









-
Photosynthetic carbon gain is an important aspect of plant performance in natural and agricultural systems[1]. Identification of the genes that mediate photosynthetic processes in plants would be critical for understanding the regulation of photosynthetic carbon gain in leaves, as well as the allocation of that carbon to flowers and fruits. Consequently, the process of photosynthesis, a fundamental biological mechanism primarily occurring in leaves, assumes a pivotal position in shaping the overall physiology of fruit trees and significantly influences the production of fruit[2]. Photosynthesis is the process by which plants, including fruit trees, capture energy from sunlight and transform it into vital chemical energy and structural building blocks in the form of sugars. This energy serves as the lifeblood of the plant, fueling growth, development, and various physiological functions[3,4]. Photosynthetic capacity, efficiency and activity can be quantified through traits such as radiative absorptance of contrasting wavelengths, chlorophyll content, and various aspects of light use and excess light quenching collectively calculated through chlorophyll fluorescence[5−7]. Recently, technological developments that allow for the non-destructive high-throughput phenotyping are becoming increasingly important to monitor crop photosynthetic responses to changing environmental conditions and to potentially model and predict plant growth and development under specific environmental constraints[8]. Therefore, we employed a novel approach to rapidly collect multiple photosynthetic parameters using leaf spectral reflectance within a mapping population of citrus to better understand the linkages between photosynthetic processes and gene function.
Physiological aspects related to chlorophyll represent one of the most fundamental methods to probe photosynthetic performance in plants. Chlorophyll content, for example, plays a pivotal role in capturing light energy[9]. It acts as the primary pigment responsible for absorbing sunlight and generating the chemical energy to promote atmospheric CO2 assimilation into organic molecules. The efficiency of this process directly influences the capacity of a tree to produce sugars, thereby impacting fruit production[10]. A higher chlorophyll content translates into increased energy production, fostering the development of healthier and more abundant fruit. On the other hand, absorbance at various wavelengths determines how proficiently leaves capture and utilize light energy[11]. In contrast, the quenching ratio of incoming light is associated with the ability of a tree to balance the absorption of light while implementing protective mechanisms to prevent damage from excessive sunlight exposure[12]. Additionally, chlorophyll fluorescence serves as an indicator of the health and vitality of the photosynthetic apparatus within the plant. It provides insights into plant responses to environmental factors and stressors that may influence fruit tree physiology[13]. Variable Fluorescence (Fv) represents the difference between the minimal (Fo) and variable (Fv) fluorescence levels in a dark-adapted leaf. It indicates the portion of the total fluorescence that can change in response to variations in the light environment, reflecting the dynamic nature of the photosynthetic process[14]. In other words, Fv measures how much the fluorescence of a leaf can change in response to varying light conditions. Maximum Fluorescence (Fm), on the other hand, is what a dark-adapted leaf can emit when all photosystem II (PSII) reaction centers are closed, and all available chlorophyll molecules are in their excited state. It serves as a fixed value for a given leaf under specific conditions and is used as a reference point for calculating other fluorescence parameters. The ratio FvP/FmP, where 'P' represents the prime values (adjusted for different measuring conditions), is employed to evaluate the efficiency of photosystem II (PSII) in converting absorbed light energy into chemical energy during photosynthesis. Non-photochemical quenching (NPQ)[15], PhiNPQ[16], and NPQt[17] describe the excited state of chlorophyll a, which is a major and extensively studied photoprotective mechanism for plants to survive under high light conditions[18]. This parameter is especially valuable in assessing the photosynthetic health of plants, providing insights into the functionality of PSII reaction centers and their capacity to perform photosynthesis under varying conditions[19,20]. Collectively, these photosynthesis-related traits make substantial contributions to the overall physiological function of fruit trees. They determine the ability of a tree to convert sunlight into energy and, consequently, into fruits. Understanding and analyzing these traits can enhance fruit production, optimize tree health, and inform decision-making in the cultivation and management of fruit orchards. This is especially true in the context of a genome-wide association study (GWAS), a powerful tool for identifying genetic variations and critical genes, which in turn can be used to develop crops with higher photosynthetic capacity and yield.
Here, we measured 37 photosynthesis-related traits from a selection of 71 citrus accessions or varieties sourced from the Givaudan Citrus Variety Collection (GCVC) at the University of California, Riverside. Additionally, we obtained genomic data comprising over 56,000 single-nucleotide polymorphisms (SNPs) using Affymetrix™ Axiom® Citrus genotyping arrays. Pairwise correlation analysis of these biophysical measurements revealed groupings of interrelated traits within distinct clusters. Notably, the variations observed in these intricate trait profiles, which represent a unique form of phenotypic outcomes arising from genetic diversity and its interaction with environmental factors, exhibited associations with the citrus phylogeny. We conducted comprehensive GWAS on these photosynthesis-related traits, with the goal of identifying genomic loci and potentially linked genes responsible for the observed trait variations. Our analysis identified a total of 125 loci and 189 genes contributing to seven specific photosynthesis-related traits drawn from different trait clusters. Additionally, we delved into a discussion regarding the potential pathways that may influence the development of these traits within the citrus population.
