
-
In addition to being a crucial component of plant nucleic acids and membranes, phosphorus is also necessary for crop growth and development, physiological metabolism, yield, and quality through respiration, glycolysis, photosynthesis, signal transduction, enzyme activity regulation, and redox reactions in a variety of plant physiological processes[1]. Thus, the uptake, storage, and metabolism of phosphorus are critical for the growth and development of plants[2]. Phosphorus can also improve crop resilience and adaptability. Phosphorus improves the hydration of cellular structures and the viscosity and flexibility of protoplasmic colloids during drought, making protoplasm more resilient to dehydration[3]. However, there is very little inorganic phosphorus present in the soil that plants can directly absorb and utilize[4]. To maintain phosphorus homeostasis in the body, plants have evolved a complex set of strategies to adapt to phosphorus-deficient environments. These strategies include adjusting phosphorus uptake, transport, and utilization at the molecular level by sensing phosphorus level signals in the internal and external environments of plants[5]. Among these, the SPX gene family members are crucial to this process.
The SPX gene family members, which contain SPX structural domains (Pfam:PF03105)[6], are extensively distributed in eukaryotes. The SPX name is derived from a homologous amino acid sequence called the SPX structural domain, which is shared by SYG1 (suppressor of yeast gpa1), PHO81 (phosphorus-regulated cell cycle-dependent protein inhibitor 81), and XPR1 (xenotropic and polytropic retrovirus receptor1)[7]. The conserved hydrophilic domain of SPX is usually located at the N-terminus. Plant proteins with SPX domains can be categorized into four subfamilies based on their distinct C-terminal domains: SPX, SPX-MFS, SPX-EXS, and SPX-RING[8]. Different subfamily members have distinct roles in the signaling, absorption, and transport of phosphorus in plants[9].
SPX subfamily members play a predominantly negative regulatory role in the phosphorus starvation response in plants. OsSPX1 and OsSPX2 block PHR2 through interactions with PHR2 in rice (Oryza sativa) phosphorus-sufficient conditions, hence negatively regulating the start of downstream phosphorus starvation-responsive genes[10]. PHO1 (Phosphate 1) and its related genes encode the proteins that make up the SPX-EXS subfamily, which are mainly involved in the movement of phosphorus from plant components below ground to above ground[11]. MFS structural domain-containing proteins can participate in the phosphorus transport pathway between the vacuole and the cytoplasm to preserve phosphorus homeostasis in plants. They are also widely involved in the transmembrane transport of chemicals and phosphorus translocation[12]. To maintain phosphorus homeostasis in plants under various phosphorus nutritional conditions, for instance, OsSPX-MFS1 and OsSPX-MFS2 in rice import phosphorus into the vacuole, whereas OsSPX-MFS3 transfers phosphorus out of the vacuole into the cytoplasm[13−15]. While phosphorus dynamic homeostasis is regulated by the SPX-RING subfamily's NLA (nitrogen limitation adaptation) in a nitrate-dependent manner, phosphorus homeostasis is regulated in vivo by modulating the phosphate transporter's degradation when phosphorus sufficiency is present[16].
The application of phosphorus fertilizers in agriculture is the primary source of phosphorus in soils in regions that are suitable for Vitis vinifera (V. vinifera) and Vitis amurensis (V. amurensis); yet, only roughly 10%–25% of phosphorus fertilizers are used seasonally[17]. The use of phosphorus fertilizers, which are frequently used in considerable quantities in conventional farming to boost yield and quality, is the most significant source of phosphorus in the soil[18]. However, because of its chemical makeup, phosphorus readily forms insoluble precipitates by adsorption, precipitation, or microbial absorption with soil particles like calcium, iron, and aluminum ions, as well as soil clay particles[19]. Therefore, the effective phosphorus contained in the soil is much lower than the phosphorus content required by plants[20]. Phosphorus deficiency in plants is often addressed by applying large quantities of phosphorus fertilizers to raise crop yields. However, in addition to eutrophying water bodies, this method overuses and depletes non-renewable resources like phosphate rock[21]. Thus, it is imperative that this issue be resolved. Numerous factors, such as variations in genotypes, individual plants, and plant organs, can impact a plant's capacity to absorb and use phosphorus. These factors can all result in variations in the uptake and utilization of phosphorus by plants[22,23].
As one of the oldest crops in the world, grapes are cultivated worldwide[24], one of the components required for the growth and development of plants is phosphorus[1], grapes require phosphorus to grow, and phosphate fertilizers are the main source of phosphorus in the soil. Nevertheless, they are only applied at a rate of roughly 10%–25%[17]. Additionally, aquatic bodies may become eutrophic due to excessive usage of phosphorus fertilizers[21]. Phosphate deprivation in Arabidopsis can be either positively or negatively regulated by the SPX gene family, which has been thoroughly researched in plants[25], the phosphate balance in tomatoes can be regulated by the SPX-PHR regulatory pathway[26], OsSPX1 in rice may be associated with cold stress and pi hunger[27], whereas the cucumber SPX gene family is involved in the pi response[28]. In eggplant, the SPX gene family is linked to auxin when there is a phosphorus deficit[29]. The uptake, transport, use, and distribution of phosphate and Pi signaling pathways are significantly influenced by the protein gene family that contains the SPX structural domain[30]. Vitis vinifera is a long-established and widely planted grape variety that can be used not only for winemaking but also for eating. Vitis amurensis is a grape variety that is widely distributed in northern China and can tolerate barrenness, but they do not have a high phosphorus utilization rate. Therefore, to solve the problem of low phosphorus fertilizer utilization in viticulture at the molecular level, it will be necessary to perform identification of the grape protein gene family throughout the entire genome that contains the SPX domain. In this study, we identified the genes of the SPX gene family in the genomes of two grape species, grapevine, and mountain grape, and explored their structures and further analyzed their expression patterns under phosphorus stress, to provide basic information for studying the gene functions of the SPX gene family in grapevine and mountain grape, using molecular approaches to alter the grapevine's response to phosphorus stress by mining genetic resources that are important in that response.
-
The plantlets of V. vinifera 'Pinot Noir' and V. amurensis 'Zuoyouhong' preserved in the laboratory of the College of Horticulture, Gansu Agricultural University (Gansu, China) were used as research materials. The plantlets were cultured on GS medium for 35 d. The GS liquid medium was chosen for this experiment in order to investigate the expression of SPX genes in grapes and mountain grapes under phosphorus stress. The concentration of all-phosphorus NaH2PO4·H2O in the GS formulation was 1 mM, with all-phosphorus serving as the control, 0 mM being low phosphorus, and 2 mM being high phosphorus. To maintain the same Na concentration in the medium for varying phosphorus concentrations of the GS medium, the same molar concentration of NaCl was substituted for NaH2PO4·H2O. Thirty five-day-old seedlings that had been carefully chosen and grown consistently were put in GS liquid media. During the day, the incubator was 25 °C with 16 h of light exposure, and at night, it was 22 °C with 8 h of darkness, all day with 60% humidity. The plantlets were treated with 0 mM (without phosphorus treatment, WP) and 2 mM (excessive phosphorus treatment, EP) phosphorus concentrations at 12 and 24 h, respectively. The treatment of 1 mM phosphorus for 0 h was set as a control (CK). Three biological replicates were set up for each treatment. For real-time fluorescence quantitative PCR (qRT-PCR) analysis, the collected plant leaves and roots were stored at −80 °C after being rapidly frozen in liquid nitrogen.
Physical identification and physicochemical property analysis
-
Full login numbers for the Arabidopsis SPX gene family are now available in the Arabidopsis database (
www.arabidopsis.org ). Using the login number, the Arabidopsis SPX gene family protein sequences were obtained from the NCBI database (www.ncbi.nlm.nih.gov ). The protein, full-length, and GFF3 files of Vitis vinifera were downloaded from Ensemblplants (http://plants.ensembl.org/index.html ) and Vitis amurensis were downloaded from the Grape (http://www.grapeworld.cn/ ). The obtained amino acid sequences of the Arabidopsis thaliana SPX gene family were performed homologous comparison with those of Vitis vinifera and Vitis amurensis using TBtools (TBtools-ll v2.096) to remove sequences less than 100 bp and duplicate sequences to obtain the initial protein sequences, and then the proteins were further screened using NCBI-CDD (www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ) to retain protein sequences containing the complete SPX structural domain. In the NCBI CDD website, the expected value and threshold was 0.01, the maximum number of hits was 500, the search mode selected was automatic, and the search against the database selected was CDD--62456 PSSMs.Protein's physical and chemical properties were analyzed by Protparam (
https://web.expasy.org/protparam/ ).Secondary structure and subcellular localization
-
Prabi (
https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_hnn.html ) was used to analyze the protein secondary structures online. The subcellular localization of SPX gene family proteins in V. vinifera and V. amurensis was predicted by WOLFPSORT (https://wolfpsort.hgc.jp/ ) online analysis. ChiPlot (www.chiplot.online ) was used to visualize the results of subcellular localization.Collinearity relationship and phylogenetic tree
-
The phylogenetic tree of the SPX gene family of Arabidopsis, V. vinifera, and V. amurensis was constructed on MEGA 7.0 by using proteins. The Evolview (
www.evolgenius.info/evolview ) was used to enhance the evolutionary tree.Conserved motifs, structural domains, and gene structure
-
The MEME (
https://meme-suite.org/meme/tools/meme ) online program was used to analyze the conserved motifs of V. vinifera and V. vinifera SPX gene families. Motif input 5, and TBtools software were used to draw protein-conserved motif maps. The structural domains were analyzed by using TBtools according to the GFF3 files of V. vinifera and V. amurensis. The online software GSDS 2.0 (https://gsds.gao-lab.org/ ) was used for gene structure analysis. Combined mapping of conserved motifs, structures, and gene structures was analyzed by using TBtools software.Cis-acting elements
-
The sequences of the first 2,000 bp of the SPX gene family promoters from V. vinifera and V. amurensis were extracted using TBtools software. Plant CARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) and New PLAC (www.dna.affrc.go.jp/PLACE/?action=newplace ) were online programs that were used to determine the types and quantity of cis-acting elements in the promoter region. The results of cis-acting elements were plotted on TBtools.Chip expression patterns
-
Tissue expression data for V. vinifera was obtained from The Bio-Analytic Resource for Plant Biology (
https://bar.utoronto.ca/ ), which covered various organs during various phases of the growth process of V. vinifera. The obtained data was further collated through Excel software and visually analyzed by using TBtools.Codon preference
-
CodonW 1.4.2 (
http://codonw.sourceforge.net ) was used to examine the codon use features of the CDS sequences of the VvSPX and VaSPX genes. Origin and TBtools were utilized to visualize the results.Expression analysis of the SPX gene family using RNA-seq
-
Extracting total RNA from roots and leaves at different phosphorus concentrations using the CTAB method. Primers (Supplementary Table S1) were synthesized from the website of Bioengineering (Shanghai) Co. The Prime Script RT reagent kit (Perfect Real Time) (TaKaRa) was used to accomplish cDNA synthesis. Using the SYBR Green I kit (Takara Biomedical Technology Co., Ltd., Beijing, China) and real-time fluorescence quantitative PCR (Mx3005p, Stratagene, La Jolla, CA, USA), the expression of each gene was measured. Real-time fluorescent quantitative polymerase chain reaction (qRT-PCR): using the Light Cycler@96 real-time quantitative PCR instrument, PCR amplification with designed primers, 3 μL of cDNA, 1 μL of each of the upstream and downstream primers, 10 μL of SYBR MIX, and 5 μL of ddH2O comprised the entire amplification reaction system, which had a volume of 20 μL. Stability of the GAPDH gene as an internal reference[31], the reaction process involved 95 °C predenaturation for 30 s; 95 °C denaturation for 10 s, 58 °C annealing for 30 s, 72 °C extension for 20 s, 40 cycles. The experiment was repeated three times. The process of the reaction, the melting curve, and the change curve of the fluorescence value were then analyzed. After normalizing the experimental data with the internal reference gene GAPDH, the 2−ΔΔCᴛ method was utilized to determine the relative expression[32]. The processed data were statistically analyzed using SPSS 22.0 software. The significance of group differences was assessed at the p < 0.05 level using the Duncan test and one-way analysis of variance (ANOVA). The use of high-quality and pure cDNA, appropriate PCR reaction conditions, correct standard curves and calibrations, and consistent methods and software for data analysis ensures reproducible and consistent results.
