










-
Brassica napus is the second-largest oil crop in the world. Rapeseed oil is not only an important edible resource, but also an industrial resource for the production of biofuel and lubricants[1]. With the demand for plant oil on the rise, increasing oil production and quality in B. napus through genetic and molecular approaches possesses important theoretical significance and practicable value[1].
The vegetable oil biosynthesis process involves multiple steps catalyzed by different enzymes in different organelles, including glycolysis that provides a carbon source for oil synthesis, de novo synthesis of fatty acid (FA) in plastid, FA modification, and triacylglycerol (TAG) assembly in endoplasmic reticulum[2]. In higher plants, oil biosynthesis is physiologically coupled with seed development. Previous studies revealed that seed development with oil synthesis is mainly regulated by the LAFL network (including LEC1, L1L, ABI3, FUS3 and LEC2) and some NF-Y transcription factors[3,4]. The NF-Y transcription factors comprise three subfamilies: NF-YA, NF-YB, and NF-YC, and perform functions involved in multiple processes, including assembly of heterodimer between the NF-YB and NF-YC in cytoplasm, the translocation of the heterodimer to the nucleus, formation of NF-YA/NF-YB/NF-YC heterotrimer that positively or negatively regulates the expressions of the downstream genes[4]. On the other hand, LEC1 has been reported to be involved in seed development and nutrient storage. Arabidopsis lec1 mutant displays the phenotype of the reduced protein and lipid bodies in seeds. Consistent with this phenotype, the transcription of the genes encoding oil body proteins, storage proteins and related transcriptional regulators (such as WRI1, etc.) is hindered in the lec1 mutant[3,5]. Overexpression of BnLEC1 and BnL1L increased the expressions of a subset of genes involved in sugar metabolism, glycolysis, and FA biosynthesis, leading to a significant increase in seed oil content of Brassica napus[6,7]. In Arabidopsis and soybean seeds, the target genes of LEC1 are indentified by ChIP and transcriptome analyses[8]. Furthermore, it has been indicated that a gibberellin signal repressor (AtDELLA) can interact with AtLEC1 to inhibit its transcriptional activity during late embryogenesis[9]. The activation of SUCROSE SYNTHASE 2 and CRUCIFERIN C promoters by LEC1/L1L-[NF-YC2] heterodimer depends on abscisic acid (ABA) and a bZIP factor (bZIP67) that bind to an ABA-response element (ABRE)[10].
ABA plays widespread roles in multiple biological processes in the plant life cycle[11−13]. In higher plants, the biosynthetic process of ABA takes place in five steps and involves two organelles. In plastids, ABA1, ABA4, and NCED are responsible for the formation of xanthoxin which is derived from zeaxanthin. The subsequent biosynthetic reactions catalyzed by ABA2 and ABA3 are localized in the cytoplasm[14]. The central module of ABA signaling contains three core components, ABA receptor pyrabactin resistance1 (PYR1)/pyrabactin resistance1-like (PYL)/regulatory components of ABA receptor (RCAR), type 2C protein phosphatases (PP2Cs), and SNF1-related protein kinase 2 (SnRK2)[15,16]. ABA is captured by PYR/PYL/RCAR when the levels of endogenous ABA are increased in plant cells. The binding of PYR1/PYL/PCAR with ABA results in the conformational changing of PYR1/PYL/PCAR[16]. Upon conformational change, PYR1/PYL/PCAR can interact with and inhibit the activity of PP2Cs which interact with and inhibit SnRK2s in the absence of ABA[17]. Upon inhibition of PP2C, the SnRK2s are released and activated. Then, the ABA signals are transduced to SnRK2s' substrates by phosphorylation[18]. The SnRK2-type protein kinase family contains 10 members divided into three subclasses I, II, and III in Arabidopsis (SRK2A–J or SnRK2.1–2.10) and rice (SAPK1–10)[11,15,19]. Subclass III SnRK2 consists of SnRK2E/SnRK2.6, SRK2D/SnRK2.2, and SRK2I/SnRK2.3, which are redundant and strongly activated by ABA and play a pivotal role in ABA signaling[11]. Arabidopsis snrk2.2/2.3/2.6 triple mutant shows extreme ABA-insensitivity during seed germination, seedling growth, and stomatal aperture compared with wild type[11,20]. In addition, SnRK2.2, SnRK2.3 and SnRK2.6 are essential for the control of seed development and dormancy[20]. SnRK2s are also involved in leaf growth via effects on the tricarboxylic acid (TCA) cycle and sugar metabolism[13]. Several SnRK2 substrates have been proposed to be ABA response element (ABRE) binding factors (ABFs/AREBs), a group of basic leucine zipper-type (bZIP) transcription factors that regulate expressions of many ABA-responsive genes[21]. ABA-responsive kinase substrates (AKS) are involved in stomatal opening in response to blue light[22], and S-type slow anion channel (SLAC1) responsible for anion transport during ABA-responsive stomatal closure is phosphorylated by SnRK2[23,24]. Besides, substrate proteins of SnRK2s are identified by comparative quantitative phosphoproteomics analyses in Arabidopsis wild type and snrk2.2/2.3/2.6 triple mutant seedlings in response to ABA treatment or dehydration stress[25,26]. [R/K-x-x-pS/pT] is a known motif phosphorylated by SnRK2 in AREB/ABF[27,28]. Motif analysis of SnRK2 substrate proteins shows that [-(pS/pT)-x-x-x-x-(D/E)-] is potential phosphor motifs of SnRK2 targets[25,26].
The SnRK2 signaling is involved in redistributing sugars from leaf starch degradation under osmotic stress and modulating metabolism by fine-tuning carbon flux through the tricarboxylic acid cycle[29]. However, little is known whether SnRK2s participate in modulating oil synthesis in detail to date. In this study, it was found that the BnSNRK2;2-BnLEC1-[BnNF-YC1/4/9A/9B] regulatory network responding to ABA and sugar signals plays a vital role in oil biosynthesis in Brassica napus. Overexpression of BnSNRK2;2 and BnLEC1 enhanced oil content and level of very long chain unsaturated fatty acids (VLCUFAs), but decreased the protein level in seeds. Further study revealed that BnSNRK2;2 phosphorylates BnLEC1 in an ABA-dependant manner. The phosphorylation of BnLEC1 is essential for its interaction with BnNF-YCs for modulating lipid biosynthesis and accumulation in seeds of Brassica napus.
-
Two Brassica napus varieties, HFA (with high oil content, approximately 43.87% oil content in seeds) and LFA (with low oil content approximately 31.74% oil content in seeds) were kindly provided by Prof. Xiao-Ming Wu in Oil Crops Research Institute, Chinese Academy of Agricultural Sciences[30]. HFA and LFA are selected from the filial generations of Brassica napus cultivars G166 and ZheShuang6 (both are typical German semi-winter rape cultivars). All transgenic plants used in this study are in Brassica napus cv. Westar background. Seeds of HFA, LFA and Westar and related transgenic plants are sown usually at a density of 45 cm × 30 cm per plant in late September, transplanted in early November and harvested in early May in an experimental field at the campus of Central China Normal University, Wuhan, China.
Construction of vectors and transformation of B. napus
-
BnNapinA promoter (BnNapinAp) and Nos terminator were amplified by PCR from Brassica napus leaf genomic DNA and pCAMBIA-2300-35S-N-eGFP-OCS vector, respectively, and were cloned into pCAMBIAI1-1300 vector to generate PC1300-BnNapinApro-Nos fusion vector. Coding sequences of BnLEC1, BnSnRK2;2, BnNF-YC1, BnNF-YC4, BnNF-YC9A and BnNF-YC9B were amplified by PCR from 25 DAP seed cDNAs of Brassica napus using primer pairs listed in Supplemental Table S6. BnLEC1 mutants (BnLEC1S17A, BnLEC1S17D, BnLEC1S153A, BnLEC1S153D, BnLEC1S158A and BnLEC1S158D) and BnSnRK2;2 mutants (BnSnRK2;2T177A and BnSnRK2;2T177D) were amplified by PCR using primers pairs listed in Supplemental Table S6. All above coding sequences were inserted into pET32a and pMAL vectors for expressing recombinant proteins in Escherichia coli BL21, PC1300-BnNapinApro-Nos vector for generation of transgenic plants, PC2300-NGFP and PC2300-NFlag for subcellular localization and CoIP analysis, and pSAT1-nEYFP-C1 and pSAT1-cEYFP-C1-B for BiFC analysis, respectively. To construct the CRISPR/Cas9 plasmid of BnSnRK2;2/BnSnRK2;2C, sgRNA target sequence was designed using the CCTop-CRISPR/Cas9 target online predictor (
https://cctop.cos.uni-heidelberg.de:8043/ ). The sgRNA was cloned into BGK01 vector (Biogle, China;www.biogle.cn ). To construct the BnABA2 RNA interference (RNAi) vector, a 300-bp specific fragment of the BnABA2 gene sequence was cloned into the pKANNIBAL vector with a PDK intron to generate an inverted repeat, and then cloned into the PC1300-BnNapinApro-Nos vector. Transformation of Brassica napus (cv. Westar) was carried out as previously described[31]. The overexpression transgenic plants were identified by PCR using the forward primer of BnNapinAp and the reverse primers of BnLEC1 and BnSnRK2;2, respectively. Homozygous transgenic lines of T2 and T3 generations were used for further experiments. For mutant identification, the PCR was performed using the specific primers (Supplemental Table S6), and the products were sequenced.Analysis of oil content, protein content, fatty acid (FA) composition and abscisic acid (ABA) content in seeds
-
Plants of the transgenic Brassica napus lines (T2 and T3 generations) and wild type (Westar) grown in the field were used for the experiments (n > 10 plants per line). The Soxhlet extraction method was used for measuring the seed oil content as previously described[6]. The total protein was measured according to a published method[32]. FA composition analysis was performed by GC-MS as previously described[6]. The seeds of HFA and LFA lines at the different developmental stages were collected and used for the quantification of endogenous abscisic acid (ABA) by the Wuhan ProNets Biotechnology Company (Wuhan, China) as previously described[31]. Triplicates were performed for each experiment.
Extraction of total protein, in-solution protease digestion and dimethyl labeling peptides
-
Total protein was extracted with urea extraction buffer (UEB) and digested by trypsin according to a previously published method[32] with some modifications. Briefly, 2 g frozen sample powder was thawed in 20 ml UEB buffer and then centrifuged at 110,000 g for 2 h at 10 °C. After a precipitating of the mixed supernatant with 3 volumes of acetone at ‒20 °C overnight, the protein pellet was collected by centrifugation (10,000 g) at 10 °C for 20 min. Then the protein was air-dried under a vacuum for digestion by trypsin in NH4HCO3 solution.
Peptides were labeled with dimethyl and dimethyl:2H(6) using the PhosQuant kit (N-Cell Technology Company, Hong Kong, China) according to the instructions. Dimethyl-labeled peptides from HFA seeds and dimethyl:2H(6)-labeled peptides from LFA seeds of Brassica napus were mixed to form the Forward replicate, and dimethyl:2H(6)-labeled HFA and dimethyl-labeled LFA peptides were mixed to form the Reciprocal replicate. Peptides were desalted with a C18 column for phosphopeptide enrichment analysis.
Quantitative phosphoproteomic analysis
-
The phosphopeptide enrichment was performed multiple times with TiO2 beads using integrated multi-step as described previously[32]. The purity of the enriched phosphopeptides reached to over 85%. The phosphopeptides were fractionated into 10 fractions on the Poly WAX LPTM Column (Item 104WX0503) via HPLC, and LC-MS/MS and quantitative phosphoproteomic analyses were performed by N-Cell technology Company (Hong Kong, China).
The nano-ultra performance liquid chromatography system (Easy nLC, Thermo Fisher Scientific) and Q-Exactive hybrid quadrupole-Orbitrap MS (Thermo Fisher Scientific) were used for analyzing the phosphopeptides, which were operated with the following parameters: positive ion mod, MS survey-scan range of 300 and 2,000 m/z, the resolution was 70,000 with an AGC target value of 1e6 ions and a maximum ion injection time of 60 ms, the resolution of dd-MS2 was 17,500 with an AGC target value of 1e5 ions and a maximum ion injection time of 100 ms. The stepped NCE was 27 with a 10 TopN.
The MS2 spectra converted to the mgf files were searched against Brassica napus Genome Resources (
www.genoscope.cns.fr/brassicanapus/ ) using the Mascot search engine (Version 2.3, Matrix Science). The PSM of phosphopeptides was selected by the false discovery rate (FDR) which was set to 1%. The reliable phosphosite was identified according to the Mascot delta score (www.matrixscience.com ), the threshold value of which was set to 10. Those phosphopeptides that presented in both HFA and LFA were selected for quantitation. The software Silique-N (Alpharomics Co., Limited, Shenzhen, China,www.alpharomics.com/Home/Product_msda ) was used for quantifying the ion chromatograms (MS1 ion intensities) from both dimethyl and dimethyl:2H(6)-labeled envelopes of all reliable phosphopeptides. The two-tailed Student t-test followed by B.-H multiple hypothesis testing correction (FDRB.-H. ≤ 5%) was employed in determining the statistical significance of the ratios.Assay of protein-protein interaction by pull-down, BiFC and Co-IP
-
For pull-down assay, 3 μg His-BnLEC1 fusion proteins were incubated with Amylose Resin (E8021L, NEB) containing MBP and MBP-BnSnRK2;2 at 4 °C for 3 h. Then proteins combined with the Amylose Resin were eluted and collected, and subsequently detected through Western blotting analysis with anti-His (2366S, CST) and anti-MBP (2396S, CST) antibodies.
