of women in the agricultural sector in the Devdaha Municipality is also clear from the 2021 population census data[23]. Their significant contributions cannot be overlooked in terms of access to resources and responses to climate change in farming activities. Moreover, the agroforestry sector in the Devdaha Municipality has immense potential for women's involvement in agriculture, but due to various social and cultural factors, women have limited access to information and resources, leading to their exclusion from the sector. Rupandehi district of Lumbini Province also falls in one of the districts with food insecure conditions. Food insecurity among women and children is more severe[24].
Data collection and treatment
-
The study employed several methods to collect the required data to answer the fundamental questions posed above. The Nepal government divides the rural agricultural population into three categories: small commercial farmers, subsistence farmers, and landless/near landless farmers. The bulk of farmers (53%) are landless/near landless farmers with less than 0.50 hectares of land[25]. They collectively account for only 19% of all accessible land. Over 27% of farmers practice 'subsistence farming' with land holdings of 0.5−1 hectare, accounting for 28% of the total usable area.
To ensure the representativeness of the findings, the purposive sampling technique was employed[26] to select a diverse group of farmers who were willing to participate in the study. Purposive sampling was employed to select a diverse group of participants, including subsistence farmers, landless/near landless farmers, and those engaged in identified agroforestry practices with the assistance of local farmer leaders. To minimize data collection bias and ensure consistency, all data collectors received training on standardized data collection protocols. Furthermore, ethical research practices were prioritized. All participating farmers provided informed consent before data collection began. The consent form clearly explained the study's purpose, data collection procedures, and participants' rights. The study protocol was reviewed and approved by the School of Development Studies and Applied Sciences (SDSAS), Lumbini Buddhist University to ensure adherence to ethical research principles.
Data collection instruments
-
The questionnaire was prepared using the ODK (Open Data Kit) tool, which is an open-source platform for data collection[27]. The questionnaire was designed to collect information on the farmers' socio-economic characteristics, their farming practices, major farm products, and their access to markets. The questionnaire was administered in the local language to ensure that all respondents could understand and respond appropriately. To ensure the validity and reliability of the questionnaire, a pilot test was conducted with a small group of farmers before the main data collection phase. Based on the pilot test results, the questionnaire was refined to improve clarity and guarantee it captured the intended information. A back-translation process was employed to ensure their accuracy and cultural relevance. The data collection period spanned January to March 2023, which aligned with the project's short-term duration.
In addition to the interview, field visits were conducted to observe existing farming practices, agroforestry systems, and potential natural resources within the municipality. These observations focused on identifying crops, trees, and other resources with the potential for market development and integration into sustainable agricultural practices. Two distinct data collection methods were employed to gather insights from various stakeholders in Devdaha Municipality.
1) Expert consultation: Organized within the premises of the SDSAS, the consultation involved agriculture and agroforestry experts, small entrepreneurs, and local government officials. This approach allowed us to gain valuable perspectives from a diverse range of stakeholders with expertise relevant to the research.
2) Focus group discussion: Conducted in a rural area of the municipality, it involved a group of approximately 15 individuals, primarily women farmers and community members from women's groups and youth clubs. These discussions aimed to collect valuable insights into the challenges faced by farmers, potential opportunities based on available resources, and local perspectives on enhancing sustainable agriculture in the region. Supplementary interviews were conducted with key stakeholders to gather in-depth information and further explore specific themes identified during other data collection activities.
Data analysis and visualization
-
Data from the focus group discussions, including expert consultation and interviews were analyzed thematically. This involved a manual process of identifying recurring themes in the responses. The responses and transcripts were carefully reviewed, highlighting and taking notes on major points and repetitive ideas expressed by participants. These highlighted sections were then examined to identify broader themes that captured the key issues and concerns raised during the discussions and interviews.
