









-
Despite rapid growth in documented fungal diversity—with over 160,000 species known (
https://nmdc.cn/fungalnames , accessed on March 6, 2025) and thousands added annually[1−4]—known species represent only a fraction of estimates, keeping species discovery and evolutionary reconstruction key priorities in mycology[5−8]. The origin and evolution of organisms are among the most important issues in life sciences, while molecular-clock analyses and biogeographic studies aim to reconstruct the origin, speciation, and distribution patterns[9−11]. While molecular clock and biogeographic analyses help reconstruct these evolutionary contexts, fungal studies still lag behind animals and plants due to challenges like cryptic morphology, insufficient sampling, and scarce fossil records[12−16]. Fortunately, significant progress has been made in the study of macrofungi such as the Agaricomycetes, Hymenochaetales, and Boletaceae[17−21].Wood-inhabiting fungi, as an important group of macrofungi, have rich species diversity, nutritional types, morphological variety, and are widely distributed in forest ecosystems. They play multiple roles, such as saprotrophy, parasitism, and symbiosis, with significant ecological and economic values[22−31]. In recent years, a series of studies have addressed the species diversity and evolutionary history of wood-inhabiting fungi[32−35], including the genus Trichaptum s.l.[36−40].
The genus Trichaptum s.l., typified by Trichaptum perrottetii (Lév.) Ryvarden (basionym: Polyporus trichomallus Berk. & Mont.), was established by Murrill in 1904[41]. It is characterized by annual to perennial, mostly pileate basidiomata with a hispid to adpressed tomentose abhymenial surface, variable hymenophore (irpicoid, lamellate, or poroid), with brownish to purplish hymenial surface, duplex context, mainly a dimitic hyphal system with clamped generative hyphae, predominantly cylindrical basidiospores, and the presence of cystidia[39,42,43]. Trichaptum s.l. species are widely distributed in forest ecosystems, functioning as dominant white-rot decomposers. Ecological surveys have highlighted their prevalence in deadwood communities; for instance, Trichaptum abietinum was identified as the most frequent species in recent field studies, recording 145 fruit-body occurrences across 571 sampled logs, significantly outperforming other wood-inhabiting fungi[44,45]. Beyond their ecological dominance, these species possess considerable economic and biotechnological potential. While traditionally noted for the medicinal values of species such as T. abietinum, T. biforme, and T. byssogenum[46,47], recent research has expanded their utility into sustainable materials. Trichaptum biforme has been successfully evaluated as a novel mycelium source for fabricating bio-foams, offering a competitive, biodegradable alternative to commercial plastic packaging[48]. Currently, Trichaptum s.l. is divided into two families, seven genera, and 49 species based on morphological characters, phylogenic analyses, and divergence times estimation[39].
The family Hirschioporaceae includes four genera: Hirschioporus, Nigrohirschioporus, Pallidohirschioporus, and Perennihirschioporus. The family Trichaptaceae consists of a single genus, Trichaptum, while two other genera, Pseudotrichaptum and Podocarpioporus, are treated as incertae sedis within the order Hymeonchaetales[39]. Resolving their phylogenetic relationship and taxonomic status is of importance for future development and utilization of resources. Furthermore, with the increase in collection sites and samples, the number of new species will continue to grow. Although Zhou et al. conducted a preliminary study on the origin and divergence time of Trichaptum s.l.[39], an in-depth study of this group is urgently needed.
Determining the spatial-distribution ability of species is crucial for understanding the environmental factors influencing them and their biodiversity. Niche modeling is a useful tool for assessing the potential geographic distribution of species and has been widely applied in the fields of biogeography, ecology, and evolutionary phylogenetics[49−51]. Many statistical models are available to simulate species distribution types and the diversity of species' spatial patterns[52−54]. The Maximum Entropy (Maxent) model is widely used to predict the potential distribution of species based on collection data and environmental information, with some advantages, including strong performance even when data are incomplete, user-friendliness, the ability to perform well with limited occurrence data, and high simulation accuracy[55−57]. Previous studies have conducted habitat suitability predictions and explored the impact of environmental factors on the distribution of fungal groups[58−60], however, Trichaptum s.l. has not yet been carried out.
In this study, we used newly collected specimens to update the classification system, and reconstruct the evolutionary history of Trichaptum s.l., combined with multi-gene phylogenetic analyses, morphological characteristics, and molecular-clock analyses. Based on this revised taxonomic framework, a global occurrence dataset was compiled for each genus. Ecological niche modeling (Maxent) was employed to quantify the climatic niche of each lineage and to identify the key environmental drivers of their respective geographical distributions, providing a clearer understanding of their unique adaptive strategies.
-
In this study, 28 new specimens of Trichaptum s.l. were collected from eight countries, including China, Brazil, Bolivia, Costa Rica, Ecuador, French Guiana, Jamaica, and the USA. All the studied specimens were deposited in the Fungarium of the Institute of Microbiology, Beijing Forestry University (BJFC, China), and the private herbarium of Josef Vlasák (JV, Czech Republic).
Morphological studies
-
Macromorphological descriptions are based on field collection records and voucher observations of basidiomata. The micromorphological data were obtained by dissecting dried specimens and observing them under a stereomicroscope (Nikon E80i microscope, Tokyo, Japan), following previous studies[39,61], and line drawings were made with the aid of a drawing tube. Special color terminology used follows Anonymous[62] and Petersen[63]. Specimen sections were prepared and mounted in Melzer's reagent, Cotton Blue, and 5% KOH.
The following abbreviations are used: KOH = 5% potassium hydroxide; CB = Cotton Blue; CB+ = cyanophilous in Cotton Blue; CB– = cyanophilous in Cotton Blue; IKI = Melzer's reagent; IKI– = neither amyloid nor dextrinoid in Melzer's reagent; L = mean basidiospore length (arithmetic average of basidiospores); W = mean basidiospore width (arithmetic average of basidiospores); Q = variation in the L/W ratios between specimens studied; n (a/b) = number of basidiospores (a) measured from the given number of specimens (b), 'n' presenting basidiospore size variation, 5% of measurements were excluded from each end of the range and these values are given in parentheses. The texture was evaluated using the following scale: Grade I (Soft corky), tissue is soft, compressible, and easily deformed; Grade II (Coriaceous), tissue is tough, leather-like, and pliable; Grade III (Hard corky), tissue is firm, resists deformation, but retains some elasticity; and Grade IV (Woody), tissue is rigid, non-deformable, and highly lignified.
DNA extraction, polymerase chain reaction, and sequencing
-
Total DNA was extracted from the dried specimens using the standard protocol of the rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd, Beijing, China), with modifications based on a previous study by Zhou et al.[39]. Five gene fragments, including internal transcribed spacer (ITS), nuclear large subunit rDNA (nLSU), translation elongation factor 1 (TEF1), mitochondrial small subunit rDNA (mt-SSU), and nuclear small subunit rDNA (nuc-SSU), were amplified using polymerase chain reaction (PCR) to obtain target products. The selected target fragments and their primer sequences are listed in Table 1. Amplification was performed in a 30 μL reaction system (2× EasyTaq® PCR SuperMix: ultra pure water : forward primer : reverse primer : template DNA = 15:12:1:1:1). PCRs were carried out using S1000™ Thermal Cycler (Bio-Rad Laboratories, California, USA), following the procedure by Zhou et al.[39]. The PCR products were sequenced on the ABI-3730-XL DNA analyzer (Applied Biosystems, Foster City, CA, USA) by Beijing Genomics Institute (Beijing, China) with the sample primers in Table 1. Finally, all the newly generated sequences were deposited in the NCBI GenBank database, and the accession numbers are listed in Table 2.
Table 1. Primer information for five gene fragments in this study.
Sequence Primers Primers sequence (5'–3') Ref. ITS ITS4 TCC TCC GCT TAT TGA TAT GC [64] ITS5 GGA AGT AAA AGT CGT AAC AAG G nLSU LR0R ACC CGC TGA ACT TAA GC [65] LR7 TAC TAC CAC CAA GAT CT mt-SSU MS1 CAG CAG TCA AGA ATA TTA GTC AAT G [64] MS2 GCG GAT TAT CGA ATT AAA TAA C TEF1 983F GCY CCY GGH CAY CGT GAY TTY AT [66] 1567R ACH GTR CCR ATA CCA CCR ATC TT nuc-SSU NS1 GTA GTC ATA TGC TTG TCT C [64] NS4 CTT CCG TCA ATT CCT TTA AG Table 2. Taxa information and GenBank accession numbers used in this study.
Family Species Voucher Locality GenBank A accession no. Ref. ITS nLSU mt-SSU TEF1 nuc-SSU Coltriciaceae Coltricia perennis Cui 10319 China KU360687 KU360653 — KY693935 — [104] Coltriciaceae Coltricia dependens Dai 10944 China KY693737 KY693757 — — — Genbank Neoantrodiellaceae Neoantrodiella gypsea Cui 10372 China KT203290 MT319396 MT326567 — — [105] Neoantrodiellaceae Neoantrodiella gypsea Yuan 5589 China KT203292 KT203313 — — — Genbank Neoantrodiellaceae Neoantrodiella thujae Dai 5065 China KT203293 MT319397 MT326568 — — [105] Nigrofomitaceae Nigrofomes melanoporus JV 1704/39 Costa Rica MF629835 MF629831 — — — [106] Nigrofomitaceae Nigrofomes melanoporus JV 1607/82 Costa Rica MF381027 — — — — [36] Nigrofomitaceae Nigrofomes sinomelanoporus Cui 5277 China MF629836 MT319398 — — — [105−106] Nigrofomitaceae Nigrofomes sinomelanoporus Dai 16286 China MF629839 — — — — [106] Hirschioporaceae Hirschioporus abietinus Cui 2667 China OQ449096 OQ449033 — OQ831431 OQ449408 [39] Hirschioporaceae Hirschioporus abietinus CBS 221.53 France MH857167 MH868703 — — — [107] Hirschioporaceae Hirschioporus abietinus Dai 1 Finland OQ449044 — — — — [39] Hirschioporaceae Hirschioporus abietinus Dai 12337 China OL504712 OL477386 OQ517075 OQ831434 OQ449409 [39] Hirschioporaceae Hirschioporus abietinus Dai 23792 China OQ449043 OQ449038 OQ517079 — OQ448998 [39] Hirschioporaceae Hirschioporus abietinus KUC20130719-05 Korea KJ668437 KJ668289 — — — [108] Hirschioporaceae Hirschioporus abietinus FSU 2734 Germany EU484271 — — — — [109] Hirschioporaceae Hirschioporus abietinus JV 0907/9 Czechia MF381023 — — — — [36] Hirschioporaceae "Hirschioporus abietinus" BFOTU23 Finland AM901859 — — — — [110] Hirschioporaceae "Hirschioporus abietinus" Cui 9627 China OQ449046 OQ449184 OQ517080 — OQ448999 [39] Hirschioporaceae "Hirschioporus abietinus" Dai 18909 China OQ449048 OQ449185 OQ449111 OQ831436 OQ449001 [39] Hirschioporaceae "Hirschioporus abietinus" Dai 23763 China OQ449131 OQ449195 OQ449116 — OQ449006 [39] Hirschioporaceae Hirschioporus acontextus Cui 11830 China OQ449138 OQ449198 OQ449120 — OQ449010 [39] Hirschioporaceae Hirschioporus acontextus Dai 19097 China OQ449140 OQ449199 OQ449122 — OQ449012 [39] Hirschioporaceae Hirschioporus acontextus Dai 23793 China OQ449141 OQ449200 OQ449123 OQ831439 OQ449013 [39] Hirschioporaceae Hirschioporus acontextus KUC20131001-03 Korea KJ668436 KJ668288 — — — [39] Hirschioporaceae Hirschioporus beijingensis Dai 18907 China OQ449142 OQ449201 OQ449124 OQ831440 OQ449014 [39] Hirschioporaceae Hirschioporus beijingensis Dai 23704 China OQ449143 OQ449202 OQ449125 OQ831441 OQ449015 [39] Hirschioporaceae Hirschioporus chinensis Cui 2488 China OQ449146 — OQ449126 OQ831442 OQ449016 [39] Hirschioporaceae Hirschioporus chinensis Dai 19110 China OQ449098 — — — — [39] Hirschioporaceae Hirschioporus chinensis Dai 19111 China OQ449099 — — — — [39] Hirschioporaceae Hirschioporus chinensis Dai 20264 China OQ449101 OQ449204 OQ449127 OQ831443 OQ449017 [39] Hirschioporaceae Hirschioporus chinensis Dai 21116 China OQ437348 OQ438001 OQ448978 OQ831444 — [39] Hirschioporaceae Hirschioporus chinensis Dai 23048 China OQ437349 OQ438002 OQ448979 OQ831445 OQ438032 [39] Hirschioporaceae Hirschioporus floridanus Dolliner 640 USA OQ437354 — — — OQ438036 [39] Hirschioporaceae Hirschioporus floridanus Dolliner 642 USA OQ437355 OQ438004 — — OQ438037 [39] Hirschioporaceae Hirschioporus floridanus MC1 Korea MW795372 — — — — Genbank Hirschioporaceae Hirschioporus fuscoviolaceus Dai 20988 Belarus OQ437357 OQ438006 OQ448983 OQ831449 OQ438039 [39] Hirschioporaceae Hirschioporus fuscoviolaceus Dai 21021 Belarus OQ437360 OQ438009 OQ448986 OQ831451 — [39] Hirschioporaceae Hirschioporus fuscoviolaceus JV 1610/32 Czechia MF381026 — — — — [36,39] Hirschioporaceae "Hirschioporus fuscoviolaceus" Cui 11870 China OQ437362 OQ438011 OQ448988 — OQ438042 [39] Hirschioporaceae "Hirschioporus fuscoviolaceus" Dai 10576 China OQ437365 — — OQ831453 OQ438043 [39] Hirschioporaceae "Hirschioporus fuscoviolaceus" Dai 22771 China OQ437368 OQ438013 — — OQ438044 [39] Hirschioporaceae "Hirschioporus fuscoviolaceus" V. Haikonen 25849 Finland MF319119 MF318985 — — — Genbank Hirschioporaceae Hirschioporus montanus Cui 17067 China OL470322 OL462836 OQ448990 OQ857930 OQ438048 [39] Hirschioporaceae Hirschioporus montanus Dai 4204 China OQ437376 OQ438018 OQ448996 OQ857936 OQ449151 [39] Hirschioporaceae Hirschioporus montanus Dai 22979 China OQ437372 OQ438015 OQ448991 OQ857931 OQ438049 [39] Hirschioporaceae Hirschioporus montanus Dai 23143 China OQ437374 OQ438016 OQ448993 OQ857933 OQ438051 [39] Hirschioporaceae Hirschioporus pubescens Dai 17064 China OQ437377 OQ438019 OQ448997 OQ857937 OQ449152 [39] Hirschioporaceae Hirschioporus pubescens Dai 23710 China OQ512026 OQ449059 OQ449169 OQ857938 OQ449153 [39] Hirschioporaceae Hirschioporus pubescens KA17-0228 Korea MN294864 — — — — [111] Hirschioporaceae Hirschioporus pubescens SFC95081517 Korea AF266680 — — — — [112] Hirschioporaceae Hirschioporus pubescens SFC96060811 Korea AF267644 — — — — [112] Hirschioporaceae Hirschioporus pubescens SFC96102811 Korea AF267645 — — — — [112] Hirschioporaceae Hirschioporus sp1 CCFC008387 Korea AF266677 — — — — [112] Hirschioporaceae Hirschioporus sp1 CBS 374.68 Canada AF266676 MH870875 AF408707 — — [107,112] Hirschioporaceae Hirschioporus sp1 UBCF20347 Canada KC581332 — — — — Genbank Hirschioporaceae Hirschioporus sp1 UBCF28393 Canada KP454016 — — — — Genbank Hirschioporaceae Hirschioporus sp2 CCFC01056 Canada AF267647 — — — — [107,112] Hirschioporaceae Hirschioporus sp2 CBS 375.68 Canada AF267648 MH870876 AF408716 — — [112] Hirschioporaceae Hirschioporus tianschanicus Dai 15944 China OQ437383 OQ449063 OQ449173 OQ857943 OQ449158 [39] Hirschioporaceae Hirschioporus tianschanicus Dai 19067 China OQ448960 OQ449067 OQ449177 OQ857947 OQ449162 [39] Hirschioporaceae Hirschioporus tianschanicus Dai 19086 China OQ448963 OQ449070 OQ449179 OQ857949 OQ449164 [39] Hirschioporaceae Hirschioporus tianschanicus KA16-1050 Kyrgyzstan MK351689 — — — — Genbank Hirschioporaceae Nigrohirschioporus bulbocystidiatus Ryvarden 42922 Costa Rica OQ448964 — — — — [39]