-
The study was conducted at the Givaudan Citrus Variety Collection (GCVC) at the University of California, Riverside, USA (33°58'32" N 117°19'52" W). The collection contains approximately 4,500 trees of nearly 1,100 cultivars and species of citrus and related taxa, including commercial and historic citrus cultivars and other related genera and species in the Aurantioideae subfamily of the Rutaceae. We measured two leaves on each of 71 citrus accessions or varieties with associated genetic sequence data.
Leaf photosynthetic and environmental measurements
-
Measurements of plant photosynthetic phenotypical parameters including photosystem II quantum yield (Phi2), non-photochemical exciton quenching (NPQ), photosystem II photoinhibition, light-driven proton translocation and thylakoid proton motive force, regulation of the chloroplast ATP synthase as well as minor parameters (Supplemental Table S1), leaf orientation, pigments, and environmental conditions such as including light intensity and quality, temperature, humidity, CO2 levels, time and location, were conducted with a MultispeQ unit (PhotosynQ Inc., East Lansing, USA), a low-cost, open-source scientific instrument designed to collect high-quality field data on a large scale[16]. Plant phenotype data are transmitted from the MultispeQ to mobile devices, laptops or desktop computers together with key metadata that gets saved to the PhotosynQ platform (
https://photosynq.org ) and provides a suite of web-based tools for sharing, visualization, filtering, dissemination and analyses.Genomic data
-
The genomic data for the 71 accessions were obtained from the Citrus56AX array, a tailored Affymetrix™ Axiom® Citrus SNP array encompassing 57,933 autosomal SNPs and 500 chloroplast SNPs[21]. This genomic dataset played a pivotal role in the genetic dissection of photosynthesis-related traits.
Genome-wide association study
-
A Genome-Wide Association Study (GWAS) is a genetic research method designed to identify genetic variations, typically single-nucleotide polymorphisms (SNPs), associated with specific traits or diseases. In our study, we utilized GEMMA[22] for this purpose. Our approach involved two key steps: First, we excluded SNP variants with a minor allele frequency below 5% and removed any SNPs with a missing rate exceeding 20%. Second, we applied the Linear Mixed Model (LMM) within GEMMA, which incorporated a kinship matrix to account for genetic relatedness and correct for population structure. We employed a p-value threshold of 10-5 to identify significant associations between photosynthesis related traits and SNPs. To identify the nearest genes associated with each SNP, we utilized bedtools closest[23].
Pathway analysis
-
Subsequent to the GWAS, we pinpointed 11 photosynthesis-related traits whose variability could be accounted for by 125 significantly associated SNPs. To investigate the genetic underpinnings of these traits, we employed 'bedtools closest', which is part of the Bedtools suite[23] to locate a total of 189 genes containing or in close proximity to these SNPs. Following this, we performed gene ontology enrichment analysis using the Citrus Pan-genome to Breeding Database (CPBD)[24], accessible at
http://citrus.hzau.edu.cn/enrichment/query.php .Other statistical analyses
-
We used Pearson correlation analysis to evaluate linear relationships between two variables, shedding light on their associations. Pairwise correlation analysis enabled us to explore correlations among pairs of variables, such as the 37 photosynthesis-related traits in this study. This analysis unveiled patterns of interdependence within the dataset. We constructed distribution histograms to visualize the frequency distribution of data, facilitating our understanding of central tendencies and the spread of values within the dataset. Principal Component Analysis (PCA) was employed to reduce data dimensionality and uncover key patterns and variations among the variables. This technique simplified complex datasets and extracted essential information. All these analytical methods were executed within the R environment, collectively contributing to a thorough exploration of the data and enhancing our understanding of the research findings.
