








-
Outline of the order Russulales Order Russulales Kreisel ex P.M. Kirk, P.F. Cannon & J.C. David 2001 Family Albatrellaceae Nuss 1980 Albatrellopsis Teixeira 1993 Albatrellus Gray 1821 (= Ovinus (Lloyd) Torrend 1920, Polyporus sect. Ovinus Lloyd 1911) (Note 1) Byssoporia M.J. Larsen & Zak 1978 (Note 2) Leucogaster R. Hesse 1882 Leucophleps Harkn. 1899 (= Cremeogaster Mattir. 1924, Leucophleps Harkn. 1899) Mycolevis A.H. Sm. 1965 Polyporoletus Snell 1936 (Note 3) Scutiger Paulet 1808 Family Aleurocystidiellaceae Y.L. Deng & C.L. Zhao, fam. nov. (Note 4) Aleurocystidiellum P.A. Lemke 1964 (Note 5) Family Auriscalpiaceae Maas Geest. 1963 Artomyces Jülich 1982 (Note 6) Auriscalpium Gray 1821(= Pleurodon Quél. ex P. Karst. 1881) (Note 7) Dentipratulum Domański 1965 (Note 8) Gloiodon P. Karst. 1879 (= Leaia Banker 1906, Sclerodon P. Karst. 1889) (Note 9) Lentinellus P. Karst. 1879 ( =Hemicybe P. Karst. 1879) Stalpersia Parmasto 2001 Family Bondarzewiaceae Kotl. & Pouzar 1957 (= Hybogasteraceae Jülich 1982) (Note 10) Amylaria Corner 1955 (Note 11) Amylonotus Ryvarden 1975 (Note 12) Amylosporus Ryvarden 1973 (= Rigidoporopsis I. Johans. & Ryvarden 1979) (Note 13) Bondarzewia Singer 1940 (= Hybogaster Singer 1964) (Note 14) Heterobasidion Bref. 1888 (= Spiniger Stalpers 1974, Spongioides Lázaro Ibiza 1916) (Note 15) Laurilia Pouzar 1959 (Note 16) Lauriliella Nakasone & S.H. He 2017 (Note 17) Stecchericium D.A. Reid 1963 (Note 18) Family Echinodontiaceae Donk 1961 (= Amylostereaceae Boidin, Mugnier & Canales 1998) Amylostereum Boidin 1958 (= Lloydellopsis Pouzar 1959, Trichocarpus P. Karst. 1889) (Note 19) Echinodontiellum S.H. He & Nakasone 2017 (Note 20) Echinodontium Ellis & Everh. 1900 (= Hydnofomes Henn. 1900, Hydnophysa Clem. 1909) (Note 21) Larssoniporia Y.C. Dai, Jia J. Chen & B.K. Cui 2015 (Note 22) Subulicystidiella Y.L. Deng & C.L. Zhao gen. nov. (Note 23) Family Gloeocystidiellaceae Jülich 1982 Gloeocystidiellum Donk 1931 (Note 24) Family Gloeodontiaceae Y.L. Deng & C.L. Zhao fam. nov. (Note 25) Gloeodontia Boidin 1966 (Note 26) Family Hericiaceae Donk 1964 Dentipellicula Y.C. Dai & L.W. Zhou 2013 (Note 27) Dentipellis Donk 1962 (= Amylodontia Nikol. 1967) (Note 28) Hericium Pers. 1794 (= Creolophus P. Karst. 1879, Dryodon Quél. ex P. Karst. 1881, Friesites P. Karst. 1879, Hericium Schrank 1786, Hericius Juss. 1789, Manina Banker 1912, Martela Adans. 1763, Martella Endl. 1836, Medusina Chevall. 1826) (Note 29) Laxitextum Lentz 1956 (Note 30) Pseudowrightoporia Y.C. Dai, Jia J. Chen & B.K. Cui 2015 (Note 31) Wrightoporiopsis Y.C. Dai, Jia J. Chen & B.K. Cui 2015 (Note 32) Family Peniophoraceae Lotsy 1907 (= Lachnocladiaceae D.A. Reid 1965) (Note 33) Amylofungus Sheng H. Wu 1996 (Note 34) Asterostroma Massee 1889 (Note 35) Asterostromella Höhn. & Litsch. 1907 Baltazaria Leal-Dutra, Dentinger & G.W. Grif 2018 (Note 36) Dendrophora (Parmasto) Chamuris 1987 (Note 37) Dichostereum Pilát 1926 (Note 38) Duportella Pat. 1915 (Note 39) Entomocorticium H.S. Whitney, Bandoni & Oberw. 1987 (Note 40) Gloiothele Bres. 1920 (Note 41) Lachnocladium Lév. 1846 (= Eriocladus Lév. 1846, Stelligera R. Heim 1938, Stelligera R. Heim ex Doty 1948) (Note 42) Metulodontia Parmasto 1968 Parapterulicium Corner 1952 (Note 43) Peniophora Cooke 1879 (= Cryptochaete P. Karst. 1889, Gloeopeniophora Höhn. & Litsch. 1907, Sterellum P. Karst. 1889) (Note 44) Sceptrulum K.H. Larss. 2014 Scytinostroma Donk 1956 (= Licrostroma P.A. Lemke 1964 = Michenera Berk. & M.A. Curtis 1868) (Note 45) Vararia P. Karst. 1898 (= Denrophysellum Parmasto 1968) (Note 46) Vesiculomyces E. Hagstr. 1977 Family Russulaceae Lotsy 1907 Boidinia Stalpers & Hjortstam 1982 (Note 47) Gloeopeniophorella Rick 1934 (Note 48) Lactarius Pers. 1797 (= Agaricus subdiv. Galorrheus Fr. 1818, Arcangeliella Cavara 1900, Galorrheus (Fr.) Fr. 1825, Gastrolactarius R. Heim 1971, Gastrolactarius R. Heim ex J.M. Vidal 2005, Gloeocybe Earle 1909, Hypophyllum Earle 1909, Lactariella J. Schröt. 1889, Zelleromyces Singer & A.H. Sm. 1960) (Note 49) Lactifluus (Pers.) Roussel 1806 (= Lactariopsis Henn. 1901, Pleurogala Redhead & Norvell 1993) (Note 50) Multifurca Buyck & V. Hofst. 2008 (Note 51) Pseudoxenasma K.H. Larss. & Hjortstam 1976 (Note 52) Russula Pers. 1796 (= Bucholtzia Lohwag 1924, Cystangium Singer & A.H. Sm. 1960, Dixophyllum Earle 1909, Elasmomyces Cavara 1897, Gymnomyces Massee & Rodway 1898, Hypochanum Kalchbr. 1876, Lactarelis Earle 1909, Macowanites Kalchbr. 1882, Martellia Mattir. 1900, Omphalomyces Battarra ex Earle 1909, Phaeohygrocybe Henn. 1901, Russulina J. Schröt. 1889) Family Stereaceae Pilát 1930 (Note 53) Acanthobasidium Oberw. 1965 (Note 54) Acanthofungus Sheng H. Wu, Boidin & C.Y. Chien 2000 Acanthophysellum Parmasto 1967 (Note 55) Acanthophysium (Pilát) G. Cunn. 1963 Aleurobotrys Boidin 1986 Aleurodiscus Rabenh. ex J. Schröt. 1888 ( =Aleurobotrys Boidin 1985, Aleurodiscus Cooke 1885, Aleurodiscus Rabenh. 1874, Gloeomyces Sheng H. Wu 1996, Gloeosoma Bres. 1920, Nodularia Peck 1871) (Note 56) Aleuromyces Boidin & Gilles 2002 Amylohyphus Ryvarden 1978 Amylosporomyces S.S. Rattan 1977 (Note 57) Conferticium Hallenb. 1980 (Note 58) Confertotrama Nakasone & S.H. He 2025 (Note 59) Coniophorafomes Rick 1934 Dextrinocystidium Sheng H. Wu 1996 Gelatinostereum S.H. He, S.L. Liu & Y.C. Dai 2025 (Note 60) Gloeocystidiopsis Jülich 1982 (Note 61) Gloeomyces Sheng H. Wu 1996 (Note 62) Gloeosoma Bres. 1920 (Note 63) Megalocystidium Jülich 1978 (Note 64) Neoaleurodiscus Sheng H. Wu 2010 (Note 65) Scotoderma Jülich 1974 Stereodiscus Rajchenb. & Pildain 2021 (Note 66) Stereum Hill ex Pers. 1794 (= Haematostereum Pouzar 1959) (Note 67) Xylobolus P. Karst. 1881 (Note 68) Family Terrestriporiaceae Y.C. Dai, B.K. Cui, F. Wu, Y. Yuan & Jia J. Chen 2020 (Note 69) Terrestriporia Y.C. Dai, B.K. Cui, F. Wu, Y. Yuan & Jia J. Chen 2020 (Note 70) Family Wrightoporiaceae Jülich 1982 Wrightoporia Pouzar 1966 (Note 71) Russulales genera incertae sedis Dentipellopsis Y.C. Dai & L.W. Zhou 2013 (Note 72) Dichantharellus Corner 1966 Dichopleuropus D.A. Reid 1965 Gloeoasterostroma Rick 1938 Gloeohypochnicium (Parmasto) Hjortstam 1987 (Note 73) Haloaleurodiscus N. Maek., Suhara & K. Kinjo 2005 (Note 74) Laeticutis Audet 2010 Neoalbatrellus Audet 2010 (Note 75) Perplexostereum Ryvarden & S. Tutka 2014 (Note 76) Polypus Audet 2010 Scopulodontia Hjortstam 1998 Scytinostromella Parmasto 1968 (= Confertobasidium Jülich 1972) Xeroceps Audet 2010
HTML
-
Russulales Kreisel ex P.M. Kirk, P.F. Cannon & J.C. David is one of the significant fungal orders, comprising wood-inhabiting fungi within Agaricomycetes, Basidiomycota [ 1− 4] . The concept of Russulales was initially proposed in 1969, and later accepted and amended by Kirk [ 4] , and it was erected with the type family Russulaceae Lotsy [ 4] . Approximately 4,500 species have been ascribed to Russulales, which contains 98 genera, within 11 families [ 1, 4− 7] . Members of Russulales exhibit a wide range of ecological strategies including saprotrophic wood rotting, tree root and heartwood pathogens, ectomycorrhizal, and association with insects as entomogenous fungi [ 4, 8] . The genus Heterobasidion Bref. was indigenous to many areas and an important ecological factor involved in nutrient cycling, forest regeneration, and forest succession, such as Heterobasidion occidentale Otrosina & Garbel. and H. irregulare Garbel. & Otrosina as entry plant quarantine fungi in North America, and H. abietinum Niemelä & Korhonen, H. annosum (Fr.) Bref., and H. parviporum Niemelä & Korhonen as entry plant quarantine fungi in Europe [ 4, 9− 13] . Species of Albatrellus have been reported as significant ectomycorrhizal fungi [ 4, 14] . They also formed mutualistic symbiotic relationships with plants, especially Pinaceae [ 4, 15] , and some angiosperms [ 4, 16] , aiding in nutrient uptake and enhancing plant growth [ 17] . Albatrellus ovinus (Schaeff.) Kotl. & Pouzar is a common edible mushroom reported in Europe and North America [ 17− 19] .
Russulales is the most conspicuous and largest group of macrofungi, characterized by highly diverse basidiomata types which can range from agaricoid to discoid, clavarioid, polyporoid, corticiod, and even gasteroid [ 1, 9− 11, 20− 22] . Among Russulales, the hymenophore types encompass lamellate, hydnoid, poroid, labyrinthoid, grandinioid, and a smooth configuration [ 17, 23, 24] . Many species of Russulales share two key morphological characteristics: (1) basidiospore walls exhibit an amyloid reaction combined with an ornamented outline (although there are a few exceptions that have non-amyloid or smooth walls); (2) many of these species possess gloeoplerous hyphae or gloeocystidia containing sesquiterpenes that can be stained by sulphuric benzaldehydes [ 25, 26] . However, it is worth noting that in certain genera within the order, these elements may be absent or transformed into homologous structures like lactiferous hyphae [ 1] .
The morphology of the basidiomata and hymenophore, together with habitat, are often regarded as important characteristics of the order Russulales [ 26− 28] . Resupinate basidiomata are common in Echinodontiaceae Donk (=Amylostereaceae Boidin, Mugnier & Canales), Peniophoraceae Lotsy, Stereaceae Pilát and Terrestriporiaceae Y.C. Dai, B.K. Cui, F. Wu, Y. Yuan & Jia J. Chen, and rarely present in Hericiaceae Donk and Russulaceae Lotsy [ 3, 21, 29− 34] . Pileate basidiomata are predominantly found in Albatrellaceae Nuss, Auriscalpiaceae Maas Geest., Bondarzewiaceae Kotl. & Pouzar, Hericiaceae and Russulaceae; while gasteroid basidiomata are mainly present in Hybogasteraceae Jülich and Russulaceae, rarely in Albatrellaceae [ 17, 35] . Clavarioid basidiomata are present in Auriscalpiaceae, Bondarzewiaceae and Peniophoraceae, while effused-reflexed basidiomata are infrequently scattered in Auriscalpiaceae, Echinodontiaceae, Hericiaceae, Peniophoraceae, and Stereaceae [ 1, 3, 12, 25, 28] .
Fungal classification was traditionally based on comparative morphology, anatomy, biochemistry, physiology, and ecology [ 2, 5, 6, 36, 37] . The systematics of fungi has been revolutionized by advancements in molecular biology, phylogenetics, and bioinformatics. Nowadays, morphology integrated with DNA sequence-based classification and identification has become the standard approach in fungal taxonomy [ 35, 38− 43] . In the past few decades, evidence supporting the monophyly of Russulales has been gathered through both morphological characteristics and DNA sequence phylogenetic analyses [ 1, 6, 10, 42] . Miller et al. [ 27] first explored the molecular phylogeny of Russulales with an emphasis on the agaricoid, gasteroid and pleurotoid taxa in family Russulaceae, according to the current accepted classification [ 25] , which indicated that these taxa were not clustered strictly according to their basidiomata morphology. Larsson & Larsson [ 26] subsequently sampled some additional species producing corticioid basidiomata, and the resulting phylogeny suggested 13 major clades in the order with strong or moderate bootstrap support, as /peniophorales, /amylostereaceae, /gloeocystidiellum I, /gloeocystidiellum II, /auriscalpiaceae, /gloeodontia, /aleurocystidiellum, /hericiaceae, /bondarzewiaceae, /albatrellus, /scytinostromella, /russulales, and /stereales. Following these studies, Russulales was classified into 12 families and 80 genera on the basis of molecular and morphological characters. However, many taxa remain unsampled in phylogenetic analyses of Russulales, and phylogenies of many sampled taxa were not resolved. Some genera such as Gloeocystidiellum Donk and Wrightoporia Pouzar, are morphologically well defined, whereas they were inferred as polyphyletic or paraphyletic [ 28] . Previous research suggested that species of Wrightoporia s.l. were mainly placed in the Bondarzewiaceae clade, and the Wrightoporiaceae clade based on ITS and nLSU sequences [ 10, 26, 44] . Phylogenetic studies have supported the monophyletic status of only a few genera, such as Aleurocystidiellum P.A. Lemke and Wrightoporia, and these genera correspond to two families of their own [ 11, 21, 45, 46] . Other genera still require deeper analysis, since they were either intermixed with other taxa or possess or lack characters that intermingle with those present in other genera [ 1, 19, 21, 46] . Thus, the question remained as to whether to treat them in a broad or narrow sense [ 46] .
Some studies have shown a relatively constant relationship between differences in the amino acid sequences and divergence time of species [ 47, 48] . Recently, divergence time was used as an important criterion for the classification and estimation of evolutionary time of Basidiomycota [ 1, 5, 13, 49− 51] . The phylogenomic analysis suggested that the subphyla diverged in a time range of 443–490 Mya (million years), classes in a time range of 312–412 Mya, and orders in a time range of 102–361 Mya, families diverged in a time range of 50–289 Mya, 76–224 Mya, and 62–156 Mya in Agaricomycotina, Pucciniomycotina, and Ustilaginomycotina, respectively [ 1] . The Bayesian evolutionary analysis based on the combined ITS + nLSU + rpb1 + rpb2 + tef1- α dataset indicated an ancient divergence of the family Terrestriporiaceae from Albatrellaceae during the Cretaceous (124.68 ± 0.39 Mya (million years ago)) [ 21] . Nevertheless, there is no comprehensive study focused on the divergence times of Russulales and its lower classification ranking.
According to geographical regions, numerous new fungal species have been found in southwest China and low-latitude tropical and subtropical regions [ 20] . These regions have many hills and mountains, extensive vegetation coverage, a high fungi species richness, a mild climate, and abundant rainfall, all of which are conducive to fungal growth, and contribute to a rich species diversity [ 20, 52] . In recent years, several studies have been conducted on wood-inhabiting fungi in Yunnan Province, China [ 20, 36, 51, 53, 54] . However, there are still many new taxa that have not been discovered yet, due to the unique geographical environment and vegetation types in Yunnan Province. In this study, the taxon sampling of wood-inhabiting fungi is expanded to investigate taxonomy and phylogeny of new samples within Russulales collected from Yunnan Province. Based on a combination of morphological and molecular evidence of internal transcribed spacer (ITS) region, the large subunit nuclear ribosomal RNA gene (nLSU), the small subunit of mitochondrial rRNA gene (mtSSU), the second largest subunit of RNA polymerase II ( rpb2), and the translation elongation factor 1- α gene ( tef1- α) markers, two new families, a new genus, and 26 new species are proposed. The data available to date were used to construct a time-framed phylogenomic tree of Russulales. Based on the recent taxonomic revisions by He et al. [ 1] and Liu et al. [ 42] , and the updated classification of fungi (Outline of Fungi 2024) by Hyde et al. [ 7] , the currently recognized suborders, families, and number of genera within order Russulales are systematically presented in Table 1.
Table 1. Number of genera within families of Russulales.
Order Families Number of genera Russulales Albatrellaceae 8 Aleurocystidiellaceae 1 Auriscalpiaceae 6 Bondarzewiaceae 8 Echinodontiaceae 5 Gloeocystidiellaceae 1 Gloeodontiaceae 1 Hericiaceae 6 Peniophoraceae 17 Russulaceae 7 Stereaceae 23 Terrestriporiaceae 1 Wrightoporiaceae 1 Incertae sedis 13
-
Fresh basidiomata of the fungi growing on angiosperm branches, and on the ground were collected from Dali, Dehong, Diqing, Honghe, Lincang, Lijiang, Puer, Qujing, Wenshan, Tengchong, Xishuangbanna, and Zhaotong in Yunnan Province, China. Voucher specimens were dried in an electric food dehydrator at 40 °C, and then deposited in the herbarium of the Southwest Forestry University (SWFC), Kunming, Yunnan Province, China. The samples were photographed in situ, and fresh macroscopic details were recorded. Photographs were recorded by a Jianeng 80D camera (Tokyo, Japan). All photos were stacked and merged using Helicon Focus Pro 7.7.5 software.
Morphological studies
-
Macromorphological descriptions and color terminology are based on field notes and photos captured in the field or laboratory, and follow those of a previous study [ 36] . Micromorphological characters were obtained from the dried specimens observed using a light microscope following a previous study [ 36] . The following abbreviations are used: KOH = 5% potassium hydroxide water solution, CB = cotton blue, CB– = acyanophilous, CB+ = cyanophilous, IKI = Melzer’s reagent, IKI+ = amyloid, IKI– = both inamyloid and indextrinoid, L = mean spore length (arithmetic average for all spores), W = mean spore width (arithmetic average for all spores), Q = variation in the L/W ratios between the specimens studied, and n = a/b (number of spores [a] measured from given number [b] of specimens) [ 55] .
Molecular procedures and phylogenetic analyses
-
The EZNA HP Fungal DNA Kit (Omega Biotechnologies Co., Kunming, China) was used with some modifications to extract DNA from the dried specimens. The DNA samples were kept at –20 °C. The thermal cycling conditions for each locus are provided in Table 2. The amplified fragments were internal transcribed spacer ITS (ITS5 and ITS4), the large subunit nuclear ribosomal RNA gene nLSU (LR0R and LR7), the small subunit of mitochondrial rRNA gene mtSSU (MS1 and MS2), the translation elongation factor 1- α gene tef1- α ( ef1-983 F and ef1-2218R), RNA polymerase II largest subunit rpb1 ( rpb1-Af and rpb1-Cf), and the second large subunit of RNA polymerase II rpb2 ( brpb2-6F and brpb2-7.1R) [ 56− 60] . The components of a 30 μL volume PCR mixture were 12.5 μL of double distilled water, 15 μL of PCR Master Mix (Sangon Biotech Shanghai Co., Shanghai, China), 1 μL of each primer and 1 μL of template DNA. The amplification followed the protocol of Dong et al. [ 52] . Amplified PCR products were examined through 1.5% agarose gel electrophoresis stained with GoldenView, and sent to Qingke Co., China, for sequencing. The PCR products were purified and sequenced at Kunming Tsingke Biological Technology Limited Company, Kunming, China. The sequences were reviewed and manually modified with Chromas v.1.0.1.1 to remove low-quality base calls from both ends. All newly generated sequences were subsequently deposited in GenBank ( Table 2).
Table 2. Gene regions and respective primers used in the study.
Gene
regionPrimer
pairsSequence (5′−3′) Annealing temperature (°C) Ref. ITS ITS5 GGAAGTAAAAGTCGTAACAAGG 55 [ 56] ITS4 TCCTCCGCTTATTGATATGC nLSU LR0R ACCCGCTGAACTTAAGC 48 [ 61] LR7 TACTACCACCAAGATCT mtSSU MS1 CAGCAGTCAAGAATATTAGTCAATG 52 [ 56] MS2 GCGGATTATCGAATTAAATAAC tef1- α ef1-983F GCYCCYGGHCAYCGTGAYTTYAT 60 [ 62] ef1-2218R ATGACACCRACRGCRACRGTYTG rpb1 rpb1-Af GARTGYCCDGGDCAYTTYGG 52 [ 59, 60] rpb1-Cf CCNGCDATNTCRTTRTCCATRTA rpb2 brpb2-6F TGGGGYATGGTNTGYCCYGC 52 [ 58] brpb2-7.1R CCCATRGCYTGYTTMCCCATDGC The PCR protocol for ITS was as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles at 94 °C for 40 s, and 58 °C for 40 s. The PCR procedure for nLSU was as follows: initial denaturation at 94 °C for 1 min, followed by 35 cycles at 94 °C for 30 s, 48 °C for 1 min and, 72 °C for 1.5 min, and a final extension of 72 °C for 10 min. The PCR procedure for mtSSU was as follows: initial denaturation at 94 °C for 2 min, followed by 35 cycles at 94 °C for 45 s, 52 °C for 45 s and 72 °C for 1 min, and a final extension of 72 °C for 10 min. The PCR procedure for tef1- α was as follows: initial denaturation at 94 °C for 2.5 min, denaturation at 94 °C for 45 s, annealing at 60 °C for 50 s (minus 1 °C per cycle), extension at 72 °C for 2 min, repeat for 6 cycles starting at step 2, denaturation at 94 °C for 30 s, annealing at 55 °C for 50 s, extension at 72 °C for 1.5 min, repeat for 34 cycles starting at step 6, leave at 72 °C for 5 min. The PCR procedure for rpb1 was 94 °C for 2 min, followed by 10 cycles at 94 °C for 40 s, 60 °C for 40 s and 72 °C for 2 min, then followed by 37 cycles at 94 °C for 45 s, 55 °C for 1.5 min and 72 °C for 2 min, and a final extension of 72 °C for 10 min. The PCR procedure for rpb2 was 95 °C for 2.5 min, followed by 40 cycles at 95 °C for 30 s, 52 °C for 1 min, and 72 °C for 1 min, then followed by 40 cycles at 72 °C for 1.5 min, and final extension of 72 °C for 5 min [ 20, 53] . The PCR products were purified and sequenced at Kunming Tsingke Biological Technology Limited Company, Kunming, China. The newly generated fungal sequences in this study were deposited in GenBank.
Phylogenetic analyses
-
Phylogenetic analyses followed the methods described in Dissanayake et al. [ 63] . Newly generated sequence data were initially subjected to a BLAST search in NCBI to obtain the most probable closely related taxa in the GenBank (
http://blast.ncbi.nlm.nih.gov ). Sequence data were retrieved from GenBank based on recent publications (www.ncbi.nlm.nih.gov/nuccore ). The sequences were aligned using MAFFT version 7 [ 64] with the G-INS-I strategy. The alignment was adjusted manually using AliView version 1.27 [ 65] . The dataset was initially aligned and later, ITS, nLSU, mtSSU, rpb2, and tef1- α sequences were combined using Mesquite version 3.51. FASTA data file formats were converted to NEXUS formats using the online tool available on the ALTER website (http://sing.ei.uvigo.es/ALTER/ ) [ 66] . Phylogenetic trees were constructed based on randomized accelerated maximum likelihood (ML) and Bayesian inference (BI) analyses.Maximum likelihood analysis was performed using the CIPRES Science Gateway (
www.phylo.org/portal2/login!input.action ) [ 67] based on the dataset using the RA × ML-HPC BlackBox tool, with setting RA × ML halt bootstrapping automatically and 0.25 for maximum hours and obtaining the best tree using ML search. Other parameters in ML analysis used default settings, and statistical support values were obtained using nonparametric bootstrapping with 1,000 replicates. Bayesian inference analysis was performed on the dataset using MrBayes v3.2.7a [ 68] . The best substitution model for the dataset was selected by ModelFinder v2.2.0 [ 69] using a Bayesian information criterion, and the model was used for Bayesian analysis. Four Markov chains were run from random starting trees. Trees were sampled every 1,000 th generation. The first 25% of sampled trees were discarded as burn-in, while the remaining trees were used to construct a 50% majority consensus tree and to calculate Bayesian posterior probabilities (BPPs). Phylogenetic trees were visualized and adjusted using FigTree v1.4.0 (http://tree.bio.ed.ac.uk/software/figtree ), and the exports were edited using Adobe Illustrator CS6 software (Adobe Systems, USA). Branches of the consensus tree that received bootstrap support for ML equal to or above 70%, and BI equal to or above 0.95, are indicated.Divergence time estimation
-
Three fossil calibrations, Archaeomarasmius leggetti Hibbett, D. Grimaldi and Donoghue, Quatsinoporites cranhamii S. Y. Sm., Currah and Stockey, and Paleopyrenomycites devonicus Taylor, Hass, Kerp, M. Krings and Hanlin, were used in the divergence time estimation. Archaeomarasmius leggetti was used as the representative of the minimum age of Agaricales at 90 Mya [ 70] ; Q. cranhamii was the representative of the minimum age of Hymenochaetaceae at 125 Mya [ 52] ; P. devonicus was used as the representative of the minimum age between Basidiomycota and Ascomycota at 400 Mya [ 71, 72] . Divergence time was estimated with the BEAST v2.6.5 software package with ITS, nLSU, rpb2 and tef1- α sequences representing main lineages in Basidiomycota ( Table 3). According to these time points, the offset age with a gamma distribution prior (scale = 20, shape = 1) for Agaricales was set as 90 Mya, and for Hymenochaetaceae as 125 Mya. After 20 million generations. The log file was analyzed in Tracer v1.6 to confirm that the estimated effective sample size (ESS) is ≥ 200 3. The first 10% of the sampled trees every 1000 th generation were removed as burn-in. The resulting log file was checked for chain convergence using Tracer 1.5.
Table 3. Taxa used in molecular clock analysis.
Order/family Species Sample no. GenBank accession no. Ref. ITS nLSU rpb2 tef1- α Agaricales Asterophora lycoperdoides CBS 170.86 AF357037 AF223190 DQ367431 DQ367424 [ 77] Gymnopilus picreus ZRL2015011 LT716066 KY418882 KY419027 KY419077 [ 77] Amylocorticiales Amylocorticium cebennense HHB-2808 GU187505 GU187561 GU187770 GU187675 [ 78] Anomoloma myceliosum MJL-4413 GU187500 GU187559 GU187766 GU187677 [ 78] Atheliales Athelia arachnoidea CBS 418.72 GU187504 GU187557 GU187769 GU187672 [ 78] Leptosporomyces raunkiaerii HHB-7628 GU187528 GU187588 GU187791 [ 78] Auriculariales Auricularia heimuer Xiaoheimao LT716074 KY418890 KY419035 KY419083 [ 78] Exidia sp . PBM2527 DQ241774 AY700191 DQ408144 [ 78] Boletales Coniophora arida FP104367 GU187510 GU187573 GU187775 GU187684 [ 78] Gomphidius roseus MB 95-038 DQ534570 DQ534669 GU187818 GU187702 [ 78] Dacrymycetales Calocera cornea AFTOL 438 AY789083 AY701526 AY536286 AY881019 [ 78] Dacryopinax spathularia AFTOL 454 AY854070 AY701525 AY857981 AY881020 [ 78] Geastrales Geastrum taylorii OSC59760 DQ218520 DQ219060 DQ219235 [ 79] Schenella pityophila OSC59743 DQ218519 DQ219057 DQ219232 [ 79] Gomphales Clavariadelphus truncatus OSC67280 AY574649 DQ219064 DQ219240 [ 78] Kavinia alboviridis 0102140 AY574692 DQ219073 DQ219250 [ 78] Hymenochaetales Fomitiporia mediterranea AFTOL688 AY854080 AY684157 AY803748 AY885149 [ 78] Phellinus hartigii Dai 11766 KT203287 KT203308 KJ651721 [ 78] Hysterangiales Aroramyces gelatinosporus H4010 DQ218524 DQ218941 DQ219118 [ 78] Chondrogaster pachysporus OSC49298 DQ218538 DQ218958 DQ219136 [ 78] Jaapiales Jaapia argillacea CBS252.74 GU187524 GU187581 GU187788 GU187711 [ 78] Polyporales Fomitopsis pinicola AFTOL 770 AY854083 AY684164 AY786056 AY885152 [ 78] Polyporus squamosus Cui 10595 KU189778 KU189809 KU189988 KU189925 [ 78] Russulales/Albatrellaceae Albatrellus ovinus PS11795 MW269673 MW269685 MW290304 MW290320 [ 26] Byssoporia terrestris Hjm18172 EU118608 EU118608 [ 45] Polyporoletus sublividus JA 030918 DQ389663 DQ389663 [ 80] –/Aleurocystidiellaceae Aleurocystidiellum bernicchiae MR12636 MT831037 MT831017 [ 46] Aleurocystidiellum bernicchiae SPG3217 MT831016 [ 46] Aleurocystidiellum subcruentatum He2886 KU559341 KU574847 KU992720 [ 81] Aleurocystidiellum subcruentatum HHB-17353-sp KU559360 KU574818 [ 81] Aleurocystidiellum tsugae He4025 KY706211 KY706223 [ 81] Aleurocystidiellum tsugae He4024 KY706210 KY706222 [ 81] –/Auriscalpiaceae Auriscalpium vulgare HKAS93484 MK211170 KY485984 KY495319 KY474614 Unpublished Dentipratulum bialoviesense GG1645 AF506389 AF506389 [ 26] Lentinellus cochleatus KGN960928 AF506417 AF506417 [ 26] –/Bondarzewiaceae Bondarzewia occidentalis HHB-14803 KM243329 KM243332 KX066163 KX066142 [ 46] Heterobasidion annosum Korhonen 06129/6 KJ583211 KJ583225 KF006499 KX252741 [ 46] Laurilia sulcata He20120916-7 KY172894 KY172909 [ 82] Lauriliella taxodii FP-105464-Sp KY172896 KY172912 [ 82] –/Echinodontiaceae Amylostereum chailletii NH8031 AF506406 AF506406 [ 26] Echinodontiellum japonicum Dai 7378 KY172887 KY172902 [ 82] Echinodontium tinctorium HHB-12866-Sp KY172888 KY172903 MH550371 [ 26] Larssoniporia tropicalis Ryvarden 45363 KJ513294 KJ807089 [ 44] Subulicystidiella murina CLZhao 30728 PV771055 PX418375 Present study Subulicystidiella murina CLZhao 35801* PV441140 PV441154 Present study –/Gloeocystidiellaceae Gloeocystidiellum clavuligerum He3376 KY860377 KY860434 Unpublished Gloeocystidiellum granulatum He4301 KY860391 KY860449 Unpublished Gloeocystidiellum membranaceum CLZhao 37038 PV940928 PX070092 PX432797 PX439082 Present study Gloeocystidiellum porosellum Hjm 8851 AY048878 AY048878 Unpublished Gloeocystidiellum porosum HHB-15589-Sp MK625627 MK625555 MN031003 Unpublished Gloeocystidiellum punctatum CLZhao 20755* PP356586 PP785346 Present study Gloeocystidiellum purpureum Wu9310-45 AF441338 AF441338 [ 26] –/Gloeodontiaceae Gloeodontia columbiensis NH 11118 AF506444 AF506444 [ 26] Gloeodontia pyramidata Ryvarden 15502 AF506446 AF506446 [ 26] Gloeodontia sinensis CLZhao 34748 PV147171 PV185857 PV400175 Present study Gloeodontia subasperispora GB/KHL8695 AF506404 AF506404 [ 26] Gloeodontia discolor KHL 10099 AF506445 AF506445 [ 26] Gloeodontia eriobotryae Dai 12080 JQ349116 JQ349103 [ 30] Gloeodontia yunnanensis CLZhao 10504 MN908252 MN908254 [ 83] –/Hericiaceae Dentipellis fragilis Dai14767 MH085943 MH085958 Unpublished Hericium americanum DAOM21467 AF506458 AF506458 [ 26] Laxitextum bicolor Dai14056 KY860393 KY860451 [ 82] –/Peniophoraceae Asterostroma muscicola He4397 MK625630 MK625563 MN030965 [ 82] Baltazaria galactina He4999 MK625618 MK625547 MN030977 [ 84] Dichostereum boidinii He4410 MH538315 MH538331 MH550361 [ 43] Gloiothele lamellosa CBS 404.83 AF506487 AF506487 [ 26] Lachnocladium schweinfurthianum KM49740 MH260033 MH260051 [ 85] Peniophora quercina CBS 408.50 MH856688 MH868205 [ 84] Scytinostroma portentosum EL 11-99 AF506470 AF506470 [ 26] Vararia trinidadensis CBS:650.84 MH873495 MH873495 [ 84] –/Russulaceae Lactarius torminosus CBS 197.72 MH860447 MH872175 [ 84] Multifurca ochricompacta JJ2010.08 (PC0723658) MH063879 MH063844 MH061176 [ 86] Russula delica FH12-272 KF432955 KR364224 KR364340 [ 61] –/Stereaceae Acanthobasidium delicatum CBS 233.86 MH861948 MH873638 [ 84] Aleurobotrys botryosus DAOM211598 AF506398 AF506398 [ 26] Aleurodiscus amorphus Ghobad-Nejhad2464 KU559342 KU574832 KU992717 [ 81] Confertotrama rugulosa LodgeSJ 110.1 AF506441 AF506441 [ 87] Gelatinostereum phlebioides He4492 MW533096 MW528942 Unpublished Gloeosoma vitellinum 646cc MT831039 MT831019 [ 46] Megalocystidium leucoxanthum HK9808 AF506420 AF506420 [ 26] Stereodiscus antarcticus MR11265 MT831048 MT831028 [ 46] Stereum hirsutum He3504 MK625629 MK625557 MN031010 Unpublished Xylobolus frustulatus He2231 MH121216 KU574825 KU992704 [ 81] –/Terrestriporiaceae Terrestriporia alba Dai 18548 MT068564 MT068560 MW290307 MW290324 [ 21] Terrestriporia alba Dai 18556 MT068565 MT068561 MW290308 MW290325 [ 21] –/Wrightoporiaceae Wrightoporia avellanea LR 41710 AF506488 AF506488 [ 26] Wrightoporiaceae Wrightoporia subavellanea Dai 11484 KJ513295 KJ807085 [ 26] Sebacinales Sebacina sp. AFTOL 1517 DQ911617 DQ521412 [ 78] Tremellodendron pallidum AFTOL 699 DQ411526 AY745701 DQ408132 DQ029196 [ 78] Sordariales Neurospora crassa OR74A HQ271348 AF286411 AF107789 XM959775 [ 78] Thelephorales Boletopsis leucomelaena PBM2678 DQ484064 DQ154112 GU187820 GU187763 [ 78] Thelephora ganbajun ZRL20151295 LT716082 KY418908 KY419043 KY419093 [ 78] Genealogical concordance phylogenetic species recognition (GCPSR) analysis
-
The pairwise homoplasy index (PHI) test was conducted in certain cases of species delineation when necessary. We used the genealogical concordance phylogenetic species recognition analysis (GCPSR) to check for significant recombination events [ 73] . The data were analyzed using the PHI test in SplitsTree 4 to determine the recombination level with closely related species [ 74− 76] . ITS and LSU datasets with closely related species were used for the analyses. The pairwise homoplasy index lower than 0.05 (Φw ≤ 0.05) indicates significant recombination in the dataset. The relationships between closely related taxa were visualized by constructing split graphs from the concatenated datasets using the LogDet transformation and split decomposition options.
Sample collection and herbarium specimen preparation
-
The aligned dataset encompassed 104 specimens representing 99 taxa. Cerioporus squamosus (Huds.) Quél. and Trametes suaveolens (L.) Fr. retrieved from GenBank were used as outgroup taxa ( Fig. 1) in previous analysis by Liu et al. [ 88] . Trees and parameters were sampled every 1,000 generations. ModelFinder v2.2.0 [ 69] was used to select the best-fit model based on the BIC criterion. The best model for the combined ITS, nLSU, mtSSU, rpb2, and tef1- α dataset was estimated as GTR + I + G, and it was applied in the Bayesian analysis. Maximum likelihood (ML) and Bayesian inference (BI) analyses yielded a similar topology, with an average standard deviation of split frequencies of 0.009610 (BI), and an effective sample size (ESS) average ESS (avg. ESS) = 754. The phylogram, based on the combined ITS + nLSU + mtSSU + rpb2 + tef1- α sequence analysis ( Fig. 1), showed that Russulales formed 13 distinct lineages, Albatrellaceae, Aleurocystidiellumaceae, Auriscalpiaceae, Bondarzewiaceae, Echinodontiaceae, Gloeocystidiellaceae, Gloeodontiaceae, Hericiaceae, Peniophoraceae, Russulaceae, Stereaceae, Terrestriporiaceae, and Wrightoporiaceae.

Figure 1.
Maximum likelihood strict consensus tree illustrating the phylogeny of the species of Russulales based on ITS + nLSU + mtSSU + rpb2 + tef1- α sequences. Branches are labeled with maximum likelihood bootstrap values higher than 70%, and Bayesian posterior probabilities more than 0.95, respectively.
The divergence time of the order Russulales based on combined ITS + nLSU + rpb2 + tef1- α sequences data ( Fig. 2)
-
The ITS, nLSU, rpb2, and tef1- α dataset included 92 collections, of which 59 belonged to Russulales. This dataset resulted in a concatenated alignment of 5,419 characters with GTR + I + G as the best-fit evolutionary model. Chain convergence was indicated by the ESS 499. The result ( Fig. 2) showed that Russulales occurred in a mean crown age of 222.49 Mya (179.97–262.75 Mya, 95% HPD). The initial diversification between Hericiaceae and Aleurocystidiellaceae at 140.94 Mya (87.46–196.71 Mya, 95% HPD). The family Gloeodontiaceae occurred in a mean stem age of 178.7 Mya (139.36–218.13 Mya, 95% HPD). Subsequently, the new genus Subulicystidiella is grouped into family Echinodontiaceae and occurred in a mean stem age of 114.84 Mya (71.1–161.48 Mya, 95% HPD). The estimated divergence times for other nodes are summarized in Table 4.
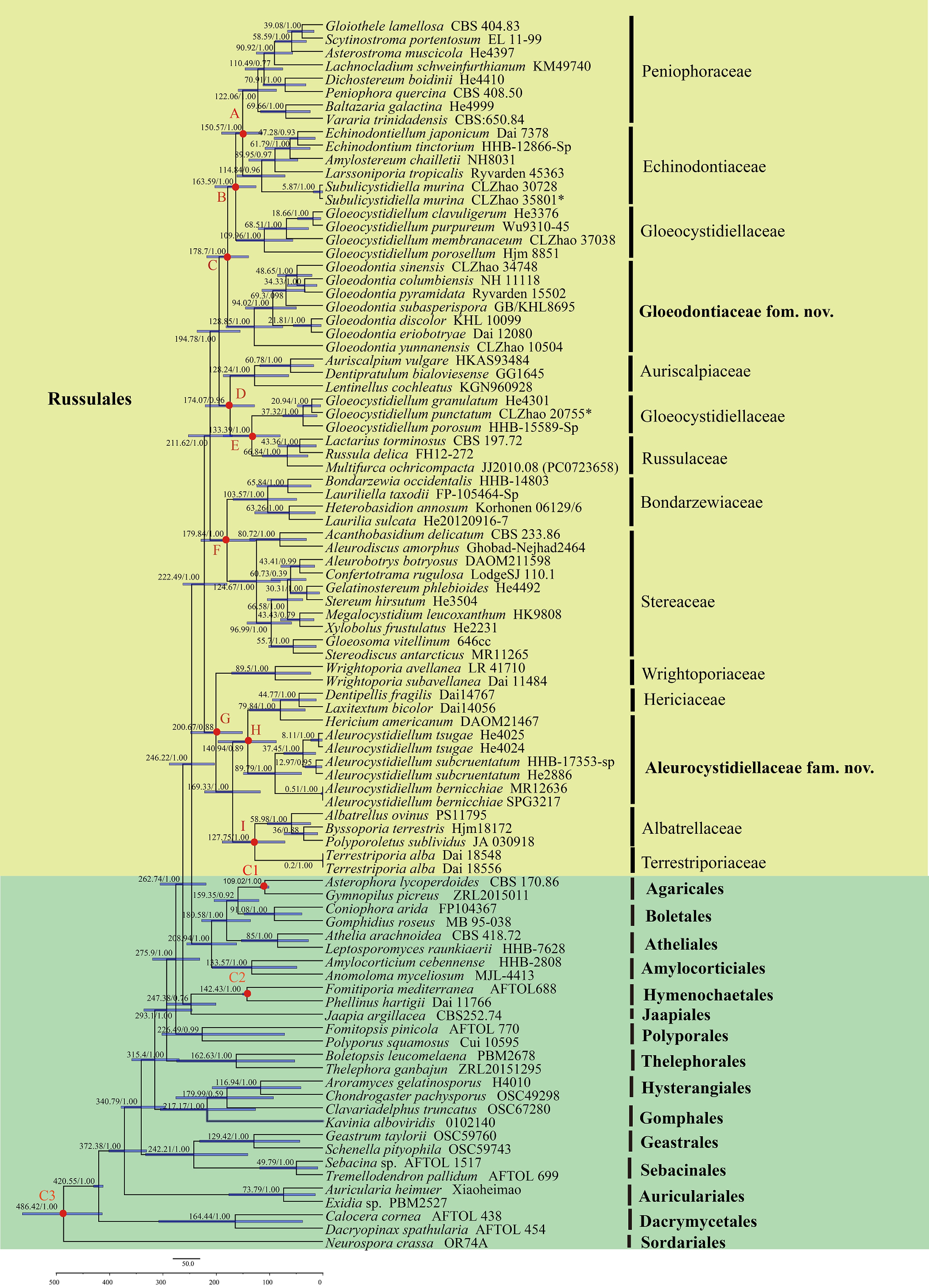
Figure 2.
Divergence time estimation of families within Russulales from molecular clock analysis sampling tree based on the combined sequence dataset of ITS, nLSU, rpb2, and tef1- α. Posterior probabilities not less than 0.80, and the mean ages of each node are annotated. The 90% highest posterior densities of divergence time estimation are marked by horizontal bars.
Table 4. Estimated divergence time of each node.
Node Means of stem age (Mya) 95% HPD (Mya) posterior probabilities A: Peniophoraceae/Echinodontiaceae 150.57 113.56–190.2 B: Gloeocystidiellaceae 163.59 125.46–202.89 C: Gloeodontiaceae 178.7 139.36–218.13 D: Auriscalpiaceae 174.07 128.22–221.01 E: Gloeocystidiellaceae/Russulaceae 133.39 79.96–186.84 F: Bondarzewiaceae/Stereaceae 179.84 126.6–228.84 G: Wrightoporiaceae 200.67 150.99–248.34 H: Hericiaceae/Aleurocystidiellaceae 140.94 87.46–196.71 I: Albatrellaceae/Terrestriporiaceae 127.75 71.29–188.91 C1: Hymenochaetales 142.43 135.31–150.67 C2: Agaricales 109.02 101.26–117.85 C3: Ascomycota/Basidiomycota 486.42 413.94–727.32 Family Gloeocystidiellaceae phylogeny based on combined ITS + nLSU sequences data ( Fig. 3)
-
The aligned dataset encompassed 43 specimens representing 20 taxa, including the two new species, Gloeocystidiellum membranaceum, G. punctatum, and Laurilia sulcata (Burt) Pouzar was retrieved from GenBank as an outgroup in using the concatenated ITS + nLSU sequences dataset analysis ( Fig. 3) following the previous study analysis [ 89] . The best fit model was estimated as TIM3ef + I + G for the ITS + nLSU dataset, and it was applied in the Bayesian analysis. Four Markov chains were run twice from a random starting tree, for 0.35 million generations of the datasets ( Fig. 3) with trees and parameters sampled every 1,000 generations. Both Bayesian analysis and ML analysis resulted in a similar topology to MP analysis with an average standard deviation of split frequencies = 0.009945 (BI), and the effective sample size (ESS) across the two runs is double of the average ESS (avg ESS) = 1224. The phylogram depicts an overall topology of the genus Gloeocystidiellum Donk ( Fig. 3) and reveals that G. membranaceum Y.L. Deng & C.L. Zhao, and G. punctatum Y.L. Deng & C.L. Zhao are grouped into this genus and G. membranaceum is retrieved as a sister to G. porosellum Hjortstam, and closely clustered with G. bisporum Boidin, Lanq. & Gilles. In addition, G. punctatum formed a separate clade.
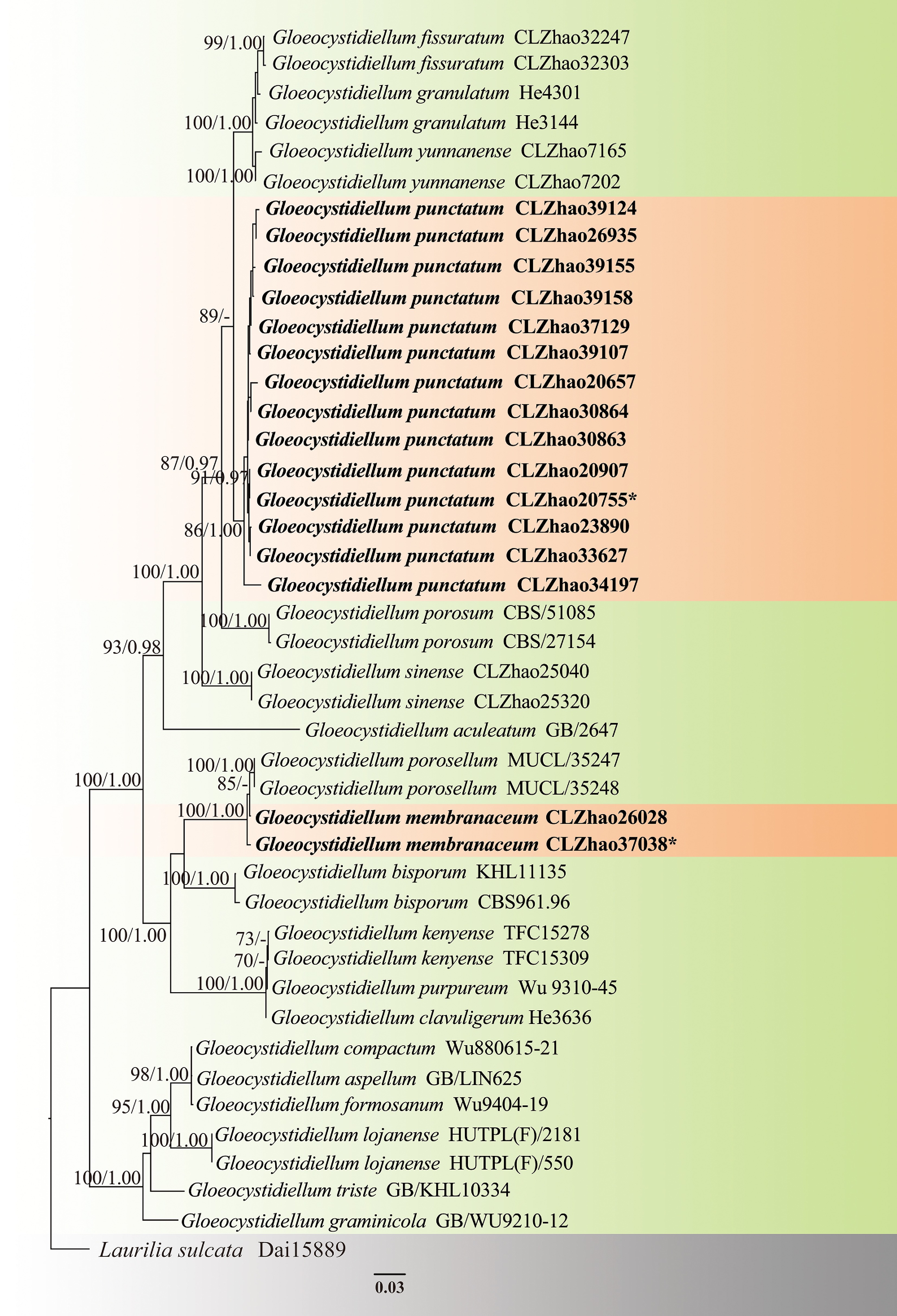
Figure 3.
Maximum likelihood strict consensus tree illustrating the phylogeny of two new species of Gloeocystidiellum based on ITS and nLSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 70%, and Bayesian posterior probabilities more than 0.95, respectively. The new species are in bold,, and type specimens are indicated with an asterisk (*).
Family Hericiaceae phylogeny based on combined ITS + nLSU sequences data ( Fig. 4)
-
The aligned dataset encompassed 75 specimens representing 53 taxa. Albatrellus ovinus (Schaeff.) Kotl. & Pouzar and A. subrubescens (Murrill) Pouzar were retrieved from GenBank as the outgroup taxa in using the concatenated ITS + nLSU sequences dataset analysis ( Fig. 4), following a previous analysis [ 11] . The estimated best fit model was GTR + I + G for the ITS + nLSU dataset, and it was applied in the Bayesian analysis. Four Markov chains were run twice from a random starting tree, for 0.25 million generations of the datasets ( Fig. 3) with trees and parameters sampled every 1,000 generations. Both Bayesian analysis and ML analysis resulted in a similar topology to MP analysis, with an average standard deviation of split frequencies = 0.009780 (BI), and the effective sample size (ESS) across the two runs is double of the average ESS (avg ESS) = 268.
The phylogram depicts an overall topology of family Hericiaceae ( Fig. 4), and shows that Dentipellis yingjiangensis and Laxitextum cremeum are grouped into genera Dentipellis Donk and Laxitextum Lentz, respectively. Dentipellis yingjiangensis Y.L. Deng & C.L. Zhao is retrieved as a sister to D. rhizomorpha Yuan Yuan & Y.C. Dai, and closely related to D. fragilis (Pers.) Donk, and D. dissita (Berk. & Cooke) Maas Geest. In addition, Laxitextum cremeum Y.L. Deng & C.L. Zhao is closely related to L. subrubrum R. Saha, A.K. Dutta & K. Acharya, L. bicolor (Pers.) Lentz, and L. incrustatum Hjortstam & Ryvarden. Furthermore, Subulicystidiella Y.L. Deng & C.L. Zhao clustered together within family Echinodontiaceae, and closely related to Larssoniporia Y.C. Dai, Jia J. Chen & B.K. Cui, and Amylostereum Boidin.