GenbankHirschioporaceae Nigrohirschioporus confertus JV 1407/75 Costa Rica MF380988 — — — — [36] Hirschioporaceae Nigrohirschioporus confertus JV 1504/101 Costa Rica MF380992 — — — — [36] Hirschioporaceae Nigrohirschioporus confertus JV 1504/32 Costa Rica MF380991 — — — — [36] Hirschioporaceae Nigrohirschioporus deviatus Ryvarden 46959 Venezuela OQ476081 — — — — [39] Hirschioporaceae Nigrohirschioporus durus Cui 11327 China OQ448965 OQ449071 OQ449180 OQ874781 OQ449165 [39] Hirschioporaceae Nigrohirschioporus durus Dai 12305 China OQ448969 OQ449073 — OQ857951 OQ449167 [39] Hirschioporaceae Nigrohirschioporus durus Dai 18419 Vietnam OQ448970 — — — — [39] Hirschioporaceae Nigrohirschioporus durus Dai 19497 Sri Lanka OQ448971 OQ449074 OQ449181 OQ857952 OQ449168 [39] Hirschioporaceae Nigrohirschioporus durus Dai 19640 Sri Lanka OQ476082 OQ504233 OQ512031 OQ857953 OQ512030 [39] Hirschioporaceae Nigrohirschioporus durus M1569 Japan LC327032 [113] Hirschioporaceae Nigrohirschioporus fissile TKC 145 Brazil MK973088 — — — — Genbank Hirschioporaceae Nigrohirschioporus fissile TKC 99 Brazil MK973089 — — — — Genbank Hirschioporaceae Nigrohirschioporus griseofuscus B3942 Brazil OQ448975 OQ438022 — OQ857957 OQ449106 [39] Hirschioporaceae Nigrohirschioporus griseofuscus JV 1808/103 French Guiana OQ448976 OQ438023 OQ449182 — — [39] Hirschioporaceae Nigrohirschioporus griseofuscus JV 1909/6 French Guiana OQ437343 OQ438024 — OQ857958 OQ449107 [39] Hirschioporaceae Nigrohirschioporus sector AS 2707 Brazil OQ437344 OQ438025 OQ449183 OQ857959 OQ449108 [39] Hirschioporaceae Nigrohirschioporus sector Dolliner 897 USA OQ437346 — — — — [39] Hirschioporaceae Nigrohirschioporus sector JV 1408/8-J Costa Rica OQ437347 OQ438027 — OQ857961 — [39] Hirschioporaceae Nigrohirschioporus sector JV 1808/108 French Guiana OQ453304 OQ453531 OQ453509 — OQ453485 [39] Hirschioporaceae Nigrohirschioporus keylargoensis JV 0904/66-J USA MF381009 PX747684 — PX766002 PX747671 [36] and present study Hirschioporaceae Nigrohirschioporus keylargoensis JV 2103/2-J USA PX741913 PX747685 — PX766003 PX747672 Present study Hirschioporaceae Nigrohirschioporus sp1 CF00108 Thailand OQ076496 — — — — [114] Hirschioporaceae Nigrohirschioporus sp1 NLB1300 Australia ON715817 — — — — Genbank Hirschioporaceae Nigrohirschioporus sp1 NLB1301 Australia ON715818 — — — — Genbank Hirschioporaceae Nigrohirschioporus submurinus Dai 18392 Vietnam OQ453305 OQ453532 — — OQ453486 [39] Hirschioporaceae Nigrohirschioporus trimiticus B696 Brazil OQ453308 OQ453535 — — OQ453488 [39] Hirschioporaceae Nigrohirschioporus trimiticus B3471 Brazil OQ453306 OQ453533 — OQ857962 — [39] Hirschioporaceae Nigrohirschioporus trimiticus B4105 Brazil OQ453307 OQ453534 OQ453510 OQ857963 OQ453487 [39] Hirschioporaceae Nigrohirschioporus wuyiensis Dai 24990 China PX741910 PX747682 PX747725 PX766000 PX747669 Present study Hirschioporaceae Nigrohirschioporus wuyiensis Dai 24991 China PX741911 PX747683 PX747726 PX766001 PX747670 Present study Hirschioporaceae Pallidohirschioporus biformis Cui 7213 Japan OQ453310 OQ453537 — — OQ453490 [39] Hirschioporaceae Pallidohirschioporus biformis CZ 411 China FJ755247 — — — — Genbank Hirschioporaceae Pallidohirschioporus biformis Dai 12746 USA OQ453311 OQ453538 OQ453512 OQ874735 OQ453491 [39] Hirschioporaceae Pallidohirschioporus biformis Dai 16076 China OQ453316 OQ453542 OQ453517 OQ874740 OQ453495 [39] Hirschioporaceae Pallidohirschioporus biformis Dai 19466 China OQ453223 OQ453548 OQ453523 OQ874743 OQ453501 [39] Hirschioporaceae Pallidohirschioporus biformis Dai 20920 Belarus OQ453226 OQ453282 OQ453526 OQ874745 OQ453504 [39] Hirschioporaceae Pallidohirschioporus biformis He 2146 USA OQ453230 OQ453286 OQ453529 OQ874747 OQ453507 [39] Hirschioporaceae Pallidohirschioporus brastagii Dai 12316 China OQ453235 OQ453289 OQ453268 OQ874749 OQ453246 [39] Hirschioporaceae Pallidohirschioporus brastagii Dai 18804 Australia OQ453236 OQ453290 OQ453269 — OQ453247 [39] Hirschioporaceae Pallidohirschioporus brastagii Dai 20459 China OQ453240 OQ453294 OQ453271 — OQ453249 [39] Hirschioporaceae Pallidohirschioporus brastagii Dai 22919 China OQ453371 OQ453297 OQ453274 OQ874751 OQ453251 [39] Hirschioporaceae Pallidohirschioporus imbricatus Cui 5384 China OQ453374 — OQ453277 OQ874754 OQ453252 [39] Hirschioporaceae Pallidohirschioporus polycystidiatus Dai 14686 China OQ453377 OQ453300 OQ453278 OQ874756 OQ453254 [39] Hirschioporaceae Pallidohirschioporus polycystidiatus Dai 19100 China OQ453378 OQ453301 OQ453279 OQ874757 OQ453255 [39] Hirschioporaceae Pallidohirschioporus polycystidiatus Dai 19103 China OQ453380 OQ474949 OQ453281 OQ874759 OQ453257 [39] Hirschioporaceae Pallidohirschioporus subchartaceus CCFC003932 USA AF266679 — AF408714 — — [112] Hirschioporaceae Pallidohirschioporus subchartaceus JV 0509/146 USA MF381010 — — — — [36] Hirschioporaceae Pallidohirschioporus versicolor Cui 9701 China OQ504334 OQ504324 OQ512029 OQ874760 OQ504328 [39] Hirschioporaceae Pallidohirschioporus versicolor Cui 16974 China OQ453383 OQ474950 OQ534100 — OQ453259 [39] Hirschioporaceae Pallidohirschioporus versicolor Dai 19331 China OQ453386 OQ474951 — OQ874762 OQ453261 [39] Hirschioporaceae Pallidohirschioporus versicolor Dai 19332 China OQ453387 OQ474952 OQ534102 OQ874763 OQ453262 [39] Hirschioporaceae Pallidohirschioporus versicolor Dai 22867 China OQ453390 OQ474954 OQ534105 OQ874766 OQ453265 [39] Hirschioporaceae Perennihirschioporus agricola JV 1407/97 Costa Rica MF380993 — — — — [36] Hirschioporaceae Perennihirschioporus agricola JV 1504/75-J Costa Rica MF380995 — — — — [36] Hirschioporaceae Perennihirschioporus daedaleus Cui 18235 Australia OQ474938 — OQ534106 OQ874767 OQ450175 [39] Hirschioporaceae Perennihirschioporus daedaleus Dai 21143 Australia OQ474939 — — — — [39] Hirschioporaceae Perennihirschioporus fumosoavellaneus JV 1607/79-J Costa Rica MF381021 — — — — [36] Hirschioporaceae Perennihirschioporus fumosoavellaneus JV 2203/80 Costa Rica OQ474940 OQ474955 OQ512028 OQ874768 OQ450176 [39] Hirschioporaceae Perennihirschioporus fumosoavellaneus JV 2203/85 Costa Rica PX747736 — — — — Present study Hirschioporaceae Perennihirschioporus perennis Dai 19295 China OQ474941 OQ474956 — OQ874779 — [39] Hirschioporaceae Perennihirschioporus sp1 CBS 455.76 India MH860992 MH872763 — — — [107] Hirschioporaceae Perennihirschioporus caymanensis JV 2208/7-J French Guiana PX741914 PX747686 PX747727 PX766004 PX747673 Present study Hirschioporaceae Perennihirschioporus variabilis B856 Brazil OQ474942 OQ474957 — OQ874769 OQ450177 [39] Hirschioporaceae Perennihirschioporus variabilis JV 1707/40-J Mexico OQ504335 OQ504325 OQ883911 OQ874770 — [39] Hirschioporaceae Perennihirschioporus variabilis Ryvarden 35177 Venezuela OQ504337 — — — — [39] Pseudotrichaptaceae Pseudotrichaptum laricinum ANT242-QFB28749 Canada MN992532 — — — — Genbank Pseudotrichaptaceae Pseudotrichaptum laricinum Dai 19455 China — OQ449019 — — OQ449394 [39] Pseudotrichaptaceae Pseudotrichaptum laricinum Dai 19457 China OQ449078 OQ449020 OQ517061 — OQ449395 [39] Pseudotrichaptaceae Pseudotrichaptum laricinum Dai 23782 China OQ449079 OQ449021 OQ517062 — OQ449396 [39] Pseudotrichaptaceae Pseudotrichaptum laricinum RLG-4665 USA U63471 — — — — [115] Pseudotrichaptaceae Pseudotrichaptum laricinum YY 99 China OQ449080 OQ449022 OQ517063 — — [39] Podocarpioporaceae Podocarpioporus podocarpi Dai 12015 China OQ449081 — — — OQ449397 [39] Podocarpioporaceae Podocarpioporus podocarpi Dai 21986 China OQ449082 OQ449023 OQ517064 OQ785648 OQ449398 [39] Podocarpioporaceae Podocarpioporus hydnoides Dai 26968 China PX741918 PX747689 PX747730 PX766006 PX747676 Present study Trichaptaceae Trichaptum byssogenum Dai 10753 China OQ449083 OQ449024 OQ517065 — OQ449399 [39] Trichaptaceae Trichaptum byssogenum Dai 15555 China OQ449085 OQ449026 OQ517067 OQ874771 OQ449401 [39] Trichaptaceae Trichaptum byssogenum Dai 16758 Thailand OQ449086 OQ449027 OQ517068 OQ874772 OQ449402 [39] Trichaptaceae Trichaptum byssogenum Dai 18850 Australia OQ449087 — OQ517069 OQ874773 — [39] Trichaptaceae Trichaptum byssogenum E7361 Indonesia AJ536654 — — — — Genbank Trichaptaceae Trichaptum cystidiolatum JV 1512/18-J Costa Rica MF380996 — — — — [36] Trichaptaceae Trichaptum cystidiolatum JV 2203/77 Costa Rica PX741921 PX747690 PX747732 PX766008 PX747678 Present study Trichaptaceae Trichaptum cystidiolatum JV 2203/78 Costa Rica PX741922 PX747691 PX747733 PX766009 PX747679 Present study Trichaptaceae Trichaptum perrottetii JV 1808/101 French Guiana OQ449091 OQ449030 OQ517072 OQ874775 OQ449404 [39] Trichaptaceae Trichaptum perrottetii JV 1908/45 French Guiana OQ449092 OQ449031 — OQ874776 OQ449405 [39] Trichaptaceae Trichaptum perrottetii B2626 Brazil OQ449093 OQ449032 — OQ874777 OQ449406 [39] Trichaptaceae Trichaptum resacarium JV 1109/56 USA MF381020 — — — — [39] Trichaptaceae Trichaptum resacarium JV 1109/57 USA MF381018 — — — — [39] Trichaptaceae Trichaptum strigosum JV 1012/2-J USA MF381011 — OQ517073 OQ874778 — [39] Trichaptaceae Trichaptum longisporum JV 0809/19-J Costa Rica PX747737 — — — — Present study Trichaptaceae Trichaptum longisporum JV 2203/104 Costa Rica PX747738 — — — — Present study Trichaptaceae Trichaptum longisporum JV 2311/13-J Bolivia PX741923 PX747692 PX747734 PX766010 PX747680 Present study Trichaptaceae Trichaptum longisporum JV 2311/18-J Bolivia PX747739 — — — — Present study Trichaptaceae Trichaptum longisporum JV 2311/19-J Bolivia PX741924 PX747693 PX747735 PX766011 PX747681 Present study Note: '–' indicates that no sequence was obtained in this study. Datasets
-
In this study, three datasets were used for the phylogenetic analyses, divergence time estimation, as well as the historical biogeography and ancestral state of the basidiomata. Dataset 1 included five gene fragments, namely ITS, nLSU, TEF1, mt-SSU, and nuc-SSU, from 157 specimens, for the phylogenetic analyses (Table 2). Dataset 2 consisted of three gene fragments, including ITS, nLSU, and TEF1, from 124 specimens, to estimate the divergence time of Trichaptum s.l. (Table 2 and Supplementary Table S1). Dataset 3 also contained three gene fragments, including ITS, nLSU, and TEF1, from 52 specimens, to infer the historical biogeography and ancestral state of the basidiomata (Table 2 and Supplementary Table S1).
Phylogenetic analyses
-
All the sequences of the five gene fragments were aligned separately using MAFFT v. 7.0[67], manual editing and adjustments of bases were performed in BioEdit v. 7.0.9[68], and then combined with the five gene fragments as a dataset. The best optimal model for the combined dataset was determined using ModelTest-NG v0.1.7[69]. Phylogenetic analyses were performed using Maximum Likelihood (ML) and Bayesian Inference (BI) using RAxML v. 8.2.12[70] and MrBayes v. 3.2.6[71], respectively. For ML analysis, bootstrap support values (BS) were estimated with 1,000 replicates using default parameters. For BI analysis, Bayesian posterior probabilities (BPP) were calculated with two million generations, and the first 25% were discarded as burn-in. Finally, the ML and BI trees were visualized using Figtree v1.4.4 (
https://tree.bio.ed.ac.uk/software/figtree ). Nodes with ML bootstrap values below 50% and BI posterior probabilities below 0.90 were excluded from consideration.Divergence time estimation of Trichaptum s.l.
-
The divergence time estimation of Trichaptum s.l. was based on three gene fragments (ITS, nLSU, and TEF1) from 124 specimens, following the methods of previous studies[18,33,39]. Briefly, XML was generated using BEAUTi v. 2.6.6[72] with the most optimal model of the GTR substitution model, which was estimated by ModelTest-NG v0.1.7[69], using a log-normal relaxed molecular clock and Yule speciation prior. Three fossil calibration points with established fossil records were selected: 1) 90 Mya representing the minimum age of Agaricales[73,74]; 2) 125 Mya representing the minimum age of Hymenochaetaceae[75,76]; and 3) 400 Mya representing the divergence time between the Ascomycota and the Basidiomycota[77,78]. The offset age with a gamma-distributed prior (scale = 20 and shape = 1) was set as 90, 125, and 400 Mya for Agaricales, Hymenochaetaceae, and Basidiomycota, respectively. Four independent Markov Chain Monte Carlo (MCMC) chains were run for 100 million generations, with tree sampling every 1,000 generations. Tracer v. 1.5 (
https://beast.bio.ed.ac.uk/Tracer ) was used to check the results log file to confirm that ESS ≥ 200. FigTree v. 1.4.4 (https://tree.bio.ed.ac.uk/software/figtree ) was used to summarize the Maximum Clade Credibility (MCC) tree with 95% Highest Posterior Density (HPD) intervals, removing 20% as burn-in and setting the posterior probability limit to 0.80.Inferring historical biogeography and ancestral state of basidiomata
-
The historical biogeography and ancestral state of basidiomata of Trichaptum s.l. were inferred using Reconstruct Ancestral State in Phylogenies v. 4.2 (RASP)[79,80] under the Statistical Dispersal-Extinction-Cladogenesis (S-DEC) model and the Bayesian Binary MCMC model. The distribution of Trichaptum s.l. species was divided into six geographic zones, viz., A) Central and South America, B) North America, C) Europe, D) Central and East Asia, E) South to Southeast Asia and Oceania, and F) Africa, as well as four climate zones, namely A) temperate zones, B) subtropical zones, C) tropical zones, and D) mountain zones. The consistency and pileal surface morphology of basidiomata were classified into four and five types, respectively. The four types of consistency were A) Soft corky, B) Coriaceous, C) Hard corky, and D) Woody. The five types of pileal surface were A) Tomentose, B) Smooth, C) Warted, D) Hispid, and E) Apileate.
Species distribution and environmental factors
-
Records of Trichaptum s.l. were gathered from global field survey sites and the online database of the Global Biodiversity Information Facility (GBIF,
www.gbif.org , accessed on 1st Oct, 2025), prioritizing samples with molecular or photographic verification. For records lacking precise geo-coordinates, Google Maps (www.google.com/maps ) was used to estimate approximate latitude and longitude based on the described geographic locations. The dataset underwent rigorous cleaning. First, duplicate distribution points were filtered based on longitude and latitude coordinates. To mitigate sampling bias and spatial autocorrelation, the data were spatially thinned to match the resolution of the environmental variables, retaining only one occurrence record per 2.5 arc-min grid cell. Second, extreme outlier values were manually identified and removed by cross-referencing with explicit climatic distribution records documented in the literature. After these quality control measures, a final set of 25,828 valid distribution records for Trichaptum s.l. was established. The species of the genera were analyzed on a genus-by-genus basis to understand the key environmental factors influencing the distribution of each genus.The maps were downloaded from the Natural Earth website (
www.naturalearthdata.com ) as the World Map (1:10 million scale). The collected geographic data of the generalized Trichaptum s.l. were processed and imported into ArcGIS software to create vector maps, ultimately producing the species' actual distribution map. Nineteen bioclimatic variables and elevation data with a spatial resolution of 2.5 arc-min were downloaded from the World Clim database (www.worldclim.org ) (Table 3). These data were used to identify the primary environmental factors influencing the distribution of Trichaptum s.l. species, including baseline climate condition data averaged over the period from 1970 to 2000.Table 3. Environmental variables used to create the species distribution model
Description Unit Bio1 Annual mean temperature °C Bio2 Mean diurnal range (mean of monthly (max tempmin temp)) °C Bio3 Isothermality (Bio2/Bio7) (× 100) C of V Bio4 Temperature seasonality (standard deviation × 100) °C Bio5 Maximum temperature of warmest month °C Bio6 Minimum temperature of coldest month °C Bio7 Temperature annual range (Bio5-Bio6) °C Bio8 Mean temperature of wettest quarter °C Bio9 Mean temperature of driest quarter °C Bio10 Mean temperature of warmest quarter °C Bio11 Mean temperature of coldest quarter °C Bio12 Annual precipitation mm Bio13 Precipitation of wettest month mm Bio14 Precipitation of driest month mm Bio15 Precipitation seasonality (coefficient of variation) C of V Bio16 Precipitation of wettest quarter mm Bio17 Precipitation of driest quarter mm Bio18 Precipitation of warmest quarter mm Bio19 Precipitation of coldest quarter mm Bio20 Elevation m -
A total of 157 specimens of Dataset 1 (comprising 5,957 base pairs across five gene fragments) were used to reconstruct the phylogenetic relationships of Trichaptum s.l., including 148 specimens of Trichaptum s.l., three specimens of Neoantrodiella, four specimens of Nigrofomes, and two specimens of Coltricia as the outgroups[39]. Phylogenetic analyses were conducted using RAxML v. 8.2.12[70] and MrBayes v. 3.2.6[71], employing the best-fit model GTR + F + I + G4.
Phylogenetic inference resolved that Trichaptum s.l. is divided into four families: Hirschioporaceae, Pseudotrichaptaceae fam. nov., Podocarpioporaceae fam. nov., and Trichaptaceae (Fig. 1). The family Trichaptaceae forms a clade with Nigrofomitaceae and is closely related to Hirschioporaceae. Podocarpioporaceae is closely related to Neoantrodiellaceae, while Pseudotrichaptaceae is an independent clade within the order Hymenochaetales. Additionally, further analyses based on divergence times and morphological evidence were performed for the four families within Trichaptum s.l.
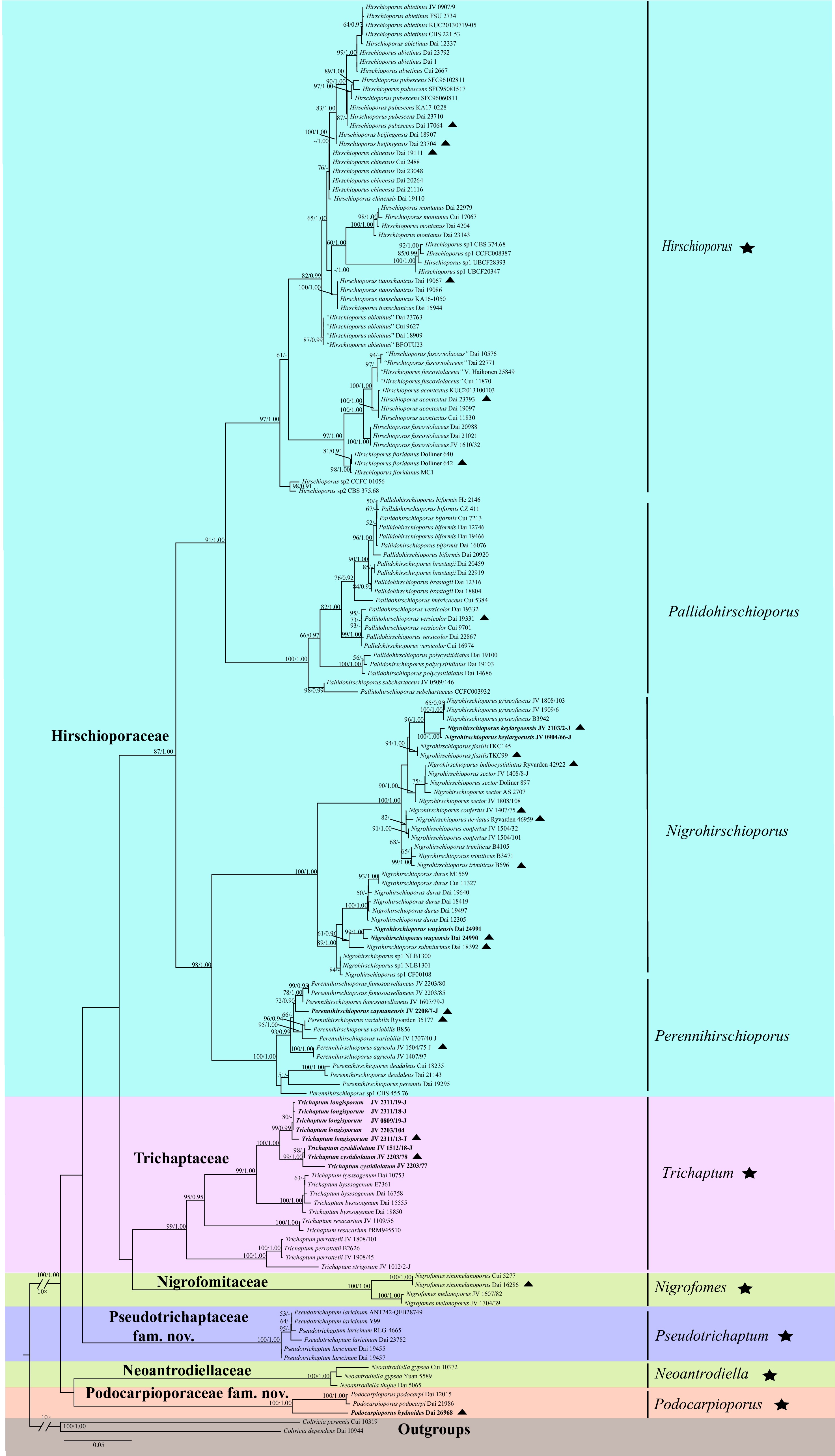
Figure 1.