-
A pairwise correlation analysis of the 37 photosynthesis-related traits revealed distinct clusters of interrelated traits is depicted in Fig. 1. These clusters can be described as follows:
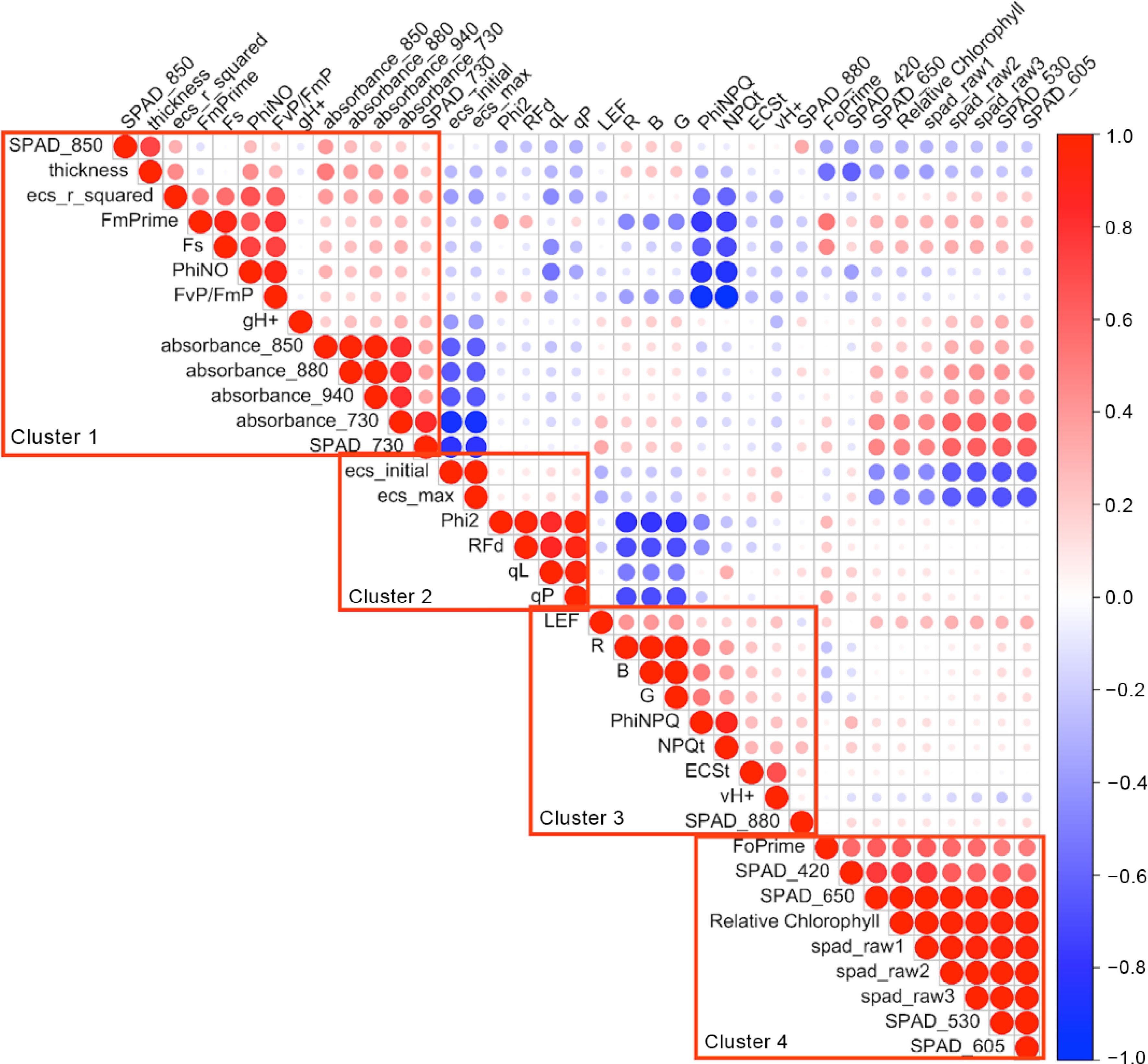
Figure 1.
Heatmap illustrating the pairwise correlations among 37 photosynthesis-related traits, with clustered traits showing interrelatedness.
Cluster 1: This cluster encompasses traits such as photochemical efficiency (FvP/FmP), leaf thickness (measured with the Hall Effect sensor), absorbance at various wavelengths, Special Products Analysis Division (SPAD, equivalent to relative chlorophyll content) at longer wavelengths, the r-squared value of electrochromic shift (ecs_r_squared), maximal fluorescence (FmPrime), steady-state fluorescence (Fs), the ratio of incoming light (PhiNO, representing excited electrons diverted towards non-photochemical quenching), and proton conductivity (gH+).
Cluster 2: In this cluster, you can find traits including initial and maximum values of electrochromic shift (ecs_initial and ecs_max), Quantum yield of Photosystem II (Phi2), Fluorescence decrease ratio (RFd), Photosystem II Redox State using the 'Lake' model (qL) and 'Puddle' model (qP), and Linear Electron Flow (LEF).
Cluster 3: Cluster 3 encompasses traits related to the raw amounts of red light (R), blue light (B), and green light (G) captured by the PAR sensor, the ratio of incoming light that is lost through non-regulated processes (PhiNPQ), non-photosynthetic quenching (NPQt), electrochromic shift (ECSt), Steady-State Proton Flux (vH+), and SPAD measurement at the highest wavelength of 880.
