








-
Bermudagrass (Cynodon spp.) is a perennial grass species of the family Poaceae. It is widely distributed throughout the tropical and subtropical areas of the world. Meanwhile, bermudagrass is a deep-rooted and sod-forming grass, which means it can spread through rooted runners, stolons, rhizomes, and seeds[1]. Owing to its strong vegetative growth, it is tolerant of various environmental stressors[2]. Therefore, bermudagrass is not only a popular turfgrass but also a slope protection plant, and a forage species worldwide[3]. According to the WFO (The World Flora Online,
www.worldfloraonline.org ), the Cynodon genus has a diversification of 15 species. However, in general, bermudagrass is usually divided into three subspecies including African bermudagrass (Cynodon transvaalensis), common bermudagrass (Cynodon dactylon), and hybrid bermudagrass (Cynodon dactylon × C. transvaalensis). Among them, African bermudagrass is diploid, hybrid bermudagrass is triploid, common bermudagrass is usually tetraploid[4]. Additionally, hybrid bermudagrass is the most widely used high quality turfgrass on sports fields, golf courses, and home lawns. It's also an important feed source for ruminants[5]. Hence, with these properties bermudagrass plays crucial roles in animal husbandry and horticulture.As a sessile organism, C. dactylon is affected by several environmental factors. Previous literature has depicted that light, water, temperature, soil, and microorganisms can affect its growth and development[6]. More recently the adverse effects of environmental stresses on bermudagrass and its response mechanisms have become research hotspots. The development of research technology significantly contributes to the systemic investigation of stress response mechanisms in plants. Here, the progress on stress response growth regulation of C. dactylon in the last decade was reviewed, and future perspectives proposed.
-
Throughout the life cycle of higher plants, almost all natural factors, especially light, temperature, water, and soil affect their growth and survival (Fig. 1). Bermudagrass is a warm-season and heliophile grass species which means that low temperature and light are crucial factors that limit its establishment and growth. In addition, heat, drought, and nutrient deficiency will also metabolically disturb and cause growth inhibition of bermudagrass. Generally, the stress conditions are classified as abiotic and biotic. The effects induced by different stresses in Cynodon dactylon have been thoroughly investigated over the past ten years.
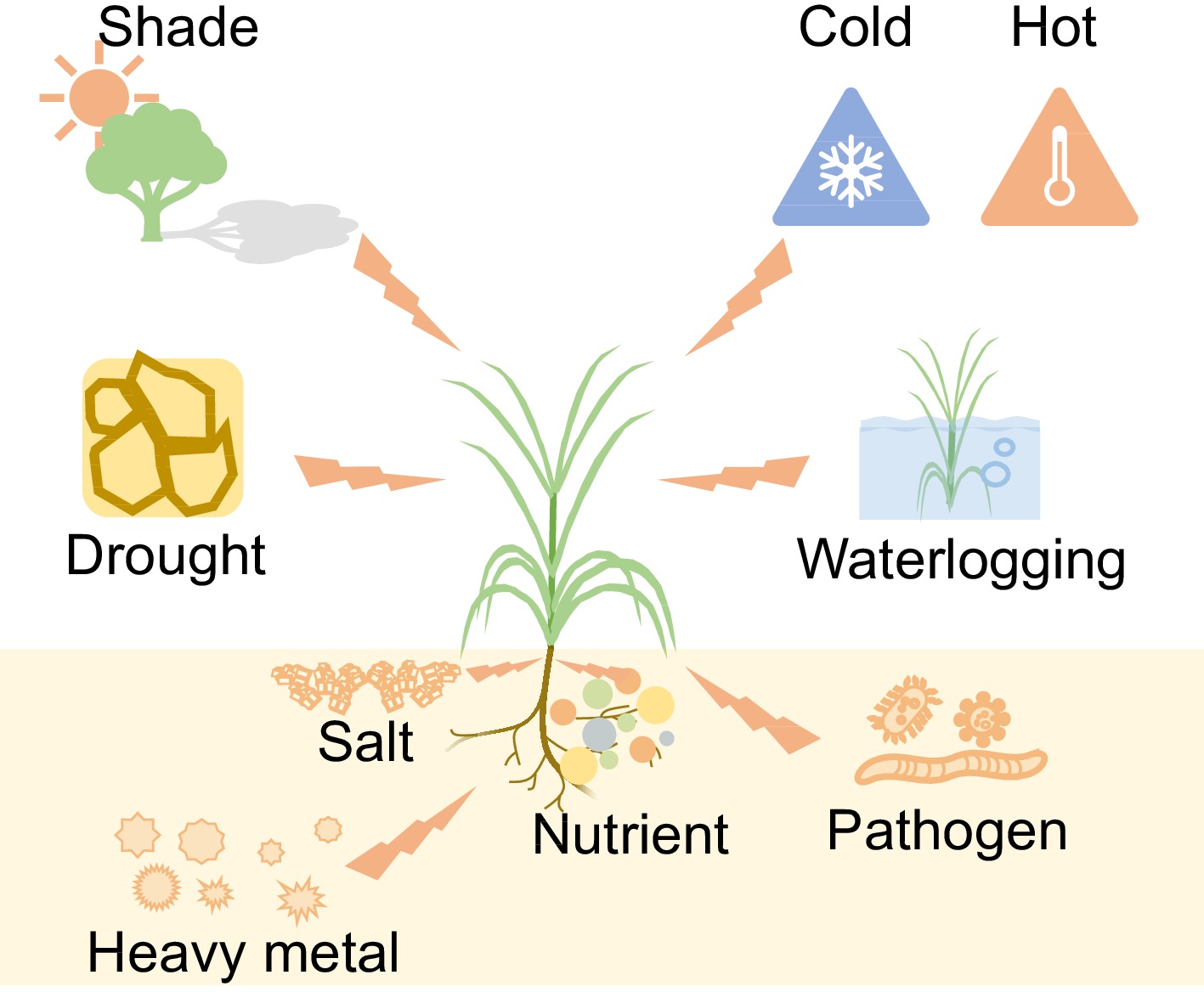
Figure 1.