For the bimolecular fluorescence complementation (BiFC) assay, pSAT1-nEYFP-C1, pSAT1-cEYFP-C1-B, and related fusion vectors containing coding sequences of BnSnRK2;2, BnNFYC1, BnNFYC4, BnNFYC9A, BnNFYC9B, and BnLEC1 and their variants were transferred into Agrobacteria tumefaciens strain GV3101, respectively. The transformed agrobacteria were injected into young leaves of tobacco (N. benthamiana) according to the corresponding combinations. After three days of growth under long-day conditions, the YFP fluorescence and DAPI (4',6-diamidino-2-phenylindole) staining in leaf lower epidermal cells was observed under the Leica SP5 confocal laser scanning microscope. DAPI staining was used to show the cell nucleus, and YFP fluorescence was excited at 488 nm and detected between 493 and 542 nm.
For Co-immunoprecipitation (Co-IP) assay, Flag-BnSnRK2;2, Flag-BnNYFC1, Flag-BnNYFC1, Flag-BnNYFC4, Flag-BnNYFC9A or Flag-BnNYFC9B with GFP, GFP-BnLEC1, GFP-BnLEC1S17A, GFP-BnLEC1S17D, GFP-BnLEC1S153A, GFP-BnLEC1S153D, GFP-BnLEC1S157A or GFP-BnLEC1S157D were coexpressed in 25 DAP seeds of Brassica napus and leaves of tobacco (Nicotiana benthamiana), respectively. The samples were ground with liquid nitrogen and resuspended in IP buffer (100 mM MOPS, pH7.6, 150 mM NaCl, 20 mM iodoacetamide, 1% TritonX-100, 0.1% NonidetP-40, 1 mM PMSF, 1 µg/L pepstin, 2 µg/L aprotinin, and 1 µg/L leupeptin). After centrifugation at 20,000 g for 30 min, supernatant were incubated with anti-BnSnRK2;2, anti-BnLEC1 or anti-GFP antibody and Protein A Agarose Beads (9863, CST) at 4 °C overnight. Then proteins combined with the beads were eluted six times with wash buffer (50 mM Tris-HCl, pH7.5, 150 mM NaCl, 0.2% TritonX-100, and 1 mM PMSF), and subsequently detected through Western blotting analysis with anti-Flag (Sigma–Aldrich) and anti-GFP antibodies (Sigma–Aldrich) or used for MS assay[32].
Polyclonal antibody preparation and immunoblotting assay
-
The oligopeptides of IPQNLGSpFKEESSNLSDLSDSEK, ASSpKDLDSQFSSLSLNSK, ASSKDLDSpQFSSLSLNSK, EGIESpDEEIR, LGTVSSpPETITAVR, ERSEDRMETSSpAEAGGGGSGIR, STpVGTPAYIAPEILLR, STVGTPAYIAPEILLR, SNSpGLNLDQHNNSIPTMTGSIGACDDKNK and SNSGLNLDQHNNSIPTMTGSIGACDDKNK were made and used for producing rabbit polyclonal antibodies by GL Biochem Ltd. (Shanghai, China). The immunoblotting analysis was performed using previously published methods[32,33]. The anti R-x-x-(pS/pT) (#9621) antibody which detects peptides and proteins containing a phospho-serine/threonine residue with arginine at the −3 position was purchased from Cell Signalling Technology, Inc (Danvers, MA, USA). Phosphate affinity SDS-PAGE was performed using Mn2+-phos-tag gel (193-16711, Wako, Japan).
-
Total RNA was isolated from Brassica napus seeds at different developmental stages, and RT-qPCR analysis was performed as described previously[30]. Relative expression levels were normalized against the expression level of the housekeeping gene BnActin2 (BnACT2). The related gene-specific primers were designed after blast (
www.genoscope.cns.fr/blat-server/cgi-bin/colza/webBlat )[34]. Sequences of homologous genes were aligned by the Clustal W. The primers used in RT-qPCR analysis are shown in Supplemental Table S6.ABA and sugar treatments
-
Fifty µM abscisic acid (ABA) (Sigma–Aldrich) and 100 mM sucrose, glucose, fructose, and mannitol were injected into pod cavities of Brassica napus (cv. Westar), respectively, and sucked out half an hour later with an injector from 20 DAP to 35 DAP every 2 d, using water as control (CK). Then oil content and fatty acid composition in mature seeds were assayed. To detect the influence of ABA and sugar on the phosphorylation of BnLEC1 and BnSnRK2;2 and expressions of the related genes, 50 µM abscisic acid (ABA) and 100 mM sucrose, glucose, fructose, and mannitol were injected into pod cavities of Brassica napus (cv. Westar), respectively, at different time points before 25 DAP, and then RNA and protein were isolated from the 25 DAP seeds for the related experiments.
In vitro kinase assay
-
Reaction solution of in vitro kinase assay consists of 500 nM kinase (BnSnRK2;2 or/and its variants), 250 nM substrates (BnLEC1 or/and its variants), 25 mM Tris HCl, pH 7.5, 1 mM EGTA, 10 mM MgCl2, and 2 mM ATP . After incubation at 30 °C for different times, the reactions were stopped by the addition of equal volume 2 × SDS loading buffer for immunoblotting assay and MS assay[32].
Cell-free degradation assay
-
BnSnRK2;2 was incubated with NGFP-BnLEC1, NGFP-BnLEC1S17A, NGFP-BnLEC1S17D, NGFP-BnLEC1S153A, NGFP-BnLEC1S153D, NGFP-BnLEC1S157A and NGFP-BnLEC1S157D, respectively, in in vitro kinase reactions. Different versions of phosphorylated BnLEC1 were collected by immunoprecipitation as described by Zhai et al.[35], and desalted in 50 mM Tris-HCl buffer, pH 7.5. Crude proteins from 3-week-old leaves were extracted in degradation buffer (50 mM Tris-HCl, pH 7.5, 10 mM NaCl, 10 mM MgCl2, 4 mM PMSF, 5 mM DTT, and 10 mM ATP which is supplemented at the start of the reaction). Crude proteins and all above BnLEC1-related proteins were quantified by the Bio-Rad protein assay[32]. Each reaction contains 1 µg different versions of BnLEC1 and 1 mg crude proteins in 200 µl extracts. After incubation at 28 °C, the reactions were stopped at different times by the addition of equal volume 2 × SDS loading buffer for immunoblotting assay.
Accession numbers
-
The accession numbers of the genes in this study are the following: BnaA03g40850D (BnSnRK2;2), BnaA07g10770D (BnLEC1), BnaC08g20340D (BnNF-YC1), LOC106347913 (BnNF-YC4), BnaA09g48620D (BnNF-YC9A), BnaC05g06520D (BnNF-YC9B), BnaC03g69290D (BnABA2).
-
Our previous study suggested that 25 d after pollination (DAP) was the key stage for seed oil accumulation in two Brassica napus lines (HFA with high oil content and LFA with low oil content)[30]. The experimental results further indicated that oil content in seeds of both HFA and LFA is dramatically increased after 20 DAP and the oil synthesis rate reached a peak value at approximately 25 DAP (Supplemental Fig. S1). To identify the phosphorylation signaling components responsible for oil accumulation, a liquid chromatography-tandem mass spectrometry (LC-MS/MS) approach was applied to compare phosphopeptides isolated from 25 DAP seeds of HFA and LFA (Supplemental Fig. S2). By quantitative phosphoproteomic analysis, 2908 non-redundant phosphopeptides including 3,123 phosphorylation sites −91.53% pSer, 6.84% pThr, and 1.63% pTyr-derived from 1,743 phosphoprotein groups were identified in the seeds (Fig. 1a, b; Supplemental Dataset S1). Among them, 76 phosphopeptides were significantly up-regulated and 143 phosphopeptides were significantly down-regulated in seeds of HFA compared with LFA (Fig. 1c, Supplemental Dataset S2). Furthermore, polyclonal antibodies against IPQNLGSpFKEESSNLSDLSDSEK (anti:BnaC06g24230DS101), ASSpKDLDSQFSSLSLNSK (anti:BnaC09g15590DY102), ASSKDLDSpQFSSLSLNSK (anti:BnaC09g15590DS342), EGIESpDEEIR (anti:BnaCnng20200DS31), LGTVSSpPETITAVR (anti:BnaC08g20440DS248), ERSEDRMETSSpAEAGGGGSGIR (anti:BnaC03g49000DS191), STpVGTPAYIAPEILLR (anti:pBnSnRK2;2), STVGTPAYIAPEILLR (anti:BnSnRK2;2), SNSpGLNLDQHNNSIPTMTGSIGACDDKNK (anti:pBnLEC1) and SNSGLNLDQHNNSIPTMTGSIGACDDKNK (anti:BnLEC1) were generated and verified by immuno-dot blotting analysis for validating the quantitative phosphoproteomic data (Supplemental Fig. S3). Furthermore, the phosphorylation levels of BnSnRK2;2, BnLEC1, BnaC06g24230DS101 and BnaC09g15590DS342 were higher, while the phosphorylation levels of BnaC09g15590DY102, BnaCnng20200DS31, BnaC08g20440DS248 and BnaC03g49000DS191 were lower in 25 DAP seeds of HFA compared with those in LFA, consistent with the phosphoproteomic analysis (Fig. 1e).
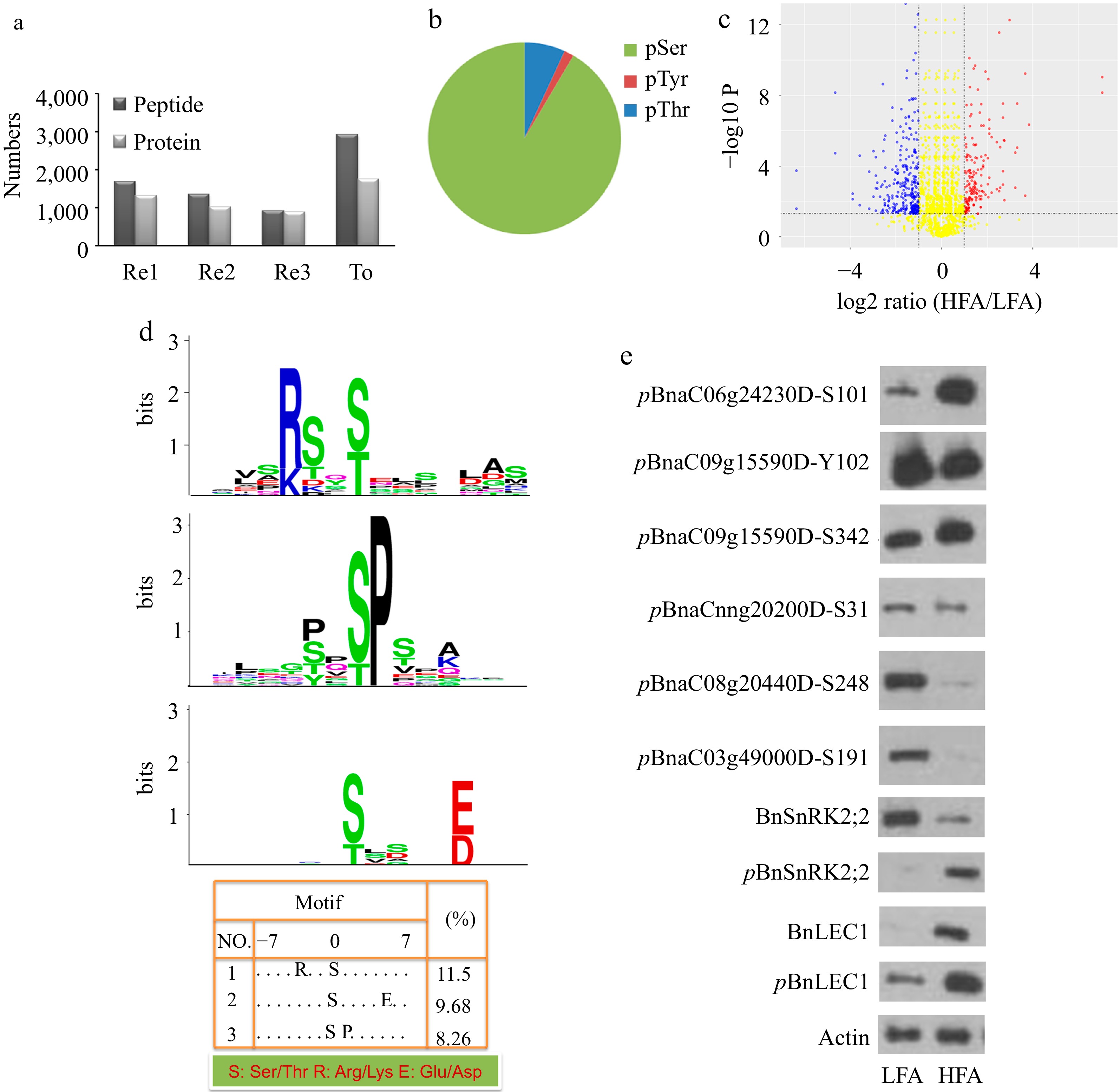
Figure 1.