Figure 4.
Maximum likelihood strict consensus tree illustrating the phylogeny of species of family Hericiaceae, and related families based on ITS and nLSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 70%, and Bayesian posterior probabilities more than 0.95, respectively. The new species are in bold, and type specimens are indicated with an asterisk (*).
Family Peniophoraceae phylogeny based on combined ITS and nLSU sequence data ( Fig. 5)
-
The aligned dataset encompassed 149 specimens representing 89 taxa. Stereum ostrea (Blume & T. Nees) Fr., and S. hirsutum (Willd.) Pers. were retrieved from GenBank as the outgroup taxa in the concatenated ITS + nLSU sequences dataset analysis ( Fig. 5) following the method of Deng et al. [ 33] . The estimated best fit model was GTR + I + G for the ITS + nLSU dataset, and it was applied in the Bayesian analysis. Four Markov chains were run twice from a random starting tree, for 7.66 million generations of the datasets ( Fig. 5) with trees and parameters sampled every 1,000 generations. Both Bayesian analysis and ML analysis resulted in a similar topology to MP analysis, with an average standard deviation of split frequencies = 0.010000, and the effective sample size (ESS) across the two runs is double of the average ESS (avg ESS) = 2,852.
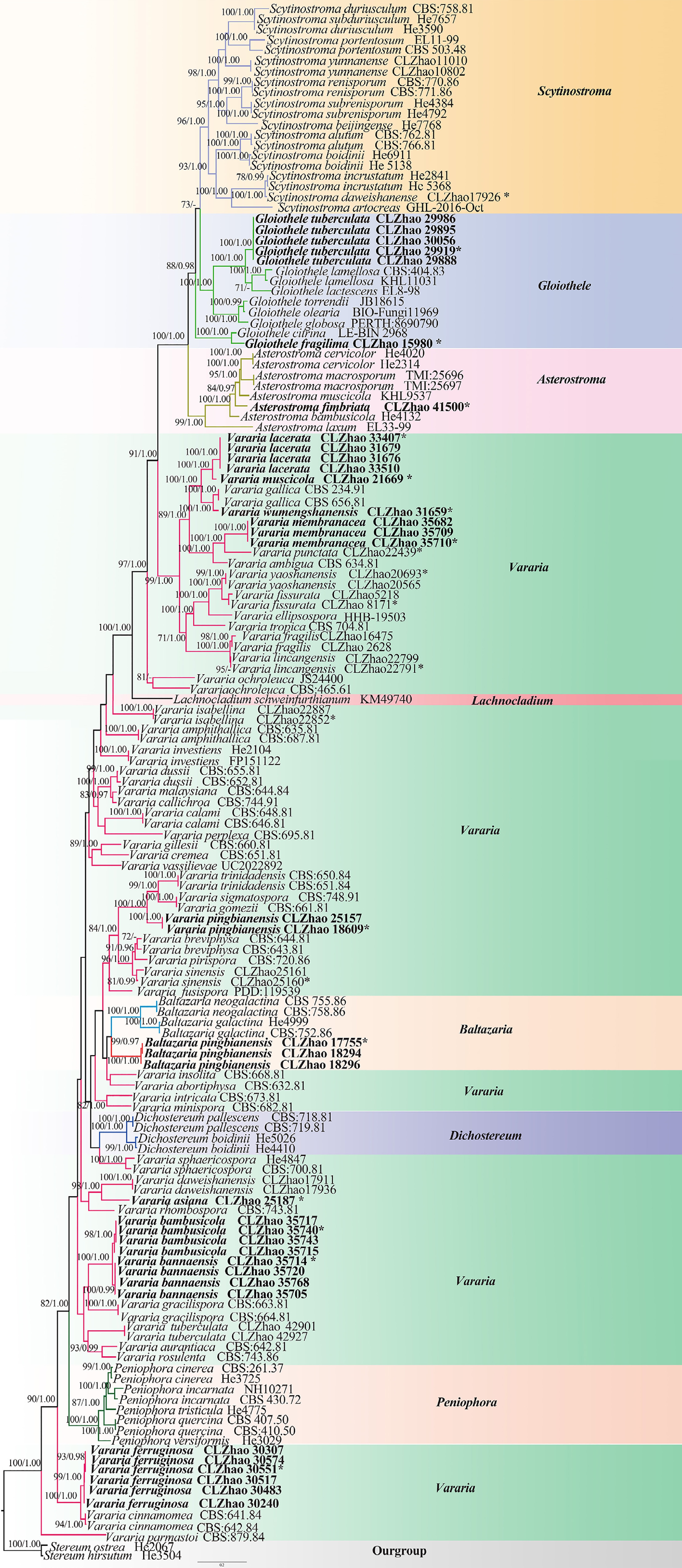
Figure 5.
Maximum likelihood strict consensus tree illustrating the phylogeny of species of family Peniophoraceae based on ITS and nLSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 70%, and Bayesian posterior probabilities more than 0.95, respectively. The new species are in bold, and type specimens are indicated with an asterisk (*).
The phylogram depicts an overall topology of family Peniophoraceae ( Fig. 5), and supports the introduction of 13 new species. Asterostroma fimbriatum Y.L. Deng & C.L. Zhao is grouped into Asterostroma, and retrieved as a sister to A. muscicola (Berk. & M.A. Curtis) Massee. Baltazaria pingbianensis Y.L. Deng & C.L. Zhao is grouped into Baltazaria, and closely related to B. galactina (Fr.) Leal-Dutra, Dentinger & G.W. Griff, and B. neogalactina (Boidin & Lanq.) Leal-Dutra, Dentinger & G.W. Griff. Two new species Gloiothele fragilima Y.L. Deng & C.L. Zhao, and G. tuberculata Y.L. Deng & C.L. Zhao are grouped into Gloiothele. They are sister to G. citrina (Pers.) Ginns & G.W. Freeman. and G. lamellosa (Henn.) Bres. and G. lactescens (Berk.) Hjortstam., respectively. Nine new Vararia species are grouped into genus Vararia, in which, V. asiana Y.L. Deng & C.L. Zhao is sister to V. daweishanensis L. Zou & C.L. Zhao, and closely related to V. rhombospora Boidin & Lanq; V. bambusicola Y.L. Deng & C.L. Zhao is sister to V. bannaensis Y.L. Deng & C.L. Zhao; V. ferruginosa Y.L. Deng & C.L. Zhao is related to V. cinnamomea Boidin, Lanq. & Gilles as a sister; V. lacerata Y.L. Deng & C.L. Zhao is sister to V. muscicola Y.L. Deng & C.L. Zhao; V. membranacea Y.L. Deng & C.L. Zhao is sister to V. punctata Y.L. Deng & C.L. Zhao; V. muscicola is sister to V. lacerata; V. pingbianensis is closely related to V. gomezii Boidin & Lanq., V. sigmatospora Boidin, Gilles & Lanq., and V. trinidadensis A.L. Welden. V. wumengshanensis Y.L. Deng & C.L. Zhao is grouped as sister to V. gallica (Bourdot & Galzin) Boidin .
Family Stereaceae phylogeny based on combined ITS and nLSU sequences data ( Fig. 6)
-
The aligned dataset encompassed 170 specimens representing 100 taxa. Laurilia sulcata was retrieved from GenBank as the outgroup taxon in the concatenated ITS+nLSU sequences dataset analysis ( Fig. 6) following a previous analysis [ 87] . The estimated best fit model was GTR + I + G for the ITS + nLSU dataset, and it was applied in the Bayesian analysis. Four Markov chains were run twice from a random starting tree, for 5.115 million generations of the datasets ( Fig. 6) with trees and parameters sampled every 1,000 generations. Both Bayesian analysis and ML analysis resulted in a similar topology to MP analysis with an average standard deviation of split frequencies = 0.009977, and the effective sample size (ESS) across the two runs is double of the average ESS (avg ESS) = 2255.
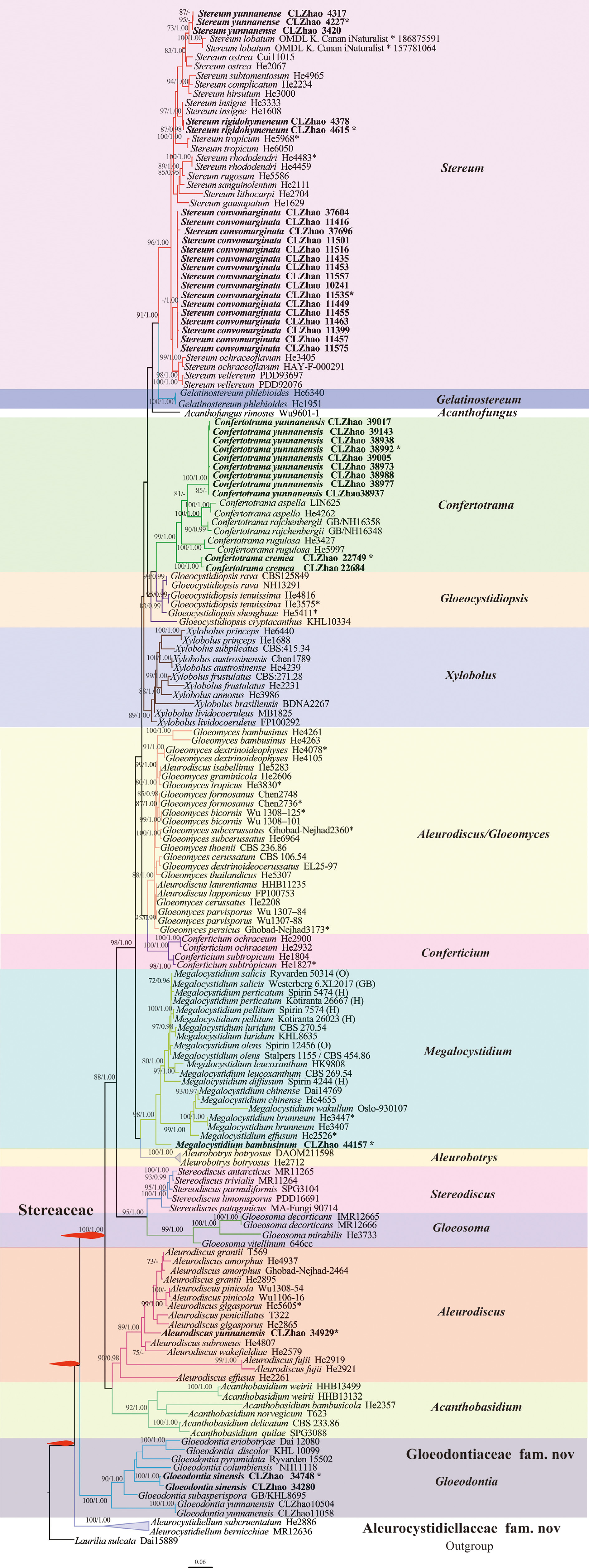
Figure 6.
Maximum likelihood strict consensus tree illustrating phylogeny of species of family Stereaceae and related families based on ITS and nLSU sequences. Branches are labeled with maximum likelihood bootstrap values higher than 70%, and Bayesian posterior probabilities more than 0.95, respectively. The new species are in bold, and type specimens are indicated with an asterisk (*).
The phylogram depicts an overall topology of family Stereaceae ( Fig. 6), and eight new species are introduced. Aleurodiscus yunnanensis Y.L. Deng & C.L. Zhao is grouped within the genus Aleurodiscus and is closely related to A. wakefieldiae Boidin & Beller, and A. subroseus S.H. He & Y.C. Dai. Two new species, Confertotrama cremea Y.L. Deng & C.L. Zhao and C. yunnanensis Y.L. Deng & C.L. Zhao grouped with C. aspella (Hjortstam) Nakasone & S.H. He, C. rajchenbergii (Gorjón & Hallenb.) Nakasone & S.H. He, and C. rugulosa (Berk. & M.A. Curtis) Nakasone & S.H. He forming a clade. Gloeodontia sinensis Y.L. Deng & C.L. Zhao is grouped into Gloeodontia and closely related to G. columbiensis Burt ex Burds. & Lombard, and G. subasperispora (Litsch.) E. Larss. & K.H. Larss. Megalocystidium bambusinum Y.L. Deng & C.L. Zhao formed a separate lineage, closely related to M. effusum S.H. He and M. brunneum S.H. He. The three new species, Stereum convolutomarginatum Y.L. Deng & C.L. Zhao, S. rigidohymeneum Y.L. Deng & C.L. Zhao, and S. yunnanense Y.L. Deng & C.L. Zhao are grouped within the genus Stereum and closely related to S. ochraceoflavum (Schwein.) Sacc. and S. vellereum Berk., S. insigne Bres., and S. lobatum (Kunze ex Fr.) Fr., respectively.
Taxonomy:
-
Russulales Kreisel ex P.M. Kirk, P.F. Cannon & J.C. David
Family Albatrellaceae Nuss 1980
Index Fungorum number: IF80437
Type genus – Albatrellus Gray
Notes – Albatrellaceae is characterized by annual, resupinate, pileate-stipitate or gasteroid basidiomata, poroid or locular (gasteroid fungi) hymenophore, monomitic hyphal system, with or without clamp connections, inamyloid or amyloid hyphae, smooth or appearing slightly rough, with a double wall separated by interwall pillar or aleveolate, hyaline, inamyloid or amyloid basidiospores. Currently, eight genera are accepted in family Albatrellaceae [ 1, 7, 17] .
Note 1 Albatrellus Gray
Index Fungorum number: IF17035
Type species – Albatrellus albidus Gray 1821
Notes – Albatrellus is a mushroom genus belonging to Russulales, and holds a unique taxonomic status due to its poroid basidiomata [ 16, 90] . The important identifying features of Albatrellus are a poroid hymenophore of mostly white or cream colour, usually inflated hyphae, and smooth and mostly amyloid basidiospores. Initially, the taxonomy of Albatrellus was based on morphological studies, and the genus was treated as a polypore [ 91− 93] . Previous phylogenetic studies of Russulales revealed some Albatrellus species clustered in the russuloid clade [ 26, 94, 95] . The taxonomy of European and North American Albatrellus has been carried out by various mycologists [ 15, 91− 93, 96] . Recently, Zhou et al. [ 17] conducted multi-locus phylogenetic analyses based on seven gene loci (ITS, nLSU, tef1- α, rpb1, rpb2, mtSSU, and nucSSU), and showed that Albatrellus formed a polyphyletic group and belonged to family Albatrellaceae. Species within Albatrellus, specifically those in Albatrellus sensu stricto, are significant ectomycorrhizal fungi [ 14] . Some Albatrellus species also have edible and medicinal functions (antioxidative and antitumor activities), such as A. ovinus (Schaeff.) Kotl. & Pouzar which is a common edible mushroom in Europe and North America [ 19, 97− 99] . Based on Index Fungorum (2025), Albatrellus consists of 54 registered names of which 32 species are accepted worldwide.
Note 2 Byssoporia M.J. Larsen & Zak 1978
Index Fungorum number: IF17210
Type species – Byssoporia terrestris (DC.) M.J. Larsen & Zak 1978
Notes – Byssoporia was typified by B. terrestris (Pers.) M.J. Larsen & Zak, a genus proposed for Poria terrestris Pers. and its varieties [ 100] . It is characterized by effused basidiomata, basidia with 4 sterigmata, and ellipsoid to subglobose basidiospores [ 100] . Based on the MycoBank database (2025), and Index Fungorum (2025), Byssoporia has seven taxa, but only B. terrestris is accepted [ 100] . The species diversity of this genus needs to be further explored. Based on the ITS and nLSU dataset, the phylogeny of Russulales showed that Byssoporia grouped together with Albatrellus and Polyporoletus into family Albatrellaceae [ 10] .
Note 3 Polyporoletus Snell
Index Fungorum number: IF18333
Type species – Polyporoletus sublividus Snell
Notes – Polyporoletus inferred from the specific spores and typified by P. sublividus Snell. The genus is characterized by annual fascicular basidiomata, hymenophore with gray to bluish-gray or olive gray pores, monomitic hyphal system, cylindrical basidia and ellipsoid to subglobose basidiospores [ 96] . Based on the MycoBank database (2025), and Index Fungorum (2025), Polyporoletus has four species, P. bulbosus Audet, P. neotropicus M. Mata & Ryvarden, P. sublividus, and P. sylvestris (Overh. ex Pouzar) Audet. Based on morphological characteristics and the phylogenetic analysis of ITS1, 5.8S, ITS2 sequence data, Polyporoletus was closely related to Albatrellopsis Teixeira in family Albatrellaceae [ 96] .
Note 4 Family Aleurocystidiellaceae Y.L. Deng & C.L. Zhao, fam. nov.
Index Fungorum number: IF861354
Type genus – Aleurocystidiellum P.A. Lemke
Description – Basidiomata annual to perennial, cupulate to substereoid, margin determinate, subcoriaceous, hymenophore smooth, abhymenial sterile surface glabrous. Hyphal system monomitic or dimitic, generative hyphae with clamps. Basidia subclavate with 4-sterigmata and a basal clamp. Basidiospores ovoid to broadly ellipsoid, apiculate, thick-walled, minutely verruculose amyloid in Melzer's [ 46, 101] .
Notes – Lemke [ 101] proposed the genus Aleurocystidiellum, and it could not be assigned to any recognized family in Russulales. Rajchenberg et al. [ 46] proposed two new species combinations, A. bernicchiae (Gorjón, Gresl. & Rajchenb.) Rajchenb. & Pildain, and A. hallenbergii (Gorjón, Gresl. & Rajchenb.) Rajchenb. & Pildain based on morphology and phylogenetic analyses. However, a phylogenetic tree analysis indicated that Aleurocystidiellum formed a separate clade, and did not affiliate in any recognized family of Russulales. In this study, based on combined ITS + nLSU + rpb2 + tef1- α sequence dataset of Russulales in the phylogenetic analyses as well as the divergence time ( Figs 1, 2), Aleurocystidiellum formed a distinct lineage with strong support within Russulales. The divergence time of the Aleurocystidiellum clade is 140.94 Mya, with a 95% highest posterior density (HPD) of 87.46–196.71 Mya, 95% HPD. Therefore, the new family Aleurocystidiellaceae is introduced for this genus.
Note 5 Aleurocystidiellum P.A. Lemke 1964
Index Fungorum number: IF17039
Type species – Aleurocystidiellum subcruentatum (Berk. & M.A. Curtis) P.A. Lemke
Notes – Aleurocystidiellum was established by Lemke [ 101] to include dimitic species with discoid basidiomata and large, minutely verrucose, amyloid spores with A. subcruentatum (Berk. & M.A. Curtis) P.A. Lemke as the type species. Aleurocystidiellum subcruentatum was previously placed in Aleurodiscus based on its discoid basidiomata and the amyloid basidiospores [ 101] . Previously, Aleurodiscus disciformis (DC.) Pat. was transferred to Aleurocystidiellum [ 26] . Morphologically, A. disciforme (DC.) Tellería has moniliform gloeocystidia rather than skeletocystidia as in A. subcruentatum (Berk. & M.A. Curtis) P.A. Lemke, otherwise these two species are highly similar. The two species formed a distinct clade distant from Aleurodiscus s.s. in the phylogenetic trees in previous studies [ 101− 103] . Aleurodiscus tsugae Yasuda ex Lloyd was originally described from Japan on bark of Tsuga, and then found on Pinus in Japan, the Russian Far East and northeastern China [ 102, 104] . Careful morphological and molecular studies of the Chinese specimens of Aleurodiscus tsugae indicated that it belongs to the genus Aleurocystidiellum, and thus a new combination was proposed [ 104] .
Family Auriscalpiaceae Maas Geest. 1963
Index Fungorum number: IF80506
Type genus – Auriscalpium Gray
Notes – Auriscalpiaceae is characterized by annual, resupinate, effused-reflexed, pileate-sessile, pileate-stipitate to clavarioid basidiomata, hydnoid, poroid, labyrinthine to daedaleoid, meruloid and lamellate hymenophore. Monomitic to dimitic hyphal system, generative hyphae with clamp-connections, inamyloid or amyloid, skeletal hyphae when present dextrinoid ( Amylonotus), gloeoplerous hyphae and gloeocystidia present or absent, with asperulate, spinulose, verrucose, hyaline to pigmented, amyloid basidiospores. The species of this family are wood decaying or ectomycorrizal fungi [ 1] .
Note 6 Artomyces Jülich
Index Fungorum number: IF17104
Type species – Artomyces pyxidatus (Pers.) Jülich
Notes – Jülich [ 105] introduced Artomyces, and typified it with A. pyxidatus (Pers.) Jülich, proposing Artomyces as a distinct genus separate from Clavicorona Doty. Lickey et al. [ 106] conducted a comprehensive phylogenetic and taxonomic study for Artomyces, accepting 15 species within the genus based on a combination of morphology, nuclear ribosomal internal transcribed spacer DNA (nrITS DNA) sequences, and mating studies. In addition, they described seven new species. Kneal & Smith [ 107] described a new species A. nothofagi R.J. Kneal & M.E. Sm., based on morphology and phylogeny. Subsequently, Dong et al. [ 52] proposed two new species, A. niveus J.H. Dong & C.L. Zhao and A. yunnanensis J.H. Dong & C.L. Zhao, based on morphological and molecular evidence. Cai et al. [ 108] reported three new species, A. brunneoalbus Zhu L. Yang & Q. Cai, A. hirtipes Zhu L. Yang & Q. Cai, and A. pteruloides Zhu L. Yang & Q. Cai, based on morphological characteristics, ecological traits and molecular phylogenetic evidence in the family Auriscalpiaceae. To date, Artomyces consists of 24 registered names of which 22 species are accepted worldwide (Index Fungorum 2025).
Note 7 Auriscalpium Gray
Index Fungorum number: IF17139
Type species – Auriscalpium vulgare Gray
Notes – Auriscalpium (Auriscalpiaceae, Russulales) has about eight widely distributed species, and is characterized by pale to dark brown basidiocarps that are laterally to centrally stipitate and amyloid-ornamented basidiospores [ 109] . The genus is commonly known as 'the cone tooth', and A. vulgare is widely reported from the Northern Hemisphere [ 109] . It is the only known species of Auriscalpium which grows and reproduces on cones of various conifers; all other species inhabit soil or deadwood [ 26, 109− 111] . Wang & Yang [ 112] described two species, A. microsporum P.M. Wang & Zhu L. Yang, and A. orientale P.M. Wang & Zhu L. Yang inferred from morphological characteristics and molecular markers (ITS, nLSU, and rpb2). Currently, 19 species are accepted in Auriscalpium (Index Fungorum 2025).
Note 8 Dentipratulum Domański
Index Fungorum number: IF17488
Type species – Dentipratulum bialoviesense Domański
Notes – Domański [ 113] described the genus Dentipratulum with D. bialoviesense Domański to accommodate this mucronelloid fungus. Domański [ 113] placed Dentipratulum in family Hericiaceae, but the presence of sulfocystidia detected by Boidin & Gilles [ 114] indicated its affinity to family Auriscalpiaceae. The genus is characterized by basidiomata forming clusters of downwards-growing individual spines, scattered or crowded, pointed and unbranched; spines connected with very thin, rudimentary, discontinuous or continuous subiculum; monomitic hyphal system, generative hyphae with clamp connections, hyaline or rarely brownish in subiculum, thin to slightly thick-walled; gloeopleurous hyphae and presenting gloeocystidia, sulfopositive; clavate basidia with 2–4 sterigmata; and broadly ellipsoidal to globose basidiospores, slightly thick-walled, apiculate, ornamented, strongly amyloid, acyanophilous [ 115] . Macro-morphological similarities of the basidiomata of Dentipratulum and Mucronella Fr. are not reflected in their phylogenetic relationships, with Mucronella belonging to order Agaricales [ 116] . Based on the MycoBank database (2025), and Index Fungorum (2025), Dentipratulum has only three species, viz. D. bialoviesense Domański, found in the Białowieża Primeval Forest in Poland, and later reported from several locations in Eurasia; D. crystallinum Karasiński from the Kuril Islands and France; and D. khuranae Karasiński & Piątek from India [ 116] .
Note 9 Gloiodon P. Karst.
Index Fungorum number: IF17677
Type species – Gloiodon strigosus (Sw.) P. Karst.
Notes – Gloiodon was introduced by Karsten and typified by G. strigosus (Sw.) P. Karst. This genus is characterized by dark brown, effused-reflexed, or sessile basidiomata; spines with very dark but acquiring a whitish or bluish bloom from the ripening spores; dimitic hyphal system with thin-walled generative hyphae and clamp connections; clavate basidia with four spores, and a basal clamp; broadly ellipsoid basidiospores, minutely spinulose, and amyloid [ 117, 118] . Gloiodon includes five known species, G. hirtus (Fr.) P. Karst., G. nigrescens (Petch) Maas Geest., G. occidentalis Ginns, G. stratosus (Berk.) Banker and G. strigosus [ 12] . According to MycoBank and He et al. [ 1] , this genus belongs to family Bondarzewiaceae. However, based on the phylogenomic relationships and divergence times of combined ITS + nLSU + mtSSU + tef1- α+ rpb2 dataset Gloiodon grouped into Auriscalpiaceae.
Note 10 Family Bondarzewiaceae Kotl. & Pouzar 1957 (=Hybogasteraceae Jülich 1982)
Index Fungorum number: IF80527
Type genus – Bondarzewia Singer
Notes – The family Bondarzewiaceae was originally introduced to accommodate wood-rotting mushrooms, with type genus Bondarzewia Singer [ 12, 21] . Bondarzewiaceae is characterized by annual to perennial, resupinate, effused-reflexed, pileatesessile, pileate-stipitate to clavarioid basidiomata, smooth, tuberculate, poroid, hydnoid hymenophore. The hyphal system is monomitic, pseudodimitic to dimitic, generative hyphae with or without clamp-connections, inamyloid, skeletal hyphae inamyloid or dextrinoid ( Amylosporus), gloeoplerous hyphae and gloeocystidia present or absent, with asperulate, spinulose, verrucose, echinulate, ridges or crests, hyaline to pigmented, amyloid basidiospores. The family comprises wood decaying fungi [ 1] . Later, other genera, such as Amylaria Corner, Amylosporus Ryvarden, Heterobasidion Bref., and Echinodontium Ellis and Everh., were added to the family [ 26] . However, it was was later suggested that Echinodontium is sister to Amylostereum Boidin and it has been treated under Echinodontiaceae Donk [ 45, 94, 119] . Members of this family are widespread and found in tropical, subtropical, and temperate climates [ 1] . Ecologically, these species are mostly associated with wood as decaying fungi; however, some such as Bondarzewia berkeleyi (Fr.) Bondartsev and Singer, B. montana (Quél.) Singer, Heterobasidion annosum (Fr.) Bref., and H. parviporum Niemelä and Korhonen, are plant pathogens [ 1, 5, 45, 120, 121] .
Note 11 Amylaria Corner
Index Fungorum number: IF17062
Type species – Amylaria himalayensis Corner
Notes – Amylaria, typified by A. himalayensis. It is characterized by clavarioid basidiomata, hyphal system dimitic with clamped generative hyphae, clavate to subventricose basidia with 2–4 sterigmata and amyloid ellipsoidal basidiospore [ 1, 25] Amylaria is a monotypic, clavorioid genus reported from Bhutan and Nepal [ 25] . The genus has been placed in Bondarzewiaceae according to recent systematics reports [ 1] . However, Hussain et al. [ 12] declared that the systematic position of this taxon in Bondarzewiaceae is questionable due to lack of sequence data and limited reports of the genus have been made after the original description.
Note 12 Amylonotus Ryvarden
Index Fungorum number: IF17069
Type species – Amylonotus africanus Ryvarden
Notes – Amylonotus was proposed by Ryvarden [ 122] based on A. africanus, but species in the genus were later treated in Wrightoporia by David & Rajchenberg [ 123] . A phylogeny based on ITS + nLSU sequence data revealed that A. labyrinthinus (= Wrightoporia labyrinthina T. Hatt.), and A. africanus (= W. pouzarii A. David & Rajchenb.) formed a well-supported lineage within family Bondarzewiaceae clade, distant from W. lenta (Overh. & J. Lowe) Pouzar, and closely related to species of Bondarzewia Singer and Heterobasidion Bref. [ 10, 11] . Amylonotus is characterized by its effused-reflexed or resupinate, sessile, pileate, soft coriaceous to brittle basidiomata with cinnamon to dark brown pileal surface; pale orange, isabelline, pale cinnamon to brown pore surface; hyphal system dimitic with clamped generative hyphae; thin- to slightly thick-walled, ellipsoid to subglobose, finely asperulate basidiospores [ 122] . Five species have been recorded in Amylonotus, viz. A. africanus Ryvarden, A. gyroporus (Corner) Y.C. Dai, Jia J. Chen & B.K. Cui, A. labyrinthinus (T. Hatt.) Y.C. Dai, Jia J. Chen & B.K. Cui, A. ramosus (A. David & Rajchenb.) Y.C. Dai, Jia J. Chen & B.K. Cui and A. tenuis G.Y. Zheng & Z.S. Bi [ 10, 11, 122, 123] .
Note 13 Amylosporus Ryvarden
Index Fungorum number: IF17072
Type species – Amylosporus graminicola (Murrill) Ryvarden
Notes – Amylosporus was introduced in 1973, initially typified with A. graminicola [ 124] . Later, A. graminicola was synonymized with A. campbellii (Berk.) Ryvarden, with the latter becoming the type species of the genus, introduced to include species having both simple septate and multi-clamped generative hyphae, and finely asperulate and amyloid basidiospores [ 125] . Amylosporus wrightii Rajchenb. is a taxonomic synonym of A. bracei (Murrill) A. David & Rajchenb. Based on ITS + nLSU sequence data and morphological characteristics, Chen et al. [ 10] proposed that Wrightoporia casuarinicola Y.C. Dai & B.K. Cui, W. efibulata I. Lindblad & Ryvarden and W. rubella Y.C. Dai are characterized by their generative hyphae lacking clamp connections, which fits the newly defined Amylosporus (clamp connections are absent in hymenium). Currently, there are 14 species of Amylosporus, namely A. annosus Y.C. Dai, P. Du & X.H. Ji, A. auxiliadorae Drechsler-Santos & Ryvarden, A. bracei (Murrill) A. David & Rajchenb., A. campbellii, A. casuarinicola (Y.C. Dai & B.K. Cui) Y.C. Dai, Jia J. Chen & B.K. Cui, A. daedaliformis G.Y. Zheng & Z.S. Bi, A. efibulatus, A. guaraniticus Campi & Robledo, A. rubellus, A. ryvardenii Stalpers, A. succulentus Jia J. Chen & L.L. Shen, A. sulcatus F.C. Huang & Bin Liu, A. wadinaheezicus S. Hussain, Al-Sadi, Al-Yahya'ei, Al-Kharousi & Al-Owaisi, A. wrightii Rajchenb. [ 10, 12, 126] . Initially, the genus was placed in Wrightoporiaceae Jülich, but recently it was treated under Bondarzewiaceae [ 1, 5, 12, 121] .
Note 14 Bondarzewia Singer
Index Fungorum number: IF17176
Type species – Bondarzewia montana (Quél.) Singer
Notes – Bondarzewia was established based on B. montana, now considered a synonym of B. mesenterica (Schaeff.) Kreisel, originally described from Abies in the Pyrenees mountains ( B. mesenterica in Germany). It is a remarkable genus because the species usually have relatively large and imbricate basidiomata. Some species are edible and have medicinal potential, such as B. mesenterica [ 11, 12, 14, 127, 128] , while others are plant pathogens [ 120] . The genus is characterized by annual growth habit, pileate basidiomata with poroid hymenophores and it is morphologically similar to many species of order Polyporales. However, it has strongly amyloid and ornamented basidiospores and recent phylogenetic analyses revealed that it belongs to Russulales [ 12, 26, 28, 45, 128, 129] . Chen et al. [ 11] conducted a taxonomic study of Bondarzewia based on many samples covering a wide geographic range. With the aid of morphological and phylogenetic analyses of ITS and nuc 28S rDNA D1-D2 domain (28S) sequences, three new species were described and three new combinations were proposed [ 11] . Hussain et al. [ 12] estimated the divergence time of Bondarzewiaceae (Russulales) based on ITS-28S dataset, revealed that the species diversified approximately 114 million years ago (Mya). The clade consisted of Bondarzewia, Heterobasidion, Gloiodon, Laurilia, Lauriliella, and Wrightoporia, which estimated stem age of the clade is approximately 90 Mya.
Note 15 Heterobasidion Bref.
Index Fungorum number: IF17745
Type species – Heterobasidion annosum (Fr.) Bref.
Notes – Heterobasidion was typed by H. annosum. It is characterized by effused-reflexed to sessile basidiomata, dimitic hyphal system with dextrinoid skeletal hyphae, generative hyphae without clamp connections, and finely asperulate and nonamyloid basidiospores, and is distributed in both Northern and Southern Hemispheres [ 9] . Based on ITS, nLSU, rpb1, rpb2, gapdh, atp6, and mtSSU, Chen et al. [ 13] suggested that ancestral Heterobasidion species originated in Eurasia during the Early Miocene, followed by dispersal and speciation to other continents during the Middle Miocene and Early Pliocene. Heterobasidion is a global complex of woody plant pathogens and saprobes whose host range comprises over 200 plant taxa, most of which are conifers [ 13] . Heterobasidion has a negative impact on conifers, both ecologically and economically, by reducing site productivity and the amount of harvestable timber [ 9, 12] . Heterobasidion abietinum Niemelä and Korhonen, distributed in Italy, is associated with Abies alba and Picea abies; H. amyloideum Y.C. Dai, Jia J. Chen and Korhonen with Abies in China; H. annosum from Italy and Russia, associated with different species of Pinus; H. araucariae P.K. Buchanan with trees of Araucaria cunninghamii, reported from Australia; H. armandii Y.C. Dai, Jia J. Chen and Yuan Yuan with Pinus armandii, found in China; H. australe Y.C. Dai and Korhonen from China, associated with Pinus species; H. insulare (Murrill) Ryvarden from China in association with Pinus massoniana; H. irregulare Garbel. and Otrosina is a South American species associated with Pinus; H. linzhiense Y.C. Dai and Korhonen is a Chinese species associated with Abies and Picea; H. occidentale is pathogenic to various conifer hosts; H. orientale Tokuda, T. Hatt. and Y.C. Dai associated with fallen conifer trunk, reported from China; H. parviporum Niemelä and Korhonen associated with Picea abies, distributed in Europe and Asia; H. subinsulare Y.C. Dai, Jia J. Chen and Yuan Yuan is reported from China, associated with wood of Pinus; H. subparviporum Y.C. Dai, Jia J. Chen and Yuan Yuan with wood of Abies and Picea, reported from China; and H. tibeticum Y.C. Dai, Jia J. Chen and Korhonen with Pinus wood from China [ 9, 12] .
Note 16 Laurilia Pouzar
Index Fungorum number: IF17912
Type species – Laurilia sulcata (Burt) Pouzar
Notes – Laurilia is a monotypic genus with L. sulcata, characterized by effuse-reflexed basidiomata with smooth to tuberculate hymenophore, and a trimitic hyphal system [ 81] . Laurilia sulcata is widely distributed in boreal conifer forests in the northern hemisphere [ 81] . It is characterized by having basidiomata of perennial, leathery or ligneous, resupinate, effused and confluent, or partly pileate, especially on vertical sides of the substrate; hyphal system is trimitic with skeletals and binding hyphae with thick walls and few clamps, and thin-walled, fibulate generative hyphae; tinder-layer mainly dimitic, composed largely of horizontal, brown, thick-walled skeletal hyphae; metuloid cystidia numerous, thick-walled, encrusted; clavate basidia with 4 sterigmata and basal clamp; and globose spores, somewhat thick-walled, echinulate and amyloid [ 130] . Only two species, L. sulcata and L. taxodii (Lentz & H.H. McKay) Pouzar are accepted (Index Fungorum 2025).
Note 17 Lauriliella S.H. He & Nakasone
Index Fungorum number: IF819211
Type species – Lauriliella taxodii (Lentz & H.H. McKay) S.H. He & Nakasone
Notes – Lauriliella was established by He & Nakasone, and typified by L. taxodii (= Stereum taxodii Lentz & H.H. McKay). It is a perennial genus with effused-reflexed, pileate or umbonate, woody basidiomata, hymenophore smooth to tuberculate, basidia with basal clamp connections, basidiospores broadly ellipsoid to subglobose, hyaline, thick-walled, echinulate, and amyloid [ 81] . The genus comprises two species, L. taxodii and L. taiwanensis S.H. He and Nakasone. Lauriliella taxodii is distributed in USA, causing white stringy rot or brown powdery rot in living Taxodium distichum. Similarly, L. taiwanensis is reported from China, causing white rot in living Chamaecyparis formosensis [ 81] . With the transfer of Laurilia taxodii into Lauriliella, Laurilia becomes monotypic. Although similar, Laurilia and Lauriliella can be distinguished by several critical features. Laurilia causes a white stringy rot or white pocket rot in dead coniferous wood, whereas Lauriliella creates large pockets of decayed wood scattered in the heartwood of Taxodium and Chamaecyparis which is somewhat stringy or laminated. The hymenophore is light yellow or pink to salmon-colored in Laurilia but gray, orange, or brown in Lauriliella. Microscopically, unbranched skeletal hyphae are dominant in the context of Lauriliella, whereas in Laurilia unbranched skeletals and richly branched binding hyphae are present. Davidson et al. [ 131] and Nakasone [ 132] noted the differences in cultures, with L. sulcata growing faster, producing strong oxidase reactions, and developing conidia (Spiniger anamorph). In contrast, L. taxodii cultures grew very slowly and produced no or weak oxidase reactions and produced chlamydospores.
Note 18 Stecchericium D.A. Reid
Index Fungorum number: IF18581
Type species – Stecchericium fistulatum (G. Cunn.) D.A. Reid
Notes – Stecchericium was established by D.A. Reid based on S. seriatum (Lloyd) Maas Geest. (= S. fistulatum G. Cunn.) as the type. It is characterized by pileate basidiomata, hydnoid hymenophore, monomitic to imperfectly dimitic hyphae system, tubular and thick-walled gloeocystida, and finely asperulate, strongly amyloid basidiospores [ 133] . This genus resembles Steccherinum Gray in macroscopic characters, and they could be easily confused with each other in the field. But Steccherinum has encrusted skeletocystidia and smooth, non-amyloid basidiospores [ 133] . According to MycoBank, Stecchericium belongs to Wrightoporiaceae; however, recent studies [ 1, 5, 134] classified it in Bondarzewiaceae. The known species are S. abditum Maas Geest., found on rotten log in Australia, S. acanthophysium T. Hatt. and Ryvarden on hardwood reported from Japan, S. isabellinum Corner associated with fallen wood in the Amazon forest, S. rusticum Maas Geest., on dead wood in Singapore, S. seriatum were found in Singapore and Malaysia, and S. dimiticum Douanla-Meli associated with angiosperm wood, reported from Cameroon [ 135] .
Family Echinodontiaceae Donk 1961
Index Fungorum number: IF80722
Type genus – Echinodontium Ellis & Everh.
Notes – Echinodontiaceae is characterized by annual to perennial, resupinate, effused-reflexed to pileate-sessile basidiomata, smooth, poroid to hydnoid hymenophore, monomitic, pseudodimitic to dimitic hyphal system, generative hyphae with or without clamp-connections, inamyloid, skeletal hyphae inamyloid or dextrinoid ( Larssoniporia), gloeocystidia present or incrusted cystidia absent or present, with smooth, asperulate, spinulose, verrucose, hyaline to pigmented, amyloid basidiospores. This family consists of wood decaying fungi [ 1] .
Note 19 Amylostereum Boidin
Index Fungorum number: IF17073
Type species – Amylostereum chailletii (Pers.) Boidin
Notes – Amylostereum with A. chailletii as its type is a fascinating genus, as some species are symbionts of mycophagus horntails [ 136] . Species are characterized by numerous thick-walled and apically encrusted cystidia in hymenium and context, nodose-septate generative hyphae and distinctly amyloid basidiospores [ 137] . Six species, A. areolatum (Chaillet ex Fr.) Boidin, A. chailletii, A. ferreum (Berk. & M.A. Curtis) Boidin & Lanq., A. laevigatum (Fr.) Boidin, A. orientale S.H. He & Hai J. Li, A. stillwellii Slippers, K.N.E. Fitza & J.D. Allison, are recognized worldwide, all associated with gymnosperm hosts [ 138− 140] . Traditionally, Amylostereum has been placed in family Stereaceae due to its morphological similarity to Stereum Hill ex Pers. [ 141, 142] . However, phylogenetic analysis based on DNA sequences showed that Amylostereum is close to Echinodontium Ellis & Everh. and should be placed in the monotypic family Amylostereaceae [ 138, 139] . However, divergence times showed that Amylostereum was placed in family Echinodontiaceae [ 1] . The present study, based on the ITS, nLSU, mtSSU, rpb2, and tef1- α dataset, confirmed that Amylostereum formed an independent lineage in Echinodontiaceae.
Note 20 Echinodontiellum S.H. He & Nakasone
Index Fungorum number: IF819204
Type species – Echinodontiellum japonicum (Imazeki) S.H. He & Nakasone
Notes – Echinodontiellum was established by S.H. He & Nakasone to accommodate E. japonicum (= Echinodontium japonicum Imazeki). It is characterized by perennial, resupinate to slightly effused-reflexed basidiomata, gray to olive gray and dentate hymenophore, dimitic hyphal system, thin- to thick-walled generative hyphae nodose-sepatate, with scattered secondary simple septa, thick-walled to subsolid skeletal hyphae light brown, clavate, hyaline to light brown, thick-walled cystidia apically encrusted, blunt, embedded or slightly projected, clavate basidia with four sterigmata, and a basal clamp connection, and ellipsoid, thick-walled, echinulate, amyloid basidiospores [ 81] . Morphological differences between Echinodontium Ellis & Everh and Echinodontiellum are few but significant. Basidiomata of Echinodontiellum are effused to effused-reflexed, whereas they are effused-reflexed to pileate, rarely effused, in Echinodontium. The context in Echinodontiellum is cinnamon to olive gray or brownish gray that darkens in KOH. In comparison, the context of Echinodontium species are brick red or brownish orange turning maroon in KOH or pale brown to brown (in Echinodontium ryvardenii Bernicchia & Piga). Furthermore, Echinodontiellum japonicum is sister to Echinodontium s.s. and segregated into a separate genus because of ecological, basidiomata, and molecular criteria [ 81] .
Note 21 Echinodontium Ellis & Everh.
Index Fungorum number: IF17540
Type species – Echinodontium tinctorium (Ellis & Everh.) Ellis & Everh.
Notes – Species of Echinodontium sensu lato are characterized by conspicuous basidiomata, dentate to smooth hymenophores, encrusted cystidia, and ornamented, amyloid basidiospores [ 81] . Based on a concatenated dataset of ITS and 28S sequences of taxa in Russulales, E. tinctorium, E. tsugicola (Henn. & Shirai) Imazeki, and E. ryvardenii were confirmed in family Echinodontiaceae [ 81] . Echinodontium sulcatum (Burt) H.L. Gross and E. taxodii (Lentz & H.H. McKay) H.L. Gross were also placed in Laurilia Pouzar and Lauriliella S.H. He & Nakasone in some studies [ 81, 143] . Currently, only five species, E. ballouii (Banker) H.L. Gross, E. japonicum Imazeki, E. ryvardenii, E. tinctorium, and E. tsugicola are accepted (Index Fungorum 2025). Tabata et al. [ 138] demonstrated that Echinodontium and Amylostereum were phylogenetically related, and belonged in Echinodontiaceae. This was confirmed by subsequent studies [ 10, 81, 94, 144, 145] .
Note 22 Larssoniporia Y.C. Dai, Jia J. Chen & B.K. Cui
Index Fungorum number: IF812223
Type species – Larssoniporia tropicalis (Cooke) Y.C. Dai, Jia J. Chen & B.K. Cui
Notes – Larssoniporia was proposed to accommodate L. tropicalis ( Wrightoporia tropicalis (Cooke) Ryvarden) and L. incrustatocystidiata Y.C. Dai, Jia J. Chen & B.K. Cui [ 10] . Larssoniporia is characterized by its woody hard basidiomata with tough tubes, dextrinoid skeletal hyphae, presence of cystidia with crystals at tips, and gloeocystidia, finely asperulate and amyloid basidiospores, and is tropical in distribution [ 10] . Phylogeny of Russulales inferred from ITS and 28S sequence data, placed Larssoniporia into family Echinodontiaceae [ 81] .
Note 23 Subulicystidiella Y.L. Deng & C.L. Zhao gen. nov.
Index Fungorum number: IF861371
Type species – Subulicystidiella murina Y.L. Deng & C.L. Zhao.
Etymology – referring to the subulate cystidia of the type species.
Notes – Subulicystidiella is characterized by annual, resupinate, membranous basidiomata, closely adnate, smooth, gray to charcoal gray hymenial surface, a monomitic hyphal system, thin-walled, generative hyphae with clamp connections, subulate, yellowish brown, thick-walled, cystidia with acute tips, encrusted with crystals in the apical part, thin-walled, smooth, barrelled cystidia with a clamp connection at base, cylindrical basidia with four sterigmata and a basal clamp connection thin-walled, smooth, globose basidiospores. Our study, based on ITS + nLSU + mtSSU + rpb2 + tef1- α sequence data and ITS + nLSU sequence data, showed that Subulicystidiella clustered within family Echinodontiaceae, closely related to Larssoniporia and Amylostereum, and formed a separate clade. However, morphologically, Larssoniporia differs from Subulicystidiella by its hard basidiomata with tough tubes, hyphal system dimitic, dextrinoid skeletal hyphae, and finely asperulate basidiospores [ 10] . Amylostereum can be distinguished from Subulicystidiella by its thick-walled and apically encrusted cystidia in hymenium and context, nodose-septate generative hyphae and distinctly amyloid basidiospores [ 10] . Thus, a new genus Subulicystidiella is introduced, based on phylogenetic analyses and morphological characteristics ( Figs. 7, 8).
Subulicystidiella murina Y.L. Deng & C.L. Zhao, sp. nov. Figures 7, 8
Index Fungorum number: IF861374

