









-
Cold and drought stresses are key abiotic environmental factors that strongly affect plant growth, development, and yield[1,2]. When plants are subjected to cold and drought stresses, it will cause an imbalance of free radical metabolism in cells, leading to the excessive accumulation of reactive oxygen species (ROS)[3]. And then leads to oxidative stress, which can cause oxidative damage to proteins, lipids, and pigments[4], and even lead to plant death. Therefore, it is of significance to research the effects of cold and drought stresses on plant growth and development and improve plant tolerance to cold and drought stresses.
A series of plant responses to environmental stress were regulated by transcription factors, which receive stress signals and modulated the expression of stress-related functional genes. The pathway 'CBF-COR' among many stress regulation pathways was the focus of researchers. CBF (C-repeat binding transcription factor), also known as DREB1, is a class of plant-specific transcription factors activated by cold stress or ICE. Plant cold-responsive (COR) genes can produce cold regulatory proteins to improve plant cold tolerance. The CCGAC sequence of CBF can bind to the CRT/DRE cis-element in the COR gene promoter to activate the cold-resistance response. Tobacco NtbHLH123 confers tolerance to cold stress by regulating the NtCBF pathway and reactive oxygen species homeostasis[5]. The bHLH transcription factor ICE1 induced the expression of CBF3/DREB1A and COR genes under cold stress by combining with the cis-acting element MYC (CANNTG) of their promoter regions, to improve the tolerance of transgenic plants to stress[6]. MaNAC1 from banana (Musa acuminata) is directly bound to the MaICE1 promoter to target MaCBF1 and enhance the cold tolerance of banana plants through a CBF-dependent pathway[7]. Overexpression of wild rice OrbHLH2, which is highly homologous to ICE1, induces the up-regulation of salt stress-related genes such as DREB1A/CBF3, COR15A, and enhances the tolerance to salt stress in transgenic plants[8]. Members of the bHLH gene family might regulate environmental stress response via the CBF-dependent pathway. However, it is not clear whether bHLH93, which belongs to the same bHLH IIIb subgroup is also involved in the CBF pathway playing a role in defense against abiotic plant stresses.
Apple is one of the most important economic fruit trees in the world. So far, there has been a certain basis for the research on the stress tolerance regulation mechanism of the bHLH gene in cultivated apple (Malus domestica). The MdCIbHLH1 protein binds to the promoter of MdCBF2 to up-regulate the expression of MdCBF2, which contributes favorably to the cold tolerance of transgenic apple plants[9]. Under cold conditions, MdbHLH4 represses the expression of MdCBF1 and MdCBF3 by directly binding to their promoters. MdbHLH4 also interacts with MdCICE1L, a homolog of AtICE1 in apple, and represses the binding of MdCCE1L to the MdCBF1/3 promoter, which inhibits its expression negatively regulating cold tolerance in plants[10]. MdABI4 positively regulated apple cold tolerance by interacting with MdICE1, which activated the downstream cold stress-crucial gene MdCBF1. While MdJAZ1/2 interferes with the interaction between MdABI4 and MdICE1 to negatively regulate apple cold tolerance. Therefore, MdABI4 accurately regulates cold tolerance of apple plants by integrating JA and ABA signals to form a JAZ-ABI4-ICE1-CBF cascade pathway[11]. Similarly, overexpression of bHLH transcription factor MdbHLH33 increased the expression levels of cold stress-related genes MdCBF2, MdCOR15A-1, and MdCOR15A-2 in apple calli, and MdbHLH33 could bind to LTR cis-acting elements of MdCBF promoter to positively regulate low-temperature stress[12].
Wild apple (Malus baccata) as an important germplasm resource of apple tolerance is rich in valuable resistance genes, which is of great significance for apple resistance breeding. MxNAS1 contributes to transgenic tobacco plants' tolerance to Fe stress by increasing the plant's antioxidant capacity and MbCBF1 also contributes to plant tolerance against cold and salt stress[13,14]. In the present study, another cold- and drought-induced transcription factor MbbHLH93 was identified and cloned. Phylogenetic and structural analysis revealed that MbbHLH93 is a typical bHLH family protein. To determine the function of MbbHLH93 in plant tolerance to stress, the MbbHLH93-overexpressed heterologous lettuce (Lactuca sativa) was obtained to investigate its effect on plant tolerance. The results confirm that MbbHLH93 functions to enhance the cold and drought tolerance of lettuce. Heterologous expression of MbbHLH93 up-regulated the expression levels of its downstream stress response crucial genes LsCBFs and increased the contents of chlorophyll and proline as well as the activities of antioxidant enzymes but reduced the accumulation of MDA, H2O2, and O2·− in lettuce leaves, alleviating cold and drought damage to lettuce plants. The present study provides important candidate genes for further study of plant tolerance regulation and lays a foundation for the genetic improvement of tolerance plants.
-
Apple plants (Malus baccata) from previous research[15] were precultured in Hoagland hydroponic solution at 25 °C and 80% relative humidity for a 16-h light/8-h dark cycle. The nutrient solutions were replaced every 3 d during the experiment. A total of 50 seedlings of M. baccata displaying similar growth (well-developed roots and 10-12 leaves) were separated into five groups for control, cold (4 °C), heat (37 °C), salt (200 mM NaCl), and drought (20% PEG6000) treatments for 12 h[16]. Roots and new leaves of various treatments were sampled at 0, 1, 3, 6, 9, and 12 h.
The Hong Kong iceberg lettuce (Lactuca sativa L.) was used in the present study. Lettuce seeds were disinfected with 75% ethanol and 10% sodium hypochlorite solution and seeded in 1/2 MS (Murashige & Skoog) solid medium. After 1 d of vernalization at 4 °C, then grown at 25 °C and 70% relative humidity for a 16-h light/8-h dark cycle.
Cloning and characterization of MbbHLH93
-
Total RNA was extracted from the leaves of M. baccata using an OminiPlant RNA Kit (Kangweishiji, Beijing, China) and the cDNA synthesis was conducted with HiFiScript gDNA Removal RT MasterMix (Kangweishiji, Beijing, China). MbbHLH93 was cloned with the specific primer shown in Supplementary Table S1.
The protein sequences of MbbHLH93 (M. baccata) were obtained according to the MdbHLH93 (Malus × domestica), and were retrieved from the Genome Database for Rosaceae (GDR) (
www.rosaceae.org ). MbbHLH93 were used as query proteins to identify their homologous proteins in pear (Pyrus communis), peach (Prunus persica), and black raspberry (Rubus occidentalis) with a protein-protein BLAST (BLASTp) in GDR[17,18]. Structural motif annotation of MbbHLH93 was analyzed via the MEME program (http://meme-suite.org/ ). Phylogenetic analyses of MbbHLH93 orthologous proteins from these four Rosaceae species were conducted via MEGA6.0 software (www.megasoftware.net ) by the maximum likelihood method based on 1,000 bootstrap replicates. Subsequently, the physiological and biochemical properties of tested proteins were conducted with the ProtParam online website (https://web.expasy.org/protparam/ ).Subcellular localization of MbbHLH93
-
The coding sequences (CDS) of MbbHLH93 without stop codon were inserted into the BamH I and Sal I sites downstream of GFP in the pSAT6-GFP-N1 vector, obtaining the fusion expression vector pro35S::MbbHLH93::GFP. Subsequently, the fusion expression vector and pSAT6-GFP-N1 empty vector were respectively transformed into tobacco leaves for 24 h according to a previous study[19]. 0.5 μg/mL 4,6-diamidino-2-phenylindole (DAPI) (Solarbio, China) served as cell nuclear dye. GFP and conjugate of DNA and DAPI fluorescence were observed respectively via a confocal laser scanning microscope (LSM 710; Carl Zeiss) at the wavelengths of 488 nm and 405 nm.