Quantitative phosphoproteomic identification of phosphoproteins related oil biosynthesis and accumulation in seeds of B. napus. (a) The number of unique phosphopeptides and phosphoproteins identified from each experimental replicate (Re1, Re2, Re3) and combination of all replicates (To). Each experimental replicate comprises the mixture of dimethyl- and dimethyl:2H(6)-labeled peptides. (b) Distribution of phosphorylation sites by amino acids. (c) Volcano plots of quantitative phosphopeptide analysis. The red and blue circles represent significantly increased and decreased phosphopeptides (HFA vs. LFA), respectively. The log2 ratio represents the average binary logarithmic ratio of MS1 dimethyl-labeled areas of a phosphopeptide between HFA and LFA, P is the P value determined using Student's t test followed by Benjamini–Hochberg multiple hypothesis test correction. The horizontal and vertical dashed line indicates the threshold for the significance (FDRB.–H. ≤ 5%) and fold change (log2 ratio ≤ −1 or > 1), respectively. (d) Phosphorylation motif analysis of upregulated phosphopeptides. Phosphorylation motifs were searched by Motif-X and depicted using LOGO by MEME. The numbers and detailed sequences of phosphopeptides in each LOGO are shown in bottom right table and Supplemental Table S1− S3, respectively. (e) Immunoblotting analysis for validating the quantitative phosphoproteomic results. Protein immunoblotting analysis of protein phosphorylation in 25 DAP seeds of HFA and LFA was performed using polyclonal antibodies against anti-pBnSnRK2;2, anti-BnSnRK2;2, anti-pBnLEC1, anti-BnLEC1, anti-pBnaC06g24230DS101, anti-pBnaC09g15590DY102, anti-pBnaC09g15590DS342, anti-pBnaCnng20200DS31, anti-pBnaC08g20440DS248, and anti-pBnaC03g49000DS191. Anti-actin was used as the internal reference. HFA, a B. napus variety with high seed oil content; LFA, a B. napus variety with low seed oil content; DAP, day after pollination.
Additionally, we extended the phosphorylation motif analysis of the identified phosphoproteins using the Motif-X algorithm to gain insights into the potential kinases participating in oil accumulation. As shown in Fig 1d and Supplemental Table S1, 12 upregulated phosphopeptides contain the motif [-(K/R)-x-x-(pS/pT)-], recognized by SnRK2. Besides, 14 and 9 of the phosphorylation sites belong to [-(pS/pT)-x-x-x-x-(D/E)-] and [-(pS/pT)-P-] motifs, respectively, that are enriched in ABA- or dehydration-responsive phosphopeptides (Fig. 1d; Supplemental Table S2 & S3).
Phosphorylation of BnSnRK2;2 is essential for its function in regulating oil accumulation in seeds
-
SnRK2s is the core component in ABA signaling and play important roles in plants[11]. To determine the function of BnSnRK2;2 in oil accumulation, we generated transgenic Brassica napus plants overexpressing BnSnRK2;2 driven by the seed-specific BnNapinA promoter (Supplemental Fig. S4a), and the double mutant of Bnsnrk2;2/Bnsnrk2;2c was acquired using the CRISPR/Cas9 system (Supplemental Fig. S5). Although the mutant of Bnsnrk2;2/ Bnsnrk2;2c had no effect on the oil and protein levels (Supplemental Fig. S5), which may be caused by functional redundancy of the SnRK2 members, the oil level was increased, but the protein content decreased in the seeds of the BnSnRK2;2 overexpression (OE) transgenic plants compared with those in wild type (WT) (Fig 2a, b; Supplemental Tables S4, S5). Furthermore, fatty acid composition analysis showed that the level of very long-chain unsaturated fatty acids (VLCUFAs) in the BnSnRK2;2 OE transgenic seeds were much higher than that in WT (Fig. 2c). Especially, the ratio of eicosenic acid (C20:1) and erucic acid (C22:1) in fatty acids (FAs) in OE seeds was remarkably higher than that in WT, and the levels of eicosadienoic acid (20:2), behenic acid (C22:0), docosadienoic acid (C22:2) and nervonic acid (C24:1) was also increased in the OE seeds, relative to WT. In contrast, the levels of most medium-long-chain fatty acids, especially oleic acid (C18:1), were reduced in the OE transgenic lines, compared with WT (Fig. 2c). On the other hand, seed-specific overexpression of BnSnRK2;2 has no effect on growth and development (including the major agronomic traits) of the transgenic plants (Supplemental Fig. S4b).
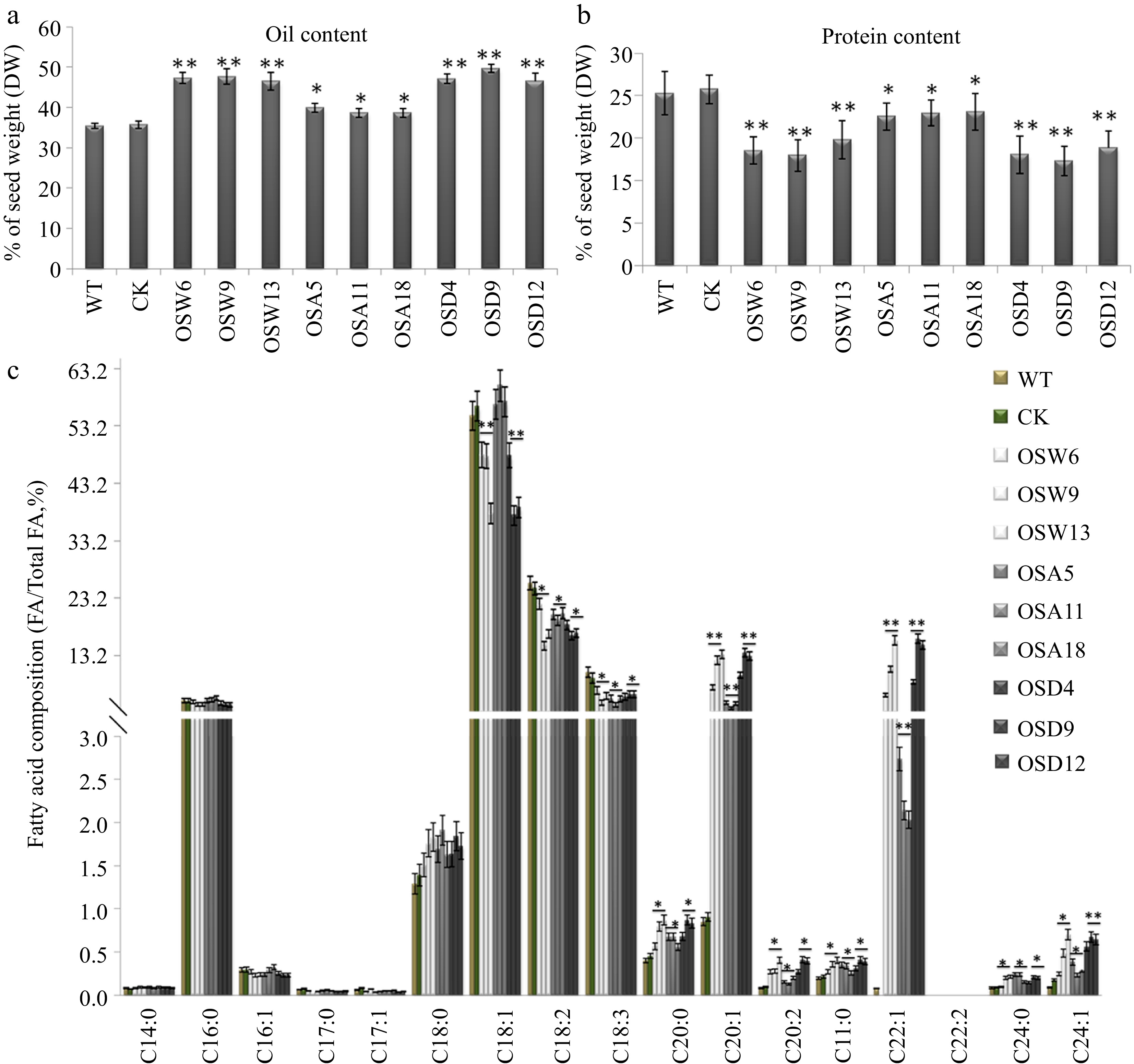
Figure 2.
Assay of oil and protein contents and fatty acid composition in seeds of the BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177D overexpression transgenic B. napus. Assay of (a) oil content and (b) protein content in seeds of the BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177D transgenic lines and controls (WT and CK). The seeds of 10 plants in each line (T3 generation) were used for analysis. Means ± SE are shown (Fisher's Least Significant Difference (LSD) test, * p < 0.05, ** p < 0.01). (c) Assay of fatty acid composition in seeds of the BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177D transgenic lines (T3 generation) and controls. Three biological replicates were performed, and means ± SE are shown (Student's t test, * p < 0.05, ** p < 0.01). WT, wild type; CK, the null transgenic lines; OSW6/9/13, BnSnRK2;2 transgenic lines; OSA5/11/18, BnSnRK2;2T177A transgenic lines; OSD4/9/12, BnSnRK2;2T177D transgenic lines; DAP, day after pollination.
To evaluate whether phosphorylation of BnSnRK2;2 at Thr177 has an effect on its role in oil synthesis, the transgenic plants harboring BnSnRK2;2T177A which removes the phosphorylatable amino acid, and BnSnRK2;2T177D which mimicked the phosphorylation were generated, respectively (Supplemental Fig. S4a). The experimental results revealed that oil and protein contents and FA composition in seeds of the BnSnRK2;2T177D transgenic plants were similar to those in the BnSnRK2;2 transgenic plants. Although the oil content and VLCFAs ratio have slightly increased in seeds of the BnSnRK2;2T177A transgenic plants compared with WT, however, it is far behind the BnSnRK2;2T177D transgenic seeds (Fig. 2). The above results indicated that Thr177 in BnSnRK2;2 protein is the key phosphorylation site, and the phosphorylation of BnSnRK2;2 has a great influence on its function in regulating oil synthesis and accumulation in seeds of Brassica napus.
BnSnRK2;2 specifically phosphorylates BnLEC1 in seeds
-
To understand the role of BnSnRK2s in oil accumulation, coimmunoprecipitation (Co-IP) assay was performed to identify the proteins interacting with BnSnRK2;2. Clarified cell lysates from 25 DAP seeds of Brassica napus were incubated with polyclonal antibodies against BnSnRK2;2, and the potential proteins (including BnLEC1) interacting with BnSnRK2;2 were identified by LC-MS/MS analysis (Fig. 3a; Supplemental Dataset S3). To verify the interaction between BnSnRK2,2 and BnLEC1, Co-IP assay was performed. Flag-BnSnRK2,2 and GFP-BnLEC1 proteins were coexpressed in the leaves of tobacco (N. benthamiana). As shown in Fig 3b, BnLEC1 protein could coprecipitate with BnSnRK2;2 protein, demonstrating BnSnRK2;2 interact with BnLEC1 in vivo. Also, pull-down assay revealed BnSnRK2;2 was able to interact with BnLEC1 in vitro (Fig. 3c). Furthermore, bimolecular fluorescence complementation (BiFC) assay showed that BnSnRK2;2 was associated with BnLEC1 strongly in the nucleus but weakly in the cytoplasm (Supplemental Fig. S6). Moreover, BiFC assay showed that BnLEC1 could also interact with BnSnRK2;3 and BnSnRK2;6 which have the highest sequence similarity with AtSnRK2;3 and AtSnRK2;6, respectively. Combined with the foregoing phosphoproteomic data and motif analysis that indicated BnLEC1 contains the motif [-(K/R)-x-x-(pS/pT)-] and its phosphorylation level was up-regulated in the HFA seeds (Fig. 1c, d), the above results implied that BnLEC1 may be the phosphorylation substrate of BnSnRK2; 2.
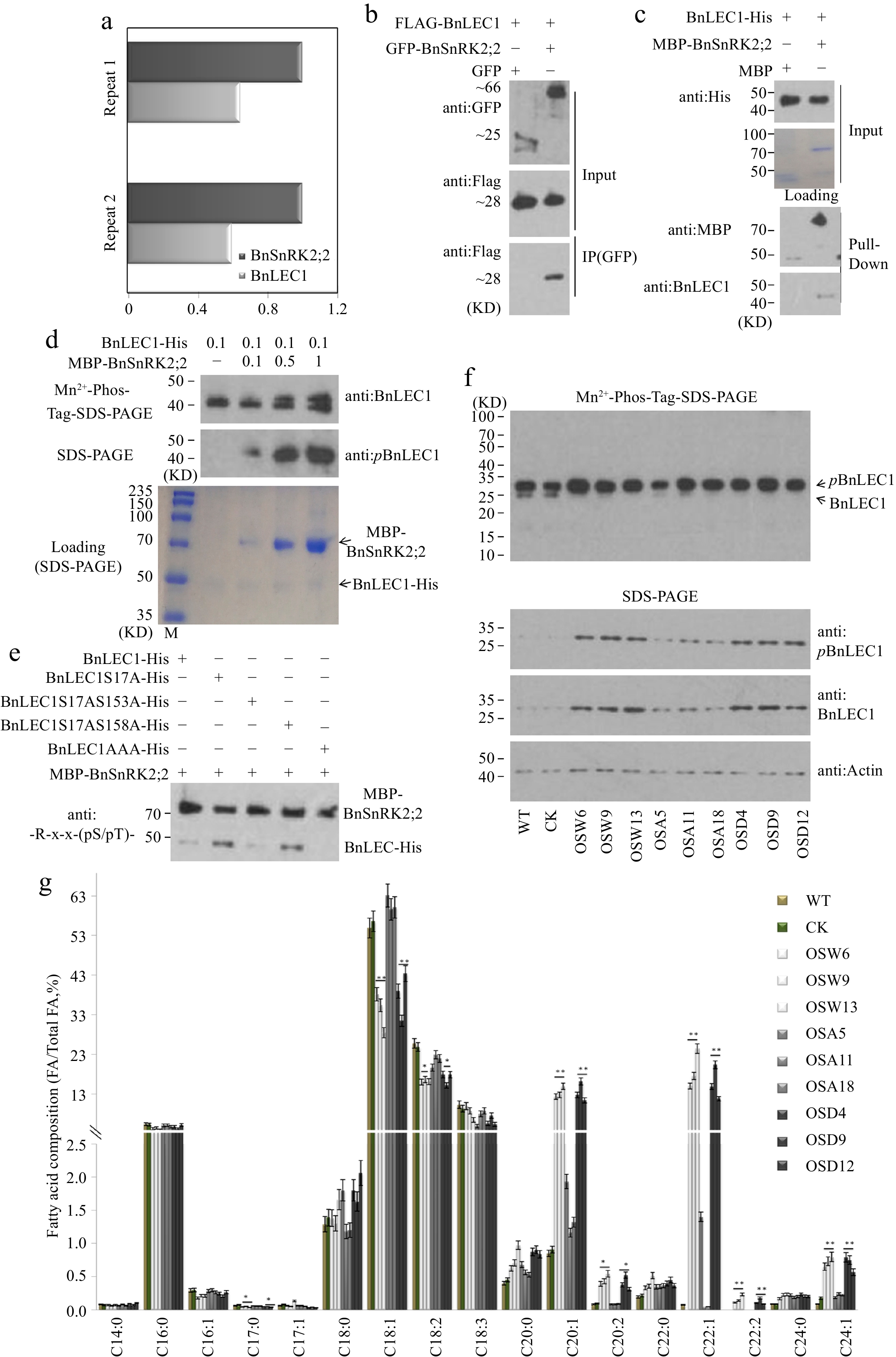
Figure 3.
