









-
The fragrance of flowers involves a complex mixture of thousands of low-molecular-weight volatile organic compounds released by plant floral tissues. It serves as one of the crucial means of communication between plants and insects[1−3]. Fragrance significantly influences the market value of commercial ornamental plants[4,5]. Fragrant metabolites from flowers can also be used practically and are widely used in the food, cosmetic, and pharmaceutical industries[6]. Floral fragrance is complex and dynamic, involving not only a myriad of components but also substantial variations in individual elements. This complexity depends on factors such as species, organ types, environmental conditions, and time[7,8]. Currently, approximately 1,900 fragrance chemicals have been detected in 90 different plant species[9]. The volatile compounds of flowers are mainly divided into three categories and their derivatives: terpenoids, benzenoids/phenylpropanoids, and fatty acids[10,11]. Studies have shown that peonies, lilies, and Freesia × hybrida are rich in terpenoid compounds such as linalool, ocimene, and α-terpinene[12−14], whereas high levels of benzene ring compounds and phenylpropanes, such as phenyl ethyl alcohol, ethyl benzoate, and butyl acetate are found in Rosa damascena and Nymphaea hybrida[15,16]. Fatty acid compounds such as (E)-cinnamaldehyde, γ-decalactone, and (2E)-hexenal are the main volatile compounds in the floral fragrance of Prunus mume and Dianthus caryophyllus[17,18].
Extraction methods for aroma compounds from plant tissues include several common techniques, such as distillation extraction, headspace sampling, solid-phase microextraction (SPME), supercritical fluid extraction, and solvent extraction[19,20]. The method of SPME of the static headspace (HS) is a straightforward and sensitive technique for collecting volatile compounds from plant tissues. The collected samples are subsequently analyzed using a gas chromatography–mass spectrometry (GC–MS) system. This method has proved to be highly effective for the qualitative and quantitative analysis of the aromatic constituents in flowers. It has been widely employed in the analysis of fragrance components in various ornamentals, such as roses, osmanthus, orchids, Jasminum spp., and Hedychium spp.[21−26].
Dendrobium is the second-largest genus of the Orchidaceae family and is of great economic importance in traditional medicine and the horticultural trade. Phalaenopsis-type Dendrobium hybrids (den-phals) are successful man-made hybrids based on D. phalaenopsis Fitzg. and are primarily grown as a major cut flower plant for export from several Southeast Asian countries, where they contribute significantly to the country's foreign exchange earnings. Although several commercial den-phals are scented, fragrance has not been marketed for den-phals, because the majority of commercial cultivars are odorless. A minority of fragrant hybrids either have only a slight aroma or an unpleasant scent. Fragrance will be a desirable target in future breeding. Many other species in the Dendrobium genus have strong and pleasant scents, and the main aroma components (e.g., in D. officinale, D. nobile, and D. bensoniae) are hexanal, heptanal, α-terpinene, and D-limonene[27]. In D. parishii, the main aroma component in the sepals is hexanal, whereas in the petals the main compounds are trans-2-octenal and 2-pentyl furan, while lip, column, and spur have a common main aroma component, which is 2-pentadecanone[28]. In D. Zhenzhu, D-limonene is widely present in the sepals, petals, lips, and column and spur, serving as its main aroma compound. The concentration of aroma compounds of D. primulinum flowers show a decreasing trend during the day, with the highest concentration at 06:00 and the lowest at 18:00[27−32]. Although our previous study detected several terpenes in flowers of seven den-phals, namely 3-carene, linalool, α-cubebene, and α-phellandrene, which acted as their major aromatic compounds[33], the mechanism of scent production by den-phals has not been well investigated to date. The contribution of an individual aromatic compound to the overall aroma depends on its concentration and odor threshold. Previous studies have evaluated the contribution of individual aromatic compounds to the overall aroma by calculating odor activity values (OAV). An OAV greater than 1 indicates that the aroma component has a certain impact on the overall aroma, with higher OAVs contributing more markedly to floral fragranc. Further detailed investigations of the aromatic products and their release patterns in den-phals is necessary.
Therefore, in this current study, 22 hybrids with various degrees and types of floral scent were identified out of 79 den-phals screened for the evaluation of aroma intensity. Subsequently, nine den-phals were further selected for sensory evaluation and characterization, the latter involving HS–SPME, coupled with GC–MS technology. The study conducted analyses on the headspace volatiles during the flowering stages, from different floral organs, and at various time points in the day across the nine den-phals. Subsequently, the research utilized variable importance in projection (VIP) values, along with OAV, to identify the volatile compounds that significantly contributed to the aroma of den-phals with various fragrances. The present work provides the basic information for illustrating the mechanism of scent production in den-phals, which could be applied for fragrance breeding and the use of aromatic dendrobium accessions.
-
Twenty-two den-phals accessions (Table 1) were obtained from the Tropical Flower Resource Nursery of Tropical Crops Genetic Resources Institute, the Chinese Academy of Tropical Agricultural Sciences, Hainan, China. Nine den-phals, namely D. Chao Praya Garnet, D. Udomsri Beauty, "DL2020-1", D. Burana Charming, D. Aridang Blue, "2018086-6", D. Emma White, D. Smile, and D. Nopporn Pink (Table 1, Fig. 1a), were used for sensory analysis and aromatic characterization. Samples of different floral organs (petals, sepals, labellum, and column and spur) from fully blooming flowers (characterized as 2 d after initial blooming) were collected between 11:30 and 12:30 from identical plants propagated through tissue culture. Three samples of each floral organ type were collected from each plant. Samples of flowers were collected at each of the three stages of flowering [early flowering stage (the initial opening of the buds), full flowering stage (5 d after the initial opening), and late flowering stage (14 to 18 d after the initial opening, the floral tissues starting to wilting)] were collected (Fig. 1b). At the full flowering stage, three samples were collected at each of four different times (08:00, 12:00, 16:00, and 20:00). The collected samples were placed in a refrigerated container and promptly transported to the laboratory.
Table 1. Names and abbreviations of den-phals used in the experiments.
Hybrid name
or designationAbbreviations Hybrid name
or designationAbbreviations "37-03" "37" D. Nopporn Pink NP "17001-S36" "17001" D. Oh Swee End OSE "2018086-6" "2018" D. Udomsri Beauty UB "DL2020-1" "DL" D. King Dragon KD "DL2020-6" "DL-1" D. Burana Charming BC "DL2021-6" "DL-2" D. Smile S D. Emma White EW D. Mount Kelly Beauty MKB D. Burana Crystal BCr D. Chao Praya Garnet CPG D. Bertha Chong BCh D. Nanyang Primary School NPS D. Give My God GMG D. Black Cat BCa D. Aridang Blue AB D. Banana Royal BRTaT 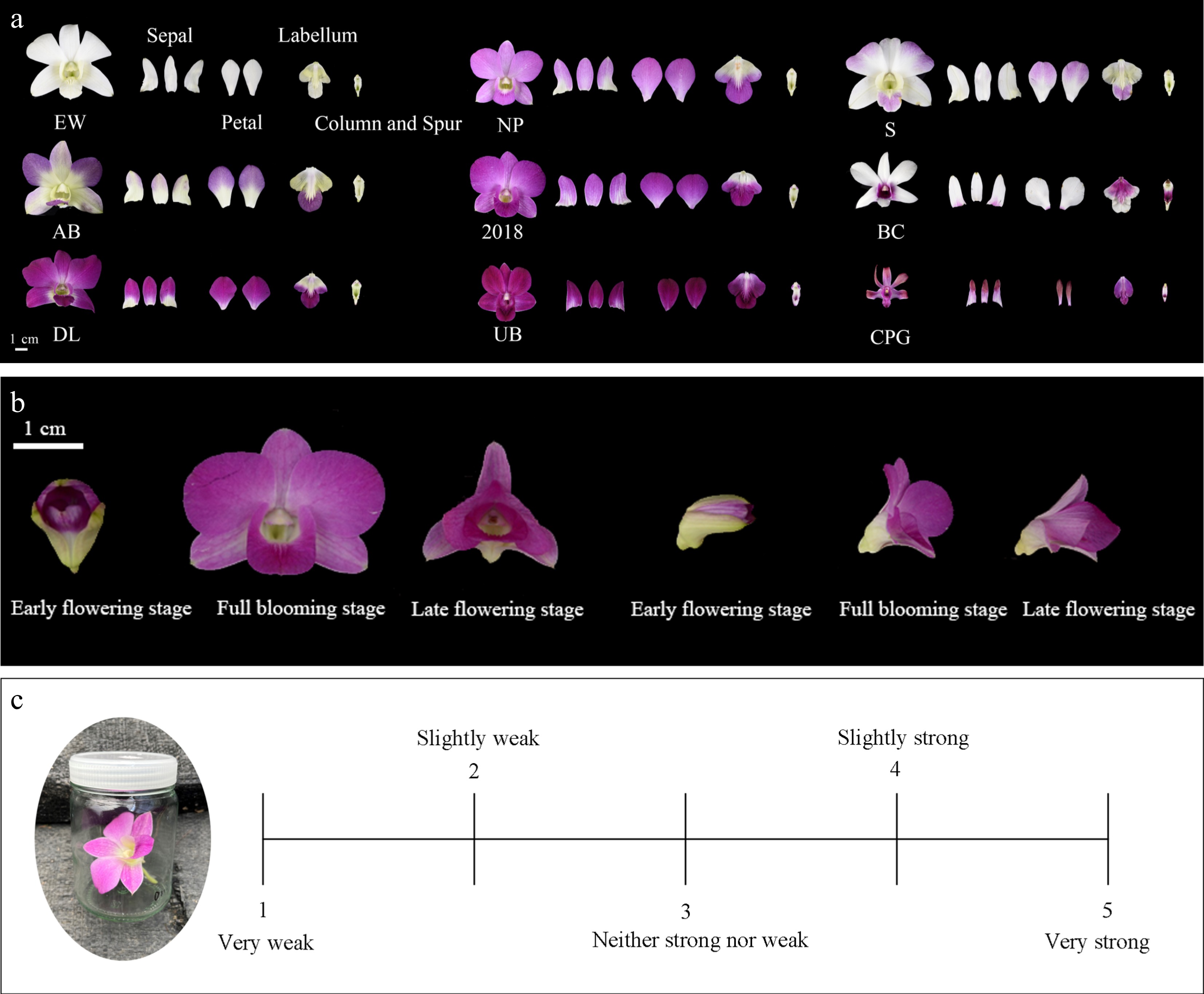
Figure 1.
