










-
The evolutionary journey of plants form ocean to terrestrial environment is accompanied with the shift of sexual reproduction manner from zoidogamy to siphonogamy. Angiosperm sperm have lost their mobility and are delivered to the female gametes through pollen tubes. When pollen grains are deposited on the stigma of flower, a tip-growing pollen tube forms and elongates into the style and then in the transmitting tract, which marks the initiation of pollen tube guidance[1]. Thereafter, in the extracellular matrix of transmitting tract, pollen tubes continuously perceive and respond to the guiding 'signals' from the female gametophytes to grow towards the ovule, which process is called pollen tube guidance. This polarized pollen tube growth, culminating in arriving at the receptive synergid and subsequent releasing two sperm cells. Over the last two decades, these guiding 'signals' are uncovered to be CRP-type peptides in dicots, including LUREs, XIUQIUs, TICKETs, as well as SALVAGERs[2−6]. Meanwhile, synergids and the central cell are validated to be the primary sources of these pollen tube attractants[7, 8].
CRPs are overrepresented in the plant peptide superfamily and play important roles in plant development, defense, symbiosis, and reproduction[4, 9, 10]. The hallmark of CRPs are the cysteines in their sequences, and intramolecular disulfide bonds between cysteine residues determines the three-dimensional structure of CRPs[4, 9]. CRPs are subdivided into subgroups with specific conserved cysteine-distribution patterns, including defensins, thionins, RALFs, and lipid-transfer proteins (LTPs), among others[11]. Moreover, CRPs among different subgroups or different species are highly polymorphic in sequences[11]. The reported defensin-like attractants in dicot plants are characterized with six cysteines that stabilize the CSαβ and γ-core motifs by disulfide bonds, including LUREs in Torenia fournieri and LURE1s, XIUQIUs, TICKETs, SALVAGERs in Arabidopsis and other relatives[3, 5, 6, 8, 9]. AtLURE1s are species-preferential attractants, which promote emergence of conspecific pollen tube from transmitting tract, while XIUQIUs, as AtLURE1-related CRPs entice pollen tube with no species preference[3]. By contrast, the LURE1-related TICKETs attract solely the conspecific pollen tubes[5]. AtLURE1s, AtXIUQIUs and AtTICKETs belong to CRP_810 group and are transcriptionally regulated by transcription factor MYB98, the master regulator in synergids[12, 13]. The recently identified SALVAGER1 and SALVAGER2 are central cell-derived CRP-type attractants, which belong to the CRP_0300 group and CRP_0220 group, respectively, and mediate fertilization recovery in the case of fertilization failure with two synergid cells consumed[8]. However, a hendecuple mutants with lesions in all four XIUQIUs and seven AtLURE1s exhibited only a slight compromise in pollen tube guidance and fertility, suggesting the existence of other unknown attractants[3]. Unlike the case in dicot, the monocots utilize non-CRP-type pollen tube attractants[14, 15]. In Zea mays, the synergids and egg cell secrete Egg Apparatus1 (EA1) peptide to attract pollen tube into the embryo sac. Moreover, transgenic down-regulation of EA1 induces severe ovule sterility[14, 15]. It appears that dicots and monocots employ different types of peptide attractants. Intriguingly, mammals apply diverse chemoattractants in guiding the sperm to the egg cell[16]. It is tempting to speculate that there are other types of attractant evolved in different angiosperm lineages.
To date, no non-defensin peptide attractant has been identified in Arabidopsis. In this study, a non-defensin peptide NPA1 is characterized to be a new pollen tube attractant in Arabidopsis. NPA1 is expressed in the synergid cells and downstream of MYB98. Expression of NPA1 in synergids increases the pollen tube attraction efficiency of myb98 mutant ovules. Taken together, this study identifies a new type of peptide attractant, reveals the diversity of attractants in plants and opens the revenue for the further study of the signaling pathway.
-
Seeds of Arabidopsis thaliana were sterilized and plated onto Murashige and Skoog media supplemented with 40 mg/L hygromycin for transgenic lines. The plants were grown at 22 °C with 16 h of light and 8 h of darkness. Arabidopsis thaliana Columbia-0 (Col-0) was used as the wild-type (WT) control. Mutant seeds of myb98 (SALK_020263) were obtained from the Arabidopsis Biological Resource Center (ABRC). npa1 npa2 mutant were generated by CRISPR/Cas9 technology in Col-0 background.
RT-PCR
-
Total RNA was extracted from mature ovule of myb98 and Col-0 using RNeasy Plant Mini Kit (QIAGEN), and reversely transcribed to cDNA after the contaminated genome DNA was removed. ACTIN was used as the internal control for quantitative normalization. Thirty PCR cycles were used for amplification. The specificity of the primers was confirmed by DNA sequencing of the products after electrophoresis. For primer information see Supplemental Table S1.
Vector construction and plant transformation
-
For the GFP reporter expression, genomic sequences containing 2-kb native promoters and genomic coding sequences for NPA1 were fused to the GFP sequence to generate pCAMBIA1300-proNPA1:NPA1-GFP-TerNOS and pCAMBIA1300-proNPA2:NPA2-GFP-TerNOS. For complementation assay, two MYB98 promoters and NOS terminator were inserted into pCAMBIA1300, respectively. The genomic sequences of NPA1/2, LURE1.2, and LURE1.5 containing the 3'-terminators were inserted after the first MYB98 promoter, respectively. GFP sequence was inserted at the 3' end of the second MYB98 promoter to generate pCAMBIA1300-proMYB98:NPA1-ter-proMYB98:GFP-TerNOS. For null mutant of npa1 npa2 constructed by CRISPR/Cas9 technology, dual spacers were amplified from pCBC-DT1T2 and cloned into vector pHEE401E[17]. For primer information see Supplemental Table S1.
Phenotypic analysis
-
For seed set scoring, the silique of 10 d after pollination (DAP) were observed and recorded with a CCD camera (Leica, DFC450). For in vivo pollen tube growth and aniline blue staining, flowers at 12c stage were emasculated and left to grow for 12−24 h to achieve pistil maturation. Then, the pistils with mature stigmas were pollinated with wild-type pollen grains. After 3, 5, 8, 12, 24, 36, and 48 h, respectively, the pistils were excised and fixed in Carnoy's fixative (75% ethanol and 25% acetic acid) for at least 4 h. The pistils were washed three times with 50 mM PBS buffer (NaHPO4/NaH2PO4, pH 7.0) and immersed in 1 M NaOH for 12 h. Then after three washes with PBS, the pistils were stained with 0.1% aniline blue (pH 8.0 in 0.1 M K3PO4) for 4 h. The stained pistils were observed with a fluorescence microscope (ZEISS, Axioskop2)[18−20].
Protein purification
-
The coding sequence of NPAs and LURE1.2 lacking the putative N-terminal signal peptides (21, 20 and 19 amino acids, respectively) were inserted to the pET28a vector in fusion with the N-terminal His tag, and then transformed into Escherichia coli strain BL21 (DE3). Cells were grown to A600 = 0.6 at 37 °C and then induced with 1 mM isopropyl-β-d-thiogalactopyranoside (IPTG) for 16 h at 20 °C. Then the peptides were purified from the E. coli lysate as described previously[21].