-
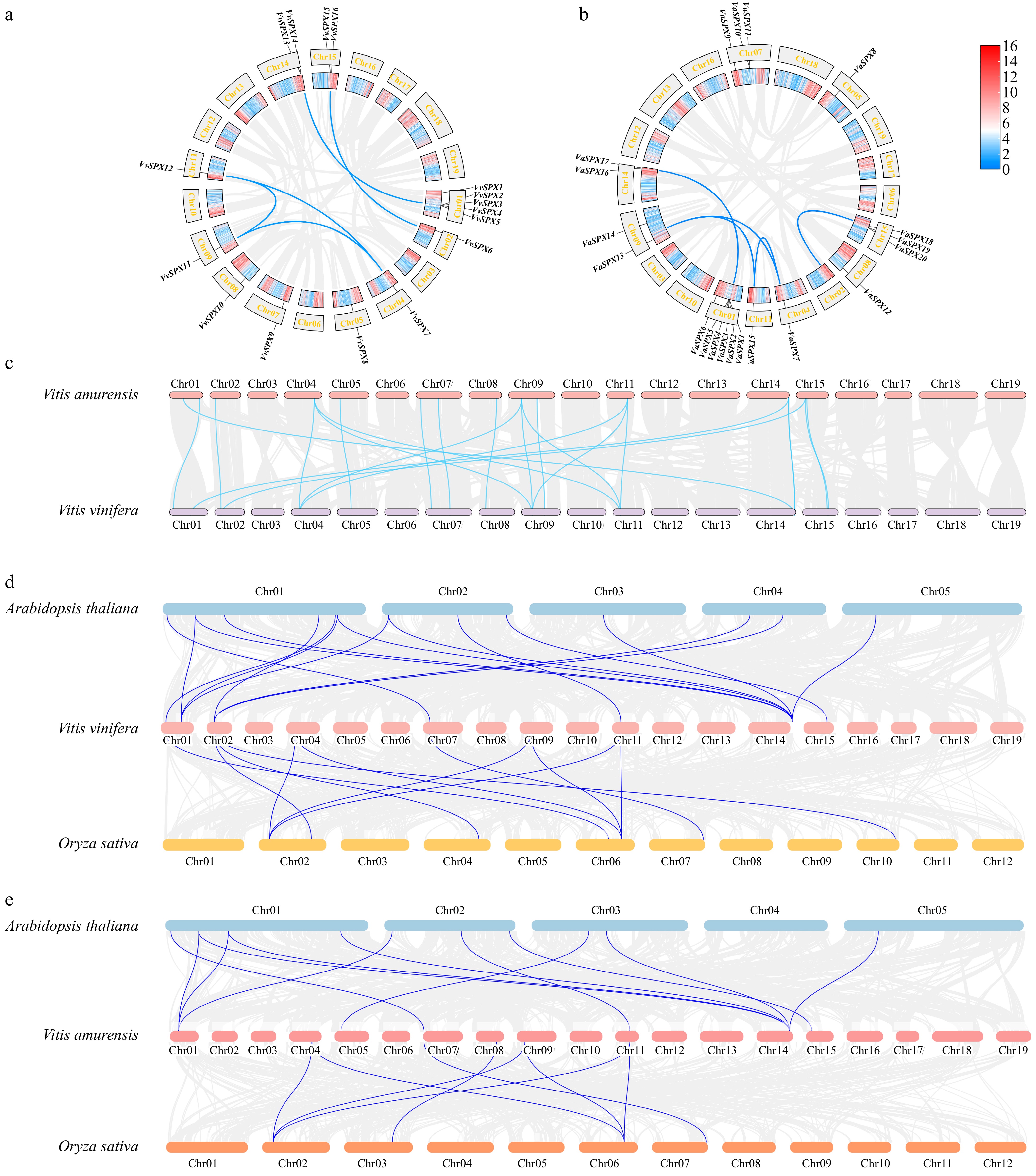
The genes were named sequentially based on their positions on chromosomes. A total of 16 VvSPX genes (VvSPX1 to VvSPX16) and 21 VaSPX genes (VaSPX1 to VaSPX21) were identified in this study. Chromosomal localization of the VvSPX genes showed that there were 5 VvSPX genes (VvSPX1, VvSPX2, VvSPX3, VvSPX4, VvSPX5) distributed on chromosome 1 and 2 genes distributed on chromosomes 14 and 15, respectively. Besides, there was only one gene distributed on chromosomes 2, 4, 5, 7, 8, 9, and 11, respectively (Fig. 1b). Chromosomal localization of VaSPX genes showed that (Fig. 1a) chromosome 1 contained six genes (VaSPX1, VaSPX2, VaSPX3, VaSPX4, VaSPX5, VaSPX6), and there were three genes distributed on chromosomes 7 and 15, respectively. In addition, there were two genes separately distributed on chromosomes 9 and 14, and only one gene distributed on chromosomes 4, 5, 8, and 11, respectively.
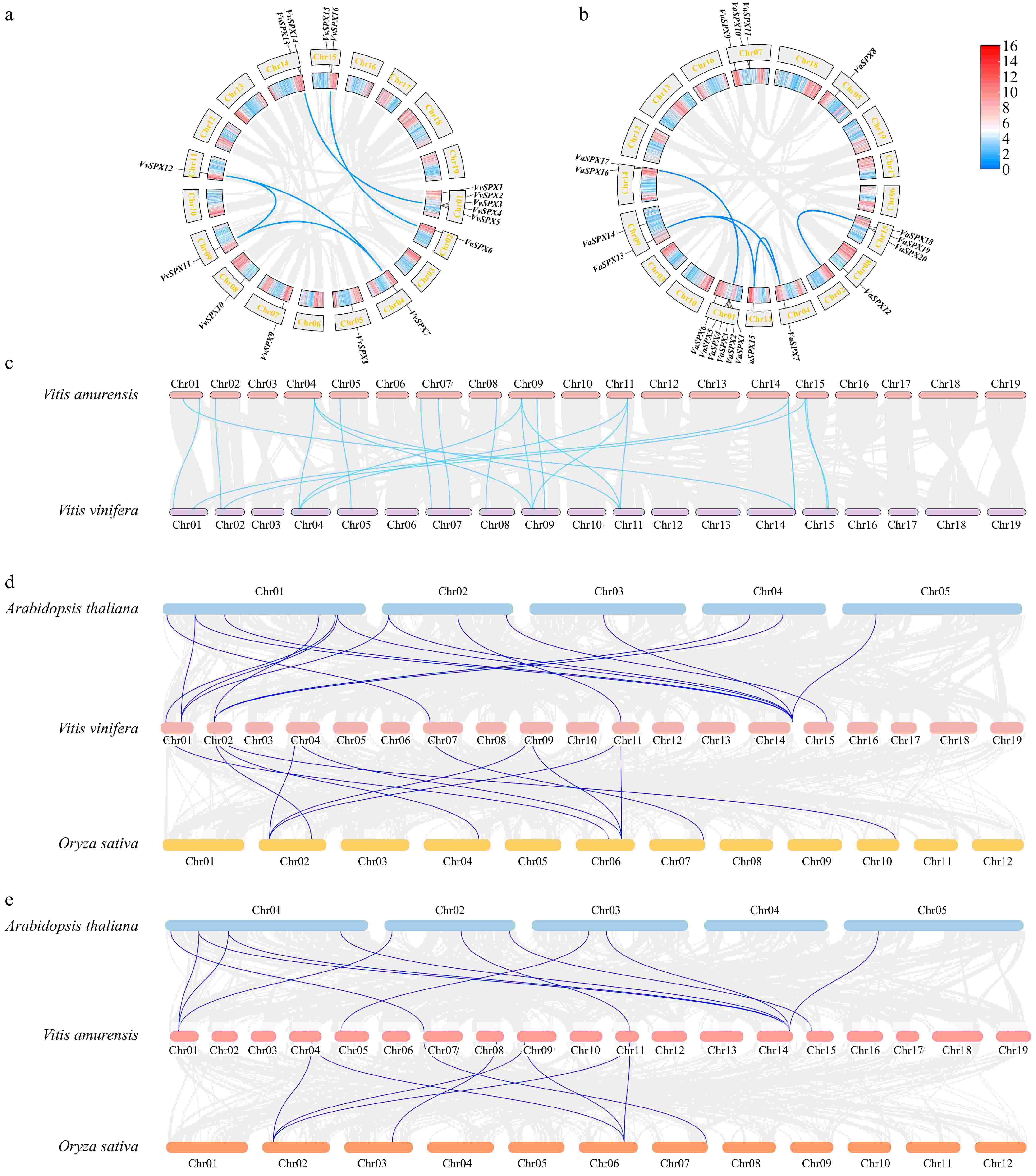
Figure 1.
Analysis of collinearity within VvSPX and VaSPX species. (a) Collinearity analysis within V. vinifera species. (b) Collinearity analysis within V. amurensis species. The gray line represents all collinear blocks in the V. vinifera and V. amurensis genomes, and the blue line represents gene pairs between VvSPX and VaSPX genes. (c) Colinearity analysis between V. vinifera and V. amurensis SPX genomes. (d) Colinearity analysis of SPX genes in V. vinifera, Arabidopsis, and Oryza sativa. Collinearity between the genomes of V. vinifera, Arabidopsis, and Oryza sativa is depicted by the gray lines in the backdrop. The blue lines show the collinearity between the VvSPX gene, Arabidopsis, and Oryza sativa. (e) Colinearity analysis of SPX genes in V. amurensis, Arabidopsis, and Oryza sativa. The gray lines in the background show the collinearity between the genomes of V. amurensis, Arabidopsis, and Oryza sativa. The blue lines show the collinearity between the VaSPX gene, Arabidopsis, and Oryza sativa.
Physicochemical properties analysis of VvSPX and VaSPX gene families
-
Physicochemical properties are important for the indepth study of functional differences and evolutionary patterns of gene families in organisms. Therefore, the physicochemical characteristics of SPX proteins were examined to comprehend their function, stability, isoelectric point, and hydrophilicity. The research in Supplementary Table S2 showed that the VvSPX proteins have between 182 (VvSPX8) and 802 (VvSPX4) amino acids. The molecular weights of the proteins ranged from 20,858.59 Da (VvSPX8) to 9,303,323.94 Da (VvSPX4), and the theoretical isoelectric points (pI) were between 4.92 (VvSPX13) and 9.46 (VvSPX3). The remaining proteins were acidic, while VvSPX1, VvSPX2, VvSPX3, VvSPX4, VvSPX5, VvSPX8, VvSPX9, VvSPX10, and VvSPX14 were basic proteins with isoelectric points greater than 7. The instability index ranged from 33.73 (VvSPX16) to 51.37 (VvSPX9). An instability index of less than 40 indicates stability, while an index of more than 40 indicates instability. The VvSPX9 protein had the most stability, while the VvSPX16 protein had the lowest. The range of the aliphatic index was 81.11 (VvSPX10) to 111.91 (VvSPX16). The grand average of hydropathicity, or GRAVY, is the ratio of the sum of the hydrophilicity values of all the amino acids in the sequence to the number of amino acids. Bigger negative values indicate better hydrophilicity, while bigger positive values indicate more hydrophobicity. VvSPX was mostly hydrophobic, only VvSPX6, VvSPX8, and VvSPX16 were hydrophilic.
According to an analysis of the physicochemical characteristics data (Supplementary Table S2), the VaSPX gene's amino acid residue count varied from 129 (VaSPX14) to 820 (VaSPX4). The molecular weights of the proteins were in the range of 14,870.21 Da (VaSPX20) to 95,386.81 Da (VaSPX4), and pI was between 4.96 (VaSPX16) and 9.48 (VaSPX5). VaSPX1, VaSPX7, VaSPX13, VaSPX15, VaSPX16, VaSPX18, and VaSPX19 were acidic proteins, and the rest were basic proteins. The range of the instability index was 32.95 to 52.52. The VaSPX14 protein was the most stable of all, whereas the VaSPX20 protein was the least stable. The range of the aliphatic index was 81.11 (VaSPX12) to 114.11 (VaSPX14). VaSPX was mostly hydrophobic, only VaSPX14 and VaSPX18 were hydrophilic.
Secondary structure and subcellular localization of VvSPX and VaSPX gene families
-
Different forms of secondary structures have distinct chemical and physical characteristics. Therefore, secondary structure prediction was performed for SPX protein. The secondary structure prediction revealed that all VvSPX proteins featured α-helices, random coils, and extended strands (Supplementary Table S3). The protein α-helix predominated, ranging from 46.15% (VvSPX8) to 60.72% (VvSPX4). Besides, the range of extended strand was 4.1% (VvSPX12) to 15.38% (VvSPX8), and the range of random coil was 27.01% (VvSPX16) to 43.79% (VvSPX13). The range of the VaSPX gene family α-helices was from 49.3% (VaSPX13) to 61.56% (VaSPX5). Besides, the range of extended strand was 4.78% (VaSPX15) to 15.02% (VaSPX13), and the range of random coil was 26.29% (VaSPX18) to 44.1% (VaSPX16).
Subcellular localization refers to the specific location of a protein within a cell. It is possible to comprehend a protein's function and connections within a cell by identifying its precise location. Therefore, subcellular localization of SPX proteins was performed. According to the subcellular localization prediction in V. vinifera and V. amurensis, the VvSPX family contained the greatest number of 12 genes in the plasma membrane, followed by 11 in the nucleus and six in the cytoplasm (Supplementary Fig. S1). There were 16 genes in the plasma membrane of the whole VaSPX family, followed by 13 in the nucleus and nine in the cytoplasm. Genes belonging to the SPX gene family were most abundantly distributed in the plasma membrane, followed by the nucleus and cytoplasm, according to predictions of subcellular localization in V. vinifera and V. amurensis.