Data obtained through field surveys, interviews with farmers and experts are supplemented with literature reviews and existing agroforestry databases, such as The Agroforestree Database (
www.worldagroforestry.org/output/agroforestree-database ). The most suitable and profitable agroforestry systems, including intercropping between fruits and vegetables currently planted in the municipality, were visually expressed.A grading system was developed based on the suitability of different fruits and vegetables for intercropping to prioritize and visualize possible agroforestry systems. The goal of this rating system was to let farmers know which combinations were most likely to succeed. The scoring process involved assessing the suitability of intercropping between different fruits and vegetables. Each combination received a score based on its potential (1), current practices (2), currently practiced and/or very good combination (3), and excellent agroforestry (4).
These scores were then used to construct a Sankey diagram using Flourish visualization tools (
https://app.flourish.studio ). The benefits of using this scoring system and the resulting Sankey diagram were twofold. First, it provided a quantitative measure of compatibility, helping us identify the most favorable intercropping combinations. Second, the Sankey diagram visually represented these combinations, making it easier for farmers and stakeholders to understand and implement agroforestry practices. The thickness of the lines in the Sankey diagram indicated the potential for successful intercropping, with thicker lines signifying higher suitability. -
Altogether 200 survey responses (125 female, 75 male) were used out of 209 total responses collected from 12 wards of the Devdaha municipality to understand local agriculture practices, market access patterns, and opportunities for improvement in Devdaha Municipality. The analysis revealed a diverse range of 57 farm-based products (Table 1). Field observations corroborated this diversity, showcasing various cropping patterns and traditional agroforestry practices, though to a lesser extent. However, thematic analysis of group discussion, expert consultation, and interviews identified key challenges limiting profitability and hindering growth for farmers. These challenges revolved around:
Table 1. List of farm based products from the Devdaha Municipality.
No. Category Name 1 Allium Garlic 2 Onion 3 Cruciferous Broccoli 4 Cabbage 5 Cauliflower 6 Cucurbit Ash gourd 7 Balsam gourd 8 Bitter gourd 9 Bitter gourd 10 Chayote 11 Cucumber 12 Ivy gourd 13 Kakari 14 Pointed gourd 15 Pumpkin 16 Ridge gourd 17 Snake gourd 18 Sponge gourd 19 Squash 20 Fruit Banana 21 Grapes 22 Mango 23 Orange 24 Papaya 25 Watermelon 26 Fruit vegetables Eggplant 27 Capsicum 28 Drumsticks 29 Okra 30 Tomato 31 Tree tomato 32 Leafy Green Amaranths 33 Broad leaf mustard 34 Coriander leaf 35 Cress 36 Fennel leaf 37 Fenugreek leaf 38 Lettuce 39 Spinach 40 Swiss chard 41 Legume Broad beans 42 Cowpea 43 French beans 44 French beans - sword type 45 French beans - bush type 46 French beans - pole type 47 Peas 48 Root Carrot 49 Radish 50 Turnip 51 Spices Chili pepper 52 Chili (round) 53 Stem Asparagus 54 Tuber Colocasia 55 Elephant foot yam 56 Potato 57 Yam Seasonal farming patterns
-
Survey data (Fig. 2a) and discussions revealed peak farming activity in January-February and April-May. Interestingly, survey respondents reported lower vegetable production throughout the year compared to their stated inputs. Notably, vegetable production declines significantly during the monsoon season, likely due to farmers prioritizing staple crop (cereal) planting, as indicated by their responses showing decreased engagement during this period (Fig. 2b). This pattern aligns with higher farm output for male farmers compared to female farmers during monsoon months (one month earlier to one month later). Thematic analysis of discussions revealed gender disparities in farming practices. Notably, both genders lacked access to technologies like raised nursery beds, potentially limiting vegetable production during the monsoon season for female farmers. Additionally, market access and connections emerged as a significant factor.