Cluster 4: This cluster comprises various raw measurements of SPAD, SPAD at shorter wavelengths, relative chlorophyll content, and minimal fluorescence (FoPrime).
These clusters reflect the interrelated nature of these photosynthesis-related traits, grouping them based on their shared correlations. Our analysis revealed the following correlations:
• A negative correlation between SPAD 850 and Phi2 (r = −0.273; p-value = 0.021), qL (r = −0.289; p-value = 0.014), and qP (r = −0.311; p-value = 0.008).
• NPQt demonstrated a significant positive correlation with PhiNPQ (r = 0.881; p-value < 0.001).
• Phi2 exhibited noteworthy positive correlations with FvP/FmP (r = 0.240; p-value = 0.043), qL (r = 0.826; p-value < 0.001), and qP (r = 0.952; p-value < 0.001).
• PhiNO showed a positive correlation with FvP/FmP (r = 0.912; p-value < 0.001) but displayed negative correlations with PhiNPQ (r = −0.839; p-value < 0.001), qL (r = −0.547; p-value < 0.001), and qP (r = −0.345; p-value = 0.003).
Understanding these trait clusters can provide valuable insights into the underlying factors influencing photosynthetic processes and plant physiology.
Phylogenetic insights through photosynthesis-related traits
-
Principal Component Analysis (PCA) was performed on both genomic data and photosynthesis-related trait data derived from a set of 71 citrus accessions and varieties. This selected citrus collection encompasses four discernible citrus groups, including citron, lemon, pomelo, and mandarin, along with hybrid varieties. In the analysis of genomic data, these four distinct citrus groups were clearly delineated (Fig. 2a). PCA analysis based on photosynthesis-related traits demonstrated the capacity to distinguish these primary citrus groups, although some level of overlap was observed (Fig. 2b). The variations observed in these biophysical traits, including SPAD, NPQt, PhiNPQ, and Fv/Fm, are influenced by genotype, reflecting the combined impact of numerous genetic factors and their intricate interactions with environmental variables.
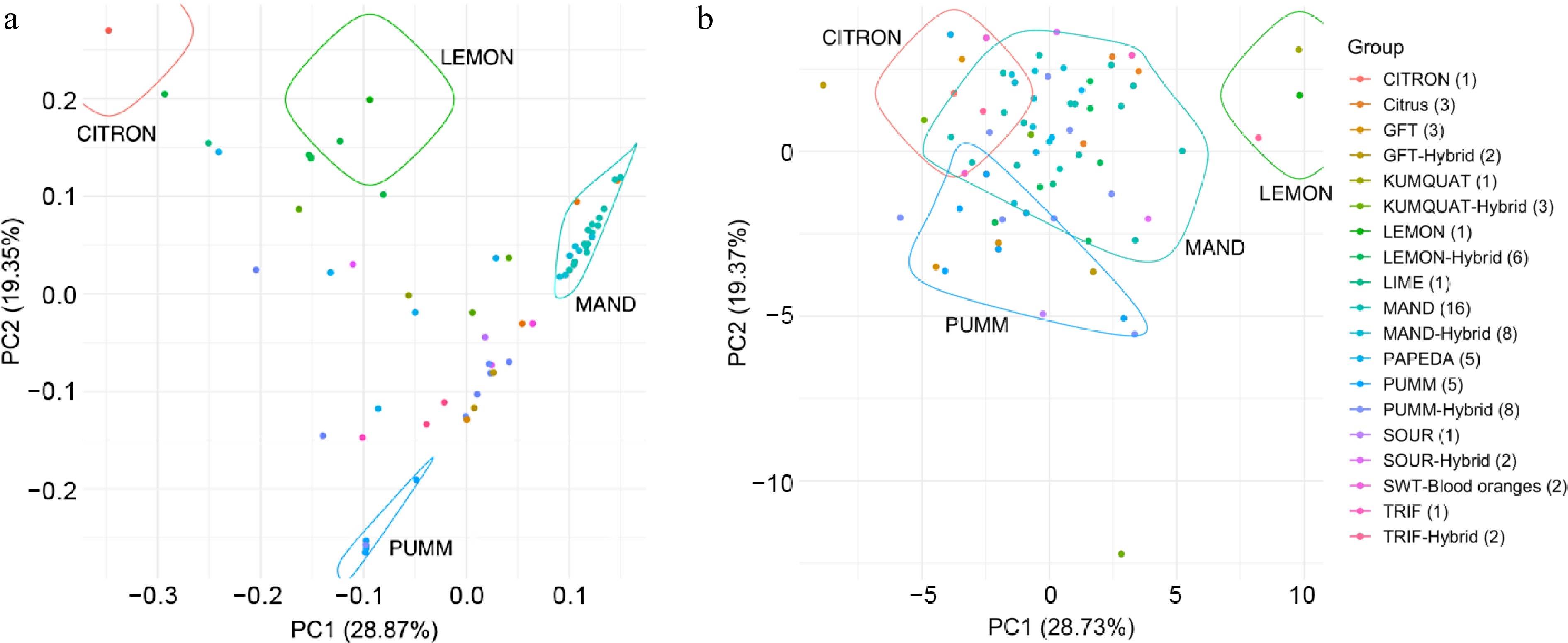
Figure 2.