Expression analysis of MbbHLH93
-
The expression analysis of MbbHLH93 was performed via qRT-PCR reactions based on a previous description[20]. Each reaction involved three biological replicates for error minimization, and the transcript level of MbbHLH93 was determined via the 2−ΔΔCᴛ method with Actin (EB127077) as an internal reference. Primer sequences crafted for this experiment are shown in Supplementary Table S1.
Gene transformation
-
To obtain MbbHLH93 transgenic lettuce, the CDS of MbbHLH93 was introduced into pCAMBIA2300 vectors under the control of the CaMV35S promoter, constructing the MbbHLH93 overexpression vector. Genetic transformation of the lettuce plant was performed by the Agrobacterium-mediated leaf disk method[21]. The positive transgenic lettuce plants were screened in a selection medium containing kanamycin and identified by semi-quantitative RT-PCR analyses. The homozygous T2 generation plants were used for subsequent experiments.
Stress treatments of lettuce plants and physiological measurements
-
A total of 30 seedlings of 25-day-old lettuce displaying similar growth were separated into three groups for control, cold (−7 °C for 6 h), and drought (no-watered for 7 d) treatments and lettuce plants were subjected respectively to 6 h and 7 d of low temperature and drought stress were collected for subsequent determination of genes and physiological indexes related to low-temperature response. The plant phenotype and survival rates were analyzed after returning the stressed lettuces to normal growth for 15 d.
The content of chlorophyll, free proline, malondialdehyde (MDA), relative electrolyte leakage (EL), active oxygen (H2O2 and O2·−), and the enzyme activity of superoxide dismutase (SOD), peroxidase POD, and catalase (CAT) were measured respectively based on previous research[22,23]. Each reaction involved three biological replicates for error minimization.
Detection of key stress-related genes
-
Total RNA extraction and cDNA synthesis from the leaves of cold- and drought-stressed lettuce refer to the methods above. The qRT-PCR was performed to analyze the expression levels of stress-related LsCBF genes in control and transgenic lettuce, with LsIPP2 and LsEIF2 as internal references[24,25]. Primer sequences crafted for this experiment are shown in Supplementary Table S1.
Statistical analysis
-
The experimental data were processed using IBM SPSS Statistics 21 software. One-way ANOVA followed by Tukey's multiple range test (p < 0.05) was used to compare significant differences between all the groups of each data set, and values are shown as the mean ± standard deviation (SD) of biological triplicates.
-
The coding region of MbbHLH93 contains 1,428 bases encoding a predicted protein of 498 amino acids (aa). Physiological and biochemical properties of MbbHLH93 indicated that MbbHLH93 protein is probably an acidic hydrophilic protein (Supplementary Table S2). MbbHLH93 contained conserved bHLH and ACT-like domains at the C-terminal, which were consistent with other tested family members in Rosaceae species, indicating MbbHLH93 belongs to the bHLH family gene (Fig. 1a). Further, phylogenetic analysis showed that bHLH93 of apple and pear are closely related (Fig. 1b).
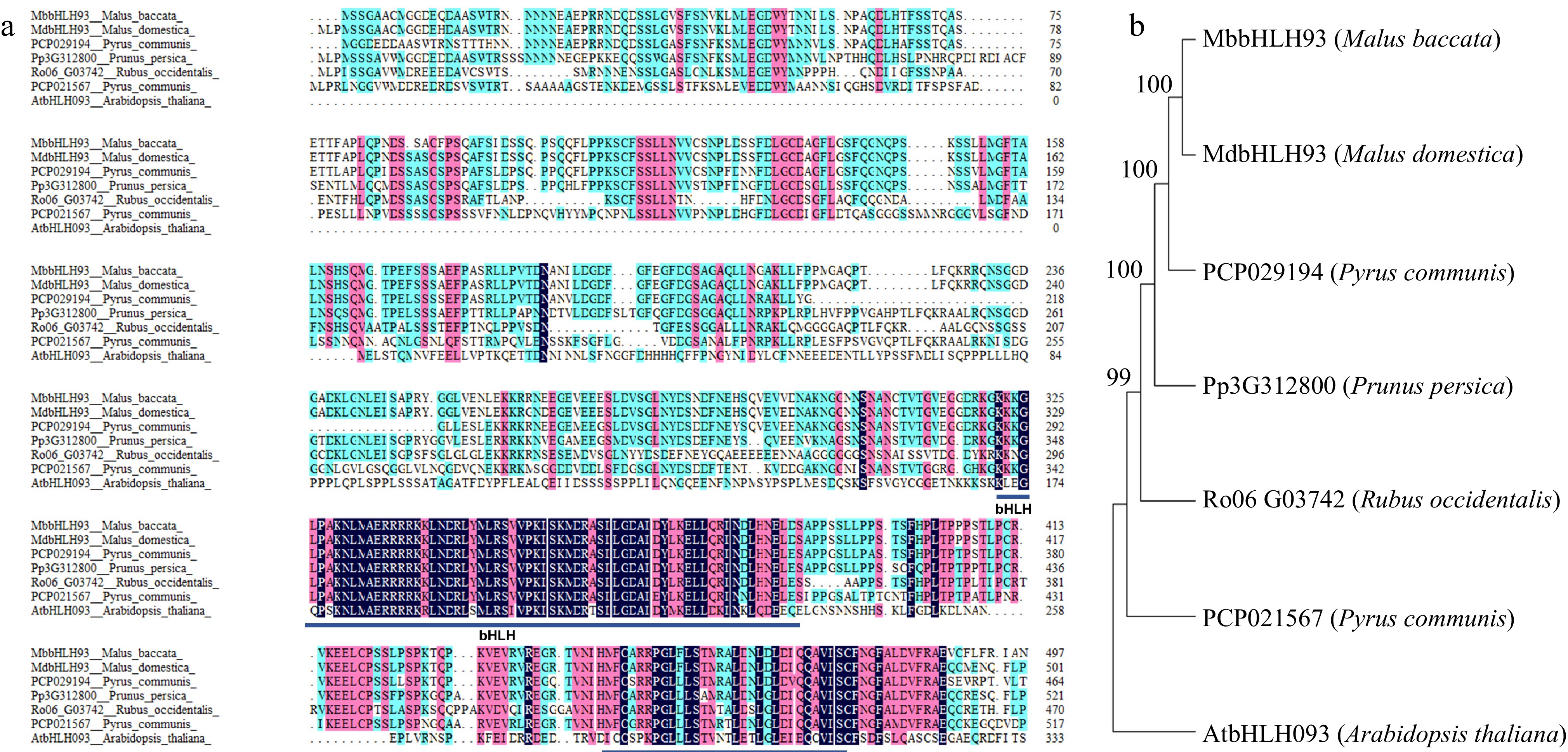
Figure 1.