Environmental stresses that affect the growth and development of bermudagrass.
Abiotic stresses
Light
-
Light, being a signal, is an indispensable environmental factor that influences turfgrass growth through plant morphogenesis and photosynthesis. It can induce, regulate, and promote plant growth and cell differentiation. Light can regulate the growth and development of plants via light intensity, quality, and photoperiod[7]. Light deficiency is also termed as shade stress, it can affect the development of the stems, stolon, and leaves of bermudagrass. For example, the stem diameter and internode length of bermudagrass decreased, but the leaf length increased under shade stress[8]. As well as morphology, physiology is also affected by shade stress. The photosynthetic pigment contents and photosynthesis efficiency of bermudagrass decreased following shade treatment[9]. The investigation on shade stress response of different bermudagrass varieties showed that the tetraploid common bermudagrass cultivar 'Chuanxi' has higher shade tolerance than that of the triploid bermudagrass cultivars 'Tifdwarf', 'Tifsport', and 'Tifway'. Given the rich germplasm resources of common bermudagrass worldwide, it is feasible to select light-tolerant genotypes. A study found that darkness can induce rapid senescence of bermudagrass and upregulation of numerous genes including plant hormones signal transduction, N-glycan synthesis, and transcription factors[10]. Photoperiod also affects the development of bermudagrass. Long photoperiods not only promote the growth and nutrient uptake but also induces precocious flowering in both common and African bermudagrass[3].
Heat and cold stress
-
Since bermudagrass is a warm-season grass species, cold stress is a limiting factor for its growth and development. The cold tolerance varies from species to species. For example, 'Chuannong-3', a new hybrid bermudagrass cultivar, is more tolerant to cold stress than 'Tifdwarf', a popular hybrid bermudagrass cultivar[11]. It is well-known that the phenotype is genotype-specific. An investigation on cold tolerance of different bermudagrass genotypes revealed that the cold-related genes expressed faster in cold-tolerant genotypes as compared to cold-sensitive genotypes. Further studies showed that the DEGs (differential expression genes) in bermudagrass after cold treatment are enriched in photosynthesis, nitrogen metabolism, and carbon fixation pathways, which suggested that these pathways might be crucial in the cold stress response of bermudagrass[12].
To improve the cold stress tolerance of bermudagrass, investigation of its cold stress response mechanisms in bermudagrass is necessary. Recent studies have found that some compounds can increase the cold stress tolerance of bermudagrass. ABA is one of the most crucial hormones to regulate the growth and development, and abiotic stress response of the plants. Application of exogenous ABA can alleviate the cold-induced damage in bermudagrass via regulation of photosystem II[13,14]. Ethylene is another key hormone in plants, but the effects of ethylene on the stress response of plants are controversial; it may play both negative or positive roles in different plants under different stresses. Studies on bermudagrass suggested that the ethylene precursor ACC (1-aminocyclopropane-1-carboxylic acid) contributes to decreasing its cold stress tolerance via regulating the antioxidants, photosystem, and CBF transcriptional regulatory cascade[15]. This implies that ethylene should play a negative role in the cold stress response of bermudagrass. Melatonin also accumulates in C. dactylon under cold conditions. The cold-tolerant genotype accumulates more melatonin and other metabolites, including carbohydrates, organic acids, and amino acids, than that in the cold-sensitive genotype after the application of exogenous melatonin. Besides, the photosystem of bermudagrass is improved by melatonin under cold stress, which suggests that melatonin enhances the cold tolerance of bermudagrass through regulation of the photosystems[16]. Except for hormones, some ions are also involved in its cold stress regulation. Calcium is a secondary messenger in the signaling networks of the cells that play comprehensive roles in developmental and in environmental stimuli response regulation. Chilling stress could induce an increase of the endogenous Ca2+ in bermudagrass. Besides, exogenous CaCl2 could improve the cold stress tolerance of bermudagrass through activation of antioxidants and regulation of the related proteins and metabolic homeostasis[17].
Compared to cold stress, the study of the heat stress response in bermudagrass is limited. Global climate changes induce elevation of temperature, and the heat stress will cause damage to the plants. Although bermudagrass is a warm-season grass, extremely high temperatures can induce injury in it. Global warming is usually accompanied by an increase in CO2 concentration and high concentrations of CO2 contribute to the alleviation of heat stress. High temperature causes damage to several metabolic pathways including light reaction, carbon fixation, TCA cycle, and amino acid metabolism[18]. Further investigation by transcriptomic analysis revealed that the redox-related genes are upregulated rapidly in bermudagrass after heat stress treatment. Meanwhile, genetic research found that CdF-box, the E3 ubiquitin ligase-related gene, played an important role in improving the heat stress tolerance of transgenic plants[19]. miRNAs are also involved in regulating the heat stress response in bermudagrass. The expression of miRNA159a, miRNA160a, and miRNA164f, and their target genes as GAMYB, ARF17, and NAC1, respectively, in leaf and roots of bermudagrass were changed significantly[20].