Figure 7.
Basidiomata and microscopic structures of Subulicystidiella murina (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Barrelled gloeocystidia. (f) Subulate cystidia (g) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 µm, (d)–(g) = 10 µm.
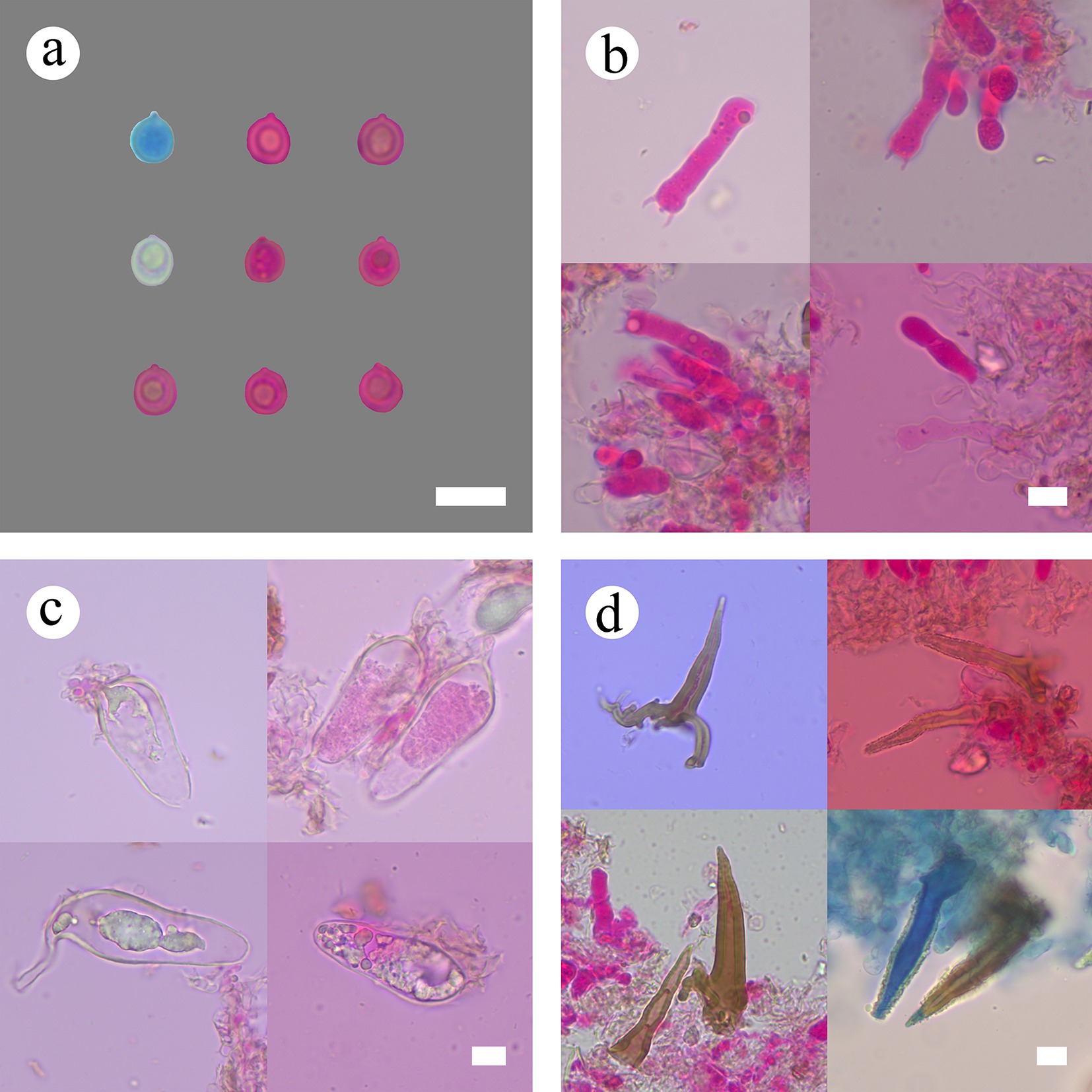
Figure 8.
Sections of hymenium of Subulicystidiella murina (holotype). (a) Basidiospores. (b) Basidia and basidioles. (c) Barrelled cystidia. (d) Subulate cystidia. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Subulicystidiella murina differs from other species by the membranous basidiomata, gray to charcoal gray hymenial surface, hyphal system monomitic with clamped generative hyphae, and globose basidiospores.
Etymology – referring to the murine hymenial surface of the type specimens.
Type – China, Yunnan Province, Xishuangbanna, Wild Elephant Valley, 22°17′ N, 100°85′ E, 900 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 25 January, 2024, CLZhao 35801 (SWFC).
Description – Basidiomata annual, resupinate, membranous, closely adnate, thin, without odor or taste when fresh, up to 5.5 cm long, 1.5 cm wide, 150 µm thick. Hymenial surface smooth, gray when fresh, gray to charcoal gray upon drying. Sterile margin gray, thinning out, up to 1 mm wide.
Hyphal system monomitic; generative hyphae with clamp connections, colorless, thin-walled, smooth, moderately branched, 2.5–5.5 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Cystidia with two types: (1) subulate cystidia, numerous, some slightly sigmoid, yellowish brown, thick-walled, with acute tips, encrusted with crystals in the apical part, 18–60 × 7–49 µm, projecting up to 36 µm above the hymenium; (2) barrelled cystidia, colorless, thin-walled, smooth, filled with refractive material, with a clamp connection at base, 41–57 × 6–10 µm. Basidia cylindrical, with one oil drop, four sterigmata, and a basal clamp connection, 20–34 × 5–8.5 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores globose, colorless, thin-walled, smooth, with some oil drops, CB–, IKI–, (5.3–)5.7–7.4(–7.8) × (4.3–)4.6–6.3(–6.9) µm, L = 6.42 µm, W = 5.55 µm, Q = 1.16 (n = 60/2).
Additional material examined – China, Yunnan Province, Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 21 July 2023, CLZhao 30728 (SWFC).
GenBank accession numbers – CLZhao 35801 ITS: PV441140, LSU: PV441154, mtSSU: PV461163; CLZhao 30728 ITS: PV771055, LSU: PX418375, mtSSU: PV774692.
Family Gloeocystidiellaceae Jülich 1982
Index Fungorum number: IF81775
Type genus – Gloeocystidiellum Donk
Notes – Gloeocystidiellaceae is characterized by annual resupinate, effused-reflexed basidiomata, smooth, rarely grandinioid or odontioid hymenial surface, a monomitic hyphal system with nodose or simple-septate generative hyphae, gloeocystidia, subcylindrical to clavate basidia with four sterigmata and suballantoid, ellipsoid, subglobose or globose, verrucose or aculeate basidiospores [ 45] .
Note 24 Gloeocystidiellum Donk
Index Fungorum number: IF17663
Type species – Gloeocystidiellum porosum (Berk. & M.A. Curtis) Donk
Notes – A corticioid genus Gloeocystidiellum belonging to order Russulales was established by Donk with G. porosum as its type species. The genus is characterized by smooth, rarely grandinioid or odontioid hymenial surface, a monomitic hyphal system with nodose or simple-septate generative hyphae, gloeocystidia, subcylindrical to clavate basidia with four sterigmata and suballantoid, ellipsoid, subglobose or globose, verrucose or aculeate basidiospores [ 146] . Recently, molecular studies involving Gloeocystidiellum based on single-gene or multi-gene datasets have been carried out [ 3, 147− 151] . Phylogenetic relationships of russuloid basidiomycetes revealed that the species of Gloeocystidiellum formed two clades based on analyses of nuclear 5.8S, ITS2, and nLSU rDNA genes, in which the generic type species G. porosum grouped into clade I [ 26] . The high phylogenetic diversity among corticioid homobasidiomycetes showed that G. subasperisporum (Litsch.) J. Erikss. & Ryvarden nested into the russuloid clade and closely grouped with Gloeodontia discolor (Berk. & M.A. Curtis) Boidin [ 116] . Phylogenetic relationships inferred from 5.8S and nuLSU rDNA sequences showed that Gloeocystidiellum is clustered into family Gloeocystidiellaceae and closely related to Russulaceae within Russulales [ 45] . Analyses based on ITS and nLSU sequences indicated that Gloeocystidiellum clustered with Russulaceae Lotsy and Gloeodontia clade [ 82, 150] . In this study, analyses inferred from ITS + nLSU + mtSSU + rpb2 + tef1- α sequences ( Figs 1, 3) showed that Gloeocystidiellum is polygeneic, formed two single clades and then grouped with Gloeodontia clade and Russulaceae. Thus, Gloeocystidiellum is placed into family Gloeocystidiellaceae. Also, two new species, G. membranaceum and G. punctatum found in China are introduced, based on phylogeny and morphological characteristics.
Gloeocystidiellum membranaceum Y.L. Deng & C.L. Zhao, sp. nov. Figures 9, 10
Index Fungorum number: IF860765

Figure 9.
Basidiomata and microscopic structures of Gloeocystidiellum membranaceum (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Gloeocystidia. (e) Basidioles. (f) Basidia. (g) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 µm, (d)–(g) = 10 µm.
Figure 10.
Sections of hymenia of Gloeocystidiellum membranaceum (holotype). (a)–(c) Basidiospores and basidia. (d) Gloeocystidia. (e) Hyphae from context. (f) Section of hymenium. Scale bars: (a)–(f) = 10 µm.
Diagnosis – Differs other species by the white to cream, smooth hymenial surface, a dimitic hyphal system with clamped generative hyphae, cylindrical basidia, and ellipsoid basidiospores.
Etymology – referring to the membranous hymenial surface of the type specimen.
Type – China, Yunnan Province, Dehong, Mang City, Tongbiguan Provincial Nature Reserve, 25°50′ N, 97°36′ E; 1,000 m asl, on fallen angiosperm branch, 30 June, 2024, CLZhao 37038 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, membranous, without odor or taste when fresh, up to 10 cm long, 1 cm wide, and 150 µm thick. Hymenial surface smooth, white to cream when fresh, buff to pale brown upon drying. Sterile margin thin, white, up to 1 mm.
Hyphal system monomitic, generative hyphae bearing simple septa, colorless, thin-walled, branched, 2.5–4.5 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Gloeocystidia abundant, cylindrical, basally inflated and slightly tapering towards the apices, thin-walled, 26.5–72.5 × 6–12.5 µm. Basidia cylindrical, colorless, thin-walled, with a basal simple septum and four sterigmata, 13–23 × 3–5 µm, basidioles dominant, similar in shape to basidia, but slightly smaller. Basidiospores ellipsoid, colorless, thin-walled, smooth, IKI+, CB–, (3.8–)4.4–5.3(–5.8) × (2.5–)3.2–3.9(–4.4) µm, L = 4.82 µm, W = 3.48 µm, Q = 1.35–1.39 (n = 60/2).
Additional material examined – China, Yunnan Province, Qujing, Qilin District, Cuishan Park, 25°54′ N, 103°69′ E; 1,875 m asl, on fallen angiosperm branch, 4 November, 2022, CLZhao 26028 (SWFC).
GenBank accession numbers – CLZhao 37038 ITS: PV940928, LSU: PX070092, rpb2: PX432797, rpb1: PX441306, tef1- α: PX439082; CLZhao 26028 ITS: PV940927.
Notes – Based on the multigene phylogeny ( Figs 1, 3), Gloeocystidiellum membranaceum clustered together with G. bisporum, G. clavuligerum (Höhn. & Litsch.) Nakasone, G. kenyense Hjortstam, and G. porosellum. However, morphologically, G. bisporum differs from G. membranaceum by having basidia with two sterigmata, oval to subglobose basidiospores [ 152] ; G. clavuligerum differs by its cream to ochraceous hymenophore, generative hyphae with clamp connections (Nakasone 1982); G. kenyense has larger fusiform cystidia (20–100 × 8–18 µm), and subcylindrical basidia [ 153] ; G. porosellum has pale cream coloured, smooth basidiomata, thin-walled, subfusiform cystidia, constricted basidia [ 154] .
Gloeocystidiellum punctatum Y.L. Deng & C.L. Zhao, sp. nov. Figure 11
Index Fungorum number: IF852412

Figure 11.
Basidiomata and microscopic structures of Gloeocystidiellum punctatum (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidioles. (e) Basidia. (f) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 µm, (d)–(f) = 10 µm.
Diagnosis – Differs from other species in the genus by grandinioid, white to cream hymenial surface, thin-walled generative hyphae with clamp connections, and thin-walled ellipsoid basidiospores.
Etymology – referring to the punctate basidiomata.
Type – China, Yunnan Province, Zhaotong, Qiaojia County, Yaoshan Town, Yaoshan National Nature Reserve, 27°08′ N, 103°09′ E, 2,220 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 23 August, 2020, CLZhao 20755 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, ceraceous, up to 6 cm long, 1.9 cm wide, and 0.2 mm thick. Hymenial surface grandinioid, white to cream when fresh, cream upon drying. Sterile margin thin, slightly cream, up to 1 mm.
Hyphal system monomitic, generative hyphae with clamp connections, colorless, thin-walled, branched, 1.5–3.8 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Gloeocystidia abundant, tubular, basally inflated and slightly tapering towards the apices, thick-walled, thinning towards apex, 51.5–106 µm long, 8.5–15 µm wide, the protoplasmic content granular and yellowish in KOH. Basidia subclavate, colorless, thin-walled, with a basal clamp connection and four sterigmata, 16–24 × 3.5–5 µm, basidioles dominant, similar shape to basidia, but slightly smaller. Basidiospores ellipsoid, colorless, thin-walled, verruculose, IKI+, CB–, 4–6.5 × 2.5–4.5 µm, L = 5.03 µm, W = 3.64 µm, Q = 1.3–1.47 (n = 60/2).
Additional material examined – China, Yunnan Province, Zhaotong, Qiaojia County, Yaoshan Town, Yaoshan National Nature Reserve, 27°08′ N, 103°09′ E, 2,220 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 23 August, 2020, CLZhao 20657 and CLZhao 20907 (SWFC); Yiliang County, Luozehe Town, Li Jiaping Village, 27°49′ N, 103°91′ E, 1,700 m asl, on dead bamboo, leg. C.L. Zhao, 25 August, 2022, CLZhao 23890 (SWFC); Qujing City, Qilin District, Cuishan, 25°54′ N, 103°69′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 6 November, 2022, CLZhao 26935 (SWFC); Zhaotong, Wumeng Mountain National Nature Reserve, 27°32′ N, 103°72′ E, 2,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 2 September, 2023, CLZhao 33627 (SWFC); Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 97°94′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 21 July, 2023, CLZhao 30863, and CLZhao 30864 (SWFC); Diqing, Weixi County, Weideng Town, Fuchuan Village, 27°10′ N, 99°16′ E, 2,000 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 12 October, 2023, CLZhao 34197 (SWFC); Dehong, Mang City, Mengjia Town, Sanxian Cave Park, 24°38′ N, 98°62′ E, 1,370 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 30 June, 2024, CLZhao 37129 (SWFC); Tengchong, Tuantian Town, Gaoligong Mountain National Nature Reserve, 25°55′ N, 98°58′ E., 2,600 m asl, on fallen angiosperm branch, 7 July, 2024, CLZhao 39107, CLZhao 39124, CLZhao 39155, and CLZhao 39158 (SWFC).
GenBank accession numbers – CLZhao 20755 ITS: PP356586, LSU: PP785346, mtSSU: PV774693; CLZhao 20907 ITS: PP356587, LSU: PP785347, mtSSU: PV774694; CLZhao 23890 ITS: PV771056, LSU: PV771607, mtSSU: PV774695; CLZhao 26935 ITS: PV771057; CLZhao 33627 ITS: PP356588; CLZhao 30863 ITS: PV771058, LSU: PP785348, mtSSU: PV774696; CLZhao 30864 ITS: PV771059, LSU: PP785349, mtSSU: PV774697; CLZhao 34197 ITS: PV771060; CLZhao 37129 ITS: PV771061, LSU: PV771608, mtSSU: PV774698; CLZhao 39107 ITS: PV771062, LSU: PV771609, mtSSU: PV774699; CLZhao 39124 ITS: PV771063, LSU: PV771610, mtSSU: PV774700; CLZhao 39155 ITS: PV771064, LSU: PV771611, mtSSU: PV774701; CLZhao 39158 ITS: PV771065, LSU: PV771612, mtSSU: PV774702.
Notes – The phylogram ( Fig. 3) revealed that Gloeocystidiellum punctatum is nested within the Gloeocystidiellum clade, as a sister to G. porosum. However, morphologically, G. porosum has yellowish to isabelline, smooth hymenophore, thin- to slightly thick-walled, tubular to sinuous gloeocystidia (80–150 × 8–15 µm), and subcylindrical to ellipsoid basidiospores (5–6 × 2.5–3 µm). Morphologically, G. kenyense, G. lojanense A. Jaram., D. Cruz & Decock and G. yunnanense Y.L. Zhao & C.L. Zhao are similar to G. punctatum. However, G. kenyense differs in its cream to ochraceous basidiomata, smaller gloeocystidia (35–85 × 7–16 µm), and longer cylindrical basidia (21–27 × 4–5 µm) [ 149] ; G. lojanense can be distinguished by its bright grayish white and slightly light yellow hymenial surface, thin- to thick-walled generative hyphae, smaller gloeocystidia (80–90 × 7–8 µm), larger basidia (25–35 × 5–6 µm), and longer basidiospores (6.5–8 µm × 3.4–4.5 µm) [ 149] ; G. yunnanense is distinct by having smaller clavate (25–48.2 × 6.4–9.6 µm) and tubular (22.6–47.2 × 4.1–9.5 µm) gloeocystidia, basidia (12.5–14.5 × 3.5–4.5 µm), and basidiospores (3.9–4.7 × 3–3.5 µm) [ 150] .
Note 25 Family Gloeodontiaceae Y.L. Deng & C.L. Zhao, fam. nov.
Index Fungorum number: IF 861348
Type genus – Gloeodontia Boidin
Etymology – referring to the type genus Gloeodontia.
Description – Basidiomata resupinate or effused-reflexed, ceraceous to membranous consistency. Hymenophore smooth to odontioid. Hyphal system monomitic; generative hyphae with clamp connections. Cystidia present, Basidia cylindrical to urniform with four sterigmata, thin to thick-walled. Basidiospores ellipsoid to globose, asperulate, acyanophilous, and amyloid.
Notes – Gloeodontia was created by Boidin [ 155] and could not be assigned to any recognized family of Russulales, and it was treated as incertae sedis. Recent research have reported some new species based on morphology and phylogenetic analyses. However, phylogenetic tree analysis indicated that Gloeodontia formed a separate clade, and it was not placed in any recognized family of Russulales [ 30, 82] . In this study, based on combined ITS, nLSU, mtSSU, rpb2, and tef1- α data of Russulales, the phylogenetic analyses and divergence time ( Figs 1, 2) also showed that Gloeodontia could not be assigned to any recognized family. The divergence time of the Gloeodontia clade occurred in 178.7 Mya with a 95% highest posterior density (HPD) of 139.36–218.13 Mya. Thus, the genus is placed into the new family Gloeodontiaceae.
Note 26 Gloeodontia Boidin
Index Fungorum number: IF17666
Type species – Gloeodontia discolor (Berk. & M.A. Curtis) Boidin
Notes – Gloeodontia Boidin was typified by G. discolor (Berk. & M.A. Curtis) Boidin [ 155] , and characterized by a combination of resupinate or effused-reflexed basidiomata of ceraceous to membranous consistency, smooth to odontioid hymenophore, a monomitic or dimitic hyphal system, clamped hyphae, presence of cystidia, cylindrical to urniform basidia, and colorless to pale yellow, thick-walled, asperulate, ellipsoid to globose, acyanophilous and amyloid basidiospores [ 155] . Phylogenetic relationships of russuloid basidiomycetes with an emphasis on non-gilled taxa revealed that four species of Gloeodontia formed a monophyletic clade that grouped with Russulales and Stereales clades [ 26, 30, 84] , and additionally, a new combination G. subasperispora was proposed [ 26] . Leal-Dutra et al. [ 84] studied Lachnocladiaceae and Peniophoraceae (Russulales) and showed that Gloeodontia formed a single clade and then grouped with the clades, Amylostereaceae, Auriscalpiaceae, Bondarzewiaceae, Gloeocystidiellum, Russulaceae, and Stereaceae. Nine species have been accepted in the genus [ 26, 30, 84] . In this study, a new species G. sinensis is proposed based on ITS + nLSU + mtSSU + rpb2 + tef1- α and ITS + nLSU data ( Figs 1, 6), and morphological characteristics ( Figs 12, 13).
Gloeodontia sinensis Y.L. Deng & C.L. Zhao, sp. nov. Figures 12, 13
Index Fungorum number: IF860766

Figure 12.
Basidiomata and microscopic structures of Gloeodontia sinensis (holotype). (a) Basidiomata on the substrate. (b) Characteristics of hymenophore. (c) Basidiospores. (d) Basidioles. (e) Basidia. (f) Cystidia. (g) Section of the hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 μm, (d)–(g) = 10 μm.
Figure 13.
Sections of hymenium of Gloeodontia sinensis (holotype). (a)–(c) Basidiospores. (d)–(f) Basidia and basidioles. (g)–(i) Cystidia. Scale bars: (a)–(i) = 10 µm.
Diagnosis – Differs from other species by the coriaceous basidiomata with white to cream hymenial surface odontoid, a monomitic hyphal system with clamped generative hyphae, and ellipsoid, verrucose basidiospores (3.4–4.5 × 2.4–3.4 µm).
Etymology – referring to the locality (China) of the type specimen.
Type – China, Yunnan Province, Diqing, Weixi County, Weideng Town, Fuchuan village, 27°09′ N, 99°17′ E, 1,910 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 12 October, 2023, CLZhao 34280 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, coriaceous, without odor or taste when fresh, up to 8 cm long, 2 cm wide, and 150 μm thick. Hymenial surface odontoid, thin, white to cream when fresh, cream to buff upon drying. Sterile margin white, up to 1 mm.
Hyphal system monomitic; generative hyphae with clamp connections, colorless, thin-walled, smooth, moderately branched, 2–3 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Cystidia cylindrical with a mastoid at the tip, 10–15 × 4–5 µm. Basidia cylindrical, with a basal clamp connection and four sterigmata, 11–18 × 3–4 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores ellipsoid, colorless, thin-walled, verrucose, amyloid, CB–, (3.3–)3.4–4.5(–4.8) × (2.3–)2.4–3.4(–3.7) µm, L = 3.98 µm, W = 2.87 µm, Q = 1.38–1.39 (n = 60/2).
Additional material examined – China, Yunnan Province, Diqing, Weixi County, Zhonglu Town, Lagaluo village, 27°15′ N, 99°15′ E, 3100 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 14 October, 2023, CLZhao 34748 (SWFC).
GenBank accession numbers – CLZhao 34280 ITS: PV147170, LSU: PV185856, mtSSU: PV283189, rpb1: PX441307; CLZhao 34748 ITS: PV147171, LSU: PV185857, mtSSU: PV399821, tef1- α: PV400175.
Notes – The phylogenetic analyses ( Figs 1, 6) revealed that Gloeodontia sinensis clustered within the genus Gloeodontia, and is closely related to G. columbiensis and G. subasperispora. However, G. columbiensis differs from G. sinensis by having thick-walled, cylindrical cystidia, both larger thin-walled, vesicular to cylindrical gloeocystidia (20–60 × 5–8 µm), clavate basidia (15–35 × 5–6 µm), and ellipsoid basidiospores (5.5–7 × 3.5–4.5 µm) [ 156] ; G. subasperispora can be distinguished from G. sinensis by its greyish-white to pale ochraceous, smooth hymenial surface, longer yellowish gloeocystidia (20–30 × 3–5 µm) [ 26] . Morphologically, G. sinensis is reminiscent of G. yunnanensis C.L. Zhao by thin-walled generative hyphae with clamp connections, sharing asperulate basidiospores. However, G. yunnanensis has cream to buff to brownish vinaceous smooth hymenial surface, larger obclavate gloeocystidia (32–57 × 6–9 µm), and thick-walled, subglobose to globose basidiospores measuring 3.3–4.3 × 2.5–3.5 µm [ 82] .
Family Hericiaceae Donk 1964
Index Fungorum number: IF80854
Type genus – Hericium Pers.
Notes – Hericiaceae is characterized by annual, resupinate, effused-reflexed, pileate-sessile to flabelliform basidiomata, smooth, poroid to hydnoid hymenophore, monomitic or dimitic hyphal system, generative hyphae with clamp-connections, inamyloid or amyloid, skeletal hyphae inamyloid or dextrinoid ( Pseudowrightoporia, Wrightoporiopsis), gloeoplerous hyphae and gloeocystidia present or absent, with asperulate, spinulose or echinulate, hyaline, amyloid basidiospores. The members of this family include wood decaying fungi [ 1] .
Note 27 Dentipellicula Y.C. Dai & L.W. Zhou
Index Fungorum number: IF564153
Type species – Dentipellicula taiwaniana (Sheng H. Wu) Y.C. Dai & L.W. Zhou
Notes – Dentipellicula was segregated from Dentipellis and proposed for two hydnoid fungal species, Dentipellis leptodon (Mont.) Maas Geest. and Dentipellis taiwaniana Y.C. Dai, G.M. Gates, X.H. Ji & P. Du which were transferred to the genus Dentipellicula with the latter designated as the generic type. Dentipellicula is characterized by its annual, effused-reflexed to pileate basidiomata, hydnoid hymenophore, soft corky, cream to ivory yellow to buff to cinnamon-buff spines, a monomitic hyphal system; generative hyphae with clamp connections, slightly thick-walled, minutely rough basidiospores [ 30] . Chen et al. [ 157] proposed D. austroafricana Jia J. Chen, L.L. Shen & Y.C. Dai, which was recognized in the Dentipellicula clade and distant from Dentipellis based on combined ITS and nLSU sequences. Another species, D. guyanensis Yuan Yuan, M. Zhou, Jia J. Chen & Vlasák shares several morphological characteristics with other Dentipellicula species, such as an annual growth habit, a monomitic hyphal system with non-amyloid, non-dextrinoid and acyanophilous generative hyphae [ 30, 158] .
Note 28 Dentipellis Donk
Index Fungorum number: IF17487
Type species – Dentipellis fragilis (Pers.) Donk
Notes – Dentipellis was typified by D. fragilis. This genus was introduced to accommodate the species characterized by an annual growth habit, hydnoid basidiomata, soft spines, a monomitic hyphal structure with clamp connections and cyanophilous hyphae, and amyloid rough basidiospores [ 30, 159] . Zhou & Dai [ 30] demonstrated that Dentipellis was polyphyletic and segregated D. leptodon (Mont.) Maas Geest. and D. taiwaniana Sheng H. Wu from Dentipellis based on ITS and nLSU rDNA sequences. Chen et al. [ 11] maintained that Dentipellis remained polyphyletic based on ITS and nLSU rDNA sequences, therefore, a broad concept for Dentipellis s.l. was adopted [ 160] . Recently, based on molecular and morphological analyses, more new taxa were described in Dentipellis sensu lato [ 157, 160, 161] . All Dentipellis spp. were found from the northern hemisphere [ 120, 157, 160, 161] . In the present study, a new species D. yingjiangensis is described based on morphological and molecular systematics analysis inferred from ITS + nLSU data ( Fig. 4).
Dentipellis yingjiangensis Y.L. Deng & C.L. Zhao, sp. nov. Figures 14, 15
Index Fungorum number: IF860767

Figure 14.
Basidiomata and microscopic structures of Dentipellis yingjiangensis (holotype). (a) Basidiomata on the substrate. (b) Characteristics of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Fusiform gloeocystidia. (f) Mushroom-shaped gloeocystidia. (g) Long cylindrical gloeocystidia. (h) Section of hymenium. Scale bars: (a) = 1 cm; (b) = 1 mm, (c) = 5 µm, (d)–(h) = 10 µm.
Figure 15.
Sections of hymenium of Dentipellis yingjiangensis (holotype). (a) Basidiospores, basidia, and basidioles. (b), (c) Fusiform gloeocystidia. (d) Mushroom-shaped gloeocystidia. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Dentipellis yingjiangensis differs from other species by the inseparable, leather basidiomata, cream to yellowish-brown hymenial surface odontioid, hyphal system monomitic; generative hyphae with clamp connections, and ellipsoid basidiospores.
Etymology – referring to the locality (Yingjiang) of the type specimen.
Type – China, Yunnan Province, Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 2 July, 2024, CLZhao 37187 (SWFC).
GenBank accession numbers – CLZhao 37187 ITS: PV441141, LSU: PV441155, mtSSU: PV461164, rpb1: PX441308
Description – Basidiomata annual, resupinate, inseparable, leather, without odor or taste when fresh, up to 7 cm long, 3 cm wide, 150 µm thick. Hymenial surface odontioid, fresh spines soft, white to cream, becoming fragile, cream to yellowish-brown when dry. Sterile margin cottony, buff to clay-buff, up to 1 mm wide. Subiculum very thin, soft corky, buff, up to 1 mm thick.
Hyphal system monomitic; generative hyphae with clamp connections, colorless, thin-walled, smooth, moderately branched, 2.5–4 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Gloeocystidia with three types: (1) cylindrical to fusoid gloeocystidia numerous, colorless, thin-walled, the top overflows and shrinks in a beak-like shape, 31–61 × 5–6.5 µm; (2) mushroom-shaped gloeocystidia, colorless, thin-walled, 17–32 × 2–4 µm; and (3) long cylindrical gloeocystidia, colorless, thin-walled, 82–131.5 × 5–7 µm. Basidia cylindrical, with one oil drops, with 4 sterigmata and a basal clamp connection, 14–31 × 3–5 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores broadly ellipsoid, colorless, thin-walled, smooth, with some oil drops, IKI–, CB–, (3.3–)3.5–4.7(–4.4) × 2.1–2.9(–3.1) µm, L = 3.79 µm, W = 2.51 µm, Q = 1.49–1.53 (n = 30/1).
Notes – Dentipellis yingjiangensis groups in Dentipellis (Hericiaceae) based on the ITS + nLSU data ( Fig. 4). Phylogenetic analysis shows that D. yingjiangensis is sister to D. rhizomorpha, and closely related to D. fragilis, and D. dissita. However, morphologically, D. rhizomorpha can be delimited from D. yingjiangensis by its cottony margin with white rhizomorphs, thin- to thick-walled generative hyphae, present fusoid cystidioles (18–22 × 2.5–3 µm), clavate basidia (20–23 × 3.5–4 µm), and thick-walled, minutely rough, ellipsoid basidiospores (3.5–4.1 × 2.6–3 µm) [ 160] . Dentipellis fragilis differs by having thin, soft, fragile basidiomata, narrow, thin-walled, cystidioles, moniliform at their apices, slender basidia, mostly four-spored but often also with only two sterigmata, larger broadly ellipsoid to subglobose, thick-walled, minutely warted basidiospores (5–5.5 × 4.2–4.9 µm) [ 162] . Dentipellis dissita differs by its effuse basidiomata with buff to pale flesh hymenial surface, pale yellow or concolorous margin with spines, thin to thick-walled generative hyphae with single clamp connections, slenderly clavate to cylindrical basidia (26–35 × 4.6–6 µm), and broadly ellipsoid to subglobose, thick-walled, minutely warted basidiospores [ 162] .
Note 29 Hericium Pers.
Index Fungorum number: IF17740
Type species – Hericium hystrix Pers.
Notes – Hericium coralloides (Scop.) Pers as the type species to establish the genus Hericium. Later, several species of Hericium were discovered and to date, 34 species are accepted [ 163− 166] . Most of Hericium species are reported from America and Eurasia in higher latitude ecosystems. A phylogeny of Hericium species from North America, together with key taxa from Europe, was accomplished using sequence data from ITS, nLSU, tef1- α, and rpb2 sequences [ 166] . In China, five Hericium species have been reported, viz. H. abietis (Weir ex Hubert) K.A. Harrison, H. cirrhatum (Pers.) Nikol., H. coralloides (Scop.) Pers., H. erinaceus (Bull.) Pers., and H. yunnanense S.M. Tang & S.H. Li [ 165, 166] .
Note 30 Laxitextum Lentz
Index Fungorum number: IF17914
Type species – Laxitextum bicolor (Pers.) Lentz
Notes – Laxitextum (Hericiaceae, Russulales, Basidiomycota) is a small genus of stereoid fungi [ 167] . Laxitextum species can be differentiated from other stereoid genera by distinctive characteristics like resupinate to subpileate basidiomata, brown, tomentose, zonate pileus upper surface; white when fresh, smooth hymenial surface; a monomitic type of hyphal system with thin-walled, clamped generative hyphae; numerous, subulate gloeocystidia; and subglobose to ellipsoid, echinulate and amyloid basidiospores [ 42, 167− 169] . Based on Index Fungorum (2025), Cerioporus has nine specific names with only five species accepted worldwide, viz. L. bicolor, L. globisporum Henkel & Ryvarden, L. incrustatum, L. lutescens Hjortstam & Ryvarden, and L. subrubrum [ 42, 167− 170] . In the present study, a new species L. cremeum is introduced from China, characterized by morphological traits and analyzed phylogenetically using multi-gene analyses.
Laxitextum cremeum Y.L. Deng & C.L. Zhao, sp. nov. Figures 16, 17
Index Fungorum number: IF860768