(a) Floral organs of nine den-phals. (b) Different flowering stages of den-phals. (c) The 5-point interval scale for evaluation of den-phals floral scent intensity. The floral scent intensity gradually increasing from point 1 to 5.
Sensory evaluation
-
The experiments were conducted in a clean room (25 ± 2 °C) between 11:30 and 12:30. Twenty-five people were recruited from the Tropical Crop Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences through a simple screening process, and consisted of 10 males and 15 females, with ages ranging from 20 to 40 years. All flower samples were assessed for fragrance using a 5-point interval scale (1 = very weak, 5 = very strong) (Fig. 1c)[34−36]. There was a 2 to 3-min interval between smelling different odors. The score for each sensory attribute represents the average of the scores from each of the 25 individuals.
Floral scent collection and compound identification
-
Collection and detection of volatile substances was carried out using methodologies based on prior research, with slight modifications[37]. Flower tissue was snap-frozen with liquid nitrogen. A 1.5 g sub-sample was swiftly added to a small vial fitted with a cap, and placed in a constant temperature water bath at 55 °C for 10 min. Next, the SPME fiber (50/30 μm, DVB/Carboxen/PDMS, Supelco, USA) was inserted into the headspace of the vial (to adsorb volatile compounds) for 30 min. Before extracting each sample, the fiber was desorbed at 250 °C for 5 min to eliminate residual substances.
Following volatile compound extraction by SPME, the fiber was quickly removed and immediately inserted into the GC injector port in a splitless mode for desorption at 250 °C for 5 min. The GC system (Thermo Fisher Scientific, Waltham, MA, USA) utilized a DB-5MS quartz capillary column (30 m × 0.25 mm × 0.25 μm, Thermo Scientific, Bellefonte, PA, USA) with a flow rate of 1 mL/min and high-purity helium as the carrier gas (> 99.999% purity) in the splitless mode. The temperature program was as follows: the initial column temperature was set at 40 °C for 3 min, followed by an increase to 70 °C at a rate of 8 °C/min with a 2-min hold, then a further increase to 170 °C at a rate of 3 °C/min with a 2 min hold. Finally, a ramp at 10 °C/min to 220 °C was applied with a 3 min hold. The MS system maintained the injector temperature at 230 °C, the ion source temperature at 150 °C, and used electron ionization with an electron energy of 70 eV. The GC–MS transfer line temperature was set at 250 °C, and the mass scan range was m/z 40 to 450 amu. The identification was based on mass spectral similarity. The obtained mass spectra of volatile compounds from GC–MS analysis were compared against standard substance mass spectra in the NIST library (
http://webbook.nist.gov/chemistry ) for retrieval. Further qualitative confirmation was conducted based on Kovats' Retention Indices[38]. The retention indices for volatile compounds in the samples were calculated using standard n-alkane samples (C8–C30) ( Sigma, St. Louis, MO, USA) under the same temperature program.Data analysis and presentation
-
Orthogonal partial least squares-discriminant analysis (OPLS-DA) was conducted using Simca 14.1, while a heatmap was generated using TBtools 2.012. Figures were created using Origin 2021 and Adobe Photoshop 2020.
-
Twenty-two aromatic den-phals were assessed by sense of smell and were used in the sensory evaluation of aroma intensity. The 22 den-phals were categorized into three categories, namely: light fragrance (five hybrids), medium fragrance (nine hybrids), and strong fragrance (eight hybrids), based on different aroma intensities (Fig. 2a). Furthermore, EB, S, and NP from the light fragrance category, AB, BC, and "2018" from the medium fragrance group, and CPG, UB, and DL from the strong fragrance category were selected for more detailed sensory evaluation of the aroma. In the light fragrance category, EW had floral, grassy, and sweet aromas, and S and NP had floral and relatively strong grassy aromas (Fig. 2b). Of the medium fragrance category, AB had floral and relatively strong grassy aromas, BC had grassy, sweet, and relatively strong floral aromas, and "2018" had grassy and spicy aromas (Fig. 2c). Among the strong fragrance category, CPG had floral, spicy, and relatively strong grassy aromas, UB had fruity, sweet, and relatively strong grassy aromas, while DL had fruity, grassy, and relatively strong floral aromas (Fig. 2d). The aroma of den-phals is mainly characterized by grassy and floral notes. In terms of fragrance preferences, hybrids that simultaneously possess higher levels of both grassy and floral notes were more favored by the testers.
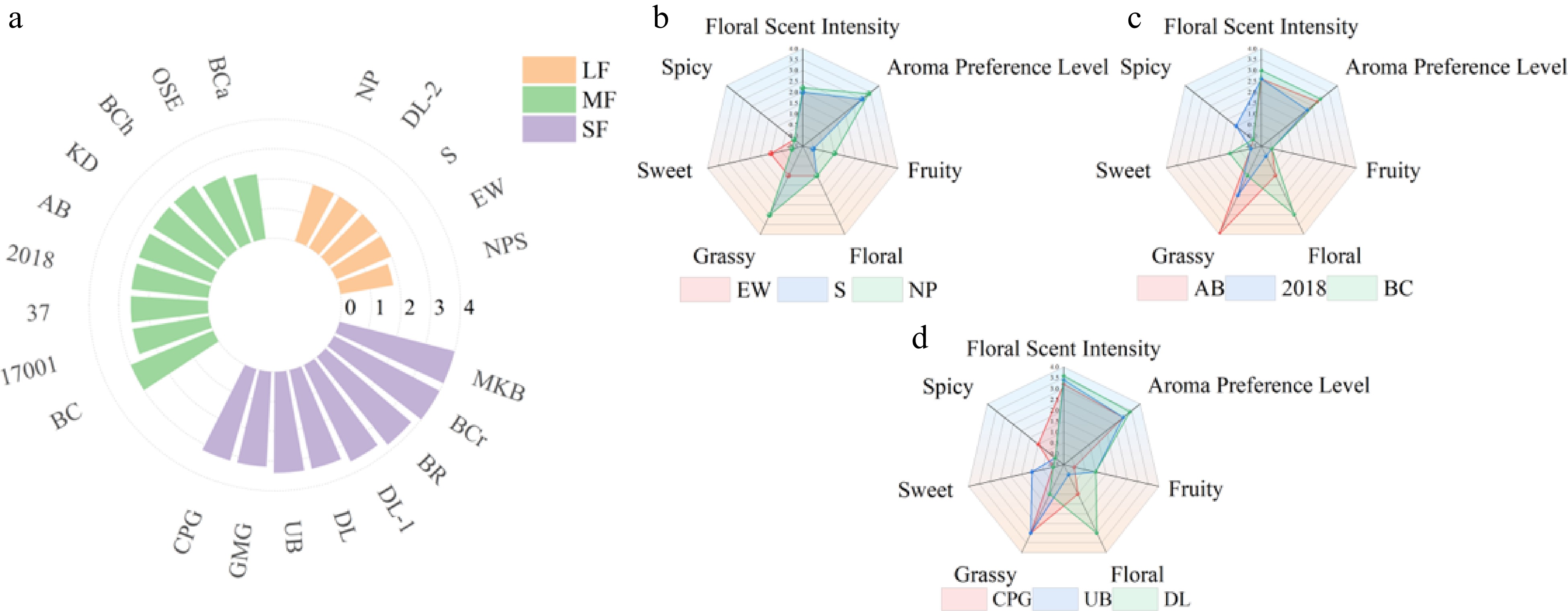
Figure 2.