Maximum Likelihood (ML) tree of Trichaptum s.l. was constructed based on five sequences (ITS, nLSU, TEF1, mt-SSU, and nuc-SSU), using Coltricia dependens and C. perennis as outgroups. ML bootstrap values (≥ 50%) and Bayesian inference (BI) posterior probabilities (≥ 0.9) are shown along the branches for each clade. The six new species, Nigrohirschioporus keylargoensis, N. wuyiensis, Perennihirschioporus caymanensis, Podocarpioporus hydnoides, Trichaptum cystidiolatum, and T. longisporum, are highlighted in bold. A scale bar in the upper left represents substitutions per site. Black stars (★) represent type genera, and black triangles (▲) represent type sequences.
At the genus level, Hirschioporaceae comprises the most genera, including Hirschioporus, Nigrohirschioporus, Pallidohirschioporus, and Perennihirschioporus, while Pseudotrichaptaceae, Podocarpioporaceae, and Trichaptaceae each contain a single genus, namely Pseudotrichaptum, Podocarpioporus, and Trichaptum, respectively. All genera were recovered as monophyletic.
Phylogenetic analyses also revealed six new species described in this study, Nigrohirschioporus keylargoensis sp. nov., Nigrohirschioporus wuyiensis sp. nov., Perennihirschioporus caymanensis sp. nov., Podocarpioporus hydnoides sp. nov., Trichaptum cystidiolatum sp. nov., and Trichaptum longisporum sp. nov. (Fig. 1). Nigrohirschioporus keylargoensis is most closely related to N. griseofuscus, with strong support (96/1.00 in ML and BI analyses). Nigrohirschioporus wuyiensis is closely related to N. submiurinus, with strong support (99/1.00 in ML and BI analyses). Perennihirschioporus caymanensis is closely related to P. fumosoavellaneus, with moderate support (72/0.90 in ML and BI analyses). Podocarpioporus hydnoides is closely related to P. podocarpi, with full support (100/1.00 in ML and BI analyses). Trichaptum cystidiolatum is most closely related to Trichaptum longisporum, with full support (100/1.00 in ML and BI analyses).
In addition, five putative species represented by single specimens, "Hirschioporus abietinus", "Hirschioporus fuscoviolaceus", Hirschioporus sp 1, Hirschioporus sp 2, and Nigrohirschioporus sp 1, require further sampling for formal description.
Divergence times of Trichaptum s.l.
-
The dataset 2 (comprising 4,320 characters from three gene fragments) was used to estimate the divergence times of Trichaptum s.l. using Beast software v. 2.6.6[72] (Table 4). The results indicated that with the following mean stem ages (Fig. 2) the Basidiomycota and Ascomycota diverged at approximately 449 Mya with a 95% highest posterior density (HPD) of 406–554 Mya, the Agaricomycetes diverged around 330 Mya with a 95% HPD of 287–371 Mya, the Agaricales at 156 Mya with a 95% HPD of 121–196 Mya, and the Hymenochaetales at 247 Mya with a 95% HPD of 213–284 Mya. In addition, the mean stem ages of families within the order Hymenochaetales ranged from 71 Mya to 219 Mya.
Table 4. The estimated divergence times of Trichaptum s.l.
Family Genus Means of stem age (Mya) 95% HPD (Mya) Hirschioporaceae 116 73–119 Hirschioporus 75 55–97 Nigrohirschioporus 74 52–97 Pallidohirschioporus 75 55–97 Perennihirschioporus 74 52–97 Pseudotrichaptaceae 71 42–102 Pseudotrichaptum 71 42–102 Podocarpioporaceae 109 72–147 Podocarpioporus 109 72–147 Trichaptaceae 96 70–125 Trichaptum 96 70–125 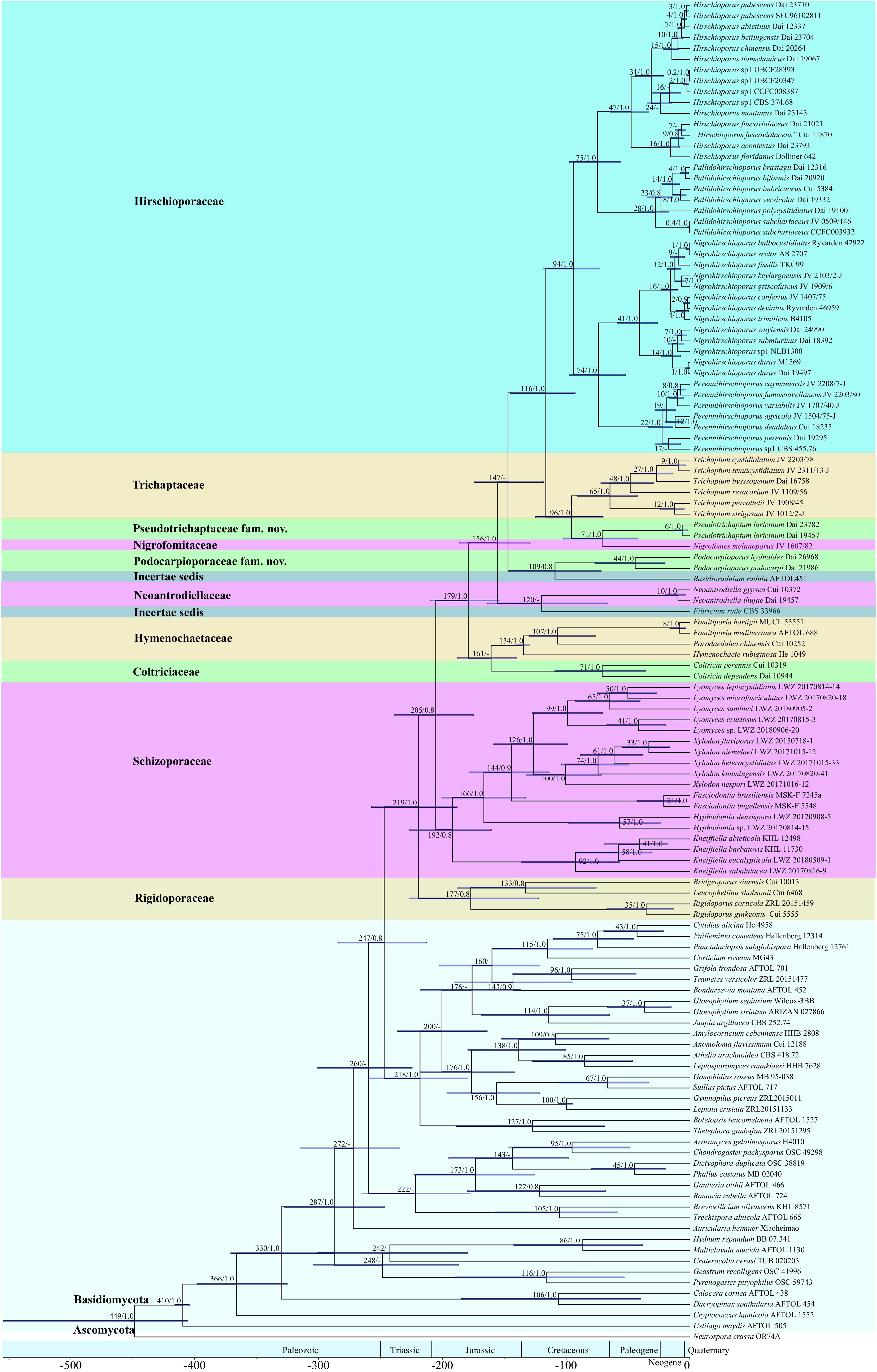
Figure 2.
The estimated divergence times for the Trichaptum s.l. are shown, derived from molecular clock analyses using three sequences (ITS, nLSU, and TEF1). Mean divergence times (Mya) and posterior probabilities (PP) > 0.8 are annotated at the internodes, with horizontal blue bars representing the 95% highest posterior density (HPD) intervals for divergence time estimates.
Within Trichaptum s.l., the mean stem ages of the four families ranged from 71 Mya (with a 95% HPD of 42–102 Mya) for Pseudotrichaptaceae to 116 Mya (with a 95% HPD of 73–119 Mya) for Hirschioporaceae, during the Cretaceous period. The mean stem ages for Podocarpioporaceae and Trichaptaceae were 109 Mya (with a 95% HPD of 72–147 Mya) and 96 Mya (with a 95% HPD of 70–125 Mya), respectively. The mean stem ages for genera and species within Trichaptum s.l. were 71–96 Mya and 1–71 Mya, respectively. In this study, with the exception of Pseudotrichaptum laricinum, all other species diverged starting in the Eocene period (1–48 Mya).
Taxonomy
-
In this study, two new families, Pseudotrichaptaceae and Podocarpioporaceae, and six new species of Trichaptum s.l. within the Hymenochaetales are proposed Nigrohirschioporus keylargoensis, N. wuyiensis, Perennihirschioporus caymanensis, Podocarpioporus hydnoides, Trichaptum cystidiolatum, and T. longisporum. These proposals are supported by phylogenetic, divergence time, and morphological evidence.
Key to families of Trichaptum s.l
-
1. Cystidia projecting out of the hymenium, apically often with long rectangular yellowish crystals; cystidia length usually more than twice that of basidia; restricted to Podocarpus Podocarpioporaceae 1. Cystidia usually within the hymenium, apically with hyaline crystals; cystidia similar in length to basidia 2 2. Hymenophore lamellate, completely without pores; growing on gymnosperm wood Pseudotrichaptaceae 2. Hymenophore poroid or hydnoid, poroid at least when young or at the margin; growing on angiosperm and gymnosperm wood 3 3. Upper surface glabrous to adpressed tomentose, mixed with glabrous zones; cystidia mostly fusoid, clavate to capitate Trichaptaceae 3. Upper surface strongly strigose to hispid; cystidia mostly subulate to ventricose Hirschioporaceae Pseudotrichaptaceae Meng Zhou, Y.J. Cui, H. Zhao, Y.C. Dai & Yuan Yuan, fam. nov.
-
MycoBank number: MB 862003
Type genus – Pseudotrichaptum Y.C. Dai, Yuan Yuan & Meng Zhou, Mycosphere 14(1): 890 (2023)
Type species – Pseudotrichaptum laricinum (P. Karst.) Y.C. Dai, Yuan Yuan & Meng Zhou, Mycosphere 14(1): 890 (2023)
Key characteristics: Basidiomata annual, pileate or effused-reflexed, coriaceous to tough; pileal surface cream to pale buff with distinct concentric sulcate zones, covered by mosses with age; hymenophore lamellate; lamella purplish, thin, entire and wavy, dichotomously forked near margin; context duplex. Hyphal system dimitic; generative hyphae with clamp connections, skeletal hyphae pale yellowish in KOH, IKI–, CB+; cystidia apically encrusted; basidiospores allantoid, IKI–, CB–.
Notes: Pseudotrichaptaceae is distinguished from other families in Trichaptum s.l. by its coriaceous pilei with purplish lamellate hymenophore and primarily inhabit coniferous trees with a circumboreal distribution in the Northern Hemisphere.
Podocarpioporaceae Meng Zhou, Y.J. Cui, H. Zhao, Y.C. Dai & Yuan Yuan, fam. nov.
-
MycoBank number: MB 862004
Type genus – Podocarpioporus Y.C. Dai, Yuan Yuan & Meng Zhou, Mycosphere 14(1): 888 (2023)
Type species – Podocarpioporus podocarpi (Y.C. Dai) Y.C. Dai, Yuan Yuan & Meng Zhou, Mycosphere 14(1): 889 (2023)
Key characteristics: Basidiomata annual, resupinate or effused-reflexed to pileate; hymenophore poroid to irpicoid. Hyphal system dimitic; generative hyphae with clamp connections; skeletal hyphae occasionally with simple septa, IKI–, CB+; tissue darkening but otherwise unchanged in KOH. Cystidia encrusted at the apex or not; basidiospores allantoid, hyaline, thin-walled, smooth, IKI–, CB–.
Notes: This family is unique in Trichaptum s.l. because of the poroid to irpicoid hymenophore, narrow allantoid basidiospores, distinctly long subulate hymenial cystidia, usually more than twice that of basidia, and growth only on species of Podocarpaceae.
Nigrohirschioporus keylargoensis Meng Zhou, Y.C. Dai, H. Zhao, Vlasák & Yuan Yuan sp. nov. (Figs 3, 4)
-

Figure 3.
Basidiomata of Nigrohirschioporus keylargoensis JV0904/66-J. Scale bars: (a) = 1 cm, (b) and (c) = 1 mm.
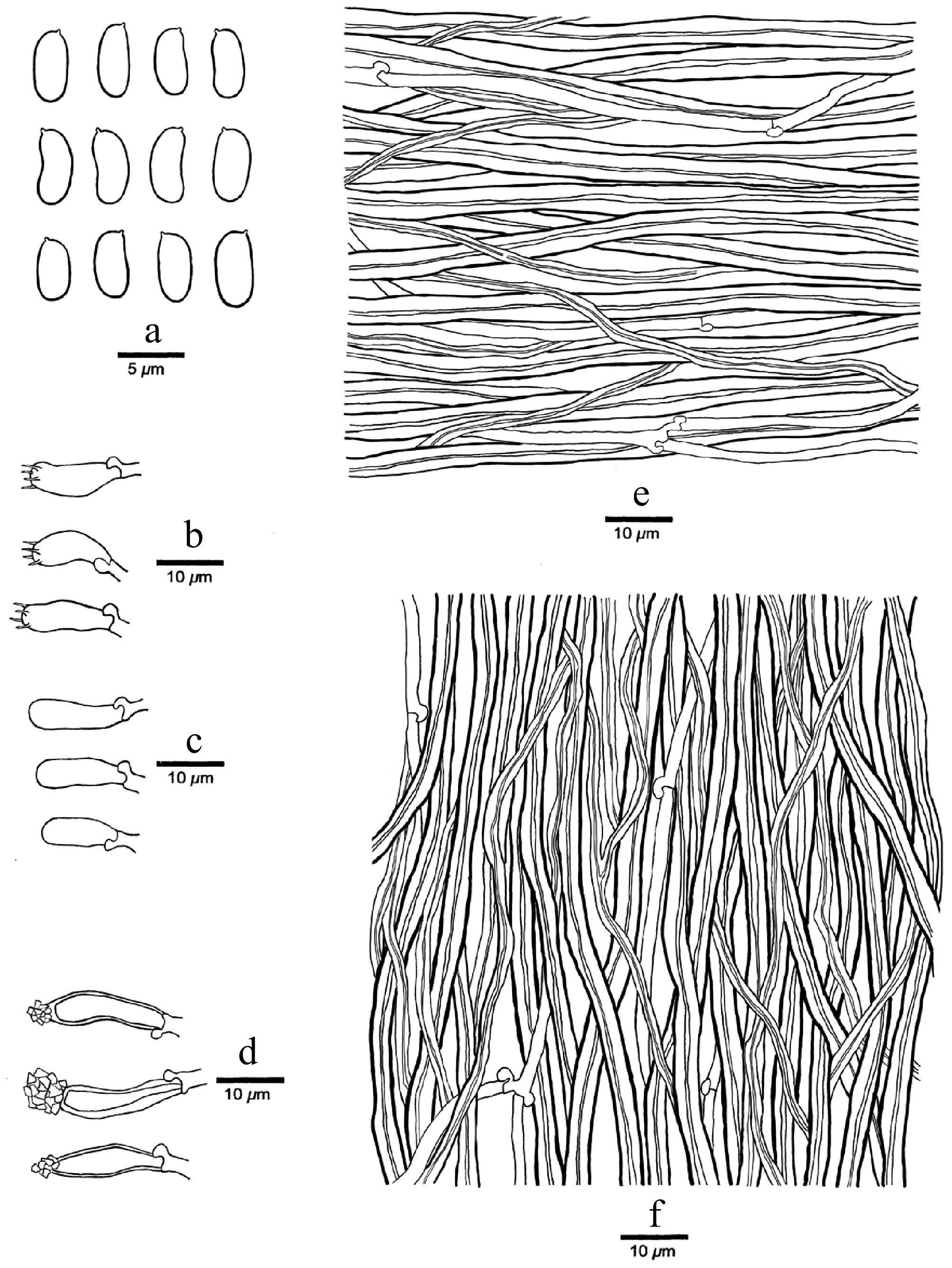
Figure 4.
Microscopic structures of Nigrohirschioporus keylargoensis (drawn from the holotype, JV0904/66-J). (a) Basidiospores. (b) Basidia. (c) Basidioles. (d) Cystidia. (e) Hyphae from context. (f) Hyphae from trama.
MycoBank number: MB 361999
Type – USA. Florida, Key Largo, on fallen angiosperm trunk, III.2021, JV 2103/2-J (Holotype BJFC039885).
Etymology – Keylargoensis (Lat.): refers to the type locality, Key Largo, Florida, USA.
Basidiomata – Annual, effused-reflexed to pileate; corky when fresh and dry. Pilei semicircular to elongate, flexible and tough, projecting up to 3 cm, 3 cm wide and 3 mm thick at the base. Pileal surface pale fuscous, vinaceous or vinaceous buff to snuff brown when fresh and honey yellow to fuscous when dry, finely tomentose to velutinate, sub-scaly towards the margin; margin entire to lobed. Hymenophore poroid to irpicoid or hydnoid, olivaceous buff when fresh, grayish brown to fuscous when dry; pores angular when juvenile, becoming split to dentate with age, 1–2 per mm; dissepiments thin, entire when juvenile, strongly lacerate with age. Context duplex, up to 1.5 mm thick, the upper layer grayish brown, pubescent, soft, up to 1 mm, the lower layer coriaceous dark brown, tubes brown, up to 2 mm long.
Hyphal structure – Hyphal system trimitic; generative hyphae with clamp connections, hyaline, thin-walled; skeletal hyphae thick-walled with a narrow lumen, slightly dextrinoid, CB+; binding hyphae thick-walled to sub-solid; tissues darkening in KOH.
Context – Generative hyphae hyaline, thin-walled, occasionally branched, 2.5–3 µm in diam.; skeletal hyphae thick-walled with a narrow lumen, unbranched, regularly arranged, 4–6 μm in diam.; binding hyphae thick-walled with a narrow lumen, rarely branched, interwoven, 1.5–2.5 μm in diam.
Tubes – Generative hyphae hyaline, thin-walled, occasionally branched, 2.5–3 µm in diam.; skeletal hyphae thick-walled, unbranched, interwoven, 3–5 μm in diam; binding hyphae thick-walled with a narrow lumen to sub-solid, rarely branched, interwoven, 1.5–2 μm in diam.
Hymenium – Cystidia fusoid to capitate, slightly thick-walled, abundant, sometimes apically encrusted, 15–25 × 4–6 µm; hyphal pegs absent; basidia clavate, with four sterigmata and a basal clamp connection, 13–18 × 4.5–6 μm; basidioles similar with basidia in shape, but smaller.
Spores – Basidiospores cylindric to slightly allantoid, hyaline, thin-walled, smooth, IKI–, CB–, (5.0–)5.1–7.0(–7.1) × (1.9–)2–3(–3.2) µm, L = 5.95 μm, W = 2.41μm, Q = 2.03–2.95 (n = 60/2).
Type of rot – White rot.
Distribution and ecology – Nigrohirschioporus keylargoensis has so far only been found in Florida, USA.