Semi-in vitro pollen germination and guidance assay
-
For semi-in vitro pollen germination, the wild-type pollen grains were pollinated on the emasculated Col-0 stigma for 20 min. Then, the stigmas were cut and put on the germination media (1 mM CaCl2, 1 mM Ca(NO3)2, 1 mM MgSO4, 0.01% H3BO4, 18% sucrose and 1% agarose) and cultured for 4 h at 28 °C in dark. For attraction activity assay, gelatin beads containing different peptides were prepared and placed beside the pollen tube tip using a micro-manipulator as previously reported[5]. The attraction of pollen tubes was monitored and recorded with a CCD camera (ZEISS, Axiocam 506 mono).
Immunofluorescence
-
For pollen tube binding assay, the wild type pollen grains were germinated in semi-in vitro conditions. The diluted His-tag fused peptides of NPA1, NPA2, LURE1.2 or TBS buffer were dropped on the pollen tubes for 2 or 5 min. Then, the samples were immediately fixed in 4% paraformaldehyde (50 mM HEPES, 1 mM CaCl2, 1 mM MgSO4, 5% Sucrose) for 30 min, and then transferred to 3% bovine serum albumin (BSA) buffer for 2 h. Subsequently, the samples were incubated with the primary monoclonal anti-His antibody (1:500, Cwbio, CW0286) in PBS buffer containing 3% bovine serum albumin (BSA), overnight at 4 °C. Then, the samples were washed three times with PBS buffer before incubation with FITC-conjugated secondary antibodies (1:100, Abmart, M212308M) in 3% BSA buffer for 2 h in darkness at 4 °C. After washing at least three times, the fluorescence signal was observed using a confocal microscope (excitation/emission wavelength: 488 nm/505–530 nm).
Microscopy
-
For localization assay of NPA1 and NPA2, Zeiss 780 two-photon laser scanning confocal microscope and Zeiss 980 laser scanning confocal microscope (Carl Zeiss) were used to acquire the images with the excitation/emission of 488 nm/505–530 nm.
Bioinformatics and phylogenetic analysis
-
The dataset of RNA-seq was obtained from NCBI: SRP160651, NCBI: PRJNA527221 and GSE8392[3, 22, 23]. Phylogenetic tree was drawn with MEGA7 using the neighbor-joining method with bootstrapping of 1,000 replicates. The sequence alignment was performed using ClustalW (
www.ebi.ac.uk/Tools/msa/clustalw2 ). Signal peptides were predicted by SignaP-4.1 Server (https://services.healthtech.dtu.dk/service.php?SignalP-4.1 ). Guide RNA design for gene editing by CRISPR/Cas9 gene editing was performed on CRISPOR (https://crispor.tefor.net/ ). The three-dimensional structure of NPA1 and NPA2 was predicted by AlphaFold Protein Structure Database (www.alphafold.ebi.ac.uk ). -
To investigate the existence of non-defensin peptides acting as pollen tube attractants in Arabidopsis, we searched for the candidate peptides in the reported transcriptome data[11, 22, 23]. Three gene clusters from published transcriptome datasets were selected to identify the synergid-derived non-CRPs, including the down-regulated genes in myb98 mutant ovules, the synergids-expressed genes, and genes of non-CRPs expressed in the wild type Arabidopsis[11, 22, 23]. As a result, six genes were identified to be shared by these three gene groups (Fig. 1a). Among them, one gene candidate (AT4G02655) exhibits high expression level in the ovule and encodes a peptide with a signal peptide. AT4G02655 was then named NON-DEFENSIN PEPTIDE ATTRACTANT 1 (NPA1) based on motif analysis of protein sequences (Fig. 2). After screening, AT1G18486 is the sole homolog of NPA1 in Arabidopsis genome and was termed NPA2 (AT1G18486). The mRNA of NPA1 and NPA2 were only detected in the pistil, instead in the root, leaf, or pollen by RT-PCR (Fig. 1b). These results are consistent with the transcriptome data, which showed NPA1 and NPA2 are expressed in ovules and early seeds[24]. Additionally, RT-PCR also validated the transcriptome data that transcription of NPA1 and NPA2 are hardly detectable in myb98 ovules in comparison to the wild type (Fig. 1c, d). To confirm the expression pattern of NPA1 and NPA2, plants expressing the genomic NPA1 or NPA2 fused with green fluorescent protein (GFP) under their native promoter were obtained. The fluorescent signal of NPA1-GFP and NPA2-GFP were initially detected in the synergids of immature ovules at FG5 stage after cellularization, and then dispersed in the filiform apparatus and the plasma membrane invagination facing the micropyle at synergid maturation stage (Fig. 1e). Moreover, the TAAC element was found to be present in the promoter of NPA1 at −925, −258, and −121 sites, and of NPA2 at −169, −56 and −34 sites, which has been reported to be the binding site of MYB98[13] (Fig. 1f).
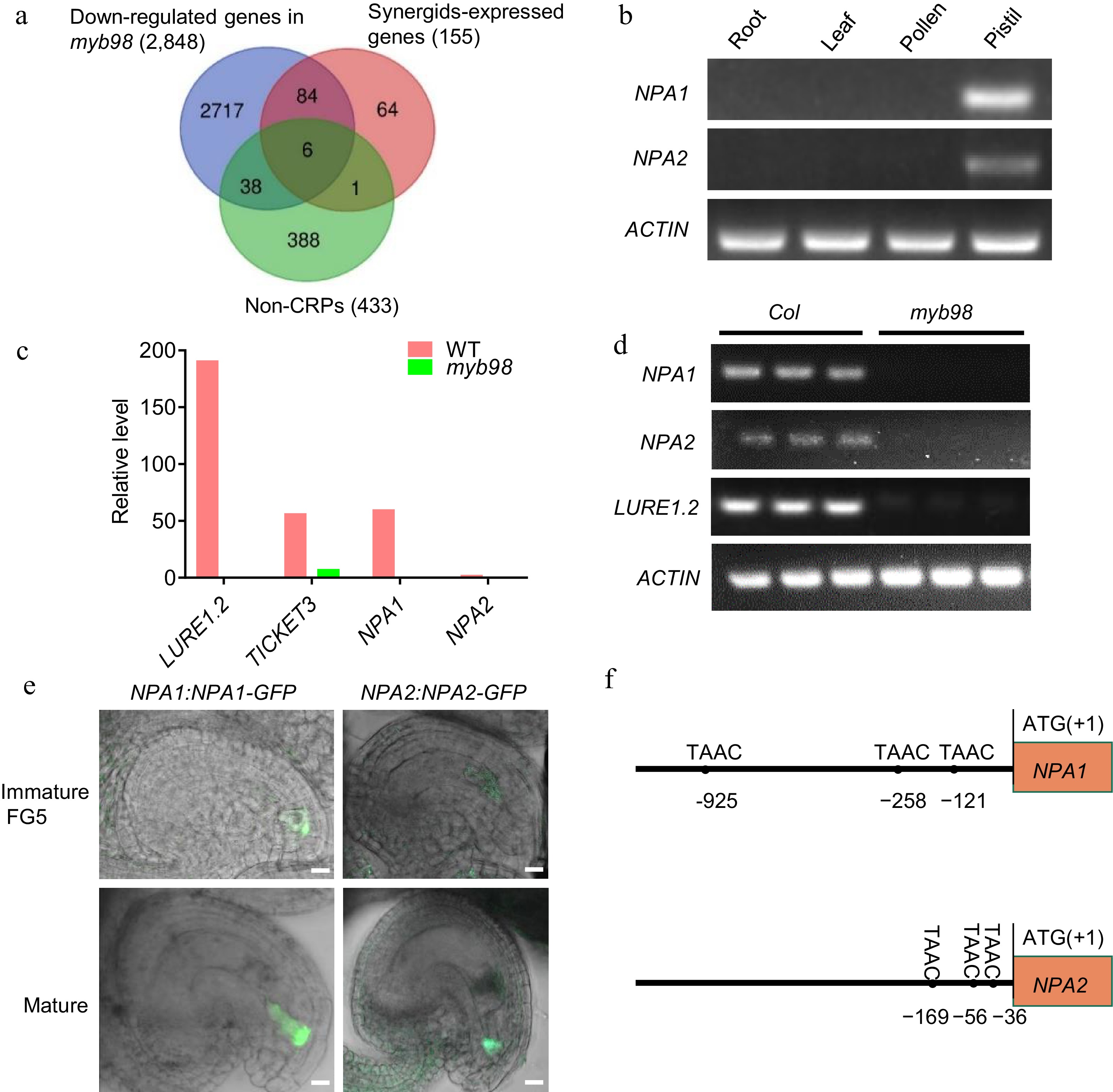
Figure 1.