Sensory evaluation of den-phals floral fragrance. (a) Sensory evaluation of aroma intensity for 22 den-phals. (b) Radar chart of sensory evaluation for the light fragrance category. (c) Radar chart of sensory evaluation for the medium fragrance category. (d) Radar chart of sensory evaluation for the strong fragrance category. (LF: light fragrance; MF: medium fragrance; SF: strong fragrance).
Identification of volatile components in den-phals
-
A total of 137 volatile substances were detected in the samples of den-phals floral organs at different flowering stages and times of the day (Supplemental Table S1). They could be categorized into nine classes, namely 80 terpenoids, 21 benzenoids/phenylpropanoids, 28 aliphatics, and eight miscellaneous cyclic compounds (Fig. 3a). In the samples taken at different flowering stages, times, and from various organs, 90, 81, and 106 volatile compounds were detected, respectively. There were 52 aromatic substances common to all three sampling criteria (Fig. 3b). Terpenoids and some aliphatics were the major components of the aromatic emissions of den-phals. Hexanal (2.43%–26.22%), (E)-2-hexenal (14.14%–58.13%), (E)-2-octenal (0%–12.53%), linalool (0%–20.26%), (E,Z)-2,6-nonadienal (0%–8.01%), (E)-2-nonenal (0%–9.95%), ylangene (0%–7.63%), copaene (0%–18.15%), caryophyllene (0%–21.07%), α-muurolene (0%–17.76%), and γ-muurolene (0%–4.02%) were consistently identified in the majority of den-phals.
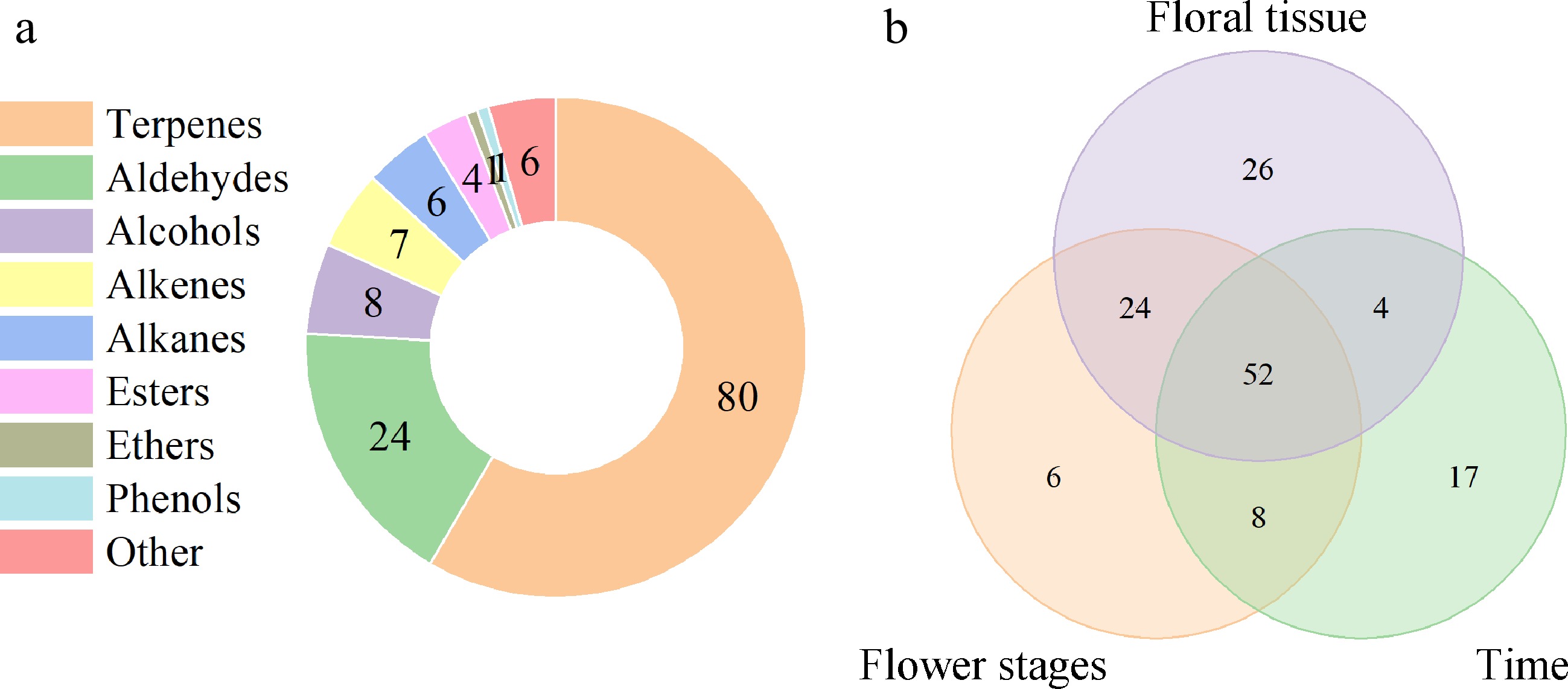
Figure 3.
Volatile components in den-phals. (a) Number of volatile compounds in different categories. (b) Venn diagram of types of volatiles at different flowering stages, times of the day, and organs.
Analysis of volatile components at different flowering stages
-
During the process of flowering, there are fewer types and lower concentrations of volatile compounds in the early flowering stage, with 57 different volatile substances. As the flowers continued to open, both the types and concentrations of volatile compounds rapidly increased, reaching 80 compounds during the full flowering stage. Subsequently, with the senescence of the flowers, both the types and quantities of aromatic compounds decreased, with 70 different volatile compounds present during the late flowering stage (Fig. 4a, Fig. 5a).
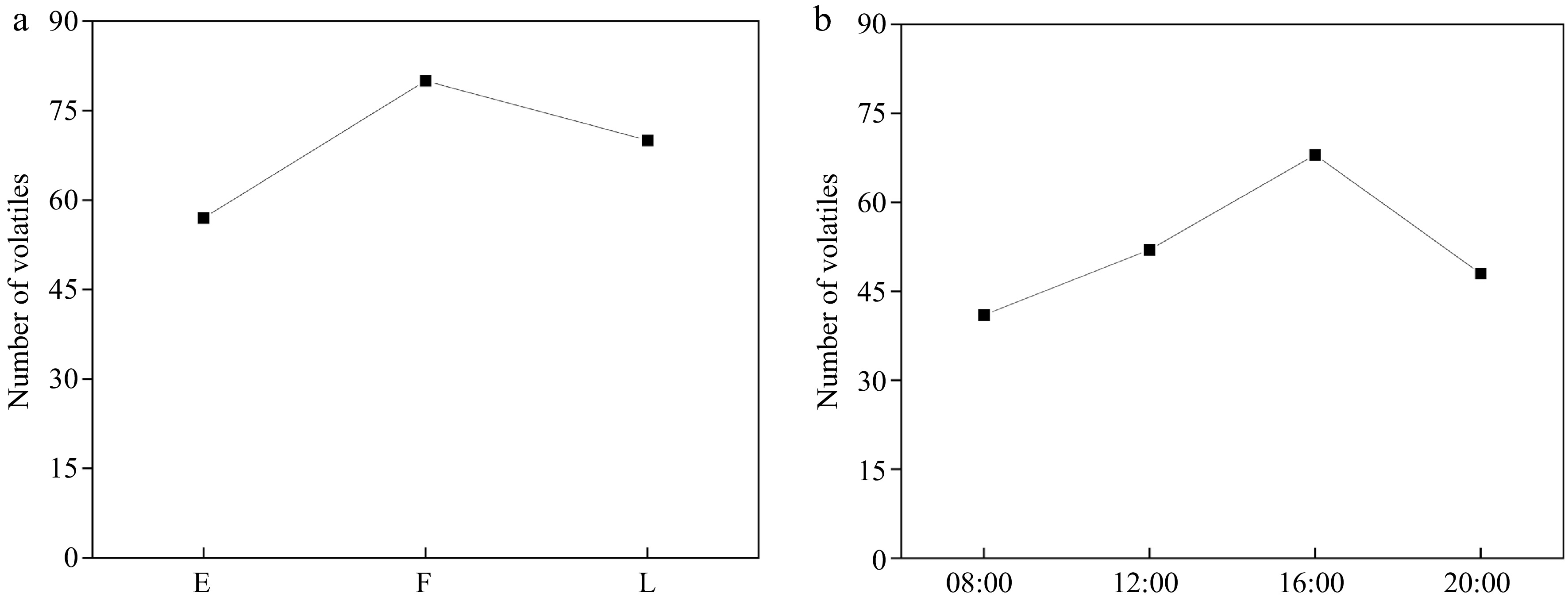
Figure 4.