Phylogenetic tree of VvSPX and VaSPX gene families
-
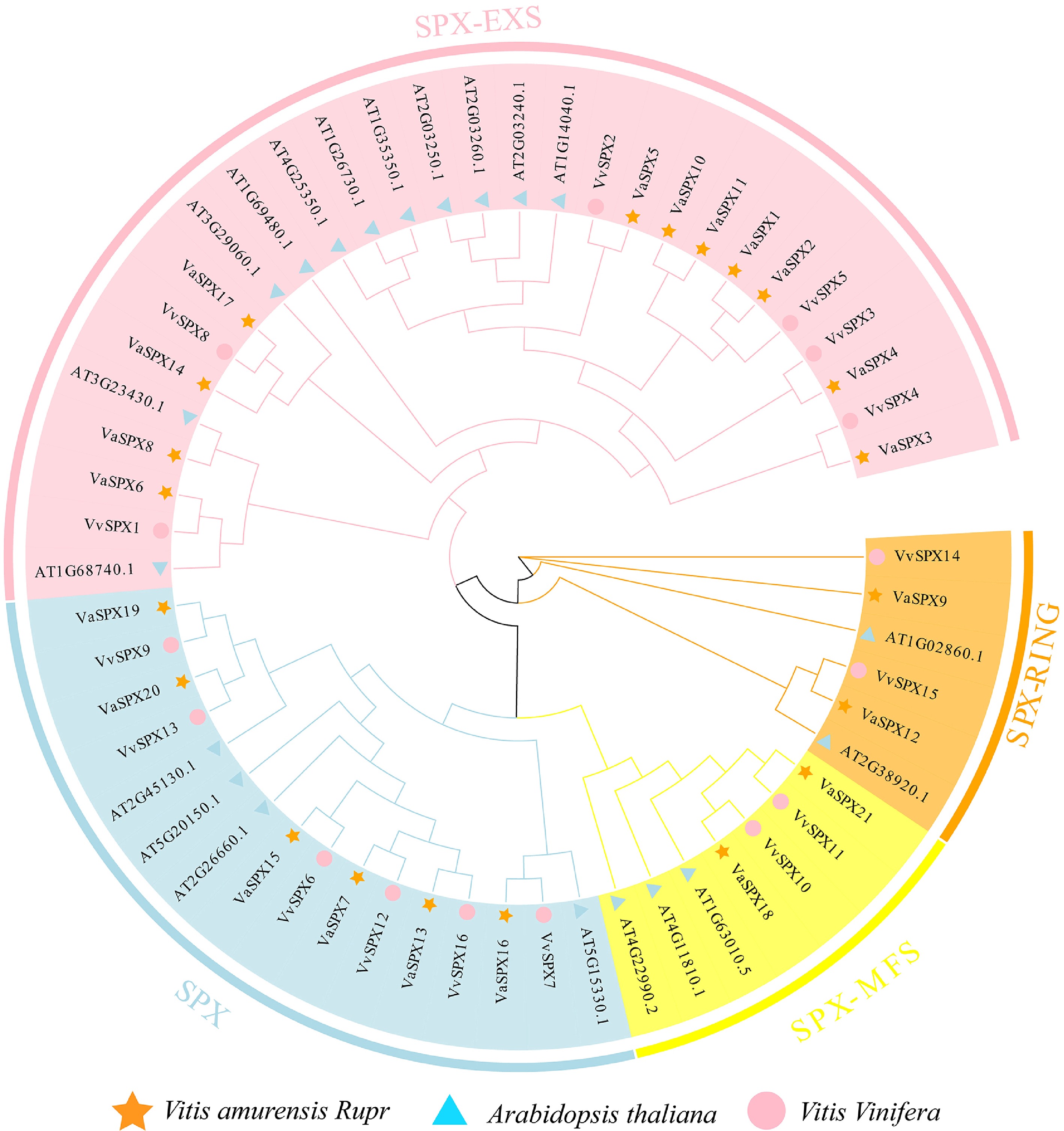
To elucidate the evolutionary relationships of SPX protein sequences in V. vinifera and V. amurensis, phylogenetic evolutionary trees were constructed through the SPX protein sequences of V. vinifera, V. amurensis, and Arabidopsis (Fig. 2). SPX-EXS, SPX, SPX-RING, and SPX-MFS were the four subfamilies into which 57 SPX protein sequences were determined to fall. Members of VvSPX and VaSPX proteins were distributed in all four subfamilies. The SPX-EXS subfamily contained six VvSPX and 11 VaSPX protein members. The SPX subfamily contained six VvSPX and six VaSPX protein members. The SPX-RING subfamily contained two VvSPX and two VaSPX protein members. The SPX-MFS subfamily contained protein members VvSPX10, VvSPX11, VaSPX18, and VaSPX21.
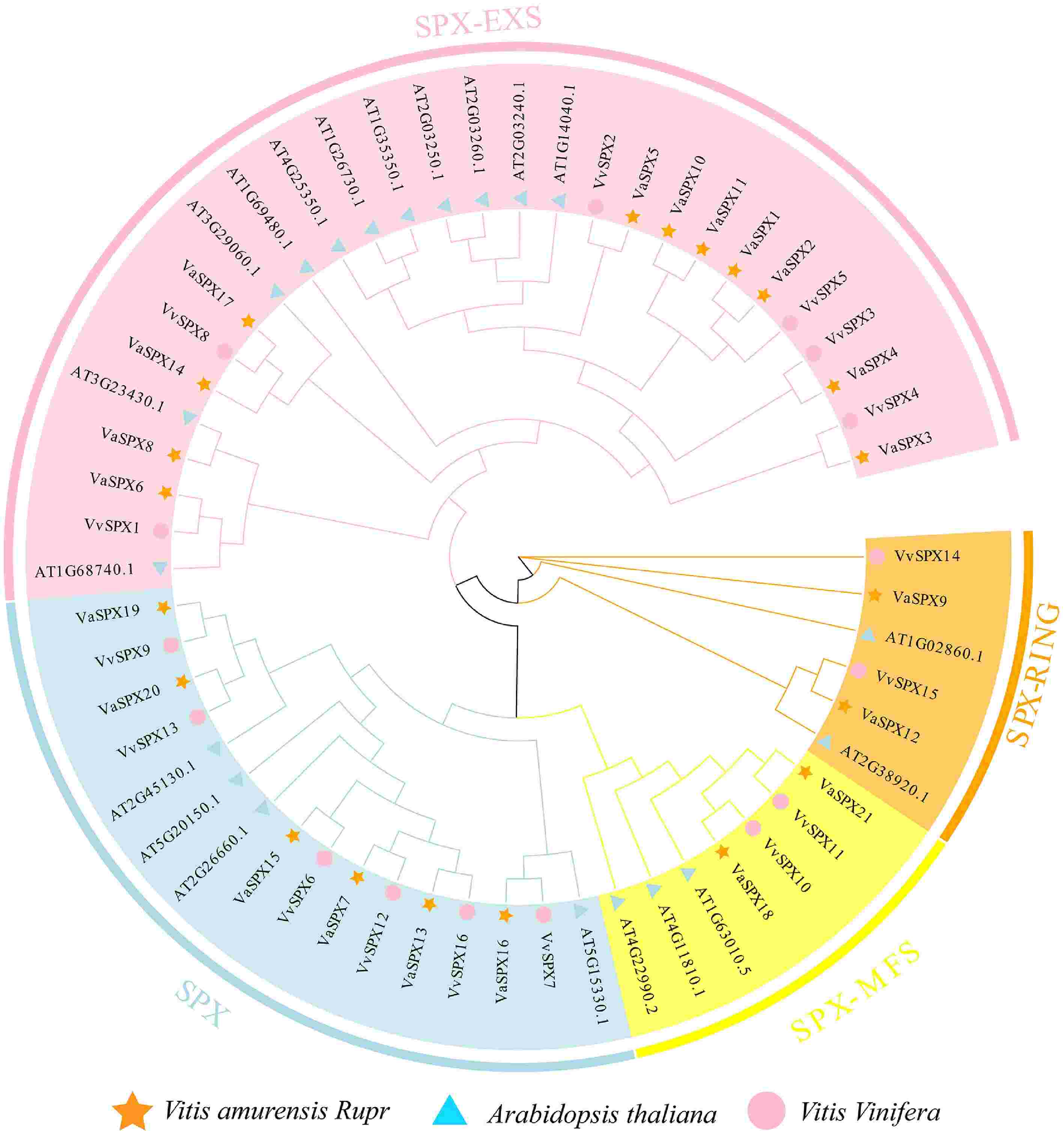
Figure 2.
Phylogenetic analysis of SPX gene families in V. vinifera, V. amurensis, and Arabidopsis thaliana. The SPX protein sequences were used to create phylogenetic trees. The bootstrap value was set at 1,000, and the NJ approach was used.
Analysis of the motif, structural domain, and gene structure of the VvSPX and VaSPX gene families
-
As shown in Fig. 3a, there were five relatively conserved motifs in the VvSPX and VaSPX gene families. Except for the absence of motif 1 in VaSPX1, VaSPX11, and VaSPX14, all other genes had motif 1. Except for VaSPX12, VvSPX15, VvSPX13, and VaSPX20, which did not have the motif 4, all other genes had the motif 4. The number and type of conserved motifs in the four VaSPX protein sequences varied considerably (Fig. 3a). VvSPX15, VaSPX12, and VaSPX20 had only motif 1, and VaSPX14 had only motif 4, which differed considerably from the other sequences.
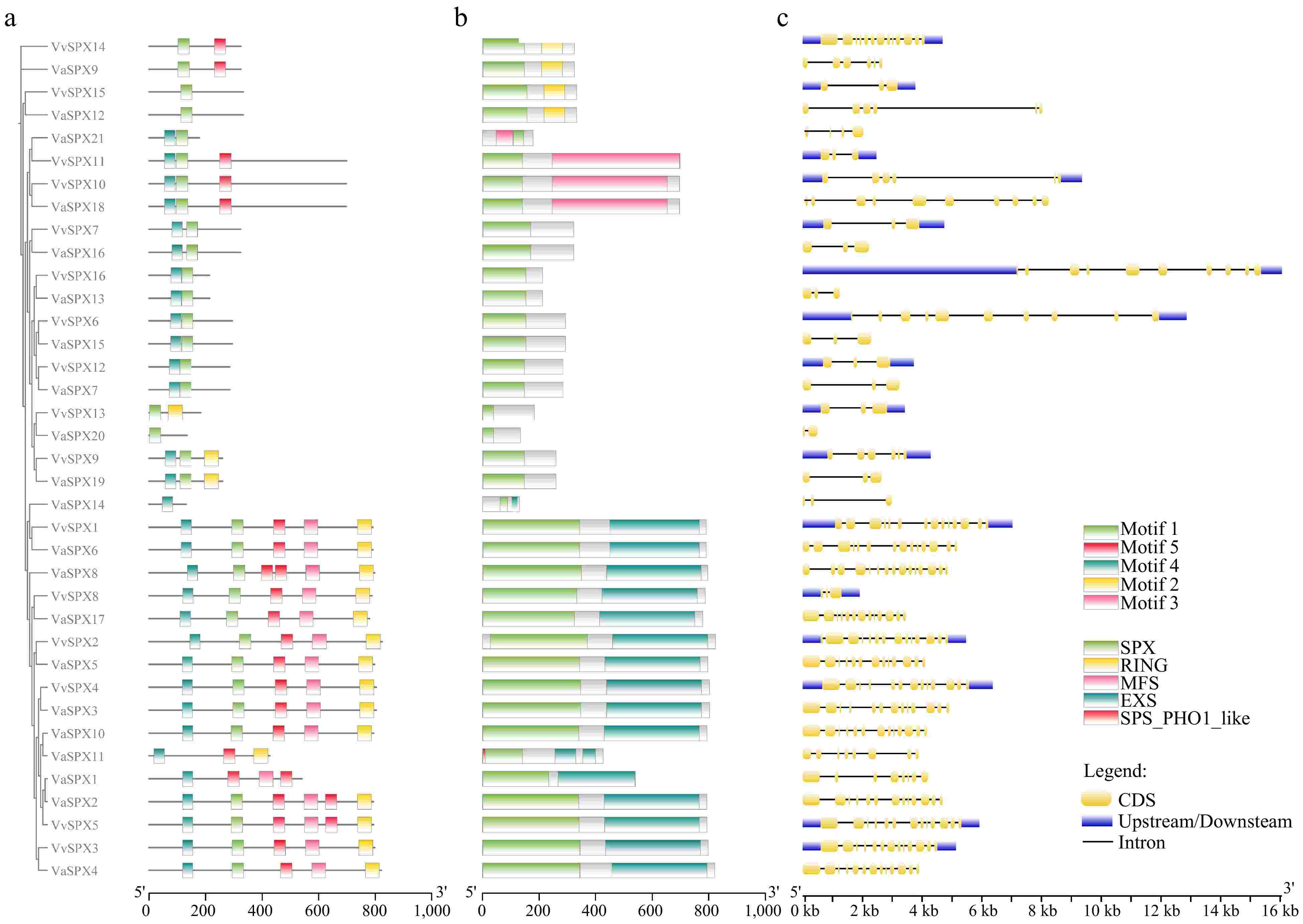
Figure 3.
Sequence, domain, and gene structure analysis of VvSPX and VaSPX genes. (a) Conserved sequence analysis of SPX gene in V. vinifera and V. amurensis. (b) Analysis of conserved domain of SPX gene in V. vinifera and V. amurensis. (c) Exon-intron structure of SPX gene in V. vinifera and V. amurensis.
Based on evolutionary relationships, joint analysis of conservative domains (Fig. 3b). The SPX structural domain is present in both VaSPX and VvSPX proteins, and the SPX subfamily contains only the SPX structural domain; the SPX-RING subfamily contains two structural domains: SPX and RING domains. The composition of the SPX-EXS subfamily domain was relatively complex. Among them, VaSPX11 contains the SPX, EXS, and SPX-PHO1-like domains, while the other proteins contain both SPX and EXS domains. The SPX-MFS subfamily proteins contain both SPX and MFS domains.
Gene structural analysis showed that (Fig. 3c), few genes had genome lengths less than 2 kb, while the great majority had genome lengths shorter than 9 kb. The 15 VvSPX genes contained upstream and downstream sequences, and the 1 VaSPX gene contained upstream and downstream sequences. Every gene that has non-coding regions has two non-coding regions; for instance, VvSPX14 has one non-coding region in front of the coding region and one after it. There were 16 genes with non-coding regions, accounting for 43.24% of the total. VvSPX and VaSPX had different numbers of introns and exons in different subfamilies. VaSPX20 had the fewest introns (2), whereas VaSPX8 had the most (15). The majority of genes have between 3~14 exons, and their gene structures are identical in length and distribution within the same subfamily.