(a) Venn diagram of the down-regulated genes in myb98, synergids-expressed genes, and non-CRPs. (b) RT-PCR showing the expression patterns of NPA1 and NPA2 in different tissues of Arabidopsis thaliana. ACTIN, internal control. (c) Relative level of transcripts of LURE1.2, TICKET3, NPA1 and NPA2 from the reported transcriptome data. (d) RT-PCR showing the mRNA level of LURE1.2, ACTIN, NPA1 and NPA2 in the WT and myb98 mature ovules. (e) Expression of NPA1 and NPA2 in the synergids of immature and mature ovules. (f) The sites of TAAC element in the promoters of NPA1 and NPA2.
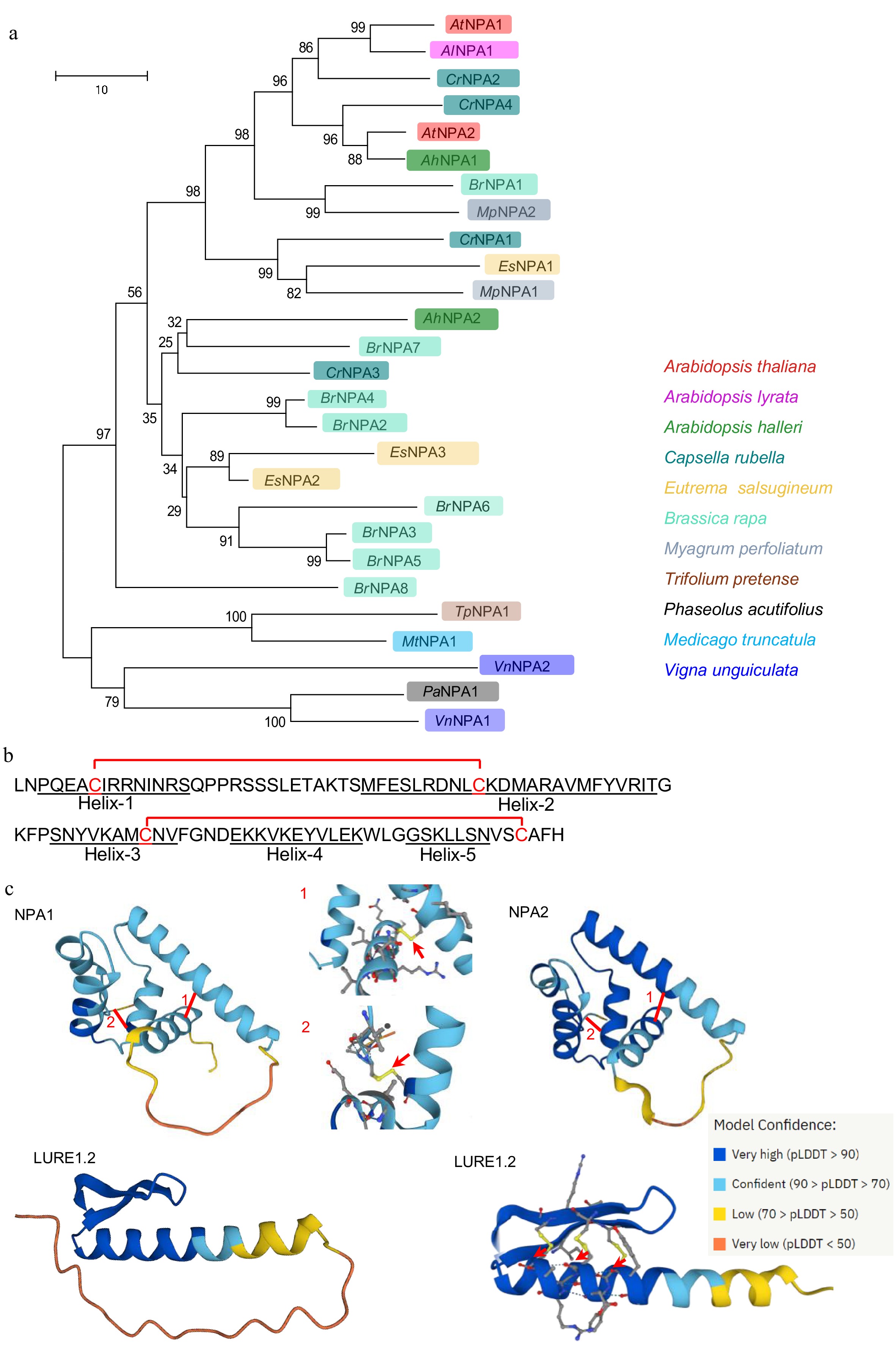
Figure 2.
The phylogeny and protein structure of NPAs. (a) Phylogenetic tree constructed with the protein sequences of NPA1 and NPA2 and their homologs. (b) Mature peptide sequence of NPA1 with five predicted helices and two disulfide bonds. (c) The structural modeling of NPA1, NPA2 and LURE1.2 by AlfaFold. Arrows indicate disulfide bonds.
Homologs of NPA were present in many close relatives of Arabidopsis thaliana in Brassicacea, such as Arabidopsis lyrata, Arabidpsis helleri, Capsella rubella, Eutrema salsugineum, Brassica rapa, as well as some Leguminosae species (Fig. 2a). NPA1 (121 aa in full length) and NPA2 (118 aa in full length) were previously classified as non-CRPs[11, 25], although NPA1 and NPA2 indeed contain four cysteines that are conserved among Brassicacea species (Supplemental Fig. S1). As the classification of CRPs is based on a range of artificial criteria, such as the number of cysteine residues and sequence conservation[11, 25], NPAs were safely classified as non-defensin peptides in this study. Structural modeling using the AlphaFold program showed that both NPA1 and NPA2 possess five α-helixes. These helixes are connected by two disulfide bonds, which is distinct from the CRP_810 peptides that form an α-helix and a two-strand anti-parallel β-sheet connected by three disulfide bonds (Fig. 2b, c)[6]. These results suggest that NPA1 and NPA2 belong to a new non-defensin-type peptide family that are specifically expressed in the synergids under the control of MYB98.