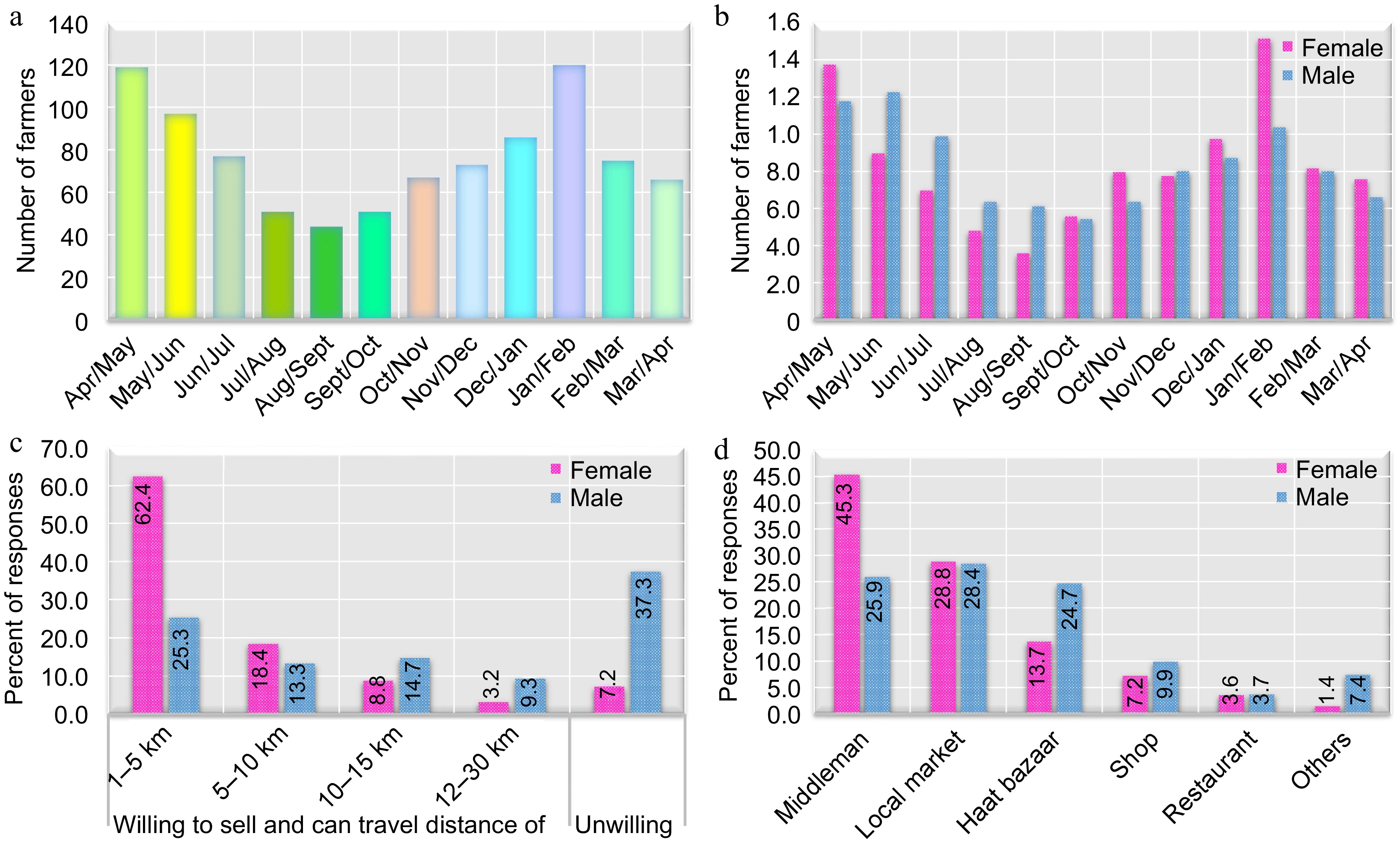
Figure 2.