Assay of BnSnRK2;2 mediated phosphorylation of BnLEC1 in seeds of B. napus. (a) Co-immunoprecipitation (Co-IP) assay of BnSnRK2;2 protein from 25 DAP seeds of B. napus. Proteins were precipitated from cell lysates using anti:BnSnRK2;2 antibody and identified by LC-MS/MS. Control precipitations were performed without antibody. Relative abundance represent the ratio of the number of peptide spectral matches derived from BnLEC1 and BnSnRK2;2. Two biological replicates were performed. (b) Co-IP assay of the interaction of BnSnRK2,2 with BnLEC1 in tobacco (N. benthamiana) leaves co-expressing Flag-BnLEC1 and GFP-BnSnRK2,2 (GFP as the negative control ). The anti-Flag and anti-GFP antibodies were used for immunoblotting analysis. (c) Pull-down assay of BnLEC1-His interacting with MBP-BnSnRK2;2. His-BnLEC1 proteins were incubated with MBP or MBP-BnSnRK2;2 proteins coupled with Amylose Resin. The anti-His and anti-MBP antibodies were used for immunoblotting analysis. (d) In vitro kinase assay was performed using BnLEC1-His and different amount of MBP-BnSnRK2;2 (µg). Immunoblotting analysis using anti:pBnLEC1 antibody was performed to detect the phosphorylation of Ser17 residue in BnLEC1. Proteins were separated by SDS-PAGE (down) or Mn2+-phos-tag gel electrophoresis (up). (e) In vitro kinase assay was performed using five versions of BnLEC1 and MBP-BnSnRK2;2. anti:-R-x-x-(pS/pT)- antibody was used for detecting the phosphorylation of BnLEC1. (f) Protein immunoblotting analysis of phosphorylation of BnLEC1 Ser17 residue in 25 DAP seeds of BnSnRK2;2 transgenic Brassica napus. Proteins were separated by SDS-PAGE (down) or Mn2+-phos-tag gel electrophoresis (up). (g) Assay of fatty acid composition in seeds of BnLEC1, BnLEC1S17A and BnLEC1S17D transgenic lines (T3 generation) and controls (WT and CK). Three biological replicates were performed, and means ± SE are shown (Student's t test, * p < 0.05, ** p < 0.01). WT, wild type; CK, the null transgenic lines; OSW6/9/13, BnSnRK2;2 transgenic lines; OSA5/11/18, BnSnRK2;2T177A transgenic lines; OSD4/9/12, BnSnRK2;2T177D transgenic lines; OLW3/9/14, BnLEC1 transgenic lines; OLA4/13/17, BnLEC1S17A transgenic lines; OLD2/9/15, BnLEC1S17D transgenic lines.
To determine whether BnSnRK2;2 specifically phosphorylates BnLEC1 in developing seeds of Brassica napus, in vitro phosphorylation assay was performed with the purified MBP-BnSnRK2;2 and His-BnLEC1 proteins. As shown in Fig. 3d, Ser17 residue of BnLEC1 protein could be phosphorylated in vitro by BnSnRK2;2. Based on polyclonal antibodies against -R-x-x-(pS/pT)-, furthermore, immunoblot analysis also showed that BnLEC1 could be phosphorylated in vitro by BnSnRK2;2 (Fig. 3e).
To confirm the phosphorylation of BnLEC1 by BnSnRK2;2 in developing plants of Brassica napus, the phosphorylation level of BnLEC1 was quantified in the BnSnRK2;2 transgenic plants and controls. Compared with the controls (WT and CK), the phosphorylation levels of BnLEC1 were remarkably enhanced in seeds of the BnSnRK2;2 and BnSnRK2;2T177D transgenic plants, but only slightly increased in the BnSnRK2;2T177A transgenic plants (Fig. 3f). Moreover, following separation by phosphate affinity SDS-PAGE with acrylamidependant phos-tag (Mn2+-phos-tag), which enables separating the phosphorylated polypeptides and non-phosphorylated polypeptides, the signals of slower-migrating bands representing the phosphorylated BnLEC1 were enhanced, while the signals of the native BnLEC1 were weakened in seeds of BnSnRK2;2 and BnSnRK2;2T177D transgenic plants compared with the controls (Fig. 3f). In vitro phosphorylation assay also showed that strong phosphorylation of BnLEC1 was observed in the presence of both BnSnRK2;2T177D and BnLEC1, but relatively weak signal of BnLEC1 phosphorylation was detected in the reactions containing BnSnRK2;2T177A and BnLEC1 (Supplemental Fig. S7b). Moreover, Thr177 residue is located in the activation domain of BnSnRK2;2 (Supplemental Fig. S7a). These results demonstrated that BnLEC1 is phosphorylated by BnSnRK2;2 in seeds of Brassica napus, and the kinase activity of BnSnRK2;2 partly relies on the phosphorylation of its Thr177. Additionally, the transgenic plants overexpressing BnLEC1 and its two mutated versions, in which the Ser17 was replaced by Ala or Asp (designated BnLEC1S17A and BnLEC1S17D), driven by seed-specific BnNapinA promoter were generated for measuring oil and protein contents and fatty acid composition (Supplemental Fig. S8a). As shown in Fig 3g, Supplemental Fig. S8b, and Supplemental Tables S4 & S5, both BnLEC1 and BnLEC1S17D transgenic plants had higher seed oil and VLCUFA contents than WT. Oppositely, there was no difference in seed oil and protein contents and fatty acid composition between BnLEC1S17A transgenic plants and WT. Moreover, seed-specific overexpression of BnLEC1, BnLEC1S17A and BnLEC1S17D has no effect on plant growth and development and the major agronomic traits of Brassica napus (Supplemental Fig. S8c). Taken together, the above data suggested that the phosphorylation of Ser17 residue in BnLEC1 by BnSnRK2;2 is crucial for BnLEC1 functioning in seed oil synthesis and accumulation.
Phosphorylation of BnLEC1 by BnSnRK2;2 is required for its interaction with BnNFYC1/4/9A/9B and its nuclear localization
-
To explore how phosphorylation of BnLEC1 affects its role in seeds, Co-IP/MS assay was performed to identify the proteins interacting with BnLEC1 in 25 DAP seeds using antibodies against BnLEC1. As shown in Fig 4a and Supplemental Dataset S4, BnSnRK2;2 and the NF-YC transcription factors (including BnNF-YC1, BnNF-YC4, BnNF-YC9A, and BnNF-YC9B) were coimmunoprecipitated with BnLEC1, which was also verified by pull-down assay in vitro (Fig. 4b).
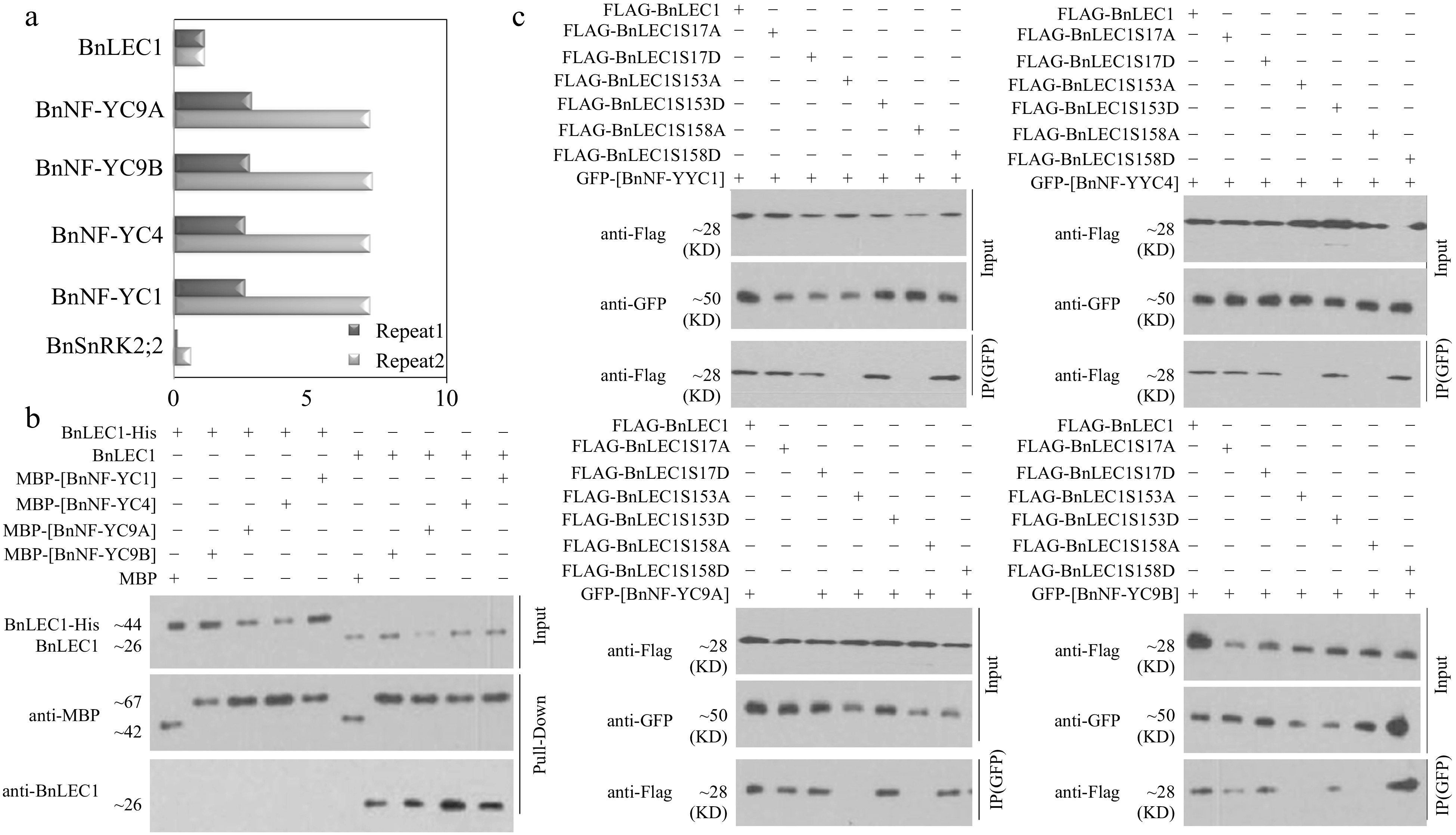
Figure 4.
Assay of BnLEC1 interacting with BnNF-YCs. (a) Co-immunoprecipitation (Co-IP) assay of BnLEC1 protein from 25 DAP seeds of B. napus. Proteins were precipitated from cell lysates using different antibodies and identified by LC-MS/MS. Control precipitations were performed without antibody. Relative abundance represent the ratio of the number of peptide spectral matches derived from the interaction of BnLEC1 with BnNF-YCs and BnSnRK2;2 and BnLEC1 itself. Two biological replicates were performed. (b) Pull-down assay of [BnNF-YCs] and BnLEC1 interaction. His-LEC1 proteins (left) or native LEC1 which was collected by IP from 25 DAP seeds of Brassica napus (right) were incubated with MBP or MBP-[BnNF-YCs] proteins coupled with Amylose Resin. The anti-BnLEC1 or anti-MBP antibody was used for immunoblotting analysis. (c) Co-IP assay of the interaction of BnNF-YCs with BnLEC1 in tobacco (N. benthamiana) leaves coexpressing GFP-[BnNF-YCs] and Flag-BnLEC1. The anti-Flag and anti-GFP antibodies were used for immunoblotting analysis.
Interestingly, the LC-MS/MS assay showed that Ser 17, Ser153 and Ser158 residues in BnLEC1 may be phosphorylated by BnSnRK2;2 in vitro. Furthermore, the MS/MS assay of the proteins coimmunoprecipitated with BnLEC1 in 25 DAP seeds also showed Ser 17, Ser153, and Ser158 residues in BnLEC1 protein may be phosphorylated in vivo. Subsequently, phosphorylation assay revealed that the site-mutated BnLEC1S17A, BnLEC1S17AS153A and BnLEC1S17AS158A could still be phosphorylated, but the triple site-mutated BnLEC1S17AS153AS158A could not phosphorylated by BnSnRK2;2 (Fig. 3e), demonstrating Ser17, Ser153 and Ser158 are three sites of phosphorylation in the BnLEC1 protein. Additionally, amino acid alignment showed that Ser153 and Ser158 are located in the NF-YC interaction domain of BnLEC1 (Supplemental Fig. S9), implying that phosphorylation of these two residues by BnSnRK2 may affect the interaction of BnLEC1 with BnNF-YCs.