NPA1 exhibits pollen tube attraction activity
-
To analyze whether NPA1 and NPA2 are capable to attract pollen tubes, mature NPA1 and NPA2 peptides, and AtLURE1.2 were synthesized using prokaryotic expression system, whereby the gene sequences of NPA1, NPA2, and AtLURE1.2 without signal peptide were fused with the His-tag and expressed in E. coli, respectively (Supplemental Fig. S1). Western blot verified the successful purification of NPA1, NPA2, and LURE1.2 peptides with the desired protein sizes (Fig. 3a). Semi-in vitro pollen tube guidance assay was then performed with gelatin beads containing the purified peptides. The results showed that wild-type pollen tube can be attracted by LURE1.2 and NPA1, but not by NPA2 and negative control (Fig. 3b, c). Of note, the attraction activity of NPA1 appears weaker than LURE1.2 (Fig. 3c). Moreover, mutation versions of NPAs (NPA1-M and NPA2-M) with substitution of four conserved cysteines by alanine were also examined in the semi-in vitro pollen tube guidance assay. The results showed that pollen tube attraction ability is fully abolished for NPA1-M (Fig. 3b, c), suggesting the importance of these four cysteines for NPA1 in attracting pollen tube. To confirm whether NPA1 binds to the pollen tubes, we performed peptide binding assay by immunostaining with the purified His-tagged peptides. Once applied to the pollen tube, His-NPA1, like LURE1.2 bound to the plasma membrane of wild type pollen tube tips in 2.5 min and was further internalized by endocytosis in 5 min (Fig. 3d). In contrast, His-NPA2 and His-NPA1-M did not bind the pollen tube plasma membrane (Fig. 3d). These results suggest that NPA1, but not NPA2 is capable of attracting pollen tubes through directly binding to the pollen tube plasma membrane, in which process the four conserved cysteine residues are pivotal.
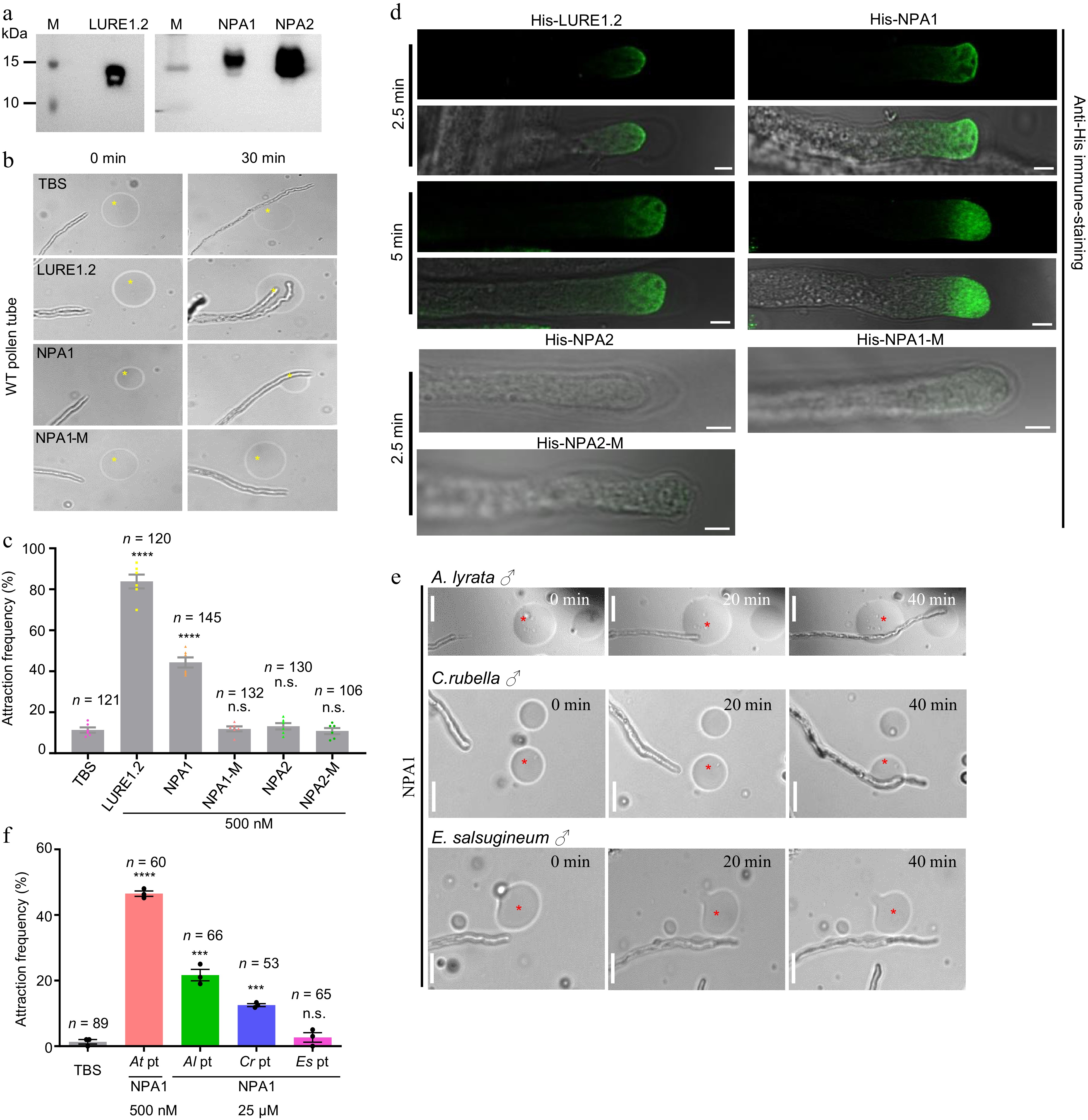
Figure 3.
NPA1 has pollen tube attraction activity. (a) Western blot of the purified His-tagged LURE1.2, NPA1 and NPA2 stained by His antibody. (b) Pollen tube attraction assay with gelatin beads containing the purified peptides, bar = 10 μm. (c) Statistics of (b), n = 121, 120, 145, 132, 130, 106 for each sample. The concentration for each assay is 500 nM. Two-tailed Student's t-test, **** p < 0.0001. Three independent biological experiments were repeated. (d) His-NPA1 binds to the pollen tube tip membrane. The LURE1.2 and NPA1 were incubated with the WT pollen tubes for 2.5 and 5 min, respectively. Endocytosis was observed at 5 min, n = 370 and 96 for each sample. NPA2, NPA1-M and NPA2-M were incubated with the WT pollen tubes for 2.5 min, n = 23, 19, 35, Bar = 5 μm. Three independent biological experiments were repeated. (e) Pollen tube attraction assay with gelatin beads containing the purified peptides. Pollen tubes of A. lyrata, C. rubella and E. salsugineum were used, bar = 10 μm. (f) Statistics of (e), n = 89, 60, 66, 53 and 65 for each sample. Two-tailed Student's t-test, **** p < 0.0001, *** p < 0.001. All data shown are the mean ± s.e.m. Three independent biological experiments were repeated.