Drought and submergence stress
-
Drought or water-deficit stress is a major environmental condition that limits plant distribution and growth in worldwide. The drought stress response mechanisms of bermudagrass have been studied extensively in the last decade. Comparative proteomic analysis of drought-tolerant and sensitive genotypes of bermudagrass suggested that many proteins are related to photosynthesis, glycolysis, N-metabolism, tricarboxylicacid (TCA) and redox pathways are involved in drought response regulation[21]. Under water deficit conditions, the tolerant genotypes showed over-expression of a dehydrin gene CdDHN4, antioxidant genes Cu/ZnSOD and APX, which leads to higher antioxidant activities to scavenge the excessive reactive oxygen species and minimize the membrane damage[22]. Contrary to drought, submergence occurs when the water supply is far beyond the threshold. Therefore, submergence-induced damage is caused by the limited oxygen availability and the inhibition of respiration. To adapt to the adverse conditions, schizo-lysigenous aerenchyma is formed in bermudagrass roots via PCD (programmed cell death)[23]. Transcriptome analysis revealed that genes related to chlorophyll biosynthesis, light-harvesting, and carbon fixation are down-regulated in bermudagrass after submergence treatment. During aerenchyma development, the genes related to ethylene signaling, Ca2+-ROS signaling, and cell wall modification are up-regulated after submergence treatment[24]. Bermudagrass is a typical clonal plant and its physiological integration can alleviate the damage induced by stress conditions. This excellent characteristic is also contributing to improve the flooding tolerance of bermudagrass. Under flooding conditions, the growth inhibition and carbohydrate accumulation were relieved by physiological integration[25]. The comparative proteomic and metabolomic analysis showed that many proteins are involved in drought and submergence regulation. Interestingly, most metabolites are upregulated by drought stress, while most are downregulated by submergence stress. This implies that drought stress can promote photosynthesis and redox metabolisms, but submergence will induce bermudagrass dormancy[26].
Salinity and alkali stress
-
Soil salinization and alkalization is a major problem that limits plant distribution across the world. When the plants are exposed to salinity and alkali stress, the growth and development is severely inhibited[27]. The damage induced by salt stress are mainly in three ways including osmotic stress, ionic stress, and secondary stress[28]. Similarly, alkali stress also affects the plants in these ways. Whereas, compared to salt, alkali stress has a more aggravated effect on plants due to the high pH[29]. Between these two stresses, salt stress has received more attention in the study of bermudagrass. It is well known that long-term exposure to adverse environments will modify the phenotypic characteristics of the plant. In bermudagrass, a saline environment causes anatomical modifications. The leaf sheath thickness and vascular bundle area are significantly increased to avoid water loss in bermudagrass under salt stress. Also, the bermudagrass from saline waterlogged areas also showed a high cortical region and strong aerenchyma in the root tissue to increase the bulk movement of salt[30]. Interestingly, different tissues showed diverse modifications in bermudagrass after salt treatment. The growth rate of shoots decreased and roots were increased, under saline conditions. Further study by metabolomic analysis revealed that the nitrogen metabolism pathway regulates the equilibrium between shoots and roots in bermudagrass[31]. Like other metabolites, hormones are reprogrammed in different genotypes of bermudagrass under salt conditions. For example, the contents of trans-zeatin riboside (tZR), and dihydrozeatin riboside (DHZR) were increased, but the content of ABA was decreased in salt-tolerant genotypes[32]. These results suggested that growth could be regulated by changing phytohormones in bermudagrass under salt stress. Furthermore, transcriptome analysis showed that many transcription factors, such as MYB, bHLH, and WRKY are involved in the salt stress response in the root tips of C. dactylon[33]. Further investigation by the time-course transcriptome analysis of bermudagrass root under salt stress suggested that the expression of salt stress-response genes changes with the stress duration. The signaling receptor kinase, transcription factors, tetrapyrrole synthesis, and lipid metabolism-related genes are upregulated rapidly after salt treatment, while, simple phenols, glucosinolates, isoflavones, and tocopherol biosynthesis-related genes changed slowly[34]. Apart from salt stress, alkali stress can induce the accumulation of higher carbohydrate concentrations in bermudagrass[35]. This implies that alkali stress could also affect the growth of bermudagrass but the molecular mechanisms are currently obscure.
Improving the salt stress tolerance of bermudagrass is also an important work in horticulture and forage studies. Given that the damages induced by salt stress in bermudagrass were significantly ameliorated after inoculation with the fungus Aspergillus aculeatus[36], and bacteria Enterobacter ludwigii B30[37], the rhizospheric microorganisms are usually applied to alleviate the injury induced by salt stress in bermudagrass.
Heavy metal stress
-
Heavy metals usually causes toxic effects in plants, and it has become a serious environmental problem for agriculture. Investigating the mechanisms of plant's response to heavy metal stress is necessary to improve the stress tolerance and phytoremediation of the contaminated soils. The heavy metal stress response in bermudagrass has been studied in recent years. Cd is one of the most phytotoxic metal pollutants in the environment due to its high mobility in the soil and it easily being transferred into the food chain. It can affect the survival, reproduction, and migration of plants. The Cd stress tolerance is usually different among various bermudagrass germplasms[38]. Cd stress tolerance diversity analysis of wild bermudagrass germplasms in China showed that the accessions with similar Cd tolerance were distributed in adjacent geographic regions. The genetic relationship analysis of these bermudagrass germplasms suggested that the bermudagrass accessions from adjacent geographic regions are clustered together. However, the accessions from Cd-Cd-contaminated regions have high genetic diversity[39]. This implies that Cd stress is the selective pressure of wild bermudagrass under a natural environment. The translocation rate of Cd in plants is related to their Cd tolerance because some studies reported that the Cd-sensitive genotype transported more Cd from root to shoot than the Cd-tolerant genotype. The metabolome analysis revealed that numerous metabolites, such as norvaline, glycine, proline, glyceric acid, and xylulose are related in the Cd response in C. dactylon[40].