Figure 16.
Basidiomata and microscopic structures of Laxitextum cremeum (holotype). (a) Basidiomata on the substrate. (b) Characteristics of hymenophore. (c) Basidiospores. (d) Basidia. (e) Basidioles. (f) Subcylindrical gloeocystidia. (g) Narrowly subcylindrical gloeocystidia. (h) Bottled gloeocystidia. (i) Piriform gloeocystidia. (g) Section of hymenium. Scale bars: (a) = 1 cm; (b) = 1 mm, (c) = 5 µm, (d)–(j) = 10 µm.
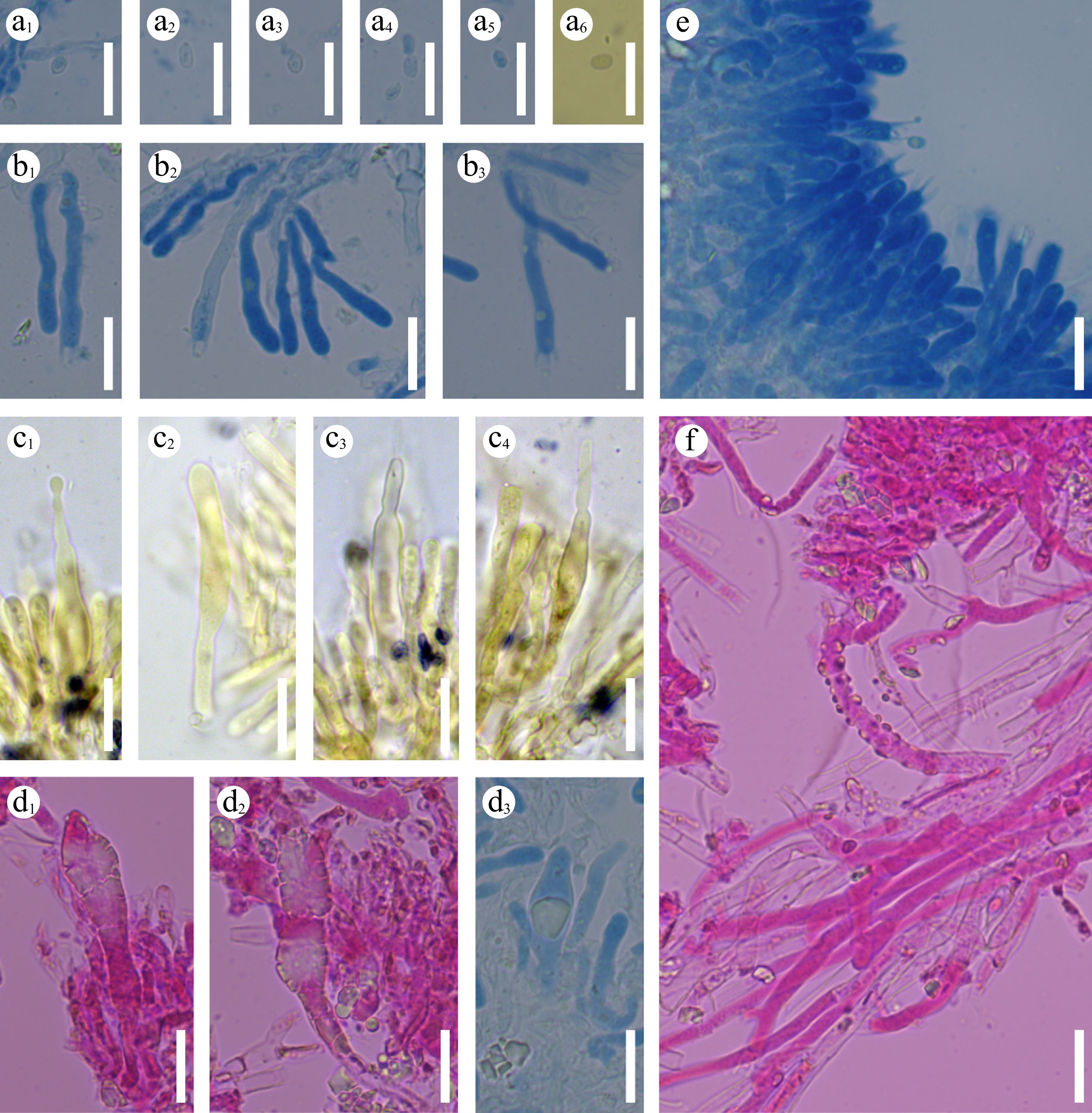
Figure 17.
Sections of hymenium of Laxitextum cremeum (holotype). a 1–a 6 Basidiospores. b 1–b 3 Basidia and basidioles. c 1–c 4 subcylindrical gloeocystidia. d 1–d 3 Pyriform gloeocystidia. (e) Section of hymenium. (f) Generative hyphae. Scale bars: (a)–(f) = 10 µm.
Diagnosis – Laxitextum cremeum differs from other speies by the coriaceous basidiomata with white to cream hymenial surface, a hyphal system monomitic with generative hyphae bearing simple septa, and apiculate ellipsoid basidiospores measuring 2.5–3.3 × 1.8–2.4 µm.
Etymology – referring to the cream color of the hymenial surface.
Type – China, Yunnan Province, Zhaotong, Wumengshan National Nature Reserve, 27°33′ N, 103°27′ E, 1,910 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 21 Septemper, 2023, CLZhao 33747 (SWFC).
GenBank accession numbers – CLZhao 33747 ITS: PV147169, LSU: PV185855, mtSSU: PV399820, rpb2: PV339601, rpb1: PX441309, tef1- α: PV400174
Description – Basidiomata annual, resupinate, closely adnate, sessile, coriaceous, soft-spongy when fresh and brittle on drying, without odor or taste when fresh, up to 1.5 cm long, 1 cm wide, 0.1 mm thick. Hymenial surface smooth, thin, white to cream when fresh, cream upon drying. Sterile margin white to cream, thinning out, up to 2 mm.
Hyphal system monomitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, moderately branched, 2–2.5 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Subhymenial hyphae densely covered by crystals. Gloeocystidia with four types: (1) subcylindrical, the tip contracts to form a long beak, colorless, thin-walled, smooth, filled with refractive oil-like matter, 23–36.5 × 3.5–5 µm; (2) narrowly subcylindrical, colorless, thin -walled, smooth, 27–39 × 4–5 µm; (3) bottled, filled with refractive matter, colorless, thin-walled, smooth, 12–28 × 5–9.5 µm; and (4) pyriform, consisting of a crystal overlay, occasionally the tip contracts to globular, colorless, thin-walled, smooth, 30–45.5 × 7–8 µm. Basidia narrowly subcylindrical, with four sterigmata and a basal simple septum, filled with oil-like matter, 20–28 × 3–3.5 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores ellipsoid, colorless, thin-walled, finely echinulate, CB–, strongly amyloid in Melzer’s reagent, (2.4–)2.5–3.3(–3.4) × (1.7–)1.8–2.4(–2.6) µm, L = 2.94 µm, W = 2.04 µm, Q = 1.45 (n = 30/1).
Notes – The phylogram based on combined ITS+nLSU data ( Fig. 4) revealed that Laxitextum cremeum, is clustered within Laxitextum (Hericiaceae), forming a clade with L. subrubrum, L. bicolor and L. incrustatum. Morphologically, L. subrubrum can be delimited from L. cremeum by having pileate, sessile, soft-spongy, brittle basidiomata, thin- to thick-walled generative hyphae with clamp connections, larger pale orange, lanceolate gloeocystidia with apex moniliform (102–156 × 8.5–15 µm), both larger narrowly clavate basidia with 4-sterigmate and a basal clamp connection (23.5–38 × 5.5–9 µm), and ellipsoid, apiculate basidiospores (4.7–5.9 × 3.1–3.5 µm) [ 42] . Laxitextum bicolor differs from L. cremeum by its resupinate or subpileately reflexed, upper side brown basidiomata, thin to thick-walled light brown generative hyphae with clamp connections, larger fusiform, subulate cystidia with a moniliform apical appendix (40–100 × 5–10 µm), wider narrowly clavate basidia with 4-sterigmate and a basal clamp connection (20–30 × 3.5–5 µm), and longer oblong-ellipsoid, finely echinulate basidiospores (4.5–5 × 2.5 µm) [ 168] . Laxitextum incrustatum can be distinguished from L. cremeum by having effuse basidiomata, white to pallid, cottony to matted tomentose margin, thin- to thick-walled, pale yellow generative hyphae with clamp connections, larger tubular or with a bulbous swelling gloeocystidia with moniliform at the apex (45–170 × 5–10 µm), both larger slenderly clavate basidia with four sterigmata (28–40 × 5.0–6.0 µm), and slightly thick-walled, broadly ellipsoid to subglobose, verrucose basidiospores (4.6–5.2 × 3.4–3.8 µm) [ 168] .
Note 31 Pseudowrightoporia Y.C. Dai, Jia J. Chen & B.K. Cui
Index Fungorum number: IF812226
Type species – Pseudowrightoporia cylindrospora (Ryvarden) Y.C. Dai, Jia J. Chen & B.K. Cui
Notes – Pseudowrightoporia was proposed to accommodate five species, P. africana (I. Johans. & Ryvarden) Y.C. Dai, Jia J. Chen & B.K. Cui, P. aurantipora (T. Hatt.) Y.C. Dai, Jia J. Chen & B.K. Cui, P. gillesii (A. David & Rajchenb.) Y.C. Dai, Jia J. Chen & B.K. Cui, P. solomonensis (Corner) Y.C. Dai, Jia J. Chen & B.K. Cui, and P. straminea (T. Hatt.) Y.C. Dai, Jia J. Chen & B.K. Cui, which were segregated from Wrightoporia [ 10] . Pseudowrightoporia is characterized by soft corky to corky basidiomata, usually with shining pores, corky to fibrous-corky tubes, a dimitic hyphal structure, oblong, broadly ellipsoid, ellipsoid to subglobose, finely asperulate and amyloid basidiospores, and a subtropical to tropical distribution [ 10] . Several new species and new combinations have been proposed, P. crassihypha Y.C. Dai, Jia J. Chen & B.K. Cui, P. cylindrospora, P. hamata Y.C. Dai, Jia J. Chen & B.K. Cui, P. japonica (Núñez & Ryvarden) Y.C. Dai, Jia J. Chen & B.K. Cui, P. oblongispora Y.C. Dai, Jia J. Chen & B.K. Cui. Subsequently, P. dominicana Angelini, Losi & Vizzini was described from the Dominican Republic based on morphological and molecular data (ITS and nLSU sequence analyses) [ 31] .
Note 32 Wrightoporiopsis Y.C. Dai, Jia J. Chen & B.K. Cui
Index Fungorum number: IF812239
Type species – Wrightoporiopsis neotropica (Ryvarden) Y.C. Dai, Jia J. Chen & B.K. Cui
Notes – Wrightoporiopsis was established by Chen et al. [ 10] and was typified by W. neotropica. Wrightoporiopsisis is characterized by pileate, yellow to yellowish-brown basidiomata, a dimitic hyphal system with generative hyphae bearing clamp connections, skeletal hyphae usually dextrinoid, basidiospores ellipsoid to subglobose, hyaline, finely asperulate, strongly amyloid, and causing a white rot [ 10] . Some taxa were previously treated under Wrightoporia Pouzar. [ 10, 44, 171] . Later, five species, Wrightoporiopsis amylohypha Y.C. Dai, Jia J. Chen & B.K. Cui, W. biennis (Jia J. Chen & B.K. Cui) Y.C. Dai, Jia J. Chen & B.K. Cui, W. irregularis Y.C. Dai, Qian Chen & X.H. Ji, W. neotropica and W. roseocontexta (Ryvarden & Iturr.) Y.C. Dai, Jia J. Chen & B.K. Cui, were accepted in the genus [ 10] . Phylogenetic analysis demonstrated that Wrightoporiopsis was distant from Wrightoporia sensu stricto, and these two genera in fact belong to two families, Hericiaceae and Wrightoporiaceae, respectively [ 10] .
Note 33 Family Peniophoraceae Lotsy 1907
Index Fungorum number: IF81123
Type genus – Peniophora Cooke
Notes – Peniophoraceae (Russulales) was established by Lotsy, with the type genus Peniophora. Peniophoraceae is characterized by basidiomata annual, resupinate, orbicular, discoid, effused-reflexed, to clavarioid. Hymenial surfaces smooth, grandinioid, tuberculate or raduloid. Hyphal system monomitic to dimitic, generative hyphae with or without clamp connections, IKI–. Skeletal hyphae or dichohyphae when present dextrinoid or inamyloid binding hyphae as 'bovista-type', dextrinoid, asterosetae present. Gloeoplerous hyphae and gloeocystidia present or absent, lamprocystidia present or absent, smooth, asperulate, spinulose, verrucose, hyaline to pigmented, IKI– basidiospores, or IKI+ suprahilar plage or IKI+ basidiospores [ 1, 172] . Leal-Dutra et al. [ 84] demonstrated the inclusion of Parapterulicium and Baltazaria in family Peniophoraceae based on morphological and molecular characters. It is a large and rather heterogeneous family with seventeen genera, viz. Amylofungus Sheng H. Wu , Asterostroma, Baltazaria, Dendrophora, Dichostereum, Duportella Pat., Entomocorticium, Gloiothele Bres., Lachnocladium, Licrostroma, Metulodontia Parmasto, Parapterulicium, Peniophora, Sceptrulum, Scytinostroma, Vararia, and Vesiculomyces E. Hagstr. currently accepted [ 1, 173] . Peniophora and Vararia, have the highest number of taxa in this family, in which they play fundamental ecological roles to drive carbon cycling in forest soils, acting as decomposers [ 25, 174] .
Note 34 Amylofungus Sheng H. Wu
Index Fungorum number: IF27556
Type species – Amylofungus corrosus (G. Cunn.) Sheng H. Wu
Notes – Amylofungus was established by Sheng H. Wu [ 175] , and typed by A. corrosus (= Corticium corrosum G. Cunn.). It is characterized by resupinate, effuse basidiomata, smooth hymenial surface, generative hyphae simple-septate, amyloid, numerous, clavate or cylindrical, amyloid gloeocystidia, utriform basidia, four sterigmata, amyloid, globose or subglobose, smooth, thin-walled, amyloid basidiospores [ 175] . Another species A. globosporus (Maekawa) Sheng H. Wu was transferred from Vesiculomyces globosporus N. Maek., it has characters typical of the genus Amylofungus Sheng H. Wu, i.e., amyloid reaction for all elements in the basidioma: hyphae, gloeocystidia, basidia, and basidiospores. Simple-septate hyphae, utriform basidia, and smooth basidiospores are also important characters of the species in this genus [ 175, 176] .
Note 35 Asterostroma Massee
Index Fungorum number: IF17113
Type species – Asterostroma apalum (Berk. & Broome) Massee
Notes – The corticioid genus Asterostroma, belonging to order Russulales, was established by Massee with A. apalum as its type species. The genus is characterized by resupinate and felted-membranous basidiomata, gloeocystidia, and dextrinoid astrosetae [ 177] , and based on the asterosetae, Asterostroma was placed in family Lachnocladiaceae [ 178] . Asterostroma belongs to Peniophoraceae of russuloid lineage in recent phylogenetic analysis of 5.8S + nLSU sequence data [ 28, 45, 53, 179] . According to the morphological characters of the basidiospores in Asterostroma [ 180] , this genus can be divided into two subgenera Austroasterostroma Parmasto (smooth and amyloid basidiospores) and Asterostroma (ornamented and amyloid basidiospores) [ 23, 34] . In recent years, three clades were correlated with two sections and a monotypic lineage: the sect. Asterostroma clade, which includes A. bambusicola, A. cervicolor (Berk. & M.A. Curtis) Massee, A. macrosporum N. Maek. & Suhara, A. medium Bres., A. muscicola (Berk. & M.A. Curtis) Massee, and A. ochroleucum Bres. ex Torrend; sect. Laevispora Parmasto contains A. vararioides S.L. Liu & S.H. He and A. laxum Bres.; and a monotypic lineage consisting of A. andinum Pat. [ 53, 179, 181] . In the present study, based on morphological traits ( Figs 18, 19) and analyzed phylogeny using multiple gene regions, a new species Asterostroma fimbriatum is described.
Asterostroma fimbriatum Y.L. Deng & C.L. Zhao, sp. nov. Figures 18, 19
Index Fungorum number: IF860769
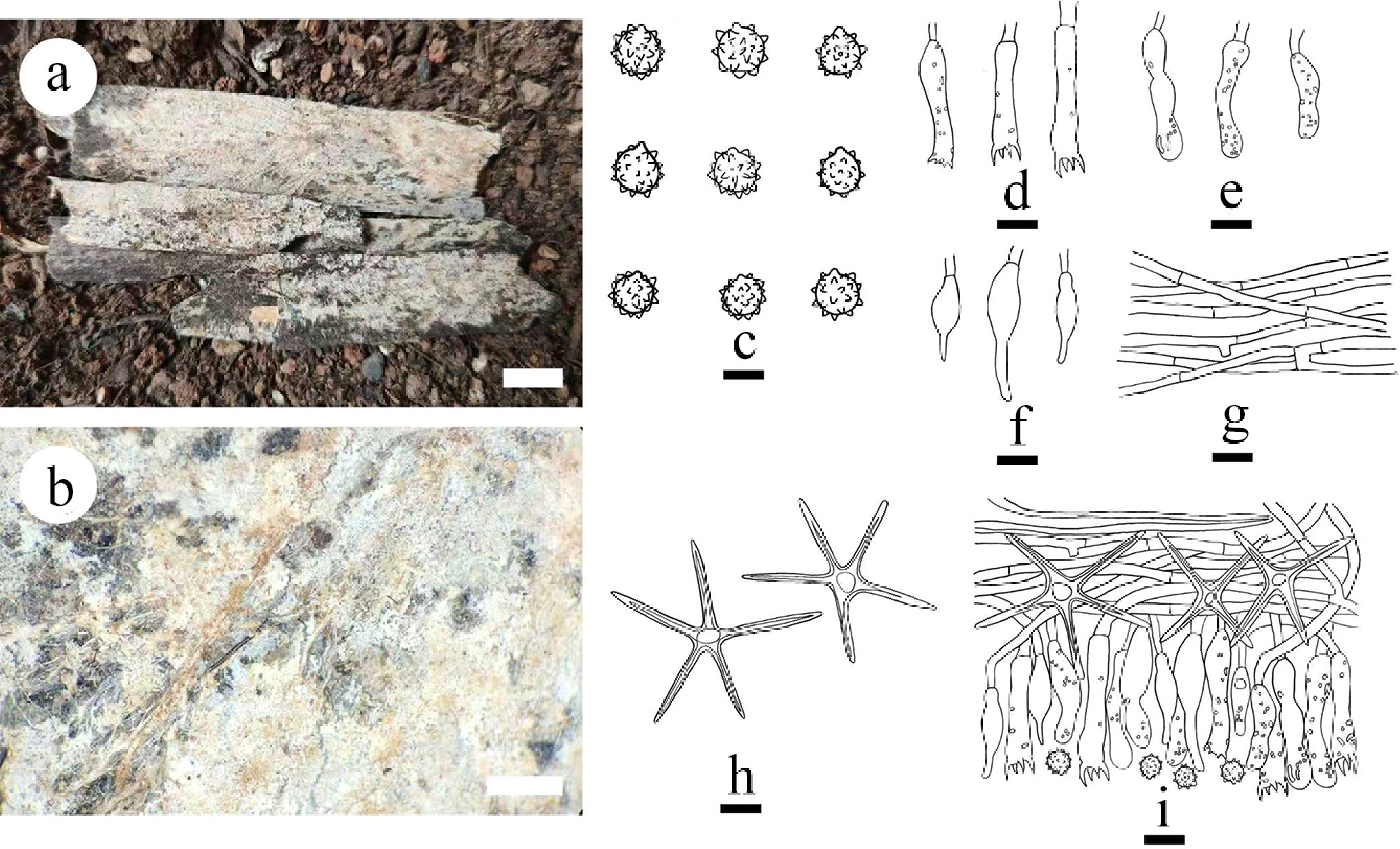
Figure 18.
Basidiomata and microscopic structures of Asterostroma fimbriatum (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia. (e) Basidioles. (f) Gloeocystidia. (g) Hyphae from context. (h) Asterosetae. (i) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 µm, (d)–(i) = 10 µm.
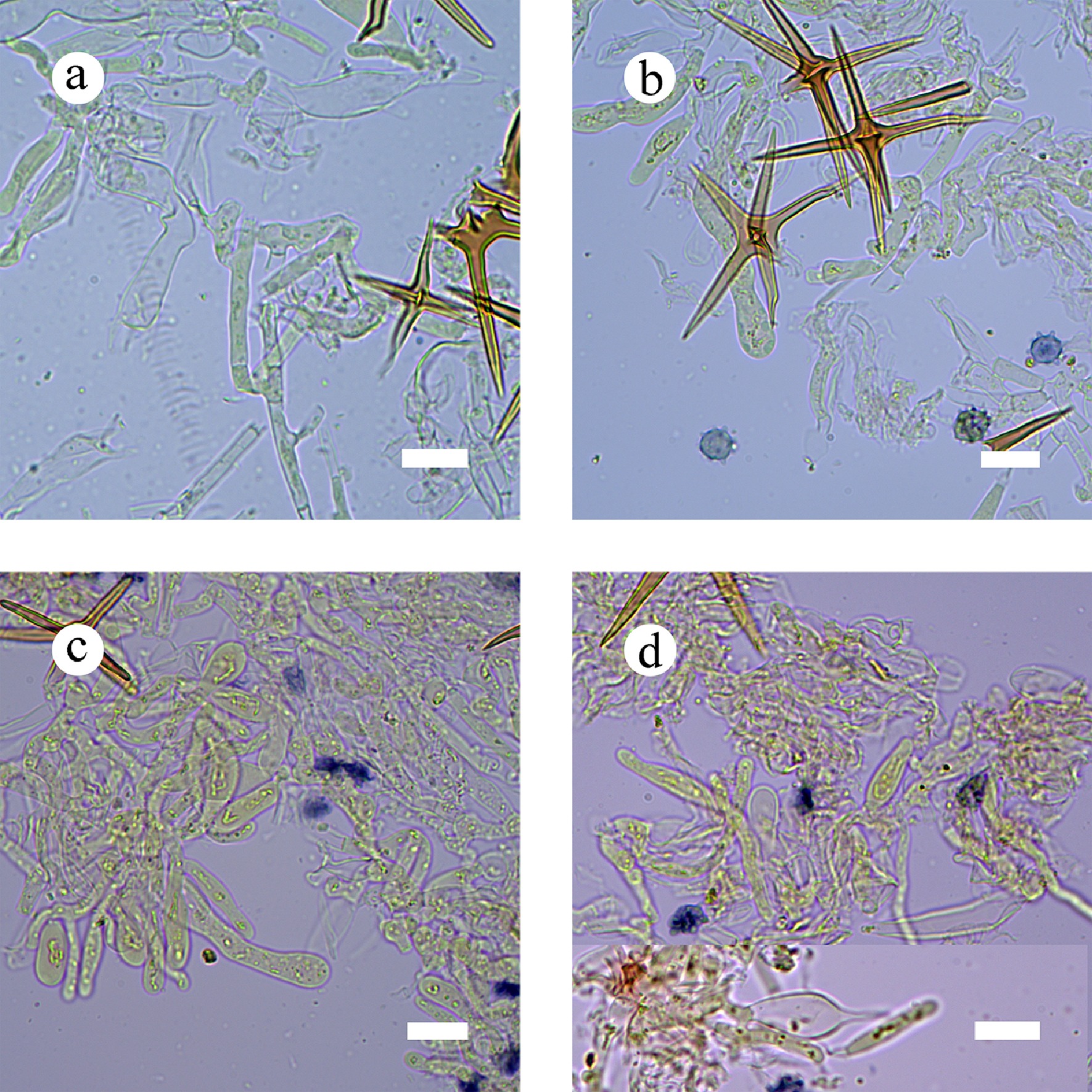
Figure 19.
Sections of hymenium of Asterostroma fimbriatum (holotype). (a) Basidia. (b) Basidiospores. (c) Basidioles. (d) Gloeocystidia. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Asterostroma fimbriatum differs from other species by the felted-membranous to pellicular basidiomata, cream to slightly buff hymenial surface, a dimitic hyphal system with generative hyphae bearing simple septa, and subglobose to globose echinulate basidiospores.
Etymology – referring to fimbriate sterile margin of the basidiomata.
Type – China, Yunnan Province, Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on dead bamboo, leg. C.L. Zhao, 21 November, 2024, CLZhao 41500 (SWFC).
GenBank accession numbers – CLZhao 41500 ITS: PV940929, LSU: PX070093, mtSSU: PX243671, rpb2: PX432789
Description – Basidiomata annual, resupinate, rhizomorphic, without odor and taste when fresh, up to 9 cm long, 2 cm wide, and 90 µm thick. Hymenial surface smooth, cream when fresh, cream to buff to slightly brown upon drying. Sterile margin thinning out, fimbriate, cream, up to 1 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, moderately branched, colorless, thin-walled, 3–4 µm in diameter; IKI–, CB–; tissues unchanged in KOH. Asterosetae in subiculum abundant, predominant, yellowish brown, thick-walled, regularly star-shaped, 3–9 µm in diameter, dextrinoid, rays up to 76 µm long, with acute tips, CB–; IKI–, tissues unchanged in KOH. Asterohyphidia in hymenium similar to asterosetae in the subiculum, but smaller and less regularly shaped, 3–5 µm in diameter, rays up to 40 µm long. Gloeocystidia subulate to fusiform, thin-walled, with a basal simple septum, 15.6–24.7 × 4.6–8.2 µm; cystidioles absent. Basidia subcylindrical, colorless, with four sterigmata and a basal simple septum, 16–35 × 4.7–7.6 µm; basidioles dominant, similar to basidia in shape, but slightly smaller. Basidiospores subglobose to globose, with a distinct apiculus, spines conical, colorless, thin-walled, echinulate, with one oil drop, IKI+, CB–, (4.7–)4.9–6(–8) × (4.5–)4.8–6(–7.8) µm, L = 5.55 µm, W = 5.34 µm, Q = 1.04 (n = 30/1).
Notes – Based on ITS + nLSU sequence data ( 5), Asterostroma fimbriatum is grouped within Asterostroma (Peniophoraceae), and it is closely related to A. bambusicola, A. cervicolor, A. macrosporum, A. muscicola, and A. laxum. However, morphologically, A. bambusicola can be delimited from A. fimbriatum by its membranaceous to pellicular, soft basidiomata, brownish yellow, greyish brown, light brown to brownish red hymenophore, larger subulate gloeocystidia (40–70 × 6–11 µm) [ 179] . Asterostroma cervicolor differs from A. fimbriatum by having larger gloeocystidia (40–50 × 4–6 µm), and basidia (30–40 × 4.5–5.5 µm) [ 180] . Asterostroma macrosporum differs from A. fimbriatum by its effused, soft, felt-like basidiomata, buff, ochreous to fulvous hymenial surface, thin- to slightly thick-walled generative hyphae, both larger subcylindrical, ventricose to subfusiform gloeocystidia (40–80 × 10–17 µm), and subcylindrical to utriform basidia (38–65 × 8–9.5 µm), globose basidiospores (8.5–11 × 7.5–9 µm) [ 181] . Asterostroma muscicola can be distinguished from A. fimbriatum by having larger gloeocystidia (20–70 × 8–12 µm), utriform basidia with 2-4 sterigmata (18–24 × 5–6 µm) [ 177] . Asterostroma laxum differs from A. fimbriatum by its effused, membranaceous basidiomata, cream to ochraceous hymenophore, larger irregularly cylindrical gloeocystidia (40–100 × 6–10 µm), utriform basidia with 4-sterigmata, and a simple basal septum (40–80 × 6–8 µm), and smooth, subglobose basidiospores [ 177] .
Note 36 Baltazaria Leal-Dutra, Dentinger & G.W. Grif
Index Fungorum number: IF825233
Type species – Baltazaria galactina (Fr.) Leal-Dutra, Dentinger & G.W. Griff.
Notes – Baltazaria was proposed by Leal-Dutra et al. [ 84] , and typified by B. galactina, to accommodate B. octopodites (Corner) Leal-Dutra, Dentinger & G.W. Griff. (= Parapterulicium octopodites Corner), B. galactina (= Scytinostroma galactinum (Fr.) Donk), B. neogalactina (Boidin & Lanq.) Leal-Dutra, Dentinger & G.W. Griff., (= S. neogalactinum Boidin & Lanq.) and B. eurasiaticogalactina (Boidin & Lanq.) Leal-Dutra, Dentinger & G.W. Griff. (= S. eurasiaticogalactinum Boidin & Lanq.) within Peniophorales [ 84] . This genus is characterized by corticioid, adherent to effused, coriaceous, or membranaceous to hard basidiomata, white, cream or pale ochraceous hymenial surface. Context densely homogeneous with thick-walled and dextrinoid skeletal-binding hyphae, sometimes bearing rows of short papillae or skeletodendrohyphidia [ 84] . In the present study, based on the characterization of morphological traits ( Fig. 20) and analyzed phylogeny using multiple gene analyses ( Figs 1, 5), the new species Baltazaria pingbianensis is described.
Baltazaria pingbianensis Y.L.Deng & C.L. Zhao, sp. nov.
Index Fungorum number: IF856165, Fig. 20

Figure 20.
Basidiomata and microscopic structures of Baltazaria pingbianensis (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidioles. (e) Basidia. (f) Skeletal hyphae. (g) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 µm, (d)–(g) = 10 µm.
Diagnosis – Differs from other species by its membranous basidiomata, a dimitic hyphal system with generative hyphae bearing simple septa, fibriform skeletal hyphae, cylindrical basidia, and ovoid basidiospores.
Etymology – referring to the locality (Pingbian) of the type specimen.
Type – China, Yunnan Province, Honghe, Pingbian County, Daweishan National Nature Reserve, 22°51' N, 103°41' E, 2,360 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 1 August 2019, CLZhao 17755 (SWFC).
Description – Basidiomata annual, resupinate, membranous, soft, and adnate, up to 130 mm long, 20 mm wide, and 90 µm thick. Hymenial surface smooth, white when fresh, white to slightly cream upon drying. Sterile margin narrow, whitish, up to 1 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, rarely branched, thin-walled, 2–2.5 μm in diameter, CB–; tissues unchanged in KOH. Skeletal hyphae fibriform, white to yellowish, moderately dextrinoid, thick-walled, 1–2 μm in diameter. Cystidia and cystidioles absent. Basidia cylindrical, colorless, thin-walled, with four sterigmata and a simple septum at base, 18–28 × 3–5 μm, basidioles dominant, in shape similar to basidia, but slightly smaller, with a refractive oil-like matter. Basidiospores oblong ellipsoid, with a beaklike extension, thin-walled, colorless, smooth, IKI–, CB–, 3.5–5 × 2–3 µm, L = 4.38 µm, W = 2.52 µm, Q =1.83–2.11 (n = 60/3).
Additional Material Examined – China, Yunnan Province, Honghe, Pingbian County, Daweishan National Nature Reserve, 22°51' N, 103°41' E, 2,360 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 3 August, 2019, CLZhao 18294 and CLZhao 18296 (SWFC).
GenBank accession numbers – CLZhao 17755 ITS: OR048814, LSU: OR510674, mtSSU: OR539677, tef1- α: PQ726901; CLZhao 18294 ITS: OR048816; CLZhao 18296 ITS: OR048815
Notes – The phylogenetic analysis based on ITS + nLSU sequence data ( Figs 1, 5) shows that Baltazaria pingbianensis is closely related to B. galactina and B. neogalactina within family Peniophoraceae. However, morphologically, B. galactina differs from B. pingbianensis by its resupinate, effused, membranaceous basidiomata, cream to isabelline or pale yellowish hymenophore, generative hyphae with clamps, hyaline to yellowish, thick-walled skeletal-binding hyphae, cylindrical gloeocystidia (30–120 × 2–5.5 µm), clavate to subcylindrical basidia with four sterigmata and a basal clamp, narrowly ellipsoid to subcylindrical basidiospores [ 84] . Baltazaria neogalactina can be distinguished from B. pingbianensis by having beige to beige ombré, isabelle, cannelle hymenophore, and larger ellipsoid basidiospores (4.5–6 × 3–3.5 µm) [ 182]
Note 37 Dendrophora (Parmasto) Chamuris
Index Fungorum number: IF25097
Type species – Dendrophora versiformis (Berk. & M.A. Curtis) Chamuris
Notes – Dendrophora was initially typified by D. versiformis, but the species was transferred to Peniophora. The genus is characterized by resupinate, effuse-reflexed or rarely sessile basidiomata, smooth, finely pruinose or velutinous, grey to brown, hymenial surface brown to black, tomentose to felty hymenophore, hyphal system monomitic (or pseudodimitic), subhyaline to brown, thin- to thick-walled generative hyphae with clamp connections, heavily encrusted cystidia (lamprocystidia), subhyaline to brown dendrohyphidia, narrowly clavate basidia with four sterigmata, and a basal clamp, and smooth, thin-walled, cylindrical to allantoid basidiospores [ 183] . Dendrophora species share similar characters with the species of Peniophora. Dendrophora mainly differs in the brown dendrohyphidia. It is phylogenetically classified in family Peniophoraceae [ 45] . Only two species are accepted, D. albobadia (Schwein.) Chamuris, and D. erumpens (Burt) Chamuris.
Notes 38 Dichostereum Pilát
Index Fungorum number: IF17504
Type species – Dichostereum durum (Bourdot & Galzin) Pilát
Notes – Dichostereum, typified with D. durum, is a small and well-delimited corticioid genus in Russulales. It is characterized by resupinate basidiomata with smooth or grandinioid hymenophore, dimitic hyphal system with dextrinoid dichohyphae and clamped generative hyphae, gloeocystidia and ellipsoid or subglobose, ornamented basidiospores with a strong amyloid reaction in Melzer’s reagent [ 45] . Previously, Dichostereum was placed in Lachnocladiaceae, which included genera with dextrinoid skeletal hyphae [ 184] . However, recent DNA-based phylogenetic analyses, showed that Dichostereum formed a monophyletic lineage within family Peniophoraceae, which included genera with or without dextrinoid hyphae [ 26, 28, 45] . Dichostereum was once treated as a subgenus of Vararia (Peniophoraceae, Russulales) in some studies since the two genera are very similar in morphology except that the latter has smooth basidiospores [ 185, 186] . Boidin & Lanquetin [ 187] emended the description of Dichostereum, and retained it as a separate genus. Later, Boidin & Lanquetin [ 188] monographed the genus and provided a key to its 11 species based on evidence of morphology, distribution, and inter compatibility tests of cultures. Based on limited taxon sampling, their results showed that D. effuscatum (Cooke & Ellis) Boidin & Lanq. and D. granulosum (Pers.) Boidin & Lanq. were widely distributed, while the other species seemed to be rather endemic [ 188] . Few studies on the genus have been carried out since then and many regions including East Asia need further collecting and studies [ 182] . Previously, five species, D. austrosinense S.H. He & S.L. Liu, D. boidinii S.H. He & S.L. Liu, D. boreale (Pouzar) Ginns & M.N.L. Lefebvre, D. eburneum S.H. He & S.L. Liu, and D. pallescens (Schwein.) Boidin & Lanq. were reported in temperate China [ 43, 173] . According to Index Fungorum (2025), and Mycobank (2025), Dichostereum has 18 specific and infraspecific names, and the number of accepted species is 15.
Note 39 Duportella Pat.
Index Fungorum number: IF17533
Type species – Duportella velutina Pat.
Notes – Duportella (Russulales, Basidiomycota) is a small genus of corticioid fungi currently including 20 records of intraspecific names. However, only eight species, D. kuehneri (Boidin & Lanq.) Hjortstam, D. kuehneroides Boidin, Lanq. & Gilles, D. lassa Spirin & Kout, D. raimundoi Pat., D. renispora Boidin, Lanq. & Gilles, D. schomburgkii (Berk.) G. Cunn., D. tristicula (Berk. & Broome) Reinking, and D. velutina have been accepted worldwide. The species mostly have a tropical distribution. Many species of Duportella have been transferred to Peniophora [ 189− 191] . This genus is characterized by resupinate, effused to effuse-reflexed, adnate basidiomata, greyish, reddish, violaceous, ochraceous or blackish hymenophore, monomitic or dimitic hyphal system, hyphae usually with clamps or some simple-septa, brown, apically encrusted pseudocystidia and gloeocystidia usually SA+, cylindrical to subclavate basidia with four sterigmata, and with a basal clamp, ellipsoid to ovoid or globose, cylindrical to allantoid, smooth, thin-walled basidiospores [ 192] . Duportella is characterized among Peniophora s.l. by the brown encrusted pseudocystidia and by presence of skeletal or skeletoid hyphae. Duportella is closely related to Peniophora and Dendrophora and it is classified in family Peniophoraceae [ 45, 191] .
Note 40 Entomocorticium H.S. Whitney, Bandoni & Oberw.
Index Fungorum number: IF23076
Type species – Entomocorticium dendroctoni H.S. Whitney 1987
Notes – Entomocorticium is comprised of 12 species, all associated with Scolytinae beetles [ 193− 195] . Entomocorticium dendroctoni, the type species of the genus, was described based exclusively on morphological characters to accommodate a cryptic species that was observed growing intermingled with a blue stain fungus [ 193] . Hsiau & Harrington [ 196] were the first to show that Entomocorticium was a diverse fungal lineage associated solely with a group of phloem-inhabiting bark beetles that feed heavily on fungi. In addition to E. dendroctoni, they identified nine putative species based on their mt-SSU, ITS, and IGS-1 phylogenetic analyses. They also suggested that the Entomocorticium clade was relatively young, likely having recently diversified, and remained uncharacterized until the early 2000s.
Note 41 Gloiothele Bres.
Index Fungorum number: IF17678
Type species – Gloiothele lamellosa (Henn.) Bres.
Notes – Gloiothele was introduced by Bresadola [ 197] and it was typified by G. lamellosa, which deviates from other members of the genus by its coarsely hydnoid-tuberculate hymenophore [ 198] . It comprises resupinate basidiomata with simple septate, generative hyphae, true gloeocystidia generated from gloeoplerous hyphae, and smooth, thin-walled, amyloid basidiospores. It belongs to order Russulales and is most closely related to Asterostroma, Scytinostroma, and Vesiculomyce [ 26] . Vesiculomyces is separated form Gloiothele by the absence of sulfovanillin reaction in gloeocystidia [ 26, 199] . According to Index Fungorum (2025, www.indexfungorum.org), Gloiothele has 18 specific names, with 14 species accepted worldwide [ 20, 26, 198− 201] . Two species were reported from Amazon Rainforest, G. incrustata Gorjón and G. lactescens growing on a frondose trees [ 202] . Gloiothele torrendii (Bres.) Boidin & H. Michel was originally described as Corticium torrendii Bres., and grows exclusively on olive trees [ 198, 201] . From China, G. yunnanensis Yang Yang & C.L. Zhao was reported on the fallen branch of angiosperms [ 20] . In the present study, based on morphological traits and phylogeny using multi-gene sequences, two new species G. fragilima and G. tuberculata are described.
Gloiothele fragilima Y.L. Deng & C.L. Zhao, sp. nov. Figures 21, 22
Index Fungorum number: IF860770

Figure 21.
Basidiomata and microscopic structures of Gloiothele fragilima (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidioles. (e) Basidia. (f) Gloeocystidia. (g) Section of hymenium. Scale bars: (a) = 1 cm; (b) = 1 mm, (c) = 5 µm, (d)–(g) = 10 µm.
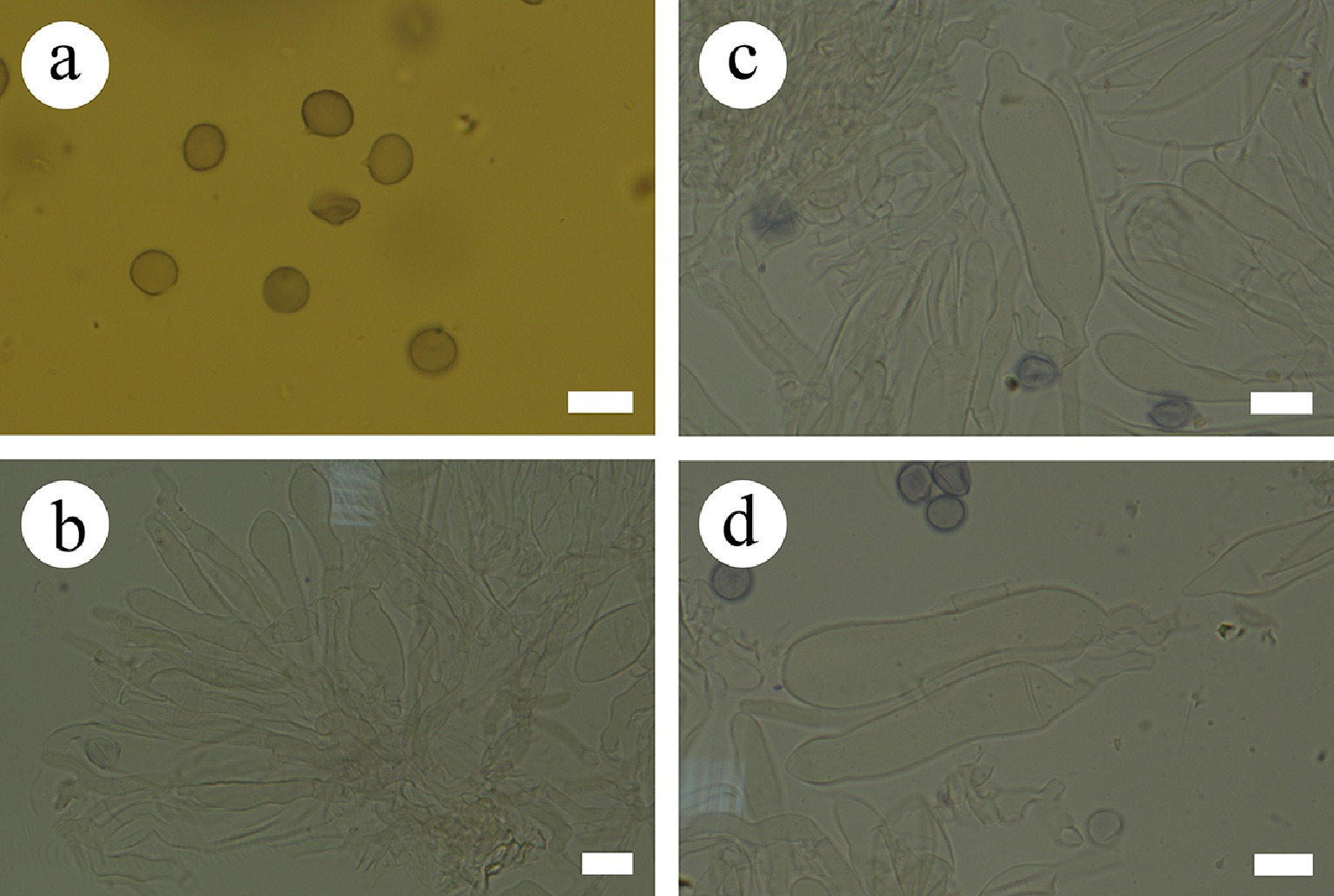
Figure 22.
Sections of hymenium of Gloiothele fragilima (holotype). (a) Basidiospores. (b) Section of hymenium. (c), (d) Gloeocystidia. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Gloiothele fragilima differs from other species by the cream to slightly yellowish hymenial surface, a monomitic hyphal system with simple septa generative hyphae, and globose basidiospores measuring 6.6–8.3 × 6.2–7.7 µm.
Etymology – referring to fragile basidiomata of the species.
Type – China, Yunnan Province, Wenshan, Bozhu Town, Wenshan National Nature Reserve, 23°36′ N, 104°25′ E; 1,500 m asl, on trunk of Camellia, leg. C.L. Zhao, 23 July, 2019, CLZhao 15980 (SWFC).
GenBank accession numbers – CLZhao 15980 ITS: PV147168, mtSSSU: PV399819
Description – Basidiomata annual, membranous, adnate, up to 6 cm long, 2 cm wide, and 100 µm thick. Hymenial surface smooth, cream to slightly yellowish when fresh, slightly yellowish upon drying. Sterile margin distinct, narrow, white, up to 2 mm.
Hyphal system monomitic; generative hyphae bearing simple septa, rarely branched, colorless, thin-walled, 3–5 μm in diameter, IKI–, CB–; tissues unchanged in KOH. Gloeocystidia fusoid to barrelled, colorless, thin-walled, smooth, contract into a mastoid shape at the top, 48.4–69.3 × 11.2–17.2 µm. Basidia subcylindrical, with four sterigmata and a basal simple septum, thin-walled, smooth, 41.1–49.2 × 9–10.5 µm; basidioles in shape similar to basidia, but slightly smaller. Basidiospores globose, thin-walled, colorless, smooth, amyloid, CB–, (6.5–)6.6–8.3(–8.4) × (5.9–)6.2–7.7(–8) µm, L = 7.57 µm, W = 6.98 µm, Q =1.08 (n = 30/1).
Notes – The combined ITS + nLSU phylogenetic analyses ( Fig. 5) revealed that Gloiothele fragilima is grouped within Gloiothele, sister to G. citrina. However, morphologically, G. citrina can be distinguished from G. fragilima by its widely effuse, margin indeterminate basidiomata with yellowish hymenial surface, irregularly cylindrical to vesicular gloeocystidia, wider tubular to narrowly clavate basidia (30–40 × 5–7 µm), and smaller subglobose basidiospores (4.5–7 × 4–6 µm) [ 203] .
Gloiothele tuberculata Y.L. Deng & C.L. Zhao, sp. nov. Figures 23, 24
Index Fungorum number: IF860771

Figure 23.
Basidiomata and microscopic structures of Gloiothele tuberculata (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Barrelled gloeocystidia. (f) Long cylindrical gloeocystidia. (g) Section of hymenium. Scale bars: (a) = 1 cm; (b) = 1 mm, (c) = 5 µm, (d)–(g) = 10 µm.
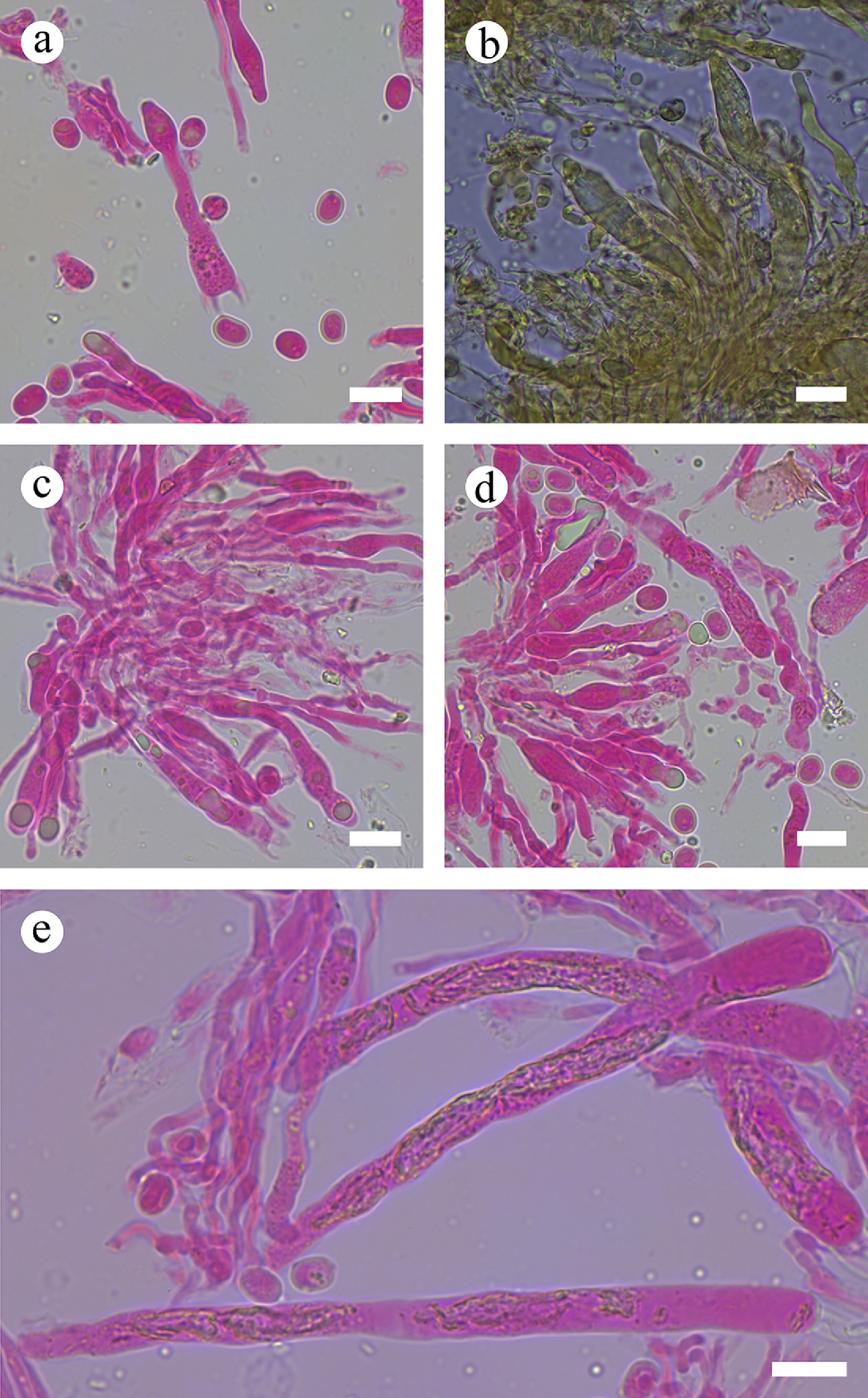
Figure 24.
Sections of hymenium of Gloiothele tuberculata (holotype). (a) Basidiospores and basidia. (b), (e) Gloeocystidia. (c), (d) Section of hymenium. Scale bars: (a)–(e) = 10 µm.
Diagnosis – Gloiothele tuberculata differs from other species by the coriaceous basidiomata with tubercules hymenial surface white to cream, and broadly ellipsoid to globose basidiospores (5.6–8.9 × 4.3–6.8 µm).
Etymology – referring to tuberculous hymenial surface of the type specimens.
Type – China, Yunnan Province, Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 18 July 2023, CLZhao 29919 (SWFC).
Description – Basidiomata annual, adnate, coriaceous, up to 8 cm long, 5 cm wide, and 200 µm thick. Hymenial surface tuberculate, white to cream when fresh, cream upon drying. Sterile margin distinct, white, up to 2 mm.
Hyphal system monomitic; generative hyphae bearing simple septa, rarely branched, colorless, thin-walled, 2–3.5 μm in diameter, IKI–, CB–; tissues unchanged in KOH. Gloeocystidia with two types: (1) barrelled, colorless, thin-walled, smooth, filled with a lot of refracted matter, 29.4–53.3 × 9.2–11.8 µm; and (2) long cylindrical, colorless, thin-walled, smooth, filled with a lot of refracted matter, 75.4–117.5 × 6.1–12.3 µm. Basidia subcylindrical, swollen at the base, with four sterigmata and a basal simple septum, thin-walled, smooth, 34.7–49.6 × 5.2–8.4 µm; basidioles in shape similar to basidia, but slightly smaller. Basidiospores broadly ellipsoid to globose, thin-walled, colorless, smooth, amyloid, CB–, (4.9–)5.6–8.9(–9.5) × (4.1–)4.3–6.8(–7.1) µm, L = 6.85 µm, W = 5.17 µm, Q =1.26–1.38 (n = 150/6).
Additional material examined – China, Yunnan Province, Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 18 July, 2023, CLZhao 29888, CLZhao 29895, CLZhao 29986, and CLZhao 30056 (SWFC).
GenBank accession numbers – CLZhao 29919 ITS: PV441136, LSU: PV441151, MTSSU: PV461161, rpb2: PV648922, tef1- α: PV648916; CLZhao 29888 ITS: PV441135, LSU: PV441150, MTSSU: PV461160; CLZhao 29895 ITS: PV441137; CLZhao 29986 ITS: PV441138, LSU: PV441152, MTSSU: PV461162, tef1- α: PV648917; CLZhao 30056 ITS: PV441139, LSU: PV441153, rpb2: PV648923, tef1- α: PV648918
Notes – In the multi-locus phylogeny, Gloiothele tuberculata grouped within Gloiothele, forming a separate lineage sister to G. lamellosa and G. lactescens. However, morphologically, G. lamellosa differs from G. tuberculata by its isabelline basidiomata, both larger irregularly cylindrical gloeocystidia (80–250 × 10–20 µm), and claviformes basidia (40–60 × 6–8 µm) [ 197] . Gloiothele lactescens can be distinguished from G. tuberculata by its smooth yellowish hymenial surface, both longer, tubular, and sinuous gloeocystidia (100–150 × 5–8 µm), and narrowly clavate basidia (45–60 × 6–7 µm) [ 200] .
Note 42 Lachnocladium Lév.
Index Fungorum number: IF17889
Type species – Lachnocladium brasiliense (Lév.) Pat.
Notes – Lachnocladium was typed by L. brasiliense (= Eriocladus brasiliensis Lév.). It is characterized by annual, often caespitose basidiomata usually strongly ramified with rather slender branch; hymenium unilateral on the underside; section shows distinct medulla and paler cortex; pale ochre or yellowish to brown; apices paler and sterile; a dimitic hyphal system; generative hyphae with simple septate, abundant dichophyses; often present gloeoplerous hyphae; always present gloeocystidia, hyaline, thin-walled; small basidia with (2–)4 sterigmata; hyaline basidiospores that are very small, smooth, not amyloid [ 204] . The distinctive characters of Lachnocladium are coriaceous and more or less hairy covering of fructifications; by these characters the genus can be distinguished from Clavaria Vaill. ex L.
Note 43 Parapterulicium Corner
Index Fungorum number: IF18185
Type species – Parapterulicium subarbusculum Corner
Notes – Parapterulicium was introduced to accommodate two Brazilian species, P. simplex Corner, and P. subarbusculum of coralloid fungi with affinities to Pterulaceae (Agaricales). Despite the coralloid habit and the presence of skeletal hyphae, other features such as the presence of gloeocystidia, dichophyses, and papillate hyphal ends differentiate this genus from Pterulaceae sensu stricto. Fieldwork in Brazil resulted in the rediscovery of two coralloid fungi identifiable as Parapterulicium, the first verified collections of this genus since Corner’s original work in the 1950s. Molecular phylogenetic analyses of ITS and nLSU sequences from these modern specimens revealed affinities with order Russulales, rather than Pterulaceae. The presence of distinctive hyphal elements, homologous to the defining features of Peniophoraceae, is consistent with the phylogenetic evidence and thus clearly distinguished Parapterulicium and its type species P. subarbusculum from Pterulaceae, placing this genus within Peniophoraceae (Russulales) [ 84] .
Note 44 Peniophora Cooke
Index Fungorum number: IF18201
Type species – Peniophora quercina (Pers.) Cooke
Notes – Peniophora was introduced in 1879, and typified by P. quercina, which was the first corticioid genus described on the basis of microscopic characteristics [ 205] . It was initially introduced to accommodate Corticium Pers. and Stereum Hill species which have lamprocystidia (metuloids) in the hymenium. It is a cosmopolitan genus with a wide distribution from boreal to tropical areas, causing a white rot on both angiosperms and gymnosperms. The genus is characterized by having membranaceous, ceraceous, or coriaceous basidiomata with a smooth hymenophore, a monomitic hyphal system with simple-septate or nodose-septate generative hyphae, both encrusted cystidia and gloeocystidia, and thin-walled, smooth basidiospores [ 206] . Based on analyses of ITS sequences of Aphyllophorales, Boidin [ 207] found that the species of Peniophora s.l. formed a relatively strongly supported group that they named ‘Peniophorales’. Later, phylogenetic studies showed that Peniophora s.l. belongs to Peniophoraceae, Russulales [ 26, 28, 45] . According to the phylogenetic analyses of ITS + nLSU sequence data, the morphological characters of Peniophora s.l. are used to delimit the three genera, Peniophora s.s., Dendrophora, and Duportella [ 206] .
Note 45 Scytinostroma Donk
Index Fungorum number: IF18530
Type species – Scytinostroma portentosum (Berk. & M.A. Curtis) Donk
Notes – Scytinostroma was established by Donk [ 146] , and typified by S. portentosum. Initially, the sexual morph of Michenera Berk. & M.A. Curtis which was typified by M. artocreas, was described as Licrostroma P.A. Lemke that was typified by L. subgiganteum (Berk.) P.A. Lemke by Lemke [ 101] . Lyman [ 208] had earlier demonstrated that cultures derived from basidiospores of Corticium subgiganteum Berk., the basionym of L. subgiganteum, produced spores he referred to as Michenera. However, he was unable to induce the cultures to form hymenia and basidiospores. Molecular data place L. subgiganteum with Scytinostroma aluta, S. portentosum, the type of Scytinostroma [ 45, 209] and S. caudisporum [ 210] . Based on a morphological study, Stalpers et al. [ 211] transferred two species of Michenera Berk. & M.A. into Scytinostroma and treated the former genus as a synonym of the latter. Thus, Scytinostroma, Michenera, and Licrostroma are synonyms. Li et al. [ 212] conducted phylogenetic analyses of Peniophoraceae based on concatenated ITS1-5.8S-ITS2-nLSU sequence data, with a focus on samples of Scytinostroma s.s. in different localities, and introduced four new species viz., S. beijingense Yue Li, S.L. Liu & S.H. He, S. boidinii Yue Li, S.L. Liu & S.H. He, S. subduriusculum Yue Li, S.L. Liu & S.H. He, and S. subrenisporum Yue Li, S.L. Liu & S.H. He. Recently, Dong et al. [ 52] reported a new species S. daweishanense J.H. Dong & C.L. Zhao from China based on morphological characteristics and phylogenetic analyses inferred from ITS + nLSU sequences. Scytinostroma includes 44 names based on MycoBank (2025) and Index Fungorum (2025).
Note 46 Vararia P. Karst.
Index Fungorum number: IF18724
Type species – Vararia investiens (Schwein.) P. Karst.
Notes – Vararia is a corticioid wood-inhabiting genus with a wide distribution, typified by V. investiens. It was described by Karsten as a subgenus of Xerocarpus P. Karst. for X. alutarius (Berk. & M. A. Curtis) P. Karst., which was later found to be a synonym of Radulum investiens Schwein [ 213] . The genus is characterized by resupinate basidiomata, a dimitic hyphal structure with simple-septate or clamped generative hyphae and often dextrinoid dichohyphae in Melzer’s reagent, presence of gloeocystidia, and variously shaped smooth basidiospores with or without an amyloid reaction [ 214− 216] . The species of Vararia are found on fallen angiosperm branches, dead wood or herbaceous stems or occasionally on gymnosperm wood [ 217] . Based on MycoBank (2025) and Index Fungorum (2025), Vararia has 107 specific and infraspecific names, with up to 83 accepted species, occurring mainly in tropical and subtropical areas of the world [ 33, 218− 221] . However, Vararia is still poorly studied in China, 18 species have been reported [ 33, 218− 222] . In the present study, eight new wood-inhabiting species belonging to order Russulales, namely, Vararia asiana, V. bambusicola, V. bannaensis, V. ferruginosa, V. lacerata, V. membranacea, V. muscicola and V. wumengshanensis are proposed based on a combination of morphoanatomical characteristics and molecular phylogenetic data. Additionally, an identification key to 25 species of Vararia in China is provided.
Vararia asiana Y.L. Deng & C.L. Zhao, sp. nov. Figures 25, 26, 27a
Index Fungorum number: IF859280

Figure 25.
Basidiomata and microscopic structures of Vararia asiana (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. e Gloeocystidia. (f) Dichohyphae. (g) Section of the hymenium. Scale bars: a = 1 cm, b = 1 mm, c = 5µm, d–g = 10 µm.