Sequence alignment and phylogenetic analysis of bHLH93 in Rosaceae species. (a) Sequence alignment of MbbHLH93. The conserved amino acid residues are shown in purple. The bHLH and ACT-like domain are displayed respectively with rectangles in dark blue. (b) Phylogenetic analysis of MbbHLH93. The MbbHLH93 orthologous proteins from Arabidopsis and four Rosaceae species were obtained to build a phylogenetic tree via the maximum likelihood method based on the 1,000 bootstrap analyses of the MEGA6.0 software.
Subcellular localization of MbbHLH93
-
To characterize the biological role of MbbHLH93 in plant stress response, a MbbHLH93::GFP construct was transiently transformed into tobacco leaves to generate a fluorescently tagged MbbHLH93 protein for observation of protein subcellular localization. Green fluorescence was present in the whole cell of positive control, while only distinct fluorescence was observed in the nucleus (Fig. 2b, f), and the DAPI (4,6-diamidino-2-phenylindole) staining also confirmed that MbbHLH93 was targeted to the nucleus in vivo (Fig. 2a, e).
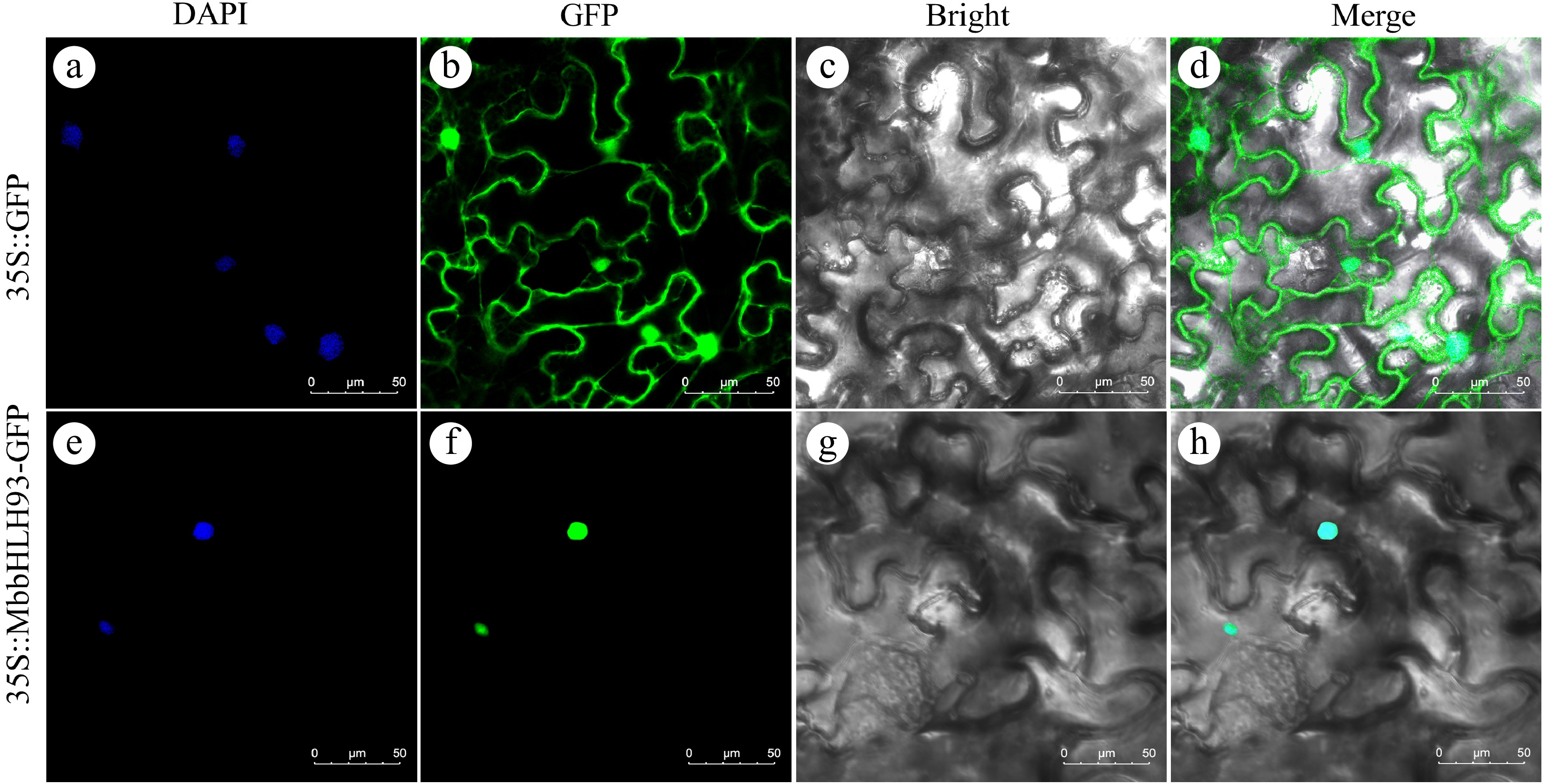
Figure 2.
Subcellular localization of MbbHLH93. 35Spro::MbbHLH93::GFP was expressed transiently into tobacco leaves with 35Spro::GFP as positive control. (a), (e) DAPI dyeing; (b), (f) GFP signals; (c), (g) bright field; (d), (h) merge. Scale bars: 50 μm.
Expression patterns of MbbHLH93 in various tissues of Malus baccata
-
To understand the expression patterns of MbbHLH93 genes in Malus baccata plants, qRT-PCR analysis was conducted for evaluating MbbHLH93 expression level in root, stem, new leaves, and mature leaves. The MbbHLH93 showed significantly different expression levels in different tissues, and a high expression amount was observed in new leaves and roots (Supplementary Fig. S1). Subsequently, the variation trend of MbbHLH93 gene expression was observed in roots and new leaves during the cold, drought, salt, and hot treatments. The results indicated that all four abiotic stresses could induce the expression of MbbHLH93, and with the extension of stress treatment time, the expression level of MbbHLH93 in new leaves and roots showed a trend of first increasing and then decreasing (Fig. 3). It was worth noting that cold and drought treatments could rapidly induce large amounts of expression of MbbHLH93 in the early stage of stress treatment. The highest expression abundance was detected in the roots induced by 6 h of low temperature and 3 h of drought, as well as in the new leaves induced by 9 h of low temperature and 9 h of drought (Fig. 3), indicating MbbHLH93 might play an essential role in plant low temperature and drought tolerance.