Specimens examined – USA. Florida, Key Largo, on a fallen angiosperm trunk, IV.2009, JV0904/66-J (BJFC039884); III.2021, JV 2103/2-J (holotype BJFC039885).
Nigrohirschioporus wuyiensis Meng Zhou, Y.J. Cui, H. Zhao, Y.C. Dai & Yuan Yuan, sp. nov. (Figs 5,6)
-

Figure 5.
Basidiomata of Nigrohirschioporus wuyiensis Dai 24990 (holotype). Scale bars: (a) = 1 cm, (b) and (c) = 1 mm.
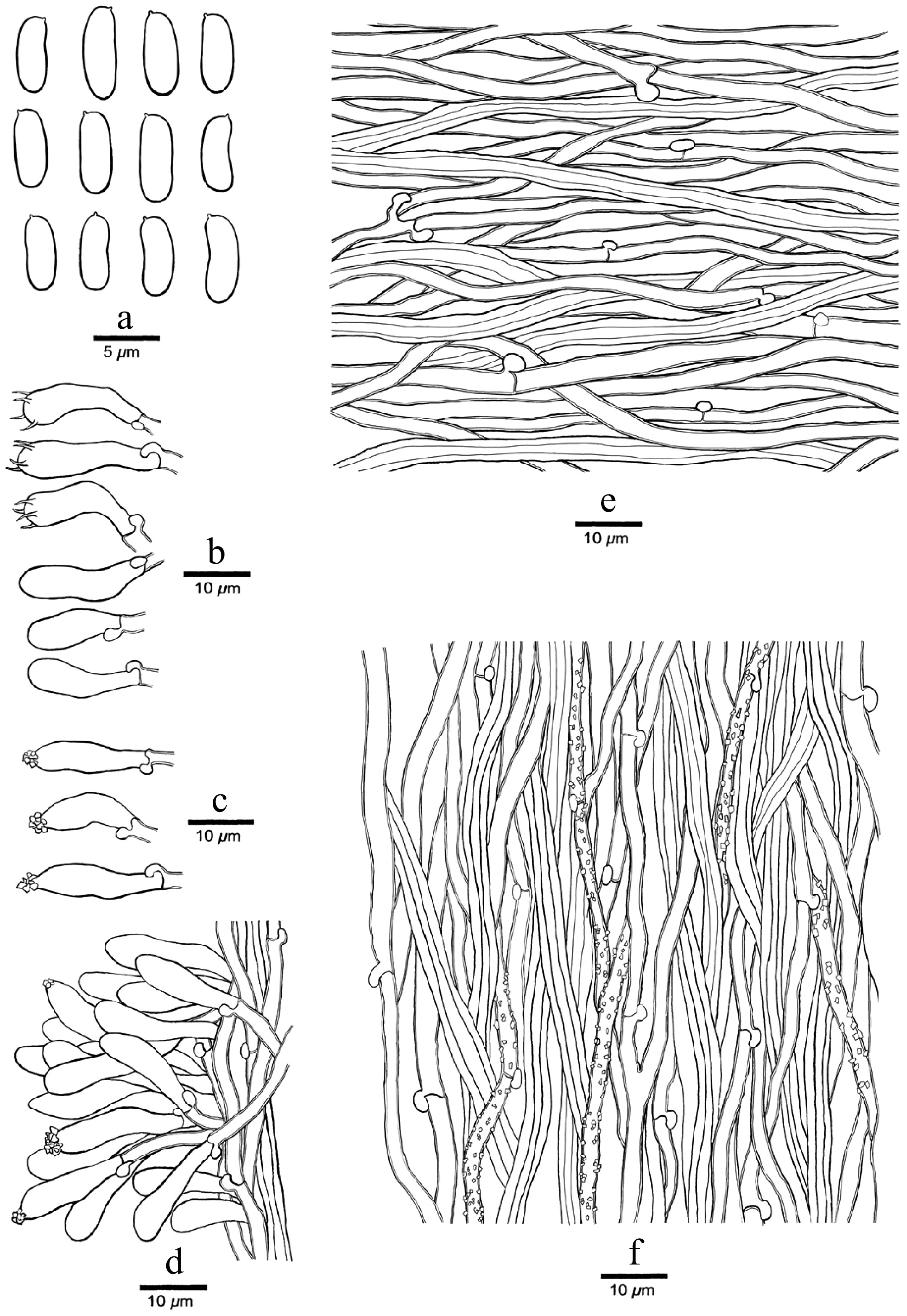
Figure 6.
Microscopic structures of Nigrohirschioporus wuyiensis (drawn from the holotype, Dai 24990). (a) Basidiospores. (b) Basidia and basidioles. (c) Cystidia. (d) Hyphal peg. (e) Hyphae from context. (f) Hyphae from trama.
MycoBank number: MB 361998
Type – CHINA. Zhejiang, Wuyi, on fallen branch of Liquidambar, 18.VI.2023, Dai 24990 (Holotype BJFC042543).
Etymology – Wuyiensis (Lat.): refers to the species being found in Wuyi, Zhejiang Province in China.
Basidiomata – Annual, resupinate; corky when fresh, becoming brittle upon drying, up to 5 cm long, 2 cm wide, and 3 mm thick at the center. Hymenophore poroid, olivaceous buff when fresh, buff to clay buff when dry; pores angular when juvenile, becoming irpicoid with age, 4–6 per mm; dissepiments thin, entire when juvenile, strongly lacerate with age. Context buff-yellow when dry, up to 0.5 mm thick. Tubes pale buff, up to 2.5 mm long.
Hyphal structure – Hyphal system dimitic; generative hyphae with clamp connections, hyaline, slightly thick-walled; skeletal hyphae distinctly thick-walled, slightly dextrinoid, CB+; tissues darkening in KOH.
Context – Generative hyphae hyaline, slightly thick-walled, rarely branched, 2–3.5 µm in diam.; skeletal hyphae thick-walled with a wide lumen, unbranched, interwoven, 3–4 μm in diam.
Tubes – Generative hyphae hyaline, slightly thick-walled, rarely branched, usually covered by abundant fine thorn-like crystals, 2.5–3.5 µm in diam.; skeletal hyphae thick-walled, unbranched, interwoven, 3–5 μm in diam.
Hymenium – Cystidia clavate to fusoid, thin-walled, abundant, sometimes apically encrusted, 17–25 × 4.5–6 µm; hyphal pegs present; basidia clavate, with four sterigmata and a basal clamp connection, 16.5–26 × 5.5–7 μm; basidioles similar to basidia in shape, but smaller.
Spores – Basidiospores cylindric to slightly allantoid, hyaline, thin-walled, smooth, IKI–, CB–, (5–)5.5–7.2(–7.3) × 2–2.5(–2.8) μm, L = 6.47 μm, W = 2.27 μm, Q = 2.79–2.92 (n = 60/2).
Type of rot – White rot.
Distribution and ecology – Nigrohirschioporus wuyiensis has so far only been found in Zhejiang, subtropical China.
Specimens examined – CHINA. Zhejiang, Wuyi, on a fallen branch of Liquidambar, 18.VI.2023, Dai 24990 (holotype BJFC042543); Dai 24991 (BJFC042544).
Key to known species of Nigrohirschioporus
-
1. Basidiomata perennial 2 1. Basidiomata annual 5 2. Basidiomata resupinate N. deviatus 2. Basidiomata effused-reflexed to pileate 3 3. Pores 5–8 per mm N. nigrivineus 3. Pores > 8 per mm 4 4. Pore surface pale orange, pores 13–16 per mm N. agglutinatus 4. Pore surface dark purplish to grayish brown, pores 8–10 per mm N. durus 5. Basidiomata completely resupinate 6 5. Basidiomata resupinate to effused-reflexed or pileate 8 6. Hymenophore poroid to irpicoid, pores 2–4 per mm N. bulbocystidiatus 6. Hymenophore poroid, pores 4–9 per mm 7 7. Pores 6–9 per mm; cystidia fusoid with a long neck N. submurinus 7. Pores 4–6 per mm; cystidia clavate to fusoid N. wuyiensis 8. Hymenophore hydnoid to irpicoid 9 8. Hymenophore poroid 11 9. Hyphal system dimitic N. griseofuscus 9. Hyphal system trimitic 10 10. Spores broadly ellipsoid, 4–4.8 × 3–3.5 µm N. molestus 10. Spores cylindric, slightly curved, 5.1–7 × 2–3 µm N. keylargoensis 11. Hyphal system trimitic 12 11. Hyphal system dimitic 13 12. Spores ellipsoid, 4–4.5 × 1.8–2.2 μm N. confertus 12. Spores cylindric to slightly allantoid, 5–6 × 2.4–2.7 μm N. trimiticus 13. Basidiospores cylindrical, 6.4–10 × 2.3–3.2 µm N. fissilis 13. Basidiospores narrowly ellipsoid, 5–6.4 × 2.2–2.5 µm N. sector Perennihirschioporus caymanensis Meng Zhou, H. Zhao, Vlasák, Y.C. Dai & Yuan Yuan, sp. nov. (Figs 7,8)
-

Figure 7.
Basidiomata of Perennihirschioporus caymanensis JV 2208/7-J (holotype). Scale bars: (a) = 1 cm, (b) and (c) = 1 mm.
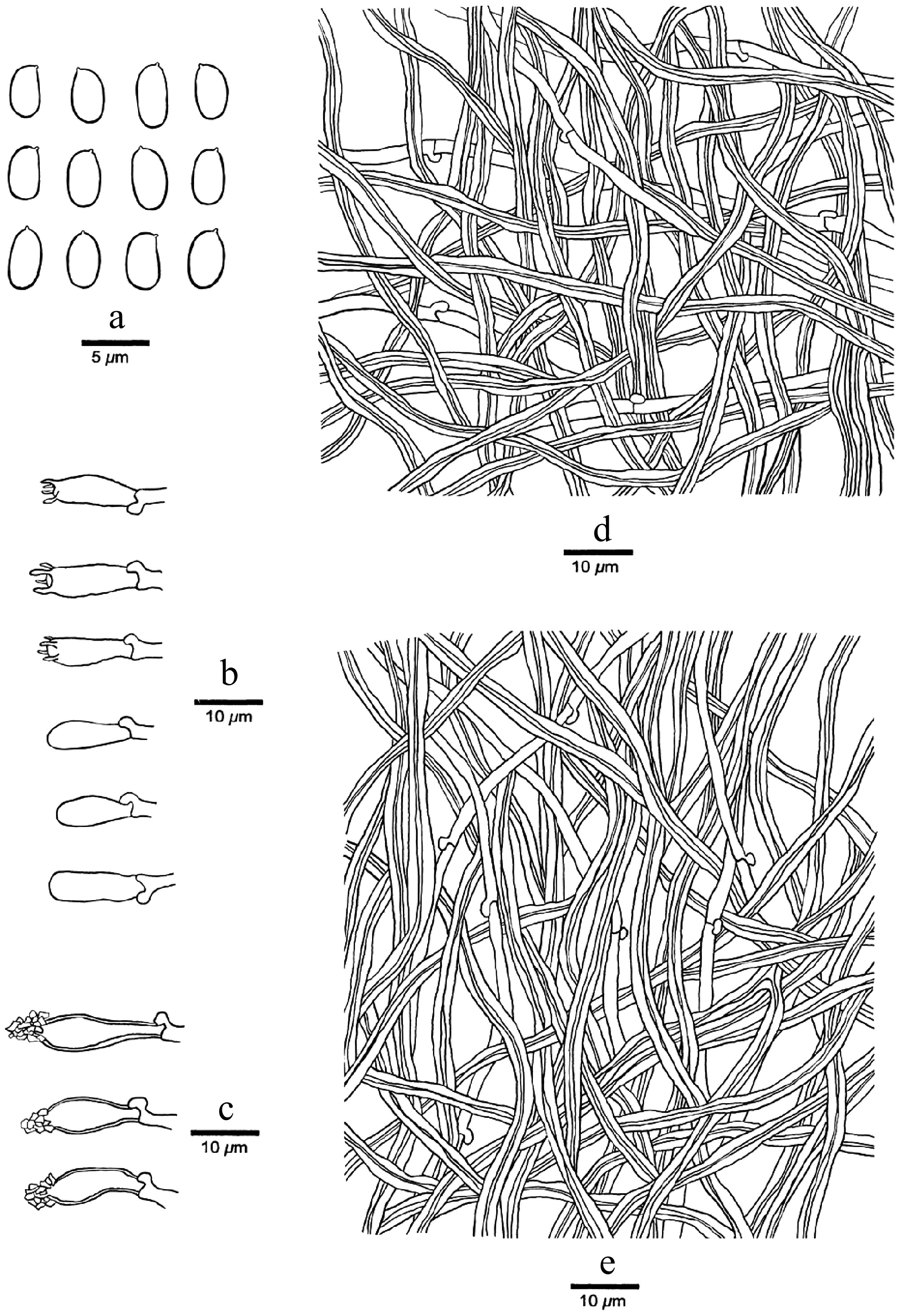
Figure 8.
Microscopic structures of Perennihirschioporus caymanensis (drawn from the holotype, JV 2208/7-J). (a) Basidiospores. (b) Basidia and basidioles. (c) Cystidia. (d) Hyphae from context. (e) Hyphae from trama.
MycoBank number: MB 861997
Type – FRENCH GUIANA, Camp Cayman, ravine, on dead stump of extremely hard, dark "ironwood", 25.VIII.2022, JV 2208/7-J (Holotype BJFC044766).
Etymology – Caymanensis (Lat.): refers to the species being found in Camp Cayman, French Guiana.
Basidiomata – Perennial, pileate, solitary, consistency hard, corky to woody; pilei applanate, projecting up to 10 cm, 10 cm wide, and 1.5 cm thick at base. Pileal surface brownish with a distinct violet tint when fresh and umber when dry, hairless, rough, slightly warted; margin brown, acute, entire. Hymenophore daedaleoid to lamellate, dark brown when fresh and grayish brown to dark brown when dry; pores or lamella 0.5–2 per mm, radially elongate or daedaleoid near the margin; dissepiments wavy, thick, entire. Sterile margin indistinct. Context homogeneous, brown, hard corky, up to 4 mm thick. Tubes concolorous with context, up to 11 mm long.
Hyphal structure – Hyphal system dimitic; generative hyphae with clamp connections, hyaline, thin-walled; skeletal hyphae dominant, yellowish, distinctly thick-walled with a narrow lumen, IKI–, CB+; tissues darkening in KOH.
Context – Generative hyphae hyaline, thin-walled, unbranched, 2–3 µm in diam.; skeletal hyphae thick-walled with a narrow lumen, unbranched, interwoven, 2–3 μm in diam.
Tubes – Generative hyphae hyaline, thin-walled, unbranched, 1.5–2.7 µm in diam.; skeletal hyphae thick-walled with a narrow lumen, rarely branched, interwoven, 2.5–4.5 μm in diam.; hyphae at dissepiment edges are smooth.
Hymenium – Cystidia fusoid, slightly thick-walled, abundant, usually apically encrusted, 17–25 × 4–5.5 µm, projecting outside of hymenium up to 7 µm, with a basal clamp connection; hyphal pegs absent; basidia clavate with four sterigmata and a basal clamp connection, 12–15 × 4.5–5.0 μm, basidioles similar to basidia in shape, but slightly smaller.
Spores – Basidiospores oblong ellipsoid to ellipsoid, hyaline, thin-walled, smooth, IKI–, CB–, (3.2–)3.5–4.3(–4.5) × (1.9–)2–2.7 μm, L =3.85 μm, W = 2.28μm, Q =1.69 (n=30/1)
Type of rot – White rot.
Distribution and ecology – Perennihirschioporus caymanensis is known from the type locality only, where several basidiomata were growing on a dead standing tree. After four years, in 2026, the fresh fungus was still growing in many pilei on the stag which indicates high subdtrate resistance to decay.
Specimens examined – FRENCH GUIANA, Camp Cayman, ravine, 25.VIII.2022, JV 2208/7-J (Holotype BJFC044766); 15.II.2026, JV 2602/8 (JV, PRM).
Key to known species of Perennihirschioporus
-
1. Hymenophore irpicoid, daedaleoid to lamellate 2 1. Hymenophore poroid 4 2. Basidiomata effused-reflexed to pileate or resupinate P. variabilis 2. Basidiomata pileate 3 3. Basidiomata applanate, hymenophore daedaleoid to lamellate P. caymanensis 3. Basidiomata triquetrous, hymenophore daedaleoid to hydnoid P. daedaleus 4. Basidiomata effused-reflexed to pileate, pore surface vinaceous brown to umber; hyphal system dimitic P. perennis 4. Basidiomata completely pileate, pore surface pale brown to avellaneus; hyphal system trimitic 5 5. Pores 1–3 per mm; basidiospores 4–5 × 2.5–3 μm P. agricola 5. Pores 3–4 per mm; basidiospores 3.6–4 × 2.5–3 μm P. fumosoavellaneus Trichaptum cystidiolatum Meng Zhou, H. Zhao, Y.J. Cui Vlasák, Y.C. Dai & Yuan Yuan, sp. nov. (Figs 9,10)
-

Figure 9.
Basidiomata of Trichaptum cystidiolatum JV 2203/78 (holotype). Note: JV 2203/78 is the whitish, eff-reflexed, large-pores Poria growing around the central, brown and massive fruitbody of P. agricola. Scale bars: (a) = 1 cm, (b) and (c) = 1 mm.

Figure 10.
Microscopic structures of Trichaptum cystidiolatum (drawn from the holotype, JV 2203/78). (a) Basidiospores. (b) Basidia and basidioles. (c) Cystidia and cystidioles. (d) Hyphae from context. (e) Hyphae from trama.
MycoBank number: MB 861996
Type – COSTA RICA. Rincón de la Vieja, Guachipelin, on hardwood, III. 2022, JV2203/78 (Holotype BJFC044764).
Etymology – Cystidiolatum (Lat.): refers to the presence of cystidioles.
Basidiomata – Perennial, effused-reflexed or resupinate; corky when fresh, hard corky when dry. Pilei are sometimes laterally fused, projecting up to 2 cm, 4 cm wide, and 4 mm thick at the base. Pileal surface consistently whitish, ochraceous to milky coffee when fresh and milky coffee when dry, adpressed velutinate; margin sharp, entire to lobed. Hymenophore poroid, whitish ochre when dry and pale buff when old; pores angular, 1–3 per mm; dissepiments thin, entire. Context gray to ochraceous or milky coffee, thin, up to 0.3 mm. Tubes concolorous with context, up to 2 mm long.
Hyphal structure – Hyphal system dimitic; generative hyphae with clamp connections, hyaline, thin-walled; skeletal hyphae distinctly thick-walled to subsolid, slightly dextrinoid, CB–; tissues unchanged in KOH.
Context – Generative hyphae hyaline, thin-walled, occasionally branched, 1–2 µm in diam.; skeletal hyphae thick-walled with a narrow lumen to subsolid, unbranched, interwoven, 1.5–2.5 μm in diam.
Tubes – Generative hyphae hyaline, thin-walled, usually branched, 1.5–2 µm in diam.; skeletal hyphae thick-walled with a wide lumen, unbranched, interwoven, 1.5–2 μm in diam.
Hymenium – Cystidia fusoid, thick-walled, apically encrusted, sometimes with several separate, 16–26 × 3–6 µm, cystidioles narrow fusoid, thin-walled, 14–22 × 3-4; hyphal pegs absent; basidia barrel-shaped, with four sterigmata and a basal clamp connection, 16–22 × 4.5–6 μm; basidioles similar with basidia in shape, but smaller.
Spores – Basidiospores oblong ellipsoid, hyaline, thin-walled, smooth, IKI–, CB–, (4.2–)4.5–6(–6.5) × (2–) 2.2–2.9 μm, L = 5.31μm, W = 2.56 μm, Q = 2.02–2.13 (n = 30/2).
Type of rot – White rot.
Distribution and ecology – To date, Trichaptum cystidiolatum is found in Costa Rica only, where it is locally not rare on wooden fence posts.