To investigate the role of NPA1 in reproductive isolation among the close relatives in Brassicaceae, we examined the attraction activity of NPA1 to pollen tubes of different species. As the pollen tubes of A. lyrata and C. rubella germinate and grow well within the transmitting tract of A. thaliana pistil[5], the pollen grains of A. lyrata, C. rubella, and E. salsugineum were pollinated to the pistil of A. thaliana in semi-in vitro pollen tube guidance assay. NPA1 attracted the pollen tubes of both A. lyrata and C. rubella with a relative lower efficiency than that of A. thaliana but showed no attraction to E. salsugineum pollen tubes (Fig. 3e, f). This result suggests that NPA1 is a species-preferential attractant, preferring conspecific pollen tube over pollen tubes of other species.
To investigate whether NPA1 is perceived by the known receptors on pollen tube plasma membrane, we performed pollen tube attraction assay with the mutants of LURE receptors and their homologous genes. The result showed that the mutants of the Pollen Receptor Kinase (PRK) family members, MALE DISCOVERER 1 (MDIS1) and MDIS2 exhibited normal attraction response to NPA1 (Fig. 4a, c). In addition, NPA1 induced wavy growth pattern of the WT and these mutant pollen tubes, like the effect of LURE1.2, while NPA1-M did not (Fig. 4b). These results suggest that NPA1 acts through a new signaling pathway.
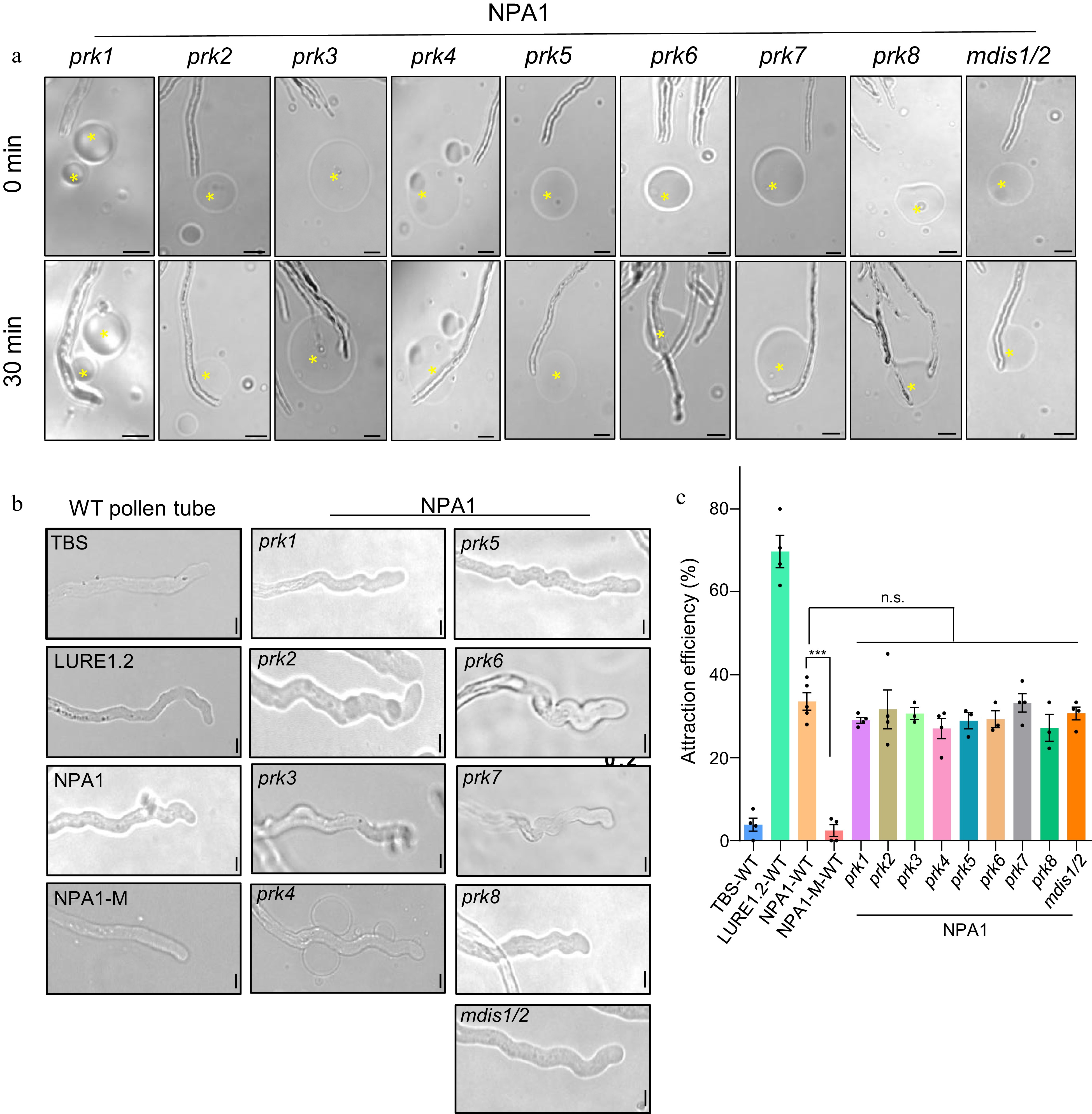
Figure 4.
NPA1 attracts the mutant pollen tubes of PRK family members and mdis1/2. (a) NPA1 attracted pollen tubes of different mutants, bar = 20 μm. (b) NPA1 induced wavy growth patterns of pollen tubes. The concentration of LURE1.2, NPA1 and NPA1-M was 500 nM, bar = 5 μm. (c) Statistics of (a), n = 82, 64, 145, 70, 55, 62, 48, 54, 51, 45, 55, 53, 69. Two-tailed Student's t-test, *** p < 0.001, n.s., no significance. The concentration of LURE1.2 was 500 nM, NPA1 and NPA1-M 500 nM. All data shown are the mean ± s.e.m.
Expression of NPA1 in the synergids alleviates the phenotype of myb98 mutant
-
To investigate the biological function of NPA1 and NPA2 in vivo, three independent npa1 npa2 null mutants (npa1 npa2-1, npa1 npa2-2, and npa1 npa2-3) were created by knocking out these two genes in the wild type plants using CRISPR/Cas9 gene-editing (Supplemental Fig. S2). These three npa1 npa2 null mutants exhibited no seed set defect (Fig. 5a, b). Moreover, there was no difference observed between wild type and these double mutants regarding pollen tube attraction at 5 h after pollination (HAP), 8 and 12 HAP (Fig. 5c, d; Supplemental Figs S3, S4). The normal seed set and pollen tube guidance of these npa1 npa2 mutants are expectable as more than ten CRP attractants are still present in the synergids[3]. MYB98 is the predominant regulator in synergids that governs the transcription of multiple pollen tube attractants[12, 13]. To test the in vivo function of NPAs, NPA1 and NPA2, together with LURE1.2 and LURE1.5 were expressed in the myb98 mutant under MYB98 promoter, respectively. In this context, the transcription of MYB98 promoter was not affected by the knockout of MYB98[13]. The results showed that the seed set defect of myb98 was significantly alleviated in the transgenic lines expressing NPA1 or LURE1.2, but not in the lines expressing NPA2 or LURE1.5 (Fig. 5e, f).