To test the above hypothesis, a Co-IP assay was employed to determine the interaction between BnNF-YCs and BnLEC1 as well as a series of mutated versions of BnLEC1 in which the Ser17, Ser153, and Ser158 were successively replaced by Ala or Asp (designated BnLEC1S17A, BnLEC1S17D, BnLEC1S153A, BnLEC1S153D, BnLEC1S158A and BnLEC1S158D), representing non-phosphorylatable or phospho-mimicking forms of BnLEC1, respectively. The GFP-BnNFYC1/4/9A/9B with Flag-BnLEC1 or its six mutated versions were coexpressed in leaves of tobacco (N. benthamiana), respectively. As shown in Fig. 4c, GFP-BnNFYC1/4/9A/9B were coimmunoprecipitated with Flag-BnLEC1, Flag-BnLEC1S153D and Flag-BnLEC1S158D, but not Flag-BnLEC1S153A and Flag-BnLEC1S158A, indicating that phosphorylation of Ser153 and Ser158 of BnLEC1 affects its interaction with BnNF-YC1/4/9A/9B. Moreover, Flag-BnNFYC1/4/9A/9B could be coimmunoprecipitated with both GFP-BnLEC1S17A and GFP-BnLEC1S17D (Fig. 4c), suggested that phosphorylation of Ser17 in BnLEC1 have no effect on its interaction with BnNFYC1/4/9A/9B.
Additionally, BiFC assay was employed to further investigate the protein-protein interaction in vivo. The N-terminal and C-terminal fragments of YFP were fused to BnNF-YCs (BnNFYC1/4/9A/9B) and mutated versions of BnLEC1, respectively. Fluorescence imaging demonstrated that BnNF-YCs could interact with BnLEC1, BnLEC1S153D, and BnLEC1S158D in the cell nuclei, but not with BnLEC1S153A and BnLEC1S158A (Supplemental Fig. S10). Interestingly, the fluorescence signal was observed in the cytoplasm for BnLEC1S17A, while in the nucleus for BnLEC1S17D (Supplemental Fig. S10), suggesting phosphorylation of Ser17 residue in BnLEC1 does not affect its interaction with BnNF-YCs but affects the subcellular localization of these heterodimers. Further analysis showed that BnLEC1 and BnLEC1S17D was located in the nuclei, while BnLEC1S17A was located in the cytoplasm of living tobacco cells (Supplemental Fig. S11). Thus, the above data suggest that phosphorylation of Ser153 and Ser158 in BnLEC1 are crucial for their interaction with BnNF-YCs, while phosphorylation of Ser17 in BnLEC1 is essential for its nuclear localization.
Phosphorylation of BnLEC1 by BnSnRK2;2 enhances the protein stability
-
When the subcellular localization of different versions of BnLEC1 were detected, it was also found that the GFP signals of phosphorylation form of BnLEC1 in the nucleus were stronger than those of its non-phosphorylation form in the cytoplasm (Supplemental Fig. S11), suggesting that phosphorylation of BnLEC1 by BnSnRK2;2 may affect the protein stability. Therefore, seven forms of BnLEC1 (BnLEC1, BnLEC1S17A, BnLEC1S17D, BnLEC1S153A, BnLEC1S153D, BnLEC1S158A and BnLEC1S158D) were fused with GFP driven by CaMV 35S promoter, and transiently expressed in leaves of tobacco (N. benthamiana). The immunoblot analysis using the GFP antibody showed that strong signals were detected in the lanes with BnLEC1, BnLEC1S17D, BnLEC1S153D and BnLEC1S158D proteins, but very weak signals were observed in the lanes with BnLEC1S17A, BnLEC1S153A, and BnLEC1S158A proteins, indicating the relative protein contents of BnLEC1 phosphorylation forms were significantly higher than those of its non-phosphorylation forms (Fig. 5a). Moreover, the level of BnLEC1 protein in tobacco leaves coexpressing Flag-BnSnRK2;2 and GFP-BnLEC1 was significantly higher than the control coexpressing the Flag (EV) and GFP-BnLEC1 (Fig. 5b). The BnLEC1 protein was also detected in 25 DAP seeds of BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177D transgenic plants. Compared with that in the BnSnRK2;2 and BnSnRK2;2T177D transgenic plants, there is a reduced protein level of BnLEC1 in the BnSnRK2;2T177A transgenic plants (Fig. 3f). However, BnLEC1 transcript level in 25 DAP seeds of the transgenic plants had no significant difference from that in WT (Supplemental Fig. S12), suggesting that BnLEC1 protein stability is affected by BnSnRK2;2 at post-transcriptional level. Moreover, the protein level of BnLEC1 in tobacco leaves coexpressing BnSnRK2;2T177A and BnLEC1 was lower than that in tobacco leaves coexpressing BnSnRK2;2/BnSnRK2;2T177D and BnLEC1, but the transcriptional level of BnLEC1 had no change (Fig. 5b), indicating the kinase activity of BnSnRK2;2 is vital for its role in maintaining the stability of BnLEC1 protein. Furthermore, the cell-free degradation assay of BnLEC1, BnLEC1S17A, BnLEC1S17D, BnLEC1S153A, BnLEC1S153D, BnLEC1S158A and BnLEC1S158D were performed which is collected from in vitro kinase reactions containing BnSnRK2;2 and seven versions of BnLEC1, respectively. Equal amounts of the above seven versions of BnLEC1 were incubated with the crude extracts from leaves of Brassica napus, and the samples were used for immunoblotting analysis at various time points (0–120 min). As shown in Fig 5c, BnLEC1S17A, BnLEC1S153A and BnLEC1S158A were degraded more rapidly than BnLEC1, but BnLEC1S17D, BnLEC1S153D and BnLEC1S158D was degraded more slowly than BnLEC1. Taken together, the above data demonstrated that phosphorylation of BnLEC1 by BnSnRK2;2 enhances the stability of BnLEC1 protein in developing seeds of Brassica napus.
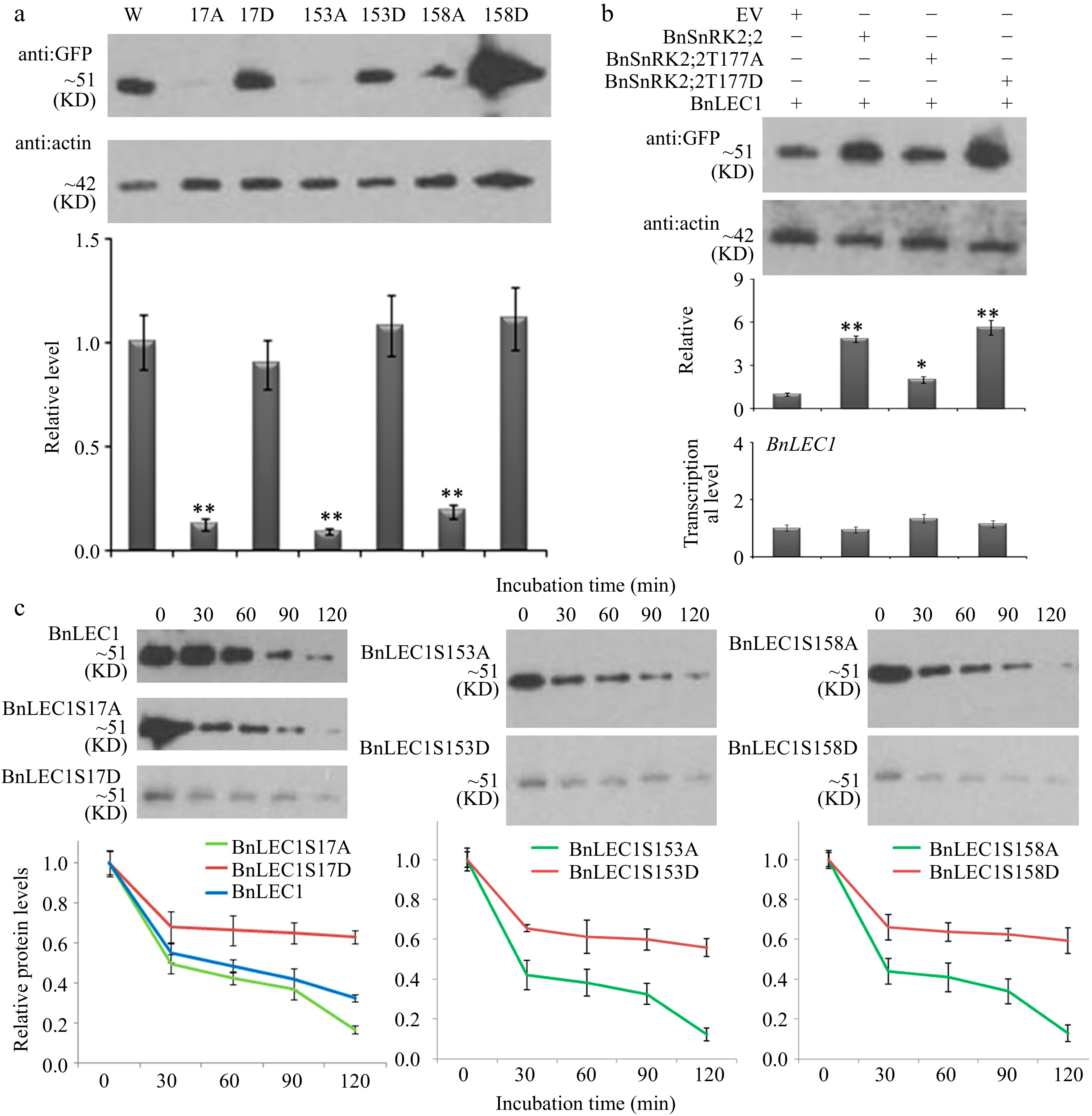
Figure 5.
Assay of BnLEC1 protein stability by its phosphorylation. (a) Protein immunoblotting analysis of the extracts of tobacco (N. benthamiana) leaves expressing NGFP-BnLEC1, NGFP-BnLEC1S17A, NGFP-BnLEC1S17D, NGFP-BnLEC1S153A, NGFP-BnLEC1S153D, NGFP-BnLEC1S158A and NGFP-BnLEC1S158D, respectively. (b) Protein immunoblotting analysis of extracts of tobacco (N. benthamiana) leaves coexpressing NGFP-BnLEC1 and EV (PC2300-NFlag), NFlag-BnSnRK2;2, NFlag-BnSnRK2;2T177A, or NFlag-BnSnRK2;2T177D. (c) Cell-free degradation assay of BnLEC1, BnLEC1S17A, BnLEC1S17D, BnLEC1S153A, BnLEC1S153D, BnLEC1S158A and BnLEC1S158D collected from in vitro kinase reactions containing BnSnRK2;2 and seven versions of BnLEC1, respectively. Equal amounts of BnLEC1, BnLEC1S17A, BnLEC1S17D, BnLEC1S153A, BnLEC1S153D, BnLEC1S158A and BnLEC1S158D were incubated with crude extracts of B. napus leaves supplemented with ATP for time as indicated (in minutes). Immunoblotting assay was performed during cell degradation assays. Three biological replicates were performed. Quantitation of BnLEC1 was achieved using Image J software. Means ± SE are shown (Student's t test, * p < 0.05, ** p < 0.01).
Abscisic acid (ABA) promotes phosphorylation of BnLEC1 by BnSnRK2;2 for oil accumulation in seeds
-
To investigate whether abscisic acid (ABA) also plays an important role in seed oil accumulation, expressions of the genes related to ABA synthesis were examined in seeds of both HFA and LFA at different developmental stages (15 to 40 DAP). As shown in Fig 6a, BnNCED4 and BnNCED9 are expressed at the highest levels in 25 DAP seeds of both HFA and LFA, but expressions of BnNCED4 and BnNCED9 in 25 and 30 DAP seeds of HFA are higher than those in LFA. Furthermore, BnNCED2/5/6 genes are highly expressed in seeds after 35 DAP (Supplemental Fig. S13). Subsequently, ABA content was detected in developing seeds of HFA and LFA. In both materials, ABA was accumulated at a peak value in 25–30 DAP seeds, just at the stages of the accelerated oil accumulation. Moreover, ABA content in 25 and 30 DAP seeds of HFA was higher than that in LFA (Fig. 6b). Consistently, expressions of the ABA-induced genes, BnRAB18 and BnRD29 also showed similar dynamic patterns (Fig. 6a). Immunoblotting analysis revealed that the strongest immunoreactive signals of the phosphorylated BnLEC1 were also detected in developing seeds (25 to 30 DAP) of both HFA and LFA, using anti-R-x-x-(pS/pT)- and anti:BnLEC1 antibodies (Fig. 6c).