Volatiles number change diagram. (a) Different flowering stages (E: early flowering stage; F: full flowering stage; L: late flowering stage). (b) Different times of the day.
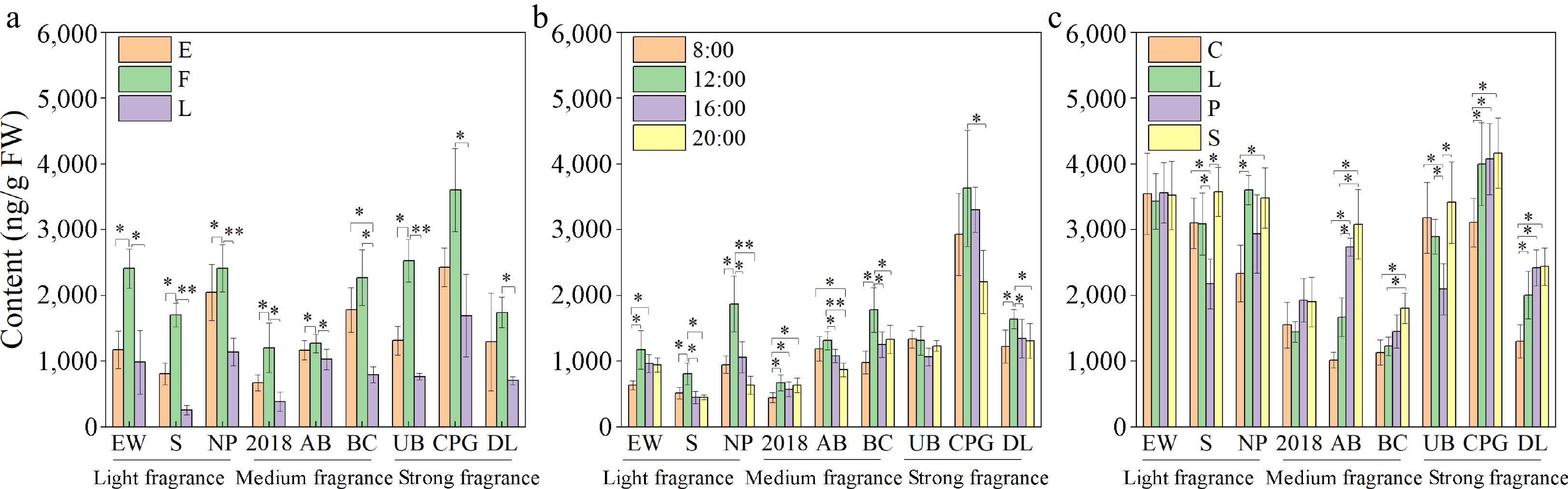
Figure 5.
Relative concentration of volatile compounds in den-phals from different fragrance categories. (a) Different flowering stages. (b) Different times of the day. (c) Different floral organs. (E: early flowering stage; F: full flowering stage; L: late flowering stage; (C: column and spur; L: labellum; P: petals; S: sepals; *, p < 0.05; **, p < 0.01).
The full flowering stage exhibited the greatest diversity and concentration of volatile compounds. Therefore, cluster analysis was conducted based on the concentration of the main volatile substances during the full flowering stage. Cultivar CPG was categorized into a subgroup due to its higher concentration of 2,4-nonadienal and caryophyllene. Another subgroup was formed from "2018", with DL, EW, and NP included, due to similar levels of ylangene and copaene concentrations. AB, S, BC and UB were grouped into one subgroup due to similarly low levels of γ-muurolene and α-muurolene (Fig. 6a). For cultivars in the light fragrance category (EW, S, and NP), aldehydes and small concentrations of terpenes dominated at the early and final flowering stages, during the full flowering stage, both the kind and the concentrations of aliphatics increased, and terpenes, such as ylangene, copaene, and humulene, present at higher concentrations, started to appear. In the medium fragrance category ("2018", AB, and BC), there was a higher concentration of aliphatics during both the early and final flowering stages when compared with the cultivars in the light fragrance category. In the early and final flowering stages of the medium fragrance category, several other compounds, including trans-α-bergamotene, trans-2-octen-ol, ocimene, and trans-β-ocimene, were detected. During the full flowering stage, along with the increase in the concentrations of the corresponding aromatic substances, the presence of linalool oxide, 2,4-nonadienal, and longifolene was also observed. In the strong fragrance category (UB, CPG, and DL), the concentration of aromatic compounds exceeded that of the other two categories. In the early and final flowering stages of the strong fragrance category, high concentrations of 1-octanol, δ-elemene, and geranylacetone were observed. Compared to all flowering stages of light aroma and medium aroma, the full flowering stage of strong aroma has a higher content of caryophyllene, γ-cadinene, and caryophyllene oxide were detected (Supplemental Table S1).
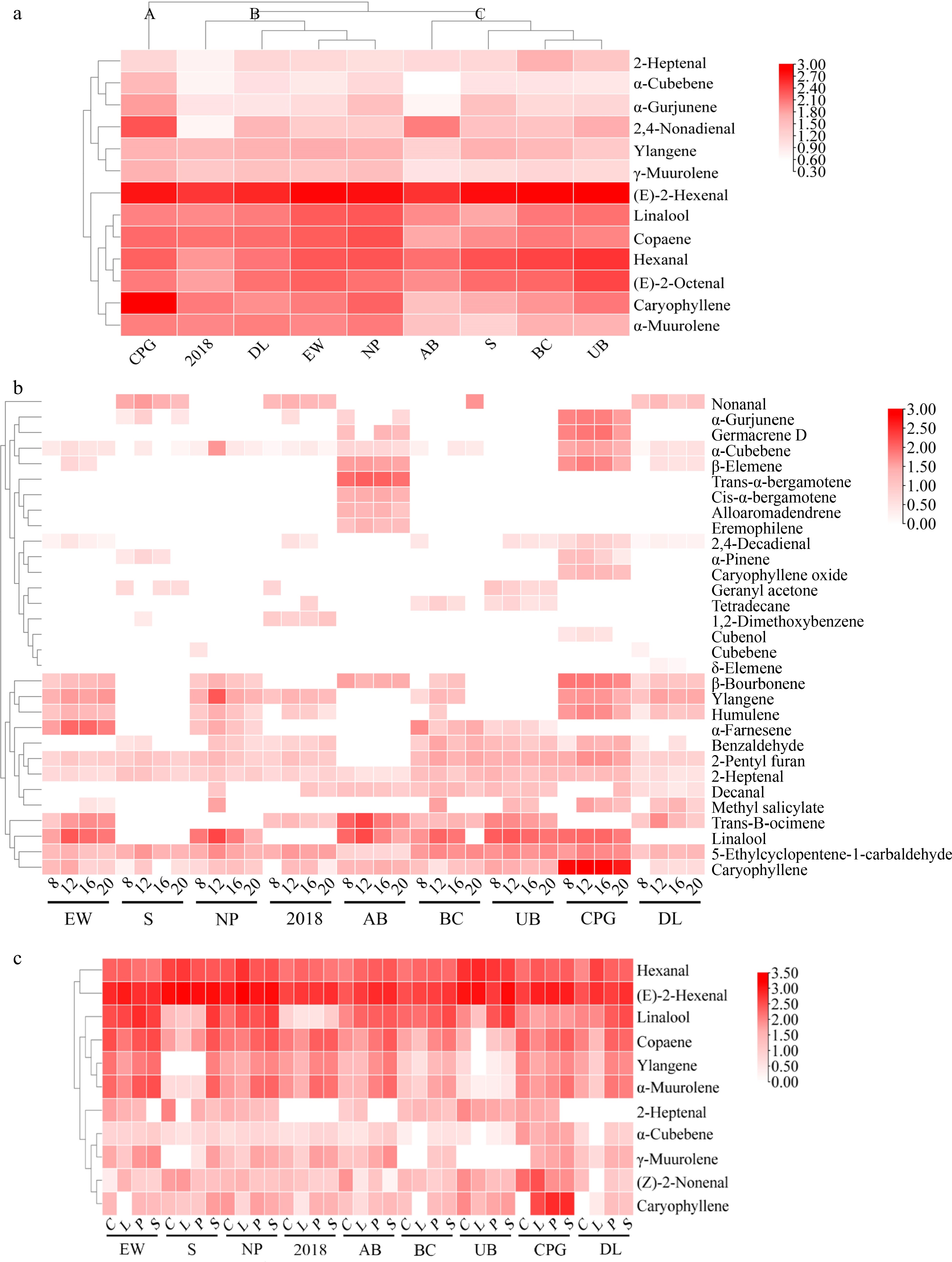
Figure 6.