Specimens examined – COSTA RICA. Guanacaste Prov., Rincón de la Vieja, Guachipelin, on angiosperm wood of fence posts, 12.III. 2022, JV2203/77 (BJFC044763), JV2203/78 (holotype BJFC044764), JV2203/101; Santa Cruz, fallen angiosperm branch, 28.XII. 2015, JV1512/18-J (JV, PRM 945507, GenBank MF380996).
Trichaptum longisporum Meng Zhou, Y.J. Cui, H. Zhao, Vlasák, Y.C. Dai & Yuan Yuan, sp. nov. (Figs 11,12)
-

Figure 11.
Basidiomata of Trichaptum longisporum JV 2311/13-J (holotype). Scale bars: (a) = 1 cm, (b) and (c) = 1 mm.
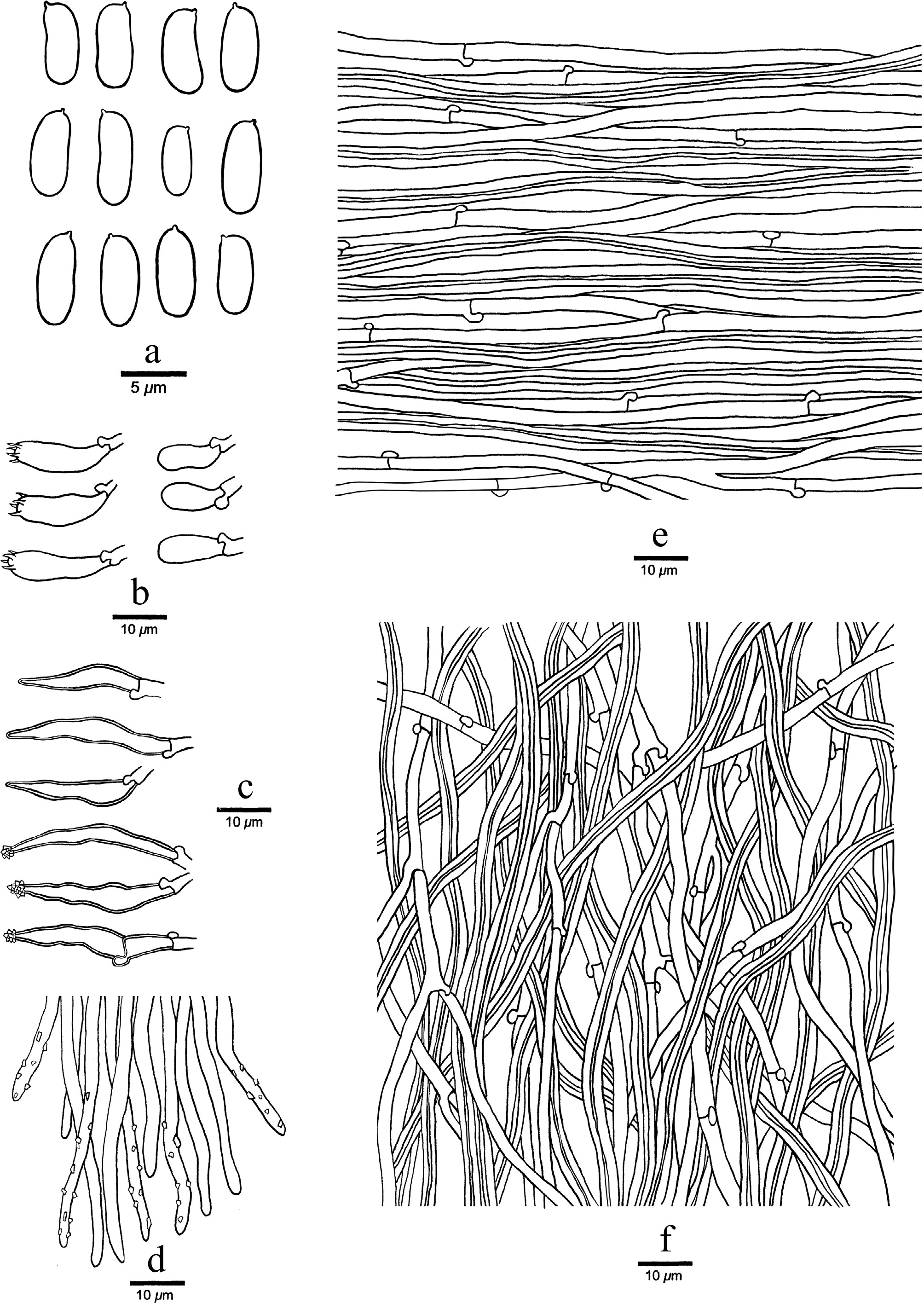
Figure 12.
Microscopic structures of Trichaptum longisporum (drawn from the holotype, JV 2311/13-J). (a) Basidiospores. (b) Basidia and basidioles. (c) Cystidia. (d) Hyphae at dissepiment edge. (e) Hyphae from context. (f) Hyphae from trama.
MycoBank number: MB 861995
Type – BOLIVIA. St. Cruz Dept., Refugio los Volcanes, on angiosperm stump, XI.2023, JV 2311/13-J (Holotype BJFC044759).
Etymology – Longisporum (Lat.): refers to the species having distinct long basidiospores.
Basidiomata – Perennial, pileate, sometimes resupinate; corky when fresh, hard corky when dry. Pilei applanate, usually imbricate or several specimens broadly attached to fused laterally in rows, projecting up to 2 cm, 6 cm wide, and 8 mm thick at base. Pileal surface distinctly flesh pink to rose when fresh, pinkish buff to grayish brown when dry, concentrically sulcate and radially striate, hispid to tomentose, more adpressed along the margin, faintly zonate; margin sharp, entire. Hymenophore poroid, ochre or wood-colored or pale vinaceous buff when dry; pores angular isodiametric, and without tendency to become lamellate towards the margin, 3–4(–5) per mm; dissepiments thin, entire. Context concolorous with tube, extremely thin, up to 0.5 mm. Tubes gray to ochraceous or milky coffee, up to 7.5 mm long.
Hyphal structure – Hyphal system dimitic; generative hyphae with clamp connections, hyaline, thin-walled; skeletal hyphae distinctly thick-walled to subsolid, slightly dextrinoid, CB–; tissues darkening in KOH.
Context – Generative hyphae hyaline, thin-walled, occasionally branched, 2–3 µm in diam.; skeletal hyphae thick-walled with a narrow lumen to subsolid, unbranched, loosely interwoven, 2–7 μm in diam.
Tubes – Generative hyphae hyaline, thin-walled, usually branched, 2–2.5 µm in diam.; skeletal hyphae thick-walled with a wide lumen, unbranched, loosely interwoven to subparallel, 2–3 μm in diam.; hyphae at dissepiment loosely encrusted with small crystals.
Hymenium – Cystidia narrow fusoid to subulate, thin- to slightly thick-walled, mostly with long narrow necks up to 5–10 × 1.5 μm, apically encrusted, sometimes with several separate, 20–30 × 3–4 µm; hyphal pegs present; basidia barrel-shaped, with four sterigmata and a basal clamp connection, 16.5–22 × 5.5–7 μm; basidioles similar to basidia in shape, but smaller.
Spores – Basidiospores cylindrical to slightly allantoid, hyaline, thin-walled, smooth, IKI–, CB–, (6–)6.3–8(–8.4) × (2.5–)2.7–3.6(–3.8) μm, L = 7.20μm, W = 3.21 μm, Q = 2.13–2.35 (n = 60/3).
Type of rot – White rot.
Distribution and ecology – Trichaptum longisporum is a common species found in northeastern South America, growing on angiosperm trunks.
Specimens examined – BRAZIL. Piauí, Caracol, 2011, URM 83620 & B530 (URM 83620 & BJFC032949). BOLIVIA, St. Cruz Dept., Refugio los Volcanes, XI. 2023, JV 2311/13-J (holotype BJFC044759), JV 2311/18-J (BJFC044760), JV 2311/19-J (BJFC044761); COSTA RICA. Guanacaste Prov., Rincón de la Vieja Volcano, Guachipelin, on fallen log, 10.III. 2022, JV2203/72-J; on fallen branch of Quercus, 12.III. 2022, JV 2203/104 (BJFC044762); ECUADOR. Arenillas, 17.II.2024, JV 2402/9-J; JAMAICA. IX. 2008, JV 0809/19-J (BJFC032949).
Key to known species of Trichaptum
-
1. Basidiomata usually resupinate T. resacarium 1. Basidiomata pileate to effused-reflexed 2 2. Basidiosporesr < 4 μm in length T. resacarium 2. Basidiospores > 4 μm in length T. cystidiolatum 3. Pores round to angular, 6–7 per mm T. perpusillum 3. Pores angular to irpicoid, < 5 per mm 4 4. Basidiospores > 4 μm in width T. anomalum 4. Basidiospores < 4 μm in width 5 5. Pore surface snuff brown to cigar brown T. perrottetii 5. Pore surface light grayish brown to wood-colored or pale vinaceous buff 6 6. Pores 3–5 per mm; basidiospores < 5.5 μm in length T. strigosum 6. Pores 0.5–3 per mm; basidiospores > 5.5 μm in length 7 7. Pore surface vinaceous brown, basidiospores 2–2.5 µm in width T. byssogenum 7. Pore surface cream to wood-colored, basidiospores 2.7–3.6 µm in width T. longisporum Podocarpioporus hydnoides Meng Zhou, Y.J. Cui, H. Zhao, Y.C. Dai & Yuan Yuan, sp. nov. (Figs 13,14)
-

Figure 13.
Basidiomata of Podocarpioporus hydnoides Dai 26968. Scale bars: (a) = 1 cm, (b) and (c) = 1 mm.
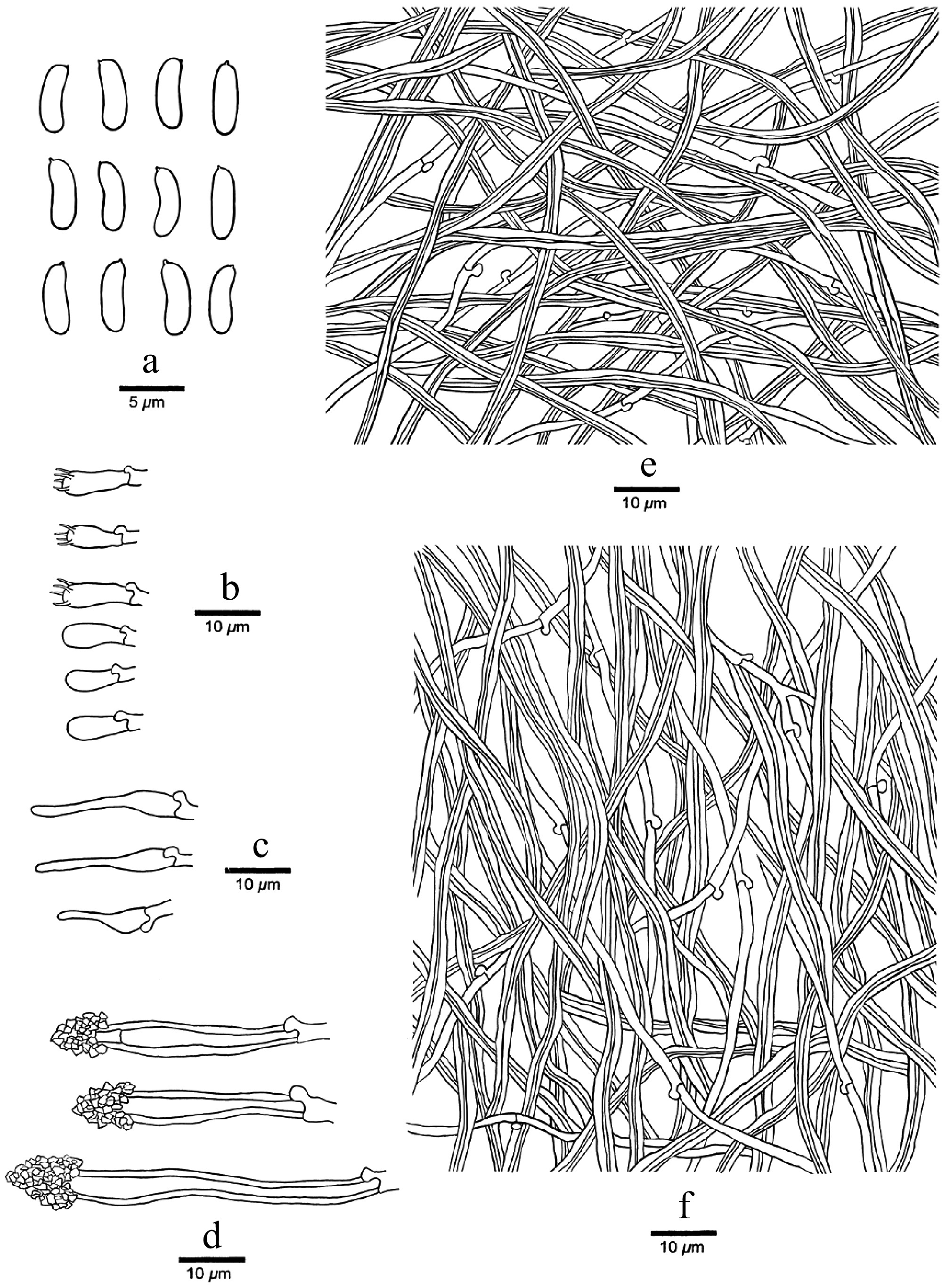
Figure 14.
Microscopic structures of Podocarpioporus hydnoides (Dai 26968). (a) Basidiospores. (b) Basidia and basidioles. (c), (d) Cystidia. (e) Hyphae from context. (f) Hyphae from trama.
MycoBank number: MB 861994
Type – CHINA. Xizang Auto. Reg., Linzhi County, on fallen trunk of Pinus yunnanensis, 27, Ⅹ, 2023 Dai 26968 (Holotype BJFC044520).
Etymology – Hydnoides (Lat.): refers to the species having a hydnoid hymenophore.
Basidiomata – Annual, resupinate, soft to soft leathery when fresh, becoming corky upon drying, up to 6 cm long, 4 cm wide, and 5 mm thick at the center. Hymenophore hydnoid, cream when fresh, buff to vinaceous brown when dry; teeth 4–6 per mm; dissepiments strongly lacerate. Context cream, up to 0.5 mm thick. Tubes pale brown, up to 1.5 mm long.
Hyphal structure – Hyphal system dimitic; generative hyphae with clamp connections, hyaline, thin-walled; skeletal hyphae distinctly thick-walled, slightly dextrinoid, CB+; tissues unchanged in KOH.
Context – Generative hyphae hyaline, thin- to thin-walled, unbranched, 1–2 µm in diam.; skeletal hyphae thick-walled with a narrow lumen, unbranched, interwoven, 1.5–3 μm in diam.
Tubes – Generative hyphae hyaline, thin-walled, occasionally branched, 1–2.5 µm in diam.; skeletal hyphae thick-walled, unbranched, interwoven, 2–3.5 μm in diam.
Hymenium – Cystidia clavate to fusoid, thin-walled, abundant, hyaline, thick-walled, apically encrusted, 37–80 × 5–6.5 µm; cystidioles present, fusoid with a long neck, thin-walled, smooth, 12–25 × 3–4 µm; basidia clavate, with four sterigmata and a basal clamp connection, 11–16 × 3.5–4.5 μm; basidioles similar to basidia in shape, but smaller.
Spores – Basidiospores cylindric to slightly allantoid, hyaline, thin-walled, smooth, IKI–, CB–, (4.3–)4.7–6.5 × (1.2–)1.3–2.1(–2.2) μm, L = 5.78 μm, W = 1.78 μm, Q = 3.20–3.35 (n = 90/3).
Type of rot – White rot.
Distribution and ecology – Podocarpioporus hydnoides to date, is found only in Xizang Auto. Reg. CHINA.
Specimens examined – CHINA. Xizang Auto. Reg., Linzhi county, on fallen trunk of Pinus yunnanensis, 27, Ⅹ, 2023, Dai 26909 (BJFC044460); Dai 26920 (BJFC044471); Dai 26968 (holotype BJFC044520); Dai 27008 (BJFC044560); Dai 27037 (BJFC044589); on charred wood of Pinus yunnanensis, Dai 26959 (BJFC044510).
Notes – Podocarpioporus hydnoides is characterized by narrow allantoid basidiospores, distinctly long subulate hymenial cystidia, which fit the definition of Podocarpioporus. Phylogenetically, four samples of Podocarpioporus hydnoides formed an independent lineage that is closely related to P. podocarpi. The latter differ from P. hydnoides in having a poroid to irpicoid hymenophore when juvenile and a lack of cystidioles.
Ancestral biogeography estimation
-
Dataset 3, comprising 52 specimens, was used to infer the historical biogeography and ancestral state of basidiomata of Trichaptum s.l. using RASP v.4.2[79,80] under the S-DEC and Bayesian Binary MCMC models. The biogeographical analyses indicated that Trichaptum s.l. has a complex evolutionary history, with its most recent common ancestor likely originating in Central and East Asia (Fig. 15, node 1). Throughout its evolutionary history, Trichaptum s.l. has experienced 49 global dispersal events, eight global vicariance events, one global extinction event, and 48 speciation events.
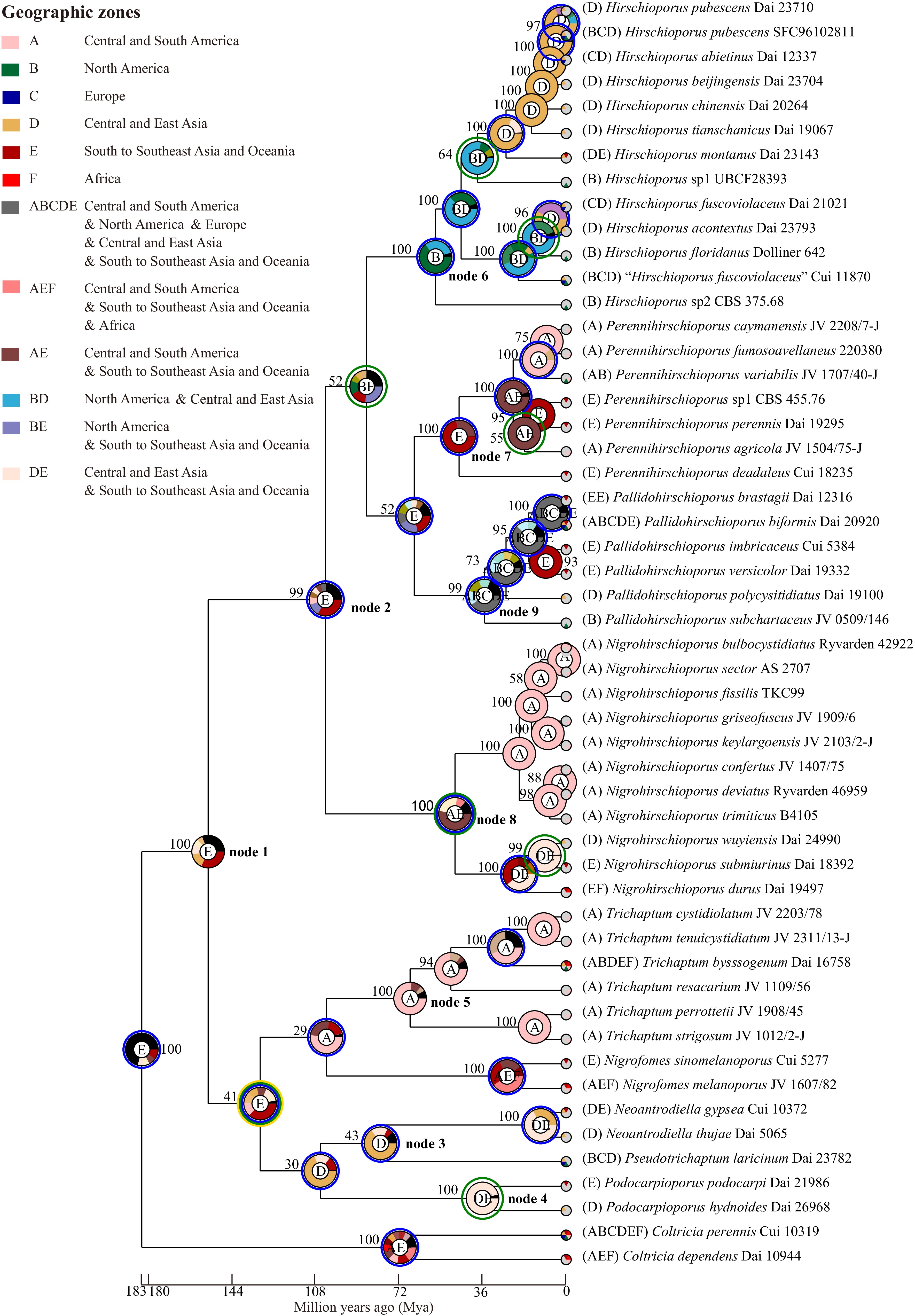
Figure 15.