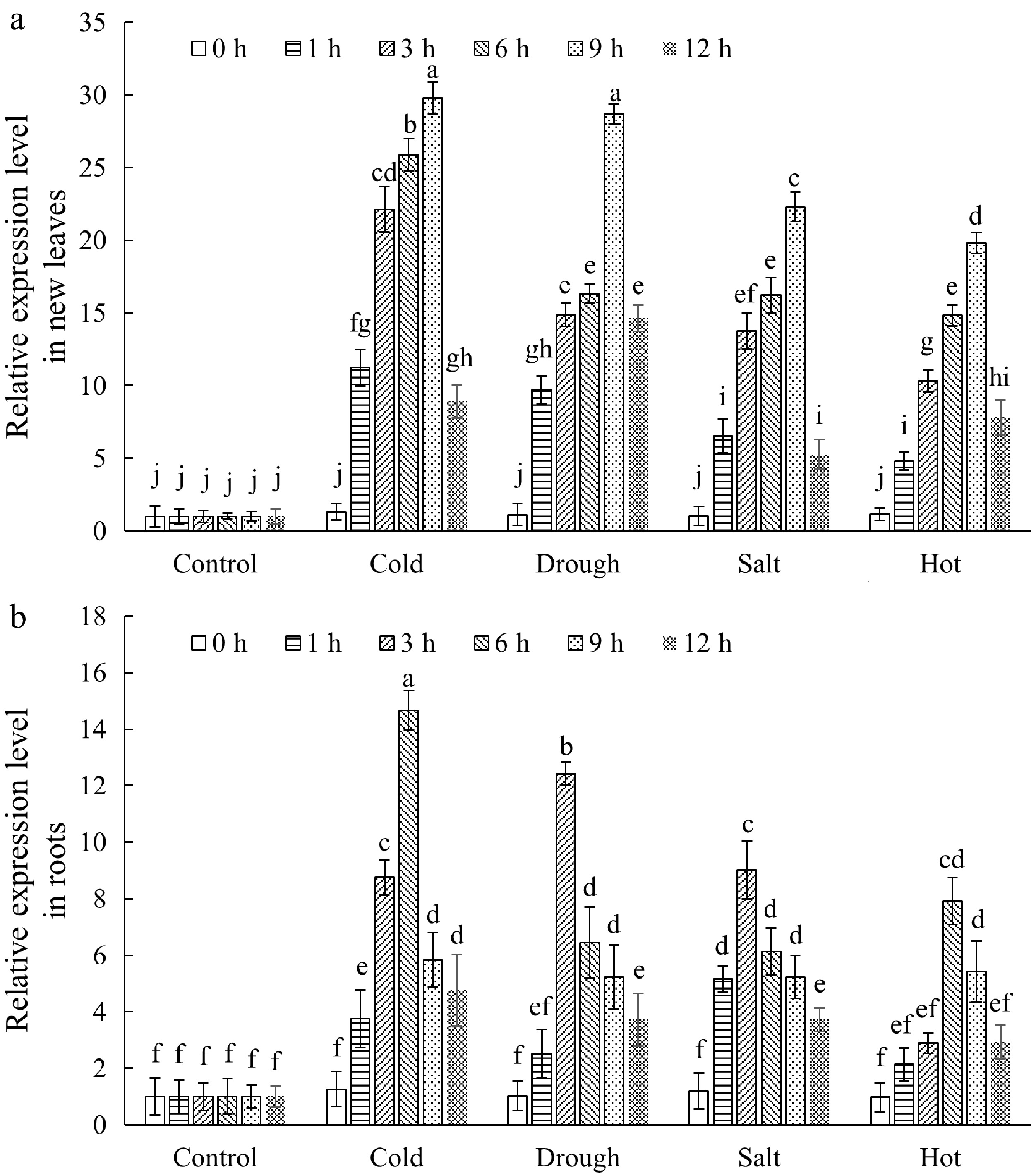
Figure 3.
The expression patterns of MbbHLH93 in Malus baccata under various stress treatments. (a) The expression levels of MbbHLH93 in new leaf under stress treatments. (b) The expression levels of MbbHLH93 in root under stress treatments. The leaf and root samples were collected 0, 1, 3, 6, 9, and 12 h after treatments. Relative expression levels of MbbHLH93 were calculated via the 2−ΔΔCᴛ method and values were shown as the mean ± standard deviation (SD) based on three repetitions. Lowercase letters displayed the significant differences at p < 0.05 (one-way ANOVA followed by Tukey's multiple range test).
Functional verification of MbbHLH93 in enhancing cold tolerance of lettuce
-
To determine the function of MbbHLH93 in affecting plant stress tolerance, the MbbHLH93 overexpression vector pCAMBIA2300-MbbHLH93 was transformed into lettuce plants. The lettuce exhibits a short life cycle and the necessity of cold-resistant production in northeast China[25]. Six MbbHLH93-overexpressed lettuce lines (L1, L2, L3, L7, L8, and L11) were identified via kanamycin tolerance and semi-quantitative RT-PCR analyses (Supplementary Fig. S2). Among them, three lines (L1, L7, L8) were screened for following research with the wild type (WT) and empty vector line (VL) transformed lettuces as controls.
Subsequently, the phenotype of control and MbbHLH93-overexpressed lettuce lines after being treated with low temperature (−7 °C for 6 h, and recovered at room temperature for 15 d) were analyzed. The results showed control and transgenic lettuces suffered from freezing injury to different extents under longer duration of cold stress, while, obviously more severe damage was observed in control plants. Most of the transgenic lettuce lines rather than control lines after cold stress were able to resume normal growth when they were transferred to room temperature (Fig. 4a). These results confirmed that overexpression of MbbHLH93 significantly improved plant adaptability to cold stress.
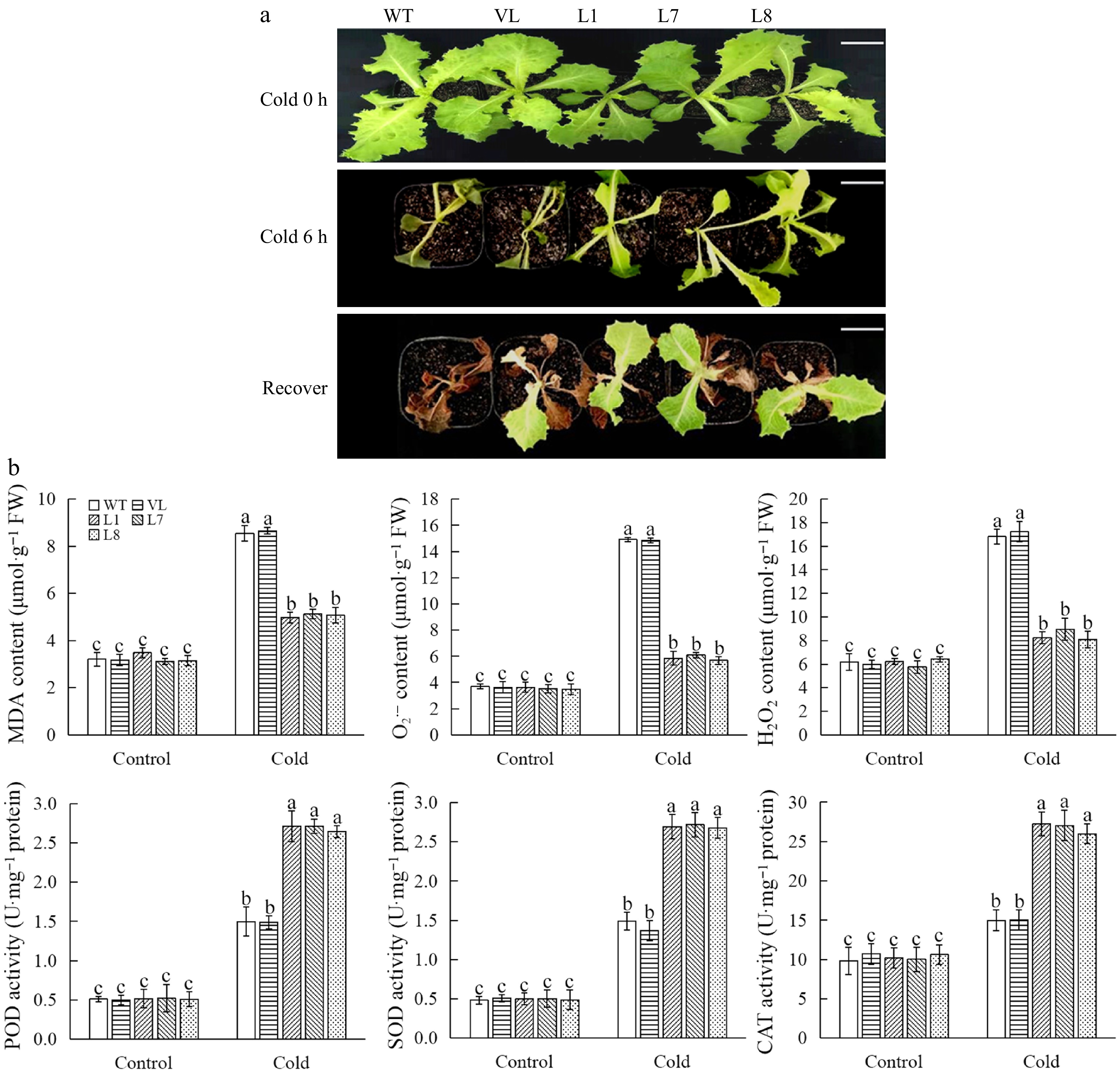
Figure 4.