The collinearity relationship of VvSPX and VaSPX gene families
-
The collinearity analysis showed that there were five collinearity pairs in 16 VvSPX genes (Fig. 1a), which were VvSPX2/VvSPX14, VvSPX6/VvSPX16, VvSPX7/VvSPX12, VvSPX7/VvSPX11, and VvSPX11/VvSPX12. The collinearity analysis showed that there were four collinearity pairs in 21 VaSPX genes (Fig. 1b), which were VaSPX1/VaSPX17, VaSPX13/VaSPX15, VaSPX7/VaSPX13, and VaSPX7/VaSPX1. Therefore, the VvSPX and VaSPX genes might have amplified family members through gene replication during evolution.
To further elucidate the phylogenetic mechanism and homology of the VvSPX and VaSPX genes, collinearity analyses between species were performed by the MCScanX tool of TBtools (Fig. 1c). There were 15 pairs of collinearity pairs between the VvSPX and VaSPX gene families. The drawing of collinear relationship maps between V. vinifera and Arabidopsis and rice (Fig. 1d) showed that there were 16 and 11 homologous genes in Arabidopsis and rice, respectively, showing that compared to monocotyledonous plants, V. vinifera, and dicotyledonous plants contained more homologous genes. A collinear relationship map was drawn between V. vinifera, Arabidopsis, and rice (Fig. 1e), with 12 and eight homologous genes in Arabidopsis and rice, respectively.
Analysis of VvSPX gene family gene chip expression patterns
-
Gene chips help research how members of a gene family express themselves in various tissues, during various stages of development, or in various environments. This aids in the prediction of these genes' functions. Since there is no gene chip database for mountain grapes, only a gene chip map of grapes was done. The results of gene expression profiling analysis (Supplementary Fig. S2) indicated that two or more genes located in the same branch of the evolutionary tree will have similar expression profiles. VvSPX1, VvSPX2, VvSPX5, and VvSPX16 showed high expression in leaves. VvSPX2, VvSPX3, VvSPX9, and VvSPX15 were all found to be expressed to a high degree in roots. VvSPX2 had the highest expression level in roots. The majority of the genes that were highly expressed in flowers were members of the SPX-EXS subfamily, while the remaining genes, known as VvSPX14 and VvSPX15, were members of the SPX-RING subfamily. It was anticipated that the SPX-EXS and SPX-RING subfamilies will regulate the growth and development of fruits and flowers.
Codon preferences in VvSPX and VaSPX gene families
-
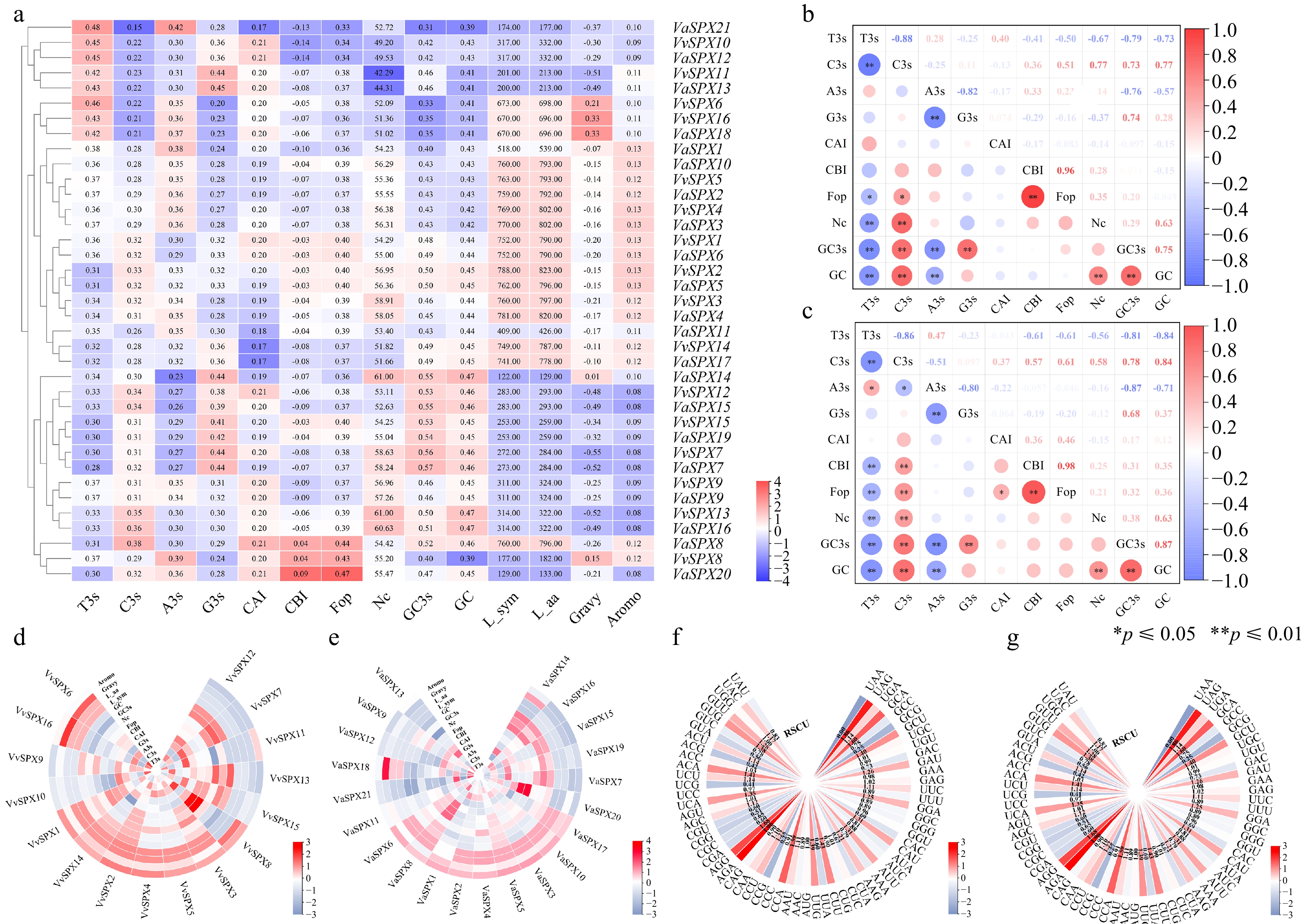
In the deviation of synonymous codon usage (Fig. 4a), VaSPX14 had a maximum Nc value of 61.00 and a minimum Nc value of 44.43, while VvSPX13 had a maximum Nc value of 61.00 and a lowest Nc value of 42.29.
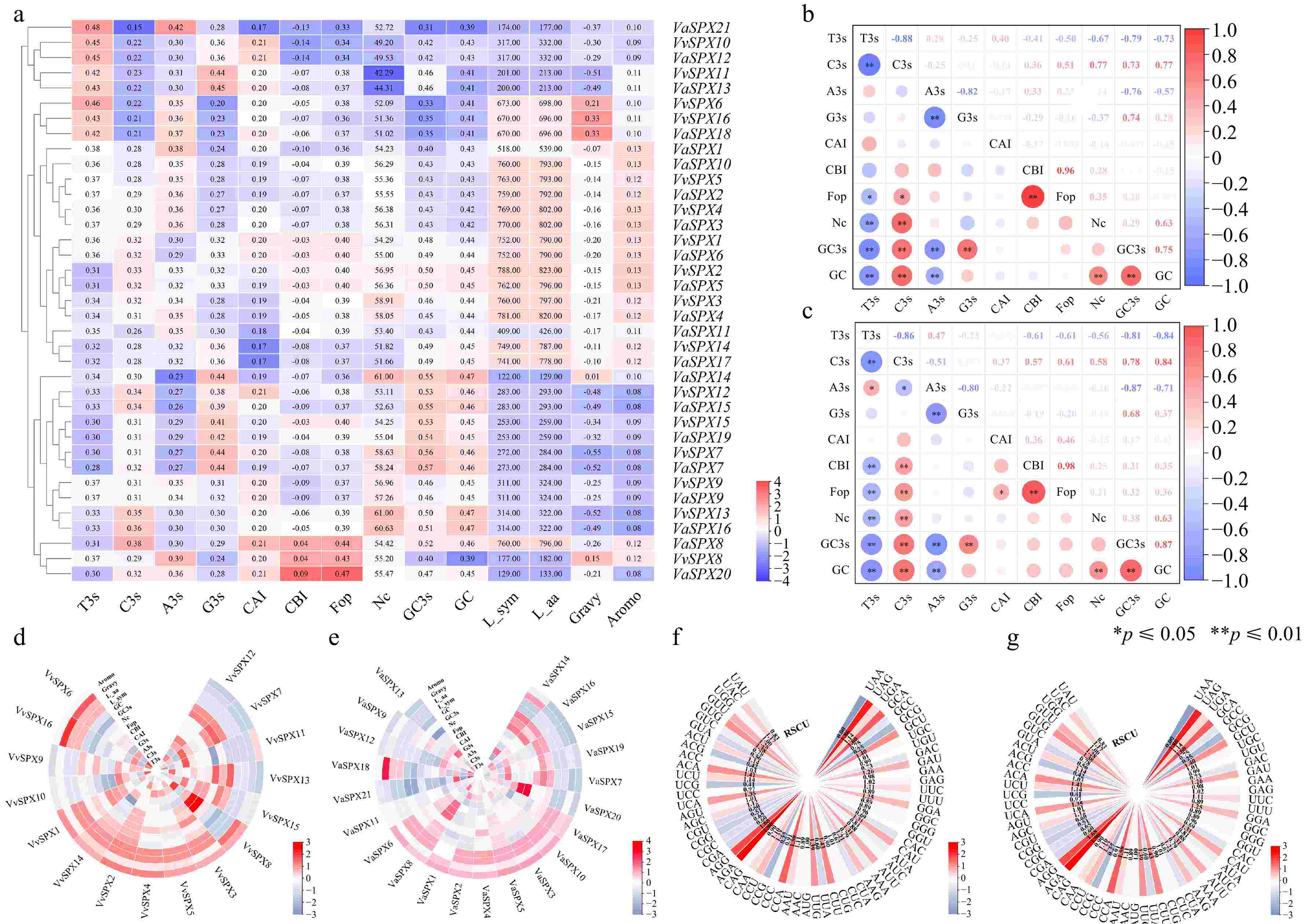
Figure 4.
VvSPX and VaSPX gene codon correlation and use bias analyses. (a) Synonymous codon preference and correlation analyses of VvSPX and VaSPX gene. (b) Correlation analysis of V. vinifera SPX gene codons. (c) Correlation analysis of SPX gene codons in V. amurensis. White denotes no association, blue denotes a negative correlation, and red denotes a positive correlation. The association is stronger and the circle is larger when the color is darker, and vice versa. (d) Analyses of the VvSPX gene's correlation and synonymous codon preference. (e) The VaSPX gene's synonymy codon preference and correlation analysis. The magnitude of the correlation coefficients is shown by different colors; the larger the correlation coefficients are in red, and the smaller numbers are in blue. 'A3s, G3s, C3s, and T3s' denote the matching base frequency of the synonymous codon on the third. CAI, codon adaptation index. CBI, codon preference index. FOP, frequency of optimal codon usage. NC: codon number that is effective. GC content of the synonymous codon for GC3 at position three. GC: the GC content. (f) Relative synonymous codon usage in V. vinifera (RSCU). (g) Relative synonymous codon usage in V. amurensis (RSCU).
Within the VvSPX family, T3s had a negative correlation with GC, GC3S, Nc, Fop, CBI, G3s, and C3s and a positive correlation with CAI and A3s. GC3s had a positive correlation with C3s, G3s, Fop, GC, and Nc and a negative correlation with T3s, A3s, CAI, and CBI (Fig. 4b). But in the VaSPX family, T3s had a negative correlation with GC, GC3S, Nc, Fop, CBI, G3s, CAI, and C3s, but a positive correlation with A3s. GC3s had a positive correlation with C3s, G3s, CAI, CBI, Fop, Nc, and GC and a negative correlation with T3s and A3s (Fig. 4c).
In the deviation of synonymous codon usage in V. vinifera (Fig. 4d), the VvSPX family members had Fop frequencies ranging from 0.36 to 0.43 and CAI values between 0.17 and 0.21. With mean values of 43.53% for GCs and 45.83% for GC3s, the VvSPX family members' GC content ranged from 39.00% to 47.00%, and their GC3s content from 32.70% to 55.90%.
On V. amurensis synonymous codon usage bias (Fig. 4e), the VaSPX family members' Fop frequencies ranged from 0.333 to 0.473, whereas the range of their CAI values was 0.174 to 0.214. With mean values of 43.98% and 46.40% for GCs and GC3s, respectively, the GC content of the VaSPX family members varied between 38.80% and 47.50%, while the GC3s concentration ranged between 31.00% and 56.80%. We discovered that VaSPX family members had higher GC and GC3 values than A3s and T3s. Four genes (VvSPX10, VvSPX11, VaSPX12, and VaSPX13) were discovered to have Nc values below 50, suggesting a comparatively high codon preference for four genes.