Heatmap analysis and hierarchical clustering analysis for den-phals. (a) Full flowering stage. (b) Heatmap analysis for VIP > 1 of different times of the day (8: 8 o' clock; 12: 12 o' clock; 16: 16 o'clock; 20: 20 o'clock). (c) Different flower organs (C: column and spur; L: labellum; P: petals; S: sepals).
To further investigate the olfactory differences among various den-phals during the individual flowering stages, an OPLS-DA model was established (Fig. 7a). The nine den-phals were categorized into three groups based on their flowering stages. The predictive ability of the model (Q2) was assessed through cross-validation, yielding a Q2 value of 0.648. Additionally, after 200 permutation tests (Supplemental Fig. S1a), the intersection point of the Q2 regression line with the vertical axis was below 0, indicating the absence of overfitting and confirming the validity of the model. Therefore, this model was deemed reliable for the analysis of volatile compounds in den-phals at different flowering stages. To further analyze the contributions of compounds from different floral organs to the aroma during different flowering stages of the various hybrids, 20 differential aromatic substances were selected based on the VIP > 1 criterion and a dual-zone plot was created (Fig. 7d). The selected compounds consisted of 13 aliphatics, four terpenes, two benzenoids/phenylpropanoids, and one miscellaneous cyclic compound. As shown in the graph, different flowering stages have different substances, during the early flowering stage, the differential volatile compounds comprised hexanal, (E)-2-hexenal, and (E)-2-octenal. The full flowering stage featured differential volatile compounds such as undecanal, methyl salicylate, and germacrene D, while the late flowering stage included stage-specific compounds like benzaldehyde, phenylacetaldehyde, and 1-octen-3-ol.
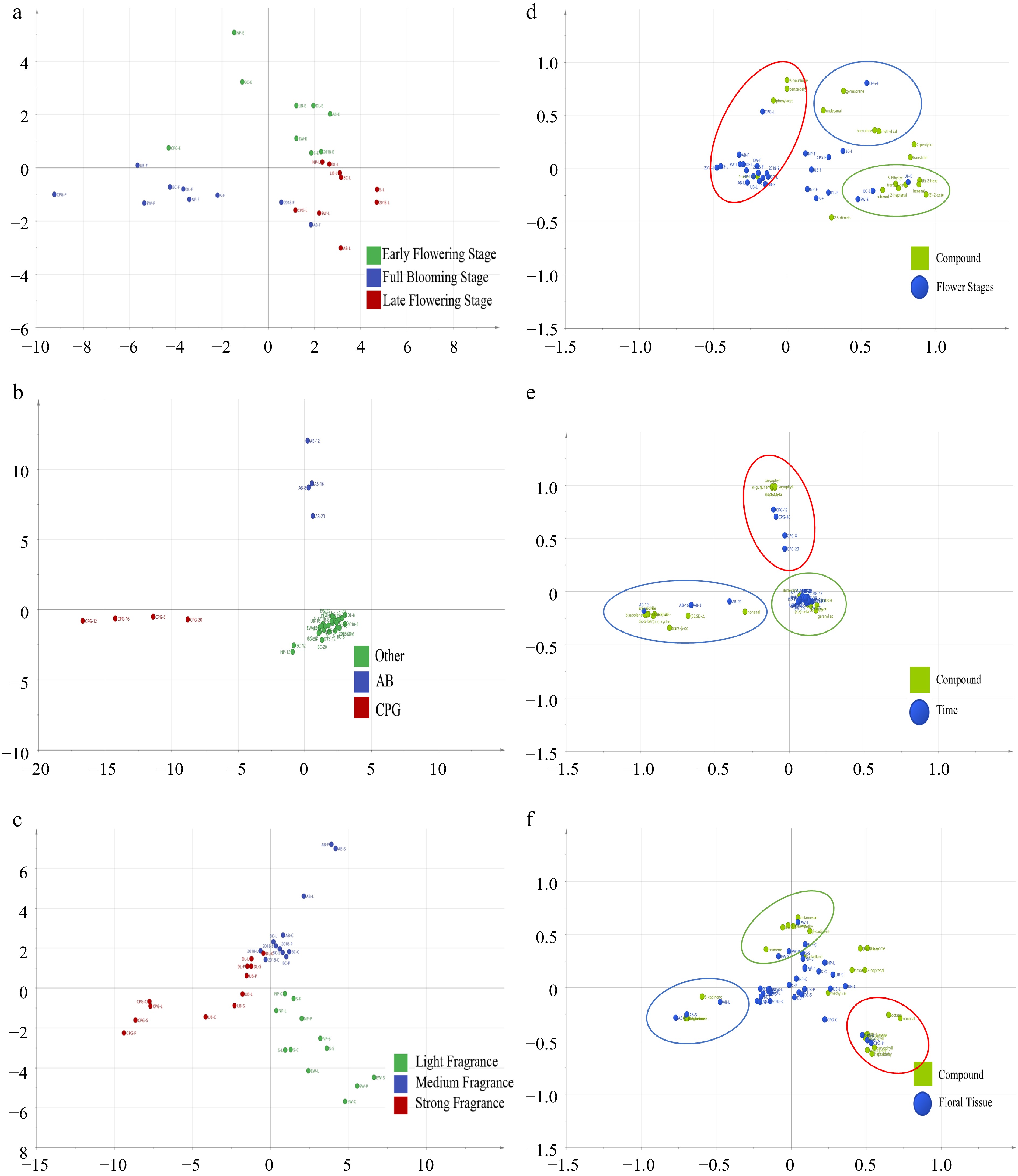
Figure 7.
OPLS-DA models of volatile compounds. (a) Different flowering stages. (b) Different times of the day. (c) Different flower organs. VIP > 1 substance double-region plot. (d) Different flowering stages. (e) Different times of the day. (f) Different flower organs. (The colors of the circles in d, e, and f correspond to the colors in the legends of a, b, and c, respectively).
Based on the reported thresholds and sensory descriptions of aroma components in the literature, a total of 26 volatile substances in all flowering stages of the nine den-phals can be queried for thresholds, which will be used in the calculation of OAV and the line chart is made up of 15 substances with OAV > 1 (Fig. 8a). There were nine compounds with relatively high OAVs across all den-phals. OAVs greater than 10 were observed for linalool (rose aroma), (E)-2-octenal (fatty aroma), hexanal (grassy aroma), and octanal (fruity aroma). OAVs greater than 1 were observed for (E,Z)-2,6-nonadienal (violet and cucumber aromas), nonanal (citrus aroma), 2-pentyl furan (vegetable aroma), heptanal (fatty aroma), and decanal (sweet aroma). The light fragrance category of scented hybrids was dominated by common aroma-contributing compounds, both the early and late flowering stages included (E)-2-hexenal (grassy aroma), and trans-2,4-decadienal (meaty aroma) during the full flowering stage. In the medium fragrance category, trans-2,6-nonadienal (floral aroma) and, during the full flowering period, trans-β-ocimene (orange blossom aroma) were present. In the strong fragrance category, methyl salicylate (herbal aroma) is present during the early flowering period. Trans-2,4-decadienal had an OAV greater than 10 in the full flowering stage of cultivar CPG, while, throughout all the flowering stages of this den-phals, caryophyllene (spicy aroma) had an OAV greater than 1, with a higher value during its full flowering stage.