Figure 26.
Sections of the hymenium of Vararia asiana (holotype). (a) Basidiospores. (b) Basidia. (c) Dichohyphae. (d) Basidioles. (e) Gloeocystidia. (f) Skeletal hyphae. Scale bars: (a)–(f) = 10 µm.
Diagnosis – Vararia asiana differs from other species by its membranous, soft basidiomata with a smooth, cream to slightly yellowish hymenial surface, generative hyphae bearing simple septa, and broadly ellipsoidal basidiospores (2.5–4 × 1.5–2.5 µm).
Etymology – referring to the continent where the species was found.
Type – China, Yunnan Province, Lincang, Yun County, Dumu Village. 24°23′ N, 101°9′ E; 1,960 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 20 October, 2022, CLZhao 25187 (SWFC).
GenBank accession numbers – CLZhao 25187 ITS: OR102488, LSU: OR510680, tef1- α: PV400170.
Description – Basidiomata annual, membranous, adnate, up to 60 mm long, 20 mm wide, and 60 µm thick. Hymenial surface smooth, cream to slightly yellowish when fresh, slightly yellowish to yellowish upon drying. Sterile margin distinct, narrow, white, up to 2 mm.
Hyphal system dimitic; generative hyphae bearing simple septa, rarely branched, colorless, thin-walled, with some crystals, 1.5–2.5 μm in diameter, IKI–, CB–; tissues unchanged in KOH. Dichohyphae predominate, frequently branched, capillary, white to yellowish, thick-walled, 2–2.5 µm in diameter, dichotomously to irregularly branched with main branch and acute tips, moderately dextrinoid in Melzer’s reagent. Hymenium skeletal hyphae fibriform, colorless, thick-walled (0.3–0.5 µm thick), 2.5–3 µm in diameter. Gloeocystidia fusiform to subcylindrical, colorless, thin-walled, smooth, with a narrow or shizophapillate apex, 15–21 × 5–7 µm. Basidia subcylindrical, with four sterigmata and a basal simple septum, thin-walled, smooth, 20–29 × 6–7.5 µm; basidioles in shape similar to basidia, but slightly smaller. Basidiospores broadly ellipsoid, thin-walled, colorless, smooth, occasionally with one or two guttules, IKI–, CB–, 2.5–4 × 1.5–2.5 µm, L = 3.21 µm, W = 2.09 µm, Q =1.54 (n = 30/1).
Notes – The phylogenetic analyses ( Fig. 5) revealed that Vararia asiana clustered within the genus Vararia sister to V. daweishanensis, and closely related to V. rhombospora. However, morphologically, V. daweishanensis differs from V. asiana by its pale yellowish hymenial surface, generative hyphae with clamps, colorless, thin- to thick-walled, and larger allantoid basidiospores (9–13 × 3.5–5 µm) [ 221] . Furthermore, V. rhombospora differs from V. asiana in having both larger basidia (30–38 × 5–7 µm) and basidiospores (15–17 × 5–6.2 µm) [ 187] . Morphologically, V. asiana is similar to V. investiens and V. isabellina Y.L. Deng & C.L. Zhao by having a smooth hymenial surface, and basidia with four sterigmata. However, V. investiens has a yellowish-cream to ochraceous hymenial surface, generative hyphae with clamps, larger fusiform gloeocystidia (40–80 × 4–8 µm), utriform basidia (30–50 × 4–5 µm), and fusoid basidiospores (8–12 × 3–3.5 µm) [ 215] . Vararia isabellina can be distinguished from V. asiana by having cream to isabelline to slightly brown hymenial surface, thin to slightly thick-walled generative hyphae bearing simple-septa, larger spindle to subcylindrical gloeocystidia (38–47 × 8–13 μm), basidia (33–39 × 7–9 μm), and sub-fusiform to navicular basidiospores (9–13 × 5–8 µm) [ 33] . The PHI ( Figs 5, 27a) analysis reveals no recombination in the new species compared to the closely related taxa. Thus, based on morphology and multigene phylogenetic evidence, V. asiana is introduced as a new species.
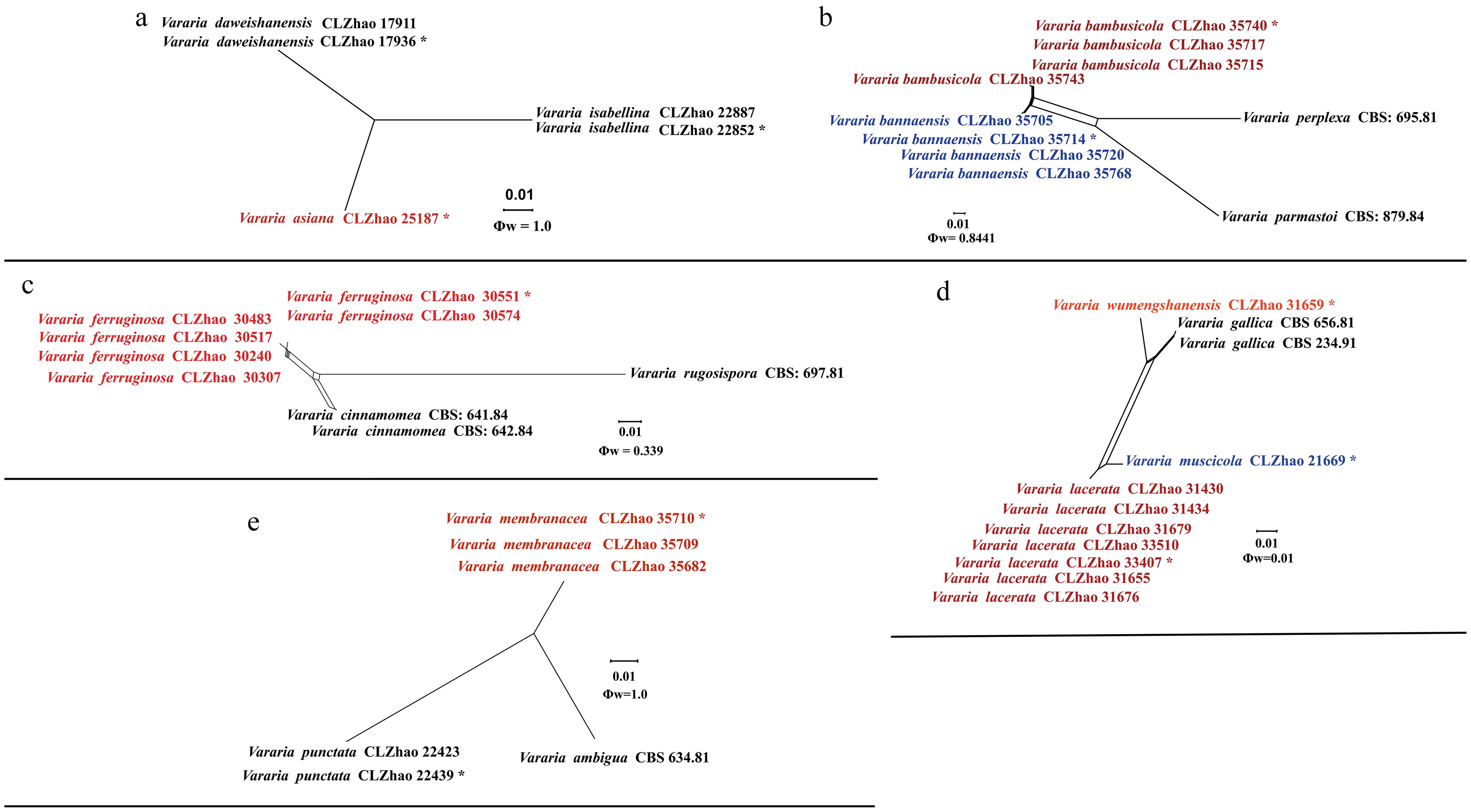
Figure 27.
Pairwise homoplasy index (PHI) test of the new species of Vararia and closely related species using both LogDet transformation and splits decomposition. PHI test results Φw ≤ 0.05 indicate that there is significant recombination within the dataset. The new taxa are in bold red type.
Vararia bambusicola Y.L. Deng & C.L. Zhao, sp. nov. Figures 27b, 28, 29
Index Fungorum number: IF859281

Figure 28.
Basidiomata and microscopic structures of Vararia bambusicola (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Urniform gloeocystidia. (f) Mushroom-shaped gloeocystidia. (g) Fusiform gloeocystidia. (h) Dichohyphae. (i) Section of the hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c)–(i) = 10 μm.
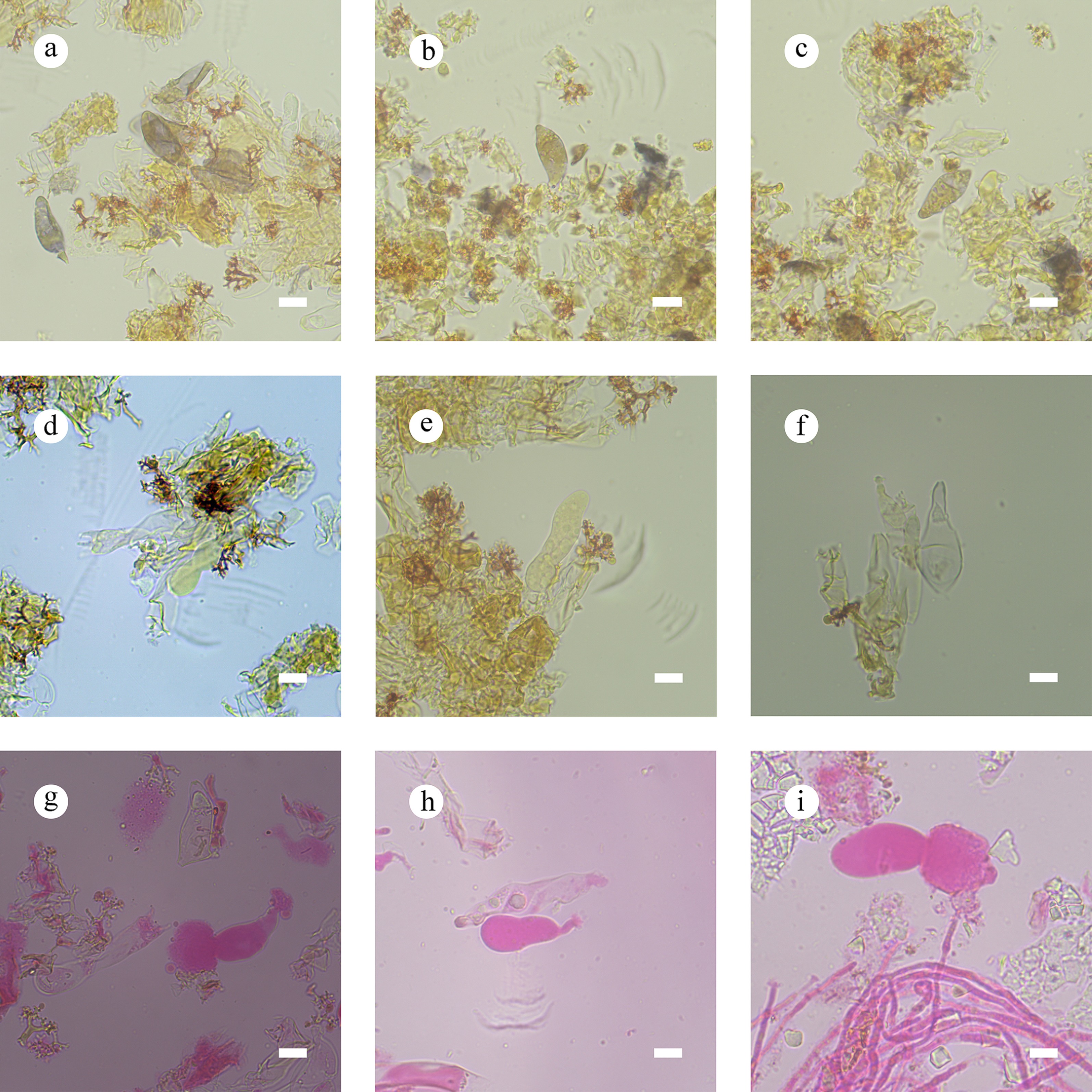
Figure 29.
Sections of the hymenium of Vararia bambusicola (holotype). (a)–(c) Basidiospores. (d) Basidia. (e) Basidioles. (f)–(i) Gloeocystidia. Scale bars: (a)–(i) = 10 µm.
Diagnosis – Vararia bambusicola differs from other species by its coriaceous basidiomata, a pink to salmon to peach hymenial surface, generative hyphae bearing simple septa, and three types of gloeocystidia (urniform, mushroom-shaped, and fusiform), as well as ellipsoid basidiospores.
Etymology – referring to the substrate of bamboo of the type specimen.
Type – China, Yunnan Province, Dehong, Xishuangbanna, Jinghong City, Rubber Plantation, 21°90′ N, 100°76′ E, 552.7 m asl, on dead bamboo, leg. C.L. Zhao, 25 January 2024, CLZhao 35740 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, coriaceous, without odor or taste when fresh, up to 10 cm long, 1.5 cm wide, and 120 μm thick. Hymenial surface smooth, pink to salmon when fresh, becoming salmon to peach when dry. Sterile margin pink, thinning out, up to 1 mm.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, frequently branched, 2.5–3 µm in diameter. Hymenium skeletal hyphae fibriform, thin-walled, colorless, 2 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Dichohyphae predominant, thin-walled, yellowish, capillary, frequently branched, dichotomously to irregularly branched with main branch and blunt tips, 2.5–3 µm in diameter, weakly to moderately dextrinoid in Melzer’s reagent, CB–; tissues unchanged in KOH. Gloeocystidia of three types: (1) urniform gloeocystidia, colorless, thin-walled, smooth, 16.3–28 × 10–14 µm; (2) mushroom-shaped gloeocystidia, colorless, thin-walled, smooth, 21.3–37 × 12.7–13 µm, flocculent at the top, up to 11–32 × 8–17 µm; and (3) fusiform gloeocystidia, colorless, thin-walled, smooth, swelling of the base and shrinkage apex, slightly flexuous, filled with refractive matter, 35.4–47.3 × 7.8–14.1 µm. Basidia cylindrical to clavate, slightly flexuous, with a basal simple septum and four sterigmata, 29–57 × 8.5–12.3 µm; basidioles numerous, in shape similar to basidia but smaller, filled with refractive oil-like matter. Basidiospores cylindrical to fusiform, colorless, thin-walled, smooth, CB–, IKI+, (20.1–)22.1–25.1(–26) × (8.6–)8.7–10.1(–10.8) µm, L = 23.19 µm, W = 9.4 µm, Q = 2.47 (n = 120/4).
Additional material examined – China, Yunnan Province, Xishuangbanna, Jinghong City, Rubber Plantation, 21°90′ N, 100°76′ E, 552.7 m asl, on dead bamboo, leg. C.L. Zhao, 25 January 2024, CLZhao 35715, CLZhao 35717, CLZhao 35743 (SWFC).
GenBank accession numbers – CLZhao 35740 ITS: PV637442, LSU: PV637449, mtSSU: PV637456, rpb1: PX441310, rpb2: PV648921, tef1- α: PV648915; CLZhao 35715 ITS: PV637440, LSU: PV637447, mtSSU: PV637454; CLZhao 35717 ITS: PV637441, LSU: PV637448, mtSSU: PV637455, tef1- α: PV648914; CLZhao 35743 ITS: PV637443, LSU: PV637450, mtSSU: PV637457
Notes – The phylogenetic analyses ( Fig. 5) reveal that the specimens of Vararia bambusicola grouped within Vararia and are sister to V. bannaensis. However, morphologically, V. bannaensis is distinguished from V. bambusicola by its white to pinkish buff to cinnamon buff hymenial surface, two types gloeocystidia (urniform, 9–20 × 8–14 µm and subcylindrical to fusiform, 23–42 × 6–10 µm), smaller subclavate basidia (20.5–26 × 5.5–7 µm), and smaller fusiform to ellipsoid basidiospores (11.8–16.1 × 5.6–7.7 µm). Morphologically, V. bambusicola is similar to V. perplexa Boidin, Lanq. & Gilles and V. parmastoi Boidin & Lanq. by sharing resupinate, adnate, coriaceous basidiomata and smooth hymenial surface. However, V. perplexa has narrower utriform basidia (30–35 × 5–6 µm) and smaller basidiospores (12.5–16 × 3–4.2 µm) [ 216] . V. parmastoi can be distinguished from V. bambusicola by narrower basidia (35–42 × 4–5 µm) and smaller basidiospores (6–7.5 × 1.8–2.4 µm) [ 223] . The PHI analysis reveals no significant evidence of genetic recombination among these species ( Fig. 27b). Thus, based on morphology and multigene phylogenetic evidence, we introduce V. bambusicola as a new species.
Vararia bannaensis Y.L. Deng & C.L. Zhao, sp. nov. Figures 27b, 30, 31
Index Fungorum number: IF859282

Figure 30.
Basidiomata and microscopic structures of Vararia bannaensis (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (d) Basidia and basidioles. (e), (f) Gloeocystidia. (g) Section of the hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) Basidiospores. Scale bars: (c)–(g) = 10 μm.

Figure 31.
Sections of hymenium of Vararia bannaensis (holotype). (a) Basidiospores. (b) Basidia. (c)–(e) Basidioles and gloeocystidia. (f) Dichohyphae. Scale bars: (a)–(f) = 10 µm.
Diagnosis – Vararia bannaensis differs from other species by thin, membranous basidiomata, white to pinkish buff to cinnamon buff hymenial surface, urniform, and subcylindrical to fusiform gloeocystidia, and fusiform to ellipsoid basidiospores.
Etymology – referring to the locality (Xishuangbanna) of the type specimen.
Type – China, Yunnan Province, Dehong, Xishuangbanna, Jinghong City, Rubber Plantation, 21°90′ N, 100°76′ E, 552.7 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 25 January, 2024, CLZhao 35714 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, thin, membranous, without odor or taste when fresh, up to 7 cm long, 2 cm wide, and 100 μm thick. Hymenial surface smooth, white when fresh and becoming pinkish buff to cinnamon buff when dry. Sterile margin white, thinning out, up to 2 mm. Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, frequently branched, 2–3.4 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Dichohyphae predominant, thin-walled, yellowish, capillary, frequently branched, dichotomously to irregularly branched with main branch and blunt tips, 1.5–2.3 µm in diameter, weakly to moderately dextrinoid in Melzer's reagent, CB–; tissues unchanged in KOH. Gloeocystidia with two types: (1) urniform gloeocystidia, colorless, thin-walled, smooth, 9–20 × 8–14 µm; and (2) subcylindrical to fusiform gloeocystidia, with a long beak in the apical part, colorless, thin-walled, smooth, 23–42 × 6–10 µm. Basidia subclavate, flexuous, with a basal simple septum and four sterigmata, 20.5–26 × 5.5–7 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores fusiform to ellipsoid, colorless, thin-walled, smooth, CB–, IKI+, (9.5–)11.8–16.1(–17.2) × (4.8–)5.6–7.7(–8.3) µm, L = 14.30 µm, W = 6.56 µm, Q = 2.09–2.28 (n = 120/4).
Additional material examined – China, Yunnan Province, Xishuangbanna, Jinghong City, Rubber Plantation, 21°90′ N, 100°76′ E, 552.7 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 25 January, 2024, CLZhao 35705, CLZhao 35720, CLZhao 35768 (SWFC).
GenBank accession numbers – CLZhao 35714 ITS: PV441143, LSU: PV441157, rpb2: PV648920; CLZhao 35705 ITS: PV441142, LSU: PV441156, mtSSU: PV461165, rpb2: PV648919; CLZhao 35720 ITS: PV441144, LSU: PV441158, mtSSU: PV461166, rpb2: PX432799; CLZhao 35768 ITS: PV441145, LSU: PV441159, mtSSU: PV461167, rpb2: PX432800
Notes – The phylogram ( Fig. 5) revealed that Vararia bannaensis is grouped within Vararia and sister to V. bambusicola. However, morphologically, V. bambusicola is distinguished from V. bannaensis by pink to salmon to peach hymenial surface, three types of gloeocystidia (urniform, 16.3–28 × 10–14 µm; mushroom-shaped, 21.3–37 × 12.7–13 µm, fusiform, 35.4–47.3 × 7.8–14.1 µm), larger cylindrical basidia (29–57 × 8.5–12.3 µm), and ellipsoid basidiospores (22.1–25.1 × 8.7–10.1 µm). Morphologically, V. bannaensis is similar to V. perplexa and V. parmastoi sharing resupinate, adnate, coriaceous basidiomata and smooth hymenial surface. However, morphologically, V. perplexa differs from V. bannaensis in having longer utriform basidia (30–35 × 5–6 µm) and narrower basidiospores (12.5–16 × 3–4.2 µm) [ 216] . V. parmastoi can be distinguished from V. bannaensis by longer basidia (35–42 × 4–5 µm) and smaller basidiospores (6–7.5 × 1.8–2.4 µm) [ 223] . To validate the species delineation, PHI analysis was performed. The analysis reveals no significant evidence of genetic recombination among these species ( Fig. 27b). Thus, based on morphology and multigene phylogenetic evidence, V. bannaensis is introduced as a new species.
Vararia ferruginosa Y.L. Deng & C.L. Zhao, sp. nov. Figures 27c, 32, and 33
Index Fungorum number: IF859283
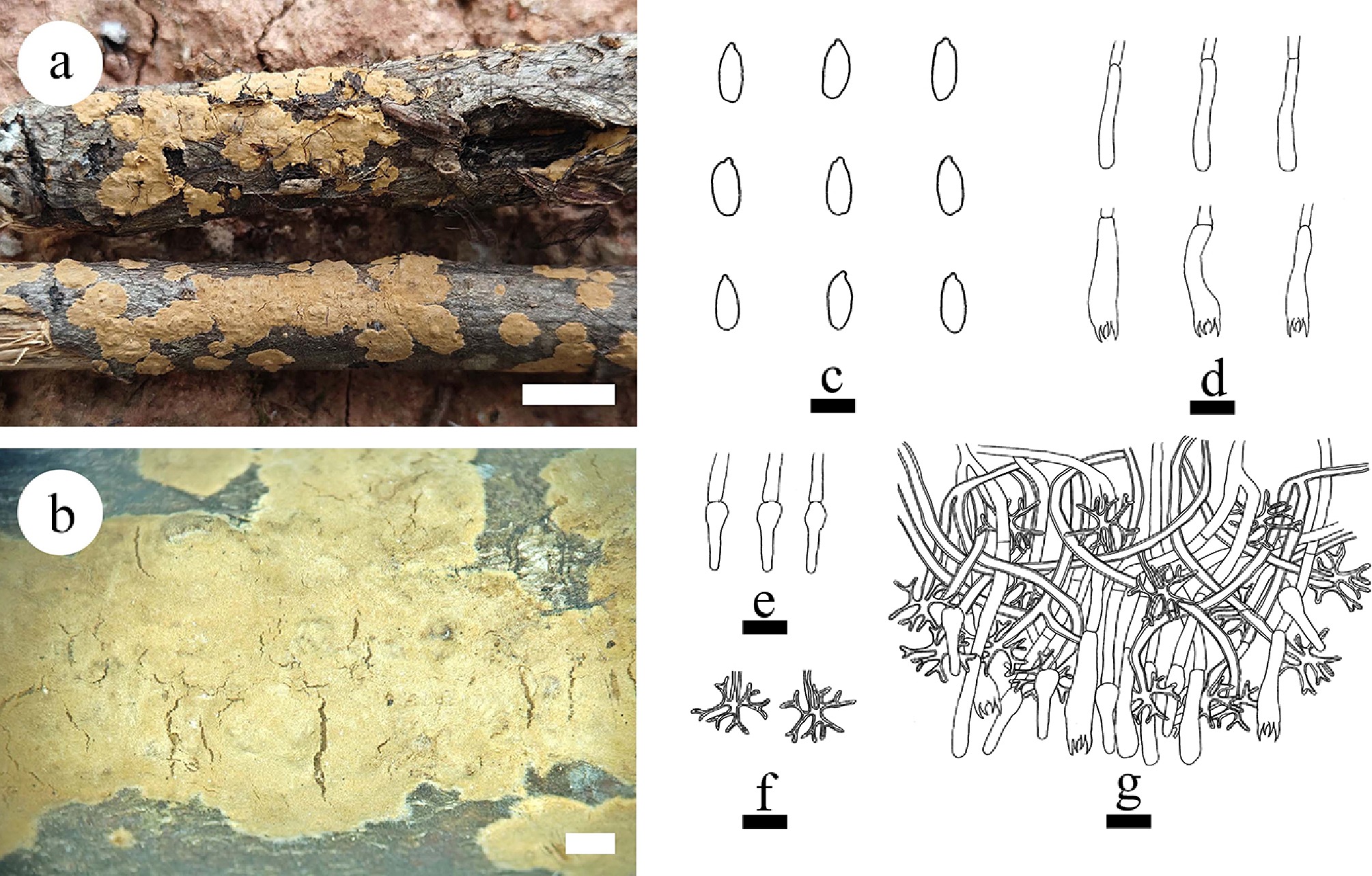
Figure 32.
Basidiomata and microscopic structures of Vararia ferruginosa (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Gloeocystidia. (f) Dichohyphae. (g) Section of the hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 µm; (d)–(g) = 10 µm.
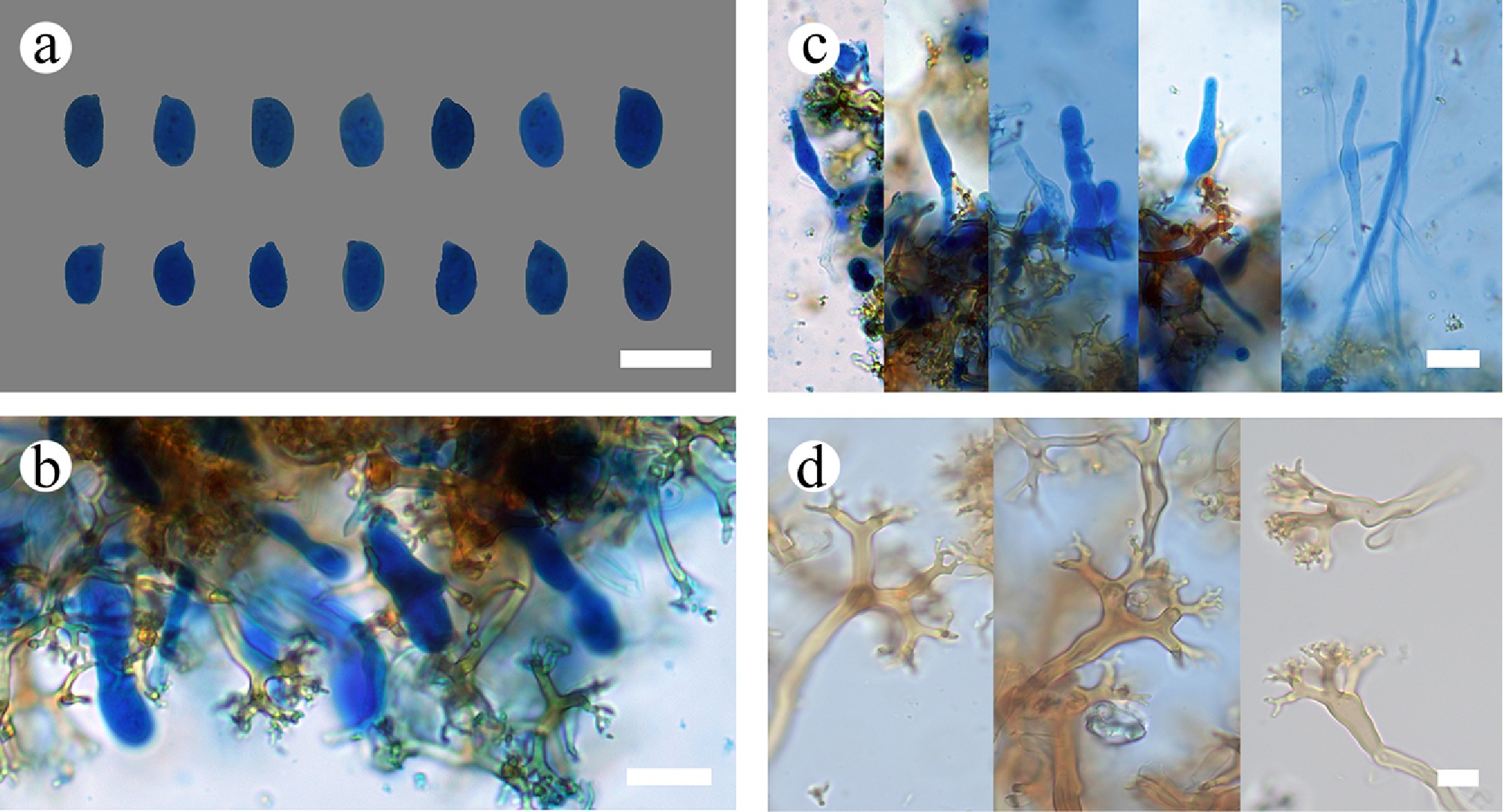
Figure 33.
Sections of the hymenium of Vararia ferruginosa (holotype). (a) Basidiospores. (b) Bbasidioles. (c) Gloeocystidia. (d) Dichohyphae. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Vararia ferruginosa differs from other species by the ferruginous to yellowish-brown hymenial surface, a dimitic hyphal system with generative hyphae bearing simple septa, and ellipsoid basidiospores.
Etymology – referring to the ferruginous color of the hymenial surface of the specimens.
Type – China, Yunnan Province, Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 19 July, 2023, CLZhao 30551 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, coriaceous, without odor or taste when fresh, up to 6 cm long, 1.5 cm wide, and 100 μm thick. Hymenial surface smooth, slightly ferruginous when fresh and ferruginous to yellowish-brown when dry. Sterile margin cream, thinning out, up to 1 mm.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, frequently branched, 2.2–3.2 µm in diameter. Hymenium skeletal hyphae fibriform, thick-walled, colorless, 2–3 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Dichohyphae predominant, thick-walled, yellowish, capillary, frequently branched, dichotomously to irregularly branched with main branch and acute tips, 1.5–3.5 µm in diameter, weakly to moderately dextrinoid in Melzer’s reagent, CB–; tissues unchanged in KOH. Gloeocystidia obclavate, colorless, thin-walled, smooth, 14–25 × 3–6 µm. Basidia fusiform to subcylindrical, flexuous, with a basal simple septum and four sterigmata, 15–28 × 3–5.3 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores ellipsoid, colorless, thin-walled, smooth, CB–, (5.1–)5.9–8.8(–9) × (2.7–)3.1–5.1(–5.3) µm, L = 7.43 µm, W = 4.25 µm, Q = 1.71–1.89 (n = 180/6).
Additional material examined – China, Yunnan Province, Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 19 July 2023, CLZhao 30240, CLZhao 30307, CLZhao 30483, CLZhao 30517, and CLZhao 30574 (SWFC).
GenBank accession numbers – CLZhao 30551 ITS: PV147175; CLZhao 30240 ITS: PV147172, LSU: PV185858, mtSSU: PV283190; CLZhao 30307 ITS: PV147177; LSU: PV185859, mtSSU: PV283192, rpb2: PV339603, tef1- α: PV400177; CLZhao 30483 ITS: PV147173, mtSSU: PV399822; CLZhao 30517 ITS: PV147174, mtSSU: PV399823; CLZhao 30574 ITS: PV147176, LSU: PV185860, mtSSU: PV283191, rpb2: PV339602, tef1- α: PV400176
Notes – The phylogenetic analyses ( Fig. 5) revealed that the six specimens of Vararia ferruginosa grouped within Vararia and are related to V. cinnamomea as sister. Morphologically, V. cinnamomea can be distinguished from V. ferruginosa by larger subcylindrical basidia (45–65 × 8–10 µm), and oblong to subellipsoid basidiospores (9–13 × 5–7.2 µm) [ 223] . Morphologically, V. ferruginosa is similar to V. rugosispora Boidin, Lanq. & Gilles by sharing smooth hymenial surface and generative hyphae bearing simple septa. However, V. rugosispora differs by larger basidia (50–55 × 9–11 µm), and oval to oblong basidiospores (12–16 × 7–8 µm) [ 216] . The PHI analysis reveals the absence of recombination in the new species compared to the closely related taxa ( Fig. 27c). Thus, based on morphology and multigene phylogenetic evidence, V. ferruginosa is introduced as a new species.
Vararia lacerata Y.L. Deng & C.L. Zhao, sp. nov. Figures 27d, 34, and 35
Index Fungorum number: IF859284
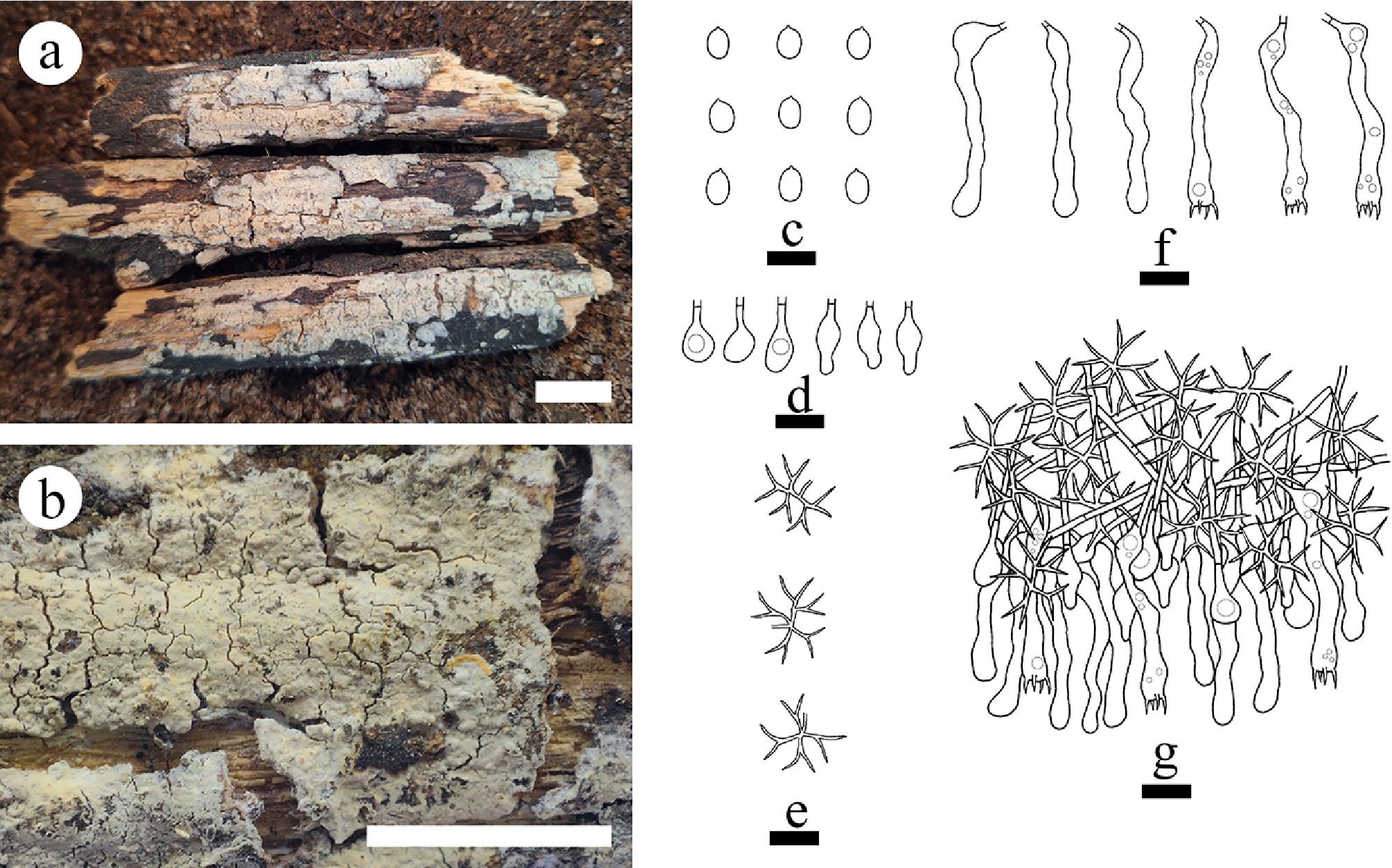
Figure 34.
Basidiomata and microscopic structures of Vararia lacerata (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Gloeocystidia. (f) Section of the hymenium. (g) Dichohyphae. Scale bars: (a) = 1 cm, (b) = 1 mm, (c)–(g) = 10 µm.
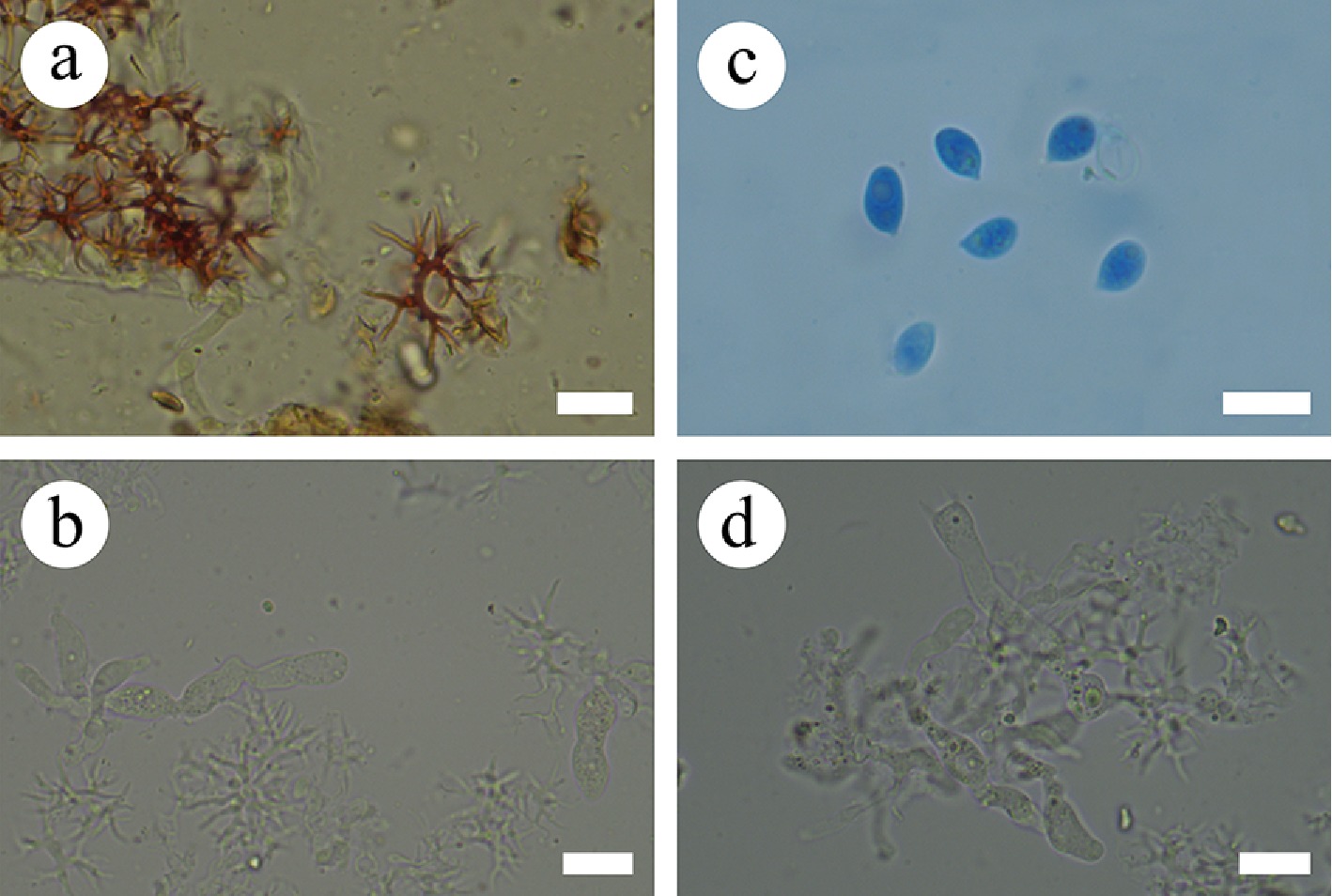
Figure 35.
Sections of the hymenium of Vararia lacerata (holotype). (a) Dichohyphae. (b) Gloeocystidia. (c) Basidiospores. (d) Basidia. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Vararia lacerata differs from other species by a cream to buff hymenial surface, a dimitic hyphal system with generative hyphae bearing simple septa, and broadly ellipsoid basidiospores.
Etymology – referring to the lacerate hymenial surface of the specimens.
Type – China, Yunnan Province, Zhaotong, Wumengshan National Nature Reserve, 27°77′ N, 104°25′ E, 1,900 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 20 September, 2023, CLZhao 33407 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, cracked, coriaceous, without odor or taste when fresh, up to 7 cm long, 2 cm wide, and 400 μm thick. Hymenial surface smooth, cream to buff in appearance when fresh, becoming buff upon drying. Sterile margin cream to buff, thinning out, up to 2 mm. Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, rarely branched, interwoven, 1.5–2 µm in diameter. Dichohyphae predominant, yellowish, capillary, frequently branched, thin-walled, dichotomously to irregularly branched with main branch and acute tips, weakly to moderately dextrinoid in Melzer’s reagent, CB–; tissues unchanged in KOH. Gloeocystidia with two types: (1) urniform, colorless, thin-walled, smooth, 12.5–16.5 × 3.5–5.5 µm; (2) capitate, colorless, thin-walled, smooth, 10.5–13 × 5–6 µm. Basidia subcylindrical to subclavate, flexuous, with a basal simple septum and four sterigmata, 34.5–37.5 × 4–5.5 µm, basidioles numerous, in shape similar to basidia but smaller. Basidiospores broadly ellipsoid, colorless, thin-walled, smooth, amyloid, CB–, 6–7.5(–8) × 3.5–5(–5.5) µm, L = 6.89 µm, W = 4.30 µm, Q = 1.47–1.60 (n = 120/4).
Additional material examined – China, Yunnan Province, Zhaotong, Wumengshan National Nature Reserve, 27°77′ N, 104°25′ E, 1,900 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 25 August, 2023, CLZhao 31430, CLZhao 31434; 26 August, 2023, CLZhao 31655, CLZhao 31676, CLZhao 31679; 20 September, 2023, CLZhao 33510 (SWFC).
GenBank accession numbers – CLZhao 33407 ITS: PQ811403, LSU: PV185851; CLZhao 31430 ITS: PQ811398; CLZhao 31434 ITS: PQ811399, LSU: PV384143, mtSSU: PV399816; CLZhao 31655 ITS: PQ811400; CLZhao 31676 ITS: PQ811401, LSU: PV185849, mtSSU: PV399817, rpb2: PV339596; CLZhao 31679 ITS: PQ811402, LSU: PV185850, tef1- α: PV400171, rpb2: PV339597; CLZhao 33510 ITS: PQ811404, LSU: PV185852, tef1- α: PV400172, rpb2: PV339598
Notes – The phylogenetic analyses ( Fig. 5) revealed that V. lacerata clustered within Vararia, and sister to V. muscicola. However, morphologically, V. muscicola is distinguished from V. lacerata by having coriaceous, fragile basidiomata growing on mossy rotting wood, white to cream hymenial surface, subcylindrical to fusiform and cylindrical (24–40 × 4–8 µm) to subclavate (12–18 × 6–7 µm) gloeocystidia, shorter fusiform to subcylindrical basidia (14–39 × 5–7 µm). The PHI test confirmed the absence of recombination in the new species compared to the closely related taxa ( Fig. 27d). Thus, based on morphology and multigene phylogenetic evidence, V. lacerata is introduced as a new species.
Vararia membranacea Y.L. Deng & C.L. Zhao, sp. nov. Figures 27e, 36, and 37
Index Fungorum number: IF859285