MbbHLH93 enhances cold resistance of lettuce plants. (a) Phenotype of wild line (WT), vector line (VL), and MbbHLH93-overexpressed lettuce (L1, L7, L8) under cold for 0 h (control), cold for 6 h, and recovery for 15 d. Scale bar: 3 cm. (b) Determinations of physiological indices associated with cold stress in wild line (WT), vector line (VL), and MbbHLH93-overexpressed lettuces (L1, L7, L8) under cold for 0 h (control) and cold for 6 h. Values are shown as the mean ± standard deviation (SD) based on three repetitions. Lowercase letters show the significant differences at p < 0.05 (one-way ANOVA followed by Tukey's multiple range test).
Cold tolerance is closely related to a series of physiological parameters in plants[26,27]. To analyze the basis for the altered tolerance to stress in MbbHLH93-overexpressed transgenic materials, cold-related physiological indicators were analyzed. Under normal growth conditions, the relative electrolyte leakage of control and transgenic lettuce plants was similar (Supplementary Fig. S3). After low-temperature stress, electrolyte leakage of control was significantly higher than that of MbbHLH93-overexpressed lettuce lines, indicating that control plants were more subjected to cell membrane damage during low-temperature stress than transgenic plants. Proline content increased and chlorophyll content declined significantly in lettuce plants after experiencing low-temperature stress, while the contents of proline and chlorophyll in MbbHLH93-overexpressed strains were both significantly higher than that of control after cold stress (Supplementary Fig. S3). Similar to the change in chlorophyll, the overexpression of MbbHLH93 also significantly increase the antioxidant enzyme (SOD, POD, CAT) activities under cold stress (Fig. 4b). Additionally, low temperature caused an increased accumulation of H2O2, O2·− as well as MDA in lettuce cells, but the heterologous expression of MbbHLH93 in lettuces relieved their accumulation to some extent (Fig. 4b).
Functional verification of MbbHLH93 in enhancing drought tolerance of lettuce
-
To further explore the function of MbbHLH93 in affecting plant drought tolerance, a waterless stress assay was conducted for 7 d to assess the phenotype and survival rate of lettuces. In keeping with the results of cold stress, after drought treatment, control lines were subjected to more serious damage than transgenic lettuces. Most of the control seedlings nearly died when they were returned to room temperature for 7 d, while MbbHLH93-overexpressed transgenic lettuces could resume growth (Fig. 5a). These results displayed the drought tolerance function of MbbHLH93.
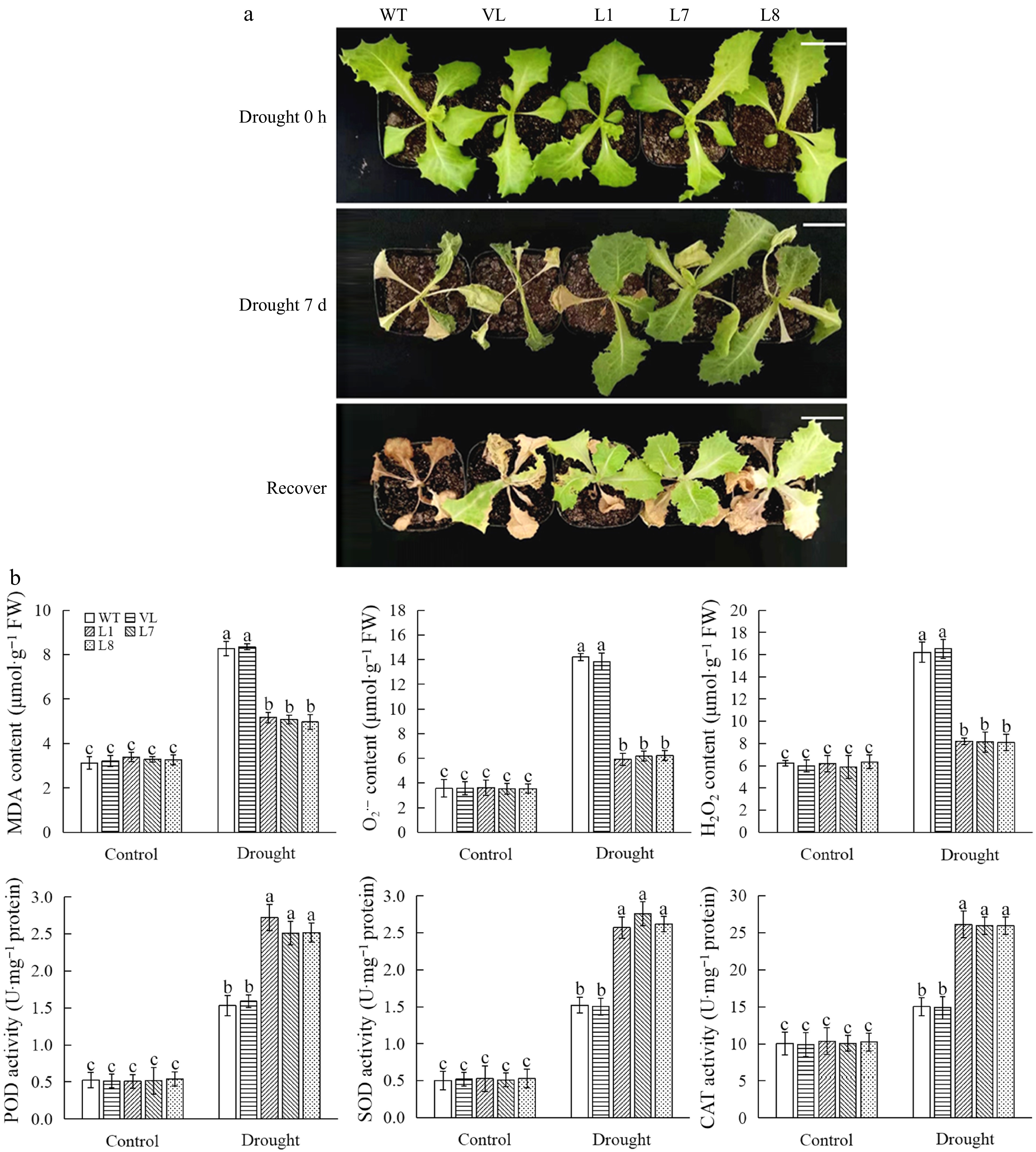
Figure 5.