RSCU was an assessment of the preference for using synonymous codons. AGA-encoded Arg was more commonly used and preferred, according to the statistics. The frequency of relative synonymous codon usage in the V. vinifera genome was analyzed (Fig. 4f), a total of 34 codons was found to have RSCU ≥ 1, namely UAC, UGG, GUU, GUG, ACU, ACC, ACA, UCU, UCA, AGU, AGC, AGG, AGA, CAA, CCU, CCA, AAU, AUG, UUG, CUU, CUC, AAG, AUU, CAU, GGU, GGA, UUC, GAG, GAU, UGC, GCU, GCA, UGA, UAG. However, one less codon in V. amurensis was GGU (Fig. 4g). In VaSPX, 11 of the third codons were U, eight were G, eight were A, and seven were C. Unlike VvSPX, six of the third codons of VaSPX were C. This suggested that the VvSPX and VaSPX proteins had a higher propensity for a U at the third codon of amino acids.
Analysis of cis-acting elements of VvSPX and VaSPX gene family
-
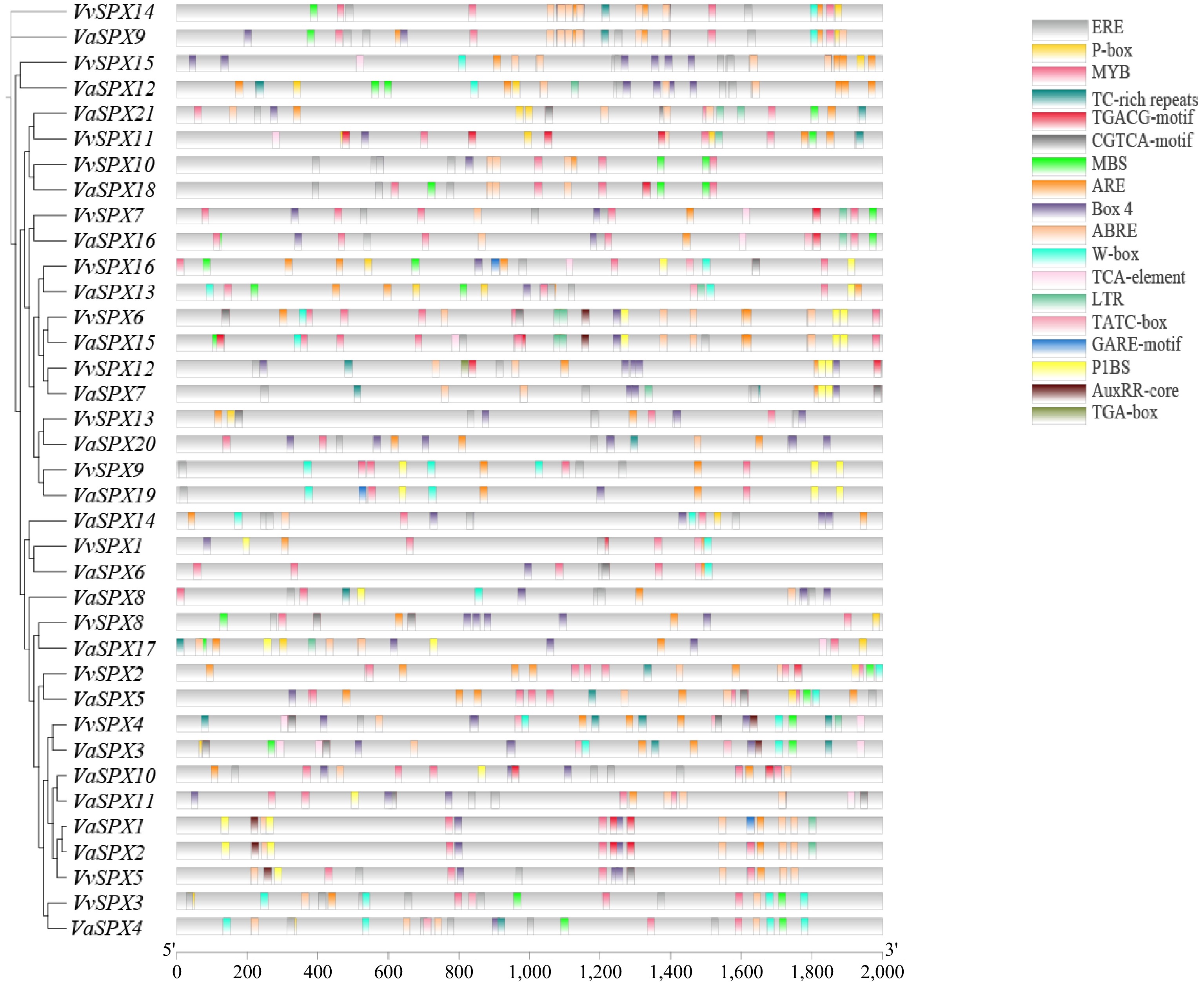
Numerous elements linked to hormones, environmental cues, and phosphorus deprivation were found when 2,000 bp promoter sequences upstream of 16 VvSPX genes and 21 VaSPX genes were analyzed for cis-acting elements (Fig. 5).
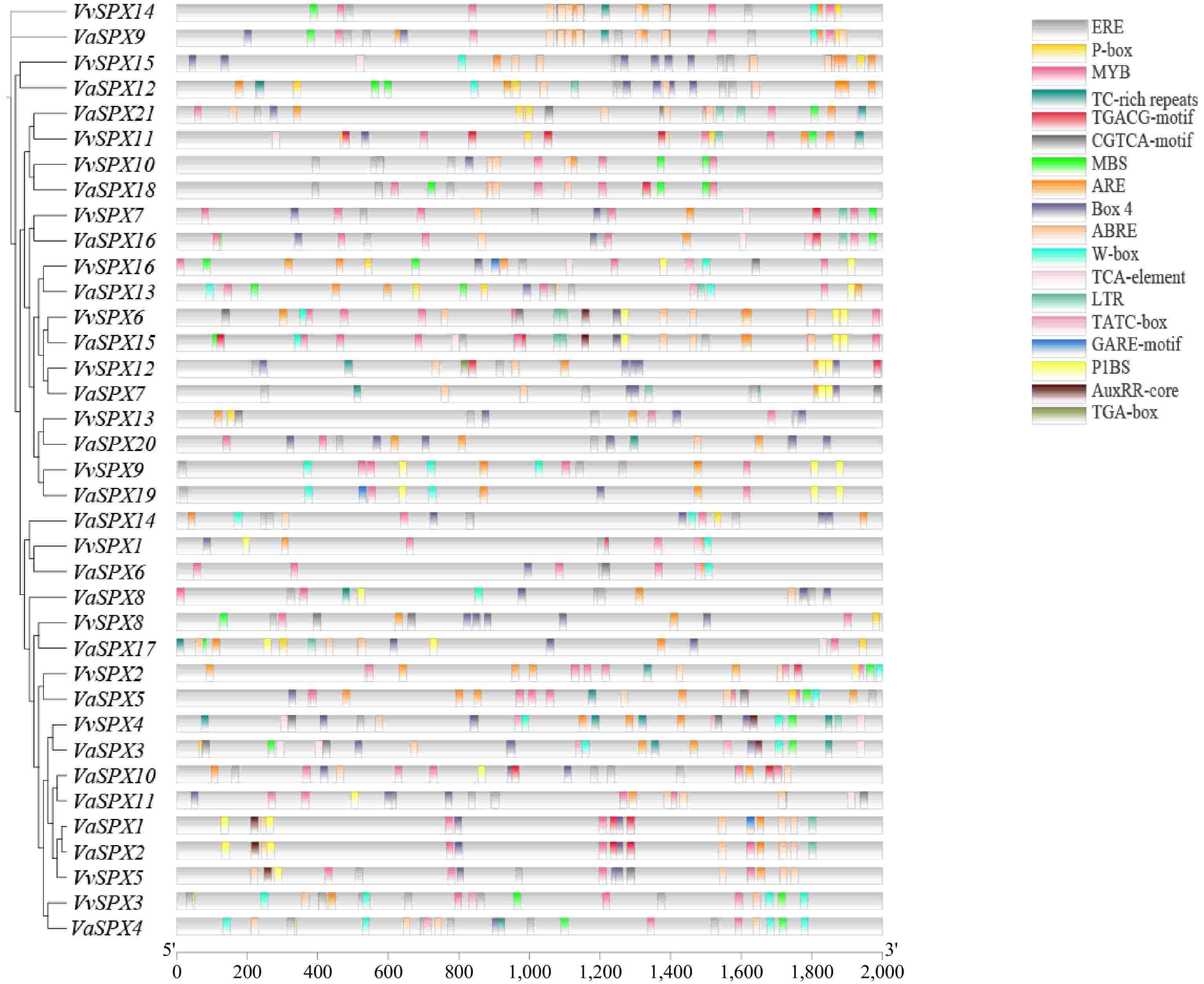
Figure 5.
Cis-regulatory element analysis of the VvSPX and VaSPX genes.
An analysis of the VvSPX gene family's cis-acting elements showed that the majority were linked to stress and hormones. The promoter region included three phosphorus stress-related and nine hormone-related elements, in addition to other related elements. There were 18 cis-acting regulatory elements. Among them, the abscisic acid response element contained more; VaSPX9 contained seven abscisic acid response elements. All genes except VvSPX10 and VaSPX4 were scattered with the response element ERE, which was associated with the stress response, and the number of EREs was also high. It implied that V. vinifera and V. amurensis growth and development might be significantly influenced by the SPX genes and have an equally important place in abiotic stresses. On the promoters of genes reacting to low phosphorus stress, the phosphorus starvation response-associated cis-acting elements P1BS (PHR1-binding sequence), MBS, and W-box were dispersed. Important phosphorus deficiency response element P1BS GNATATNC, with the VvSPX6 and VaSPX13 promoters containing the most P1BS elements with four. On other promoters containing P1BS elements, the number of P1BS elements varies from one to three, suggesting that they might be directly regulated by the gene PHR1 related to phosphorus deficiency response. Furthermore, 20 additional members had a WRKY transcription factor binding element (W-box) on their promoters, which suggested that some WRKY family members are involved in the operation of the VvSPX and VaSPX gene families.
The presence of additional environmental signal elements in the SPX gene's promoter region, such as light-responsive elements (Box4), defense/stress-responsive (TC-rich repeats) elements, anaerobic progenitors like ARE and TCA-box, indicated that the SPX gene might be involved in the response to other signals in addition to controlling phosphorus absorption and homeostasis in plants.
Expression pattern analysis of VvSPX and VaSPX gene families
-
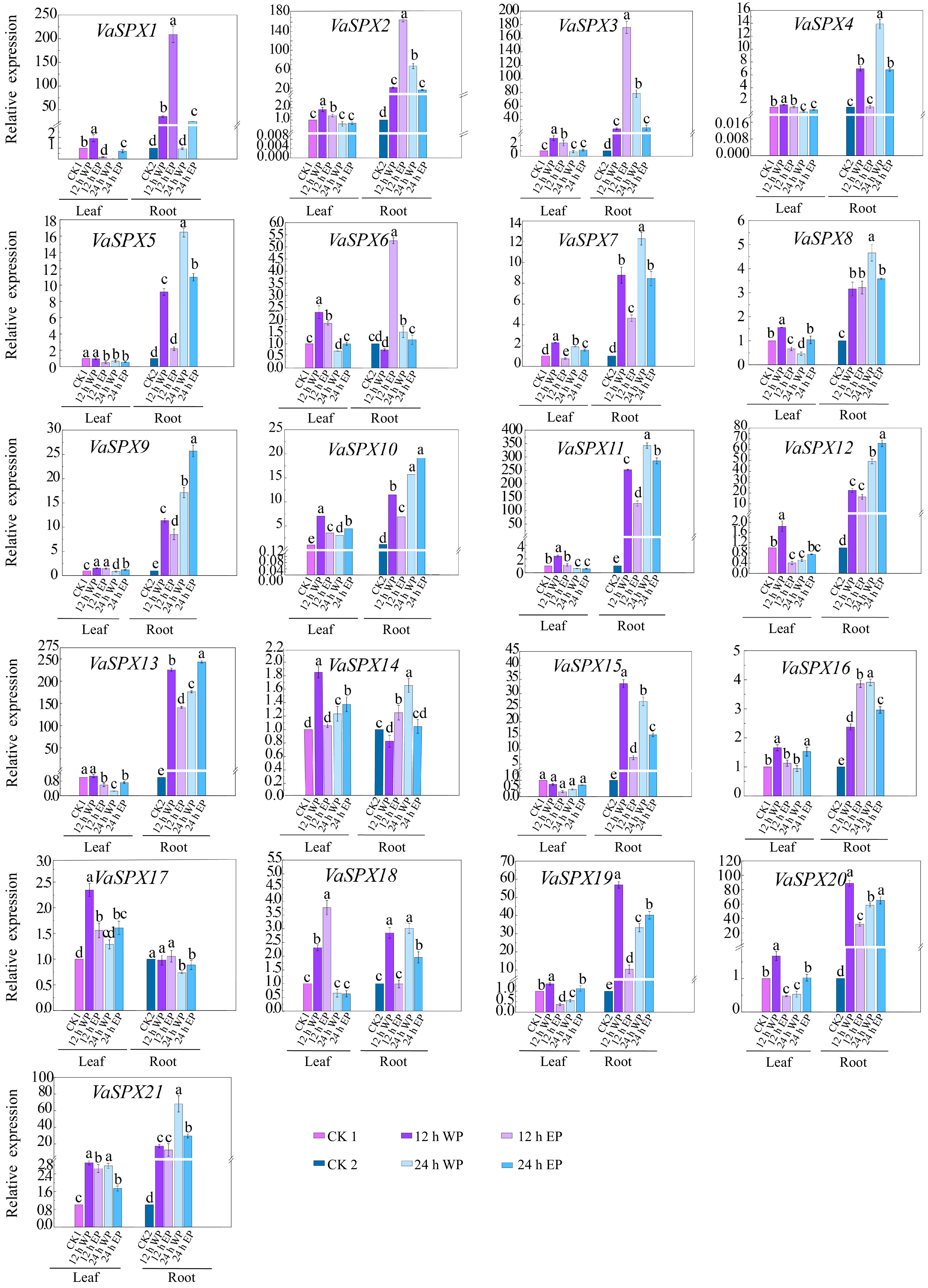
Real-time fluorescence quantification of 37 members of the SPX gene family in the roots and leaves of 'Pinot Noir' and 'Zuoyouhong' showed that (Figs 6, 7), the expression levels of these genes were different in roots and leaves, and different tissue-specific patterns were observed in response to phosphorus stress.