(a) Season of farm products in Devdaha Municipality; (b) gender differences in season of farm products; (c) gender differences in willingness to travel to market for selling farm products; (d) gender differences in places of selling farm products.
Market access and challenges
-
Limited reach, competition from imports, and reliance on middlemen were common themes. Observations during field visits confirmed limited market infrastructure, particularly for farmers located further from central areas.
Local farmers primarily prefer selling their produce close to home, with nearly half willing to travel up to 5 km and another 40% venturing between 5−10 km (Fig. 2c). Survey data (Fig. 2c) indicated a higher tendency for female farmers to sell locally, likely due to balancing farm work with household responsibilities. Conversely, the discussion suggested some male farmers may have access to larger markets or agents, as evidenced by their higher willingness to travel longer distances to sell produce (Fig. 2c).
While most market-driven produce is vegetables, a significant portion is for self-consumption. Farmers typically store grains for a year, selling the surplus. In terms of selling channels, middlemen are the most common, followed by local markets (neighborhood markets accessible daily). Notably, a significant number of farmers utilize traditional agricultural markets called haat bazaars, while some sell directly to shops, restaurants, and other buyers. Figure 2d illustrates that female farmers primarily rely on middlemen and local markets, with fewer utilizing haat bazaars and shops, while male farmers predominantly sell at haat bazaars alongside local markets and middlemen. However, discussions highlighted several obstacles, particularly for women.
• Limited market knowledge: Women farmers reported lacking proper knowledge of alternative markets beyond middlemen, restricting their options.
• Unfair pricing by middlemen: Discussions revealed concerns about the dominance of unmanaged middlemen who often lacked proper registration and offered lower prices, especially to women with limited market connections.
The present analysis based on the expert consultation identified weak supply chains as another significant challenge. Farmers lack adequate storage facilities, transportation options, and opportunities for value addition. Additionally, vendors report inconsistent product supply from farmers, hindering their ability to meet market demand effectively. Furthermore, variations in quality, and appearance across different producers make it difficult to establish a consistent brand and market presence.
Many farmers shared their experiences and opinions on why most people currently do not engage in vegetable commercial production. Competition from cheaper imports further discourages local vegetable production. Farmers perceive locally produced vegetables to be more expensive, forcing them to become consumers themselves rather than commercial producers. To address these challenges, a comprehensive inventory of farm and agroforestry products was conducted.
Inventory and prioritization
-
To identify marketing opportunities, we conducted a comprehensive inventory of farm and agroforestry products currently produced in Devdaha Municipality (Table 1). This included 57 farm-based products, ranging from fruits and vegetables to spices and legumes. The present survey revealed that few agroforestry practices are currently implemented in the traditional way. However, farmers using these methods reported a wider variety of products than vegetable farmers. Additionally, based on expert consultation, interviews, and literature review three promising agroforestry products with potential for future development were identified.
This inventory, along with discussions with farmers and experts, led to the categorization of products based on marketability and potential profitability:
• High: fruits, vegetables, spices, and potential agroforestry products;
• Moderate: leafy greens, cruciferous, cucurbits, stems, allium, and legumes;
• Low: root and tubers.
Based on this assessment, the following products were prioritized for further marketing efforts:
• Fruits such as mango, banana, papaya, grapes, watermelon and citrus fruits;
- Fruit vegetables such as tomato, eggplant, pumpkin, and okra;
• Spices such as chili, ginger, and turmeric.
Potential agroforestry products listed at this stage such as curry leaf (Murraya koenigii), miracle tree (Moringa oleafera) and medicinal plants.
Potential agroforestry solutions
-
Focus group discussions highlighted the potential of adopting improved agroforestry systems to diversify and increase farm products. This diversification could empower farmers to leverage opportunities with local government in establishing well-managed local farmers' markets closer to farms, enhancing market access and income potential.