Interestingly, soil nutrient heterogeneity can affect the Cd absorption of bermudagrass. The soils enriched with ammonium nitrogen (NH4+-N) and available phosphorus have a higher ability to enhance the Cd absorption and translocation from root to stem and leaf tissues than soils enriched in nitrate nitrogen (NO3−-N)[41]. Different from other heavy metals, DU (depleted uranium) is the residual of a uranium source after the enrichment process. It has lower radioactivity than natural uranium, but it causes chemical toxicity to organisms. Evaluation of DU tolerance of bermudagrass and purple three awn (Aristida purpurea) suggested that bermudagrass is very sensitive to DU, and the main DU enrichment tissue is the root[42]. Besides, zinc (Zn) is a micronutrient necessary for plant growth and development, but excessive concentrations of Zn will hinder the growth. The investigation of Zn tolerance of different bermudagrass cultivars revealed that common bermudagrass has a stronger tolerance of Zn than hybrid bermudagrass[43].
However, some compounds and microorganisms are reported to alleviate the injuries induced by heavy metals. For example, after the application of exogenous melatonin, the biomass and the Pb accumulation in the roots of bermudagrass increased remarkably. This suggests that melatonin can increase Pb stress tolerance and its potential use in phytoremediation of lead pollution soils[44,45]. Simultaneously, some gaseous signaling molecules, such as endogenous NO (nitric oxide) and H2S (hydrogen sulfide), are induced by Cd stress tolerance upon exogenous application of these molecules. Interestingly, the NO signal can be blocked by both NO and H2S inhibitors, but the H2S signal can only be blocked by H2S inhibitors. This implies there are some associations between these two signal pathways to regulate the Cd stress response in bermudagrass[46]. In addition, soil microorganisms, such as AMF (arbuscular mycorrhizal fungi) and PGPR (plant growth-promoting rhizobacteria), can influence the heavy metal stress adaptive capacity of the plants via mobility and bioavailability of the metals. The AMF of Funneliformis mosseae, and Diversispora spurcum have proved to contribute to increase the translocation factor of Zn in bermudagrass[47]. Some Cd-tolerant AMF are also isolated by ribosomal internal transcribed spacer region sequence analysis from Cd contaminated soil, and they are involved in regulation of the Cd stress response. Among these AMFs, Penicillium janthinellum is found to facilitate plant growth and increase Cd uptake in the shoot and root of bermudagrass under Cd stress conditions[48]; and Aspergillus aculeatus improve the Cd stress tolerance by increasing the photosynthetic activity and decreasing the Cd transportation from root to shoot[49]. The Cd-tolerant fungus, A. aculeatus, can improve the Cd stress tolerance of bermudagrass through regulation of metabolites, i.e., the contents of citric acid and sucrose are decreased but glucose content is increased by A. aculeatus under Cd stress conditions[50]. Additionally, the AMF Rhizophagus irregularis can be used to improve the Cr stress tolerance of bermudagrass[51]. Similar to AMF, PGPR is also an excellent candidate to facilitate the uptake of heavy metals from contaminated soils. When inoculated with Bacillus cereus, which is the popular genera of rhizobacteria that promote plant development, the uptake of various heavy metals, including Cu, Ni, Pb, As, Cd, and Zn in bermudagrass are significantly improved[52]. Interestingly, although antagonistic interactions are found between AMF and PGPR or between AMF and earthworms on the growth of plants, the triple inoculation of AMF with PGPR and epigeic earthworms can enhance Pb bioaccumulation and uptake in bermudagrass[53].
Nutrient stress
-
The main purpose of fertilizer is to maintain the turf stand, quality, and growth regulation. However, a large portion of fertilizer is wasted in the practice of turfgrass maintenance. To achieve the best effect with the least amount of fertilizer is crucial in turfgrass management. Therefore, investigating the nutrient deficiency response of turfgrass is very important. Nitrogen (N) is one of the most important nutrients for plants. The tolerance to low N stress is different among the wild bermudagrass germplasms. In the low-N-tolerant genotype, the genes related to protein synthesis, such as ribosomes and aminoacyl-tRNA synthetase, are upregulated under low N stress. While, most of these genes are downregulated in the low-N-sensitive genotype, which causes decreased protein content in the plant[54]. Interestingly, the abilities of uptake, assimilation and remobilization of nitrogen (N) are improved in bermudagrass after a small N treatment[55]. In addition, some substances are found to be involved in regulating nutrient deficiency stress. With the application of exogenous melatonin, the accumulation of K+ are increased in the shoot and root tissues of bermudagrass, and photosynthesis is improved[56]. Microbes are also involved in nutrient deficiency stress response regulation in bermudagrass. For instance, the AMF A. aculeatus, which is isolated from the rhizosphere of bermudagrass in Cd-contaminated areas, contributes to facilitating the uptake of N, P, and K in bermudagrass under K or P deficient conditions[57].
Biotic stresses
-
In addition to abiotic, biotic stresses also induce severe injuries in bermudagrass. Nematodes are one of the most prevalent pathogens that can cause serious damage to plants. The resistance against nematodes varies in different bermudagrass cultivars. The microbial community structures of bermudagrass cultivars with low resistance ('Latitude 36') and high resistance ('TifTuf') to nematodes are different[58]. This interesting result implies the important function of microbes in regulating the adaptation of biotic stress conditions in bermudagrass. Besides, mole crickets (Neoscapteriscus spp.) usually induce damage to grass roots and disrupt the soil structure via tunneling behavior. Improving the resistance to injurious pathogens and insects is an important research orientation in turf management. Recently, PGPRs have been reported to be involved in increasing the white grub resistance of hybrid bermudagrass[59]. The rhizobacteria contributes to increasing resistance of bermudagrass to tawny mole crickets (Neoscapteriscus vicinus Scudder)[60].
In addition, the combination of different stresses may have different effects on bermudagrass. For example, reduction of N level in bermudagrass will alleviate the salt stress-induced damage[61]. Cold priming could improve the salt stress tolerance of bermudagrass[62]. However, the combined stress of cold and salt will induce severe damage in bermudagrass[63]. Currently, studies that focus on the effects of combined stresses on bermudagrass are not sufficient; given the interesting results that are reported, it is worthy of further investigation.