The origin of geographical distribution of the Trichaptum s.l., A. Central and South America, B. North America, C. Europe, D. Central and East Asia, E. South to Southeast Asia and Oceania, and F. Africa. The phylogenetic tree was generated with Bayesian Inference algorithms using BEAST, while the trait of the pie chart at each node was evaluated using RASP under the Bayesian Binary MCMC model. The trait represented by each color and letter in the pie chart is indicated in the upper left with the supported values.
At the family level, Hirschioporaceae and Pseudotrichaptaceae are reconstructed with the highest probability as originating in Central and East Asian (Fig. 15, nodes 2 and 3), while the origin of Podocarpioporaceae (Fig. 15, node 4) remains ambiguous, with analyses showing comparable support for origins in Central and East Asia or South to Southeast Asia and Oceania. In contrast, Trichaptaceae appears to have an origin in Central and South America (Fig. 15, node 5).
At the genus level, Pseudotrichaptum, Podocarpioporus, and Trichaptum align with their respective families, as each corresponds to a monotypic family. The genera within Hirschioporaceae exhibit distinct biogeographical origins: Hirschioporus likely originated in North America (Fig. 15, node 6), Perennihirschioporus in South to Southeast Asia and Oceania (Fig. 15, node 7), Nigrohirschioporus shows a primary probability for a Central and South American origin (Fig. 15, node 8), alternative scenarios involving Central and East Asia cannot be ruled out. This uncertainty likely reflects the complex pantropical distribution of these lineages. Lastly, Pallidohirschioporus is inferred to have origins in regions excluding Africa (Fig. 15, node 9).
Climate distribution patterns estimation
-
A total of 44 global dispersal events and eight global vicariance events were inferred using RASP v.4.2[79,80] under the S-DEC and Bayesian Binary MCMC models. The most recent common ancestor of Trichaptum s.l. was likely distributed in tropical regions, with subsequent dispersal into subtropical, tropical, and mountain zones (Fig. 16 node 1). Hirschioporaceae is most likely of tropical origin (Fig. 16 node 2), while Pseudotrichaptaceae (with the single genus Pseudotrichaptum) is inferred to have originated in temperate to subtropical zones (Fig. 16 node 3). Both Podocarpioporaceae (with the single genus Podocarpioporus) and Trichaptaceae (with the single genus Trichaptum) are inferred to have originated in subtropical to tropical zones (Fig. 16 nodes 4 and 5). Within Hirschioporaceae, Hirschioporus likely originated in temperate zones (Fig. 16 node 6), while Nigrohirschioporus and Perennihirschioporus are inferred to have originated in tropical zones (Fig. 16 nodes 7 and 8). The origin of Pallidohirschioporus remains unresolved, with all climatic zones being equally plausible (Fig. 16 node 9).
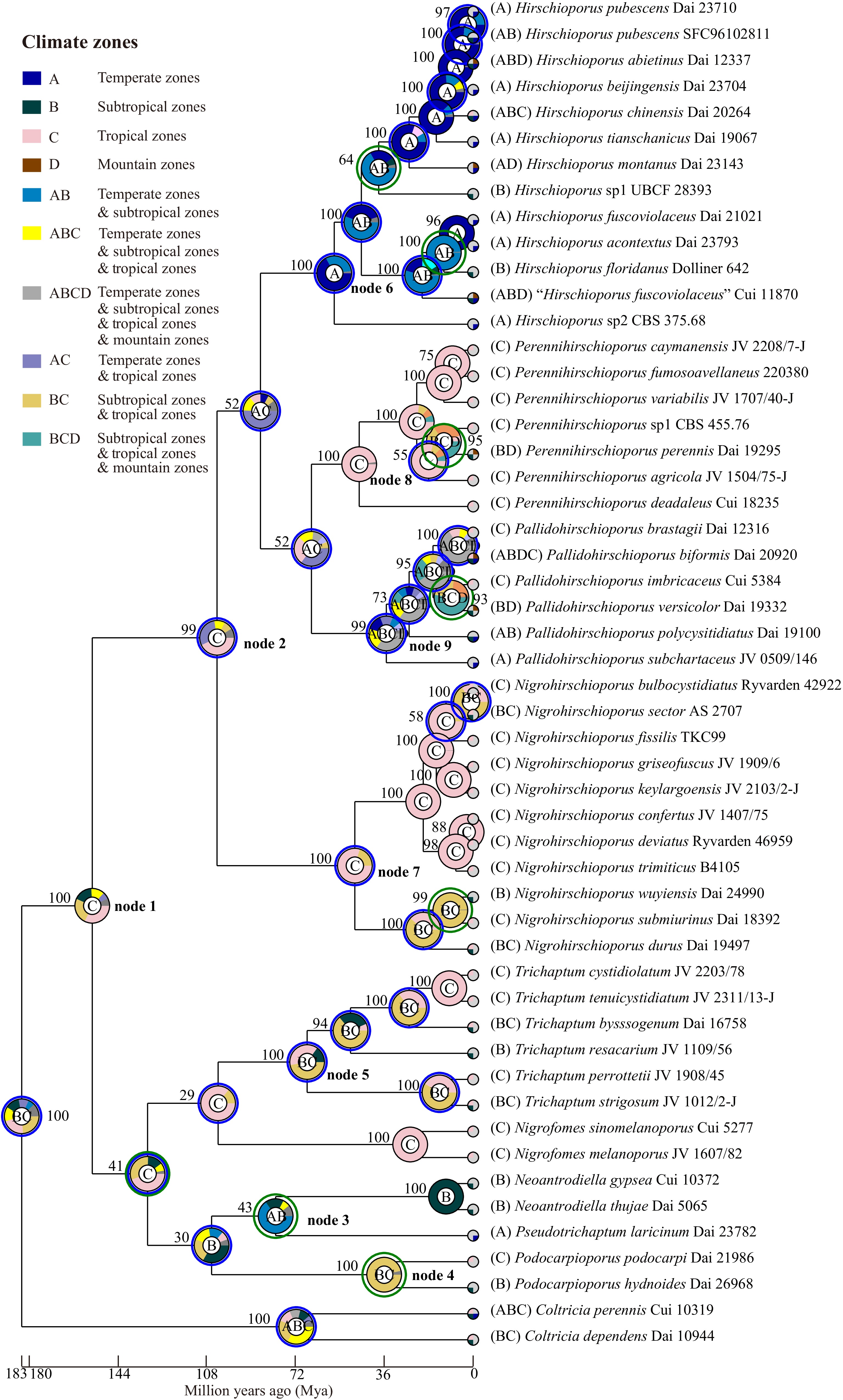
Figure 16.
The origin of climate distribution of the Trichaptum s.l., A. Temperate zones, B. Subtropical zones, C. Tropical zones, and D. Mountain zones. The phylogenetic tree was generated with Bayesian Inference algorithms using BEAST, while the trait of the pie chart at each node was evaluated using RASP under the Bayesian Binary MCMC model. The trait represented by each color and letter in the pie chart is indicated in the upper left with the supported values.
Ancestral characters evolution
-
Ancestral character reconstruction was used to examine the evolutionary configuration of basidiomata in Trichaptum s.l. using RASP v.4.2[79,80] under the S-DEC and Bayesian Binary MCMC models. RASP analyses suggested that the ancestral morphology of Trichaptum s.l. most likely consisted of a coriaceous consistency with a tomentose surface or apileate (Fig. 17). However, node support values indicate some uncertainty at deeper nodes, suggesting that alternative ancestral states (e.g., soft corky consistency) are possible. Despite this ambiguity, a general trend toward increasing basidiomata hardness is observable throughout the evolutionary history of the group.
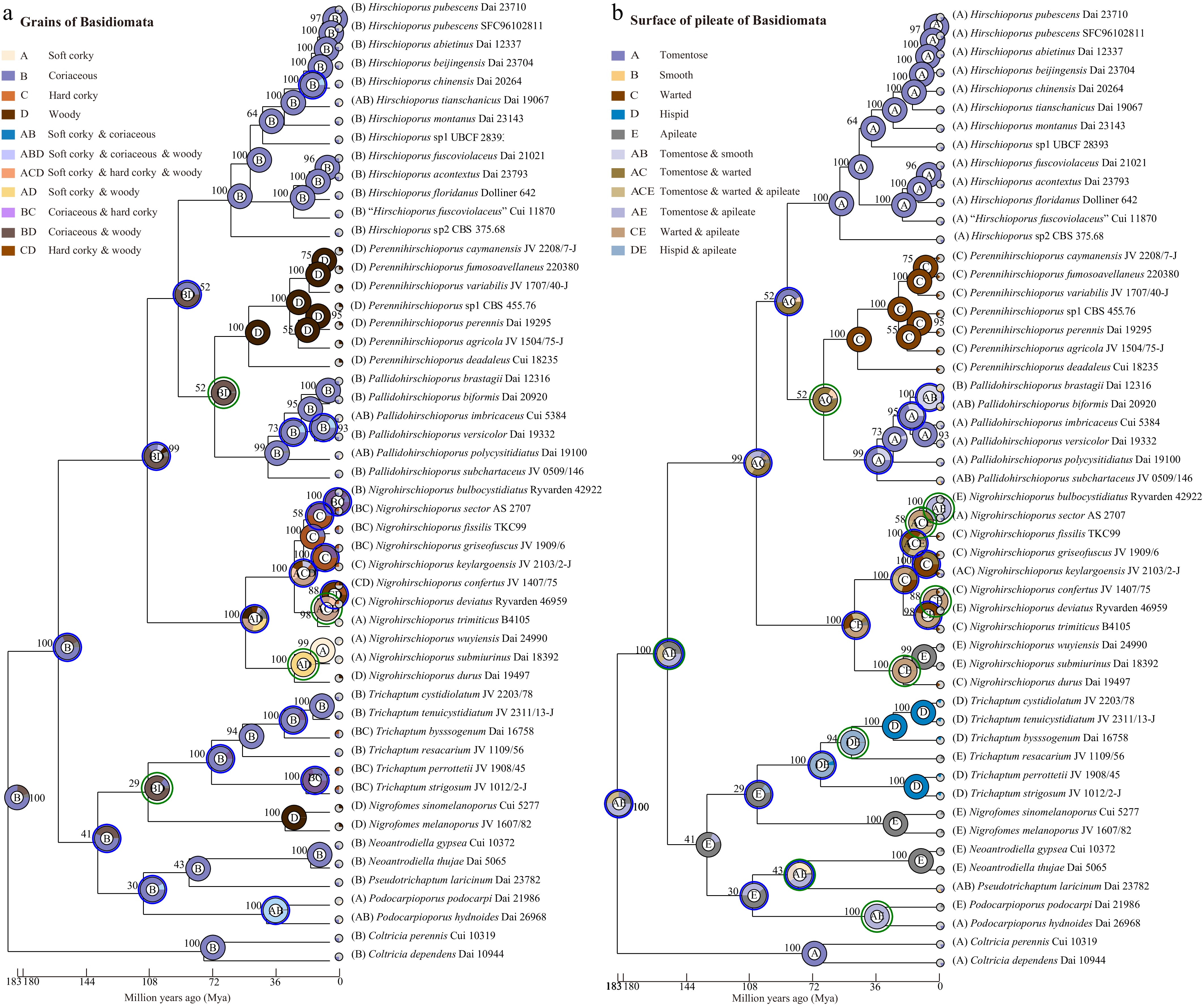
Figure 17.
The configuration evolution of basidiomata of Trichaptum s.l., (a) consistency and (b) surface of pileus. The phylogenetic tree was generated with Bayesian Inference algorithms using BEAST, while the trait of the pie chart at each node was evaluated using RASP under the Bayesian Binary MCMC model. The trait represented by each color and letter in the pie chart is indicated in the upper left with the supported values. Four types of consistency, A. Soft corky to brittle when dry, B. Coriaceous, C. Hard corky, and D. Woody hard. Five types of surfaces of pileate, A. Tomentose, B. Smooth, C. Warted, D. Hispid, and E. Apileate.
Regarding basidiomata consistency, the ancestral state of Hirschioporaceae was inferred to be either coriaceous or woody hard, whereas that of Pseudotrichaptaceae and Trichaptaceae was most likely coriaceous. The ancestral consistency of Podocarpioporaceae was inferred to be either soft corky (becoming brittle when dry) or coriaceous. Within Hirschioporaceae, the ancestral consistency was reconstructed as coriaceous for Hirschioporus, Pallidohirschioporus, and Perennihirschioporus, while for Nigrohirschioporus it was either woody hard or soft, corky to brittle when dry.
The ancestral surface morphology of the pileal surface in Hirschioporaceae was likely tomentose or warted. In Pseudotrichaptaceae and Podocarpioporaceae, the ancestral condition is inferred to be tomentose or apileate. Trichaptaceae likely originated with a hispid surface or was apileate. Within Hirschioporaceae, the ancestral morphology was reconstructed as tomentose for Hirschioporus and Pallidohirschioporus, warted or apileate for Nigrohirschioporus, and warted for Perennihirschioporus.
Species distribution of Trichaptum s.l. and environmental factors
-
To investigate the distribution patterns of each genus within the broad Trichaptum s.l., global occurrence data at the genus level were compiled and categorized, based on the revised taxonomic system of Trichaptum s.l. revised in this study (Fig. 18, Table 5). This dataset integrated both publicly available records and data from specimens examined herein. Subsequently, a comprehensive suite of environmental variables was screened to identify the primary climatic drivers for each genus. The objective was to determine the factors with the highest contribution rates in the models, through which the adaptive relationships between the fungal lineages and their climatic niches could be explored.
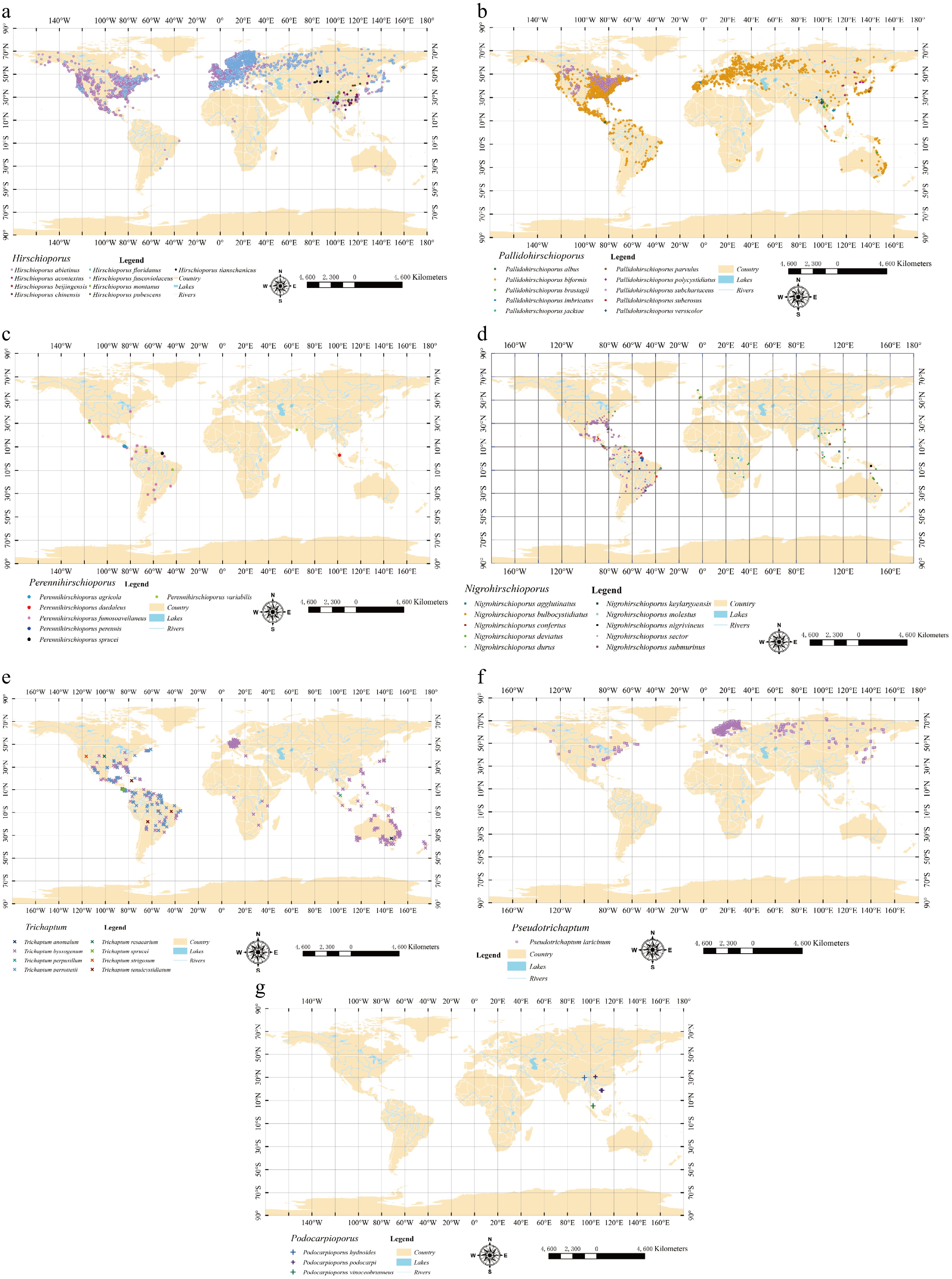
Figure 18.
Global distribution map of Trichaptum s.l. genera based on spatially thinned and verified GBIF occurrence records. (a) Hirschioporus, (b) Pallidohirschioporus, (c) Perennihirschioporus, (d) Nigrohirschioporus, (e) Trichaptum, (f) Pseudotrichaptum, and (g) Podocarpioporus. Source: World Map (1:10,000,000), the Natural Earth website,
www.naturalearthdata.com .Table 5. Contribution of various environmental variables to the distribution of Trichaptum s.l.