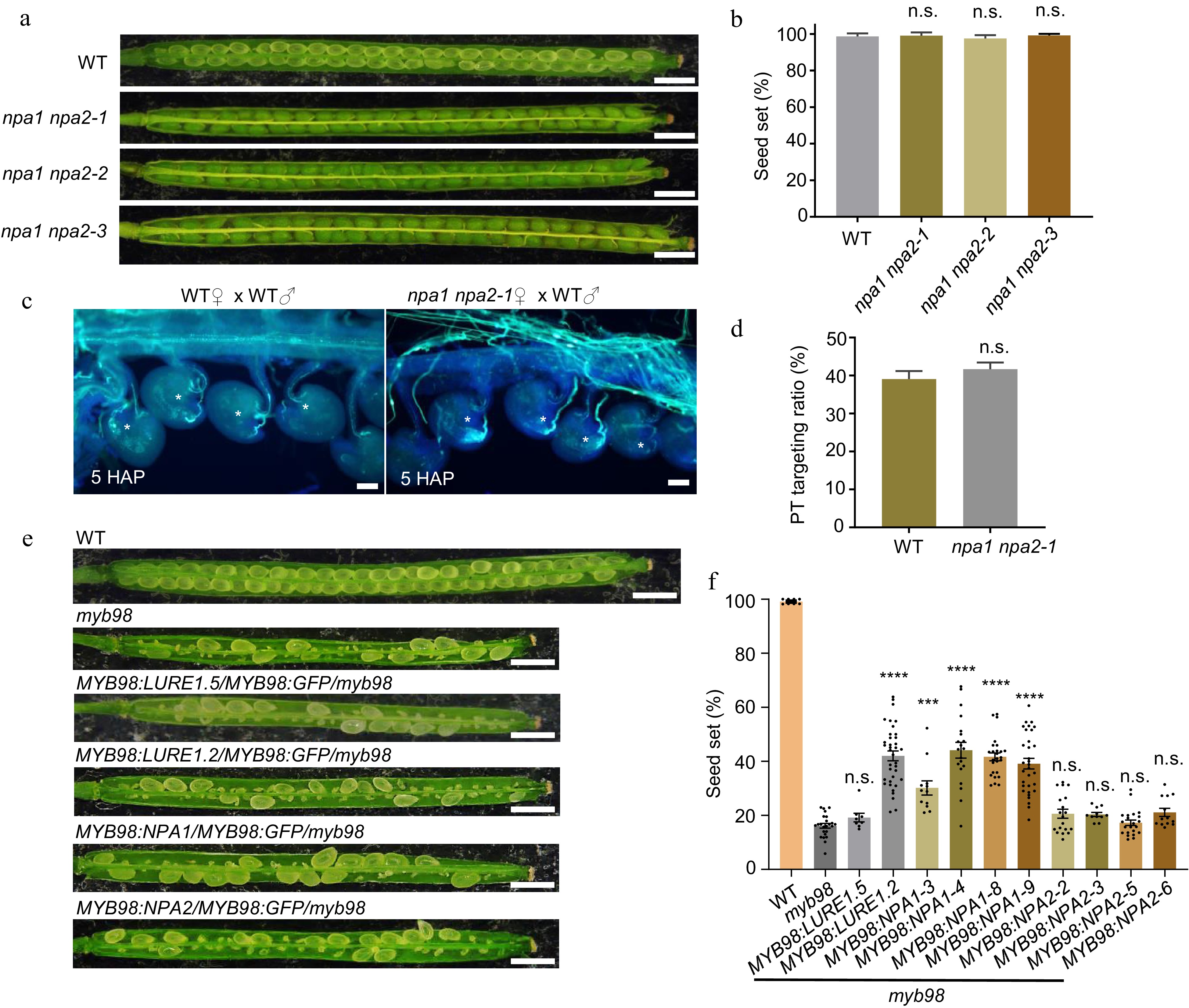
Figure 5.
The function of NPA1 in seed production. (a) npa1 npa2 null mutant shows full seed set, bar = 1 mm. (b) Statistics of (a), n = 1,190, 865, 907, 905 for each sample. Two-tailed Student's t-test. (c) The pollen tube attraction efficiency at 5 HAP, bar = 50 μm. (d) Statistics of (c), n = 736, 1007 for each sample. Two-tailed Student's t-test. (e) Seed set of myb98 was increased by the expression of NPA1 in the synergids, bar = 1 mm. (f) Statistics of (e), n = 1,068, 1,294, 1,401, 587, 832, 1,043, 1,264, 927, 606, 942, 971 for each sample. Two-tailed Student's t-test, **** p < 0.0001. All data shown are the mean ± s.e.m. Three independent biological experiments were repeated.
To confirm whether NPA1 increases the seed set rate of myb98 by alleviating the pollen tube guidance defect, pollen tube attraction efficiency of these transgenic plants was evaluated. The results showed that pollen tube attraction efficiency of the transgenic ovules expressing NPA1 or LURE1.2 was significantly higher than that of myb98 mutant ovules at 12, 24, and 36 HAP (Fig. 6). These results indicate that NPA1 can compensate the loss of pollen tube guidance ability in myb98, thereby increasing the seed set rate of myb98.