MbbHLH93 enhances drought resistance of lettuce plants. (a) Phenotype of wild line (WT), vector line (VL), and MbbHLH93-overexpressed lettuces (L1, L7, L8) under drought for 0 h (control), drought for 7 d and recovery for 15 d. Scale bar: 3 cm. (b) Determinations of physiological indices associated with drought stress in wild line (WT), vector line (VL), and MbbHLH93-overexpressed lettuce lines (L1, L7, L8) under drought for 0 h (control) and drought for 7 d. Values are shown as the mean ± standard deviation (SD) based on three repetitions. Lowercase letters indicate the significant differences at p < 0.05 (one-way ANOVA followed by Tukey's multiple range test).
Moreover, drought-related physiological indicators were analyzed in control and transgenic lines. The results showed that drought stress led to an increase in the relative electrolyte leakage and proline, MDA, H2O2, and O2·− content as well as antioxidant enzyme activity, while a decline of chlorophyll content was seen. After drought treatment, MbbHLH93-overexpressed lettuces showed a lower relative electrolyte leakage and MDA content but higher proline and chlorophyll contents compared to the control (Supplementary Fig. S3). In addition, drought stress contributed to the activity of antioxidant oxidase increased more and the accumulation of reactive oxygen species (ROS) decreased in transgenic lettuce compared to the control (Fig. 5b).
Over-expression of MbbHLH93 enhanced the expression of the stress response gene
-
To genetically clarify if the effect of MbbHLH93 overexpression on plant tolerance is dependent on the classical CBF pathway, the expression levels of LsCBF genes were determined in transgenic lines. Low temperature induced the up-regulated expression of most LsCBF genes, except for LsCBF2 and LsCBF13. After low-temperature stress, the expression level of most LsCBFs in transgenic lettuce was significantly higher than that in control. Similarly, the expression levels of most CBF genes were positively induced after drought treatment, and drought treatment also significantly increased the expression level of LsCBFs in transgenic lettuce compared to the control (Fig. 6). These results indicated that MbbHLH93 could positively regulate the expression of LsCBF genes, thereby improving the cold and drought tolerance of plants.
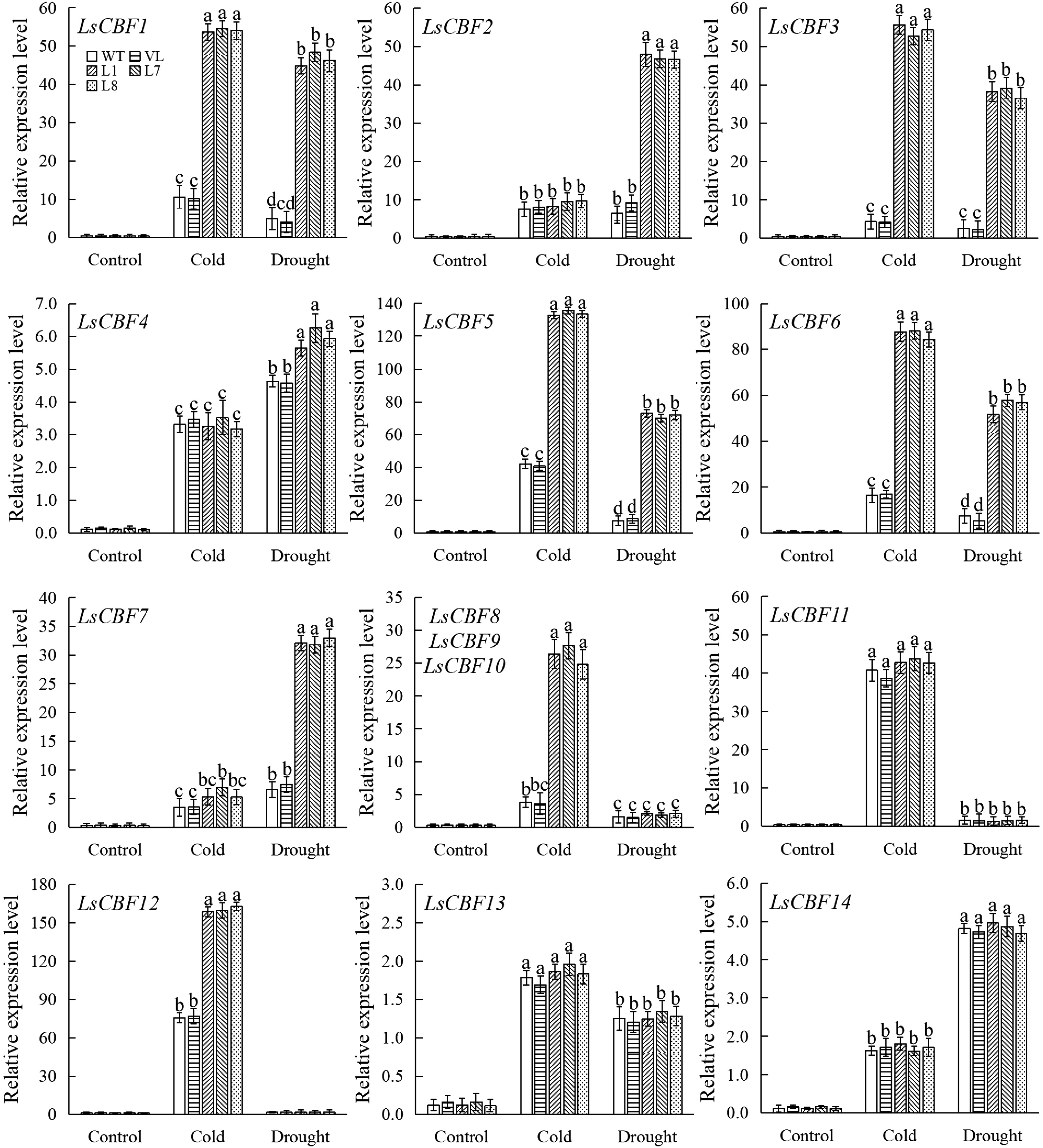
Figure 6.
Expression of LsCBF genes in wild line (WT), vector line (VL), and MbbHLH93-overexpressed lettuce lines (L1, L7, L8) after control, cold and drought stresses. LsCBF8, LsCBF9, and LsCBF10 were quantitatively amplified with the same set of primers. Relative expression levels of LsCBFs were calculated via the 2−ΔΔCᴛ method and values are shown as the mean ± standard deviation (SD) based on three repetitions. Lowercase letters indicate the significant differences at p < 0.05 (one-way ANOVA followed by Tukey's multiple range test).
-
Abiotic stress seriously damages plant growth and crop yield[28]. Transgenic technology has become a mature and rapid method to cultivate resistant crops with the development of biotechnology, which highly relies on the extraction and functional identification of key genes associated with stress regulation[29,30]. To date, researchers have demonstrated that bHLH family proteins play an important role in abiotic stress responses, but there are few studies on the anti-stress application of bHLH family proteins in wild apple (Malus baccata). In the present study, the wild apple bHLH family gene MbbHLH93 was cloned and transferred into lettuce to identify its biological function of regulating plant cold and salt tolerance. The analyses of gene structure and phylogenetics showed that MbbHLH93 was a bHLH family gene (Fig. 1a). Under low temperature and drought stress, the MbbHLH93 was significantly induced and the survival rate of MbbHLH93 transgenic lettuces was significantly higher than that of controls (Figs 4a & 5a), indicating that MbbHLH93 functioned in significantly enhancing plant low temperature and drought tolerance.