Figure 36.
Basidiomata and microscopic structures of Vararia membranacea (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidioles. (e) Gloeocystidia. (f) Dichohyphae. (g) Section of the hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c)–(g) = 10 μm.
Figure 37.
Sections of the hymenium of Vararia membranacea (holotype). (a) Basidiospores. (b) Basidioles. (c), (d) Gloeocystidia. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Vararia membranacea differs from other species by thin, membranous basidiomata, white to slightly cream to slightly greyish hymenial surface, gloeocystidia of two types (urniform and fusiform), and ellipsoid basidiospores.
Etymology – referring to the membranous basidiomata of the type specimen.
Type – China, Yunnan Province, Dehong, Xishuangbanna, Jinghong City, Rubber Plantation, 21°90′ N, 100°76′ E, 552.7 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 25 January, 2024, CLZhao 35710 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, thin, membranous, without odor or taste when fresh, up to 13 cm long, 1 cm wide, and 90 μm thick. Hymenial surface smooth, white to slightly cream when fresh, and becoming cream to slightly greyish when dry. Sterile margin cream, thinning out, up to 0.5 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, frequently branched, 2.3–3.2 µm in diameter. Dichohyphae predominant, thick-walled, yellowish, capillary, frequently branched, dichotomously to irregularly branched with main branch and blunt tips, 1–2 µm in diameter, weakly to moderately dextrinoid in Melzer’s reagent, CB–; tissues unchanged in KOH. Gloeocystidia with two types: (1) urniform gloeocystidia, colorless, thin-walled, smooth, 10–18 × 7–10 µm; and (2) fusiform gloeocystidia, colorless, thin-walled, smooth, 26.1–36.3 × 4.2–7.9 µm. Basidioles cylindrical, slightly flexuous, thin-walled, smooth, thin-walled, smooth, 17–29 × 5–8 µm. Basidiospores ellipsoid, colorless, thin-walled, smooth, CB–, IKI+, (6.8–)8.1–9.2(–9.4) × (4.6–)4.8–6.9(–7.5) µm, L = 8.53 µm, W = 5.70 µm, Q = 1.49 ± 0.04 (n = 30/3).
Additional material examined – China, Yunnan Province, Xishuangbanna, Jinghong City, Rubber Plantation, 21°90′ N, 100°76′ E, 552.7 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 25 January, 2024, CLZhao 35682, CLZhao 35709 (SWFC).
GenBank accession numbers – CLZhao 35710 ITS: PV637446, LSU: PV637453, mtSSU: PV637460, rpb1: PX441313, rpb2: PX432803, tef1- α: PX439084; CLZhao 35682 ITS: PV637444, LSU: PV637451, mtSSU: PV637458, rpb1: PX441311, rpb2: PX432801; CLZhao 35709 ITS: PV637445, LSU: PV637452, mtSSU: PV637459, rpb1: PX441312, rpb2: PX432802, tef1- α: PX439083
Notes – The phylogram ( Fig 5) revealed that Vararia membranacea grouped within Vararia, and is sister to V. punctata, and shows a close phylogenetic affinity to V. ambigua Boidin, Lanq. & Gilles. However, morphologically, V. punctata is distinguished from V. membranacea by its thick-walled, clavate to cylindrical gloeocystidia (12–21 × 5–9 µm) [ 33] . V. ambigua can be distinguished from V. membranacea by utriform basidia (27–40 × 3.5–4 µm), shorter basidiospores (6–7.3 × 3.4–5 µm) [ 216] . The PHI analysis revealed no significant evidence of genetic recombination among these species ( Fig. 27e). Thus, based on morphology and multigene phylogenetic evidence, we introduce V. membranacea as a new species.
Vararia muscicola Y.L. Deng & C.L. Zhao, sp. nov. Figures 27d, 38, 39
Index Fungorum number: IF859286

Figure 38.
Basidiomata of and microscopic structures Vararia muscicola (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidioles. (e) Basidia. (f) Gloeocystidia. (g) Dichohyphae. (h) Section of the hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 μm, (d)–(h) = 10 µm.
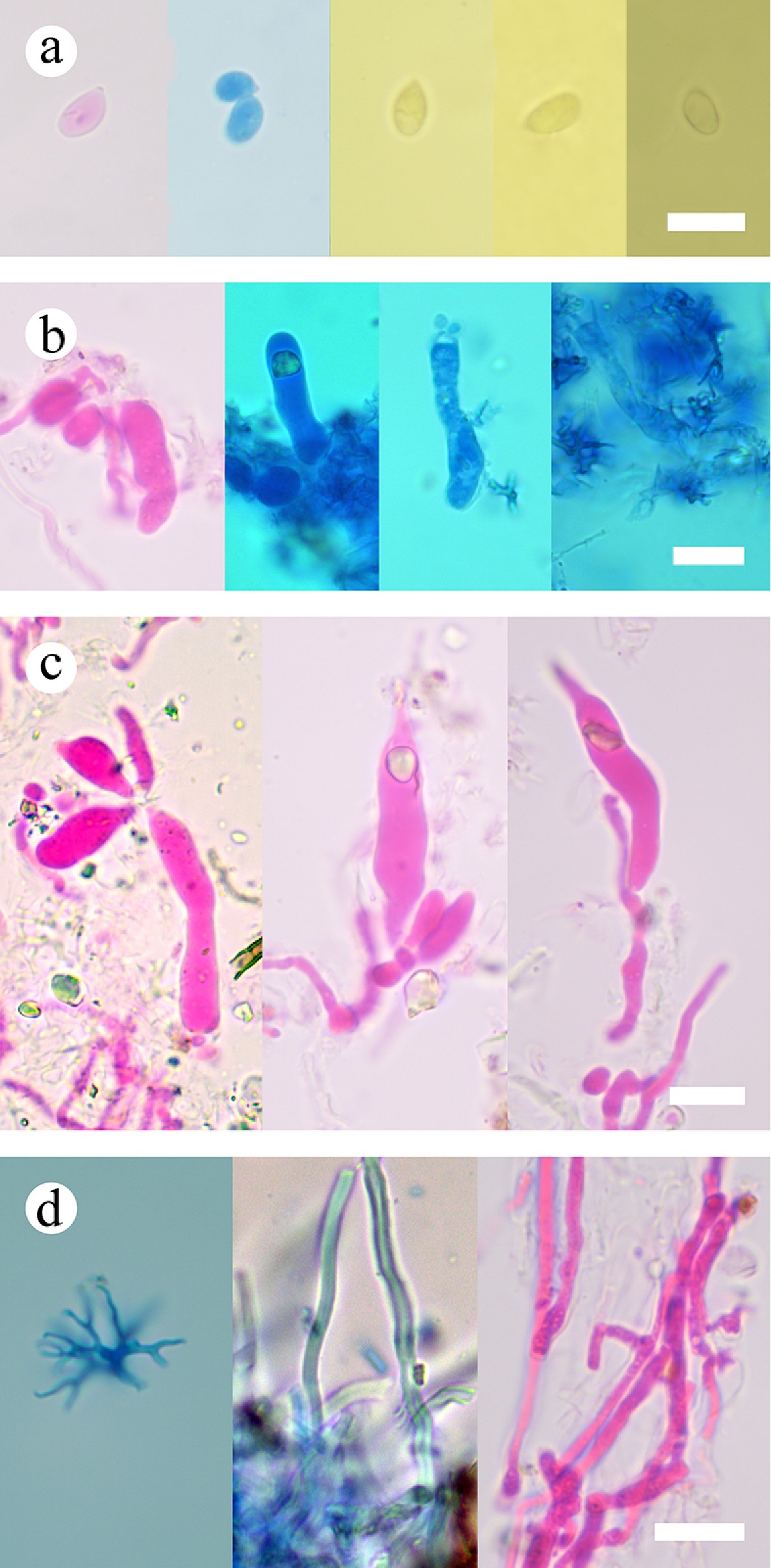
Figure 39.
Sections of the hymenium of Vararia muscicola (holotype). (a) Basidiospores. (b) Basidia and basidioles. (c) Gloeocystidia. (d) Hyphae. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Vararia muscicola differs from other species by the membranous, fragile basidiomata with a white to cream hymenial surface, generative hyphae bearing simple septa, and ellipsoid basidiospores (5.8–8.9 × 3.5–5.5 µm).
Etymology – referring to it dwelling on moss, on rotting wood.
Type – China, Yunnan Province, Lijiang, Helongtan Park, Xiangshan, 26°88′ N, 100°22′ E, 2,600 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 21 July, 2021, CLZhao 21669 (SWFC).
GenBank accession numbers – CLZhao 21669 ITS: PV147167, LSU: PV185854, mtSSU = PV399818, rpb2: PV339600
Description – Basidiomata annual, resupinate, closely adnate, membranous, fragile, growing on mossy rotting wood, without odor or taste when fresh, up to 8 cm long, 2.5 cm wide, and 200 μm thick. Hymenial surface smooth, thin, white to cream when fresh, cream upon drying. Sterile margin white, thinning out, up to 2 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, frequently branched, with some crystals, interwoven, 1.6–2 µm in diameter. Hymenium skeletal hyphae fibriform, thick-walled, colorless, 1.5–2.2 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Dichohyphae predominant, slightly thick-walled, yellowish, capillary, frequently branched, dichotomously to irregularly branched with main branch and acute tips, 1–1.6 µm in diameter, weakly to moderately dextrinoid in Melzer’s reagent, CB–; tissues unchanged in KOH. Gloeocystidia two types: (1) subcylindrical to fusiform, with a narrow or shizophapillate apex, colorless, with some refractive matter, thin-walled, smooth, 24–40 × 4–8 µm; and (2) cylindrical to subclavate, colorless, thin-walled, smooth, with some refractive matter, 12–18 × 6–7 µm. Basidia fusiform to subcylindrical, flexuous, with some refractive matter, with a basal simple septum and four sterigmata, 14–39 × 5–7 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores ellipsoid, colorless, thin-walled, smooth, CB–, (5.2–)5.8–8.9 × (3.1–)3.5–5.5(–5.7) µm, L = 7.21 µm, W = 4.77 µm, Q = 1.51 (n = 30/1).
Notes – The phylogenetic analyses ( Fig. 5) revealed that V. muscicola is clustered within Vararia and sister to V. lacerata. However, morphologically, V. lacerata is distinguished from V. muscicola by coriaceous basidiomata with cream to buff hymenial surface, urniform (12.5–16.5 × 3.5–5.5 µm) and capitate (10.5–13 × 5–6 µm) gloeocystidia, narrower subcylindrical to subclavate basidia (34.5–37.5 × 4–5.5 µm), and broadly ellipsoid basidiospores (6–7.5 × 3.5–5 µm). The PHI test confirmed the absence of recombination in the new species compared to the closely related taxa ( Fig. 27d). Thus, V. muscicola is introduced as a new species, based on both morphological and multigene phylogenetic evidence.
Vararia pingbianensis Y.L. Deng & C.L. Zhao, sp. nov. Figure 40
Index Fungorum number: IF856163

Figure 40.
Basidiomata of and microscopic structures Vararia pingbianensis (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Cystidia. (f) Dichohyphae hyphae. (g) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm. Bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 µm, (d)–(g) = 10 µm.
Diagnosis – Vararia pingbianensis differs from other species by resupinate, membranous basidiomata, a dimitic hyphal system, thin-walled generative hyphae bearing simple septa, slightly thick-walled, hyaline to light buff dichohyphae predominate in hymenium, thin-walled, ellipsoidal basidiospores.
Etymology – referring to the locality (Pingbian) of the type specimens.
Type – China, Yunnan Province, Honghe, Pingbian County, Daweishan National Nature Reserve, 22°51′ N, 103°41′ E, 2,360 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 7 June, 2020, CLZhao 18609 (SWFC).
Description – Basidiomata annual, resupinate, membranous, closely adnate, up to 20 cm long, 2 cm wide, and 150 µm thick. Hymenial surfaces smooth, sparsely and deeply cracked with age, cream to rose pink when fresh, slightly brown to dark brown upon drying. Sterile margin slightly brown, up to 0.5 mm. Hyphal system dimitic, generative hyphae bearing simple septa, thin-walled, 2–3 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Dichohyphae predominate in hymenium, frequently branched, capillary, colorless to light buff, thick-walled, 1–1.5 µm in diameter, weakly to moderately dextrinoid. Skeletal hyphae in context rarely branched, interwoven, colorless, slightly thick-walled, 1–2 µm in diameter. Cystidia pyramid, smooth, colorless, thin-walled, the apex papillaceous, 22.5–25.0 × 6–7 µm. Basidia barred-shaped, with four sterigmata and a basal simple septum, thin-walled, smooth, 16.5–23.0 × 4.5–5.5 µm, basidioles in shape similar to basidia, but slightly smaller, occasionally with some guttules. Basidiospores ellipsoid, with a distinct apiculus, thin-walled, colorless, smooth, occasionally with one or two oil drops, IKI–, CB–, 4–6 × 2.5–4.0 µm, L = 4.90 µm, W = 3.21 µm, Q =1.51–1.53 (n = 30/2).
Additional material examined – China, Yunnan Province, Lincang, Yun County, Dumu Village. 24°23′ N, 101°9′ E; 1,960 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 20 October, 2022, CLZhao 25157 (SWFC).
GenBank accession numbers – CLZhao 18609 ITS: OR195736, LSU: OR539690; CLZhao 25157 ITS: OR195737, LSU: OR510677, mtSSU: OR539689; tef1- α: PQ774593
Notes – The phylogenetic tree ( Fig. 5) inferred from the ITS + nLSU sequences shows that V. pingbianensis is grouped within the genus Vararia, and closely related to V. gomezii, V. sigmatospora, and V. trinidadensis. However, morphologically, V. gomezii differs from V. pingbianensis by having both larger fusiform basidia (30–40 × 7–8 µm), and subellipsoid basidiospores (12–14 × 4.5–5.5 µm) [ 216] ; V. sigmatospora can be distinguished by its larger basidiospores (13–22 × 2.5–3.5 µm) [ 182] ; while V. trinidadensis has larger gloeocystidia (27–50 × 7–12 µm), narrower fusiform basidia (25–30 × 4.2–5 µm), longer fusiform, and subcylindrical basidiospores (13–14.8 × 2.5–3.2 µm) [ 224] . Thus, based on both morphological and multigene phylogenetic evidence, V. pingbianensis is introduced as a new species.
Vararia wumengshanensis Y.L. Deng & C.L. Zhao, sp. nov. Figures 27d, 41, 42
Index Fungorum number: IF859287

Figure 41.
Basidiomata and microscopic structures of Vararia wumengshanensis (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Gloeocystidia. (f) Section of the hymenium. (g) Dichohyphae. Scale bars: (a) = 1 cm, (b) = 1 mm, (c)–(g) = 10 µm.
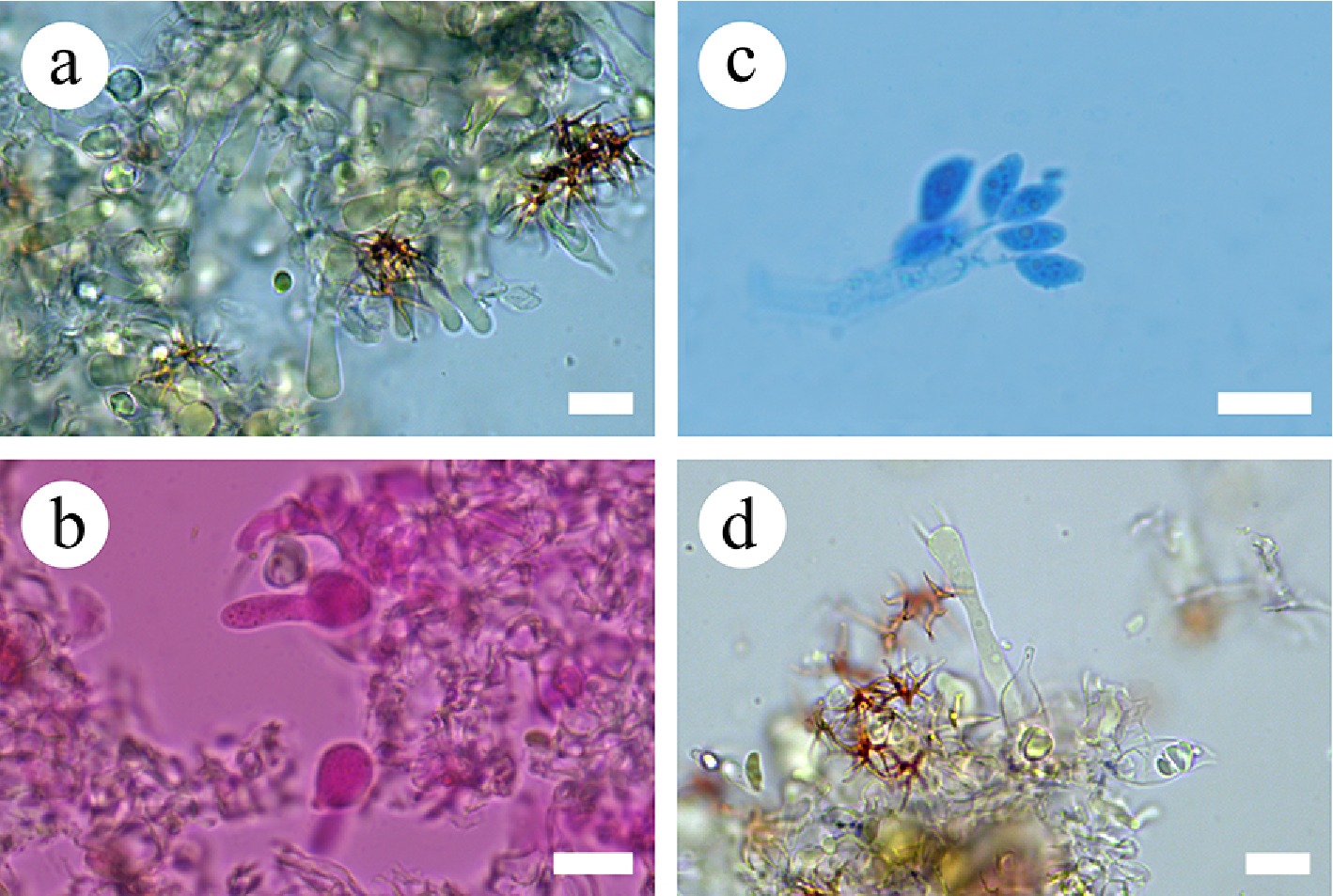
Figure 42.
Sections of the hymenium of Vararia wumengshanensis (holotype). (a) Section of the hymenium. (b) Gloeocystidia. (c) Basidiospores. (d) Basidia. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Vararia wumengshanensis differs from other species by its white to cream-colored hymenial surface, generative hyphae bearing simple septa, gloeocystidia of two types (ventricose and obclavate), and subcylindrical to narrowly smooth ellipsoid basidiospores.
Etymology – referring to the locality (Wumengshan National Nature Reserve) of the type species.
Type – China, Yunnan Province, Zhaotong, Wumengshan National Nature Reserve, 27°77′ N, 104°25′ E, 1,900 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 26 August, 2023, CLZhao 31659 (SWFC).
GenBank accession numbers – CLZhao 31659 ITS: PQ811405, LSU: PV185853, mtSSU; PV283188, rpb2: PV339599, tef1- α: PV400173
Description – Basidiomata annual, resupinate, closely adnate, membranous, without odor or taste when fresh, up to 8 cm long, 2 cm wide, and 300 μm thick. Hymenial surface smooth, thin, white to cream when fresh, cream upon drying. Sterile margin white to cream, thinning out, up to 2 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thin-walled, smooth, rarely branched, interwoven, with some crystals, 2–3.5 µm in diameter. Dichohyphae predominant, yellowish, capillary, frequently branched, thick-walled, dichotomously to irregularly branched with main branch and acute tips, weakly to moderately dextrinoid in Melzer’s reagent, CB–; tissues unchanged in KOH. Gloeocystidia with two types: (1) ventricose, with a shizopapilla in the apical part, filled with refractive oil-like matter, colorless, thin-walled, smooth, 16–19.5 × 5.5–7.5 µm; and (2) obclavate, colorless, thin-walled, smooth, 15–18 × 5.5–7 µm. Basidia subcylindrical to subclavate, flexuous, with a basal simple septum and four sterigmata, 33.5–40 × 4.5–6 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores subcylindrical to narrowly ellipsoid, colorless, thin-walled, smooth, amyloid, CB–, (7–)7.5–9.5(–10.5) × (3.5–)4–5(–5) µm, L = 8.77 µm, W = 4.12 µm, Q = 2.07 (n = 30/1).
Notes – The phylogenetic analyses ( Figs 5) revealed that V. wumengshanensis clustered within Vararia and is sister to V. gallica. However, morphologically, V. gallica differs from V. wumengshanensis by whitish hymenial surface, ventricose gloeocystidia with a shizopapilla in the apical part (15–20 × 4–7 µm), wider utriform basidia (20–40 × 5–7 µm), and longer fusiform to narrowly ellipsoid basidiospores with a very prominent apiculus (10–12 × 3–5.5 µm) [ 225] . The PHI test confirmed the absence of recombination in the new species compared to the closely related taxa ( Fig. 27d). Thus, V. wumengshanensis is introduced as a new species, based on both morphological and multigene phylogenetic evidence.
Key to 25 species of Vararia s.l. in China 1 Generative hyphae with clamp connections 2 – Generative hyphae with simple septa 8 2 Basidia with 2 sterigmata 3 – Basidia with 4 sterigmata 4 3 Subcylindrical to fusiform basidiospores (12–17 × 4.5–5.5 µm), slightly thick-walled gloeocystidia two types: subglobose (15–30 × 6–8 µm), and subcylindrical or gradually narrower (25–40 × 4.5–6 µm) Vararia amphithallica – Fusiform to cylindrical basidiospores (18–22 × 6–7.2 µm), thick-walled, ventricose, gloeocystidia with an apical papilla (20–40 × 9–12 µm) V. bispora 4 Thin to thick-walled generative hyphae, subcylindrical basidia (26–46 × 5–8 µm), allantoid basidiospores (9–13 × 3.5–5 µm), ellipsoid to ovoid to subcylindrical gloeocystidia (9–23× 7–10.5 µm) V. daweishanensis – Thin-walled generative hyphae 5 5 Gloeocystidia two types: thin-walled, subglobose (11–23 × 6–12 µm), and subulate (25.5–43 × 7–11 µm), ellipsoid to broadly ellipsoid basidiospores (5–10 × 3–7 µm) V. fissurata – Gloeocystidia one type 6 6 Basidiospores < 5 µm wide V. investiens – Basidiospores > 5 µm wide 7 7 Broadly ellipsoid basidiospores (12–16 × 9.5–13 µm), clavate basidia (70–110 × 10–16 µm), clavate gloeocystidia (50–100 × 4–9 µm) V. montana – Spherical basidiospores (8–10 × 7.5–8.5 µm), cylindrical to clavate basidia (40–45 × 6–7.5 µm), subcylindrical to fusiform gloeocystidia (48–80(–105) × 7–11(–14) µm) V. sphaericospora 8 Dichohyphae distinctly thick-walled 9 – Dichohyphae thin-walled or slightly thick-walled 20 9 Generative hyphae thin- to slightly thick-walled 10 – Generative hyphae thin-walled 14 10 Gloeocystidia one type 11 – Gloeocystidia two or three types 12 11 Gloeocystidia clavate to cylindrical, thick-walled (12–21 × 5–9 µm), subcylindrical basidia with 4 sterigmata (11–25 × 4–7 µm), ellipsoid basidiospores (6–10 × 4–6 µm), white to cream hymenial surface V. punctata – Gloeocystidia spindle to subcylindrical (38–47 × 8–13 μm), basidiospores sub-fusiform to navicular (9–13 × 5–8 µm), subcylindrical basidia with four sterigmata (33–39 × 7–9 μm), spindle to subcylindrical gloeocystidia (38–47 × 8–13 μm), cream to isabelline to slightly brown hymenial surface V. isabellina 12 Gloeocystidia three types: subcylindrical (16.5–58.5 × 4–10 µm), fusiform (18.5–43.5 × 7–9 µm), and tapering (27.5–42 × 5.5–9 µm), slightly thick-walled, ellipsoid basidiospores (5.9–11.5 × 4.7–8.6 µm), cylindrical basidia (17.5–32 × 5–9.5 µm), thin- to slightly thick-walled V. yunnanensis – Thin-walled ellipsoid basidiospores 13 13 Gloeocystidia two types: elliptical to ovoid (5.8–16 × 3.5–7 µm), subulate (16.5–27 × 4–7 µm), subcylindrical basidia with 4 sterigmata (13–23.5 × 3–4.5 µm), ellipsoid basidiospores (3.5–5.5 × 2.5–3.5 µm), buff to ochraceous hymenial surface V. fragilis – Gloeocystidia two types: thick-walled, fusiform (28.5–50 × 6–12.5 µm), subglobose (11–27 × 7–11 µm), ellipsoid basidiospores (7.6–10.8 × 5.7–7.8 µm), subclavate to subcylindrical basidia (23–46 × 5–8 µm) V. yaoshanensis 14 Thick-walled fibriform skeletal hyphae absent 15 – Thick-walled fibriform skeletal hyphae present 16 15 Hymenial surface white to cream, gloeocystidia ventricose (16–19.5 × 5.5–7.5 µm) and obclavate (15–18 × 5.5–7 µm), subcylindrical to subclavate basidia (33.5–40 × 4.5–6 µm), subcylindrical to narrowly ellipsoid basidiospores (7.5–9.5 × 4–5 µm) V. wumengshanensis – Hymenial surface white to slightly cream to slightly greyish, gloeocystidia urniform (10–18 × 7–10 µm) and fusiform (26.1–36.3 × 4.2–7.9 µm), ellipsoid basidiospores (8.1–9.2 × 4.8–6.9 µm) V. membranacea 16 Gloeocystidia two types: subcylindrical to fusiform (24–40 × 4–8 µm), cylindrical to subclavate (12–18 × 6–7 µm), fusiform to subcylindrical basidia with 4 sterigmata (14–39 × 5–7 µm), ellipsoid basidiospores (5.8–8.9 × 3.5–5.5 µm), white to cream hymenial surface V. muscicola – Gloeocystidia one type 17 17 Basidia clavate 18 – Basidia fusiform to subcylindrical 19 18 Basidia larger (25–35 × 6–7 μm), hymenial surface white to slightly pink, gloeocystidia subulate (17–35 × 6–7 μm), sub-fusiform to navicular basidiospores (6–11 × 4–6 µm) V. sinensis – Basidia smaller (11–17.5 × 2–4 μm), hymenial surface white to cream, gloeocystidia subglobose, and clavate to fusiform (6.5–16 × 3–5 µm), ellipsoid basidiospores (3.5–5.5 × 2.5–4 µm) V. lincangensis 19 Basidia subcylindrical (20–29 × 6–7.5 µm), hymenial surface cream to yellowish, gloeocystidia fusiform to subcylindrical (15–21 × 5–7 µm), broadly ellipsoid basidiospores (2.5–4 × 1.5–2.5 µm) V. asiana – Basidia fusiform to subcylindrical (15–28 × 3–5.3 µm), hymenial surface ferruginous to yellowish-brown, gloeocystidia obclavate (14–25 × 3–6 µm), basidiospores ellipsoid (5.9–8.8 × 3.1–5.1 µm) V. ferruginosa 20 Thick-walled fibriform skeletal hyphae present 21 – Thick-walled fibriform skeletal hyphae absent 22 21 Gloeocystidia three types: urniform (16.3–28 × 10–14 µm), mushroom-shaped (21.3–37 × 12.7–13 µm), fusiform (35.4–47.3 × 7.8–14.1 µm), cylindrical to clavate basidia (29–57 × 8.5–12.3 µm), cylindrical to fusiform basidiospores (22.1–25.1 × 8.7–10.1 µm) V. bambusicola – Gloeocystides absent, oblong to subellipsoid basidiospores (9–13 × 5–7.2 µm), subcylindrical basidia (45–65 × 8–10 µm) V. cinnamomea 22 Rose to orange subfusiform basidiospores ((14–)16–19(–21.5) × 4.2–6 µm), cylindrical basidia (30–53 × 6.5–7.5 µm), thick-walled, subcylindrical gloeocystides (50–65 × 6–7(–8.5) µm) V. breviphysa – Basidiospores colorless 23 23 Gloeocystidia subcylindrical to fusiform (26–40 × 4.5–9 µm), cylindrical basidiospores (6–8 × 2–3 µm), cylindrical basidia (30–40 × 4–5 µm) V. racemosa – Gloeocystidia two types 24 24 Hymenial surface cream to buff, gloeocystidia two types: urniform (12.5–16.5 × 3.5–5.5 µm), capitate (10.5–13 × 5–6 µm); subcylindrical to subclavate basidia (34.5–37.5 × 4–5.5 µm), broadly ellipsoid basidiospores (6–7.5 × 3.5–5 µm) V. lacerata – Hymenial surface white to pinkish buff to cinnamon buff, gloeocystidia two types: urniform (9–20 × 8–14 µm), subcylindrical to fusiform (23–42 × 6–10 µm), subclavate basidia (20.5–26 × 5.5–7 µm), fusiform to ellipsoid basidiospores (11.8–16.1 × 5.6–7.7 µm) V. bannaensis Family Russulaceae Lotsy 1907
Index Fungorum number: IF81358
Type genus – Russula Pers.
Notes – Russulaceae is characterized by annual, resupinate, corticioid, pleurotoid, canthareloid ( Multifurca), agaricoid, gasteroid (sequestrate), and some with veiled basidiomata, with or without milk, smooth, lamellate, regularly forked and globular or irregular hymenophore, Also, they have a monomitic to dimitic hyphal system, generative hyphae with or without clamp-connections, heteromerous trama in context, lamellae and stipe with sphaerocysts and hyphae mixed ( Lactarius, Russula and some sequestrate fungi), gloeoplerous hyphae and gloeocystidia present or absent, lamprocystidia present or absent, pseudocystidia and macrocuystidia present or absent, with asperulate, spinulose, verrucose, echinulate, ridges or crests, subreticulate to reticulate, amyloid basidiospores. Wood decay or ectomycorrizal fungi belong to this family [ 1] .
Note 47 Boidinia Stalpers & Hjortstam
Index Fungorum number: IF17162
Type species – Boidinia furfuracea (Bres.) Stalpers & Hjortstam
Notes – Boidinia was segregated from Gloeocystidiellum as a monotypic genus [ 226] , and typified by B. furfuracea. It is characterized by annual, resupinate, separable, thin, porulose or furfuraceus, sometimes pellicular to soft membranaceous basidiomata and thin- to thick-walled, globose or subglobose, echinulate or verrucose, ornamentation disappearing in KOH, strongly amyloid basidiospores with distinct apiculus as the main diagnostic features. Later authors have pointed to the suburniform shape of basidia with 1–2 sterigmata, and a basal clamp connection as an additional characteristic [ 105, 203, 227] . Several species subsequently have been placed in Boidinia. Both Boidinia and Gloeopeniophorella belong to family Russulaceae. Contrary to Lactarius and Russula that are mycorrhizal, Boidinia is saprobic and it clusters in a basal position in the clade, suggesting that the ancestor in the russuloid clade to the agaricoid forms had a corticioid basidiome and was saprotrophic [ 26, 228] .
Note 48 Gloeopeniophorella Rick
Index Fungorum number: IF22169
Type species – Gloeopeniophorella rubroflava Rick
Notes – Gloeopeniophorella was introduced with a single species G. rubroflava [ 229] , but has been largely ignored since its introduction. This genus is characterized by resupinate basidiomata, a smooth to tuberculate hymenophore surface, a monomitic hyphal system with simple-septate generative hyphae, present lamprocystidia and gloeocystidia, and amyloid ellipsoid basidiospores [ 180] . Gloeopeniophorella is characterized among Gloeocystidiellum s.l. by the presence of lamprocystidia. Boidin [ 152] restudied the type specimen and concluded that it shares similarities with Gloeocystidiellum convolvens (P. Karst.) Donk and G. laxum Sheng H. Wu, and they also made the appropriate combinations. To date, Gloeopeniophorella includes five species, namely G. bambusicola Yang Yang & C.L. Zhao, G. griseolutea Rick, G. rubroflava, G. sacrata (G. Cunn.) Hjortstam & Ryvarden, and G. singularis (Boidin, Lanq. & Gilles) Hjortstam & Ryvarden. It belongs to family Russulaceae, closely related to Boidinia based on ITS+nLSU sequence data [ 20, 26, 45] .
Note 49 Lactarius Pers.
Index Fungorum number: IF17895
Type species – Lactarius torminosus (Schaeff.) Pers
Note 50 Lactifluus (Pers.) Roussel
Notes – Lactarius is a well-known ectomycorrhizal genus that is characterized by the presence of latex, hence the common name milk caps [ 230] . Until recently, species identification of Lactarius was based on morphological characteristics such as latex color, and color change, pileus features, spore ornamentation, the presence/absence of cystidia and pileipellis structures [ 231] . Classification of Lactarius species has historically been problematic, as mycologists often use different morphological characters for infrageneric classification. Pileipellis structures have been the most frequently used character for infrageneric classification in Lactarius [ 230] . The genus has long been recognized as one of two major groups (together with the larger genus Russula Pers.) in family Russulaceae. A recent multi-locus phylogenetic study revealed that species identified as Lactarius spp. represent three genera: Multifurca Buyck & V. Hofst, Lactarius, and Lactifluus (Pers.) Roussel [ 86, 230− 235] .
Index Fungorum number: IF17896
Type species – Lactifluus piperatus (L.) Roussel
Notes – Lactifluus belongs to Russulaceae (Russulales) and is a genus of milk caps, predominantly found in subtropical and tropical regions [ 174, 232, 233, 236] . The genus was previously classified under Lactarius due to the presence of latex exudation when the basidiomata are damaged. Although there are no synapomorphic characters to distinguish between them, some morphologies tend to be well-represented in either Lactarius or Lactifluus [ 237, 238] . Upon closer inspection, Lactifluus and Lactarius have some key morphological differences. Although no synapomorphic characteristic has been found to consistently distinguish both genera, the distinction can usually be made on a combination of characters and trends. First, Lactifluus species, relative to Lactarius, are more likely to have dry pilei with a velvety to pruinose texture. Also, Lactifluus species lack zonate, hairy, viscid, or scrobiculate pilei [ 236, 237, 239, 240] . After the proposal to conserve Lactifluus as a separate genus, Lactifluus was divided into six subgenera, 13 sections, and several unclassified species [ 234, 235, 241] . A multi-locus phylogeny using global samples revealed a new classification scheme of the genus and consequently four subgenera were proposed; Lactifluus subg. Gymnocarpi (R. Heim ex Verbeken) De Crop, subg. Lactariopsis (Henn.) Verbeken, subg. Lactifluus (Pers.) Roussel, and subg. Pseudogymnocarpi (Verbeken) De Crop [ 241] . Each subgenus was further divided into four or more sections, together with undescribed clades and species in isolated phylogenetic positions [ 240] .
Note 51 Multifurca Buyck & V. Hofst.
Index Fungorum number: IF511332
Type species – Multifurca ochricompacta (Bills & O.K. Mill.) Buyck & V. Hofst.
Notes – Multifurca was described to accommodate a well-supported monophyletic clade comprising species of Russula subsect. Ochricompactae Bills & O.K. Mill. and Lactarius furcatus Coker. Mainly based on Russula subsect. Ochricompactae and sharing the same type with the subsection, as a 'hybrid' of the two genera, the small genus can nevertheless be recognized by a combination of various characters: concentrically zonate pale-colored pileus, regularly forked gills that are yellow to salmon at maturity, zonate context, dark-colored spore print, and very small and faintly ornamented spores. The known geographic distribution of Multifurca species are from southeastern North America, Central America, Oceania, and Southeast and South Asia [ 232, 242] . Multifurca is of phylogenetic importance for the phylogeny of Russulaceae as it is the only genus that exhibits features that are reminiscent of the earlier diversified resupinate genera Gloeocystidiellum Donk and Boidinia Stalpers & Hjortstam [ 232, 242] . Two species M. furcata (Coker) Buyck & V. Hofst. and M. zonaria (Buyck & Desjardin) Buyck & V. Hofst., which were originally described from USA and Thailand, respectively, were found to exist in subtropical-tropical China [ 242] . Multifurca stenophylla (Berk.) T. Lebel, C.W. Dunk & T.W. May was described and illustrated by Lebel et al. [ 243] . Subsequently, using samples from different localities and three loci (ITS, 28S rDNA and rpb2), Wang et al. [ 85] demonstrated that Multifurca is split into two highly supported major clades that are here recognized at the subgeneric level: subg. Furcata subg. nov. exclusively included lactarioid species, while subg. Multifurca included species with a russuloid habit. Meanwhile, using phylogenetic species recognition and comparison of genetic distances five new and six previously described species were recognized [ 85] . So far, 12 species have been accepted in the genus (Index Fungorum 2025).
Note 52 Pseudoxenasma K.H. Larss. & Hjortstam
Index Fungorum number: IF18416
Type species – Pseudoxenasma verrucisporum K.H. Larss. & Hjortstam
Notes – Pseudoxenasma verrucisporum has clear affinities to Russulales, and occurs as a sister taxon to that clade in phylogenetic trees [ 26] . However, the connection is not supported by bootstrap. Pseudoxenasma was established as a monotypic genus because it has pleurobasidia (lateral basidia), a feature not known from any other genus with ornamented amyloid spores and sulfo-positive cystidia. In some species with a dense, more or less gelatinized hyphal texture, generative hyphae tend to grow horizontally in the hymenium and produce basidia laterally. Both gelatinization of the hyphae and lateral production of basidia seem to facilitate quick response to favorable conditions for sporulation. Pseudoxenasma grows on dead but still attached branches of Picea abies and is often found fertile in mild periods of winter; its globose, coarsely ornamented, and strongly amyloid spores fit well with the general spore morphology in Russulales [ 26] .
Note 53 Family Stereaceae Pilát 1930
Index Fungorum number: IF81424
Type genus – Stereum Hill ex Pers. 1794
Notes – Stereaceae, typified by Stereum, formed a well-supported monophyletic clade in Russulales [ 26, 28, 45] . Stereaceae is characterized by annual, resupinate, discoid, cupulate, stereoid, auricularioid (gelatinous to fleshy, Gloeosoma) pulvinate, disciform, effused-reflexed, pileate-sessile to flabelliform basidiomata, smooth, rugose to tuberculate hymenophore. It also shows a monomitic to dimitic hyphal system, generative hyphae with or without clamp-connections, amyloid ( Amylohyphus) or inamyloid; skeletal hyphae occasionally branched, nonamyloid, nondextrinoid or binding hyphae distributed in the subiculum ( Gloeomyces); gloeoplerous hyphae and gloeocystidia present or absent, pseudocystidia present or absent, Skeletocystidia present or not, amyloid ( Amylohyphus), dextrinoid ( Dextrinocystidium) or not, acanthophyses present or absent, acanthobasidia present or absent, botryophyses with tips amiloid present in Aleurobotrys; with smooth, asperulate, spinulose, verrucose, echinulate, aculeate, ridges or crests, subreticulate to reticulate, inamyloid ( Amylohyphus) or amyloid basidiospores [ 1] . Species in the family are all saprobic, causing a white rot (or white pocket rot in some species) on both conifers and hardwoods. Although mushrooms and polypores are commonly present in different clades within the order Russulales, so far, Stereaceae contains only corticioid and stereoid wood-inhabiting fungi with a smooth or occasionally tuberculate or merulioid hymenophores. Micro-morphologically, all species in the family have gloeocystidia (skeletocystidia in Stereum), and amyloid basidiospores with smooth or ornamented walls [ 26] . Species in the family mostly belong to Stereum, Xylobolus P. Karst., Aleurodiscus s. lat., and Gloeocystidiellum s. lat. While taxa of Stereum s. str. and Xylobolus formed monophyletic clades in published phylogenetic trees, the latter two genera were highly polyphyletic, which left the generic arrangements within Stereaceae partly unresolved [ 26, 87, 103, 244, 245] . Recently, phylogenetic analyses based on a concatenated ITS1-5.8S-ITS2 (ITS) and the D1-D2 domains of nuc 28S rDNA (LSU) sequence data of Stereaceae, reported two new genera, Confertotrama and Gelatinostereum, nine new species and 20 new combinations [ 87] . In the study, based on the ITS + nLSU + rpb2 + tef1- α dataset, the divergence time of family Stereaceae occurred in a mean crown age of 179.84 Mya, with a 95% highest posterior density (HPD) of 126.6–228.84 Mya.
Note 54 Acanthobasidium Oberw. 1965
Index Fungorum number: IF17002
Type species – Acanthobasidium delicatum (Wakef.) Oberw. ex Jülich
Notes – Acanthobasidium was segregated from Aleurodiscus s.l. based on various morphological characters and supported by phylogenetic analyses [ 103] . It is characterized by thin, delicate basidiomata, a monomitic hyphal system with clamped generative hyphae, gloeocystidia moniliform, acanthocystidia and/or acanthophyses, acanthobasidia present, basidia with spines and usually ornamented subglobose basidiospores [ 246] . Acanthobasidium is a well-circumscribed group of species displaying good phylogenetic support. The presence of acanthobasidia is not unique within Aleurodiscus s. lat. [ 247] ; however, it is a strong indication of relationship with the genus [ 46, 246] . Recently, Acanthobasidium weirii (Burt) L.D. Dai & S.H. He was proposed as a new combination, and A. bambusicola L.D. Dai & S.H. He was described and illustrated by Dai & He [ 80] . Acanthobasidium quilae (Gorjón, Gresl. & Rajchenb.) Rajchenb. & Pildain was proposed based on morphology and phylogenetic analyses of ITS and 28S [ 46] . Currently, seven species are accepted in Acanthobasidium (Index Fungorum 2025) .
Note 55 Acanthophysellum Parmasto
Index Fungorum number: IF17004
Type species – Acanthophysellum lividocoeruleum (P. Karst.) Parmasto
Notes – Acanthophysellum was introduced to accommodate resupinate basidiomata, a monomitic hyphal system; generative hyphae with clamps or with simple-septa, present gloeocystidia and acanthocystidia, clavate to cylindrical basidia with 2–4 sterigmata, and cylindrical or ellipsoid to allantoid, smooth non-amyloid basidiospores, with Corticium lividocoeruleum P. Karst. as the generic type species [ 248] . However, the statement about non-amyloid spores must be regarded as a typographical error because later included Acanthophysellum in the tribe Aleurodisceae defined by the amyloid basidiospores. Corticium lividocoeruleum and Thelephora frustulata Pers., the generic type of Xylobolus, are phylogenetically and morphologically closely related [ 26] , and this connection could suggest that Acanthophysellum should be regarded as synonym of Xylobolus. However, Acanthophysellum differs from Xylobolus in some morphological characters and in having white rot (white pocket rot in Xylobolus), phenoloxidase positive reaction and nuclear behavior astatocoenocytic (holocoenocytic in Xylobolus). Genus Acanthophysium is a synonym of Xylobolus, but conserved Acanthophysellum. Xu et al. [ 87] showed that C. lividocoeruleum is the type species of Acanthophysellum, but phylogenetic analyses showed that it nested within the Xylobolus lineage. Morphologically, X. lividocoeruleus shared characters with other species in the genus a pseudo-parenchymatic subhymenium, numerous acanthohyphidia and smooth basidiospores, but is unique for its resupinate basidiomata, nodose-septate hyphae and typical gloeocystidia. With this disposition, Acanthophysellum becomes a synonym of Xylobolus. Although, four species, A. canadense (Skolko) Sheng H. Wu, Boidin & C.Y. Chie, A. fennicum (Laurila) Bernicchia & Gorjón, A. propinquum (Boidin & Gilles) Boidin & Gilles, and A. verecundum Duhem are remaining in the genus Acanthophysellum.
Note 56 Aleurodiscus Rabenh. ex J. Schröt.
Index Fungorum number: IF840214
Type species – Aleurodiscus amorphus (Pers.) J. Schröt.
Notes – Aleurodiscus belongs to family Stereaceae (Russulales). Aleurodiscus s.l. is characterized by cupulate, effused or effused-reflexed basidiomata, a monomitic or dimitic hyphal system with simple-septate or clamped generative hyphae, smooth or ornamented amyloid basidiospores, and sterile organs such as acanthophyses, gloeocystidia, hyphidia, and dendrohyphidia may be present [ 102, 103] . However, it has long been a complicated and unsettled problem as how to delimit the segregated genera among Aleurodiscus s.l. [ 19, 26, 28, 45, 103, 244] , due to highly variable diagnostic characters among species. In recent times, new species of Aleurodiscus s.l. have been proposed by Ryvarden et al. [ 249] , Gorjón et al. [ 148] , Maninder et al. [ 250] , Dai & He [ 80] , Dai et al. [ 251, 252] , Ghobad-Nejhad & Langer [ 253] , Tian et al. [ 254] , and Wu et al. [ 19] . Although Aleurodiscus s.l. had been divided into several small genera, based on different combinations of morphological characters, the phylogenetic analyses did not fully support these separations [ 80, 103] . Accordingly, the inter- and intrageneric phylogeny of Aleurodiscus s.l. in Stereaceae is still unclear and no reliable morphological characters can be used to recognize the small segregated genera. Thus, the broad concept of the genus has often been adopted by mycologists when describing new species [ 102, 148, 251, 252] .
Aleurodiscus yunnanensis Y.L. Deng & C.L. Zhao, sp. nov. Figures 43, 44
Index Fungorum number: IF860772