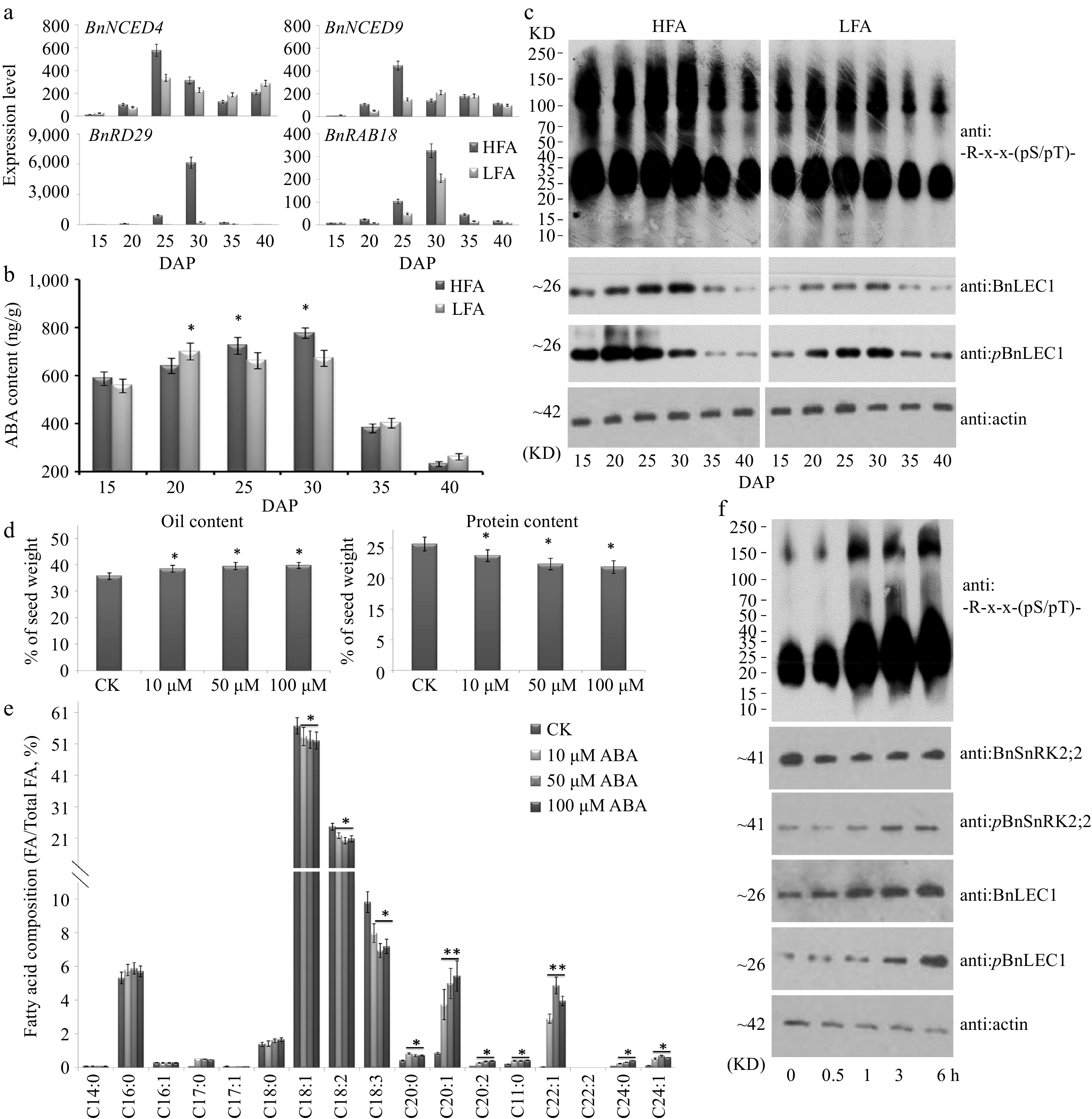
Figure 6.
Assay of abscisic acid (ABA) biosynthesis and physiological function in seed oil accumulation of B. napus. (a) RT-qPCR analysis of time-course expression of ABA biosynthesis genes BnNCED4 and BnNCED9 and ABA-induced maker genes BnRAB18 and BnRD29 in developing seeds of the B. napus varieties HFA and LFA. (b) Quantitation of ABA time-course levels in developing seeds of HFA and LFA. (c) Time-course protein immunoblotting analysis during seed development of HFA and LFA using anti: -R-x-x-(pS/pT)- antibody. Assay of (d) oil and protein contents and (e) fatty acid composition in seeds after ABA treatment. Fifty µM ABA and water (CK) was injected into pod cavities of B. napus (cv. Westar) and sucked out half an hour later with injector from 20 DAP to 35 DAP every 2 d. Then mature seeds were used for analyzing oil and protein contents and fatty acid composition. (f) Protein immunoblotting analysis were performed to detect the influence of exogenous ABA in 25 DAP seeds using antibody of anti:BnLEC1, anti:pBnLEC1, anti:BnSnRK2;2, anti:pBnSnRK2;2 and anti: -R-x-x-(pS/pT)-. Three biological replicates were performed, and means ± SE are shown (Student's t test, * p < 0.05, ** p < 0.01). HFA, a Brassica napus line with high seed oil content; LFA, a Brassica napus line with low seed oil content; DAP, day after pollination.
In addition, the exogenous ABA (10–100 µM) was injected into pod cavities of Brassica napus (cv. Westar) plants from 20 to 35 DAP every 2 d. Compared with controls (treated with water), we found that exogenous ABA could promote oil accumulation and VLCUFAs synthesis in seeds (Fig. 6d,e), in agreement with the results of seed-specific overexpression of BnSnRK2;2 and BnLEC1. Immunoblotting analysis revealed that phosphorylation levels of both BnSnRK2;2 (at Thr177 site) and BnLEC1 (at Ser17 site) were remarkably enhanced in 25 DAP seeds treated with ABA for 0.5–6 h, and the -R-x-x-(pS/pT)- immunoreactive signals were also rapidly heightened in seeds with ABA treatments (Fig. 6f). However, the transcriptional levels of BnSnRK2;2 and BnLEC1 were not altered in seeds after ABA treatments, compared with the controls (Supplemental Fig. S14).
Next, the transgenic Brassica napus plants with seed-specifically downregulated transcript levels of BnABA2 (the key enzyme gene in ABA biosynthesis) were generated through RNA interference (RNAi) technique. Compared with WT, We found expression levels of BnABA2 were remarkably decreased in seeds of the RNAi lines (Supplemental Fig. S15). Immunoblotting analysis revealed that phosphorylation levels of both BnSnRK2;2 (at Thr177 site) and BnLEC1 (at Ser17 site) were remarkably reduced in seeds of the BnABA2 RNAi plants compared with those in wild type control (WT) (Fig. 7a, b). ABA content in 25 DAP seeds of BnABA2 RNAi transgenic lines was lower than that in WT (Fig. 7c). The oil level was decreased, but protein content was increased in seeds of the BnABA2 RNAi plants compared with those in WT (Fig. 7c). Furthermore, levels of VLCUFAs in the BnABA2 RNAi seeds were lower than those in WT, but levels of most medium-long-chain fatty acids (C16:0, C18:0, and C18:1) were increased in the transgenic lines, compared with WT (Fig 7d). Taken together, the above data indicated that ABA plays an important role in promoting phosphorylation of BnLEC1 by BnSnRK2;2 for oil accumulation in seeds of Brassica napus.
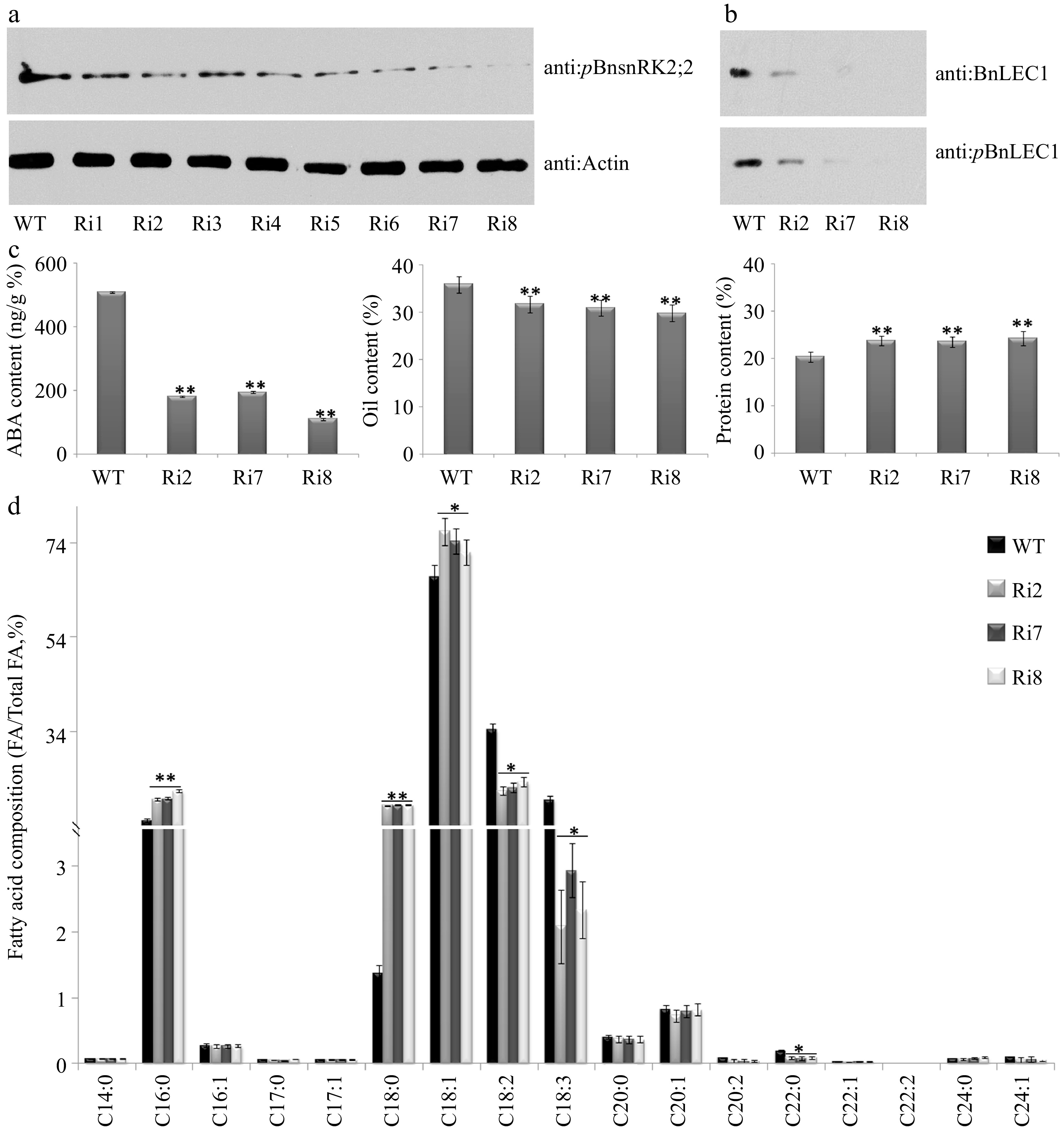
Figure 7.
Assay of oil and protein contents and fatty acid composition in seeds of the BnABA2 RNAi transgenic B. napus. (a) Protein immunoblotting analysis of phosphorylation of BnSnRK2;2 in 25 DAP seeds of BnABA2 RNAi transgenic Brassica napus. Proteins were separated by SDS-PAGE. The anti-pBnSnRK2;2 or anti-actin (control) antibody was used for immunoblotting analysis. (b) Protein immunoblotting analysis of phosphorylation of BnLEC1 at Ser17 residue in 25 DAP seeds of BnABA2 RNAi transgenic B. napus. Proteins were separated by SDS-PAGE as in (a). The anti-BnLEC1 or anti-pBnLEC1 antibody was used for immunoblotting analysis. (c) Assay of ABA content, oil content and protein content in 25 DAP seeds of the BnABA2 RNAi transgenic lines and wild type. The seeds of 10 plants in each line (T2 generation) were used for analysis. Three biological replicates were performed, and means ± SE are shown (Fisher's Least Significant Difference (LSD) test, * p < 0.05, ** p < 0.01). (d) Assay of fatty acid composition in seeds of the BnABA2 RNAi transgenic lines (T2 generation) and WT. Three biological replicates were performed, and means ± SE are shown (Student's t test, * p < 0.05, ** p < 0.01). WT, wild type; Ri, BnABA2 RNAi transgenic lines; DAP, day after pollination.
BnSnRK2;2 mediated phosphorylation of BnLEC1 affects the expressions of the genes related to sucrose photo-assimilation, glycolysis, and FA biosynthesis and modification in seeds
-
Theoretically, if the phosphorylation of BnLEC1 by BnSnRK2;2 enhances the role of BnLEC1 in seed oil accumulation, expressions of those genes regulated by BnLEC1 would be affected when the phosphorylation level of BnLEC1 is changed in seeds. To test this possibility, expressions of several genes related to sucrose photo-assimilation, glycolysis, and FA biosynthesis and modification in 25 DAP seeds of BnLEC1, BnLEC1S17A, BnLEC1S17D, BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177D transgenic Brassica napus and wild type were analyzed. As shown in Fig. 8, the transcriptional levels of these genes were remarkably increased in the BnLEC1 and BnLEC1S17D transgenic seeds, compared with those in the wild type. On the contrary, there was no significant difference in expression levels of these genes between BnLEC1S17A transgenic lines and wild type. Similarly, overexpression of BnSnRK2;2 also enhanced expressions of these genes in seeds. The expressions of these genes in BnSnRK2;2 and BnSnRK2;2T177D transgenic seeds were significantly higher than those in BnSnRK2;2T177A transgenic lines and wild type (Fig. 8). It was noting that expression of BnFAE1 was dramatically increased in seeds of the BnLEC1 and BnSnRK2;2 transgenic lines in which the phosphorylation level of BnLEC1 was enhanced (Fig. 8), consistent with the alteration of FA compositions in these transgenic seeds that displayed significantly higher levels of VLCUFAs. In addition, the expressions of the genes encoded diacylglycerol acyl transferase (DGAT), especially BnDGAT2, were also remarkably increased in the BnLEC1 transgenic seeds with the enhanced phosphorylation level of BnLEC1 (Fig. 8). Collectively, the data suggested that phosphorylation of BnLEC1 by BnSnRK2;2 may be vital for increasing carbon flux to the FA biosynthesis and modification in seeds of Brassica napus.