The bHLH transcription factors play crucial roles in various biological processes, such as plant flowering, pollen fertility, plant stomata, embryo and seed development, and abiotic stress response[31,32]. AtbHLH093, belongs to the ICE1 family bHLH-LZs, has a role in controlling flowering time and is required for apical meristem function[33,34]. Furthermore, overexpression of AtbHLH093 leads to a weak decrease in the number of mature stomatal phenotype[35]. Some studies have also pointed out that bHLH93 plays an important role in tolerance to plant stress. Ding et al. predicts that Prunus mume PmbHLH06 (bHLH93) can directly interact with PmbHLH38 (FBH4) thereby forming a dimer to function under low temperature stress[36]. Knockdown of bHLH93 in tobacco impaired disease tolerance by reducing the expression of the defense gene PDF1.2[37]. MdbHLH093 increases H2O2 accumulation and activates the SA signaling pathway in interaction with MdMYB116 to improve apple tolerance to powdery mildew[38]. Another recent study showed that MdbHLH093 in apple positively regulates dopamine accumulation through transcriptional regulation of MdTyDC conferring drought tolerance[39]. To further understand the function of MbbHLH93 in plant stress response, the expression levels of LsCBF genes, and a series of stress-related physiological indicators in MbbHLH93-overexpressed lettuce were analyzed, indicating that MbbHLH93 changed the stress-related physiological indicators in lettuce cells by up-regulating the expression of LsCBFs, thereby positively regulating the cold and drought tolerance of plants (Fig. 7).
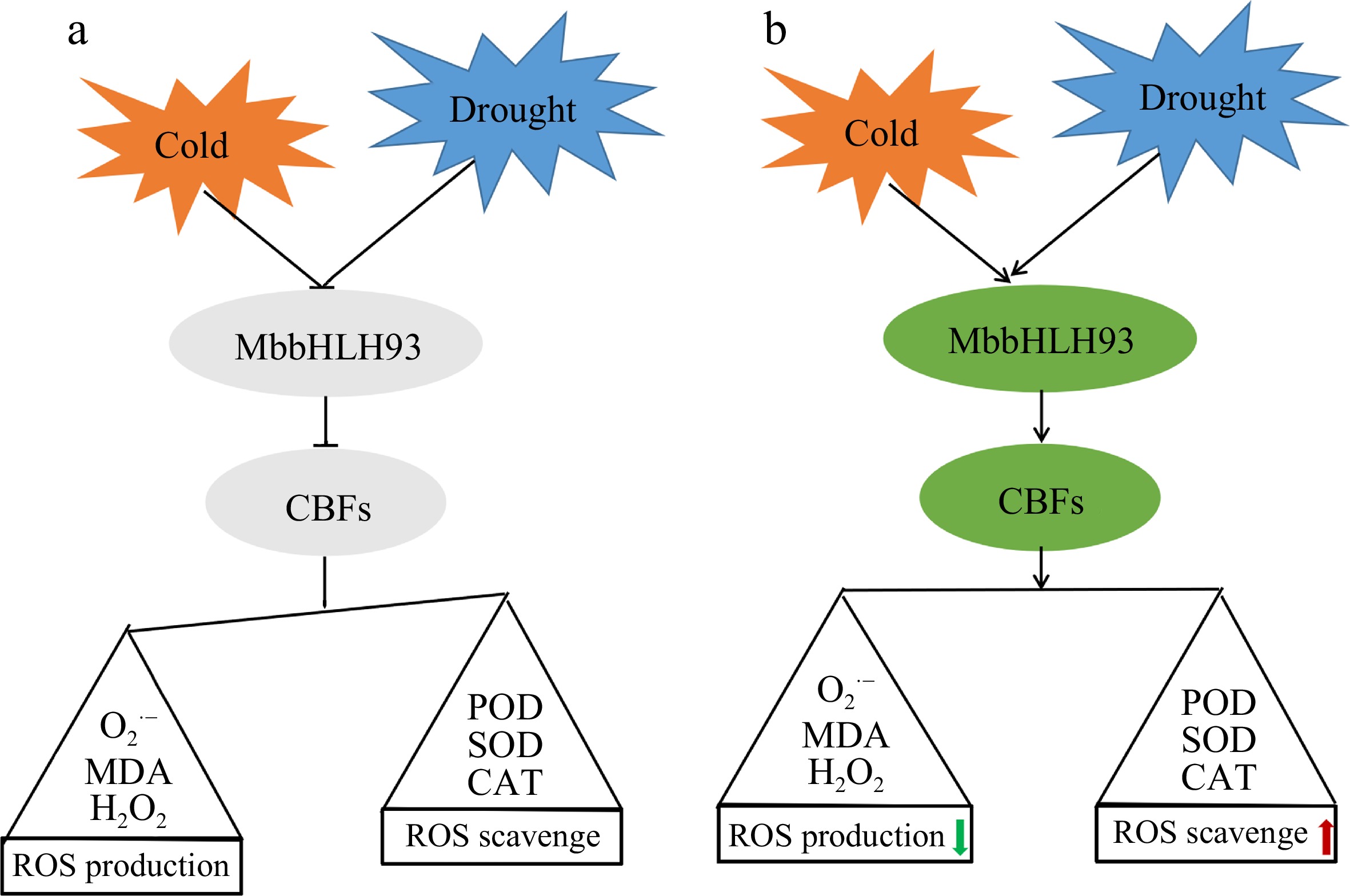
Figure 7.
Working model of MbbHLH93 function on the cold and drought adaptation in lettuce. (a) There is no MbbHLH93 in wild lettuce lines, and a large number of ROS accumulate in the cells under low temperature and drought treatment, threatening the normal growth of plants. (b) Cold and drought stress treatments rapidly induced large expression of transcription factor MbbHLH93 in MbbHLH93-overexpressed lettuce lines, and regulated lettuce plants' adaptation to stress through the classic CBF pathway. Primarily, membrane lipid peroxidation caused by large accumulation of reactive oxygen species in cells was reduced via improving the activities of antioxidant enzymes in transgenic lines, increasing plant stress resistance.
CBF cold response pathway plays a crucial role in the cold regulatory network[14,40]. Heterologous expression of AtCBF3 or AtCBF1 could increase the low-temperature tolerance of transgenic eggplant[41], potato[42], and petunia[43]. At low temperature, ICEs can bind to the MYC recognition site of the CBF gene promoter to enhance the expression of the CBF gene and its downstream stress-related target gene[44]. Heterologous overexpression of grapevine VabHLH1 and VvbHLH1 in Arabidopsis could induce the expression of AtCBF1, AtCBF2, AtCBF3, and other cold response genes[45]. In this study, MbbHLH93 could not induce the expression of CBF genes in lettuce plants grown at room temperature, which was consistent with previous findings[6]. After low temperature and drought treatment, the overexpression of MbbHLH93 induced significantly the up-regulated expression of a large number of LsCBFs gene in lettuce (Fig. 6), suggesting that MbbHLH93 could enhance cold tolerance by activating LsCBFs expression. Research conducted on different plants has shown variations in the response of CBF proteins to different environmental stresses. Cold-treated Arabidopsis plants showed higher expression of CBF1, CBF2, and CBF3 genes, in contrast to salinity and drought treatments[46]. While CBF1, CBF2, and CBF3 are more tolerance to drought, the CBF4 gene in grapes is typically activated by cold treatment[47]. In MbbHLH93 transgenic lines, LsCBF1/3/5/6 were significantly induced by both cold and drought, whereas LsCBF2/7 and LsCBF8/9/10/12 responded better to a single drought or cold (Fig. 6). This result suggested functional differences among different LsCBF genes in stress response and provided potential CBFs for the MbbHLH93 transgenic lines in response to drought and cold stress.