Figure 43.
Basidiomata and microscopic structures of Aleurodiscus yunnanensis (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Clavate gloeocystidia. (f) Acanthocystidia. (g) Moniliform cystidia. (h) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c)–(h) = 10 µm.
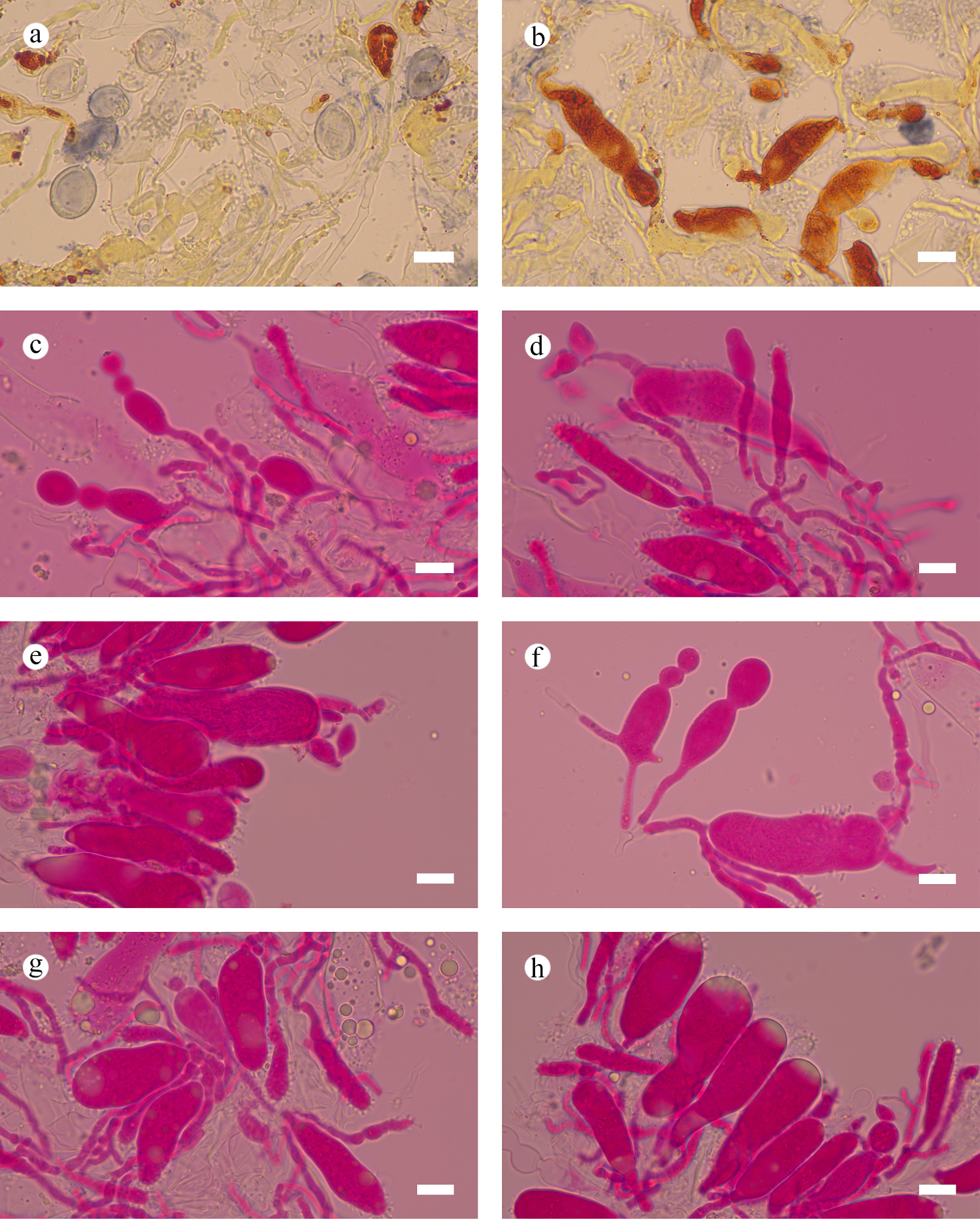
Figure 44.
Sections of hymenium of Aleurodiscus yunnanensis (holotype). (a) Basidiospores. (b), (g), (h) Clavate gloeocystidia and acanthocystidia. (c) Moniliform cystidia. (d)–(f) Basidia and basidioles. Scale bars: (a)–(h) = 10 µm.
Diagnosis – Aleurodiscus yunnanensis differs from other species by the punctate basidiomata, white to cream hymenial surface smooth, a dimitic hyphal system, generative hyphae bearing simple septa, and broadly ellipsoid to globose basidiospores.
Etymology – referring to the locality (Yunnan Province) of the type specimen.
Type – China, Yunnan Province, Diqing, Weixi County, Zhonglu Town, Lagaluo village, 27°15′ N, 99°15′ E, 3,100 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 14 October, 2023, CLZhao 34929 (SWFC).
GenBank accession numbers – CLZhao 34929 ITS: PV940926, LSU: PX070091, rpb2: PX432788, rpb1: PX441314, tef1- α: PX439085
Description – Basidiomata resupinate, adnate, membranous, punctate, without odor and taste when fresh, up to 7 cm long, 1 cm wide, and 120 µm thick. Hymenial surface smooth, white to cream when fresh, cream to slightly buff upon drying. Sterile margin white, thinning or fairly determinate, arachnoid or shortly filamentous, up to 2 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, interwoven, moderately branched, colorless, thin-walled, usually with small oily drops, 2–3 µm in diameter. Quasi-binding hyphae numerous, embedded in woody substrate, richly ramified, thin-walled, rarely branched, colorless, IKI–, CB–; tissues unchanged in KOH. Gloeocystidia numerous, immersed or slightly projecting, irregularly cylindrical, clavate, moniliform, flexuous, usually moniliform toward apices or with a schizopapillate constriction, sometimes laterally or apically protruding, usually with several big oily drops, colorless, 34.5–50 × 11–17 µm, thin-walled, weakly to moderately dextrinoid in Melzer’s reagent, CB–; tissues unchanged in KOH. Basidia cylindrical to subclavate, usually laterally with several aculei, with four sterigmata and a basal simple septum, 33–65.5 × 8–19 μm; basidioles numerous, in shape similar to basidia but smaller, 28–44.5 × 5–10 μm. Basidiospores broadly ellipsoid to globose, thin-walled, colorless, smooth to echinulate, with several minute oily drops, IKI+, CB–, (14–)14.7–17(–18) × (8.5–)9.7–12.5(–14) µm, L = 15.88 µm, W = 11.02 µm, Q = 1.44 (n = 30/1).
Notes – The phylogenetic analysis ( Fig. 6) revealed that Aleurodiscus yunnanensis is grouped within the genus Aleurodiscus in family Stereaceae, and closely related to A. wakefieldiae and A. subroseus. However, morphologically, A. wakefieldiae differs from A. yunnanensis by its bright pink to cream to pale ochraceous hymenophore, generative hyphae with clamps, hyphoid to distinctly clavate acanthophyses (30–80 × 4–12 µm), both larger slightly thick-walled and slightly moniliform gloeocystidia with a few constrictions, negative in sulphoaldehyde solution (90–180 × 5–12 µm), clavate basidia (80–180 × 10–20 µm), and ellipsoid ornamented basidiospores (20–28 × 14–20 µm) [ 185] . Aleurodiscus subroseus can be distinguished from A. yunnanensis by pinkish white to pale orange to brownish orange hymenophore, generative hyphae with clamp connections, both larger slightly thick-walled and moniliform gloeocystidia with one to several constrictions (45–70 × 6–12 µm), slightly thick-walled, subclavate to subcylindrical acanthophyses (acanthocystidia) (30–60 × 6–20 µm), and clavate basidia (52–80 × 13–17 µm), and wider basidiospores (16–20 × 11–14 µm) [ 254] . Thus, morphology and multi-gene phylogenetic analysis confirmed that the present collection is a new species, Aleurodiscus yunnanensis.
Note 57 Amylosporomyces S.S. Rattan
Index Fungorum number: IF17071
Type species – Amylosporomyces echinosporus S.S. Rattan
Notes – Hjortstam & Ryvarden [ 227] referred Gloeocystidiellum subasperisporum to Amylosporomyces, a genus described as lacking any kind of cystidia [ 255] . It is characterized by effuse basidiomata with white, pallid to pale yellow, lignicolous or corticolous hymenial surface, a monomitic hyphal system; generative hyphae with clamp connections, suburniform basidia with four sterigmata, and reniform, verruculose, amyloid basidiospores [ 255] . Only two species, A. camelicolor Khara and A. echinosporus S.S. Rattan, are accepted in Amylosporomyces [ 26] .
Note 58 Conferticium Hallenb.
Index Fungorum number: IF17349
Type species – Conferticium insidiosum (Bourdot & Galzin) Hallenb.
Notes – Conferticium (Stereaceae, Russulales) was erected in 1980 and typified by C. insidiosum, which is characterized by the resupinate basidiomata with membranaceous to ceraceous, smooth to tuberculate hymenophore, a monomitic simple-septate hyphae, and the numerous cylindrical, sinuous gloeocystidia [ 256] . Based on MycoBank (2025) and Index Fungorum (2025), nine specific and infraspecific names are registered under Conferticium, of which seven species are widely recognized [ 256, 257] . Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa revealed that C. ochraceum (Fr.) Hallenb. did not cluster with C. ravum (Burt) Ginns & G.W. Freeman, but it occurred on a single clade [ 9, 26] . The research showed that the similarities in basidiomata structures and basidia regeneration among species in Conferticium were of uncertain value for genus characterization [ 26] . Sequences of ITS and nLSU nrRNA gene regions of the studied samples were generated, in which C. ochraceum nested into family Stereaceae, and then grouped with Gloeocystidiellum, Stereum, and Gloeomyces Sheng H. Wu [ 150, 256, 257] .
Note 59 Confertotrama Nakasone & S.H. He
Index Fungorum number: IF843412
Type species – Confertotrama rugulosa (Berk. & M.A. Curtis) Nakasone & S.H. He
Notes – Confertotrama was established by Xu et al. [ 87] , and typed by C. rugulosa. Larsson & Larsson [ 26] indicated that Megalocystidium chelidonium (Pat.) Boidin, Lanq. & Gilles, Boidinia macrospora Sheng H. Wu, Gloeocystidiellum aspellum Hjortstam, G. compactum Sheng H. Wu and G. formosanum Sheng H. Wu formed a strongly supported lineage (chelidonium) in Stereaceae. Phylogenetic analyses also recovered this lineage inferring from ITS + nLSU data [ 87] . Morphologically, these species have some similar characteristics, such as yellowish basidiomata with compact texture, clamped generative hyphae and numerous large gloeocystidia with a positive sulphovanillin reaction. Basidiospores are reported as smooth in M. chelidonium and verrucose to indistinctly verrucose in the other species [ 26] . Since there are no other morphological characters that can be used to separate M. chelidonium from other species, Xu et al. [ 87] treated the lineage as a single genus and propose the new generic name Confertotrama, and proposed four newly combined species, namely C. aspella (Hjortstam) Nakasone & S.H. He, C. macrospora (Sheng H. Wu) Nakasone & S.H. He, C. rajchenbergii (Gorjón & Hallenb.) Nakasone & S.H. He, and C. rugulosa. In the present study, based on morphology and phylogenetic evidence, two new species are introduced, C. yunnanensis and C. cremea collected from China.
Confertotrama cremea Y.L. Deng & C.L. Zhao, sp. nov. Figures. 45, 46
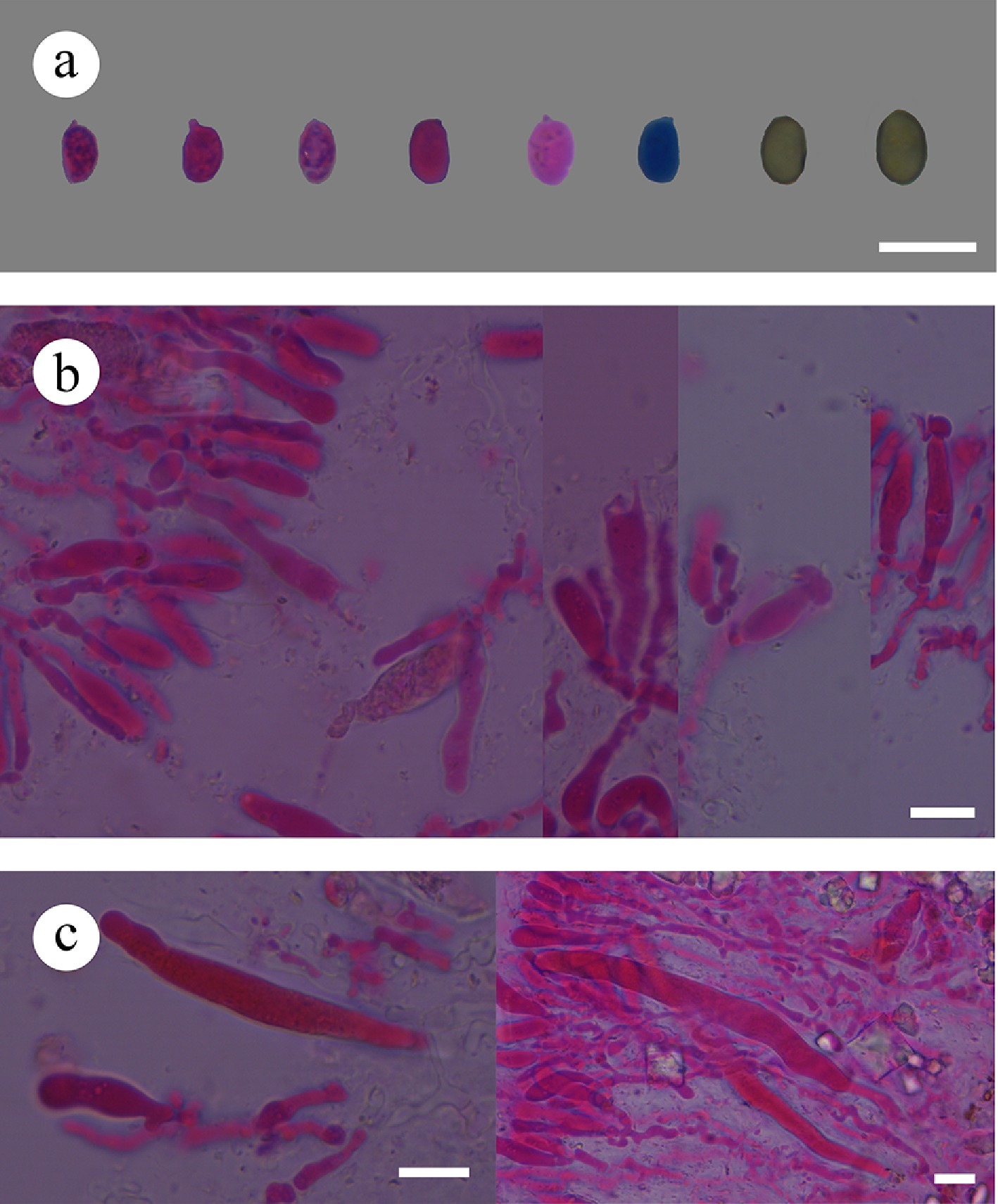
Figure 46.
Sections of hymenium of Confertotrama cremea (holotype). (a) Basidiospores. (b) Basidia and basidioles. (c) Cystidia. Scale bars: (a)–(c) = 10 µm.
Index Fungorum number: IF860773

Figure 45.
Basidiomata and microscopic structures of Confertotrama cremea (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidia and basidioles. (e) Halocystidia cystidia. (f) Cylindrical cystidia. (g) Tubular cystidia. (h) Section of the hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c) = 5 μm, (d)–(h) = 10 μm.
Diagnosis – Confertotrama cremea differs from other species by the resupinate coriaceous basidiomata with cream to slightly cream hymenial surface with tuberculate, a monomitic hyphal system with clamped generative hyphae, three types cystidia, and ellipsoid basidiospores (5.8–7.2 × 3.4–4.4 µm).
Etymology – referring to the cream basidiomata of the type species.
Type – China, Yunnan Province, Dali, Weishan County, Leqiu Town, Zhongyao village, 24°57′ N, 100°16′ E, 1,910 m asl, on angiosperm trunk, leg. C.L. Zhao, 19 July 2022, CLZhao 22749 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, coriaceous, without odor or taste when fresh, up to 7 cm long, 3 cm wide, and 200 μm thick. Hymenial surface tuberculate, cracking, slightly cream when fresh, cream upon drying. Sterile margin cream, up to 2 mm.
Hyphal system monomitic; generative hyphae with clamp connections, colorless, thin-walled, smooth, moderately branched, 2–2.5 µm in diameter, IKI–, CB–; tissues unchanged in KOH; subhymenial hyphae densely covered by crystals. Cystidia with three types: (1) Halocystidia, colorless, thin-walled, smooth, 15–27 × 4–5 µm; (2) Cylindrical cystidia, filled with refractive matter, colorless, thin-walled, smooth, 37–48 × 5–6 µm; and (3) Tubular cystidia, filled with refractive matter, colorless, thin to slightly thick-walled at the top, smooth, 72–105 × 7–10 µm. Basidia cylindrical to subclavate, with a basal clamp connection and four sterigmata, 25–28 × 5–6.5 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores ellipsoid, colorless, thin-walled, smooth, amyloid, CB–, (5.3–)5.8–7.2(–7.9) × (3.3–)3.4–4.4(–4.6) µm, L = 6.52 µm, W = 3.83 µm, Q = 1.70–1.71 (n = 60/2).
Additional material examined – China, Yunnan Province, Dali, Weishan County, Leqiu Town, Zhongyao village, 24°57′ N, 100°16′ E, 1,910 m asl, on angiosperm trunk, leg. C.L. Zhao, 19 July, 2022, CLZhao 22684 (SWFC).
GenBank accession numbers – CLZhao 22749 ITS: PV147178, LSU: PV384144, mtSSU: PV283193, rpb1 : PX441315; CLZhao 22684 ITS: PV147179, LSU: PV384145, mtSSU: PV399824, rpb1: PX441316, rpb2: PV648924
Notes – Based on the ITS + nLSU sequence data ( Fig. 6), Confertotrama cremea grouped within genus Confertotrama, forming a separate clade, and closely related to C. rugulosa. However, C. rugulosa differs from C. cremea by having folded hymenophore and strongly encrusted cystidia and has considerably smaller spores [ 258] . Based on morphology and multi-gene phylogeny, we introduced this collection as a new species, Confertotrama cremea.
Confertotrama yunnanensis Y.L. Deng & C.L. Zhao, sp. nov. Figures 47, 48
Index Fungorum number: IF860774

Figure 47.
Basidiomata and microscopic structures of Confertotrama yunnanensis (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidioles and basidia. (e), (f) Cystidia. (g) Hyphidia. (h) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 2 mm, (c)–(h) = 10 µm.
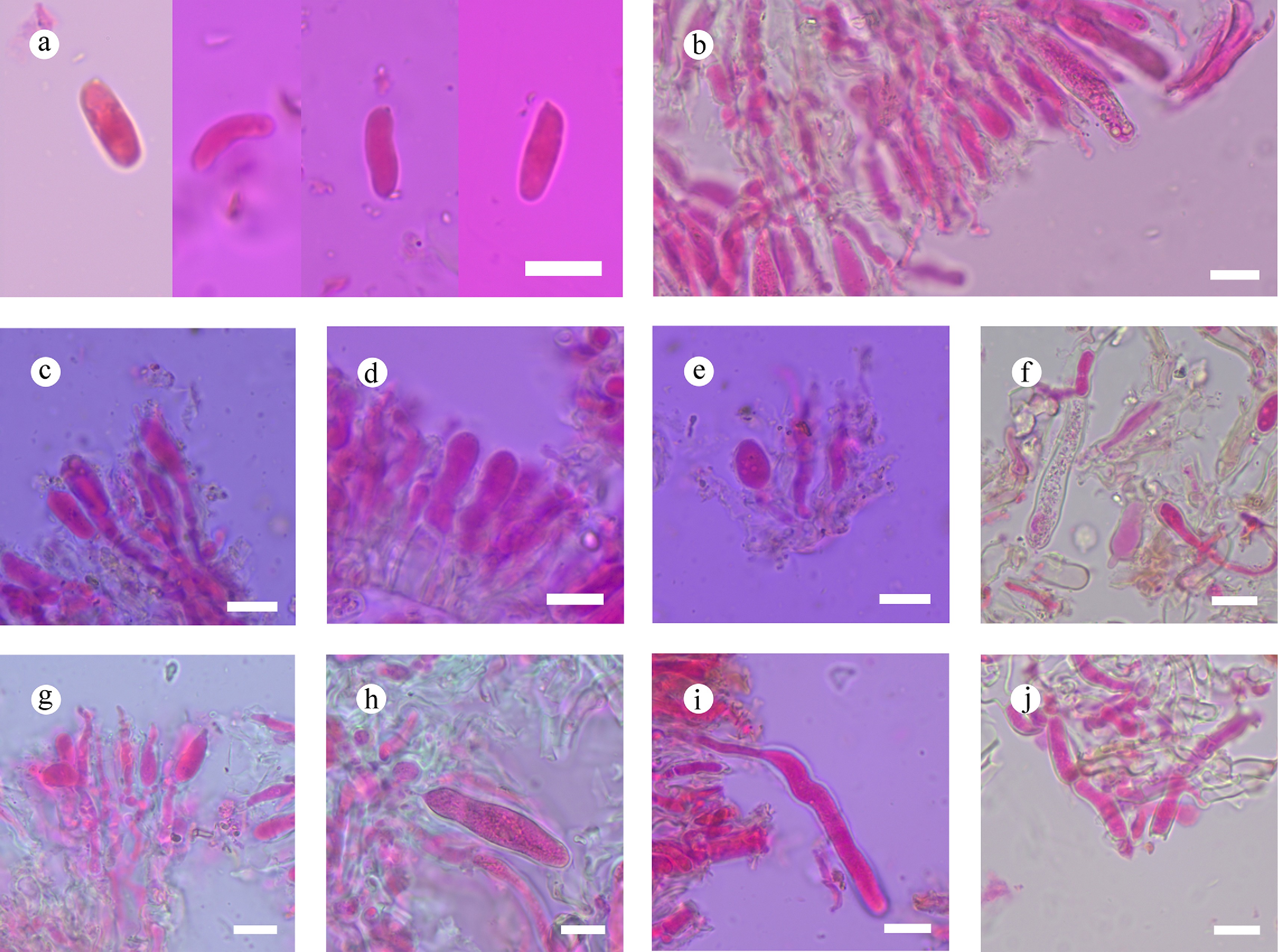
Figure 48.
Sections of hymenium of Confertotrama yunnanensis (holotype). (a) Basidiospores. (b) Section of hymenium. (c) Basidia and basidioles. (e)–(i) Cystidia. (j) Tramal hyphae. Scale bars: (a)–(j) = 10 µm.
Diagnosis – Differs from other species by the slightly ferruginous to yellowish-brown hymenial surface smooth, a hyphal system monomitic with clamped generative hyphae, allantoid basidiospores.
Etymology – referring to the locality (Yunnan Province) of the type specimen.
Type – China, Yunnan Province, Tengchong, Tuantian Town, Gaoligong Mountain National Nature Reserve, 25°55′ N, 98°58′ E, 2,600 m asl, on fallen angiosperm branch, 7 July, 2024, CLZhao 38992 (SWFC).
Description – Basidiomata annual, resupinate, closely adnate, coriaceous, without odor or taste when fresh, up to 6 cm long, 2 cm wide, and 100 μm thick. Hymenial surface smooth, slightly ferruginous when fresh and ferruginous to yellowish-brown when dry, cracked. Sterile margin slightly ferruginous, up to 1 mm wide.
Hyphal system monomitic; generative hyphae with clamp connections, colorless, thick-walled, branched, 2–3.5 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Hyphidia arising from generative hyphae, rarely branched, colorless, thin-walled, 1–3 µm in diameter. Gloeocystidia cylindrical numerous, embedded, colorless, thick-walled, containing granular refractive material, 36–45 × 5–7.5 µm; cystidia fusiform, colorless, thin-walled, with several contractions at the apex, 16.5–25 × 2–3 µm. Basidia clavate, colorless, thin-walled, with a basal clamp connection and four sterigmata, 18.5–25 × 4.5–6 µm; basidioles dominant, in shape similar to basidia, but slightly smaller. Basidiospores allantoid, colorless, thin-walled, smooth, IKI–, CB–, (6.8–)8–13.4(–15.2) × (2.8–)3.2–4.3(–4.6) µm, L = 10.89 µm, W = 3.77 µm, Q = 2.76–3.03 (n = 270/9).
Additional material examined – China, Yunnan Province, Tengchong, Tuantian Town, Gaoligong Mountain National Nature Reserve, 25°55′ N, 98°58′ E, 2,600 m asl, on fallen angiosperm branch, 7 July 2024, CLZhao 38937, CLZhao 38938, CLZhao 38973, CLZhao 38977, CLZhao 38988, CLZhao 39005, CLZhao 39017, CLZhao 39143 (SWFC).
GenBank accession numbers – CLZhao 38992 ITS: PX243636, LSU: PX243652, mtSSU: PX243667, rpb2: PX432794; CLZhao 38937 ITS: PX243633, LSU: PX243649, mtSSU: PX243664, rpb2: PX432790; CLZhao 38938 ITS: PX243632, LSU: PX243648, mtSSU: PX243663, rpb2: PX432791; CLZhao 38973 ITS: PX243633, LSU: PX243649, mtSSU: PX243664, rpb2: PX432792; CLZhao 38977 ITS: PX243634, LSU: PX243650, mtSSU: PX243665, rpb2: PX432793; CLZhao 38988 ITS: PX243635, LSU: PX243651, mtSSU: PX243666; CLZhao 39005 ITS: PX243637, LSU: PX243653, mtSSU: PX243668, rpb2: PX432795; CLZhao 39017 ITS: PX243638, LSU: PX243654, mtSSU: PX243669; CLZhao 39143 ITS: PX243639, LSU: PX243655, mtSSU: PX243670, rpb2: PX432796.
Notes – The multi-locus phylogeny ( Fig. 6) revealed that Confertotrama yunnanensis formed a separate lineage sister to C. aspella and C. rajchenbergii. However, morphologically, C. aspella differs from C. yunnanensis by having greyish orange to light orange hymenial surface, larger basidia (25–35 × 6–7 µm), and shorter ellipsoid, verrucose basidiospores (7–8 × 3.5–4 µm) [ 87] . Confertotrama rajchenbergii can be distinguished from C. yunnanensis by its yellowish ochraceous basidiomata, larger cylindrical gloeocystidia (40–70 × 5.5–7.5 µm), and shorter cylindrical to ellipsoid, verruculose, basidiospores with a straight adaxial side (6–7 × 3–3.5 µm) [ 87] . Based on morphological differences and evidence from the phylogenetic analysis, we introduce our collections, as a new species Confertotrama yunnanensis.
Note 60 Gelatinostereum S.H. He, S.L. Liu & Y.C. Dai
Index Fungorum number: IF843417
Type species – Gelatinostereum phlebioides S.H. He, S.L. Liu & Y.C. Dai
Notes – Gelatinostereum was introduced by Xu et al. [ 87] , and typified by G. phlebioides. It is characterized by resupinate to effused-reflexed, gelatinous, hygrophanous basidiomata, smooth, tuberculate to merulioid, greyish orange, brownish orange, light brown to brown hymenophore, dimitic hyphal system, generative hyphae simple-septate, smooth, amyloid basidiospores. Gelatinostereum is similar to Stereum by sharing effused-reflexed basidiomata, a dimitic hyphal system with simple-septate generative hyphae and smooth amyloid basidiospores but differs in having gelatinous and brittle basidiomata with a smooth to merulioid hymenophore and thickened subhymenium, thin-walled gloeocystidia and numerous hyphidia. In the phylogenetic tree inferring from ITS + nLSU data, Gelatinostereum formed a sister lineage to Stereum with high support values [ 87] .
Note 61 Gloeocystidiopsis Jülich
Index Fungorum number: IF17664
Type species – Gloeocystidiopsis flammea (Boidin) Jülich
Notes – Gloeocystidiopsis was introduced for Gloeocystidiellum-like species, and exhibits resupinate, effused basidiomata with ceraceous to firm-membranaceous hymenial surface, simple septate hyphae and a coenocytic nuclear behavior [ 105] . Two species, G. flammea and G. heimii (Boidin) Jülich, were originally included. Wu [ 175] moved G. heimii to Conferticium Hallenb. mainly based on his assumption on the dense hyphal structure in this species is typical for Conferticium but not present in the type of Gloeocystidiopsis. Recent research has introduced three species, viz. G. tenuissima (L.D. Dai & S.H. He) S.H. He & Y.F. Cao, G. shenghuae S.H. He & Y.F. Cao, and G. rava (Burt) S.H. He & Y.F. Cao based on morphology and phylogenetic evidence [ 87] .
Note 62 Gloeomyces Sheng H. Wu
Index Fungorum number: IF27592
Type species – Gloeomyces graminicola Sheng H. Wu
Notes – Gloeomyces graminicola, the type of Gloeomyces, lacks acanthophyses and has simple septate hyphae, while the other two species in the genus have hyphae that are nodose septate. Morphologically, species of Gloeomyces are characterized by having resupinate basidiomata, well-developed acanthohyphidia, and smooth basidiospores [ 102] . They could be compared to binding hyphae but are few and do not lend any tough consistency to the basidioma. With these characteristics Gloeomyces looks quite different from the other two species in the clade [ 26] . The colored thick-walled binding hyphae in some species might be a special form of the acanthophyses. Aleurodiscus s. str. can be easily separated from Gloeomyces by possessing ornamented basidiospores or thick-walled smooth basidiospores in A. fujii (Sheng H. Wu) S.H. He & Yun L. Xu. Based on morphological and molecular evidence, a new species, G. subcerussatus S.H. He & Ghob.-Nejh. was described and illustrated, and 11 new combinations were proposed in Gloeomyces [ 87] . More species of Aleurodiscus s. lat. will be moved to Gloeomyces after sequencing of authentic specimens [ 87] .
Note 63 Gloeosoma Bres.
Index Fungorum number: IF17672
Type species – Gloeosoma vitellinum (Lév.) Bres.
Notes – Gloeosoma was introduced by Bresadola [ 197] and typified by G. vitellinum. It is characterized by having discoid, cupulate to large and auricularioid, gelatinous to fleshy basidiomata becoming cartilaginous to hard, pink to salmon-colored hymenial surface, a monomitic hyphal system with clamped generative hyphae, present acanthocystidia and gloeocystidia, clavate basidia with four sterigmata, aculeate to echinulate, amyloid basidiospores [ 46, 197] . Gloeosoma was defined as a taxon growing on dead branches of Nothofagaceae in Patagonia, and remained monotypic over the years. A lack of information on G. vitellinum in phylogenetic studies precluded any conclusion regarding its taxonomic status. It was a matter of personal decision whether to accept the genus or consider it as the synonym of Aleurodiscus, as the microscopic characters were quite typical of the latter in a broad sense. The present study includes sequences of G. vitellinum for the first time and showed that together with G. zealandicum (Cooke & W. Phillips) Rajchenb., Pildain & C. Riquelme, G. mirabile (Berk. & M.A. Curtis) Rajchenb., Pildain & C. Riquelme, and G. decorticans Rajchenb., Pildain & C. Riquelme, they formed a well-defined group of species with strong phylogenetic support. This group clustered in the Stereaceae clade but was distant from Aleurodiscus amorphus (Pers.) J. Schröt., the type species of Aleurodiscus. The latter differs from Gloeosma morphologically by the simple-septate generative hyphae [ 46, 87] .
Note 64 Megalocystidium Jülich
Index Fungorum number: IF18027
Type species – Megalocystidium leucoxanthum (Bres.) Jülich
Notes – Megalocystidium is a genus of corticioid fungi typified by M. leucoxanthum (syn. Corticium leucoxanthum Bres.). Initially the genus was introduced to accommodate three species, and Megalocystidium currently embraces 15 species [ 87, 259, 260] . As redefined in phylogenetic studies, it belongs to family Stereaceae in order Russulales and encompasses crust-like fungi with effused basidiomata, clamped hyphae, long and deeply rooted gloeocystidia, without acanthohyphidia, as well as narrowly ellipsoid or cylindrical, smooth, strongly amyloid basidiospores [ 45, 116] . Spirin et al. [ 260] transferred Peniophora diffissa Sacc., which has abundant acanthohyphidia, into the genus. Xu et al. [ 87] showed M. chinensis (Pilát) S.H. He & Nakasone, M. wakullum (Burds., Nakasone & G.W. Freeman) E. Larss. & K.H. Larss. and other two undescribed species with simple-septate generative hyphae and acanthohyphidia formed a sister subclade to the M. leucoxanthum subclade.
Megalocystidium bambusinum Y.L. Deng & C.L. Zhao, sp. nov. Figures 49, 50
Index Fungorum number: IF860775
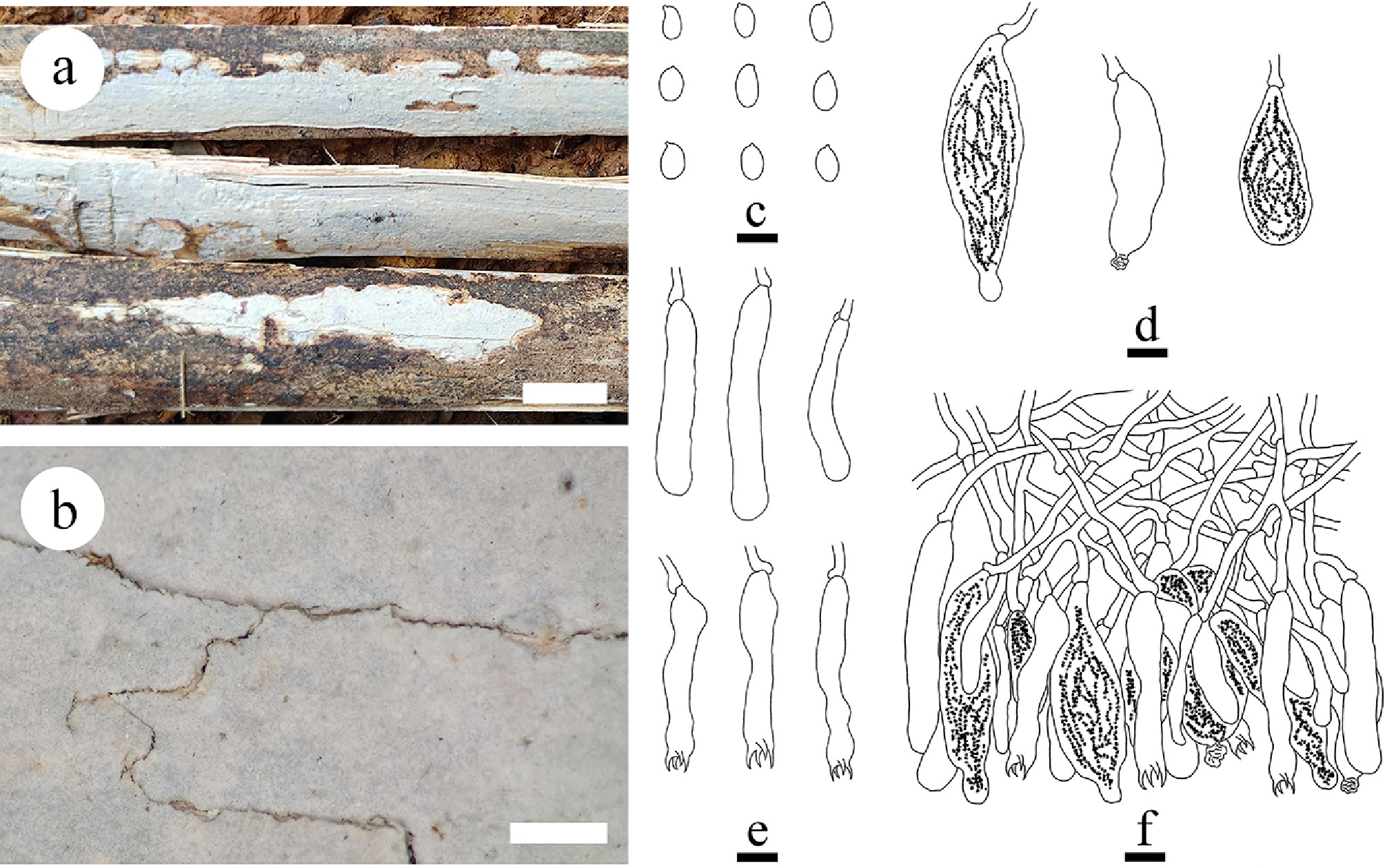
Figure 49.
Basidiomata and microscopic structures of Megalocystidium bambusinum (holotype). (a) Basidiomata on the substrate. (b) Section of hymenophore. (c) Basidiospores. (d) Basidioles and basidia. (e) Gloeocystidia. (f) Section of hymenium. Scale bars: (a) = 1 cm, (b) = 1 mm, (c)–(f) = 10 µm.
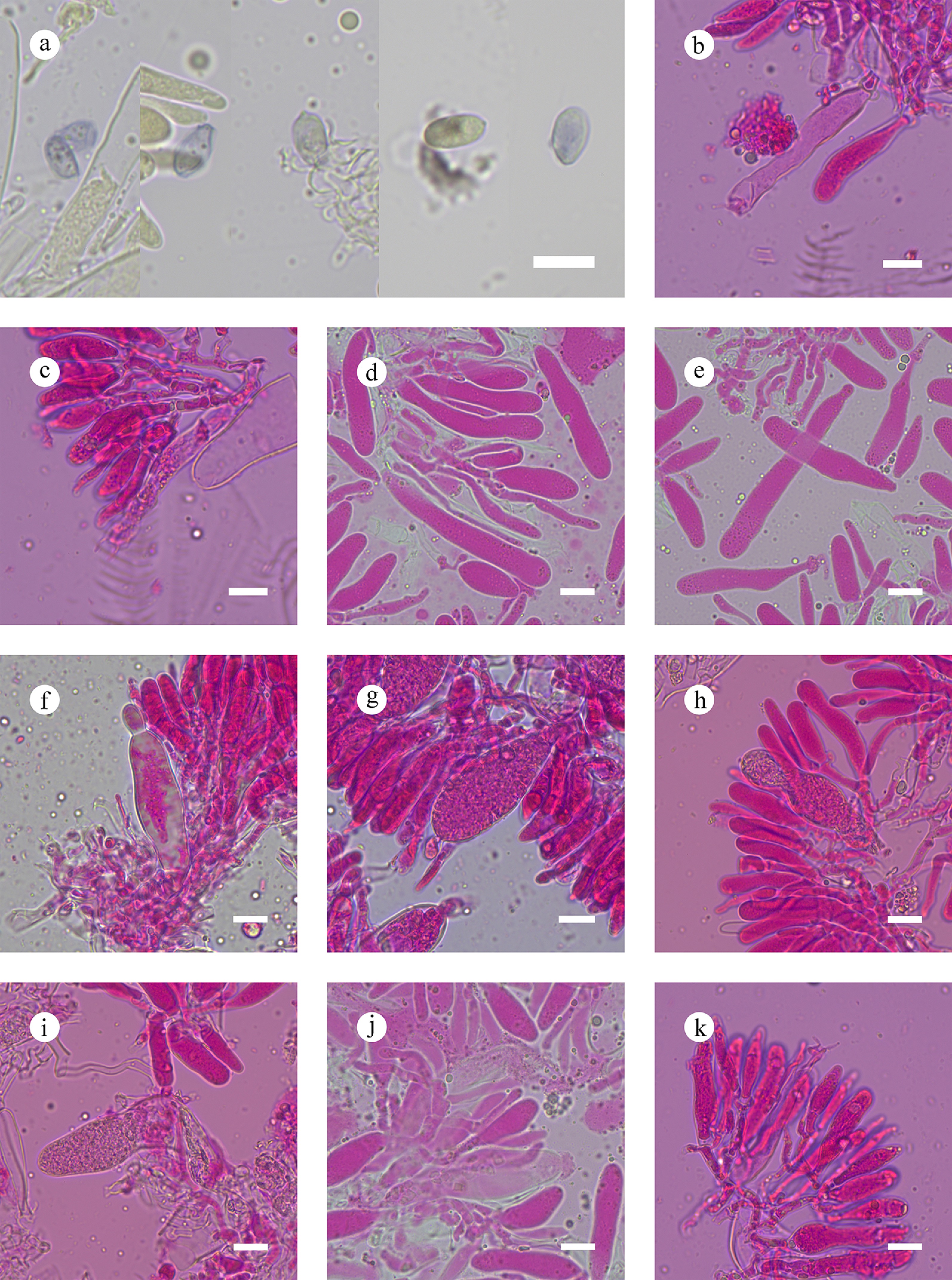
Figure 50.
Sections of hymenium of Megalocystidium bambusinum (holotype). (a) Basidiospores. (b), (c) Basidia. (d), (e) Basidioles. (f)–(j) Gloeocystidia. (k) Section of hymenium. Scale bars: (a)–(k) = 10 µm.
Diagnosis – Differs from other species by the white to cream hymenial surface smooth, a dimitic hyphal system with clamped generative hyphae, ellipsoid basidiospores (7.2–10.3 × 4–6.5 µm).
Etymology – referring to grow on bamboo of the type specimen.
Type – China, Yunnan Province, Dehong, Ruili County, Tongbiguan Provincial Nature Reserve, 23°38' N, 97°51' E, 1,500 m asl, on dead bamboo, leg. C.L. Zhao, 14 January, 2025, CLZhao 44157 (SWFC).
GenBank accession numbers – CLZhao 44157 ITS: PX243630, LSU: PX243646, mtSSU: PX243661, rpb2: PX432798, tef1- α: PX439086
Description – Basidiomata annual, resupinate, closely adnate, membranous, without odor or taste when fresh, up to 10 cm long, 1.5 cm wide, and 90 μm thick. Hymenial surface smooth, white when fresh, cream upon drying. Sterile margin thin, white, up to 1 mm wide.
Hyphal system monomitic; generative hyphae with clamp connections, colorless, thin-walled, branched, 2.5–4 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Gloeocystidia numerous, vesicular to racket-shaped, embedded, colorless, thin-walled, containing granular refractive material, with several contractions at the apex, 24–66 × 9.5–21 µm. Basidia cylindrical, flexuous, colorless, thin-walled, with a basal clamp connection and four sterigmata, 43–54.5 × 6–9 µm; basidioles dominant, in shape similar to basidia, but slightly smaller. Basidiospores ellipsoid, colorless, thin-walled, smooth, IKI+, CB–, (6.9–)7.2–10.3(–10.8) × (3.5–)4–6.5(–7) µm, L = 8.95 µm, W = 5.08 µm, Q = 1.76 (n = 30/1).
Notes – The phylogeny ( Fig. 6) revealed that Megalocystidium bambusinum formed a separate lineage and closely related to M. effusum and M. brunneum. However, morphologically, M. effusum differs from M. bambusinum by having a pale orange to light orange to greyish orange hymenophore, slightly thick-walled generative hyphae without clamps, slightly thick-walled, subclavate to subcylindrical pseudocystidia, subulate acutocystidia, smaller slightly thick-walled, cylindrical to clavate basidia (30–35 × 4–4.5 μm), and narrower oblong ellipsoid to subcylindrical basidiospores (7.5–10 × 2–3.5 μm) [ 87] ; while M. brunneum has a greyish brown to brownish orange to light brown hymenophore, thin- to thick-walled generative hyphae without clamps, slightly thick-walled, vesicular to racket-shaped gloeocystidia, and longer subcylindrical to subfusiform basidiospores (10–12 × 4–5 μm) [ 87] . Based on these morphological and phylogenetic information, it is concluded that the present collection is a new species, Megalocystidium bambusinum.
Note 65 Neoaleurodiscu s Sheng H. Wu
Index Fungorum number: IF513514
Type species – Neoaleurodiscus fujii Sheng H. Wu
Notes – Wu et al. [ 244] described the new genus, Neoaleurodiscus, based on the type species N. fujii collected from Mount Fuji in Japan on Rhododendron. Later Dai & He [ 80] also found this species in southwestern China. Morphologically, the genus mainly differs from Aleurodiscus s. str. by having thick-walled smooth basidiospores. However, the phylogenetic results by Xu et al. [ 87] , and Rajchenberg et al. [ 46] showed that N. fujii was nested within the Aleurodiscus s. str. lineage with strong support values. According to the phylogenetic delimitation of Aleurodiscus s. str., the genus is still morphologically heterogenous with simple- or nodose-septate hyphae, smooth or ornamented basidiospores, and with or without acanthohyphidia. More evidence is needed to prove whether this delimitation of Aleurodiscus s. str. is natural and reasonable.
Note 66 Stereodiscus Rajchenb. & Pildain
Index Fungorum number: IF836692
Type species – Stereodiscus antarcticus (Speg.) Rajchenb. & Pildain
Notes – Stereodiscus was established by Rajchenberg et al. [ 46] , and typified by S. antarcticus. Stereodiscus is characterized by smooth, thin-walled, cylindric, broadly ellipsoid to citriform basidiospores up to 25 μm long and absence of any of the typical cystidia in Aleurodiscus s. lat., i.e., acanthocystidia, acanthophyses, gloeocystidia, and dendrohyphidia. However, skeletocystidia are present in three species ( S. antarcticus, S. parmuliformis [G. Cunn.] Rajchenb. & Pildain, and S. trivialis [Speg.] Rajchenb. & Pildain). The hyphal system is monomitic with simple-septate hyphae, except in the recently described S. patagonicus (as Aleurodiscus), which features scattered clamps on the hyphae and at the bases of basidia [ 245] . The latter is consistent with its phylogenetic position, as it is excluded from the main cluster of species. Otherwise, the five species grouped with strong support [ 46] . Rajchenberg et al. [ 46] showed Stereodiscus to be distantly related to species of Stereum, showing features that could be synapomorphic within Stereaceae. Both Gloeosoma and Stereodiscus were shown to be phylogenetically related to each other, but distant from other clades in Stereaceae, and from other genera segregated from Aleurodiscus s. lat. A recent study proposed a new species, S. pseudotrivialis Gorjón & Gresl. based on morphological and molecular evidence [ 261] .
Note 67 Stereum Hill ex Pers.
Index Fungorum number: IF18596
Typ species – Stereum hirsutum (Willd.) Pers.
Stereum convolutomarginatum Y.L. Deng & C.L. Zhao, sp. nov. Figures 51, 52
Notes – Stereum (Stereaceae) was established in 1794 and typified with S. hirsutum. It is among the oldest described corticioid fungi and is characterized by annual to perennial, resupinate, effused, effuse-reflexed to pileate basidiomata often umbonate, seldom wholly resupinate; glabrous to tomentose or hirsute upper sterile surface, with appressed hairs and shiny, in concentric bands or completely to expose underlying cutis; smooth to slightly tuberculate hymenial surface, sometimes with concentric ridges, or with radial wrinkles on the marginal zone, usually with light colours, bruises red, orange-yellow or yellow; a dimitic hyphal system, generative hyphae with simple septa, thin- to thick-walled, skeletal hyphae; several kinds of cystidia, oleiferous pseudocystidia, skeletocystidia, acutocystidia, acanthocystidia; clavate to tubular basidia, with 4-sterigmata, simple septa at the base; and ellipsoid to cylindrical, smooth, thin-walled, amyloid basidiospores [ 26, 262] . Stereum and Xylobolus are closely related genera which form the core of family Stereaceae, which share pseudocystidia, but the latter has shorter and more ellipsoid basidiospores. Furthermore, Xylobolus has a brown context and a dense zone separating the context from a usually brown tomentum [ 263] . Three new species are introduced, S. convolutomarginatum, S. rigidohymeneum and S. yunnanense found in China, based on the ITS + nLSU data ( Fig. 6), and morphological characteristics.