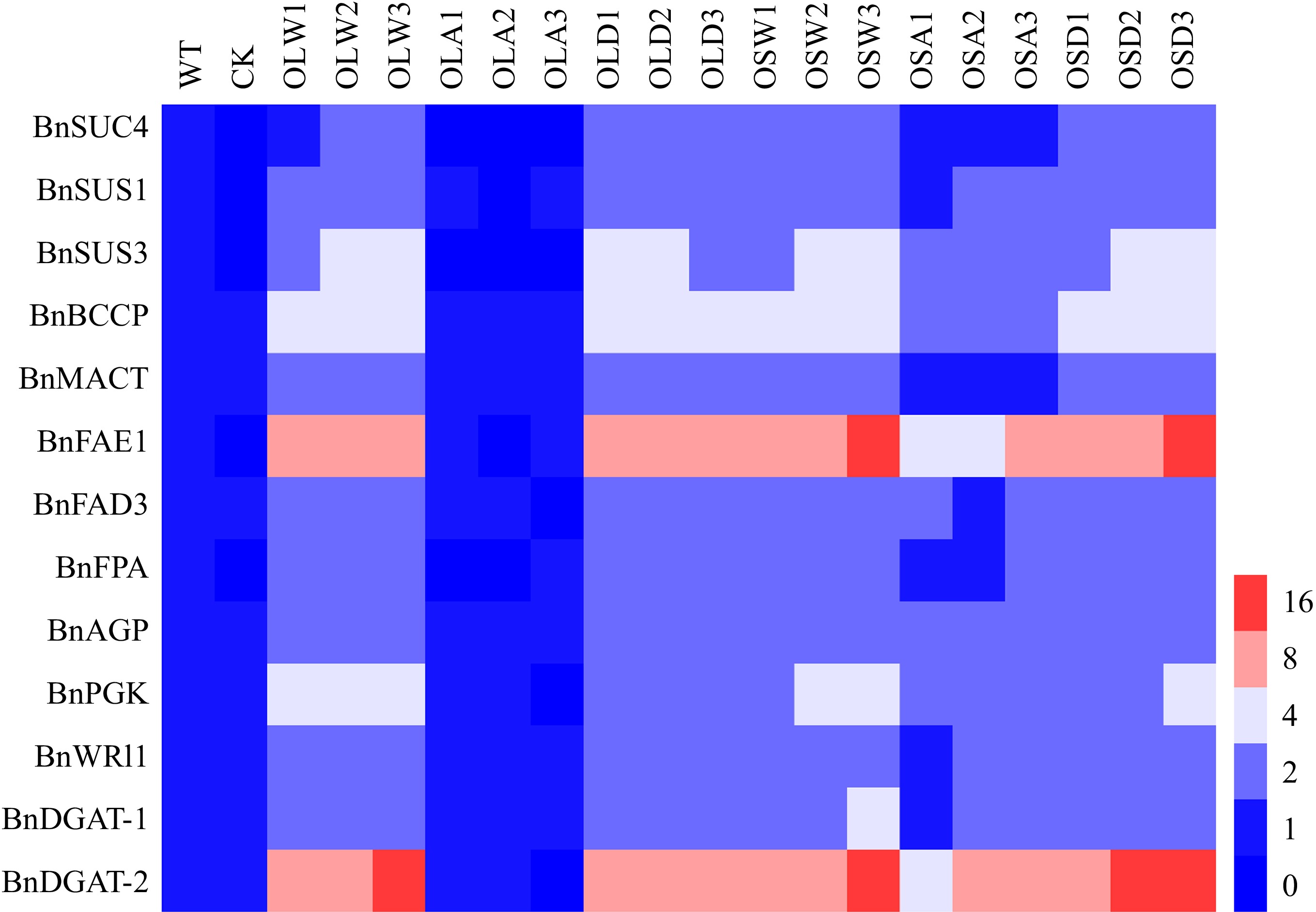
Figure 8.
BnSnRK2;2 mediated phosphorylation of BnLEC1 affects the expressions of the genes related to sucrose photo-assimilation, glycolysis, and fatty acid (FA) biosynthesis and modification. RT-qPCR analysis of the expressions of BnSUS1, BnSUS3, BnSUC4, BnAGP, BnPGK, BnFPA, BnBCCP, BnMACT, BnDGAT1, BnDGAT2, BnFAE1 and BnFAD3 in 25 DAP seeds of the BnLEC1 and BnLEC1S17A, BnLEC1S17D, BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177D overexpression transgenic lines (T3 generation) and controls (WT and CK). WT, wild type; CK, the null transgenic lines; OLW3/9/14, BnLEC1 transgenic lines; OLA4/13/17, BnLEC1S17A transgenic lines; OLD2/9/15, BnLEC1S17D transgenic lines; OSW6/9/13, BnSnRK2;2 transgenic lines; OSA5/11/18, BnSnRK2;2T177A transgenic lines; OSD4/9/12, BnSnRK2;2T177D transgenic lines.
Glucose enhances ABA biosynthesis to positively regulate phosphorylation of BnLEC1 in seeds
-
To investigate whether sugar signal also participates in regulating phosphorylation of BnLEC1 by BnSnRK2;2 for oil accumulation in seeds, exogenous sucrose, glucose, and mannitol were injected into pod cavities of Brassica napus at 25 DAP. Compared with the control, transcriptional levels of ABA synthesis-related genes BnNECD1, BnNECD2, BnNECD4, BnNECD6 and BnNECD9 were remarkably enhanced in seeds treated with glucose, but expressions of these genes were not significantly changed in seeds treated with sucrose and mannitol (Supplemental Fig. S16). Moreover, expression levels of these genes were gradually increased in seeds with the increasing treatment duration (Fig. 9a). On the other hand, exogenous glucose did not affect expressions of BnSnRK2;2 and BnLEC1 (Supplemental Fig. S16). However, immunoblotting analysis showed that the BnLEC1 protein level and its phosphorylation level exhibited a sharp increase in seeds treated with glucose. The immunoreactive signals of -R-x-x-(pS/pT)-, which is recognized by BnSnRK2;2 for BnLEC1 phosphorylation, were also rapidly heightened in seeds treated with glucose (Fig. 9b). Collectively, these results suggested that glucose may promote BnSnRK2;2 phosphorylating BnLEC1 through enhancing ABA biosynthesis in seeds of Brassica napus.
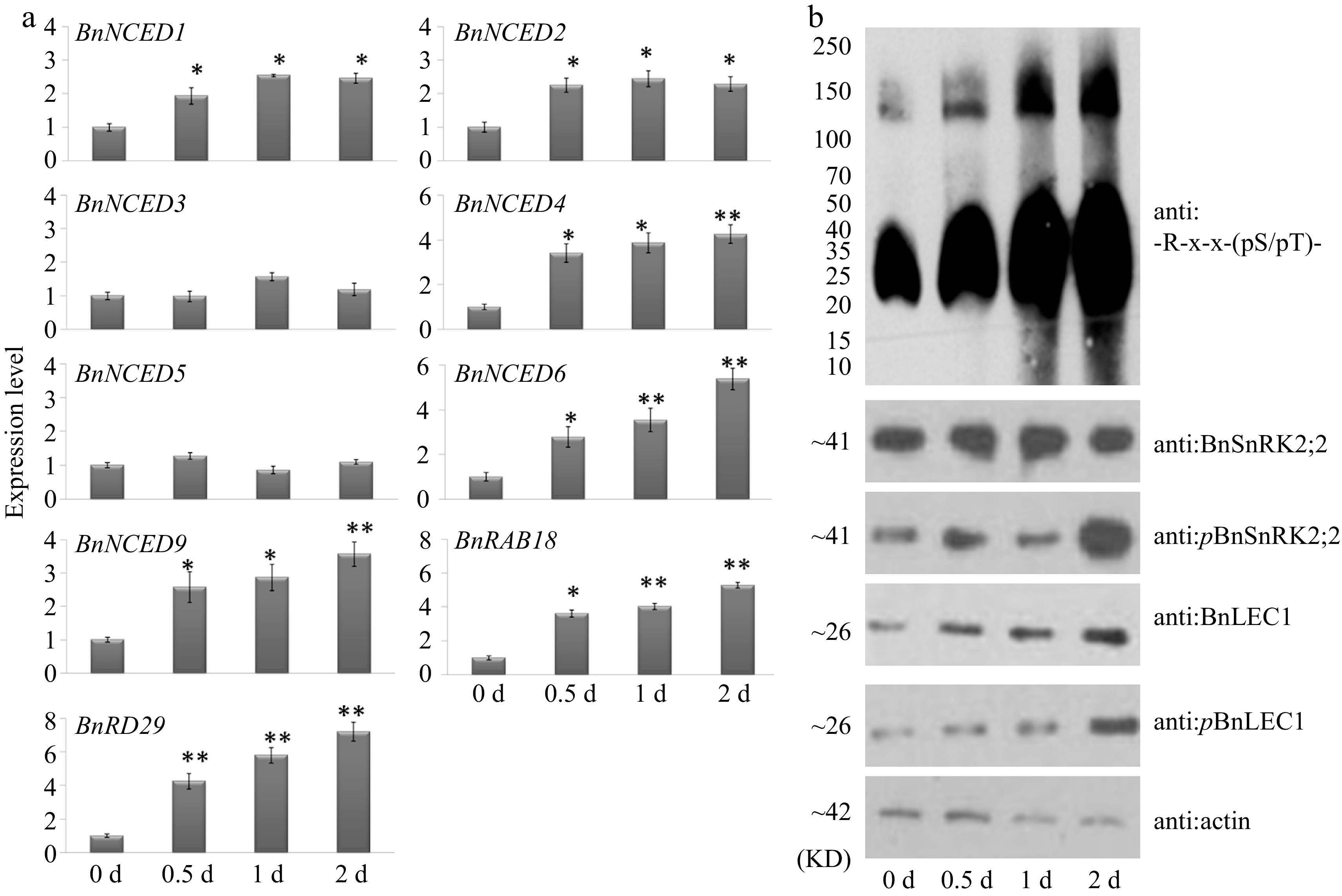
Figure 9.
Glucose promotes expressions of ABA synthesis genes and phosphorylation of BnLEC1 by BnSnRK2;2 in seeds of B. napus. (a) RT-qPCR analysis of the expressions of ABA biosynthesis genes in 25 DAP seeds after glucose treatment for 0–2 d. Three biological replicates were performed, and means ± SE are shown (Student's t test, * p < 0.05, ** p < 0.01). (b) Protein immunoblotting analysis were performed to detect the influence of exogenous glucose in 25 DAP seeds using antibodies of anti:BnLEC1, anti:pBnLEC1, anti:BnSnRK2;2, anti:pBnSnRK2;2 and anti:-R-x-x-(pS/pT)-. Three biological replicates were performed.
-
Protein phosphorylation modification, one of the most widespread posttranslational modifications have been proposed to coordinate multi-levels of both intracellular and extracellular signals during the lipid synthesis. In mammalian, multiple phosphorylation pathways including PDK1/AKT/FoxO1, MAPK signaling pathway and mTORC1-S6K1-SRPK2 signaling pathway have been well-studied in lipid metabolism[36−38]. In Arabidopsis, serine/threonine/tyrosine protein kinases phosphorylate oil body proteins to modulate oil accumulation in seeds[39]. Arabidopsis energy sensor kinase KIN10 phosphorylates WRINKLED1, a regulator involved in partitioning carbon into lipid synthesis in seeds, leading to its proteasomal degradation[35]. Furthermore, a previous study revealed that three oil body proteins (caleosins, steroleosins, and b-glucosidase) were phosphorylated for their functions in B. napus[40]. Through phosphoproteomic analysis, 956 phosphorylation proteins were identified from whole seeds at five sequential stages of development in Arabidopsis thaliana, Glycine max, and Brassica napus[41]. In this study, 76 up-regulated phosphosites and 143 down-regulated phosphosites in seeds were identified at a critical period of oil synthesis via phosphoproteomic analysis of two lines (HFA and LFA) of Brassica napus (Fig. 1c). Therefore, it is speculated that protein phosphorylation might transduce intracellular and extracellular signals during oil accumulation. Additionally, our quantitative phosphoproteomic analysis produces a group of candidate proteins with an arbitrarily defined confidence level (95% of positive rate). As a result, an acceptable small percentage of false-positive results (≤ 5%) is usually present among the selected phosphoproteins (FDR after the Benjamini–Hochberg correction, hereafter FDRB.–H., ≤ 5%).
It has been indicated that the phosphorylation substrates of SnRK2 contain the -R/K-x-x-pS/pT- and/or -pS/pT-x-x-x-x-D/E- motifs that are the phosphorylation sites by SnRK2[25,26]. Similarly, the present data revealed that the motifs [-R/K-x-x-pS/pT-] and [-pS/pT-x-x-x-x-D/E-] are widespread in the identified proteins from the quantitative phosphoproteomics of Brassica napus seeds (Fig. 1d). On the other hand, the role of SnRK2;2/3/6-mediated ABA signaling under abiotic stresses has been well-expounded in different species[11]. In Arabidopsis, it was reported that SnRK2s involved in ABA signaling modulate metabolic balance for leaf growth under nonstress conditions[13]. In this study, it was found that overexpression of BnSnRK2;2 resulted in significantly increasing oil accumulation, decreasing protein levels and increasing the VLCUFAs ratio in seeds of Brassica napus (Fig. 2 & Supplemental Fig. S4). The phosphorylation of BnSnRK2;2 at Thr177 residue is crucial for its role in oil synthesis and accumulation. Moreover, restraining ABA biosynthesis through RNA interference of BnABA2 resulted in a significant decrease of oil accumulation and increase of protein levels. It is known that acetyl coenzyme A derived from glycolytic metabolism and pyruvate metabolism have two different fates: fluxing into tricarboxylic acid (TCA) cycle for respiration or participating in fatty acid biosynthesis. Previous study demonstrated that respiration through the TCA cycle is enhanced in the srk2dsrk2esrk2i mutant under nonstress conditions, suggesting AtSnRK2s play a negative role in fluxing acetyl coenzyme A into TCA cycle in Arabidopsis leaves[13]. In apple and tomato, the SnRK2.3-AREB1-TST1/2 cascade regulates sugar accumulation across tonoplasts[42]. Here, our data showed that BnSnRK2;2 might positively regulate the acetyl coenzyme A participating in fatty acid biosynthesis and oil accumulation in seeds of Brassica napus.