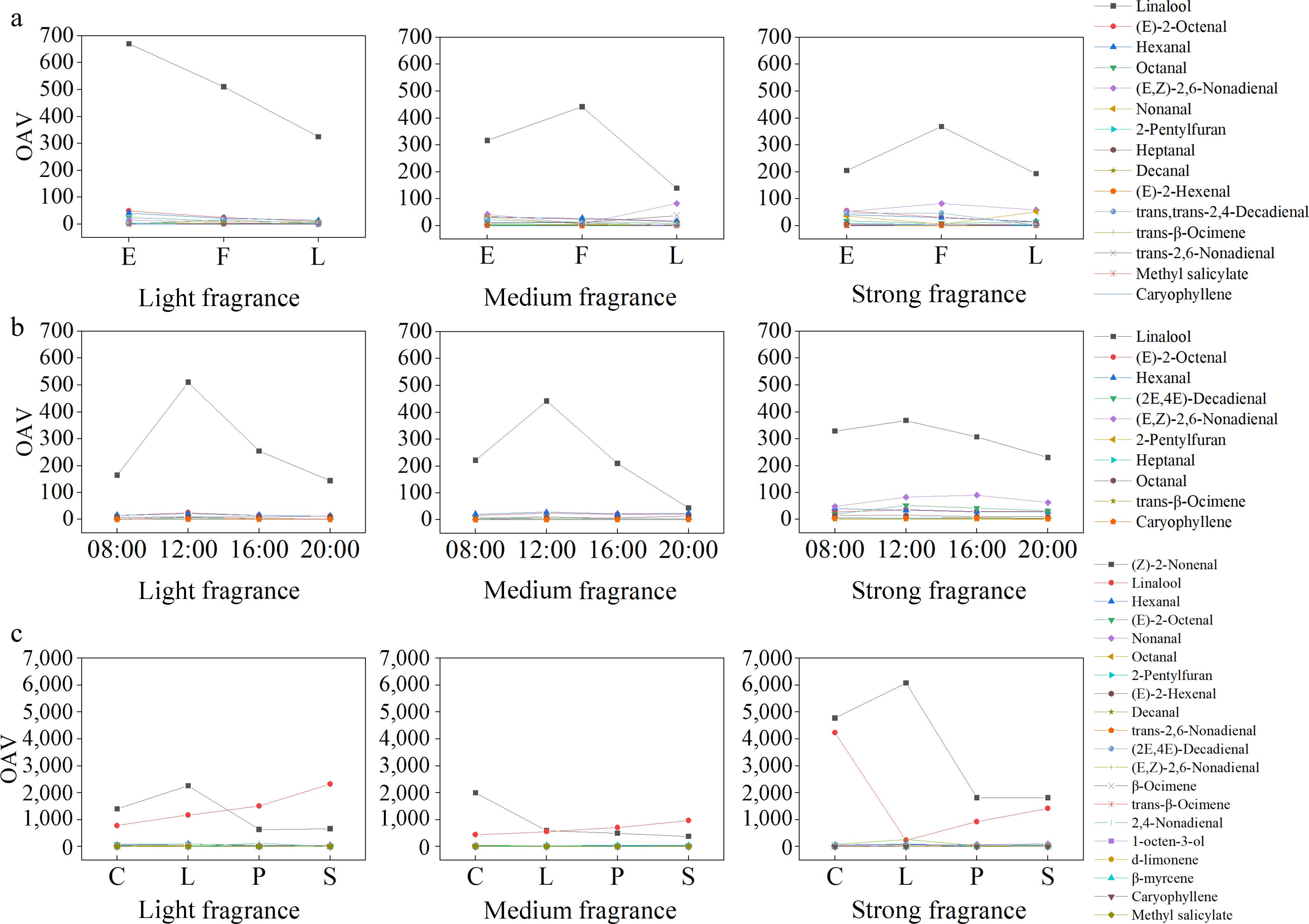
Figure 8.
Line graphs showing the relationship between the OAV of aromatic compounds of nine den-phals from light, medium, and strong fragrance categories and (a) flowering stage, (b) times of the day, and (c) floral organs. (a) E: early flowering stage; F: full flowering stage; L: late flowering stage. (c) C: column; L: labellum; P: petal; S: sepal.
Analysis of volatile components at different times of day
-
During different time points in the day, there were 32 volatile compounds, consisting of 13 terpenoids, 14 aliphatics, one benzenoids/phenylpropanoids, and four miscellaneous cyclic compounds common to the nine den-phals. The composition and concentrations of aromatic compounds exhibited a trend of initially increasing and then decreasing throughout the day, with the highest concentration of volatile compounds occurring at 12:00, and the greatest diversity of volatile compounds occurring at 16:00 (Figs 4b & 5b). At 08:00, flowers of the light fragrance category consisted mainly of aliphatics and a small number of terpenes. In the middle fragrance category, there were additional compounds, such as 2-pentyl furan, trans-β-ocimene, nonanal, and 1,2-dimethoxybenzene. Flowers in the strong fragrance category included α-pinene, cubebene, benzaldehyde, and geranyl acetone (Fig. 6b). The diversity and concentration of volatile compounds increased at 12:00, in addition to the compounds that were present at 08:00. All three fragrance categories included 2-heptenal, 5-ethylcyclopentene-1-carbaldehyde, linalool, caryophyllene, α-cubebene, ylangene, β-bourbonene, humulene, α-farnesene, and α-gurjunene. In the medium fragrance category, decanal, β-elemene, cis-α-bergamotene, trans-α-bergamotene, alloaromadendrene, germacrene D, eremophilene, and bisabolene were detected in all samples. Furthermore, methyl salicylate, caryophyllene oxide, and cubenol were also detected in the strong fragrance category. At 16:00 and 20:00, the volatile compounds were similar but the diversity and concentration of volatile compounds decreased. In the light fragrance category at these times, methyl salicylate was detected, whereas, in the medium fragrance category, tetradecane, and 2,4-decadienal were detected, and in the strong fragrance category, δ-elemene was present.
The OPLS-DA model was established with 83 aroma components as dependent variables and different time points as independent variables (Fig. 7b). The results of cross-validation analysis (Q2 = 0.649) substantiate the feasibility of this model (Supplemental Fig. S1b). The results indicate significant differences between cultivars CPG and AB at different time points, compared with other hybrids. To further analyze the contribution of different hybrids and floral organs to the aroma, 35 different aroma compounds were selected based on the VIP > 1 criterion and presented in a double-region plot (Fig. 7e). These compounds consisted of 20 terpenes, six aliphatics, one benzenoids/phenylpropanoids, and eight miscellaneous cyclic compounds. In the medium fragrance category, cultivar AB was characterized by significant differences in cis-α-bergamotene, trans-α-bergamotene, eremophilene, (+)-cyclosativene, bisabolene, alloaromadendrene, and nonanal. On the other hand, the content of caryophyllene, caryophyllene oxide, α-gurjunene, and (E,Z)-2,6-nonadienal in cultivar CPG is higher than other hybrids. Apart from CPG and AB, the other fragrance category hybrids showed relatively small differences between the different times of the day, mainly characterized by dodecane, linalool oxide, (Z,E)-α-farnesene, veratrole, geranyl acetone, β-farnesene, and trans-β-ocimene.
A total of 26 volatile substances in all times of the nine den-phals can be queried for thresholds, which will be used in the calculation of OAV and the line chart is made up of 10 substances with OAV > 1 and the flowering time of different den-phals (Fig. 8b). Notably, the OAV for linalool exceeded 100, while (E)-2-octenal and hexanal had OAVs above 10 in all hybrids. Additionally, trans-2,4-decadienal, (E,Z)-2,6-nonadienal, 2-pentyl furan, heptanal (oily aroma), and octanal had OAVs exceeding 1. The major volatiles contributing to the aroma in the flowers from the light fragrance category were the common compounds mentioned above. In the medium fragrance category, in addition to the common compounds, trans-β-ocimene also appeared at 08:00, 12:00, and 16:00 in hybrids AB and "2018". In the strong fragrance category, octanal (oily scent) exhibited high OAVs at 08:00 (11.01), 12:00 (9.8), 16:00 (13.13), and 20:00 (11.86), while (E,Z)-2,6-nonadienal had OAVs above 100 at all four time points in cultivar CPG. Methyl salicylate exhibited an OAV greater than 1 only at 16:00 (1.24) in cultivar DL.
Analysis of volatile components from different floral tissues
-
To investigate the aroma released from different parts of the flowers of den-phals, the volatile compounds from four different flower organs were analyzed separately. Petals had a greater diversity of volatile compounds, while sepals had a higher concentration of volatile compounds (Fig. 5c). According to the heatmap results (Fig. 6c), the concentrations of hexanal, (E)-2-hexenal, and (Z)-2-nonenal from the column, spur and labellum were higher than those in the petal and sepal. The concentrations of linalool, copaene, ylangene, and α-muurolene in the petal and sepal were higher than those in the column, spur, and labellum. Ylangene had a lower concentration in the column, spur, and labellum of cultivar S, while the concentration of 2-heptanal was higher in concentration only in the column, and spur of cultivars S and UB. All floral organs of hybrids "2018" and DL had only a low concentration of 2-heptanal. All floral organs of cultivar CPG contained higher concentrations of α-cubebene than other hybrids. The caryophyllene concentration in CPG labellum, petal, and sepal was significantly higher than in any other hybrids.