Figure 51.
Basidiomata and microscopic structures of Stereum convolutomarginatum (holotype). (a), (c) Basidiomata on the natural substrates. (b), (d) A character hymenophore. (e) Basidiospores. (f) Acanthocystidia. (g) Basidia and basidioles. (h) Section of hymenium. Scale bars: (a), (c) = 1 cm, (b), (d) = 1 mm, (e)–(h) = 10 µm.
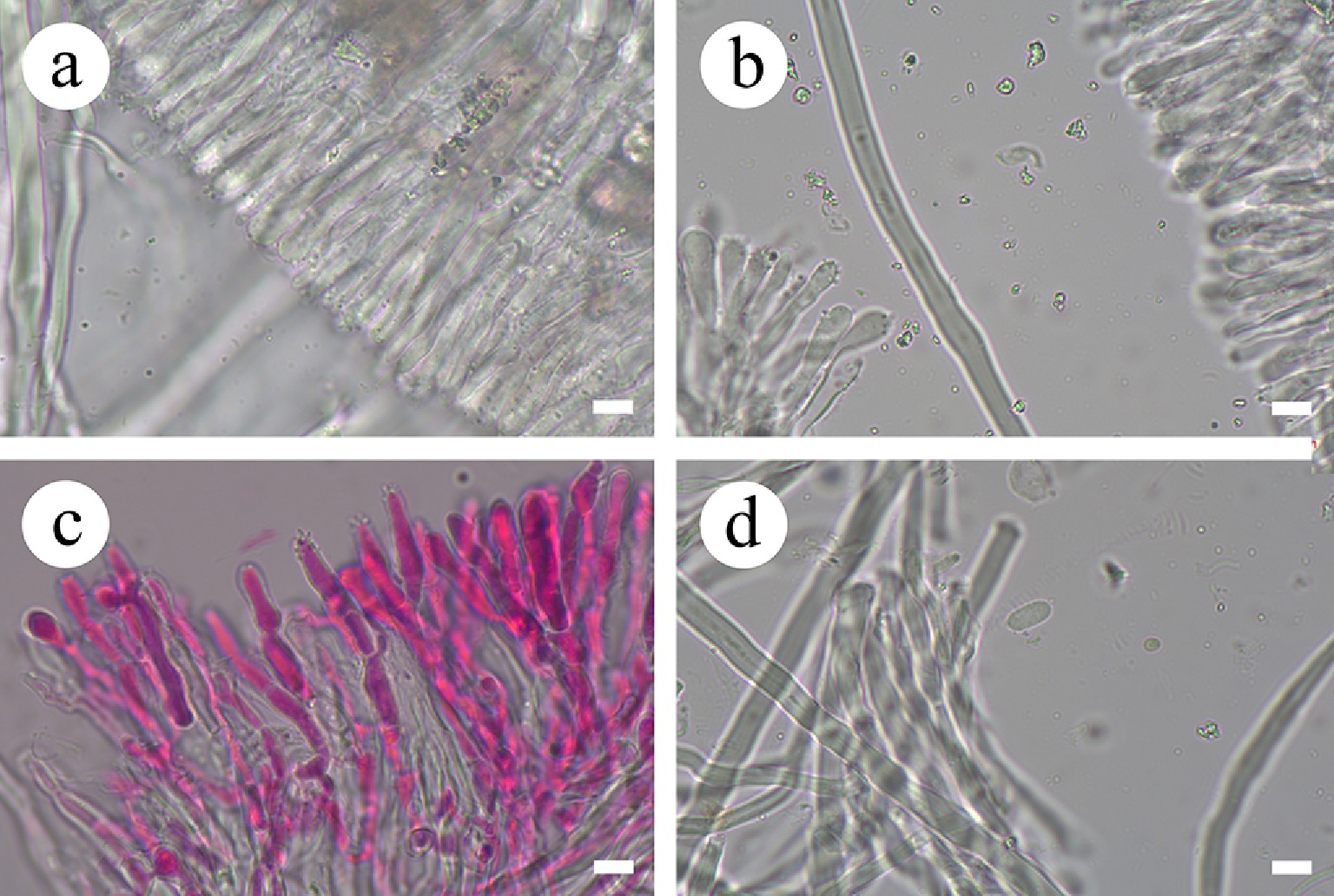
Figure 52.
Sections of hymenium of Stereum convolutomarginatum (holotype). (a), (c) Section of hymenium. (b) Skeletal hyphae. (d) Basidiospores. Scale bars: (a)–(d) = 10 µm.
Index Fungorum number: IF860776
Diagnosis – Stereum convolutomarginatum differs from other species by the resupinate to effused, coriaceous basidiomata, cream to buff hymenial surface smooth, convolutional margin, a dimitic hyphal system, generative hyphae bearing simple septa, and ellipsoid basidiospores.
Etymology – referring to the convolutional sterile margin of the specimens.
Type – China, Yunnan Province, Wenshan, Malipo County, Pingzhai Village, Forest farm, 104°42' E, 23°7'32" N, 1,057 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 18 January, 2019, CLZhao 11535 (SWFC).
Description – Basidiomata annual, resupinate to effused, coriaceous, first orbicular, later fusing together, without odour or taste when fresh, becoming hard coriaceous upon drying, up to 10.5 cm long, 2 cm wide, and 1 mm thick. Upper surface tomentose or hirsute, with appressed hairs, in concentric bands or completely to expose underlying cutis, cream when fresh, becoming cream to buff becoming, projecting up to 2 cm, 3 cm wide at center. Hymenial surface smooth, cream to buff when fresh, becoming buff to fawn upon drying. Sterile margin cream, convolutional, up to 2 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thick-walled, smooth, moderately branched, 2.8–3.3 µm in diameter. Skeletal hyphae dominant, colorless, distinctly thick-walled, narrow chamber, smooth, 5–5.5 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Acanthocystidia dominant, subclavate to subcylindrical, colorless, 20–34.4 × 4.7–7.9 µm. Basidia barrelled, with four sterigmata and a basal simple septum, 22.8–29.4 × 4.2–8 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores oblong ellipsoid, colorless, thin-walled, smooth, CB–, amyloid, (6.9–)8.9–13(–14.1) × (3.1–)3.9–5.6(–6.5) µm, L = 10.69 µm, W = 4.79 µm, Q = 4.57–4.76 (n = 420/14).
Additional material examined – China, Yunnan Province, Dali, Nanjian County, Lingbao Mountain National Forest Park, 100°30' E, 24°46' N, 2,000 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 9 January, 2019, CLZhao 10241 (SWFC); Wenshan, Malipo County, Pingzhai Village, Forest farm, 104°42' E, 23°7' N, 1,057 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 18 January, 2019, CLZhao 11416, CLZhao 11435, CLZhao 11449, CLZhao 11453, CLZhao 11455, CLZhao 11463, CLZhao 11501, CLZhao 11557, and CLZhao 11575; on angiosperm trunk, CLZhao 11399, CLZhao 11457 and CLZhao 11516 (SWFC); Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°71′ N, 94°52′ E, 1,500 m asl, on fallen angiosperm branch, leg. C.L. Zhao, 4 July, 2024, CLZhao 37604 and CLZhao 37696 (SWFC).
GenBank accession numbers – CLZhao 11535 ITS: PV283209; CLZhao 10241 ITS: PV283198; CLZhao 11416 ITS: PV283200; CLZhao 11435 ITS: PV283201, mtSSU: PV399825, rpb2: PV648927; CLZhao 11449 ITS: PV283202, LSU: PV384149, mtSSU: PV399826, rpb1: PX441317, tef1- α: PV400178; CLZhao 11453 ITS: PV283203; CLZhao 11455 ITS: PV283204, LSU: PV384150, mtSSU: PV399827, rpb1: PX441318; CLZhao 11463 ITS: PV283206, LSU: PV384152, mtSSU: PV399828; CLZhao 11501 ITS: PV283207; CLZhao 11557 ITS: PV283210; CLZhao 11575 ITS: PV283211, LSU: PV384153; CLZhao 11399 ITS: PV283199, LSU: PV384148; CLZhao 11457 ITS: PV283205, LSU: PV384151, rpb1: PX441319; CLZhao 11516 ITS: PV283208; CLZhao 37604 ITS: PX415429, LSU: PX418376, tef1- α: PX439087; CLZhao 37696 ITS: PX415428
Notes – Phylogenetic analysis ( Fig. 6) indicated that Stereum convolutomarginatum grouped within Stereum, forming a separate lineage, closely related to S. ochraceoflavum and S. vellereum. However, morphologically, S. ochraceoflavum differs from S. convolutomarginatum by its effuse-reflexed basidiomata, cream to pale yellowish orange hymenophore, slightly thick-walled, hyaline to yellowish pseudocystidia (80–100 × 5–7 µm), acutocystidia (25–35 × 3–4 µm), and smaller cylindrical to narrowly ellipsoid basidiospores (6–9 × 1.5–2.5 µm) [ 264] . Stereum vellereum can be distinguished from S. convolutomarginatum by its both smaller acutocystidioles (15–22 × 3.5–5 µm), and subcylindrical basidiospores (5–7 × 2–3 µm) [ 265] . Based on these morphological and phylogenetic evidence, we introduce our collection as a new species, Stereum convolutomarginatum.
Stereum rigidohymeneum Y.L. Deng & C.L. Zhao, sp. nov. Figures 53, 54
Index Fungorum number: IF860777

Figure 53.
Basidiomata and microscopic structures of Stereum rigidohymeneum (holotype). (a), (b) Section of hymenophore. (c), (d) The back of the basidiomata. (e) Basidiospores. (f) Basidia and basidioles. (g) Acanthocystidia. (h) Section of hymenium. Scale bars: (a), (c) = 1 cm, (b), (d) = 1 mm, (e) = 5 µm, (f)–(h) = 10 µm.
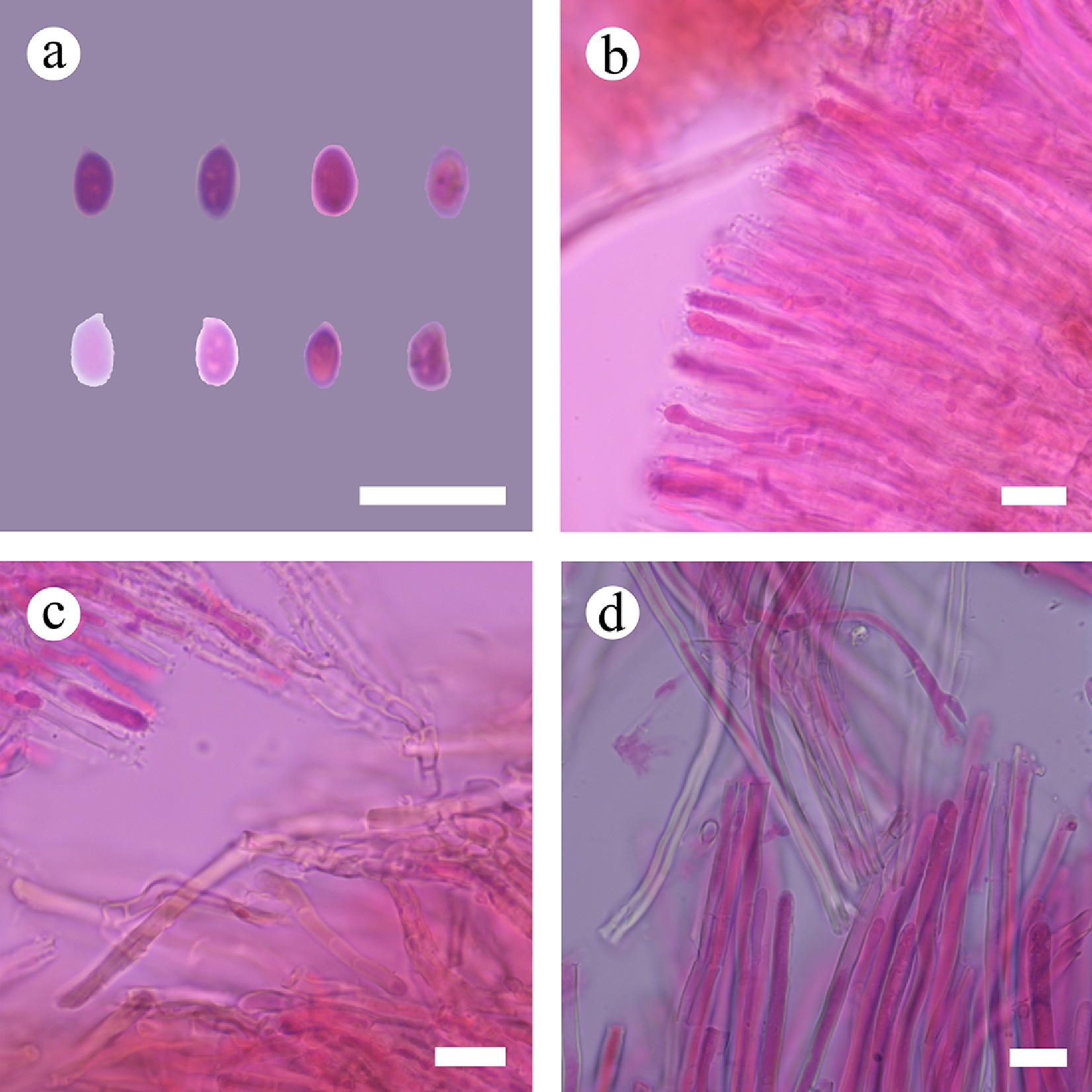
Figure 54.
Sections of hymenium of Stereum rigidohymeneum (holotype). (a) Basidiospores. (b), (c) Acanthocystidia. (d) Skeletal hyphae. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Stereum rigidohymeneum differs from other species by the perennial, rigid basidiomata, effused to pileate, cream to light orange hymenial surface with tuberculate, a dimitic hyphal system, generative hyphae bearing simple septa, ellipsoid basidiospores.
Etymology – referring to the rigid hymenial surface of the specimens.
Type – China, Yunnan Province, Puer, Jingdong County, Wuliang Mountain, 100°29' E, 24°38' N, 1,800 m asl, on angiosperm trunk, leg. C.L. Zhao, 6 October, 2017, CLZhao 4615 (SWFC).
Description – Basidiomata perennial, effused to pileate, separate from the substrate, coriaceous, rigid, without odor or taste when fresh. Pileus projecting up to 5 cm, 12 cm wide, and 4 cm thick at center. Hymenial surface slightly tuberculate, cream when fresh, cream to light orange upon drying; sterile margin brownish orange, distinct, up to 0.3 cm wide. Pileal surface brown to black, aquamarinus, in concentric bands or completely to expose underlying cutis, woody hard. Sterile margin blunt, up to 4 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thick-walled, smooth, moderately branched, 2.5–3 µm in diameter. Skeletal hyphae dominant, colorless, distinctly thick-walled, smooth, unbranched, straight, with scattered secondary simple septa, 4.9–5.3 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Acanthocystidia dominant, cylindrical, colorless, 13.8–16.4 × 2.5–3.2 µm. Basidia clavate to cylindrical, with four sterigmata and a basal simple septum, 13.3–16 × 2.7–4 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores cylindrical to ellipsoid, colorless, thin-walled, smooth, CB–, amyloid, (3.6–)3.9–5.8(–6.7) × (1.3–)1.7–2.7(–3.3) µm, L = 4.64 µm, W = 2.16 µm, Q = 1.85–2.43 (n = 60/2).
Additional material examined – China, Yunnan Province, Puer City, Jingdong County, Wuliang Mountain, 100°29' E, 24°38' N, 1,800 m asl, on angiosperm trunk, leg. C.L. Zhao, 6 Octobe,r 2017, CLZhao 4378 (SWFC).
GenBank accession numbers – CLZhao 4615 ITS: MT214936; CLZhao 4378 ITS: MT214935, mtSSU: PV283197
Notes – In the multi-locus phylogeny ( Fig. 6), the new collection of Stereum rigidohymeneum grouped within Stereum, and sister to S. insigne. However, morphologically, S. insigne can be distinguished from S. rigidohymeneum by its effuse-reflexed, coriaceous to corky basidiomata, thin- to thick-walled, hyaline to yellowish brown generative hyphae, thick-walled, brown pseudocystidia tubular with obtuse apex, both larger hyaline to brown acanthocystidia (20–30 × 4–5 µm), and narrowly clavate basidia (25–30 × 4–5 µm) [ 266] . Thus, based on morphological characters and phylogenetic analysis, the present collections are introduced as a new species, Stereum rigidohymeneum.
Stereum yunnanense Y.L. Deng & C.L. Zhao, sp. nov. Figures 55, 56
Index Fungorum number: IF860778

Figure 55.
Basidiomata and microscopic structures of Stereum yunnanense (holotype). (a), (b) Section of hymenophore. (c), (d) The back of the basidiomata. (e) Basidiospores. (r) Basidia and basidioles. (g) Cystidia. (h) Section of hymenium. Scale bars: (a), (c) = 1 cm, (b), (d) = 1 mm, (e)–(h) = 10 µm.
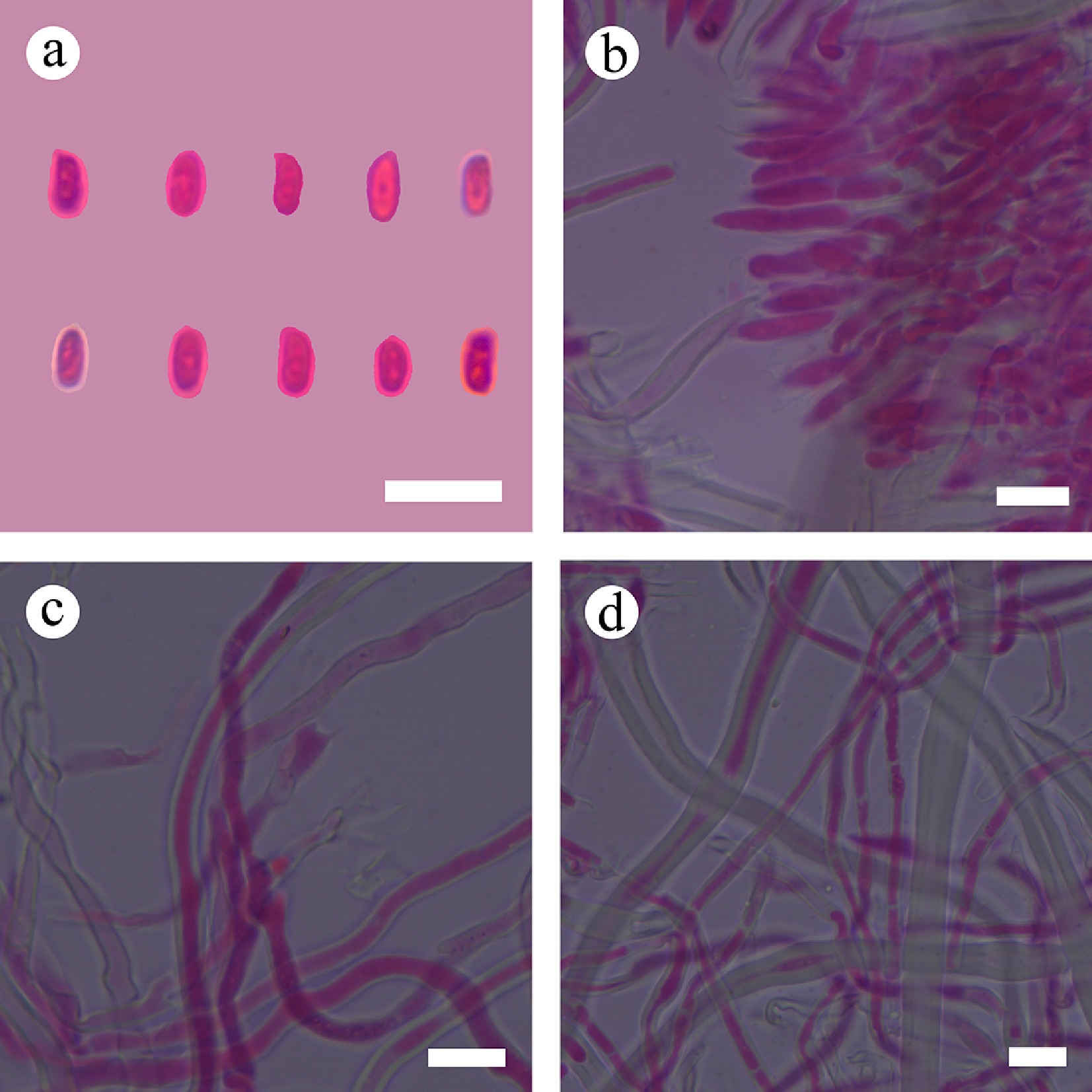
Figure 56.
Sections of hymenium of Stereum yunnanense (holotype). (a) Basidiospores. (b) Section of hymenium. (c) Basidia. (d) Skeletal hyphae. Scale bars: (a)–(d) = 10 µm.
Diagnosis – Stereum yunnanense differs from other species by the perennial, effused to pileate basidiomata, slightly cream to cream hymenial surface, cream to brownish orange pileal surface with short tomentum, ellipsoid basidiospores (4.7–8 × 2.4–3.2 µm).
Etymology – referring to the locality province (Yunnan) of the type specimen.
Type – China, Yunnan Province, Puer City, Jingdong County, Wuliang Mountain, 100°29' E, 24°38' N, 1,800 m asl, on angiosperm trunk, leg. C.L. Zhao, 5 October, 2017, CLZhao 4227 (SWFC).
Description – Basidiomata perennial, effused to pileate, separate from the substrate, coriaceous, without odor or taste when fresh. Pileus projecting up to 3.5 cm, 4.5 cm wide, and 1 mm thick at center. Hymenial surface smooth, slightly cream when fresh, becoming cream, sterile margin brownish orange, distinct, up to 1 mm wide. Pileal surface short tomentum, cream when fresh, becoming cream to brownish orange, in concentric bands or completely to expose underlying cutis. Sterile margin blunt, lightly orange, up to 1 mm wide.
Hyphal system dimitic; generative hyphae bearing simple septa, colorless, thick-walled, smooth, moderately branched, 2.5–3 µm in diameter. Skeletal hyphae dominant, colorless, distinctly thick-walled, smooth, unbranched, straight, with scattered secondary simple septa, 4.5–10 µm in diameter, IKI–, CB–; tissues unchanged in KOH. Cystidia dominant, cylindrical to fusiform, colorless, 14–21 × 3.5–5 µm. Basidia cylindrical, with four sterigmata and a basal simple septum, 12–15 × 3.5–4.5 µm; basidioles numerous, in shape similar to basidia but smaller. Basidiospores ellipsoid, colorless, thin-walled, smooth, amyloid, CB–, (4.6–)4.7–6(–8) × (2.2–)2.4–3.2(–3.5) µm, L = 5.2 µm, W = 2.84 µm, Q = 1.72–2.06 (n = 90/3).
Additiional material examined – China, Yunnan province, Puer City, Xieqipo Park, 101°5' E, 24°0' N, 1,302 m asl, fallen angiosperm branch, leg. C.L. Zhao, 1 October, 2017, CLZhao 3420; Jingdong County, Wuliang Mountain, 100°29' E, 24°38' N, 1,800 m asl, on angiosperm trunk, leg. C.L. Zhao, 5 October, 2017, CLZhao 4317 (SWFC).
GenBank accession numbers – CLZhao 4227 ITS: MT214932, LSU: PV384146, mtSSU: PV283194, rpb2: PV648925; CLZhao 3420 ITS: MT214929, LSU: PV384147, mtSSU: PV283195, rpb2: PV648926; CLZhao 4317 ITS: MT214930, mtSSU: PV283196
Notes – Phylogenetic analysis ( Fig. 6) indicated that Stereum yunnanense grouped within Stereum, sister to S. lobatum. However, morphologically, S. lobatum differs from S. yunnanense by having pseudoacanthophyses (16–27 × 2.5–3 µm), and subcylindrical basidiospores (5.2–7 × 2–3 µm) [ 267] . Thus, based on morphology and phylogenetic analysis, we concluded our collection is a new species, Stereum yunnanense.
Note 68 Xylobolus P. Karst.
Index Fungorum number: IF18770
Type species – Xylobolus frustulatus (Pers.) P. Karst.
Notes – Xylobolus is a small, globally distributed wood-inhabiting genus classified within family Stereaceae (Russulales) [ 268] . The type species of the genus is X. frustulatus, a distinctive corticioid species that grows on dead or decaying oak wood, and forms perennial basidiomata, which crack into small, angular polygons [ 269] . Another species X. subpileatus which has been widely studied, causes white rot in angiosperms, and grows mainly in southeastern USA, with a smaller distribution. The basidiomata of X. subpileatus are perennial, and acanthocystidia and can be observed in its hymenium under microscopic examination; this species was described from South Carolina by Berkeley & Curtis [ 266] , but it has since been reported from all continents except Antarctica. In Europe, X. subpileatus is mostly distributed in southern regions, and is considered a rare species in several countries [ 266] . There are few taxonomic studies on Xylobolus, although the chemical composition of Xylobolus has been studied [ 269] .
Note 69 Family Terrestriporiaceae Y.C. Dai, B.K. Cui, F. Wu, Y. Yuan & Jia J. Chen 2020
Index Fungorum number: IF834539
Type genus – Terrestriporia Y.C. Dai, B.K. Cui, F. Wu, Y. Yuan & Jia J. Chen
Notes – Terrestriporiaceae was established to accommodate Terrestriporia in order Russulales, based on its distinct lineage in the phylogenetic analysis [ 21] . The family is characterized by annual, resupinate basidiomata, a poroid hymenophore, monomitic hyphal structure, mostly simple septate generative hyphae, but occasionally with clamp connections, presence of gloeoplerous hyphae, and hyaline, thin-walled, smooth, amyloid basidiospores. The combined phylogeny of ITS, nLSU, rpb1, rpb2, and tef1- α sequence data, demonstrated that Terrestriporiaceae is related to Albatrellaceae, but showed no affinity to the other families of Russulales. Albatrellaceae includes mycorrhizal species that are mostly stipitate [ 21] .
Note 70 Terrestriporia Y.C. Dai, B.K. Cui, F. Wu, Y. Yuan & Jia J. Chen
Index Fungorum number: IF834540
Type species – Terrestriporia alba Y.C. Dai, B.K. Cui, F. Wu & Jia J. Chen
Notes – Terrestriporia was established by Wu et al. [ 21] and typified by T. alba. It is characterized by annual, resupinate basidiomata, poroid and white hymenophore, a monomitic hyphal structure, mostly simple septate generative hyphae, but occasionally with clamp connections, with gloeoplerous hyphae and gloeocystidioles, and hyaline, thin-walled, smooth, amyloid, acyanophilous basidiospores [ 21] . Terrestriporia resembles Anomoporia Pouzar and Anomoloma Niemelä & K.H. Larss. by sharing annual and resupinate basidiomata, a monomitic hyphal structure, and hyaline, thin-walled, smooth and amyloid basidiospores [ 16] , but the latter two genera have clamp connections only, lack gloeoplerous hyphae and cystidioles, and belong to Amylocorticiales [ 21] .
Family Wrightoporiaceae Jülich 1982
Index Fungorum number: IF80052
Type genus – Wrightoporia Pouzar
Note 71 Wrightoporia Pouzar
Index Fungorum number: IF18750
Type species – Wrightoporia lenta (Overh. & J. Lowe) Pouzar
Notes – The type species of Wrightoporia is W. lenta [ 92] . According to the recent studies, Wrightoporia s.str. is distinguished by an annual growth pattern, resupinate to effused-refluxed basidiomata, soft and cottony at fresh condition, membranous to cottony at dry condition, rounded to angular pores, mostly having rhizomorphs margins, finely asperulate basidiospores, mostly found on dead logs of gymnosperms, rarely on angiosperms. There are six species distributed in Asia: W. austrosinensis Y.C. Dai, W. avellanea (Bres.) Pouzar, W. borealis Y.C. Dai, W. lenta, W. srilankensis Y.C. Dai and Yuan Yuan, and W. subavellanea Jia J. Chen and B.K. Cui [ 10, 44, 270] . Cysidia are absent in most of these species, however, cystidia and cystidiols are present in W. austrosinensis [ 173] . Wrightoporia is a large and diverse genus of wood decaying tropical polypores and has been shown to be polyphyletic [ 45] . The systematic position of the genus has been in flux, initially it was placed in Wrightoporiaceae, and later transferred to Hericiaceae [ 259] . According to Index Fungorum (2025) and recent studies, the genus has been placed in Bondarzewiaceae [ 1, 5, 121] . The generic circumscription has recently been revised and placed it into family Wrightoporiaceae [ 10, 270] . Interestingly, in a recent phylogenetic analysis, W. austrosinensis formed an independent lineage outside of the genus [ 270] . A similar phylogenetic position of W. austrosinensis outside of Wrightoporia was observed by Hussain et al. [ 12] . Perhaps this species should be included either in Amylosporus or be treated as an independent genus [ 12] .
Molecular phylogeny
Russulales phylogeny based on the ITS + nLSU + mtSSU + rpb2 + tef1- α sequences data ( Fig. 1)
-
Note 72 Dentipellopsis Y.C. Dai & L.W. Zhou
Index Fungorum number: IF564151
Type species – Dentipellopsis dacrydiicola Y.C. Dai & L.W. Zhou
Notes – Dentipellopsis was erected to accommodate a newly described species D. dacrydicola by Zhou & Dai [ 30] . The genus is characterized by annual, resupinate basidiomata, hydnoid hymenophore, a monomitic hyphal system; generative hyphae with clamp connections, and distinctly thick-walled, densely echinulate, strongly amyloid basidiospores [ 30] . Dentipellopsis is similar to Dentipellis in macro-morphology but differs from the latter by its distinctly thick-walled and cyanophilous basidiospores. Furthermore, the two genera are not closely related, and Dentipellopsis was isolated from all accepted families in Russulales in the phylogeny using ITS and nLSU rDNA sequence data. Therefore, its family level taxonomic position is uncertain, and Zhou & Dai [ 30] named this clade as Dentipellopsis family for convenience.
Note 73 Gloeohypochnicium (Parmasto) Hjortstam
Index Fungorum number: IF15544
Type species – Gloeohypochnicium analogum (Bourdot & Galzin) Hjortstam
Notes – Gloeohypochnicium (Russulales) is typified by G. analogum, and it is characterized by the resupinate basidiomata with smooth to tuberculate hymenophore, a monomitic hyphal system with clamps on generative hyphae, the numerous cylindrical, sinuous gloeocystidia, and globose to ellipsoid, thick-walled, warted basidiospores [ 1, 5] . Based on MycoBank (2025) and Index Fungorum (2025), Gloeohypochnicium has three species, G. analogum, G. versatum (Burt) Tellería, M. Dueñas, Melo & M.P. Martín, and G. yunnanense Lu Wang & C.L. Zhao. The taxonomic status of the genus is unclear, and it was only placed in order Russulales, and it is a mystery genus, and lacks new taxa in the genus for nearly a decade [ 1, 5, 6] . The phylogeny of Gloeohypochnicium is ambiguous due to a lack of molecular evidence and morphological data. The phylogenetic tree inferred from ITS + nLSU sequences revealed that Gloeohypochnicium grouped within order Russulales [ 20] .
Note 74 Haloaleurodiscus N. Maek., Suhara & K. Kinjo
Index Fungorum number: IF28938
Type species – Haloaleurodiscus mangrovei N. Maek., Suhara & K. Kinjo
Notes – Haloaleurodiscus was described from Japanese mangrove forests with one species, H. mangrovei. The genus is morphologically characterized by having resupinate basidiomata, nodose-septate hyphae, sulphoaldehyde-positive gloeocystidia, dendrohyphidia and amyloid basidiospores with minute warts. These morphological characters are similar to those of Aleurodiscus s. lat., but H. mangrovei differs from Aleurodiscus primarily in occurring in white pocket-rot and is not closely related based on ribosomal DNA sequence analyses. Molecular data suggested that this species is phylogenetically placed in the root of the ‘Peniophorales’ clade [ 271] .
Note 75 Neoalbatrellus Audet
Index Fungorum number: IF511206
Type species – Neoalbatrellus caeruleoporus (Peck) Audet
Notes – Neoalbatrellus was separated from Albatrellus s. l. by Audet [ 96] , and typified by N. caeruleoporus. It was distinguished from Albatrellus s.s. by several peculiar features such as the unique hymeniderm, and blue, black, and brown color of the basidiomata [ 96, 272] . Four species, N. caeruleoporus, N. odorus Yuan Y. Chen & B.K. Cui, N. subcaeruleoporus Audet & B.S. Luther, and N. yasudae (Lloyd) Audet, have been recorded [ 96, 272, 273] . The phylogenetic tree based on ITS and nLSU dataset revealed that the genera Albatrellopsis, Polypus, Xeroceps, Neoalbatrellus, and Xanthoporus clustered within separate lineages with high branch supports [ 273] . However, Albatrellopsis differs from Neoalbatrellus by its large, convex and highly confluent basidiomata, clamped generative hyphae and small, amyloid basidiospores [ 96, 274, 275] . Polyporus is characterized by pale yellow petaloid pileus sharing a common base, its fragrant odor when fresh, and thin-walled, non-amyloid basidiospores [ 96, 274, 275] . Xeroceps showed unique fuliginous to black pileus, clamped generative hyphae and larger basidiospores than Neoalbatrellus [ 96, 274, 275] , whereas Xanthoporus is distinguished from Neoalbatrellus by its yellowish-ochraceous to yellowish-brown pileus, clamped generative hyphae, and thin-walled basidiospores [ 96] . Albatrellopsis differs from Neoalbatrellus by its large, convex and highly confluent basidiomata, clamped generative hyphae and small, amyloid basidiospores [ 96, 275] .
Note 76 Perplexostereum Ryvarden & S. Tutka
Index Fungorum number: IF807398
Type species – Perplexostereum endocrocinum (Berk.) Ryvarden & Tutka
Notes – Ryvarden [ 276] proposed Perplexostereum for Stereum endocrocinum Berk. Like Echinodontium and Laurilia. Perplexostereum develops large, pileate basidiocarps, ornamented amyloid basidiospores, and inhabits gymnosperms but differs in lacking encrusted cystidia. Phylogenetic tree, based on sequence data of the nuc rDNA ITS1-5.8S-ITS2 (ITS) and D1–D2 domains of nuc 28S rDNA (28S), Perplexostereum formed a distinct clade from Echinodontiaceae and Bondarzewiaceae within Russulales [ 81] .
-
Fungi include mushrooms, molds, lichens, yeasts, and zoosporic forms that occur as free-living or symbiotic organisms in every ecosystem on Earth, and about 155,000 species of fungi have been described, and possibly millions more remain to be named [ 2, 7, 121] . Fungal systematists have made remarkable progress toward resolving the major branches of the phylogeny, although some deep nodes have proven recalcitrant, in which fungal taxonomists steadily describe about 3,000 new species per year, and fungal molecular ecologists routinely detect many thousands of unidentifiable ‘dark fungi’ through metagenomic analyses [ 277] . In recent years, numerous taxa of wood-inhabiting fungi have been reported and documented worldwide [ 2, 7, 20, 54, 121, 134] . Over the past ten years, global studies have explored the diversity, ecology, and systematics of wood-inhabiting fungi, with different studies emphasizing various taxonomic groups [ 37, 52, 134, 220] . However, many fungi in unexplored geographic areas are threatened by climate change, forest expansion, deforestation, and human interactions. Therefore, numerous extant taxa may become extinct before they are discovered [ 7] . Consequently, it is now urgent to recognize and conserve fungal biodiversity as it may result in new discoveries [ 2] . Additionally, the diversity of wood-inhabiting fungi in the Yunnan-Guizhou Plateau of China holds significant importance.
Fungi include model systems for cell and molecular biology and they represent one of the few clades that has evolved complex multicellularity, which they have done at least eight times during their billion-year history [ 277] . Fruiting bodies (sporocarps, sporophores or basidiomata) of Russulales are among the most complex structures produced by fungi and these sexual reproductive organs facilitate efficient spore dispersal. While in simple Basidiomycota have spores that are born on naked basidia, mushroom-forming fungi have evolved complex three-dimensional structures that offer tremendous advantages in effective spore dispersal [ 278] . Ancestral basidiomata of the Agaricomycotina were probably crust-like, ‘resupinate’ forms [ 279, 280] , which then evolved into increasingly more complex forms, including derived ‘pileate-stipitate’ types, which are differentiated into a cap, stipe, and hymenophore (spore-bearing surface) [ 280] . Extensive mycological studies have confirmed that the species of Russulales exhibit pileate, clavarioid, and gasteroid basidiomata [ 1] . The diversity of Russulaceae is extremely rich, including discoid, claviform, resupinate-effuse and gasteroid basidiomata, with smooth, pleated, poroid, and odontoid hyphal surface [ 17, 281] . The distribution of corticioid basidiomata in Russulales was early studied by Larsson [ 45] . According to He et al. [ 1] , the resupinate basidiomata are present in seven families Echinodontiaceae, Hericiaceae, Peniophoraceae, Russulaceae ( Boidinia and Gloeopeniophorella were separate from Gloeocystidiellum s.l.), Stereaceae and Terrestriporiaceae. Surprisingly, in the present summary, the resupinate basidiomata type of Russulales, are distributed in 13 branches (families), In addition to the above families, Albatrellaceae, Aleurocystidiellaceae fam. nov., Auriscalpiaceae, Bondarzewiaceae ( Laurilia and Lauriliella), Gloeocystidiellaceae, Gloeodontiaceae fam. nov., and Wrightoporiaceae are included. Meanwhile, In the present study, we propose a new genus and two new families, and we introduce 26 new species with resupinate basidiomata. These taxa belong to six families, Amylostereaceae, Gloeocystidiellaceae, Gloeodontiaceae, Hericiaceae, Peniophoraceae, and Stereaceae, based on morphology, molecular phylogenetic analyses, and divergence time estimates.
Aleurocystidiellaceae
-
Lemke [ 101] established Aleurocystidiellum which could not be assigned to any recognized family of Russulales, and its family level classification was treated as incertae sedis. Subsequently, phylogenetic studies supported the monophyletic status of only a small group of the genus Aleurocystidiellum, and it was shown that the genus corresponds to a family of its own infered from ITS + 28S sequence data [ 10, 21, 45, 46] . Furthermore, Aleurocystidiellum is similar to Aleurodiscus (Stereaceae) by sharing the characters of discoid to effused-reflexed basidiomata, moniliform gloeocystida and verrucose basidiospores. However, species of Aleurodiscus can be distinguished from Aleurocystidiellum by the presence of numerous acanthocystidia. In the present study, for phylogenetic analyses, Aleurocystidiellum species were placed in an alignment of ITS and 28S sequences of Stereaceae and allied fungi, as previously generated by Wu et al. [ 103] , Dai & He [ 80] , Phookamsak et al. [ 245] , Rajchenberg et al. [ 46] , and Xu et al. [ 87] . The phylogram depicts an overall topology of family Stereaceae ( Fig. 6), and order Russulales ( Fig. 1), confirming that Aleurocystidiellum cannot be placed into any recognized family, including Stereaceae. The analysis of divergence times was also carried out ( Fig. 2), which reveals the divergence time of the Aleurocystidiellum clade and Hericiaceae at 140.94 Mya with a 95% highest posterior density (HPD) of 87.46–196.71 Mya, 95% HPD, which is similar to the differentiation time of each family in Russulales. Thus, the family name Aleurocystidiellaceae was proposed.
Echinodontiaceae
-
The family Echinodontiaceae includes four genera, Amylostereum, Echinodontiellum, Echinodontium, and Larssoniporia. Except for Larssoniporia, which has a poroid basidiomata, the other three genera have corticioid basidiomata [ 10] . The divergence time of family Echinodontiaceae and the genus Subulicystidiella are at 150.57 Mya (113.56–190.2 Mya, 95% HPD), and 114.84 Mya (125.46–202.89 Mya, 95% HPD), based on ITS + nLSU + rpb2 + tef1- α sequence data ( Fig. 2). Inference from the ITS + nLSU + mtSSU + rpb2 + tef1- α ( Fig. 1) sequence data showed that Subulicystidiella formed a monophyletic clade with statistical support of 87/1.00, and the ITS + nLSU ( Fig. 4) sequence data showed that Subulicystidiella formed an alone clade with statistical support of 100/100. These indicate that S. murina is distinct from all other species sampled in the phylogenetic analysis. However, due to the limited taxa of Subulicystidiella, the topology may be influenced by taxon sampling. Meanwhile, morphologically, Subulicystidiella was similar to the genus Amylostereum (Echinodontiaceae) by its dimitic hyphal system, thick-walled, yellowish brown subulate, encrusted cystidia, and smooth basidiospores, but the latter has effused-reflexed basidiomata, first as small colonies, later confluent, azonate, pale mouse gray to grayish-brown hymenophore, and distinctly amyloid basidiospores [ 139] . In addition, the type genus Echinodontium of the family Echinodontiaceae is characterised by annual to perennial, resupinate, effused-reflexed to pileate-sessile basidiomata, dimitic hyphal system, generative hyphae with or without clamp-connections, skeletal hyphae inamyloid or dextrinoid, gloeocystidia present or incrusted cystidia absent or present, with smooth basidiospores [ 1] . The characteristics of the new genus Subulicystidiella are consistent with those of Echinodontiaceae. Thus, we introduce Subulicystidiella as a new genus and belongs to this family Echinodontiaceae based on phylogenetic analyses and morphological characteristics.
Gloeocystidiellaceae
-
The family Gloeocystidiellaceae was established by Jülich [ 105] , and typified by the genus Gloeocystidiellum which was established in 1931 based on morphological classification, including all monophyletic species characterized by gloeocystidia and amyloid basidiospores. All species in this group are corticioid basidiomata and include only the genus Gloeocystidiellum, and the genus is not recovered as monophyletic based on the phylogenetic evidence. The situation in Gloeocystidiellum s. lat. is similar to Aleurodiscus s. lat., but its lineages have wider distributions in Russulales with only certain species nested within Stereaceae [ 3, 45, 89, 149, 151] . A complete taxonomy and phylogenetic study of the family focusing on the phylogenetic relationships at generic level is urgently needed. We attempted to classify Gloeocystidiellum into Stereaceae, but from molecular evidence did not support this view. The result also confirms that Gloeocystidiellum is polygeneic, and does not place it in any recognized family of Russulales. The divergence time of family Gloeocystidiellaceae is at 133.39 Mya with a 95% highest posterior density (HPD) of 79.96–186.84 Mya, 95% HPD. based on ITS + nLSU + rpb2 + tef1- α sequence data ( Fig. 2). Therefore, based on the divergence times and the molecular phylogenetic relationship, family Gloeocystidiellaceae accommodates Gloeocystidiellum s. str. based on the type sequences.
Peniophoraceae
-
Previous studies of family Peniophoraceae (Russulales) have shown that Vararia was closely related to Asterostroma, Baltazaria, Dichostereum, and Scytinostroma [ 33, 34, 43, 172, 219, 221] . In the present study, the phylogenetic analysis of Vararia and other related genera, inferred from the dataset of ITS + nLSU + mtSSU + rpb2 + tef1- α ( Fig. 1), reveals that Vararia forms different clades, and three genera Baltazaria, Dichostereum, and Peniophora are distributed within Vararia. According to the lineage delimitation, some genera are morphologically well-circumscribed, but some have wide ranges in several aspects, and most genera exhibit their specific characteristics. Asterostroma is strongly supported to a single clade characterized by asterosetae and ornamented basidiospores. Baltazaria and Dichostereum are both monotypic, in which the former is characterized by fibriform skeletal hyphae without simple septa and smooth basidiospores, the latter is characterized by dichohyphae and ornamented basidiospores. Scytinostroma s. str. comprises species with fibriform skeletal hyphae, abundant gloeocystidia and smooth basidiospores. Vararia s. str. remains a complex assemblage of species with diverse morphology characterized by dichohyphae, gloeocystidia and smooth basidiospores. At present, we prefer to treat it in a rather broad sense to avoid more lineages and new names. Peniophora s. str. is characterized by the numerous cystidia encrusted with crystals and smooth basidiospores.
Recalcitrant nodes notwithstanding, reconstructing most of the major branches of the fungal tree of life appears to be a tractable problem but, the goal of achieving a complete catalog of fungi is aspirational at best [ 277] . Most major branches of the fungal tree of life have been resolved and the Russulales will follow as more barcodings are sequenced. The harder part, for which no immediate solution is apparent, will be to resolve the fine branches of the phylogeny in detail within Russulales and translate their structure into an accessible classification for different families of Russulales.
The present study addressed and resolved several important taxonomic and phylogenetic issues, and provides a solution framework for further studies on the phylogeny of wood-inhabiting fungi in Russulales. Additional taxa in Russulales should be included into the phylogenetic analyses to improve the generic delimitation with better and more accurate taxonomic resolution. The present study aims to fill the knowledge gap concerning wood-inhabiting fungi within Russulales by introducing new taxa, and providing detailed morphological characteristics, comprehensive descriptions and phylogenetic analyses of these fungal species. In addition, this work contributes to the enrichment of fungal diversity in Asia, particularly expanding the knowledge of Chinese fungal diversity. Further, we suggest that future investigations on the species diversity of Russulales should be prioritized in habitats in underexplored regions and hosts such as monocots in subtropical and tropical areas to uncover the hidden fungal diversity.
-
No ethical statement was reported.
-
The authors confirm contribution to the paper as follows: study conception and design: Zhao CL; data collection: Zhao CL, Deng YL, Chen M, Zhang SC; investigation: analysis and interpretation of results: Deng YL, Chen M, Zhang SC, Wang KS, Liu WT, Qiu YH, Dou YT, Wijesinghe SN, Zhou HM, Jabeen S, Liu XF, Zhao CL.; draft manuscript preparation: Deng YL, Chen M, Zhang SC. Wang KS, Liu WT, Qiu YH, Dou YT, Wijesinghe SN, Zhou HM, Jabeen S, Liu XF, Zhao CL. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
The research was supported by the Forestry and Grass Science and Technology Innovation Joint Project of Yunnan Province (Project No. 202404CB090008), High-level Talents Program of Yunnan Province (YNQR-QNRC-2018-111), Modern Industry School of Edible-fungi, Southwest Forestry University (SYJ25), the Science Foundation of Education Department of Yunnan Province (2025Y0845), Science and Technology Projects of Yunnan Universities Serving Key Industries (FWCY-BSPY2025093), Southwest Forestry University Graduate Research Innovation Fund Project (YJS-KCJJ-2025-03), and Yunnan Province College Students Innovation and Entrepreneurship Training Program (Project No. S202510677111).
-
The authors declare that there is no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Deng Y, Chen M, Zhang S, Wang K, Liu W, et al. 2026. Notes, taxonomy, and phylogeny of wood-inhabiting fungi in Russulales.
Mycosphere 17: e003 doi: 10.48130/mycosphere-0026-0003


|