-
Phenotypic plasticity is an important trait for plants to adapt to their environment. Morphological variation is associated with the climate and the geographical regions. The morphological traits of bermudagrass are significantly affected by geographic distribution and environmental factors. The investigation on wild bermudagrass germplasms in China showed that the size of the bermudagrass that originates from high and low latitudes are usually larger than that of mid-latitudes[64]. Recently, the variations of morphological traits in bermudagrass germplasms of China have been observed along longitudes. The leaf length, leaf width, internode length, and stolon length are correlated with the different environments along the longitudes[65]. Moreover, physiological integration is usually a specific characteristic of clonal plants[66], and bermudagrass has flourishing stolon systems, so it is a typical clonal organism. This interesting characteristic enables the plastic response of bermudagrass to various environmental conditions, and the physiological integration enhances the salt stress tolerance of bermudagrass. Although the Na+ is not translocated among the ramets, the salt stress response-related genes, such as SOS and antioxidant activities, such as CAT and POD, are upregulated in bermudagrass under salt stress conditions[67]. Since adverse conditions will induce stomatal closure, which decreases CO2 supply and consequentially increase the δ13C (stable carbon isotope) in plants. When exposed to cold stress, the δ13C is observed to increase in bermudagrass[14]. Recently, several indexes have been used to evaluate the morphological traits of bermudagrass. Radiation reflectance is usually used to estimate the health status of plants under different conditions[68], and the normalized difference vegetation index (NDVI) is commonly used as a reflectance index. Besides, the water index (WI) and normalized difference water index (NDWI) are robust indexes of plant water content. Through investigation of hybrid bermudagrass with a handheld spectroradiometer under drought stress, it is found that the spectral reflectance WI (900/970) and WI/NDVI are effective indicators of water stress[69]. The plant cuticle is composed of polyester scaffolds and corneum which are a crucial protective barrier to avoid mechanical damage induced by environmental stresses in plants. A study on the cuticular wax of bermudagrass revealed that its characteristics are variable in different germplasms, and the change of cuticular wax is associated with different habitats[70]. In addition, because of its high palatability, bermudagrass is preferred by cattle. Hence, the selection of forage-type bermudagrass is important for husbandry. Through comprehensive analysis of the agronomic characteristics, such as biomass and feed quality, five germplasms of bermudagrass are selected as candidates for high-quality feed breeding[71]. Besides, the morphological traits of bermudagrass are usually used to assess the phenotypic diversity and characterize the novel germplasms. Through QTL (quantitative trait loci) marker-trait association analysis of 110 individuals of hybrid bermudagrass, 74 QTLs that are associated with canopy height, internode length, leaf blade length, and leaf blade width are identified[72].
Molecular markers and omics techniques
-
With the rapid development of molecular biological techniques, the investigations in breeding and molecular regulation of bermudagrass have made advancements in recent years. Not being influenced by environmental conditions, molecular markers are a useful tool to evaluate the genetic diversity of plants rather than morphological and isozyme markers. Nowadays, different types of molecular markers are applied in the genetic diversity analysis of bermudagrass. The genetic diversity and population structure analysis of bermudagrass germplasms in China with SRAP (sequence-related amplified polymorphism) markers revealed that they are genetically diverse, and can be clustered into different sub-populations. The germplasms originate from the same or nearby geographic locations are usually clustered into the same group[73,74]. Like SRAP markers, the SSR (simple sequence repeat) markers are another molecular marker which are widely used in the investigation of inheritance patterns, linkage analysis, and QTL mapping in plants. The large set of codominant SSR markers of African bermudagrass and common bermudagrass are developed sequentially[75,76]. The analysis of the qualitative inheritance mode in common bermudagrass using the progenies of two tetraploid variations, 'Zebra' and A12359, with SSR markers revealed that the tetraploid common bermudagrass has an allotetraploid genome with two distinct sub-genomes[77]. Moreover, five genomic regions are identified to be related to the ground coverage phenotype with the SSR markers[76]. In addition, because of the primer length and high annealing temperature, ISSR (inter-simple sequence repeat) markers can produce enough reliable and reproducible bands to detect the polymorphism DNA sequence information. Hence, it is appropriate to perform a genetic analysis of bermudagrass. The ISSR analysis of bermudagrass germplasms in Iran suggested that the germplasms can be clustered into six groups[78]. Before the whole genome identification of bermudagrass, the linkage maps were important molecular tools in genetic and genomic research. With the construction of a high-resolution linkage map for African bermudagrass, four genomic regions are identified to be related to sod establishment rate in bermudagrass[79]. Furthermore, a comparison of this linkage map against the genome sequences of Oropetium thomaeum revealed that the intrachromosomal rearrangements of the paleo-chromosome event induced the reduction of chromosome number in African bermudagrass[80]. Additionally, with the technique of genotyping-by-sequencing (GBS), 37,496 raw single nucleotide polymorphisms (SNPs) were developed as genetic markers to characterize the genetic diversity of bermudagrass. Analysis of these markers revealed that the bermudagrass could be divided into four different subpopulations, and the African bermudagrass accessions are mainly clustered into one subpopulation. Considering that there are also several common bermudagrass genotypes clustered in the same subpopulation, this might suggest the polyploidization origin of common bermudagrass[81].