In addition, in the column and spur, the concentrations of α-terpineol, citral, trans-2-octen-1-ol, longifolene, α-cadinene, trans, trans-2,4-heptadienal, and (E,E)-2,4-dodecadienal were relatively high. In the labellum, the concentrations of tetradecane, trans-2,4-decadienal, phenyl ethanol, (E,E)-2,4-hexadienal, α-pinene, benzaldehyde, 1-octen-3-ol, and undecanal were relatively high. In petals with more volatile substances, cultivar EW contained δ-cadinene, β-pinene, and geraniol; cultivar S was rich in cubebene; BC was rich in alloaromadendren, AB contained higher concentrations of β-myrcene, α-phellandrene, terpinene, and D-limonene; cultivar CPG was rich in methyl salicylate, heptanal, octanal, and β-elemene; and cultivar DL had higher levels of (+)-aromadendrene and cadinene. In sepals, with their high volatile matter content, cultivar EW contained higher levels of germacrene D and linalool oxide, S contained higher levels of α-cubebene and β-cadinene, and NP contained guaiazulene. Cultivar AB contained significant concentrations of (+)-sativen and cis-murola-3,5-diene, UB was rich in ocimene and (3E)-4,8-dimethyl-1,3,7-nonatriene, DL contained methyl salicylate, and CPG contained trans-2-decenal, γ-cadinene, and caryophyllene oxide.
To further investigate the aromatic volatile differences among flower organs of different hybrids, OPLS-DA was conducted using 106 aroma components as the dependent variables and different flower organs as the independent variables (Fig. 7c). The predictive index (Q2 = 0.723) in this analysis confirmed the feasibility of the model (Supplemental Fig. S1c). According to the VIP > 1 criterion, 47 different aroma compounds were selected to establish a double-region plot (Fig. 7f), namely 23 terpenoids, 14 aliphatics, three benzenoids/phenylpropanoids, and seven miscellaneous cyclic compounds. Different types of flower organs were clustered according to the intensity of the fragrance of the cultivar concerned. Among the cultivars, EW, AB, and CPG exhibited significant differences in the volatiles from each flower organ. In EW flowers, β-phellandrene was the main organ-specific compound in the column and spur, while α-farnesene, phenethyl alcohol, and β-cadinene were the main differential volatiles in the labellum; and ocimene was the main organ-specific volatile in the petals and sepals. In AB flowers, the labellum mainly contained δ-cadinene as the differential compound, while the petals and sepals contained β-myrcene, D-limonene, and terpinene as the differential compounds. In the flowers of cultivar CPG, methyl salicylate was the main differential substance in the column and spur, while heptanal, caryophyllene, caryophyllene oxide, and α-gurjunene were the main differential volatiles in the labellum, petals, and sepals.
A total of 46 aromatic substances was considered for OAV calculation, and the line chart is made up of 20 substances with OAV > 1 and the flowering time of different den-phals (Fig. 8c). Among the volatiles from all floral organs, (Z)-2-nonanal (violet, cucumber aromas), linalool, hexanal, (E)-2-octenal, nonanal, and octanal exhibited OAVs > 10; whereas 2-pentyl furan, (E)-2-hexenal and decanal had OAVs > 1, suggesting that these different aromatic substances play a significant role in shaping the overall aroma profile of the den-phals. In the labellum of cultivar EW, trans-β-ocimene and 2,6,6-trimethyl-1-cyclohexene-1-acetaldehyde (green lemon aroma) were significant aroma-contributing substances. The volatile 2,4-nonadienal (floral aroma) had an OAV greater than 100 in the labellum of cultivar S. In the labellum of cultivar NP, 1-octen-3-ol (mushroom aroma) had an OAV greater than 10. Trans, trans-2,4-decadienal was only a contributory volatile to the aroma in the column and spur of BC, whereas (2E,6E)-nonadienal had an OAV greater than 50 in the four floral tissues of only cultivar AB, with higher contributions in the column and spur (134.61) and labellum (120.65). Additionally, in the floral organs of cultivar AB, excluding the column and spur, D-limonene (lemony aroma) and β-myrcene (sweet orange aroma) were important aroma-contributing compounds. (E,Z)-2,6-nonadienal with an OAV > 10 contributed significantly to the floral tissue aroma of cultivars EW, AB, and CPG. In CPG, (2E,6E)-nonadienal with an OAV > 100 and heptanal with an OAV > 10 made a significant contribution to the floral aroma. Additionally, 2,4-nonadienal in the column and spur (193.05) and labellum (260.82) of CPG had an OAV > 100, and the labellum, petals, and sepals of CPG, where caryophyllene had an OAV > 1. The petals and sepals of DL contained methyl salicylate with an OAV > 5, making a significant contribution to the floral aroma.
-
In this study, flowers of 22 out of 79 den-phals that were screened were assessed in terms of their fragrance and were subjected to sensory evaluation. The majority of den-phals lack aroma. Sensory evaluation results indicated that the overall aroma intensity of the studied den-phals was relatively low, with only a few reaching strong intensities. The grassy scent was the predominant aroma type in the 22 den-phals studied. Further analysis of the aroma compounds revealed a total of 137 volatile components, predominantly aliphatics and terpenes. Based on the frequency of occurrence and concentration of each volatile compound across all 22 den-phals, the primary volatile compounds were linalool, hexanal, (E)-2-hexenal, (E)-2-octenal, nonanal, octanal, (E)-2-nonenal, 2-pentyl furan, decanal, (E,Z)-2,6-nonadienal, ylangene, copaene, γ-muurolene, α-muurolene, and α-farnesene. Previous studies had reported that the main floral volatile components of den-phals and related plants included 3-carene, linalool, α-copaene, hexanal, and (E)-2-hexenal[33,39]. The primary floral volatile compounds identified in the present study partially overlaps with those previously reported. However, due to variations in the types and concentrations of volatile compounds among different hybrids, some major compounds differed. In terms of the release of volatile substances from Osmanthus fragrans varieties, the total emissions of 'Gecheng Dangui', 'Houban Yingui', and 'Liuye Jingui' were 718.42, 803.42, and 703.47 μg/g, respectively[40], whereas the total emissions of Lilium longiflorum Thumb. was 23.60 μg/g[41]. In the current study, the aromatic substance emission of cultivar CPG in the strong fragrance category was the highest at 3.60 μg/g. Compared with other fragrant flowers, the concentration of aromatic volatiles in den-phals is extremely low. Increasing the concentration of aromatic compounds emitted from the flower is an important aspect of breeding fragrant den-phals.
This study analyzed the volatile compounds from den-phals emitted from the flower at different flowering stages and times of the day. The results revealed that the highest fragrance release occurred during the full flowering stage and at 12:00. Subsequently, the difference in volatile compounds emitted from four types of flower tissue was separately investigated under the above optimum conditions. Terpenoids were the most abundant class of floral volatile and the metabolites among all the metabolites detected at different flowering stages, time points, and tissues. This finding indicated that various terpenoid compounds, including monoterpenes, sesquiterpenes, and diterpenes, played significant roles in the composition of floral fragrances in den-phals. The emission of terpenoid volatiles is typically time-specific in plants[42]. Previous studies suggested that some specific terpenoids are expressed only at certain times in other Dendrobium species[31]. The present study suggested that, although there were differences in the floral scent compounds emitted during the early, full, and late flowering stages, certain specific compounds, such as linalool, ylangene, and copaene were present in large quantities early in the flowering process. This is similar to previous reports[43], where linalool was found to repel aphids, promoting the health of newly developed floral tissue[44,45]. Terpenoids were released during the initial opening stage of the flowers, indicating that the establishment of the floral scent characteristics of den-phals may begin in the early flowering period. With the opening of the flowers and the passage of time, there were significant decreases in the concentrations and diversity of terpenoid substances in the floral emissions.
The diurnal variation of floral compounds emitted varies among plant species. The release of the fragrance of three aroma intensity categories of den-phals exhibited an initial increase followed by a decrease over time. The maximum release of volatile compounds occurred at 12:00, which differs from some reported patterns of floral scent emission in Dendrobium spp. For example, in D. primulinum, the fragrance emission showed a decreasing trend followed by an increase, with the highest release at 06:00, gradually decreasing thereafter, reaching its lowest point at 18:00, and rising again at 21:00[32]. The trend of fragrance release in D. chrysotoxum was similar to that in the present study, both showing an initial increase followed by a decrease. However, in D. chrysotoxum, the fragrance release peaked at 14:00[31], whereas in the den-phals of the present study it peaked at 12:00. The release pattern of plant fragrance is related to environmental factors. An increase in white light enhances the release of monoterpenes in Phalaenopsis orchids[46]. Temperature variations also impact the fragrance of flowers; for instance, jasmine releases higher volatile compound concentrations at 25 or 30 °C, whereas, at 20 and 35 °C, the release of floral volatiles not only decreases but there is also a significant decrease in their endogenous levels[47]. This outcome is attributed partly to the regulation of volatile levels by biosynthetic enzymes and, partly, to vaporization caused by high temperatures[9]. The greatest aroma release from den-phals flowers at 12:00, therefore, was possibly due to optimal temperature and/or light conditions at that time.