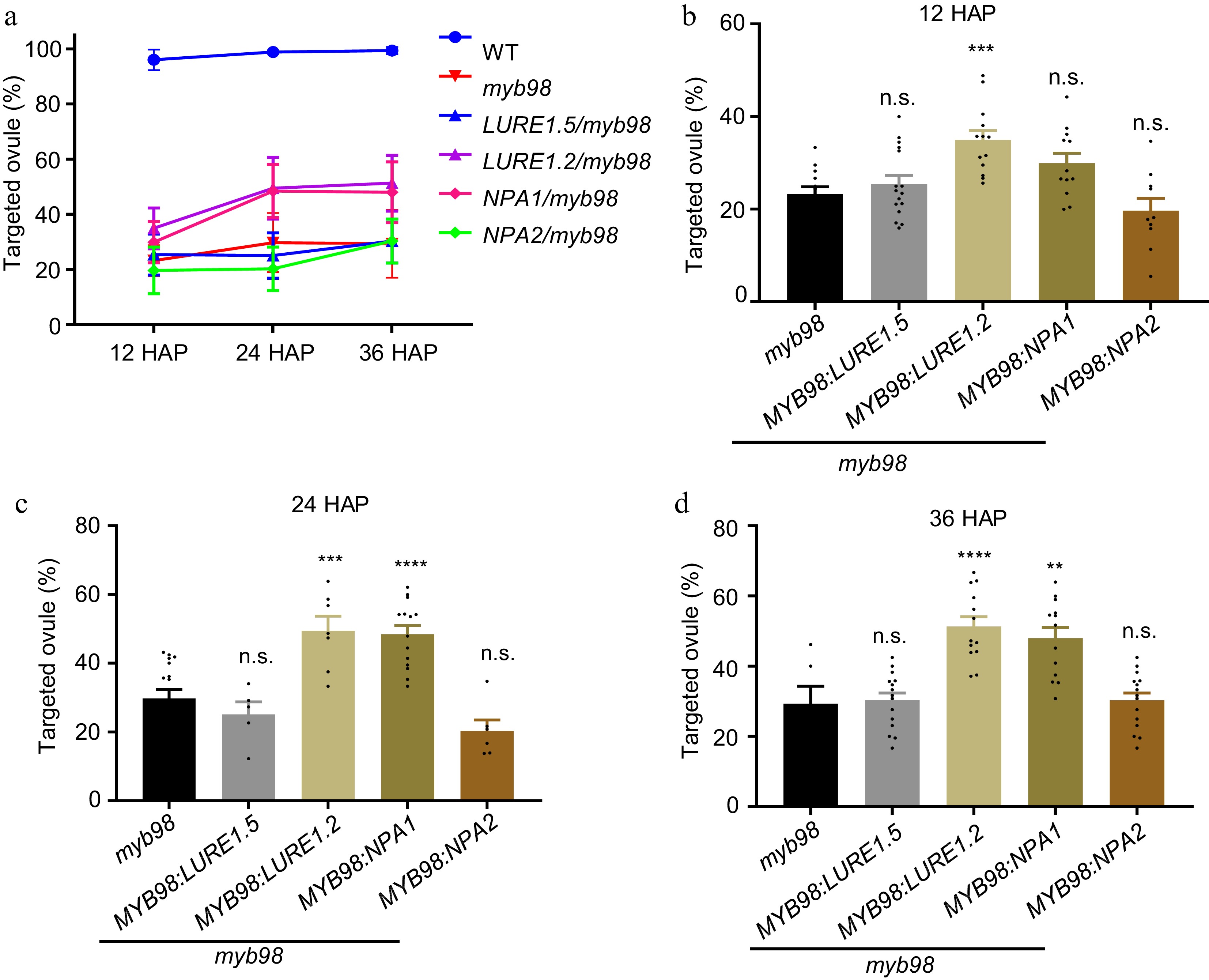
Figure 6.
Expression of NPA1 in the synergids alleviates the defective pollen tube attraction of myb98 mutant. (a) The percentage of targeted ovules. Statistics of the pollen tube attraction efficiency at (b) 12, (c) 24, (d) 36 HAP. Data in (b), n = 604, 595, 794, 477 for each sample. Two-tailed Student's t-test, *** p = 0.0002. Data in (c), n = 723, 241, 406, 660, 361 for each sample. Two-tailed Student's t-test, *** p = 0.0005, **** p < 0.0001. Data in (d), n = 249, 699, 544, 495, 701 for each sample. Two-tailed Student's t-test, **** p < 0.0001, ** p = 0.0040. All data shown are the mean ± s.e.m. Three independent biological experiments were repeated.
NPA1 does not alleviate the defective pollen tube reception and seed development of myb98 mutant
-
During analyzing the fertility rescuing effect of NPA1 in myb98, we noticed that the expression of NPA1 increased the pollen tube targeting rate of myb98 from 30% to more than 50% at 36 HAP (Fig. 6d). However, the seed setting rate of myb98 was only rescued to 40% in MYB98:NPA1 and MYB98:LURE1.2 plants. This percentage discrepancy between targeted-ovule and seed set implies that myb98 has fertility defect post pollen tube targeting.
To confirm this speculation, we carefully examined and quantified the pollen tube behaviors and seed development in myb98 and the rescued lines. Among them, pollen tube overgrowth is a sign of failed pollen tube reception post pollen tube guidance. The results showed that myb98 plants exhibited pollen tube overgrowth within the embryo sac and aberrant seed development (Fig. 7a−c). The statistical data showed that expression of NPA1 and LURE1.2 in myb98 mutant only decreased the percentage of pollen tube guidance defect, but not the pollen tube overgrowth and seed development (Fig. 7d). Together, a new type of secreted peptide NPA1 expressed in the synergid functions as a pollen tube attractant in planta.
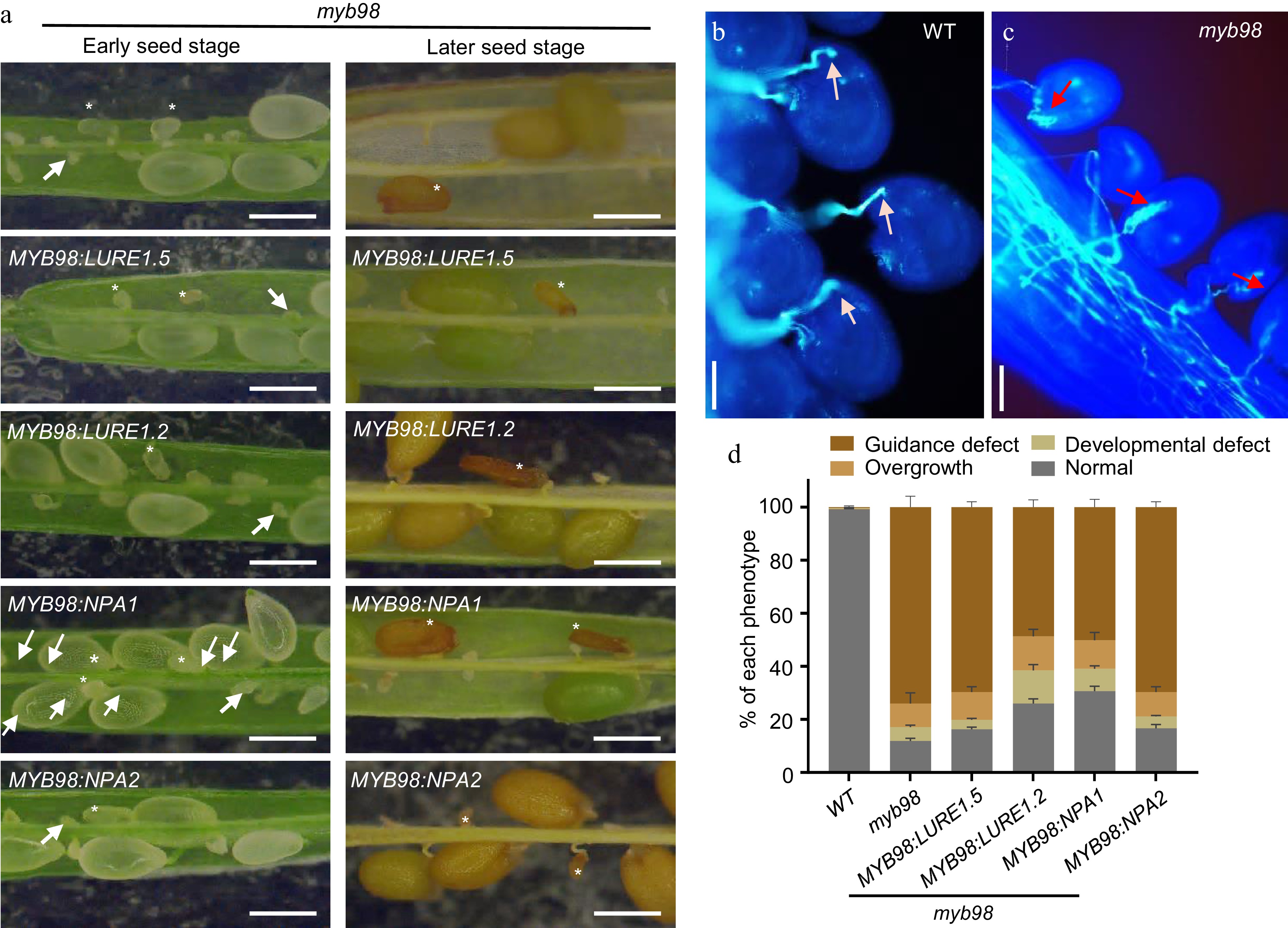
Figure 7.