Genera Abbreviation Environmental factors Units Rate of
contribution (%)Permutation
importance (%)Hirschioporus Bio14 Precipitation of driest month mm 51.1 0.8 Bio12 Annual precipitation mm 23.4 18.3 Bio1 Annual mean temperature 6 4.6 Bio11 Mean temperature of coldest quarter °C 6 1 Bio6 Maximum temperature of warmest month °C 3.8 7.8 Bio5 Maximum temperature of warmest month °C 2.9 29.3 Bio7 Temperature annual range (Bio5-Bio6) °C 1.9 5.7 Bio19 Precipitation of coldest quarter mm 0.9 8.7 Bio10 Mean temperature of warmest quarter °C 1.2 8.5 Bio16 Precipitation of wettest quarter mm 0.3 5.3 Pallidohirschioporus Bio14 Precipitation of driest month mm 41.5 0.7 Bio12 Annual precipitation mm 22 5.2 Bio17 Annual mean temperature mm 11.3 13.6 Bio 5 Maximum temperature of warmest month °C 7.8 25.5 Bio11 Mean temperature of coldest quarter °C 7.5 10.1 Bio10 Mean temperature of warmest quarter °C 0.9 18.4 Bio16 Precipitation of wettest quarter mm 0.5 9.2 Bio19 Precipitation of coldest quarter mm 0.4 7.4 Perennihirschioporus Bio16 Precipitation of wettest quarter mm 58.7 3.9 Bio3 Isothermality (Bio2/Bio7) (× 100) C of V 10.8 10.3 Bio4 Temperature seasonality (standard deviation x 100) °C 7.1 2.3 Bio18 Precipitation of warmest quarter mm 2.6 5.4 Bio19 Precipitation of coldest quarter mm 4.8 12.5 Bio7 Temperature annual range (Bio5-Bio6) °C 1 10.7 Bio8 Mean temperature of wettest quarter °C 0.7 14.6 Bio9 Mean temperature of driest quarter °C 0.4 10.7 Nigrohirschioporus Bio12 Annual precipitation mm 61.3 5.8 Bio3 Isothermality (Bio2/Bio7) (× 100) C of V 13.3 10.7 Bio11 Mean temperature of coldest quarter °C 6.9 46.3 Bio14 Precipitation of driest month mm 6.6 5.8 Bio4 Temperature seasonality (standard deviation x 100) °C 1.2 5.2 Trichaptum s.s. Bio6 Minimum temperature of coldest month °C 20.1 0.1 Bio12 Annual precipitation mm 20.1 11.2 Bio14 Precipitation of driest month mm 15.3 4.5 Bio3 Isothermality (Bio2/Bio7) (× 100) C of V 12 8.2 Bio19 Precipitation of coldest quarter mm 7.7 4.2 Bio4 Temperature seasonality (standard deviation x 100) °C 4.7 10 Bio18 Precipitation of warmest quarter mm 3.5 8.8 Bio7 Temperature annual range (Bio5-Bio6) °C 2.3 5.3 Bio13 Precipitation of wettest month mm 0.8 5.6 Pseudotrichaptum Bio14 Precipitation of driest month mm 57.6 1.1 Bio1 Annual mean temperature °C 17.2 8.9 Bio5 Max temperature of warmest month °C 7.7 42.3 Bio18 Precipitation of warmest quarter mm 4.1% 6 Bio4 Temperature seasonality (standard deviation x 100) C of V 2.5% 15.1 Bio8 Mean temperature of wettest quarter °C 1.2 6.3 Bio20 Elevation mm 0.7 8.8 Bio10 Mean temperature of warmest quarter °C 0.5 5.9 Genus Hirschioporus
-
The genus Hirschioporus exhibits a North Temperate distribution pattern, with its species primarily inhabiting coniferous wood throughout Eurasia, North America, and the Himalayas. Species within this genus are typically saprophytic, functioning as pioneer decomposers that colonize dead trees and newly fallen logs.
The distribution of the genus is primarily concentrated in temperate and subtropical regions. However, influenced by geographical and historical factors, its range has expanded southward along mountain ranges to occupy alpine zones in the tropics and temperate regions of the Southern Hemisphere. Nevertheless, our ancestral biogeography analysis reveals that the ancestral origin and center of distribution for Hirschioporus remains in the North Temperate zone (Fig. 15). The hosts of Hirschioporus are exclusively gymnosperms, primarily pine (Pinus spp.), spruce (Picea spp.), fir (Abies spp.), and larch (Larix spp.). For the environmental niche analysis, we aggregated occurrence data from the GBIF and field specimens collected in this study. The initial dataset of 45,937 records was refined by removing duplicate entries and applying a spatial thinning filter with a 2.5 arc-min resolution. This process resulted in a final dataset of 36,607 unique occurrence points, which were used to model environmental factors. The global distribution of these points is mapped in Fig. 18a.
Table 5 presents the average relative contributions of the predictor variables from the Maxent model (i.e., those with a contribution > 5%, permutation importance > 5%). The species distribution model for the genus Hirschioporus achieved a fair predictive performance, with an average Area Under the Curve (AUC) value of 0.693 ± 0.003 across 15 replicate runs (Supplementary Fig. S1a). The analysis of variable contributions identified climatic factors related to summer thermal stress and annual moisture availability as the primary drivers of the genus's distribution (Table 5). According to the permutation importance metric, the maximum temperature of the warmest month (Bio5) was the most critical limiting factor, contributing 29.3% to the model's predictive power. The second most important variable was annual precipitation (Bio12), with a permutation importance of 18.3%. While precipitation of the driest month (Bio14) showed the highest percent contribution (51.1%), its negligible permutation importance (0.8%) suggests its influence is likely an artifact of its correlation with other more causally-related variables. The response curves for the key environmental variables further defined the ecological niche of Hirschioporus (Supplementary Fig. S1). The genus exhibited a clear intolerance to high summer heat, with habitat suitability peaking where the maximum temperature of the warmest month (Bio5) was between 20–25°C, and declining sharply thereafter (Supplementary Fig. S1b). The model also demonstrated a strong positive response to moisture, with suitability increasing steadily with annual precipitation (Bio12) (Supplementary Fig. S1c). The Jackknife test indicates that annual mean temperature (Bio1) is the variable with the greatest amount of information when used alone (Supplementary Fig. S1e). The response toBio1 showed a broad optimum centered around 0–15°C, consistent with adaptation to temperate climates (Supplementary Fig. S1d).
Genus Pallidohirschioporus
-
The genus Pallidohirschioporus is widely distributed across temperate and subtropical regions worldwide, with documented occurrences in North America, Europe, Asia, and Australia. It primarily exhibits a temperate to subtropical distribution pattern. While individual species within the genus show varied host and climatic preferences, the majority are found on angiosperm wood, including oak (Quercus spp.), birch (Betula spp.), poplar (Populus spp.), and plum (Prunus spp.). The main distribution pattern is exemplified by temperate-subtropical species such as P. biformis, P. jackiae, P. parvulus, P. polycystidiatus, P. subchartaceus, and P. versicolor. However, a subset of species, including P. albus, P. brastagii, P. imbricatus, and P. suberosus, is distributed in tropical regions.
According to GBIF records and field data from specimens collected for this study, there was a total of 53,793 valid occurrence records for Pallidohirschioporus species. After removing duplicates and overlapping points within a 2.5 arc-minute resolution, 10,393 occurrence points were retained to screen for the primary environmental factors influencing this fungal genus. Its global distribution map is shown in Fig. 18b. The Maxent model developed for Pallidohirschioporus demonstrated good predictive accuracy, achieving an average test AUC of 0.788 ± 0.004. The analysis of environmental variable contributions identified a combination of seasonal temperature and precipitation factors as primary drivers. According to permutation importance, the most influential variables were the maximum temperature of the warmest month (Bio5; 25.5%), the mean temperature of the warmest quarter (Bio10; 18.4%), the precipitation of the driest quarter (Bio17; 13.6%), and the mean temperature of the coldest quarter (Bio11; 10.1%) (Table 5).
Response curves revealed that the genus thrives in warm conditions, with habitat suitability peaking when the maximum temperature of the warmest month (Bio5) is approximately 30 °C (Supplementary Fig. S2c). The genus also requires moisture, showing a strong positive response to annual precipitation (Bio12) and a preference for regions where the precipitation of the driest quarter (Bio17) remains above a minimum threshold (Supplementary Fig. S2b, S2d). Furthermore, its distribution is limited by cold winters, as suitability drops sharply when the mean temperature of the coldest quarter (Bio11) falls below 0 °C (Supplementary Fig. S2e).
Genus Perennihirschioporus
-
The genus Perennihirschioporus is distributed in tropical and subtropical regions, primarily within semi-arid climate zones. The genus exhibits a tropical Asian-American disjunct distribution pattern, meaning its species are found in the warm regions of Asia and the Americas, but not in the intervening areas. In the Eastern Hemisphere, its range may extend from Asia to northeastern Australia and potentially to the islands of the southwestern Pacific. Species of Perennihirschioporus grow on various angiosperm wood. According to GBIF records and field data from specimens collected for this study, there was a total of 70 valid occurrence records for Perennihirschioporus species. After removing duplicates and overlapping points within a 2.5 arc-min resolution, 43 occurrence points were retained to screen for the primary environmental factors influencing this fungal genus. Its global distribution map is shown in Fig. 18c.
The Maxent species distribution model for Perennihirschioporus achieved excellent predictive accuracy, with an average test AUC of 0.924 ± 0.020. The high performance of the model suggests a highly specialized and narrowly defined climatic niche for the genus. The analysis of environmental variable contributions revealed that precipitation of the wettest quarter (Bio16) was the overwhelmingly dominant factor driving the distribution, accounting for 58.7% of the model's predictive power (Table 5). The next most important variables, isothermality (Bio3; 10.8%) and temperature seasonality (Bio4; 7.1%), also point to the importance of a stable tropical climate. The critical role of Bio16 was further confirmed by the Jackknife test, which showed it possessed the highest gain when used in isolation by a large margin (Supplementary Fig. S3). Response curves indicated that the habitat suitability of Perennihirschioporus increases dramatically with precipitation of the wettest quarter (Bio16), peaking in regions that receive 1,000–2,000 mm of rain during the wet season (Supplementary Fig. S3b). The genus also showed a clear preference for areas with high isothermality (Bio3) and low temperature seasonality (Bio4), characteristics of climates with minimal temperature fluctuation (Supplementary Fig. S3c, S4d).
Genus Nigrohirschioporus
-
The genus Nigrohirschioporus, a group of poroid fungi colonizing angiosperm wood, exhibits a classic pantropical rainforest distribution pattern, which was concentrated in the world's three major tropical rainforest basins: the Amazon Basin in South America, the Congo Basin in Africa, and the Malesian region in Southeast Asia.
According to GBIF records and field data from specimens collected for this study, there was a total of 628 valid occurrence records for Nigrohirschioporus species. After removing duplicates and overlapping points within a 2.5 arc-min resolution, 510 occurrence points were retained to screen for the primary environmental factors influencing this fungal genus. Its global distribution map is shown in Fig. 18d. The Maxent model for the genus Nigrohirschioporus achieved excellent predictive accuracy, with a high average test AUC of 0.954 ± 0.004. This result indicates a well-defined and highly specialized climatic niche. The analysis of variable importance identified the mean temperature of the coldest quarter (Bio11) as the primary limiting factor for the genus's distribution. It had the highest permutation importance by a large margin, at 46.3% (Table 5). While annual precipitation (Bio12) had the highest percent contribution (61.3%), its lower permutation importance (5.8%) suggests it is a necessary but less critical limiting factor at the global scale compared to thermal constraints. Response curves illustrated the genus's strict climatic requirements (Supplementary Fig. S4). Habitat suitability was highest where the mean temperature of the coldest quarter (Bio11) remained above 18 °C, dropping precipitously at lower temperatures (Supplementary Fig. S4b). The genus also showed a strong positive response to high annual precipitation (Bio12) and high isothermality (Bio3), underscoring its adaptation to stable, consistently warm, and humid environments (Supplementary Fig. S4c, S4d).
Genus Trichaptum s.s.
-
The genus Trichaptum exhibits a pantropical distribution pattern, with a majority of its species occurring in tropical and subtropical regions. Ecologically, it is primarily found on angiosperm wood. According to GBIF records and field data from specimens collected for this study, there was a total of 555 valid occurrence records for Trichaptum s.s. species. After removing duplicates and overlapping points within a 2.5 arc-min resolution, 554 occurrence points were retained to screen for the primary environmental factors influencing this fungal genus. Its global distribution map is shown in Fig. 18e.
The Maxent model for the genus Trichaptum achieved excellent predictive accuracy, with a high average test AUC of 0.941 ± 0.006. This result indicates a well-defined and highly specialized climatic niche for the genus. The analysis of variable contributions identified a combination of thermal limits and moisture availability as the primary drivers of distribution. Minimum temperature of coldest month (Bio6) and annual precipitation (Bio12) showed the highest percent contributions (both 20.1%). However, permutation importance analysis highlighted the mean temperature of the warmest quarter (Bio10) as the most critical variable (18.0% importance), followed by annual precipitation (Bio12, 11.2%) (Table 5). Response curves illustrated the genus's strict climatic requirements (Supplementary Fig. S5). Habitat suitability was strongly limited by winter cold, dropping to near zero when the minimum temperature of the coldest month (Bio6) fell below freezing (Supplementary Fig. S5b). The genus also requires high moisture, with suitability increasing with annual precipitation (Bio12) (Supplementary Fig. S5c). Furthermore, Trichaptum thrives in warm climates, showing a clear preference for areas where the mean temperature of the warmest quarter (Bio10) exceeds 25 °C (Supplementary Fig. S5d).
Genus Pseudotrichaptum
-
The genus Pseudotrichaptum comprises species that primarily inhabit coniferous trees. It exhibits a circumboreal distribution, with species widely distributed throughout the coniferous forests of the Northern Hemisphere. These fungi demonstrate low host specificity, capable of colonizing the wood of fir (Abies), larch (Larix), spruce (Picea), and pine (Pinus). According to GBIF records and field data from specimens collected for this study, there was a total of 6,242 valid occurrence records for Pseudotrichaptum species. After removing duplicates and overlapping points within a 2.5 arc-min resolution, 4,764 occurrence points were retained to screen for the primary environmental factors influencing this fungal genus. Its global distribution map is shown in Fig. 18f.
For our model, the mean value of the average AUC for the training set is 0.892 with a low standard deviation of 0.002, which indicated favorable model performance. The average relative contributions of the predictor variables from the Maxent model (more than 5%) are presented in. The analysis of variable importance identified temperature as the primary driver of the genus's distribution. According to permutation importance, the most critical variables were the maximum temperature of the warmest month (Bio5; 42.3% importance) and temperature seasonality (Bio4; 15.1%). Elevation also emerged as a significant factor (8.8% importance) (Table 5). Response curves illustrated the species' adaptation to cold climates (Supplementary Fig. S6). Habitat suitability peaked at an annual mean temperature (Bio1) near 0 °C (Supplementary Fig. S6b). The species demonstrated a strong intolerance to heat, with suitability dropping sharply when the maximum temperature of the warmest month (Bio5) exceeded 20 °C (Supplementary Fig. S6c). This thermal limitation defines its southern distribution boundary. Furthermore, its habitat suitability showed a positive correlation with elevation (Supplementary Fig. S6d), confirming its preference for high-altitude environments.
Genus Podocarpioporus
-
Podocarpioporus grows on the wood of Dacrydium, a genus within the Podocarpaceae family. The Podocarpaceae family itself is distributed in tropical and subtropical regions, with genera such as Dacrydium being sources of high-quality timber. Podocarpioporus exhibits a remarkable host specificity, growing exclusively on members of the Podocarpaceae, which strictly limits the dispersal. The primary host genus, Dacrydium, is mainly distributed in the mountainous regions of central and southern Hainan, China, as well as in Malaysia, Vietnam, Laos, Cambodia, Myanmar, and Thailand.
-
Trichaptum s.l. was established in 1904, belonging to Hymenochaetales of Agaricomycetes within Basidiomycota, but its family-level classification has been controversial, and it has been considered a genus as incertae sedis[81−86]. The establishment of Trichaptum s.l. as a complex comprising two families and seven genera resolved long-standing taxonomic uncertainties[39]. The family Hirschioporaceae consisted of four genera, including Hirschioporus, Nigrohirschioporus, Pallidohirschioporus, and Perennihirschioporus, and Trichaptaceae included a single genus Trichaptum. However, previous studies focused on Asia, and the two genera Podocarpioporus and Pseudotrichaptum have not clearly defined their family-level taxonomic status.
In this study, four new species, Nigrohirschioporus keylargoensis, Perennihirschioporus caymanensis, Trichaptum cystidiolatum, and T. longisporum, collected from the subtropical to tropical North and South America, are described, greatly enriching the species diversity of Trichaptum s.l. in these regions. Additionally, two new species, Nigrohirschioporus wuyiensis and Podocarpioporus hydnoides are reported from temperate and subtropical China. The discovery of six new species from the Americas and Asia—supported by distinct molecular and morphological evidence—underscores the underestimated diversity of this group in tropical and subtropical regions.
Hirschioporaceae
-
In the phylogenetic analyses, two samples of Nigrohirschioporus keylargoensis formed a highly supported lineage (99/MP, 1/BI), which is sister to N. griseofuscus, and closely related to N. fissilis and N. sector. However, N. griseofuscus differs from N. keylargoensis in having a dimitic hyphal system and narrower basidiospores (Q = 3.17–3.30 vs. Q = 2.03–2.95). Both N. fissilis and N. sector differ from N. keylargoensis in having a regularly poroid hymenophore and a dimitic hyphal system. Morphologically, Nigrohirschioporus keylargoensis may be confused with N. molestus with hydnoid to irpicoid hymenophore and trimitic hyphal system; however, N. molestus is distinguished from N. keylargoensis by having smaller pores (3–4 per mm vs 1–2 per mm) and shorter ellipsoid basidiospores (4–4.8 × 3–3.5 µm vs 5.1–7.0 × 2–3 µm).
Nigrohirschioporus wuyiensis is recovered as an independent lineage sister to N. submurinus. Morphologically, Nigrohirschioporus wuyiensis may also be confused with N. submurinus and N. deviatus in its completely resupinate basidiomata and small pores. However, N. submurinus differs from N. wuyiensis in having smaller pores (6–9 per mm vs 4–6 per mm) and smooth fusoid cystidia with a long neck. N. deviatus is distinguished from N. wuyiensis by its narrow basidiospores (< 2μm in width vs > 2μm in width) and its angular or even split pore surface with poroid to irpicoid hymenophore.
Phylogenetically, Perennihirschioporus caymanensis forms an independent lineage that is related to P. agricola, P. fumosoavellaneus, and P. variabilis. Perennihirschioporus variabilis was originally described from Venezuela[87], and later found in French Guiana (JV1909/4-J). It has sometimes similar pilei as P. caymanensis, with poroid, irpicoid to daedaleoid hymenophore, but never lamellate, and basidiospores are bigger in P. variabilis than in P. caymanensis (4.5–6 × 2.2–2.5 μm vs 3.5–4.3 × 2–2.7 μm). Perennihirschioporus agricola, P. fumosoavellaneus have poroid hymenophores that are readily distinguished from P. caymanensis. P. daedaleus may also be confused with P. caymanensis in its perennial, pileate hymenophores. However, P. daedaleus is distantly related to P. caymanensis phylogenetically and is only found in Malaysia, which is far away from P. caymanensis geographically. Morphologically, P. daedaleus differs from P. caymanensis in having typically triquetrous basidiomata, with daedaleoid to hydnoid hymenophore and thinner context (0.1 mm vs 4 mm).
Trichaptaceae
-
In the phylogenetic analyses, three sequenced samples of Trichaptum cystidiolatum formed an independent lineage, closely related to T. byssogenum and T. longisporum. The latter two species differ from T. cystidiolatum in having distinctly pileate basidiomata. In addition, T. longisporum differs from T. cystidiolatum by smaller pores (3–4 per mm vs 1–3 per mm) and larger basidiospores (6.3–8 × 2.7–3.6 μm vs 4.5–6 × 2.2–2.9 μm). Morphologically, T. cystidiolatum and T. resacarium share similar resupinate to effused-reflexed basidiomata. However, T. resacarium is distinct by dark, greyish colors and smaller basidiospores (3.5–4 × 2 μm vs 4.5–6 × 2.2–2.9 μm).
Trichaptum cystidiolatum is so far only found in Costa Rica, but the same sequence, however, is present several times in GenBank as 'Uncultured fungus' (see Discussion). Ryvarden & Johansen[81] show a picture of Trametes versatilis Berk 1842 features (synonym of Trichaptum byssogenum, collected in the USA), where the spores correspond perfectly to our T. cystidiolatum spores. We have not seen its type in Kew, and so we cannot decide now if our T. cystidiolatum is identical with Trametes versatilis or not.
Phylogenetically, five samples of Trichaptum longisporum formed an independent lineage, which is closely related to T. cystidiolatum and T. byssogenum. Morphologically, T. byssogenum has a vinaceous brown pore surface and narrower basidiospores (2–2.5 µm in width vs 2.7–3.6µm in width) that can be distinguished from T. longisporum. Both Funalia bouei and Polystictus hariotianus are synonyms of T. byssogenum from South America, which has large pores (2 per mm) in the original description and is distinctly different from the small pores (3–4 per mm) of T. longisporum. T. cystidiolatum differs from T. longisporum by its effused-reflexed or resupinate basidiomata, larger pores (1–3 per mm vs 3–4 per mm), and smaller basidiospores (4.5–6 × 2.2–2.9 μm vs 6.3–8 × 2.7–3.6 μm).