There are differences in the aroma components emitted from different flower tissues. In Michelia crassipes, the aroma concentration is highest from the pistil, followed by tepals and stamens. Terpenes are the volatiles characteristic of pistils and tepals, whereas stamens are characterized by benzenoids[48]. In the petals, sepals, labella, and pistil columns of Cymbidium sinense, glutaraldehyde, hexanal, E-2-hexenal, and E-2-heptaneal are the main volatile components, respectively, with sepals being the main floral organ that releases volatile components[49]. The results of the current study indicate that the main aroma compounds in the fragrant den-phals are similar to those in C. sinense. The composition of volatile emissions from different floral organs of den-phals was similar, whereas the concentrations of volatiles emitted from different tissues varied significantly. Sepals are also the floral organs with the highest release of aroma compounds in den-phals.
Different volatile substances exhibit distinct aromatic characteristics, as determined by their OAVs[50]. The concentration of aromatic components cannot be the sole criterion for determining aroma characteristics. Typically, aromatic characteristics are attributed to compounds with higher OAVs. In the current study, compounds such as linalool, hexanal, (E)-2-hexenal, (E)-2-octenal, nonanal, octanal, (E)-2-nonenal, 2-pentyl furan, decanal, (E,Z)-2,6-nonadienal, and β-ocimene, characterized by elevated levels and lower thresholds, were identified as distinctive aromatic constituents of den-phals. In addition to shared aromatic components, the presence of different aromatic compounds contributed significantly to the respective aromatic profiles of the flower. These compounds collectively constituted the olfactory perception of den-phals fragrance. For example, due to the high OAV of (E)-2-hexenal present in the fragrance of cultivar S, exhibits a higher grassy aroma in the sensory performance, whereas cultivar BC has a more floral aroma due to the presence of trans-β-ocimene and β-ocimene, the OAV of ocimene being relatively high. Cultivar UB has a fruity aroma as a result of the presence of octanal and nonanal with high OAVs. Caryophyllene in cultivar CPG has a high OAV, contributing to the noticeable spicy aroma. We compared the concentrations of volatile compounds in the aroma of den-phals flowers in the light, medium, and strong fragrance categories. In the light fragrance category, the total concentrations of volatile compounds in cultivar EW exceeded that of some den-phals in the medium and strong fragrance categories, supporting the observation that the concentrations of volatile compounds cannot serve as the sole criterion for distinguishing the different levels of fragrance intensity in den-phals. Inter-cultivar differences in fragrance intensity are also related to the presence/absence of specific aromatic volatiles. For instance, although the total aroma of the strong fragrance cultivar DL is lower than that of hybrids in the medium and light fragrance categories, the OAV > 10 of methyl salicylate, DL still shows a relatively strong aroma in the sensory aspect. In the medium fragrance category, represented by cultivar AB, trans-β-ocimene had an OAV > 10, resulting in a moderate aroma. CPG, with a higher total volatile concentration, also exceeded other hybrids in terms of the concentration of caryophyllene, with OAV > 100, exhibiting strong aroma characteristics.
Floral fragrance is a highly complex trait. Changes in biochemical functions have led to the convergent evolution of biosynthetic enzymes[51,52], resulting in the production of various volatile compounds. In the experimental den-phals materials used in this study, the light fragrance cultivar EW, the medium fragrance cultivar AB, and the strong fragrance cultivar CPG exhibited a rich diversity of volatile compounds and relatively high concentrations of volatiles, making them excellent hybrid with fragrant den-phals flowers. This diversity contributes to significant differences in the emission of floral scents among hybrids. The first step in breeding varieties with improved fragrance-related traits is to compile comprehensive information about the volatile compounds involved. For the first time, the current study has comprehensively investigated the changes in volatile aromatic compounds associated with fragrance in den-phals concerning flowering period, time of the day, and floral organ. This research provides a strong reference base for future exploration of the synthesis of floral scents in den-phals and subsequently the breeding of commercially significant fragrant varieties.
-
Twenty-two den-phals were grouped into three categories, light fragrance (five hybrids), medium fragrance (nine hybrids), and strong fragrance (eight hybrids), based on different aroma intensities. The grassy aroma is primarily contributed by hexanal, and (E)-2-hexenal. The floral aroma is characterized by linalool, β-ocimene, and trans-β-ocimene. Octanal and nonanal are the main aroma compounds, providing the fruity aroma, while caryophyllene contributes to the spicy aroma. The concentration of volatile compounds was highest during the full flowering stage, at 12:00, and in the sepals. In response to the opening of the flowers or the progression of time during the day, the concentration of volatiles first increased and then decreased. Linalool, hexanal, (E)-2-hexenal, (E)-2-octenal, nonanal, octanal, (E)-2-nonenal, 2-pentyl furan, decanal, (E,Z)-2,6-nonadienal, and β-ocimene were common fragrance compounds in den-phals. In addition to the aforementioned common fragrance substances, the light fragrance type contained aromatic compounds, such as (2E,6E)-nonadienal, 2,6,6-trimethyl-1-cyclohexene-1-acetaldehyde, β-phellandrene, and (E,E)-2,4-hexadienal. The medium fragrance type featured fragrance compounds such as β-myrcene, trans-β-ocimene, and D-limonene, while the strong fragrance type contained fragrance compounds like heptanal, caryophyllene, methyl salicylate, and 2,4-nonadienal. The light fragrance type cultivar EW, the medium fragrance type cultivar AB, and the strong fragrance type cultivar CPG were all very rich in both the diversity and concentration of aromatic compounds, making them excellent germplasm resources for the breeding of den-phals with improved fragrance characteristics.
-
The authors confirm contribution to the paper as follows: writing-original daft, writing – review & editing, investigation: Yang X; funding acquisition, investigation: Yin J; resources, formal analysis: Lu S; investigation, resources: Liao Y; project administration, supervision: Zheng R; project administration, supervision, writing – review, funding acquisition: Li C. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This work was supported by the Innovational Fund for Scientific and Technological Personnel of Hainan Province (Grant No. KJRC2023C19); the Central Public-interest Scientific Institution Basal Research Fund (Grant No. 1630032022002); and the earmarked fund for CARS-23-G60.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Volatile substances of den-phal.
- Supplemental Fig. S1 Permutation test results.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang X, Yin J, Lu S, Liao Y, Zheng R, et al. 2024. The characterization of volatile floral scent in Phalaenopsis-type Dendrobium hybrids. Ornamental Plant Research 4: e023 doi: 10.48130/opr-0024-0023 

The characterization of volatile floral scent in Phalaenopsis-type Dendrobium hybrids
- Received: 07 May 2024
- Revised: 28 June 2024
- Accepted: 22 July 2024
- Published online: 09 August 2024
Abstract: Phalaenopsis-type Dendrobium hybrids (den-phals) are globally important ornamental plants. Aromatic commercial den-phals are rare, but the fragrance of their flowers has gradually become a significant trait influencing consumer preferences. In this study, 22 aromatic den-phals were screened by sensory evaluation. Then headspace solid–phase microextraction and gas chromatography-mass spectrometry (GC–MS) were employed to investigate the aroma profiles of nine out of the 22 aromatic den-phals at different flowering stages, times of the day, and floral organs. The results showed that the 22 den-phals could be categorized into three groups, based on different aroma intensities: light fragrance (five hybrids), medium fragrance (nine hybrids), and strong fragrance (eight hybrids). A total of 137 volatile compounds were detected in all samples according to the GC–MS results. Both the quantity and variety of volatile compounds exhibit an initial increase followed by a decrease during different flowering stages and times of day. Full flowering stage reached its peak, with its highest content at 12:00, with the highest types at 16:00. Petals have the highest variety of volatile compounds, while sepals have the highest content. The odor activity values indicated that linalool, hexanal, (E)-2-hexenal, (E)-2-octenal, (E,Z)-2,6-nonadienal, (E)-2-nonenal, octanal, nonanal, 2-pentyl furan, decanal, and β-ocimene contributed significantly to the aroma of den-phals. The grassy aroma of den-phals was primarily contributed by hexanal and (E)-2-hexenal, and the floral aroma was characterized by linalool, β-ocimene, and trans-β-ocimene. Octanal and nonanal were the main aroma compounds associated with a fruity aroma, while caryophyllene contributed to a spicy aroma.
-
Key words:
- Characterization /
- Volatiles /
- Floral /
- Scents /
Phalaenopsis-type Dendrobium hybrids