Principal component analysis: (a) Genomic data, (b) photosynthesis-related traits.
Genome-wide association study
-
A genome-wide association study was conducted on all 37 photosynthesis-related traits. Figure 3 demonstrates the detection of 125 genomic loci associated with 11 traits. Detailed information about these 125 loci is provided in Supplemental Table S2. Notably, among these traits, the critical photosynthesis-related trait FvP/FmP, with a bell-shaped phenotypic distribution, is highlighted in panel a. These 11 traits encompass a diverse array of parameters, including FvP/FmP, FmPrime, ecs_r_squared, and Fs from cluster 1, ecs_initial and ecs_max from cluster 2, and B, NPQt, vH+, SPAD 880 from cluster 3, along with SPAD raw 3 from cluster 4. The heatmap in Fig. 1 offered valuable insights by revealing positive or negative correlations between these clusters, signifying a collaborative role among these traits in the context of photosynthesis.
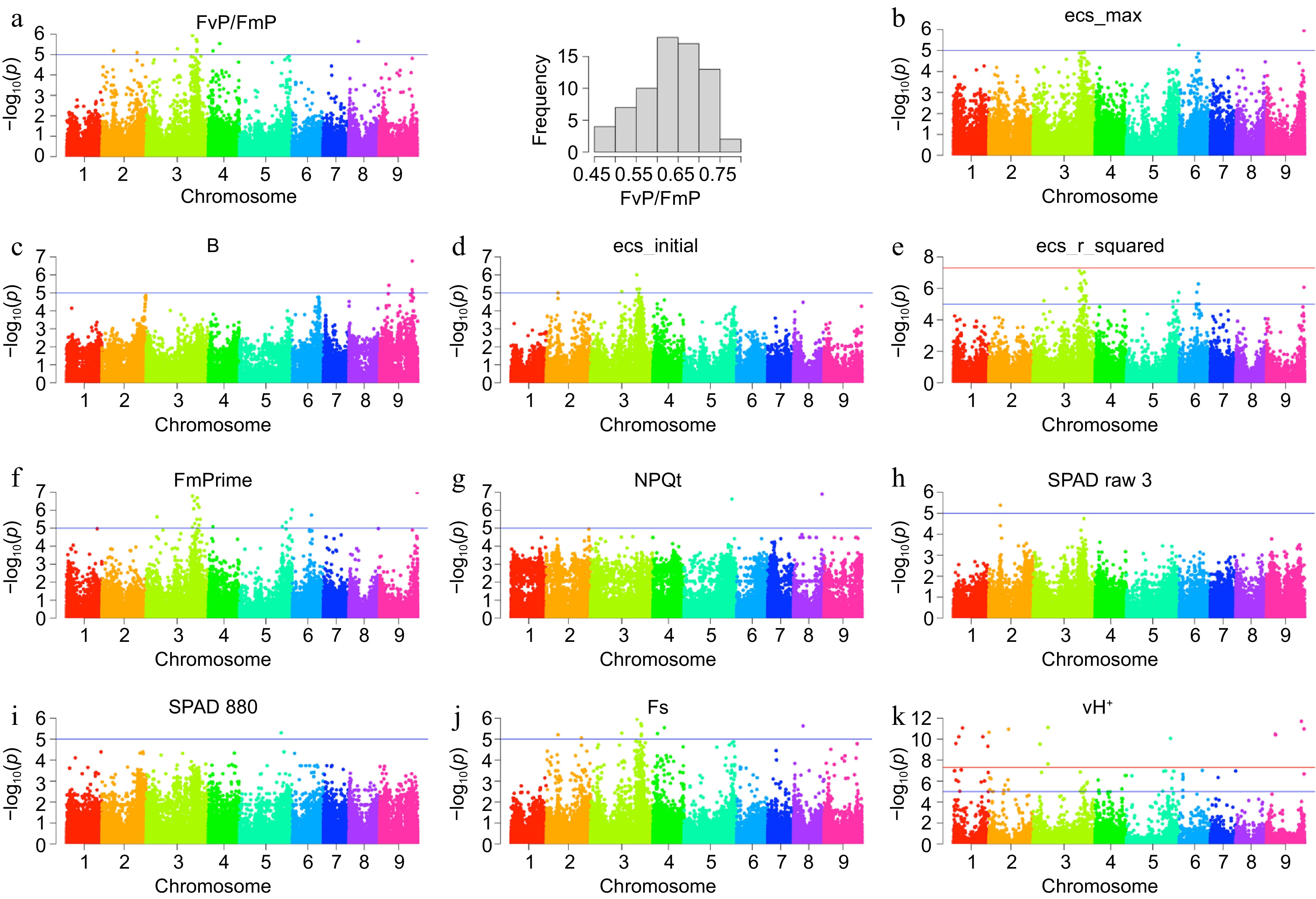
Figure 3.