In addition to molecular markers, high-throughput transcriptome analysis is also widely used in molecular regulation mechanisms studies of bermudagrass. Initially, microarray analysis is used to identify the candidate genes. With this method, 587 DEGs are identified in the crown tissue of bermudagrass after freezing temperature treatment, and these DEGs contribute to develop molecular markers in bermudagrass for its cold hardiness breeding[82]. NGS (next generation sequencing) became a popular method for transcriptome analysis subsequently. With this method, many genes were identified to be involved in stem development regulation of bermudagrass. Among these genes, the light- and gravity-responsive genes are preferentially expressed in the prostrate-growing accession of bermudagrass, whereas the lignin synthesis-related genes are usually identified in erect-growing accession[83]. Simultaneously, the organ-specific transcriptome analysis of bermudagrass identified 3,028 organ-enriched unigenes, and 171, 22, and 19 unigenes are specifically expressed in shoot, stolon, and rhizome, respectively[84]. Distinguished from NGS, full-length RNA sequencing is a single-molecule long-read sequencing technology that has been successively used in many species. With this technology, numerous genes and pathways were identified to be associated with bermudagrass flourishment[85]. Moreover, microRNAs (miRNAs) have been reported as regulatory molecules in plants. A total of 536 miRNAs were found to be involved in the salt stress response of bermudagrass. Among them, miRNA171f could increase the salt tolerance by improving the photosynthetic performance[86].
Like transcriptome, the proteome also changes remarkably with changing environmental conditions. The 2-DE (two- dimensional electrophoresis) combined with MALDI-TOF-MS analysis identified 77 proteins related to various pathways, such as photosynthesis, oxidative pentose phosphate, glycolysis, and redox metabolism against drought and salt stress in bermudagrass[87]. The development of high-throughput sequencing also contributes to the proteomic analysis. iTRAQ (isobaric tags for relative and absolute quantitation) is an effective tool to evaluate the changes at the proteomic level. Recently, a total of 228 differentially accumulated protein species (DAPs) have been identified in rhizomes and stolons of bermudagrass. Interestingly, the stolons that grow underground showed different DAP expression as that of rhizomes which implied that light should play a very important role in the organ development of C. dactylon[88]. Also, 376 DAPs were identified in shoots and stolons of bermudagrass. The starch synthesis-related proteins accumulated in stolons, by contrast, glycolysis- and transport processes-related proteins mainly accumulated in shoots[89]. This technology also analyzes the proteomic change induced by melatonin and oxidative stress in bermudagrass. A total of 76 proteins and several pathways, such as polyamine metabolism, major carbohydrate metabolism, photosynthesis, redox status, and amino acid metabolism, are identified to be related to oxidative stress response in bermudagrass[90]. With the advances in proteomic analysis, PTMs (protein post-translational modifications) analysis, such as lysine acetylation, succinylation and phosphorylation, is widely conducted in many plants. The PTMs analysis in bermudagrass revealed that lots of lysine acetylation, succinylation, and phosphorylation sites are related to the growth regulation of the plant[91,92].
Whole genome information is essential for the genetic and molecular investigation of plants. The studies on stress response, growth, and development regulation mechanisms in bermudagrass were limited due to the lack of whole genome information. This situation changed after the identification of bermudagrass genome sequence. At present, there are three versions of the bermudagrass genome, including one version of African bermudagrass and two versions of common bermudagrass. The size of African bermudagrass, a diploid Cynodon species, is 454.4 Mb[4], and the size of common bermudagrass cultivars 'Yangjiang' and 'A12359', two tetraploid Cynodon species, are 984 Mb and 604 Mb, respectively[93,94]. In addition to whole genome, the chloroplast genome of common bermudagrass was also studied. It is reported that the genome structure of bermudagrass chloroplast is similar to other Poaceae species, and the events of genome rearrangements occurred just once after the Poaceae varied from other monocots[95]. The identification of the bermudagrass genome also facilitates its genetic analysis. The genome-wide association studies (GWAS) of 91 wild bermudagrass germplasms based on the RNA-seq data and the genome of African bermudagrass, 16 SNPs were identified to be associated with the plant height and IAA (indole acetic acid) content[96]. The comparative transcriptome analysis of C. dactylon germplasm from 16 different latitudinal gradients revealed that several genes are involved in plant growth, photosystem, and redox system against cold stress. Among the genes, HSP70/90, and HsfA3/A8 may be the key genes in bermudagrass to regulate cold stress tolerance[97].
Genetic transformation system establishment
-
Genetic engineering is the key technology to investigate the regulation mechanisms of interested genes and proteins to produce new varieties of plants. Agrobacterium-mediated transformation is a popular method for transgenic manipulation. It is widely used in the studies of model plants, such as Arabidopsis, Oryza sativa, and Nicotiana benthamiana. However, this method is not effective in all plants. Bermudagrass is one of the plants that is difficult for genetic modification because of its genome constitution and resistance to Agrobacterium. To resolve this problem, researchers focused on establishing an effective and stable genetic transformation system in bermudagrass, and remarkable achievements have been made in previous years. It has been reported that a chlorophyll biosynthesis-related gene, CdHEMA1, was successfully transferred into bermudagrass by Agrobacterium tumefaciens and it enhanced the photosynthetic pigments[98]. Besides, virus-induced gene silencing (VIGS) is also an efficient tool to investigate the function of genes in plants including bermudagrass. For example, the CdERF1 gene is successfully silenced in bermudagrass[99].
-
Although numerous genes are found to be involved in various biotic processes such as growth, development, and stress response regulation by high-throughput sequencing and bioinformatic analysis, their functions should be identified further by transgenic technology. In recent years, many genes related to transcription factors, enzymes, and other proteins have been identified through this technology in bermudagrass (Fig. 2). CdWRKY2 is reported as a key gene that plays an important role in regulating cold stress response in bermudagrass via activating the sucrose-related gene CdSPS1 and the cold response-related gene CdCBF1[100]. However, salt stress sensitivity increased in CdWRKY2 overexpression lines of bermudagrass, which showed growth inhibition of the root system under stress conditions[101]. Besides, another transcription factor, CdERF1 (ethylene-responsive factor 1), also regulates the cold stress response of bermudagrass via regulating expressions of cold-responsive genes, such as CBF2, pEARLI1 (lipid transfer protein), PER71 (peroxidase), and LTP (lipid transfer protein)[99]. TB1 (teosinte branched 1) is an important transcription factor that regulates the development of a plant's branches. The study of this gene in bermudagrass revealed that there are many SNPs and indel mutations in the promoter region and protein-coding region in different germplasms of bermudagrass, and the variations of the TB1 gene are related to the geographical distributions of bermudagrass[102]. NF-Y (nuclear factor Y) is a transcription factor that plays crucial roles in many biological processes regulation, such as growth and development, and abiotic stress tolerance. This protein is a heterotrimer composed of NF-YA, NF-YB, and NF-YC. The gene expression of Cdt-NF-YC1 that is identified in hybrid bermudagrass is induced by different abiotic stresses such as drought, salt, and H2O2. Overexpression of this gene will improve the tolerance against drought and salt stress through ABA-dependent and ABA-independent pathways[103]. Additionally, the photochemical efficiency of photosystem II and K+/ Na+ ratio also improved in Cdt-NF-YC1 transgenic seashore paspalum (Paspalum vaginatum) under saline conditions[104].