To address identified challenges like the need for diversified and productive harvests, a suitability assessment analyzed potential agroforestry combinations. This assessment culminated in a Sankey diagram (Fig. 3), visually representing the suitability of different intercropping combinations between fruits and vegetables. The thickness of lines indicates the level of suitability, with thicker lines signifying a higher potential for success.
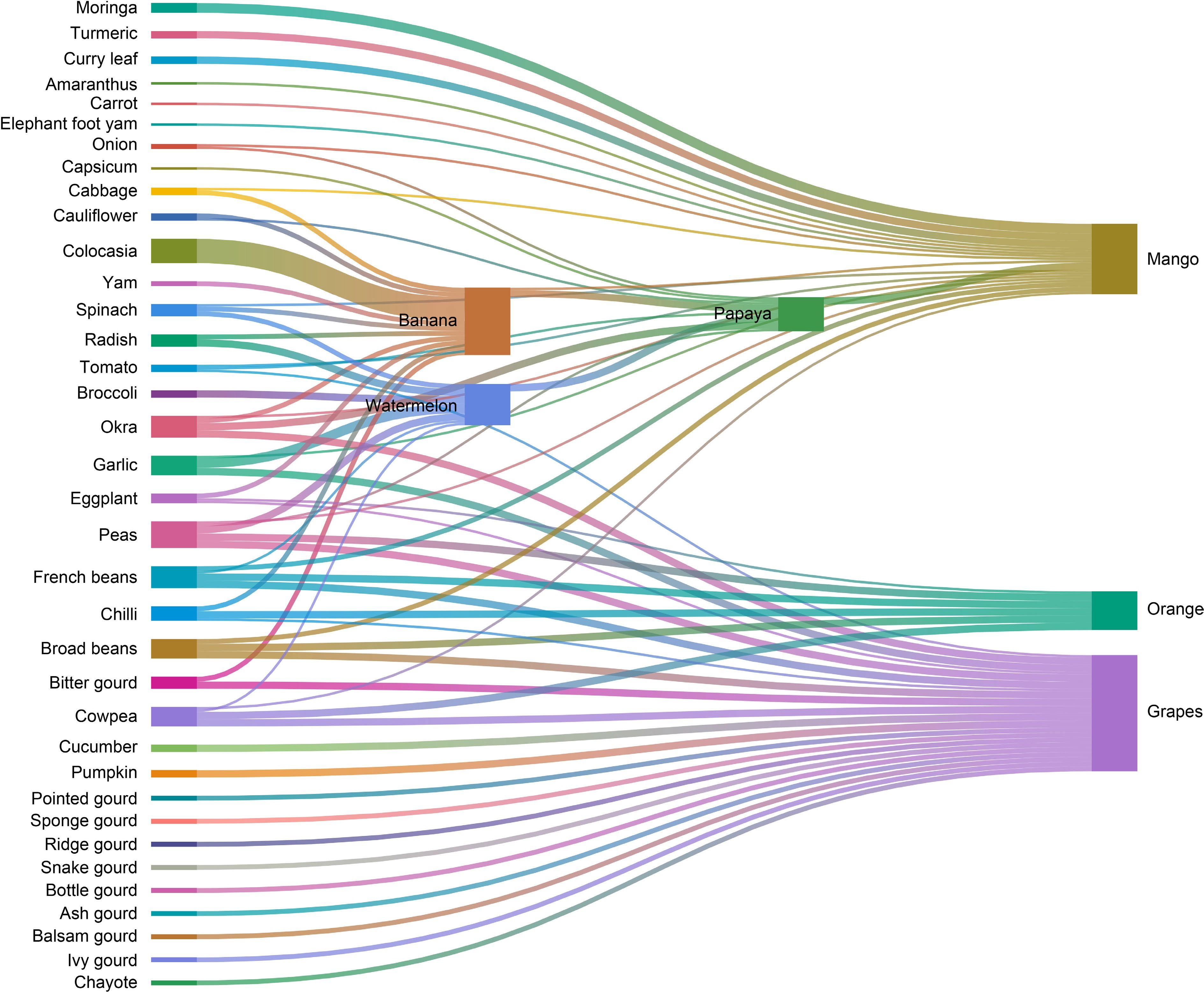
Figure 3.