Genome-wide association study. (a) Manhattan plot for FvP/FmP, the most significant photosynthesis-related trait, and its phenotypic variation across 71 citrus accessions. (b)−(k) Manhattan plots for the remaining 10 traits, which include ecs_max, B, ecs_initial, ecs_r_squared, FmPrime, NPQt, SPAD raw 3, SPAD 880, Fs, and vH+.
Biological annotation of associated genes
-
To annotate all significant SNPs identified in the GWAS, we employed the 'bedtools closest' tool in conjunction with the Cclementina_182_v1.0.gene.gff3 file[25]. This method resulted in the identification of 189 genes. Detailed information about these 189 potentially associated genes is provided in Supplemental Table S3. Among these, 82 genes (comprising a total of 12,556 within the genome) were linked to the 'biological process (BP)' Gene Ontology (GO) term, as documented in the database accessible at
http://citrus.hzau.edu.cn/index.php . Furthermore, out of the 189 genes, 111 (constituting a total of 20,076 within the genome) were associated with the 'molecular function (MF)' GO term, as indicated by the same database. The significantly represented GO terms are detailed in Table 1, encompassing a range of biochemical and molecular activities in cells and organisms. While some of these terms may have indirect connections to processes associated with photosynthesis, such as the glycolytic process, mitochondrial ADP transmembrane transport, mitochondrial ATP transmembrane transport, multicellular organism development, protein insertion into the mitochondrial outer membrane, protein phosphorylation, protein dephosphorylation, RNA-directed 5'-3' RNA polymerase activity, and transferase activity involving glycosyl groups, others are primarily linked to broader cellular functions.Table 1. Pathway analysis annotated with Biological Process (BP) and Molecular Function (MF) terms based on 189 genes.
Description Count p-value FDR BP GO:0006470 Protein dephosphorylation 7/82 86/12556 1.3E-06 3.18E-03 GO:0006096 Glycolytic process 6/82 65/12556 3.72E-06 4.54E-03 GO:0140021 Mitochondrial ADP transmembrane transport 3/82 9/12556 2.17E-05 1.77E-02 GO:1990544 Mitochondrial ATP transmembrane transport 3/82 9/12556 2.17E-05 1.33E-02 GO:0007275 Multicellular organism development 4/82 29/12556 3.43E-05 1.68E-02 GO:0045040 Protein insertion into mitochondrial outer membrane 2/82 3/12556 1.26E-04 5.11E-02 GO:0000184 Nuclear-transcribed mRNA catabolic process, nonsense-mediated decay 3/82 19/12556 2.35E-04 8.21E-02 GO:0006468 Protein phosphorylation 2/82 1611/12556 9.04E-04 2.76E-01 MF GO:0047334 Diphosphate-fructose-6-phosphate 1-phosphotransferase activity 5/111 9/20076 5.82E-10 1.42E-06 GO:0003872 6-phosphofructokinase activity 5/111 17/20076 2.74E-08 3.35E-05 GO:0004722 Protein serine/threonine phosphatase activity 7/111 64/20076 6.02E-08 4.9E-05 GO:0005471 ATP:ADP antiporter activity 3/111 11/20076 2.6E-05 1.59E-02 GO:0016791 Phosphatase activity 8/111 233/20076 4.02E-05 1.97E-02 GO:0043169 Cation binding 5/111 82/20076 8.56E-05 3.48E-02 GO:0003968 RNA-directed 5'-3' RNA polymerase activity 2/111 3/20076 9.04E-05 3.15E-02 GO:0016757 Transferase activity, transferring glycosyl groups 6/111 205/20076 8.36E-04 2.55E-01 -
The photosynthesis-related traits examined in this study collectively play a defining role in shaping the overall health and productivity of fruit trees. They are known to hold the key to the ability of a tree to harness sunlight and transform it into the energy necessary for fruit production[26]. Importantly, these traits exhibit variations among different genotypes, encompassing a diverse array of citrus cultivars and accessions. This insight is substantiated by the data we have gathered, wherein the distinct profiles of these biophysical variables offer valuable clues about the underlying citrus phylogeny. By scrutinizing these photosynthesis-related traits, we not only enhance our comprehension of the biophysical distinctions among various citrus cultivars and accessions, but we also open doors to the prospect of elevating fruit production, optimizing tree health, and making well-informed choices in the realms of fruit tree cultivation and orchard management.