Podocarpioporaceae
-
Podocarpioporus hydnoides is characterized by narrow allantoid basidiospores, distinctly long subulate hymenial cystidia, which fit the definition of Podocarpioporus. Phylogenetically, four samples of Podocarpioporus hydnoides formed an independent lineage that is closely related to P. podocarpi. The latter differ from P. hydnoides in having a poroid to irpicoid hymenophore when juvenile and a lack of cystidioles.
Evolutionary timeline and biogeographical history
Estimating the divergence times of Trichaptum s.l.
-
Currently, phylogenetic and morphological analyses are the main criteria for fungal species identification, and clock-dating analysis has gradually become a key basis for defining higher taxonomic units in fungi due to its important role in inferring fungal divergence times[17−19,21]. For example, He et al.[19] suggested that the divergence times within the phylum Basidiomycota for subphyla, classes, orders, and families are estimated to be 443–490 Mya, 312–412 Mya, 102–361 Mya, and 50–289 Mya, respectively. In recent years, the estimated divergence time of the order Hymenochaetales has been showed to range from 167 to 289 Mya[17,35,39,88]. Varga et al.[17] suggests that the rapid radiation of the orders within the class Agaricomycetes occurred during the Jurassic period. Additionally, Zhao et al. estimated that the divergence time of families within Hymenochaetales occurred between 123 and 163 Mya, corresponding to the Early Cretaceous to Late Jurassic periods[35]. These studies laid the foundation for estimating the divergence times of Trichaptum s.l. Zhou et al. estimated the mean stem ages of the families Hirschioporaceae and Trichaptaceae as 121 Mya and 103 Mya[39], respectively, which is in agreement with the results of this study. The mean stem ages of the four families within Trichaptum s.l. ranged from 71 Mya to 116 Mya, overlapping with results from previous studies[19,35,39,88] and supporting the legitimacy of the two newly proposed families. Meanwhile, the families in Trichaptum s.l. occurred during the Cretaceous, whereas individual species originated mainly during the Eocene to the Pleistocene, a period when the present-day continental configuration had largely taken shape. During this time, the global climate was warm and humid, and the diversity of both gymnosperms and angiosperms increased, providing favorable climatic conditions and abundant host trees that facilitated the rapid diversification of macrofungi[17,21,35,89−92].
The integration of divergence time estimates with biogeography and morphology represents an important approach for reconstructing the origin and evolution of macrofungi, as demonstrated in previous studies[17,21,35]. The centers of origin of macrofungi appear to be complex, with some lineages likely originating in Eurasia, such as Onnia, Phaeolus, Porodaedalea, and Heterobasidion, while others may have originated in Africa or the Americas, such as ectomycorrhizal Russulaceae and Hirschioporaceae[21,32,35,93−95]. In this study, the biogeographic reconstruction of Trichaptum s.l. revealed complex paleogeographic processes. Early divergences around 100–120 Mya coincide with the fragmentation of Gondwana, supporting possible African or South American origins for lineages such as Trichaptaceae[96]. In contrast, the wide distribution of Hirschioporaceae across the Northern Hemisphere may be associated with Laurasian connections and intercontinental dispersals facilitated by land bridges such as Beringia or the North Atlantic Land Bridge. This pattern has also been inferred for other macrofungi[16,33,97−99]. These patterns highlight the interplay between continental drift and fungal evolution. In addition, Trichaptum s.l. species distributed in the mountain zones of Central and East Asia exhibit relatively recent divergence times, later than 24 Mya (Figs 2, 15, and 16), and the uplift of the Xizang Plateau may have played an important role in their speciation[100].
Biogeographical history
-
Climatic adaptations further supported fungal evolutionary scenarios; some fungal groups may have originated in the tropical zone, such as ectomycorrhizal Russulaceae, and some possibly in the temperate zone, such as Hymenochaetales[21,35,92,99]. While Trichaptum s.l. species had a wide range of climate zones, from tropical to temperate, and mountain zones. Temperate-adapted clades from Hirschioporus and Pallidohirschioporus may have expanded during global cooling events of the Eocene–Oligocene transition (about 34 Mya), when temperate forests became widespread in the Northern Hemisphere[101].
With the Trichaptum s.l. species expanding from tropical zones to other climatic zones, their basidiomata gradually became more hardened. The coriaceous consistency evolved toward harder forms, including hard corky and woody hard, and the tomentose surface evolved into more specialized forms including smooth, warted, and hispid. The ancestral morphologies suggest a transition from softer to harder basidiomata, indicating an evolutionary trend towards increased structural rigidity. The convergence of similar traits across different clades inhabiting similar climates, such as woody basidiomata in both Hirschioporus (temperate) and Perennihirschioporus (tropical and mountain zones), suggest instances of parallel adaptation. These patterns likely reflect selection pressures shaped by macroecological factors[102], reinforcing the notion that basidiomata structure is not merely a taxonomic character but an adaptive trait molded by climate and substrate availability over evolutionary timescales. Additionally, similar to other groups of macrofungi, the morphological diversity of basidiomata may also contribute to the species diversity of Trichaptum s.l.[103]. Moreover, some Trichaptum s.l. species exhibit both extremely distant phylogenetic relationships and wide geographic separation, such as Perennihirschioporus caymanensis and P. daedaleus, yet share highly similar morphological characteristics that may be attributed to convergent evolution or atavistic evolution of different species under similar environmental conditions.
Adaptive radiation and ecological divergence
-
This study reveals three profoundly different climatic adaptation strategies among the three fungal families. The Hirschioporaceae exemplifies a classic adaptive radiation across various climate zones. In contrast, the warm-temperate Trichaptaceae and the boreal-alpine Pseudotrichaptaceae each exhibit unique and fundamentally distinct ecological strategies.
Family Hirschioporaceae: a case study in adaptive radiation
-
The family Hirschioporaceae exhibits the most remarkable ecological plasticity, showing a clear evolutionary transition from tropical origins to boreal colonization through its four constituent genera, with a clear pattern of adaptive radiation from tropical to temperate and boreal zones.
Firstly, Nigrohirschioporus represents the ancestral ecological state, exhibiting strong niche conservatism. (Fig. 16). It is strictly confined to pantropical rainforests, governed by a 'thermal floor' (Bio11 > 18°C) and high year-round humidity. It lacks the physiological plasticity to tolerate cold or drought, effectively trapping it in stable, aseasonal tropical environments. This 'thermal floor' acts as a powerful biogeographical filter, strictly preventing its dispersal into subtropical or montane regions, even those that are sufficiently wet. Therefore, the distribution of Nigrohirschioporus is a classic example of a species whose range is defined not by the search for a resource (like water), but by the avoidance of a physiological stressor (cold). The strict adherence to a thermally stable, 'aseasonal' tropical niche. also confirms that Nigrohirschioporus has a long evolutionary history deeply rooted within these rainforest environments. Notably, Nigrohirschioporus is primarily distributed in the tropical regions of Southeast Asia and South America. It is considered an ecologically conservative lineage within the family, preserving ancestral ecological niches and distribution patterns. The discovery of Nigrohirschioporus wuyiensis in this study, a rare subtropical species within the genus, provides robust evidence for an evolutionary trend involving the gradual expansion of this genus towards subtropical zones.
This study provides strong quantitative support for the classic tropical Asian-American disjunct distribution of the genus Perennihirschioporus. This distribution could be the result of vicariance, where a once-continuous ancestral population was fragmented by continental drift (e.g., the breakup of Gondwana), or it could be the product of long-distance dispersal events. Compared to Nigrohirschioporus, Perennihirschioporus marks an initial evolutionary step towards adapting to seasonal climates, namely through its adaptation to seasonal drought. With a contribution of nearly 60% to the model, precipitation of the wettest quarter (Bio16) is the absolute master regulator of the genus's distribution. This indicates an extremely high degree of niche specialization. Perennihirschioporus is not merely a 'tropical' genus; it is a genus adapted specifically to tropical climates with a pronounced and intense wet season. This extreme dependence likely reflects a life cycle strategy that is tightly synchronized with seasonal monsoons. The massive influx of water during the wettest part of the year may be a critical trigger for breaking dormancy, facilitating rapid mycelial growth, and inducing the formation of its perennial basidiomata. In contrast, the ability to survive the pronounced dry season (as suggested by its presence in 'semi-arid' tropical climates) is a necessary secondary adaptation. This 'feast and famine' ecological strategy, tied to seasonal water availability, defines its highly restricted habitat. While precipitation is the primary driver, the importance of variables like isothermality (Bio3) and temperature seasonality (Bio4) highlights that this intense seasonal rainfall must occur within a stable thermal environment. The genus thrives where the temperature is consistently warm year-round, without the sharp fluctuations characteristic of temperate or high-altitude regions. This intolerance to temperature variability acts as another layer of filtering, preventing its colonization of otherwise suitably wet subtropical or montane areas. Its physiology appears to be finely tuned to the predictable warmth of the lowland tropics.
Unlike the strictly tropical genera, Pallidohirschioporus demonstrates remarkable ecological plasticity, representing a transitional evolutionary lineage that has successfully bridged tropical and temperate biomes. Our Maxent analysis suggests its distribution is governed not by annual averages, but by the interplay of seasonal extremes, and specifically, the availability of moisture during the dry season and sufficient warmth during the growing season. This points to a 'grow-and-survive' life history strategy: the genus capitalizes on warm, wet summers for rapid vegetative growth and fruiting, while possessing the physiological resilience to endure seasonal drought. However, this plasticity has limits. The sharp decline in habitat suitability where the mean temperature of the coldest quarter (Bio11) drops below freezing indicates a critical physiological barrier: intolerance to severe frost. This 'cold wall' effectively prevents Pallidohirschioporus from expanding into the high-latitude boreal forests dominated by Hirschioporus. Thus, ecologically, Pallidohirschioporus can be defined as a heat-adapted lineage that has expanded poleward as far as the frost line permits, occupying a broad niche distinct from both tropical specialists and boreal endemics.
In stark contrast, Hirschioporus represents a radical evolutionary niche shift from the ancestral tropical state of Trichaptum s.l. to a derived temperate-boreal specialization. This genus has evolved a strict dependency on cool, mesic environments, a trait that tightly correlates with its co-evolution with gymnosperm hosts (pines, spruces, firs) in the Northern Hemisphere. The most significant finding from our model is that the southern distribution limit of Hirschioporus is defined by a 'thermal ceiling' rather than a lack of resources. The high permutation importance of the maximum temperature of the warmest month (Bio5) reveals a fundamental physiological intolerance to summer heat. While saprotrophic fungi generally require moisture, Hirschioporus specifically requires moisture in conjunction with moderate temperatures to sustain its enzymatic activity. This thermal sensitivity effectively excludes the genus from lowland subtropical and tropical ecosystems, confining it to the 'climate islands' of high-altitude mountains or high-latitude boreal zones. Consequently, Hirschioporus exemplifies a trade-off in evolutionary adaptation: by specializing in cold-climate gymnosperm decomposition, it has lost the ancestral ability to tolerate the thermal stress of lower latitudes.
Family Trichaptaceae: the warm-temperate strategist
-
The family Trichaptaceae operates on a climatic principle that is fundamentally different from the adaptive radiation seen in the Hirschioporaceae. Its distribution is dictated not by seasonal averages, but instead by a hard physiological limit to extreme values. A key finding of our study is that the distribution of Trichaptum is governed by a dual thermal control mechanism. The minimum temperature of the coldest month (Bio6) acts as a hard physiological barrier, or a 'cold wall', preventing its northward and southward expansion into temperate zones. The sharp decline in habitat suitability below freezing temperatures (Supplementary Fig. S5b) indicates a fundamental intolerance to frost, a classic trait of tropical-subtropical lineages. This mechanism effectively defines the latitudinal limits of its range. Simultaneously, the high importance of the mean temperature of the warmest quarter (Bio10) reveals that the genus requires a 'heat engine' for its growth and reproduction. It doesn't just tolerate warmth; it thrives in it, with optimal conditions occurring in climates with hot summers (Supplementary Fig. S5d). This requirement for high summer temperatures explains its concentration in lowland tropical and subtropical areas, distinguishing it from species that might tolerate cool summers. This dual dependence of avoidance of winter cold and affinity for summer heat paints a picture of a quintessential warm-climate specialist. While annual precipitation (Bio12) is undeniably crucial, as evidenced by its high contribution and the response curve showing a preference for very humid conditions (Supplementary Fig. S5c), its role appears to be that of a foundational requirement rather than a primary range-limiting factor at the global scale. The permutation importance analysis places thermal variables (especially Bio10) ahead of it. This implies that within the thermally suitable zones (i.e., areas that are not too cold in winter and are hot enough in summer) Trichaptum will then be filtered by moisture availability, preferentially occupying the wettest locations like rainforests. This hierarchical filtering, first by temperature, then by moisture, is a common pattern in niche structuring. This survival strategy, based on absolute extreme values rather than seasonal averages, is fundamentally different from the adaptive models of the four preceding genera.
Family Pseudotrichaptaceae: the extreme cold specialist
-
The monotypic family Pseudotrichaptaceae (genus Pseudotrichaptum) exhibits the most extreme specialization. In contrast to all other lineages, it is restricted to a highly specialized boreal-alpine specialist whose ecological niche is constrained by a strict 'two-way thermal filter'. A central finding of this study is that temperature acts as a 'two-way filter', setting both the southern and northern limits of the genus's range. The permutation importance analysis unequivocally identifies the maximum temperature of the warmest month (Bio5) as the most critical limiting factor. The sharp, cliff-like decline in habitat suitability above 20 °C (Supplementary Fig. S6c) acts as a 'heat barrier', effectively preventing Pseudotrichaptum from colonizing temperate regions with warmer summers. This intolerance to summer heat is a powerful physiological constraint that defines its southern boundary more strongly than any other factor. Conversely, while not the top-ranked variable, the response to annual mean temperature (Bio1), with a peak below freezing (Supplementary Fig. S6b), defines its core thermal environment. The genus is not just tolerant of cold; it is adapted to and likely requires a long, cold winter period for its life cycle, a common feature in boreal organisms. This adaptation to cold, combined with its intolerance to heat, creates a narrow thermal 'corridor' in which it can thrive. The significance of elevation as a predictor reinforces this, demonstrating that Pseudotrichaptum tracks these cold climates vertically into montane regions at lower latitudes. The seemingly paradoxical importance of precipitation of driest month (Bio14) is best understood as a proxy for these thermal conditions. In a circumboreal or alpine climate, the driest month is invariably the coldest winter month, where precipitation is locked away as snow and ice. The model's preference for low Bio14 values is therefore not an affinity for drought, but rather a reflection of the genus's profound adaptation to the arid conditions of a continental frozen winter.
The ecological strategy of Pseudotrichaptum is that of a quintessential boreal-alpine specialist. Its entire life history appears to be finely tuned to the rhythms of these cold, often harsh environments. It grows on a narrow range of hosts (conifers), is adapted to long periods of freezing temperatures, and is competitively excluded or physiologically constrained by the conditions of warmer climates. This high degree of specialization explains why, despite having spores that can travel long distances, its distribution remains strictly confined.
-
Overall, this study re-evaluated the species diversity and evolutionary origins of Trichaptum s.l. by collecting specimens from the Americas and Asia, using phylogenetical, morphological, molecular clock dating analyses, biogeographical data, morphological characteristics of basidiomata, as well as environmental factors, laying the foundation for future resource development and utilization.
-
The authors confirm contribution to the paper as follows: study conception and design: Zhou M, Yuan Y; data collection: Zhou M, Vlasák J; analysis and interpretation of results: Zhou M, Cui Y, Wu Y, Li C; draft manuscript preparation: Zhou M, Cui Y, Zeng G, Zhuang L. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated in the study are available in Table 2.
-
The research is supported by the National Natural Science Foundation of China (Project Nos 32400006, U23A20142, 32300013, 32370013), grants from Hainan Province Science and Technology Special Fund (ZDYF2023RDYL01), the Hainan Institute of National Park (HINP, KY-24ZK02), the Postdoctoral Fellowship Program (Grade C) of China Postdoctoral Science Foundation (GZC20230254) and the Young Elite Scientists Sponsorship Program by CAST (No. 2023QNRC001). We express our sincere gratitude to Heng Zhao (Institute of Applied Ecology, Chinese Academy of Sciences), Yucheng Dai (Beijing Forestry University), and Josef Vlasák Jr. (207 Silverbrook Dr., Schwenksville) for their generous provision of specimens and their strong support throughout this study.
-
All authors declare that they have no competing interests.
-
accompanies this paper online at: https://doi.org/10.48130/mycosphere-0026-0005.
-
#Authors contributed equally: Meng Zhou, Yujin Cui
- Supplementary Table S1 Species used in molecular clock analysis.
- Supplementary Fig. S1 Maxent model of Hirschioporus.
- Supplementary Fig. S2 Maxent model of Pallidohirschioporus.
- Supplementary Fig. S3 Maxent model of Perennihirschioporus.
- Supplementary Fig. S4 Maxent model of Nigrohirschioporus.
- Supplementary Fig. S5 Maxent model of Trichaptum.
- Supplementary Fig. S6 Maxent model of Pseudotrichaptum.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou M, Cui Y, Vlasák J, Wu Y, Li C, et al. 2026. Global phylogeny, divergence times and evolutionary history of Trichaptum s.l. (Hymenochaetales, Basidiomycota): two new families and six new species. Mycosphere 17: e004 doi: 10.48130/mycosphere-0026-0005 

Global phylogeny, divergence times and evolutionary history of Trichaptum s.l. (Hymenochaetales, Basidiomycota): two new families and six new species
- Received: 22 December 2025
- Revised: 29 January 2026
- Accepted: 03 February 2026
- Published online: 26 March 2026
Abstract: The species diversity of Trichaptum s.l., a significant group within the polypore fungi, has expanded substantially in recent years. However, its higher-level taxonomy and evolutionary origins remain underexplored. In this study, the evolutionary history of Trichaptum s.l. is presented using phylogenetic, morphological, and molecular-clock analyses. The results resolve Trichaptum s.l. into four families: Hirschioporaceae, Trichaptaceae, and two newly proposed families, Podocarpioporaceae fam. nov. and Pseudotrichaptaceae fam. nov. Consequently, the Trichaptum complex s.l. now comprises seven genera (Hirschioporus, Nigrohirschioporus, Pallidohirschioporus, Perennihirschioporus, Pseudotrichaptum, Podocarpioporus, and Trichaptum s.s.), and 55 taxa. The six new species from the tropical and subtropical regions are described: Nigrohirschioporus keylargoensis sp. nov., Nigrohirschioporus wuyiensis sp. nov., Perennihirschioporus caymanensis sp. nov., Podocarpioporus hydnoides sp. nov., Trichaptum cystidiolatum sp. nov., and Trichaptum longisporum sp. nov. Molecular clock dating analysis suggests that familial divergence within Trichaptum s.l. occurred during the Cretaceous period (71–119 Mya). Ancestral state reconstruction analysis indicates a complex evolutionary history, potentially originating in Central and East Asia. However, the origins of the four families show notable differences, with two major origin groups in Asia and the Americas. Ecological niche modeling (Maxent) reveals that Trichaptum s.l. likely originated in tropical regions and subsequently expanded into subtropical, temperate, and montane zones, a transition accompanied by the evolution of harder basidiomata. Climatic factors, such as the maximum temperature of the warmest month (Bio5), were identified as primary drivers of the distribution of Hirschioporus, Pallidohirschioporus, and Pseudotrichaptum. The precipitation of the wettest quarter (Bio16) was the dominant factor driving the distribution of Perennihirschioporus. The mean temperature of the coldest quarter (Bio11) was the primary limiting factor for Nigrohirschioporus. The minimum temperature of the coldest month (Bio6), and the annual precipitation (Bio12) showed the highest percentage contribution to the distribution of Trichaptum s.s. This study provides a robust framework for understanding the systematics, evolution, and biogeography of Trichaptum s.l.
-
Key words:
- Clock-dating analysis /
- Evolution /
- Species diversity /
- Wood-inhabiting fungi