Existing and potential agroforestry (intercropping) between fruit and vegetables in the Devdaha Municipality expressed in a Sankey diagram. The thickness of the line at vegetable (left) end indicate higher potential for intercropping.
The diagram reveals several promising agroforestry systems:
• Mango has high potential when intercropped with Moringa, papaya, banana, and various vegetables.
• Intercropping papaya with banana, watermelon, and other vegetables also shows good suitability.
• Banana can be profitably intercropped with colocasia, yam, and others.
• Grapes and cucurbits make a suitable pairing.
• Growing legumes under orange trees enhances the system.
The diagram aligns with observations made during field visits regarding existing agroforestry practices, while also suggesting potential for expansion. The findings suggest incorporating tree-vegetable combinations based on identified suitability can benefit local farms. For instance, mango-Moringa intercropping provides economic and ecological advantages, as Moringa fixes nitrogen while also generating income. Additionally, mango systems allow intercropping with papaya, banana, and various vegetables, maximizing land use and income potential through diversification.
Other favorable combinations are papaya-banana-watermelon and banana-colocasia-yam intercropping. The assessment encourages exploring grape-cucurbit and orange-legume systems as well. Implementing these suitable intercropping strategies can increase farm resilience, diversify income, and promote sustainable agriculture in the Municipality.
-
Local farmers in Devdaha face compounding challenges, including limited production, restricted market access for their products, and fierce competition from cheaper imports. This vulnerability, particularly pronounced for female farmers with limited travel options and connections, manifests as a concerning shift from self-sufficiency to reliance on imported food. This aligns with the observations by Sunam & Adhikari[28] and Adhikari et al.[29] on the rise of imports across Nepal's markets. The decline in production and shift to imported foods risks further deepening food insecurity. Declines in agricultural production can have serious consequences, including reduced food availability, heightened food insecurity, economic hardship, and negative social impacts[13−16,19]. According to NPC and WFP[17], Rupandehi district continues to suffer from chronic to minimal food insecurity. This trend not only threatens local livelihoods and economies, but also exacerbate existing food insecurity, as evidenced by the district's chronic food shortages, and increases dependence on volatile global food prices, as seen during the COVID-19 disruptions[29].
However, the findings also reveal opportunities for promoting sustainable local agriculture through:
(1) Locally Managed Farmers' Markets: This study's findings suggest that establishing well-managed farmers' markets close to farms offers a promising path toward boosting sustainable local agriculture. Such markets would not only address accessibility challenges, especially for women but also create a win-win situation for farmers and the community. By ensuring fair prices and reducing transportation costs (including carbon emissions from long-distance food transportation), these markets can directly support local economies and encourage small business incubation[30]. Additionally, they would promote the use of indigenous crops and food varieties, preserving local food cultures and traditions[31−33]. Research suggests that proximity to markets motivates farmers to increase production and activity[22,34,35]. Ultimately, the nexus of sustainable agriculture, local agro-production and well-managed farmers' markets can contribute to food security by facilitating access to healthy and nutritious food, promoting sustainable livelihoods for farmers, and reducing reliance on imported food items[18,31,32].
(2) Promoting high-marketability products: Diversified local agro-production are key components of sustainable agriculture[36,37]. Diversification based on market demand can increase farmer income and address reliance on imports. The study conducted a comprehensive inventory, identifying a total of 57 farm-based products currently cultivated in the region. By strategically prioritizing items like mangoes, bananas, papayas, grapes, watermelons, citrus fruits, ginger, and turmeric within an agroforestry framework for marketing, the research suggests a viable strategy for local farmers to augment their income through diversification[38,39].