NPA1 does not rescue the defective pollen tube reception and seed development of myb98 mutant. (a) The seed set of myb98 and the rescued plants at the early and late developmental stages. Asterisks, aborted seeds. Arrow, aborted ovules, bar = 1 mm. (b) and (c) Pollen tube overgrowth in myb98 ovules, bar = 20 μm. White arrow, normal pollen tubes. Red arrow, overgrowing pollen tubes. (d) Statistics of pollen tube guidance defect, pollen tube overgrowth, seed development defect, and normal seeds. Data shown are the mean ± s.e.m. Three independent biological experiments were repeated.
-
Secreted peptides constitute a large gene family and function in diverse cell-to-cell communication in plants. In the model plant Arabidopsis, > 1,000 putative peptides have been predicted[26]. However, partially due to the functional redundancy caused by gene duplication, only a small fraction of them have been functionally studied[4, 27]. To date CRPs, like defensin-like peptides and RALFs, and cleaved small peptides, like CLE and phytosulfokine (PSK), constitute the most characterized secreted peptides[4, 27]. NPA1 as a new type of peptide, its characterization would expand our knowledge on the function of the large repertoires of secreted peptides.
The identification of NPA1 suggests diversity of the pollen tube attractants and its underlying biological relevance. Distinct from the synergid-derived CRP_810 group defensin-like attractants and the central cell-derived CRP-type attractants[8], NPA1 has fewer cysteines and a totally different conformation, which is consistent with its new perception mechanism. Due to the high functional redundancy of the more than a dozen attractants, selection constraint could be alleviated to allow the accumulation of mutations that contribute to species-specificity and gene loss with high efficiency. For instance, in the CRP_810 subgroup, LUREs and TICKETs are species-specifically active, while LURE1.5 and TICKET1 lose the pollen tube attraction activity[3, 5, 6]. Compared to NPA1, NPA2 does not have attraction activity and shows much lower expression level. AtNPA1 shows attraction activity to pollen tubes of Arabidopsis and its close relatives A. lyrata and C. rubella, which may contribute to possible interspecific gene flow and the consequent speciation of new species. It is of note that AtNPA1 shows preferential attraction of the conspecific pollen tubes than the pollen tubes of sister species and no attraction activity to the E. salsugineum, suggesting that AtNPA1 may also contribute to the reproductive isolation between Arabidopsis and the distantly related species. NPA1 is supposed to function redundantly with the other pollen tube attractants, like LUREs, XIUQIUs and TICKETs in the CRP_810 group. This conjecture is based on two facts that CRP_810 members have functional redundancy in enticing pollen tubes to the micropyle[3], and NPA1 also function in this process. The presence of a large amounts of attractant-encoding genes ensures reproductive success in a plant species by rendering precise and timely pollen tube targeting into the ovule and reduces the risk of fertilization by alien pollen. The diversity of these attractants raises an intriguing question of how the different types of attractant peptides were evolved to regulate a single process. Peptides evolve rapidly compared to other genes via duplications and mutations emerging during evolution, leading to their abundance among species. Most duplicated peptides including CRPs arised from whole genome duplication or segmental duplication with 22%−39% of CRPs in most species exhibit clustering patterns with frequent tandem duplications[28−30]. The aforementioned evolutionary pattern results in the repeats of peptide genes in plants, thereby serving as an intrinsic driving force behind the abundance of pollen tube attractants. And the abundance of these peptides could alleviate the selection constraint to allow accumulations of mutations that contribute to species-specificity and gene loss. The function of the homologs of NPAs encoded in Leguminosae species, such as Trifolium pretense, Phaseolus acutifolius, Medicago truncatula, and Vigna unguiculata still need further experimentation. It can be expected that other types of signals would be identified in plants in the future and would further help to explain the signal diversity and species isolation.
-
The authors confirm contribution to the paper as follows: data analysis and interpretation of results: Wang WQ, Meng JG, Yang F, Xu YJ; study conception and design, Supervision, draft manuscript preparation: Li HJ; manuscript revision: Li SZ. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated and analyzed during this study are available from the corresponding author on reasonable request.
The work was supported by the National Key Research and Development Program of China (2022YFF1003500), National Natural Science Foundation (32170343), and CAS Project for Young Scientists in Basic Research (No.YSBR-078).
-
The authors declare no competing interest. Li HJ is the Editorial Board member of Seed Biology who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
-
# Authors contributed equally: Wei-Qi Wang, Jiang-Guo Meng
- Supplemental Table S1 Primer sequence.
- Supplemental Fig. S1 Protein sequence alignment of NPA1 and NPA2 and their homologs in Brassicaceae species.
- Supplemental Fig. S2 Genotype of the npa1 and npa2 mutants generated by CRISPR/Cas9 technology.
- Supplemental Fig. S3 Pollen tube growth of npa1 npa2 mutant.
- Supplemental Fig. S4 Pollen tube attraction efficiency of npa1 npa2 mutant.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang WQ, Meng JG, Yang F, Xu YJ, Li SZ, et al. 2024. A non-defensin peptide NPA1 attracts pollen tube in Arabidopsis. Seed Biology 3: e003 doi: 10.48130/seedbio-0024-0001 

A non-defensin peptide NPA1 attracts pollen tube in Arabidopsis
- Received: 10 October 2023
- Revised: 20 December 0202
- Accepted: 03 January 2024
- Published online: 21 February 2024
Abstract: In angiosperms, female gametophyte secretes a range of attractants to entice the pollen tube for fertilization. In dicots, all the identified attractants are defensin-like cysteine-rich peptides (CRPs) family members, while monocots, like Zea mays in Gramineae utilize non-CRP-type Egg Apparatus 1-like peptides as pollen tube attractants. However, whether dicots have non-CRP attractants is still unclear. Here we characterize a non-defensin peptide attractant NON-DEFENSIN PEPTIDE ATTRACTANT 1 (NPA1) in Arabidopsis. NPA1 is transcriptionally regulated by MYB98 in synergids. Besides the conspecific pollen tube, AtNPA1 is also capable to attract pollen tubes of sister species A. lyrata and C. rubella, but not E. salsugineum. Furthermore, when NPA1 is introduced to complement myb98, it restores pollen tube attraction and fertility to a level comparable to the complementation with LUREs. Together, this study identifies a new type of peptide attractant in dicots and highlights the diversity of the attraction cues and signaling pathways.
-
Key words:
- Non-defensin /
- Peptide /
- Pollen tube attraction