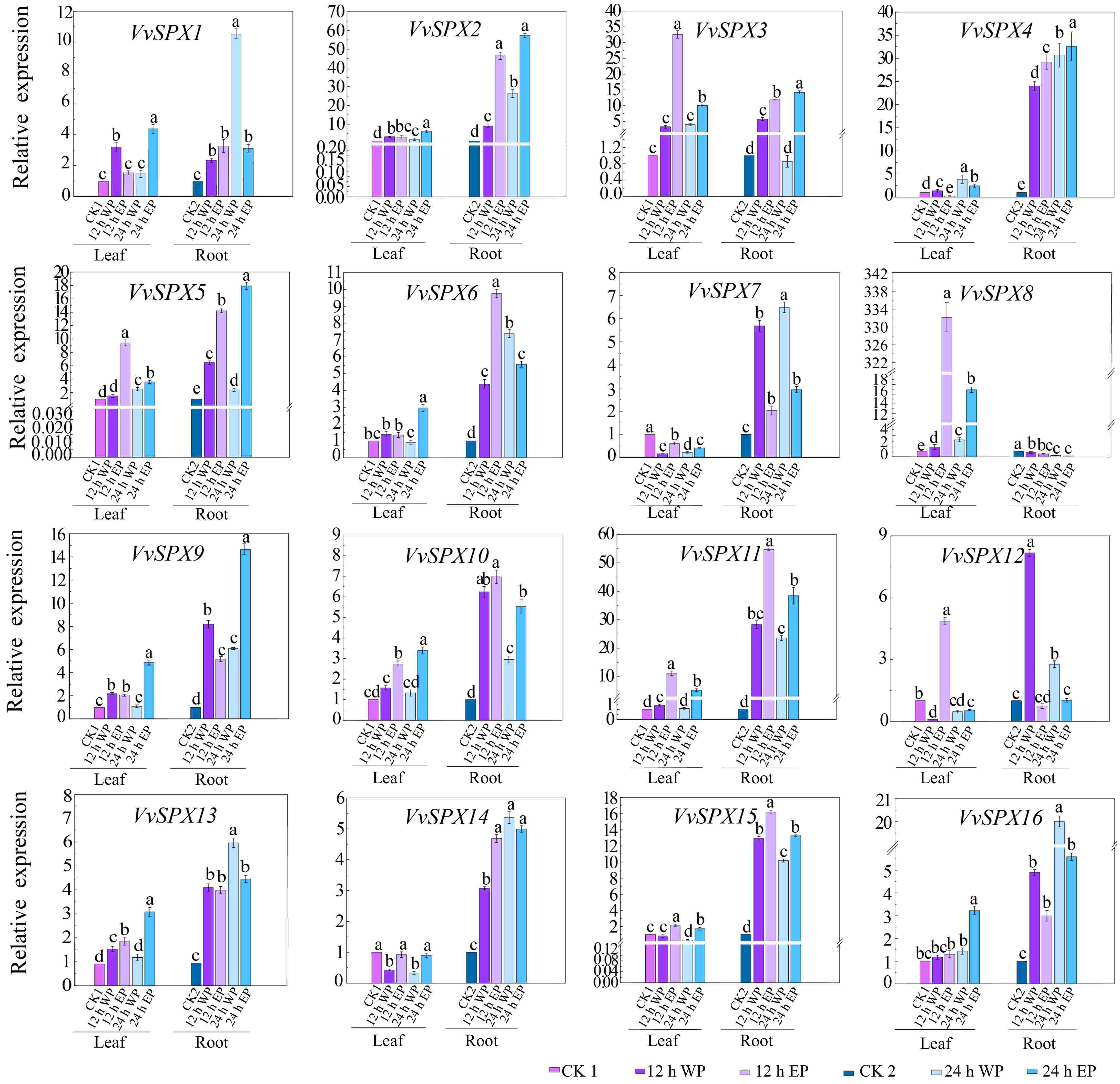
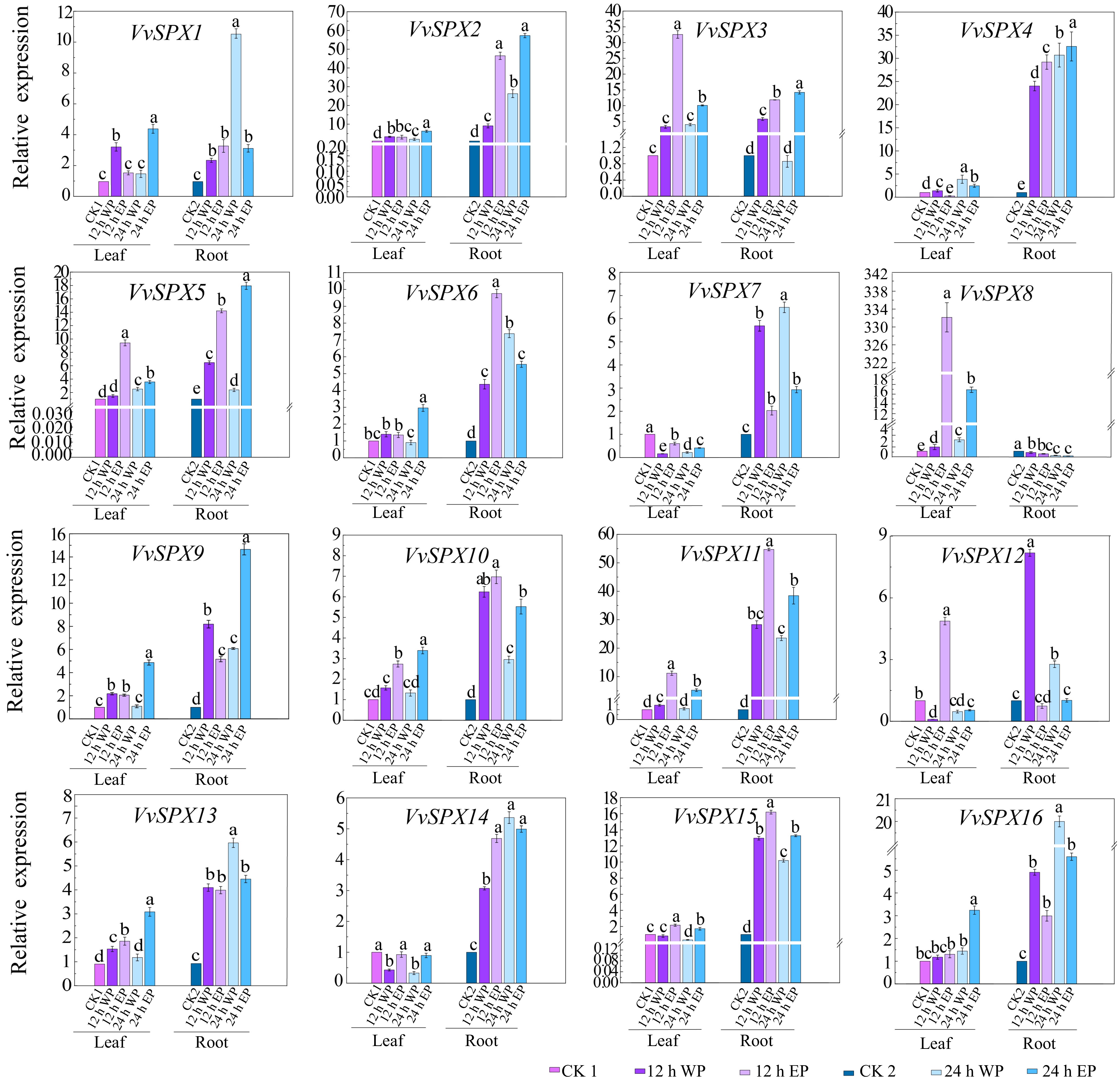
Figure 6.
Relative expression levels of SPX genes in V. vinifera with different phosphorus concentrations and different treatment times. CK1 was used as a control for leaves and CK2 as a control for roots. CK1, 1 mM Pi treated leaves for 0 h; 12 WP, 0 mM Pi treated for 12 h; 12 EP, 2 mM Pi treated for 12 h; 24 WP, 0 mM Pi treated for 24 h; 24 EP, 2 mM Pi treated for 24 h; CK2, 1 mM Pi treated roots for 0 h; WP: without phosphorus (0 mM Pi), EP: excessive phosphorus (2 mM Pi). The relative expression was computed using the 2−ΔΔCᴛ technique[32]. The mean ± SE of three biological repetitions is shown by the error bars. The significance of group differences was assessed using Duncan's test and analysis of variance (ANOVA). While the same lowercase letters indicate no statistical difference (p < 0.05), different letters indicate significant differences.
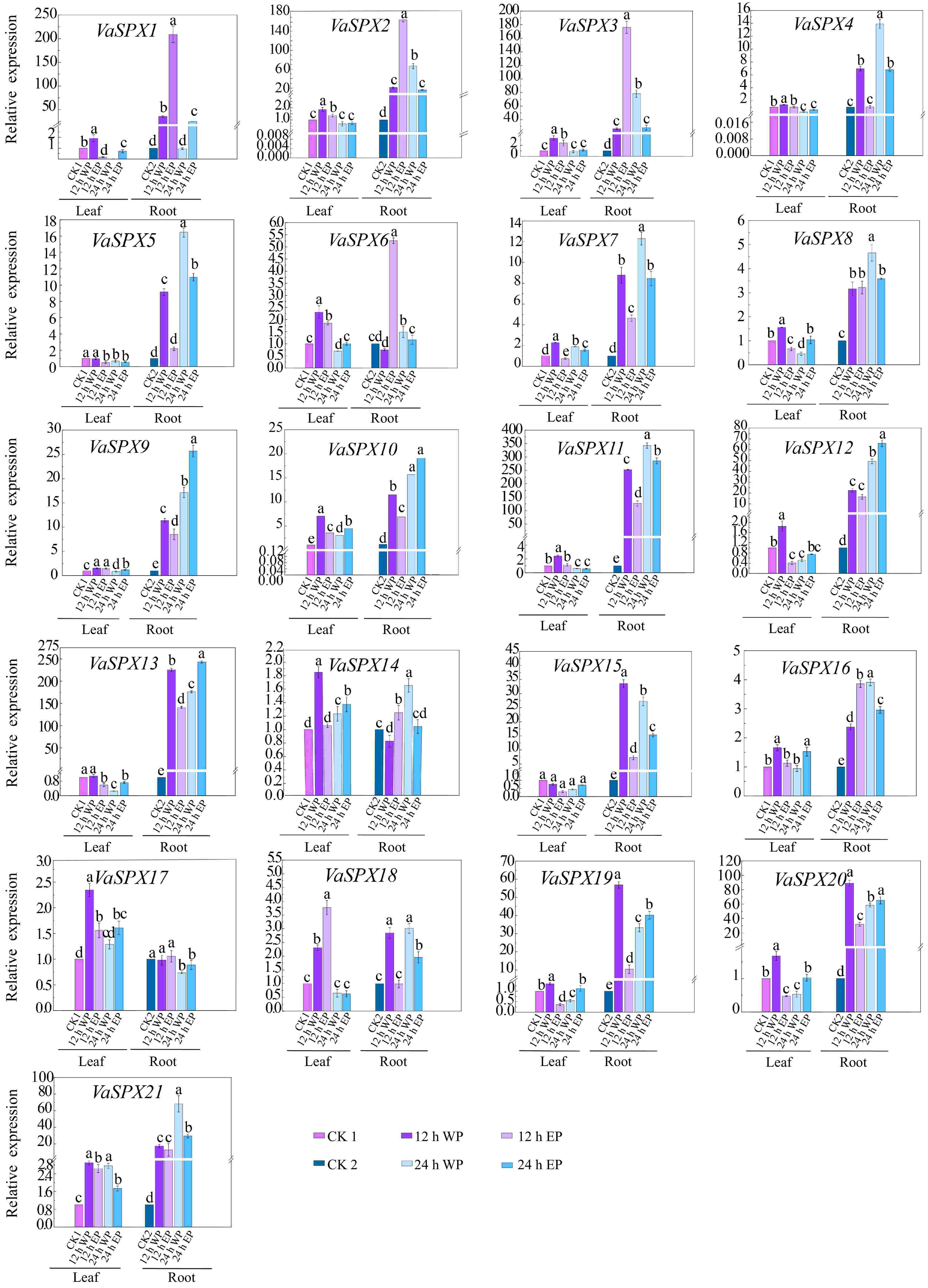
Figure 7.
Relative expression levels of SPX genes in V. amurensis with different phosphorus concentrations and different treatment times. The abbrviations ets are the same as for Fig. 6.