(3) Implementing agroforestry practices: Prioritizing high marketability and profitability products and promote them in an agroforestry system could increase opportunity in improving market linkages that help farmers increase their incomes and improve their livelihoods[15]. These systems offer benefits like soil fertility, carbon sequestration, and income diversification, contributing to climate change mitigation and sustainable agriculture. The adoption of agroforestry systems emerges as a crucial tactic in addressing climate change and fostering sustainable agriculture[36,39]. These systems bring multifaceted benefits, including erosion control, enhanced soil fertility, and carbon dioxide sequestration, thereby playing a pivotal role in mitigating climate change.
Trade-offs
-
While agroforestry and local markets present promising avenues for sustainable agriculture and increased farmer income, they are not without drawbacks[40,41]. Agroforestry systems, despite offering long-term benefits like soil health and income diversification, require higher initial investments and patience due to slower tree maturation[40]. Balancing land use between tree and annual crops also poses challenges. Additionally, managing diverse agroforestry systems can be labor-intensive. On the other hand, local markets, while reducing transportation costs and supporting local economies may have limited reach and face fluctuations in seasonal demand[41]. Some farmers may also remain reliant on intermediaries, impacting their profit margins. Recognizing these trade-offs is crucial for a comprehensive evaluation of these strategies and their effectiveness in addressing the challenges faced by small-scale farmers.
Policy implications
-
This study's findings offer valuable insights for policymakers aiming to achieve Sustainable Development Goals (SDGs) related to food security, poverty reduction, and sustainable communities. To support sustainable agriculture and promote local agro-production, well-managed farmer's markets close to farms are essential[22,34,35]. Such markets can stimulate local economies, reduce transportation-related carbon emissions, and preserve indigenous food cultures and traditions, all of which contribute to achieving SDGs related to sustainable consumption and production (SDG 12) and sustainable communities (SDG 11). Additionally, promoting agroforestry practices, prioritizing high marketability and profitability products, and improving market linkages can help farmers increase their incomes and improve their livelihoods[15]. These actions align with SDG 1 (No Poverty) by reducing poverty among small-scale farmers and SDG 2 (Zero Hunger) by ensuring food security and zero hunger. Furthermore, reducing dependency on imported food items through local agro-production can contribute to SDG 2, reducing the risks associated with volatile global food prices and the vulnerability of the food system to supply chain disruptions. By emphasizing gender-sensitive interventions and improving access to credit, agricultural subsidies, and extension services, policymakers can support female farmers in accessing markets, thereby advancing gender equality (SDG 5). Adopting these strategies not only fosters sustainable agriculture but also contributes to the achievement of multiple SDGs, making them valuable components of holistic policy frameworks for agricultural development.
-
This study delved into the agricultural landscape of Devdaha Municipality, Nepal, uncovering valuable insights into current practices, market dynamics, and promising avenues for fostering sustainable agriculture. Examining production seasons, market barriers, commercialization opportunities, and existing sustainable farming practices yielded two key findings.
First, over 50 high-marketability fruits, vegetables, and agroforestry crops were identified, showcasing the potential for diversifying production and boosting farmer income. This diversity can contribute to a more circular agricultural practice by reducing reliance on external inputs like fertilizers and pesticides. By utilizing various crops within the farm, farmers can potentially create a more closed-loop system where waste from one crop can become a resource for another.
The second key finding revealed persistent challenges for farmers in accessing markets and competing with cheaper imports. These challenges and competition highlight the need for closing loops within the local food system. The identified solutions, such as establishing local farmers' markets and promoting agroforestry, directly address these concerns and contribute to a more circular approach.
To address these challenges and to capitalize on the identified opportunities, this study recommends the following approaches:
• Cultivating high-value crops: Provide targeted training, inputs and incentives (subsidies, tax benefits) to encourage farmers to adopt the study's identified high-value fruits, vegetables, and agroforestry systems. This could include promoting intercropping techniques that enhance soil fertility and reduce reliance on external inputs, fostering a more closed-loop system.