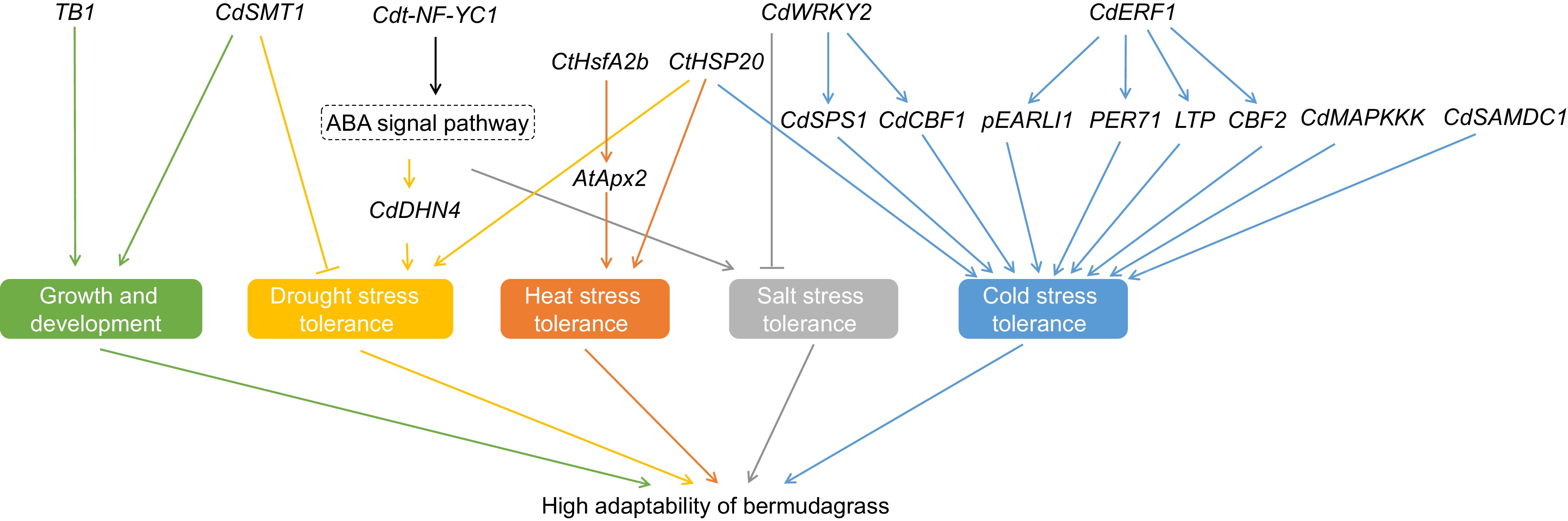
Figure 2.
Genes related to the regulation of environmental stress response and growth in bermudagrass.
Other than TFs several functional genes like DHNs (dehydrins) are LEAII (late embryogenesis abundant II) proteins are also reported. These proteins are thermostable in that they can even maintain their integrity at boiling temperature. Due to this excellent characteristic, DHNs play important roles in against stress tolerance. In 'Tifway' bermudagrass, CdDHN4 is remarkably upregulated by drought stress. Further studies on this gene revealed that it is regulated by the ABA-signaling pathway[105]. Meanwhile, overexpression of CdDHN4 can improve multiple stress tolerance, such as salt, drought, osmotic, and cold, of transgenic plants[106]. Identification of the functions of CdDHN4-L and CdDHN4-S in hybrid bermudagrass revealed that although overexpression of these two genes can increase the abiotic stress tolerance of transgenic plants, the CdDHN4-L transgenic plants exhibited better growth status than that of CdDHN4-S ones. Compared to CdDHN4-L, CdDHN4-S lacks of φ-segment. Hence, the φ-segment plays an important role in dehydrins for protecting plants against abiotic stress[107]. HSP (heat shock protein) is a molecular chaperone and is involved in the regulation of many biological processes in plants including development and stress response. With the assembly of the C. dactylon genome, 41 members of the HSP20 gene family are identified in African bermudagrass. The expression profile analysis of CtHSP20s showed that they play a key role in African bermudagrass resistance against heat, cold, drought, and salt stress[108]. CtHsfA2b, also identified in African bermudagrass, exhibits a rapid response to high temperature. It can improve the heat tolerance of transgenic plants via binding to HSE (heat shock element) on the promoter of AtApx2 (ascorbate peroxidase) and enhance the expression of AtApx2 and other downstream genes[109]. SMT1 (sterol-C-24 methyltransferase 1) is a key enzyme in catalyzing the biosynthesis of phytosterols that play important roles in plant growth and development and in stress defense systems. Investigation on Cdsmt1 mutant line of bermudagrass showed that it has a dwarf phenotype but the drought tolerance enhanced. Besides, the putrescine, spermidine, and spermine are accumulated in the Cdsmt1 mutant line. The results suggest that CdSMT1 regulates the growth and drought stress response via regulating the polyamines[110]. Simultaneously, polyamines, such as sperimidine and spermin, accumulation in plant cells contributes to the stress tolerance of the plants. SAMDC (S-adenosylmethionine decarboxylase) is a key enzyme for sperimidine and spermin formation. Overexpression of CdSAMDC1, a SAMDC gene from bermudagrass, in centipedgrass (Eremochloa ophiuroides) can enhance its cold stress resistance[111]. Besides, the kinase-related genes, such as CdMAPKKK, are enriched in signal transduction, and hormone-mediated signaling pathways are significantly induced by cold treatment[112].