In roots treated with both phosphorus excess and phosphorus deficit for 12 and 24 h, members of the SPX subfamily showed a tendency to be induced by phosphorus stress. As a member of the SPX-MFS subfamily involved in vacuolar phosphorus transport, VvSPX10, VvSPX11, VaSPX18, and VaSPX21 all showed a trend of being induced by phosphorus stress. Members of the SPX-RING subfamily mainly participate in the regulation of phosphorus balance in plants. VvSPX14, VvSPX15, VaSPX9, and VaSPX12 all showed a trend of being induced under both phosphorus deficiency and phosphorus excess.
In roots treated with phosphorus deficiency at 12 and 24 h, in the SPX-EXS subfamily, VaSPX6 and VaSPX14 were downregulated at 12 h of phosphorus deficiency and upregulated at 24 h of phosphorus deficiency. Other subfamily members were induced in the roots treated with phosphorus excess for 12 and 24 h, with the exception of VvSPX8 and VaSPX17. The differential expression patterns of different members during different stages of phosphorus deficiency suggest that they may respond to different levels of phosphorus stress in plants and exert different functions. VvSPX8 was downregulated in roots and upregulated in leaves at 12 and 24 h of phosphorus deficiency and polyphosphate treatment, but VaSPX5 was the opposite. Perhaps because the fact that members of the SPX-EXS subfamily mainly participate in the transportation of phosphorus from underground to above ground.
The relative expression level of leaves is generally lower than that of roots. In the leaves, only VvSPX9, VvSPX13, VaSPX7, VaSPX19, and VaSPX20 in the SPX subfamily were significantly induced at 12 h of phosphorus deficiency. Only VvSPX8 in the SPX-EXS subfamily was significantly induced by phosphorus excess for 12 h. All of the genes in the SPX-MFS subfamily were upregulated, except VaSPX18, which was downregulated 24 h after phosphorus excess and deficiency treatment. Every member of the SPX-RING subfamily was upregulated at 12 and 24 h of phosphorus excess, except VaSPX12, which was downregulated.
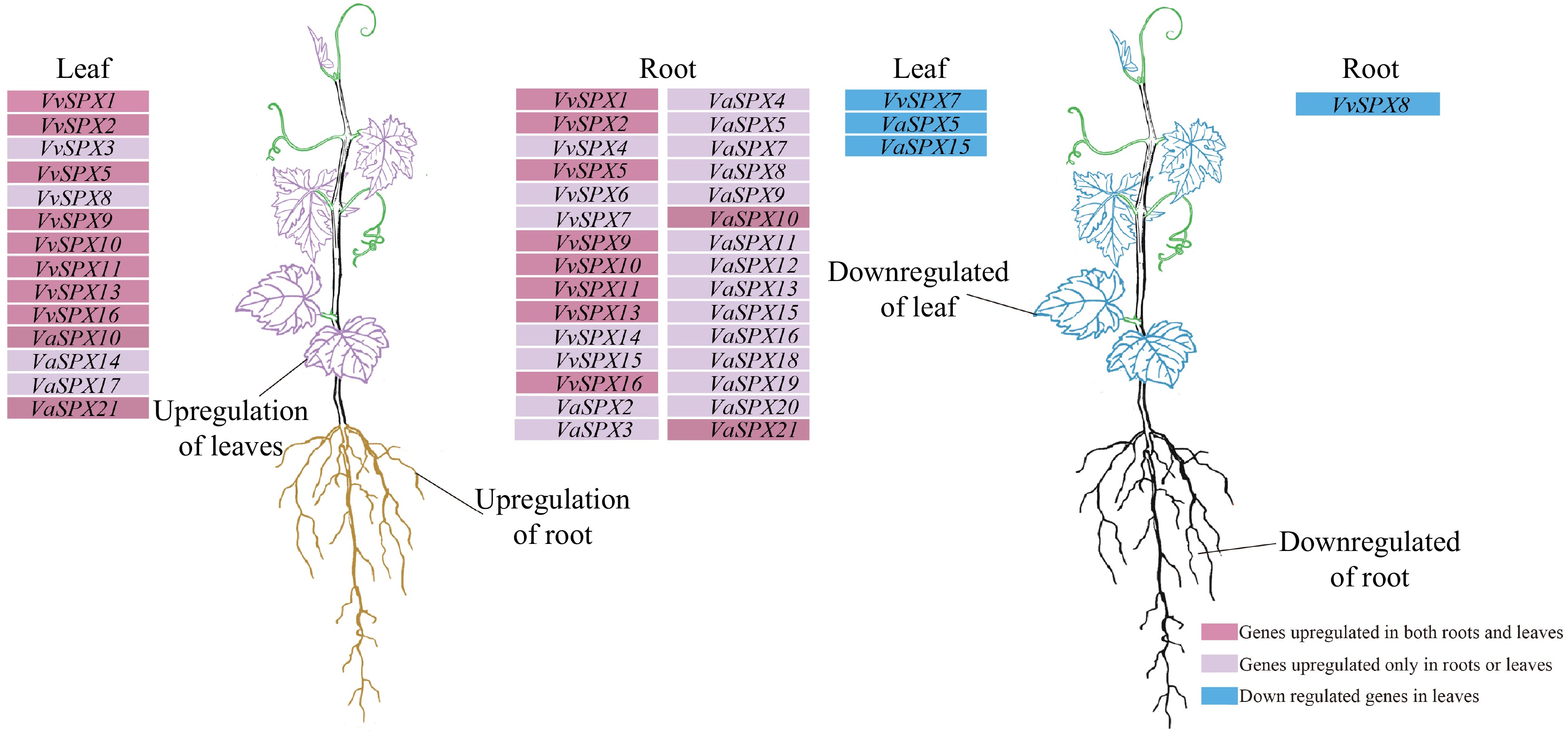
Genes with elevated or downregulated relative expression levels in roots and leaves among 16 VvSPX genes and 21 VaSPX genes were aggregated based on qRT-PCR results. As shown in Fig. 8, the roots and leaves of VvSPX1, VvSPX2, VvSPX5, VvSPX9, VaSPX10, VvSPX11, VvSPX13, VvSPX16, VaSPX10, and VaSPX21 were upregulated under different concentrations of treatment, whereas the expression levels of the remaining 27 genes varied with time and concentration.
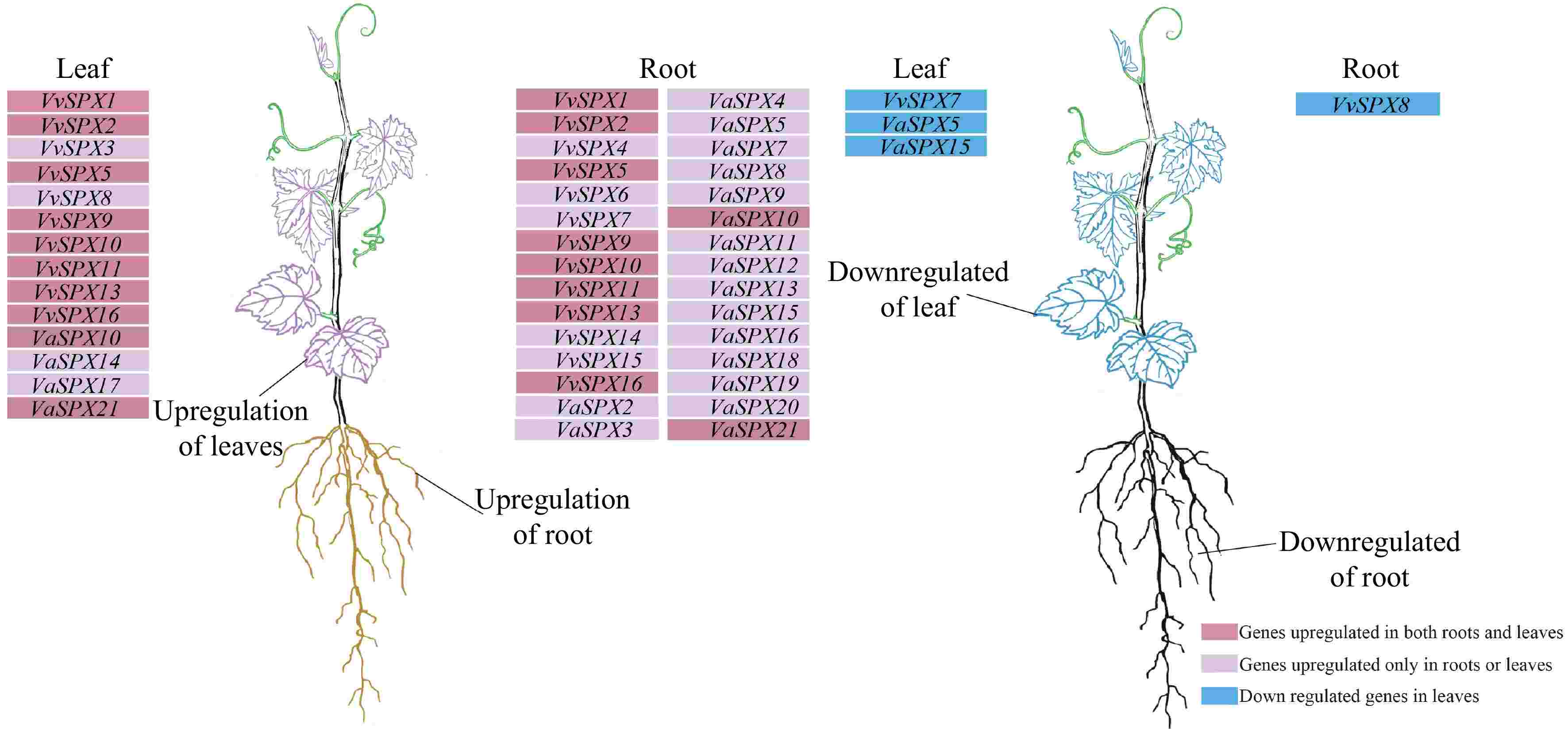
Figure 8.
Cluster analysis of upregulated and downregulated expression trends of VvSPX and VaSPX genes in roots and leaves. The genes in the deep purple background were upregulated in both roots and leaves, the genes in the purple background were upregulated in roots or leaves, and the genes in the blue background were downregulated in roots or leaves following treatment with 0 and 2 mM phosphorus for 12 and 24 h.
-
A group of proteins with the SPX structural domain are known as the SPX family; under phosphorus nutritional stress in plants, the SPX gene family was crucial for controlling phosphorus homeostasis, preserving plant phosphorus homeostasis, and phosphorus signaling[7]. Additionally, the SPX structural domain, by attaching itself to inositol polyphosphate signaling molecules (InsPs), which functioned as phosphorus sensors, was demonstrated to take part in the molecular regulatory pathways of phosphorus signaling[9].
Phosphorus homeostasis in plants under phosphorus stress was regulated by SPX genes, which were found in a variety of crops. For instance, Arabidopsis[25], tomato[26], maize[33], and soybean[7] each possessed 20, 17, 33, and nine members of SPX genes. Despite some progress, the function and evolution of the grape SPX genes have not been explored. Based on other domains that were already known[7], proteins with the SPX domain were categorized into four groups, including the SPX subfamily, the SPX-EXS subfamily, the SPX-MFS subfamily, and the SPX-RING subfamily, which was in line with the tomato study[34]. Subcellular prediction revealed that VvSPX and VaSPX family members were mainly localized in the plasma membrane, nucleus, and cytoplasm, which was consistent with the reported SPX family localization in Arabidopsis thaliana[25].
According to secondary structure analysis, proteins of all VvSPX and VaSPX gene families had more than 25% α-helices and irregular convolutions. This suggested that α-helices and irregular convolutions contributed to the formation of the SPX structure. Given their comparatively high content, it is possible to conclude that α-helices are linked to this gene family's function in the construction of the SPX structure. This was consistent with Liu et al.'s research[35].
Gene duplication, which encompasses tandem, local, and genome-wide duplication, is crucial to an organism's evolution[36]. Subfunctionalization and neofunctionalization can preserve redundant genes at the expression or coding sequence level[27]. Chromosomal localizations showed that 16 VvSPX genes and 21 VaSPX genes were distributed on 10 and 9 chromosomes, respectively (Fig. 3a & b), and there was a heterogeneous distribution. A phenomenon known as gene clustering occurrs when several genes in a family are closely together in a hotspot area[23]. Some of the VvSPX and VaSPX genes formed gene clusters on the chromosome; for example, V. vinifera Chr01 had five genes, followed by Chr14 and Chr15 with two genes. V. amurensis Chr01 had six genes, followed by Chr07 and Chr15 with three genes. Of the gene pairs of the VvSPX and VvSPX gene families (Fig. 4), all but VaSPX1/VaSPX17 belonged to the same family, indicating that these genes might have similar functions. This suggested that tandem duplication might be the main factor behind the growth of the SPX gene family in grapes.
In the collinearity between species, there were 16, 11, 12, and eight pairs of homologous genes in V. vinifera/Arabidopsis thaliana, V. vinifera/Oryza sativa, V. amurensis/Arabidopsis thaliana, and V. amurensis/Oryza sativa, respectively. The homologous genes, which were mainly concentrated in Chr01 and Chr14 of V. vinifera and V. amurensis (Fig. 3d & e), suggested that SPX genes were evolutionarily distantly related to monocotyledonous rice and more closely related to dicotyledonous Arabidopsis thaliana. It was assumed that the homologous gene sequences of the SPX gene family were conserved during evolution, as homologous genes from a common ancestor were found in many species. Homologous genes from a common ancestor existed in different species, and therefore, it was surmised that the homologous gene sequences of the SPX gene family were conserved during evolution. Furthermore, whilst certain SPX genes had no homologous gene pairs, others had 1−6 homologous gene pairs. This could be because different species have different rates of evolution and genetic specificity.