The correlation analysis unveiled four primary clusters of photosynthesis-related traits, as depicted in Fig. 1. Cluster 1 encompasses traits closely interconnected due to their roles in light absorption, energy conversion, and the overall functionality of the photosynthetic apparatus. Cluster 2 includes traits that collectively offer insights into the dynamics, efficiency, and health of the photosynthetic apparatus, specifically Photosystem II. Traits in cluster 3 collectively provide information about the plant's responses to the light environment, its protective mechanisms against excess light exposure, and the efficiency of photosynthesis, reflecting the dynamic nature of photosynthesis processes and the plant's adaptability to changing environmental conditions. Finally, cluster 4 focuses on chlorophyll content and fluorescence, fundamental to photosynthesis, offering information about the plant's capacity to capture and utilize light energy efficiently. The heatmap illustrated in Fig. 1 proved enlightening. It underscored positive affiliations between clusters 1 and 4, while negative associations between clusters 1 and 2 or between clusters 2 and 3. This intricate pattern underscores the physiological coordination among these traits within the broader framework of plant function[27]. The correlations, either positive or negative, among these four clusters emphasize their collaborative role in influencing the efficiency, functionality, and adaptability of photosynthesis within the citrus population, as well as in response to varying environmental conditions, highlighting the intricate web of interactions at play.
Previous studies on rice, including one by Chen et al.[28] and one by Liu et al.[29], have reported the following correlations: (1) SPAD, serving as an indicator of chlorophyll content, showed negative correlations with Phi2, qL, and qP. (2) NPQt, reflecting non-photochemical quenching, exhibited a significant positive correlation with PhiNPQ. (3) Phi2, an indicator of photochemical efficiency, displayed notable positive correlations with Fv/Fm, qL, and qP. (4) PhiNO showed a significant positive correlation with Fv/Fm but exhibited negative correlations with PhiNPQ, qL, and qP. (5) A positive association was found between SPAD and leaf thickness. (6) Despite higher chlorophyll content, increased leaf thickness reduced translucency, resulting in a negative correlation between SPAD and Phi2. Most of these correlations were also observed in our citrus dataset.
Integrating the intricate web of photosynthesis-related traits with comprehensive genomic data has proven to be an invaluable key that can unlock the genetic determinants that underpin these vital biophysical variables[30]. In our investigation, a GWAS was executed on 37 photosynthesis-correlated traits, yielding promising findings. As depicted in Fig. 3, we detected 125 genomic loci intertwined with 11 distinct traits. One of the standout traits among these is FvP/FmP, a pivotal facet of photosynthesis, showcased with a bell-shaped phenotypic distribution in panel a. These traits spanned varied parameters across four clusters. Delving deeper into the genomic intricacies, we utilized the 'bedtools closest' tool to annotate the significant SNPs unveiled by the GWAS, referencing the Cclementina_182_v1.0.gene.gff3 file. This rigorous approach brought to the fore 189 genes potentially impacting photosynthesis and therefore these relevant traits, the specifics of which have been elucidated in Supplemental Table S3. A closer examination revealed that 82 out of these 189 genes were resonant with the 'biological process' GO term, according to the database hosted at
http://citrus.hzau.edu.cn/index.php . In a parallel vein, 111 genes were associated with the 'molecular function' GO term. A comprehensive list of these GO terms, shedding light on a spectrum of cellular and molecular activities, is detailed in Table 1. Some bear indirect ties to photosynthesis-related processes, including the glycolytic process and mitochondrial ATP transport. In contrast, others align more broadly with overarching cellular functions.Among the listed terms, the following may be indirectly associated with the photosynthesis process:
(1) Glycolytic process: The glycolytic process involves the breakdown of glucose to produce energy, and photosynthesis is a process that captures energy from sunlight. While they are not directly related, both processes are part of the overall energy balance within a cell or organism;
(2) Mitochondrial ADP transmembrane transport and mitochondrial ATP transmembrane transport: These processes relate to the movement of ADP and ATP (adenosine diphosphate and adenosine triphosphate), which are involved in cellular energy transfer. Photosynthesis is a fundamental energy-capturing process in plant chloroplasts, so these processes may indirectly affect the overall energy balance within a cell;
(3) Multicellular organism development: While not directly linked to photosynthesis, the development and growth of multicellular organisms depend on energy, some of which is derived from photosynthesis;
(4) Protein insertion into mitochondrial outer membrane: This process involves cellular localization and membrane transport of proteins, which may be indirectly related to the overall functioning of mitochondria and energy processes, including photosynthesis in plant cells;
(5) Protein phosphorylation and protein dephosphorylation: These processes involve the addition and removal of phosphate groups on proteins, which can modulate their activity and may have implications for signal transduction pathways related to photosynthesis;
(6) RNA-directed 5'-3' RNA polymerase activity: This process is related to transcription and the synthesis of RNA. RNA is involved in various cellular processes, including gene expression, which can indirectly impact photosynthesis regulation;
(7) Transferase activity, transferring glycosyl groups: This process may involve enzymes related to glycosylation and carbohydrate metabolism, which can play a role in energy storage and overall cellular function.