• Building vibrant local food systems: Establish well-managed farmers' markets close to farms to improve accessibility, reduce food transportation distances and minimize food waste. This fosters a more localized food system, keeping resources circulating within the community.
• Demonstrating the benefits: Implement demonstration pilots showcasing the advantages of the identified agroforestry systems, allowing farmers to witness improved soil health, increased yields, and long-term economic benefits. By the time of completing this paper, the agroforestry demonstration plots along the riverbed have been established in 22 ha using the fruits and vegetables as suggested in the results.
• Facilitating knowledge exchange: Initiate knowledge exchange programs among farmers to share best practices and encourage wider adoption of sustainable agroforestry techniques.
By implementing these focused interventions, this study provides a roadmap for empowering Devdaha's farmers, enhancing food security, and promoting sustainable agricultural practices. This holistic approach holds the potential to revitalize the local agricultural landscape, improve farmer livelihoods, and create a more resilient and prosperous future for the community.
-
The authors confirm contribution to the paper as follows: study conception and design: Ranjitkar S, Bhuju UR; data collection: Ayub M, Ranjitkar S; analysis and interpretation of results: Ranjitkar S, Sujakhu NM; draft manuscript preparation: Ranjitkar S, Ayub M. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
We would like to express our sincere gratitude and appreciation to the Asia Foundation for their financial support, which made this study possible. We would also like to extend our thanks to the local farmers in the Devdaha Municipality for their valuable contribution to the study by providing us with the necessary information and insights into their farming activities. Without their cooperation, this study would not have been possible. We are also thankful to the local authorities of Devdaha Municipality for their support in facilitating the study, and providing us with the necessary permits and clearances.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ranjitkar S, Ayub M, Bhuju UR, Sujakhu NM. 2024. Promoting sustainable agriculture in Devdaha Municipality, Nepal: leveraging agroforestry to address challenges and enhance food security. Circular Agricultural Systems 4: e013 doi: 10.48130/cas-0024-0012 

Promoting sustainable agriculture in Devdaha Municipality, Nepal: leveraging agroforestry to address challenges and enhance food security
- Received: 16 February 2024
- Revised: 08 May 2024
- Accepted: 21 May 2024
- Published online: 02 August 2024
Abstract: Ensuring food security through sustainable agriculture is critical for achieving the Sustainable Development Goals (SDGs). This study aimed to examine farming practices, assess the marketability of agricultural products, and propose interventions to promote sustainable agriculture in Devdaha Municipality, Nepal, with a focus on overcoming gender disparities. Utilizing purposive sampling, interviews, focus group discussions, expert consultations, and direct observations followed by mixed methods analysis, farming practices were examined, the marketability of agricultural products were assessed, and interventions for improvement were proposed. Analysis revealed a diverse range of farm products, identified key challenges limiting profitability (e.g., seasonal variations, market access), and highlighted the need for targeted interventions. Diversifying production towards high-profit crops like mango, banana, papaya, ginger, and turmeric integrated into agroforestry systems was identified as a potential strategy to increase farmer incomes and livelihoods. Findings revealed that cheaper imports contributed to a decline in local production, making efficient marketing systems and gender-sensitive interventions crucial to reverse this trend and empower women's participation in commercial agriculture. The study recommends establishing well-managed local farmers' markets, implementing agricultural subsidies, and launching campaigns to promote indigenous crops to reduce import dependency and boost incomes. Additionally, promoting suitable agroforestry combinations like mango-Moringa-vegetables intercropping can enhance sustainability. By informing on localized strategies aligned with SDG 2, this study offers policymakers potential pathways to strengthen sustainable agriculture, food security, and gender equity in Nepal's rural municipalities.
-
Key words:
- Local production /
- Sustainable farming /
- Food security /
- Farmers market /
- Agroforestry