Among the four subfamilies of the evolutionary tree SPX (Fig. 1), the VvSPX and VaSPX genes showed high homology and close evolutionary relationships, with presumably similar functions of the homologous genes, but also some differences, which tentatively suggested that they were evolutionarily conserved.
In the present study, seven SPX genes were found in the tomato genome[34], four SPX genes were found in Arabidopsis[25], six SPX genes were found in rice[27], and six SPX genes were found in VvSPX and VaSPX, with a comparable number of genes in the SPX subfamily. As signaling proteins that sense phosphate, the SPX subfamily frequently performs a crucial function[25,34]. In this study, both VvSPX and VaSPX genes were induced at 12 h and 24 h treatments with low phosphorus. Consistent with studies that all members of the three Arabidopsis AtSPXs and the five rice OsSPXs were induced by low phosphorus in recipient roots, except for AtSPX4 and OsSPX4, which were inhibited by low phosphorus stress[37]. Components that respond to phosphorus deficiency. The promoters of genes that react to phosphate-deficient stress frequently contain P1BS, MBS, and W-box, which balance phosphorus intake and transport by controlling the expression of genes linked to phosphorus stress[38]. To maintain phosphorus homeostasis in plants, for instance, rice Os-SPX4 binds to the P1BS element on the promoter of the phosphorus starvation response gene to regulate appropriate responses[39]. To control plant responses to phosphorus shortage, AtMYB2 bound to the MBS cis-acting element on the miR399f promoter and facilitated phosphorus starvation signaling[40]. Phosphorus shortage increased the expression of AtWRKY45 and AtWRKY75, and their proteins were activated by directly binding to the promoter's W-box motif[41,42]; However, the expression of this gene is inhibited when AtWRKY6 and AtWRKY42 bind to the W-box on the PHO1 promoter[43,44]. It was found that members of the SPX subfamily, at low phosphorus levels, can both inhibit as much as possible the deleterious effects of the phosphorus starvation response on plant growth and development and act as inhibitors of phosphorus uptake as soon as the phosphorus starvation conditions are lifted, to prevent excessive phosphorus accumulation from causing damage to the plant[45−47]. Among the members of the SPX subfamily, except for VvSPX6, VvSPX9, VvSPX12, VvSPX16, VaSPX7, VaSPX13, VaSPX15, and VaSPX19, VvSPX7, VvSPX13, VaSPX16, and VaSPX20 did not have a P1BS element, while the VvSPX7 promoter, although it did not have a P1BS element, contained one MBS element. The VaSPX16 promoter contained one MBS element. W-box and MBS elements were also distributed on the promoters of VaSPX13 and VaSPX15. W-box components were also distributed on VvSPX6, VvSPX9, VaSPX19, and VvSPX16. The promoters of VvSPX and VaSPX genes also had one or more of these three elements distributed, suggesting that the three cis-acting elements, P1BS, W-box, and MBS, might play an important role in the regulation of phosphorus stress. However, VvSPX13 and VaSPX20 were the exceptions; their promoters lacked these three cis-acting elements, but phosphorus shortage stress still caused their expressions. Whereas VvSPX12 had a P1BS element on its promoter, VvSPX7 also had an MBS element on its promoter, and all three cis-acting elements of VaSPX15 were present but negatively regulated in leaves treated with phosphorus deficiency for 12 and 24 h. This illustrates the intricacy of SPX gene regulation under phosphorus shortage stress.
The second group was the SPX-MFS subfamily. The SPX-MFS subfamily's genes primarily functioned as phosphorus transporters on the vacuole membrane[48]. When the cell lacks phosphorus, phosphorus that has been stored in the vacuole is carried out to be used. On the other hand, phosphorus was transferred into the vacuoles when the cell's phosphorus concentration was too high, protecting the cell from damage caused by elevated cytoplasmic phosphorus levels[13,49]. Phosphorus deficit was shown to stimulate high expression of OsSPX-MFS2 while inhibiting the expression of OsSPX-MFS1 and OsSPX-MFS3. Phosphorus was transported from vacuoles to the cytoplasm via OsSPX-MFS3, as demonstrated by Arabidopsis plants expressing OsSPX-MFS1[15,49]. Expression of SPX-MFS subfamily members was upregulated in both phosphorus-deficient and excessive phosphorus-treated roots at 12 and 24 h. The MBS element was present in the promoters of VaSPX18, VaSPX21, VvSPX10, and VvSPX11. Whereas VvSPX10 and VaSPX18 also had MBS elements on their promoters, negative response regulation was observed in leaves of both V. vinifera and V. amurensis treated with phosphorus deficiency for 24 h. This is highly likely related to their different functions in participating in vacuolar phosphorus transport.
The third group was the SPX-EXS subfamily, which was primarily in charge of moving phosphorus from root epidermal cells to the xylem and via the phloem to the roots[50]. Several research studies have shown that the PHO1 gene family controls phosphorus levels and influences plant growth and development in Arabidopsis thaliana, rice, maize, Physcomitrium patens, and tomato[51−55]. The primary function of the SPX-EXS subfamily was that of phosphorus transporters, which were in charge of moving phosphorus from the root system to the aboveground as well as between various aboveground tissues and organs[52,56]. Signals involved in phosphorus transport may cause distinct EXS genes to react[57]. In roots treated with both phosphorus deficiency and phosphorus excess for 12 and 24 h. After 12 and 24 h of phosphorus shortage and polyphosphate treatment, VvSPX8 was downregulated in roots and upregulated in leaves, however, VaSPX5 showed the reverse effect. Phosphorus might be transported from the phloem to the roots by VaSPX5, whereas it might be transported from the roots to the xylem by VvSPX8. The expression of VvSPX3 increased after 24 h of phosphorus surplus and decreased after 12 h of phosphorus deficiency, indicating that varying phosphorus levels control VvSPX3 in different ways, enabling it to balance phosphorus in the plant.
The fourth category was the SPX-RING subfamily, the members of in which mediated the breakdown of phosphorus transporters to preserve the plant body's phosphorus homeostasis when there was an adequate supply of phosphorus[58]. The SPX-RING subfamily was expressed in a range of tissues in Solanum lycopersicum[29], and 0SNLA1 was essential for preserving Pi homeostasis in rice[16]. The roots of VvSPX14, VvSPX15, VaSPX9, and VaSPX12 were upregulated under both phosphorus deficiency and phosphorus excess stress. This suggests that the SPX-RING subfamily has a role in regulating phosphorus homeostasis in the body in the presence of phosphorus imbalance induced by both phosphorus excess and low phosphorus.
Low temperature, drought, high salt, and several phytohormone signal transduction pathways have been demonstrated to have reciprocal effects with the phosphorus stress response, hence influencing plant growth and development[59]. The promoters of the VvSPX and VaSPX family genes contain, in addition to the phosphorus stress-responsive elements P1BS, MBS, and W-box, elements that respond to low temperature (BOX4), growth hormone (AuxRR-core), gibberellins (GARE-motif and P-box), salicylic acid (TCA-element), ethylene (ERE), methyl jasmonate (TGACG-motif and CGTCA-motif), and ABA (ABRE) elements, it suggested that VvSPX and VaSPX genes could not only respond to phosphorus deficiency stress but also participate in a variety of important physiological processes such as abiotic adversity and hormone response, and the SPX genes might also mediate the process of interactions between these adversity factors and the response to phosphorus starvation in plants.
SPX genes regulate phosphorus transport and contain phosphorus stress response elements, hormone response elements, and abiotic adversity response elements on the promoter, and these findings provide direction for breeding grape varieties that are adapted to phosphorus stress environments and can help to enhance the phosphorus starvation response of plants in low-phosphorus soils through precise regulation of the SPX genes using gene editing and other means, to cultivate new grape varieties with excellent traits such as high phosphorus uptake and utilization and high-stress resistance.
The SPX subfamily, based on its properties as phosphate-sensing signaling proteins, can also be used to develop phosphorus nutrition diagnostic techniques based on monitoring changes in the expression or activity of this subfamily of proteins, to timely and accurately determine the phosphorus nutritional status of plants and to achieve precision management in agriculture. SPX-MFS subfamily contributes to the development of novel plant phosphorus nutritional regulators that mimic or interfere with the function of this subfamily of proteins and affect the process of phosphorus transport between the vacuole and the cytoplasm. More effective phosphorus fertilizer synergists can be developed to increase the proportion of phosphorus fertilizer absorbed and utilized by plants by promoting the phosphorus transport process mediated by the SPX-EXS subfamily. The SPX-RING subfamily provides ideas for the development of novel plant growth regulators. For example, by regulating the expression or protein activity of genes related to the SPX-RING subfamily, it is possible to artificially intervene in the degradation process of phosphorus transport proteins.
In kale-type oilseed rape, the SPX-MFS subfamily was significantly upregulated during Pi starvation and recovered after Pi recovered[60]. In Arabidopsis, AtPH01 and AtPH01;H1 were able to move pi to the xylem[57], gene-edited SlPHO1;1 in the genome of Solanum lycopersicum, which is essential for the maintenance of phosphorus homeostasis in tomato seedlings[54], in rice roots, 0sPH01;1 and 0sPH01;2 were substantially expressed[55]. In this study, most of V. vinifera and V. amurensis SPX genes were up-regulated under excess phosphorus treatment, suggesting that SPX genes played an important role in maintaining phosphorus homeostasis under phosphorus stress. This was a unique aspect of the V. vinifera and V. amurensis SPX gene family study. Despite being expressed in both the roots and the leaves, the SPX genes were most prominent in the roots and were linked to the fact that the roots were the primary site of phosphorus uptake in grapes. These insights had important implications for grape crop management or genetic breeding under phosphorus-limited conditions. In crop management, more scientific fertilization strategies could be developed to improve the efficiency of phosphorus use based on the phosphorus requirements of grapes at various phases of development and the SPX genes' pattern of expression. In terms of genetic breeding, it provided a direction for the grape industry to tap into key genetic resources. It might be possible to genetically alter the SPX gene to produce grape varieties that are more resilient to abiotic stressors and hormone response, hence boosting grape viability under unfavorable conditions.
-
In this study, 16 V. vinifera SPX genes and 21 V. amurensis SPX genes were found to be distributed on 10 chromosomes, respectively. Physicochemical property analyses showed that most of the SPX genes were hydrophobic proteins. The VvSPX and VaSPX genes all contained the SPX structural domain, and, based on their evolutionary ties, could be separated into four subfamilies. Analysis of promoter cis-acting elements revealed that SPX genes contained many cis-acting elements that responded to hormonal and environmental signals, such as the phosphorus deficiency response element P1BS, the light response element BOX4, the defence and stress response cis-acting elements, the growth hormone, salicylic acid, gibberellin, ethylene, abscisic acid, and methyl jasmonate response elements. The qRT-PCR results showed the upregulation level of VvSPX2, VvSPX9, VvSPX10, VvSPX11, VaSPX16, VaSPX1, VaSPX2, VaSPX3, VaSPX11, VaSPX13, and VaSPX21 was relatively high of phosphorus stress, these genes could be candidates for subsequent functional studies. This work will serve as a guide for future investigations into the mechanism and function of the SPX gene family in response to phosphorus stress.
This research was financially supported by the Key Project of Gansu Provincial Natural Science Fund (22JR5RA831).
-
The authors confirm contribution to the paper as follows: experiments conception and design: Mao J, Huang X; experiments supervision: Mao J, Lu X; experiments conducting and draft manuscript preparation: Huang X, Feng Y; plant materials collecting: Huang X, Lu X; experimental data organizition and analysis: Huang X, Gou H, Feng Y; manuscript writing: Huang X; manuscript revision: Mao J, Shao M, Liang G. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Real-time fluorescence quantitative primers for the expression analysis of ‘Pinot Noir' SPX gene family.
- Supplementary Table S2 Information and physicochemical properties of SPX gene family in Vitis vinifera and Vitis amurensis.
- Supplementary Table S3 Prediction of secondary structure of SPX gene family proteins in Vitis vinifera and Vitis amurensis.
- Supplementary Fig. S1 Subcellular localization of SPX protein in Vitis vinifera and Vitis amurensis. The numbers in the bar chart represent the number of subcellular localization.
- Supplementary Fig. S2 Heat map of VvSPX gene expression in different tissues of V. vinifera at different developmental stages. Red and blue could be used to symbolize the intensity of gene expression, with red denoting high intensity and blue denoting low intensity.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Huang X, Lu S, Shao M, Feng Y, Gou H, et al. 2025. Analysis of the SPX gene family in Vitis vinifera and Vitis amurensis and its expression under phosphorus stress. Fruit Research 5: e016 doi: 10.48130/frures-0025-0007 

Figure 1.
Analysis of collinearity within VvSPX and VaSPX species. (a) Collinearity analysis within V. vinifera species. (b) Collinearity analysis within V. amurensis species. The gray line represents all collinear blocks in the V. vinifera and V. amurensis genomes, and the blue line represents gene pairs between VvSPX and VaSPX genes. (c) Colinearity analysis between V. vinifera and V. amurensis SPX genomes. (d) Colinearity analysis of SPX genes in V. vinifera, Arabidopsis, and Oryza sativa. Collinearity between the genomes of V. vinifera, Arabidopsis, and Oryza sativa is depicted by the gray lines in the backdrop. The blue lines show the collinearity between the VvSPX gene, Arabidopsis, and Oryza sativa. (e) Colinearity analysis of SPX genes in V. amurensis, Arabidopsis, and Oryza sativa. The gray lines in the background show the collinearity between the genomes of V. amurensis, Arabidopsis, and Oryza sativa. The blue lines show the collinearity between the VaSPX gene, Arabidopsis, and Oryza sativa.