Abiotic stress will cause many physiological and biochemical changes in plant cells, such as electrolyte leakage and MDA content are indexes to assess membrane damage[15,48], and proline content affects osmotic regulation to protect proteins and cell membranes in response to environmental stresses[49,50]. SlbHLH96 and a bHLHm1 transcription factor gene MdSAT1 caused declined electrolyte leakage and MDA contents, and increased cold tolerance[51]. The overexpression of MdbHLH130 in tobacco also enhanced the plant tolerance via regulating the level of physiological indexes[52]. In accordance with the above results, overexpression of MbbHLH93 also decreased the electrolyte leakage and contents of MDA, but increased proline contents in lettuce plants after cold and drought treatment (Supplementary Fig. S3). It suggests that the higher tolerance of MbbHLH93 overexpression lines to low-temperature and drought may be attributed to the regulation of osmotic homeostasis, which may be one of the mechanisms of tolerance to abiotic stresses.
In addition, ROS accumulation was used to evaluate stress damage[53]. Excessive accumulation of ROS led to cell oxidative damage, affecting plant growth and development. The present study found that cold and drought stress induced excessive ROS accumulation in lettuce plants (Figs 4b & 5b). The lower H2O2 and O2·− accumulation in MbbHLH93 overexpressed lines indicated that these transgenic materials suffered less stress damage. These results were consistent with lower electrolyte leakage and MDA accumulation in transgenic plants[54]. Plants have evolved essential defense mechanisms, such as enzyme scavenging systems, aimed at preventing the excessive accumulation of ROS[55]. The enzyme activities of CAT, POD, and SOD in MbbHLH93 transgenic lettuces were significantly higher than those of the control under cold and drought stress (Figs 4b & 5b), indicating that MbbHLH93 can improve the activity of antioxidant enzymes in lettuce cells under stress, which is the main reason for less ROS accumulation in transgenic materials. SOD, as an important protective enzyme for ROS scavenging in plants, scavenged excess O2·− and directly mitigated O2·− induced damage to cell membranes in the MbbHLH93 transgenic line. CAT and POD were able to act synergistically to break down H2O2, and the increase in their activity in MbbHLH93 lines maximized the scavenging of potentially harmful ROS and protected the structure of the cell membrane. These results also indicated that MbbHLH93 could enhance the scavenging ability of ROS by regulating cell osmotic potential and enhancing plant tolerance to low temperature and drought.
-
In summary, the present study demonstrates the function of transcription factor gene MbbHLH93 in enhancing plant tolerance to cold and drought. MbbHLH93 was rapidly induced under low temperature and drought stress, and its overexpression up-regulated the expression of a series of LsCBFs, the key genes that respond to abiotic stress, in lettuce and probably regulated the physiological and biochemical changes of cells via the 'CBF' pathway to cope with stress response. Specifically, the overexpression of MbbHLH93 increased the content of intracellular osmotic regulatory substances and maintained the balance of cellular and external osmosis. Moreover, membrane lipid peroxidation caused by a large accumulation of ROS in cells was reduced via improving the activities of antioxidant enzymes in transgenic lines, increasing plant stress tolerance. These findings enrich our understanding of tolerance regulation of Malus baccata and lay a foundation for plant tolerance regulation and genetic improvement.
This research was supported by the National Natural Science Foundation of China (32172521), the Outstanding Youth Science Foundation of Heilongjiang Province (YQ2023C006), the China Postdoctoral Science Foundation (2023MD744175), and Modern Agricultural Industrial Technology Collaborative Innovation and Promotion System of Heilongjiang Province. The authors would like to thank Mr. Li Dalong from Northeast Agricultural University for providing the instruments.
-
The authors confirm contribution to the paper as follows: study conception and design: Han D, Huo J; data collection: Zhang L, Xu Y, Lv L, Wang T; analysis and interpretation of results: Zhang L, Liu W, Li X, Li W; draft manuscript preparation: Zhang L, Xu Y. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Lihua Zhang, Yu Xu
- Supplementary Table S1 Primers designed in the present study.
- Supplementary Table S2 Physiological and biochemical properties of test proteins. CDS, coding sequence; MW, molecular weight of the amino acid sequence; GRAVY, grand average of hydropathicity; pI, theoretical isoelectric point.
- Supplemental Fig. S1 The expression patterns of MbbHLH93 in root, stem, new leaf and mature leaf of Malus baccata. Relative expression levels of MbbHLH93 were calculated via the via 2−△△Ct method and values were shown as the mean ± standard deviation (SD) based on the three repetitions. Lowercase letters displayed the significant differences at p < 0.05.
- Supplemental Fig. S2 Genetic transformation and screening of MbbHLH93 overexpressed lettuce. (a) the genetic transformation process of MbbHLH93 overexpressed lettuce; including coculture stage, calli acquisition, resistant bud screening, rooting stage, transplanting of transgenic seedlings, and seed acquisition stage. (b) Identification of MbbHLH93-overexpressed lettuce lines by semi-quantitative RT-PCR analyses.
- Supplemental Fig. S3 Relevant physiological indicators of MbbHLH93 transgenic lettuces (L1, 7, 8) under cold and drought stress. Values were shown as the mean ± standard deviation (SD) based on the three repetitions. Lowercase letters displayed the significant differences at p < 0.05 (one-way ANOVA followed by Tukey's multiple range test).
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang L, Xu Y, Lv L, Wang T, Liu W, et al. 2024. MbbHLH93, a transcription factor associated with cold and drought tolerance in Malus baccata. Fruit Research 4: e038 doi: 10.48130/frures-0024-0032 

MbbHLH93, a transcription factor associated with cold and drought tolerance in Malus baccata
- Received: 04 September 2024
- Revised: 25 September 2024
- Accepted: 27 September 2024
- Published online: 02 December 2024
Abstract: The bHLH transcription factor is known to regulate cold signals and stress tolerance. In the present study, a new bHLH gene MbbHLH93, located in the nucleus, was isolated from Malus baccata, whose up-regulated expression were strongly induced by cold and drought treatment, and MbbHLH93-overexpressed heterologous lettuce plants displayed cold and drought stress-tolerant phenotypes. Determination of physiological and biochemical indexes associated with abiotic stress responses showed that overexpression of MbbHLH93 increased the activities of antioxidant enzymes superoxide dismutase, peroxidase, and catalase in lettuce plants treated with cold and drought stress, and decreased the contents of H2O2, O2·−, and malondialdehyde, which contributed to reducing cell membrane lipid peroxidation. Meanwhile, the accumulation of proline in transgenic plant cells increased, regulating cell osmotic pressure. Furthermore, quantitative expression analysis indicated that overexpression of MbbHLH93 improved the expression levels of LsCBFs, which were positive functional genes in response to cold and drought stress, enhancing plant tolerance. This research demonstrates that the MbbHLH93 is a key regulator in plant tolerance to cold and drought stresses, providing new knowledge for plant tolerance regulation.
-
Key words:
- Malus baccata /
- MbbHLH93 /
- Cold and drought stress /
- Transcriptional regulation