-
Bermudagrass is a widespread plant because of its wide adaptability. It is also an excellent turfgrass species used in parks, lawns, and sports fields. In addition, bermudagrass has flourishing stolons and rhizomes, so becoming suitable for landscape engineering, especially for wind prevention, sand fixation, and soil erosion. Therefore, its environmental adaptation and growth regulation mechanisms are very important for the breeding of high-quality varieties. Although significant progress has been achieved in bermudagrass research in recent years, there are some remaining problems that need to be solved in the future.
The genotypic milieu of bermudagrass germplasms is very complex, and the illumination of its genetic information is a fundamental work for molecular investigations and breeding. The aneuploidies of bermudagrass include diploid, triploid, tetraploid, pentaploid, and hexaploidy[113], and even for tetraploid germplasms; the genomic information is different[93,94]. To reveal the complexity of genetic diversity in bermudagrass, the approaches of whole genome resequencing and pan-genome sequencing can be adopted. Whole genome resequencing has been used in many plants such as chickpea (Cicer arietinum)[114], squash (Cucurbita pepo)[115], and Moso bamboo (Phyllostachys edulis)[116]. Meanwhile, pan-genome sequencing is also widely reported in different plants, including tomato (Solanum lycopersicum)[117], rapeseed (Brassica napus)[118], barley (Hordeum vulgare)[119], and pearl millet (Pennisetum glaucum)[120]. Therefore, the genetic diversity, evolution and adaptation of bermudagrass can be further studied in the future by whole genome resequencing and pan-genome sequencing.
Having a stable and efficient genetic transformation system is the prerequisite for gene function identification and molecular breeding of a plant. However, the transformation system of bermudagrass still needs a breakthrough at present. The transgenic engineering of bermudagrass was initiated more than thirty years ago[121], but the progress is slow as compared to other plants such as Arabidopsis, rice, and alfalfa. The obstacle that hinders the development of bermudagrass genetic transformation may be its complex genomes and genotypes as well as ploidies. Agrobacterium-mediated transformation is preferred in plants including bermudagrass, but the transformation system with high efficiency is still lacking. The fact that transformation efficiency is affected by Agrobacterium strains, plant tissues, genotypes, and culture medium. Hence, the potential work on bermudagrass genetic transformation in the future are isolating an appropriate Agrobacterium strain and exploring an efficient culture medium formulation. Additionally, biolistics is another efficient technology to generate transgenic plants. This method has been applied in bermudagrass previously[122].
Gene functional identification is vital for revealing the molecular regulation mechanisms of growth and development, and in stress response. While, each gene does not function independently, they will form a network via the feedback regulation. Recently, many genes have been identified to play important roles in bermudagrass, but still, their regulation pathways have not been reported in bermudagrass. Numerous gene modules and networks have been reported in plants. For example, the mediator kinase module of CDK8 (cyclin-dependent kinase 8) and MED12 contributes to the regulation of SA (salicylic acid) level in Arabidopsis[123]. OsPUB24 (U-box-type E3 ligase) is involved in OsBZR1 (brassinazole resistant 1) turnover in rice (Oryza sativa)[124]. SlSPX1 (SYG1/Pho81/XPR1) and SlPHRs regulate the arbuscular mycorrhizae symbiosis under different Pi-availability conditions by forming a SlSPX1-SlPHRs Pi-sensing module in tomato (Solanum lycopersicum)[125]. Therefore, identification of the genes or proteins interaction, and sequentially revealing the regulation pathway in bermudagrass is a significant work in the future.
-
The authors confirm contribution to the paper as follows: study conception and design: Yan X, Zhang B; document collection: Fan J, Xiang Y, Zhang B, Zhang J; draft manuscript preparation and revision: Fan J, Xiang Y, Zhang B, Noor M, Zhang J; figure preparation: Fan J, Xiang Y. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
This work was supported by the National Natural Science Foundation of China (Grant Nos 32171672 and 31702165) and the Project of Forestry Science and Technology Innovation and Promotion of Jiangsu (Grant No. LYKJ[2021]09).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fan J, Xiang Y, Zhang B, Noor M, Zhang J, et al. 2024. Progress and prospects of bermudagrass research in the last decade. Grass Research 4: e017 doi: 10.48130/grares-0024-0015 

Progress and prospects of bermudagrass research in the last decade
- Received: 18 April 2024
- Revised: 15 July 2024
- Accepted: 24 July 2024
- Published online: 16 August 2024
Abstract: Bermudagrass is a widely distributed grass worldwide. The well-developed stolons and rhizomes, good palatability, and strong tolerance against diverse environmental stresses collectively, make bermudagrass a high-quality turf and a valuable forage of animals. Recently, with the development of modern experimental techniques, significant progress has been made in bermudagrass studies. The regulation mechanisms of growth and stress responses in bermudagrass are becoming more clear with the use of advanced research strategies. In this review, research progress in environmental stress responses at physiological and molecular levels in bermudagrass are summarized using different strategies. The challenges and perspectives of research on bermudagrass in the future are also proposed. However, as the emphasis of this review is the investigation of growth and environmental adaptation regulation mechanisms in bermudagrass, only basic research not management reports are reviewed.
-
Key words:
- Bermudagrass /
- Environmental stress /
- Regulation mechanism /
- Functional genes