However, it is worth noting that the CPBD, the database used for functional annotation in our study, is relatively new and still expanding. The gene annotation for citrus remains incomplete; For instance, only 82 out of the 189 genes have been fully annotated in terms of biological processes, and 111 out of the 189 genes have been annotated for molecular function. This incompleteness presents challenges in identifying enrichment for numerous candidate genes due to the absence of information in the database. As a result, our analysis is limited to the genes currently documented in the database, potentially affecting the robustness of our findings. We have provided a comprehensive list of all candidate genes in Supplemental Table S3 for reference. As genomic methodologies continue to advance and citrus-related databases expand, we anticipate that future researchers will utilize these tools and resources to validate the functions of these genes more definitively.
This synergy between phenotypic traits and genomic insights demonstrated in this study not only enriches our understanding of the complex interplay between genetics and physiology but also holds profound implications for practical applications[30−34]. By delving into the genetic intricacies that govern photosynthesis in fruit trees, this research extends a helping hand to the realm of fruit tree breeding and orchard management. It paves the way for the cultivation of fruit trees that are not only more productive but also more resilient, adapting to diverse environmental challenges, and yielding bountiful, high-quality fruits. This intricate weave of genomic information and trait correlations beckons further research to decipher the complex puzzle of photosynthesis regulation and functionality, revealing the intricate interplay of genetics in shaping these essential traits.
-
The authors confirm contribution to the paper as follows: study conception and design: Jia Z, Santiago L, Roose ML; data collection: Hiraoka Y, Roose ML, Santiago L, Rangel AA, Vieira TO, Kahn TL; analysis and interpretation of results: Wang X, Traband R, Ferrante SP, Yu L, Jia Q, Wang Z, Wang S, Jia Z, Roose ML, Santiago L; draft manuscript preparation: Jia Z, Wang X, Traband R, Santiago L, Roose ML. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study, incluidng phenotypic data (Supplemental Table S1) and genomic data (Supplemental Table S4), are included in this published article and its supplementary information files.
This project was supported by the following grants: USDA NIFA FACT grant 2019-67022-29930 to Zhenyu Jia and Mikeal L Roose.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xuesong Wang, Ryan Traband
- Supplemental Table S1 Characteristics of 71 citrus accessions and their 37 phenotypic traits related to leaf photosynthesis. This table presents each accession along with detailed phenotype data that quantifies aspects of photosynthetic efficiency and capacity.
- Supplemental Table S2 Identification of 125 candidate SNPs in citrus genome with corresponding chromosomal positions. This table lists SNPs identified as potentially significant, providing their exact locations within the genome to facilitate further genetic analysis.
- Supplemental Table S3 Comprehensive listing of all candidate genes identified in the study. This table includes names, loci, and functional annotations for each gene considered relevant to the traits studied, aiding in further genetic and functional studies.
- Supplemental Table S4 Genotypic data for citrus accessions presented in HapMap format. This table provides detailed genotype information for each accession, formatted to align with HapMap standards for ease of comparison and analysis.
- Supplemental Table S4
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang X, Traband R, Hiraoka Y, Ferrante SP, Yu L, et al. 2024. Revealing genetic determinants of photosynthesis-related traits in citrus via genome-wide association studies. Fruit Research 4: e020 doi: 10.48130/frures-0024-0013 

Revealing genetic determinants of photosynthesis-related traits in citrus via genome-wide association studies
- Received: 22 November 2023
- Revised: 17 February 2024
- Accepted: 18 March 2024
- Published online: 23 May 2024
Abstract: Photosynthesis-related traits, encompassing aspects such as absorbance, chlorophyll content, fluorescence, quenching ratio of incoming light, and photochemical efficiency, are pivotal characteristics directly related to photosynthesis carbon gain with important implications for overall tree physiology and fruit production. In this study, we conducted an examination of 71 citrus accessions and varieties, utilizing over 56,000 single nucleotide polymorphisms and 37 leaf reflectance parameters. Pairwise correlation analysis revealed clusters of interrelated traits organized into four distinct groups. The observed trait variations were found to have associations with the citrus phylogeny. Through genome-wide association studies, we identified a total of 125 genomic loci and 189 potentially associated genes responsible for the observed trait variations. The potential network and biological pathways influencing the variability of these biophysical variables within the citrus collection are also discussed.
-
Key words:
- Genetic /
- Photosynthesis /
- Citrus