The evidence that BnSnRK2;2 directly phosphorylates BnLEC1 is supported by the following observations: (1) BnSnRK2;2 directly interacted with BnLEC1; (2) BnSnRK2;2 could phosphorylate the S17, S153 and S158 residues of BnLEC1 protein in vitro; (3) Seed-specific overexpression of BnSNRK2;2 resulted in a significant stimulation of phosphorylation of BnLEC1. Furthermore, BnSnRK2;2-medicated phosphorylation of BnLEC1 partly depends on Thr177 phosphorylation of BnSnRK2;2. Similarly, previous studies revealed that phosphorylation of the Thr177 residue in the activation loop of the kinase domain promotes SnRK2;2/3/6 activities in Arabidopsis[11]. These data suggested that BnLEC1 phosphorylation by BnSnRK2;2 might be crucial for oil accumulation in seeds of Brassica napus. However, the phosphorylation of BnLEC1 and oil and protein levels in seeds of double mutant of Bnsnrk2;2/Bnsnrk2;2c have no change, which maybe caused by the functional redundancy of BnSnRK2s. As shown in Supplemental Fig. S6, BnSnRK2;3 and BnSnRK2;6 could also interact with BnLEC1, other BnSnRK2 subfamily protein kinases may also interact with and phosphorylate BnLEC1. However, we failed to acquire the functionally multiple mutations because there are 31 nominated BnSnRK2 subfamily protein kinases with high similarity to AtSnRK2 subfamily and the comprehensive role of SnRK2 in different physiological processes[43].
The role of the LEC1 transcription factor in seed development and nutrient storage has been reported in Arabidopsis and Brassica napus[5−7]. However, the molecular mechanism of LEC1 regulated fatty acid biosynthesis and oil accumulation in Brassica napus seeds is still unclear in detail so far. In this study, seed-specific expression of BnLEC1 driven by BnnapinA promoter resulted in prominently increasing the oil content and VLCUFAs ratio in seeds of the transgenic Brassica napus (Fig. 3). The present data revealed that the role of BnLEC1 in seed oil synthesis and accumulation depends on BnSnRK2;2-medicated phosphorylation of BnLEC1, and the Ser17 residue in BnLEC1 protein is the key phosphorylation site by BnSnRK2;2. The two specific mechanisms of NF-Y regulating gene expression mainly as a protein complex have been studied in plants. On the one side, NF-YB associates with NF-YC to form a heterodimer in the cytoplasm, and then the heterodimer was translocated into the nucleus, where the active heterotrimer of [NF-YA]-[NF-YB]-[NF-YC] were assembled to regulate expressions of target genes[44−46]. On the other side, specific transcription factors interact with [NF-YB]-[NF-YC] heterodimer to form heterotrimer ([NF-YB]-[NF-YC]-transcription factor) to perform a function, but NF-YA can prevent the formation of this hereotrimer[10,47]. In Arabidopsis, co-expression of LEC1 or L1L with NF-YC2 activated the promoter of CRC, depending on the presence of ABA[10]. Although LEC1/L1L-[NF-YC2] can strongly activate the CRC promoter in the absence of ABA when co-expressed with a seed-specific ABA-response element (ABRE)-binding factor bZIP67, the transcriptional activation could be further enhanced after ABA treatment, suggesting that ABA plays multiple roles in the transcriptional activation by LEC1/L1L-[NF-YC2]-bZIP67[10]. In this report, it was found that BnLEC1 can interact with BnNF-YC1, BnNF-YC4, BnNF-YC9A and BnNF-YC9B and the formation of BnLEC1-[BnNF-YC1], -[BnNF-YC4] -[BnNF-YC9A] or -[BnNF-YC9B] dimers depend on the phosphorylation of Ser153 and Ser158 residues in BnLEC1 (Fig. 4). The mutation of the two Ser residues to Ala destructed BnLEC1 interacting with BnNF-YCs. Contrastive analysis found that Ser153 and Ser158 residues in BnLEC1 protein are located in the interaction domain with BnNF-YCs, indicating phosphorylation of the two Ser residues is essential for the formation of BnLEC1-[BnNF-YCs] dimers. Moreover, the Ser17 residue located in the N-terminal of BnLEC1 is also phosphorylated by BnSnRK2;2, which have a vital effect on the subcellular localization of BnLEC1-[BnNF-YCs] complex.
Previous studies revealed ABA is involved in regulating seed development[11,15]. Here, the data elucidated ABA plays an essential role in seed oil biosynthesis possibly via BnSnRK2;2-mediated BnLEC1 phosphorylation pathway. It was found that ABA could elevate oil content and VLCFAs ratio, heighten the -R-x-x-(pS/pT)- -immunoreactive signals in developing seeds, and enhance the phosphorylation levels of BnSnRK2;2 and BnLEC1 (Fig. 6). Additionally, previous reports provided some knowledge about ABA biosynthesis in Arabidopsis seeds[15]. The present data showed that ABA is largely accumulated in seeds at the stage of rapid lipid biosynthesis and then the ABA level is gradually decreased in seeds (Fig. 6). Here, we identify the link between ABA homeostasis and lipid biosynthesis that endogenous ABA accelerate the carbon flow from sugar to lipid biosynthesis through BnSNRK2;2-BnLEC1-[BnNF-YC1/4/9A/9B] cascade which promoted the transcription of sucrose photo-assimilation, glycolysis, FA biosynthetic and FA modification genes during the process of rapeseed seed maturation (Fig. 8). This cascade may provide clues in increasing seed oil accumulation, especially for these cultivars with low levels of endogenous ABA (such as LFA). Sugars derived from photosynthesis in pod walls are transported to seeds for lipid biosynthesis. In Arabidopsis, it had been reported that the elevated glucose levels inhibited the phosphorylation of KIN10, which relieves the KIN10-dependent degradation of WRI1 and then leads to accelerated lipid biosynthesis[35]. In this study, a new homeostatic mechanism is provided that favors oil accumulation when intracellular glucose levels are elevated and BnLEC1 is phosphorylated in seeds, which is supported by the following observations: Exogenous glucose promoted the biosynthesis of ABA; ABA could enhance the phosphorylation level of BnLEC1 and BnSnRK2;2; Exogenous glucose could also heighten the [-R-x-x-(pS/pT)-]-immunoreactive signals in developing seeds; Enhancing expressions of BnLEC1 and BnSnRK2;2 in seeds promoted the transcription of sucrose photo-assimilation, glycolysis, FA biosynthetic, and FA modification-related genes. Although both the present signaling cascade and the KIN10-WRI1 promote the flow of carbon from glucose to lipid biosynthesis, and the target genes of LEC1 partly overlap with that of WRI1, the present signaling cascade has more extensive functions: LEC1 but not WRI1 is required for embryo maturation[3]; SnRK2;2-LEC1 inhibited the protein biosynthesis, but KIN10-WRI1 does not affect protein biosynthesis; Considering the role of ABA in abiotic stress[13], the ABA-BnSnRK2;2-BnLEC1 provides clues about how plants switch resources from growth to reproduction and thus seed ripening in advance under adverse environmental conditions.
In conclusion, a homeostatic mechanism was highlighted that ABA-BnSnRK2;2-BnLEC1-[BnNF-YCs] signaling cascade promotes flow of carbon source to lipid biosynthesis in response to intracellular glucose state (Fig. 10). Thus, the present findings will provide novel insights in designing strategies to increase oil accumulation and VLCFAs level in seeds of Brassica napus.
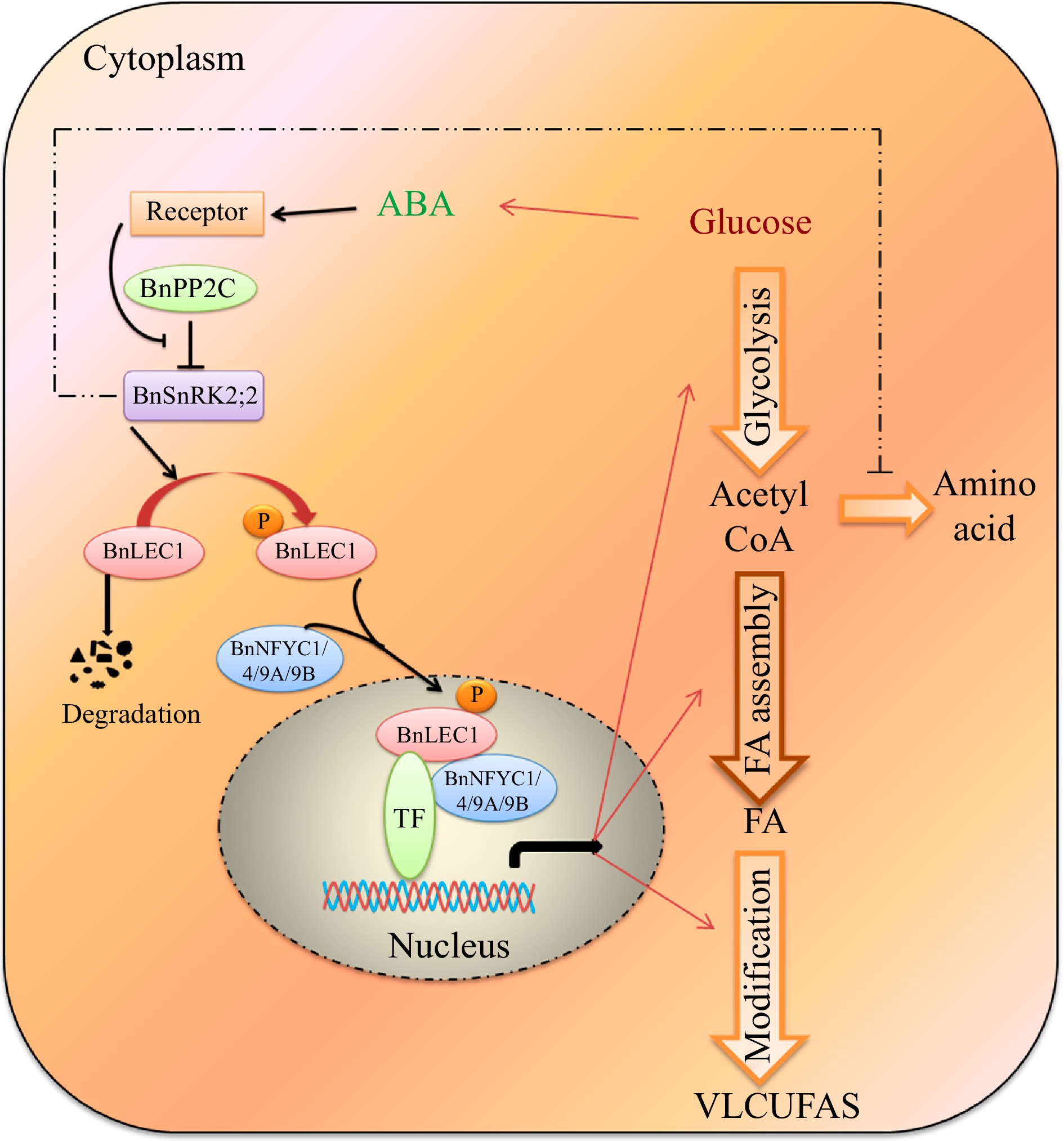
Figure 10.
A proposed model for ABA-dependent BnSNRK2;2 BnLEC1-[BnNF-YCs] signaling cascade promote flow of carbon from glucose to lipid biosynthesis in response to intracellular sugar state. During seed development of Brassica napus, glucose may promote the ABA biosynthesis, and ABA signaling enhances the phosphorylation of BnLEC1 by BnSnRK2;2. Then, the phosphorylated BnLEC1 forms heterodimer with BnNF-YC1/4/9A/9B in the cytoplasm, and is translocated into the cell nucleus, where the BnLEC1 complex activates the expressions of the genes related sucrose photo-assimilation, glycolysis, and FA biosynthetic and modification required for lipid biosynthesis and accumulation in seeds. Thus, ABA-BnPP2C-BnSnRK2;2-BnLEC1-[BnNF-YC1/4/9A/9B] cascade promotes flow of carbon from glucose to lipid biosynthesis in response to intracellular sugar state. On the other hand, BnSnRK2;2 might suppress protein synthesis by inhibiting the flux through the tricarboxylic acid cycle. The fine and thick lines represent transcriptional level and post-translational regulation, respectively. The imaginary line represents the regulation need to be further verified in seeds of B. napus.
-
The authors confirm contribution to the paper as follows: study conception and design: Li XB, Huang KL; data analysis: Huang KL, Zheng Y, Li XB; experiments performed: Huang KL, Li Y, Wang H, Tian J, Fu YF; draft manuscript preparation: Huang KL, Li XB. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was supported by Project of Transgenic Research from the Ministry of Agriculture of China (Grant No. 2016ZX08009003).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Up-regulated phosphopeptides contain [-(K/R)-x-x-(pS/pT)-] motif.
- Supplemental Table S2 Up-regulated phosphopeptides contain [-(pS/pT)-x-x-x-x-(D/E)-] motif.
- Supplemental Table S3 Up-regulated phosphopeptides contain [-(pS/pT)-P-] motif.
- Supplemental Table S4 Assay of oil content in seeds of the transgenic Brassica napus (T2 generation).
- Supplemental Dataset S1
- Supplemental Table S5 Assay of oil content in seeds of the transgenic Brassica napus (T3 generation).
- Supplemental Dataset S2
- Supplemental Table S6 Primers used in vector constructs and RT-qPCR analysis.
- Supplemental Fig. S1 Analysis of oil content in seeds of two Brassica napus lines HFA and LFA at different developmental stages.
- Supplemental Fig. S2 Overview of the quantitative phosphoproteomics analysis.
- Supplemental Fig. S3 Assay of polyclonal antibodies.
- Supplemental Fig. S4 Agronomic traits of BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177A overexpression transgenic Brassica napus.
- Supplemental Fig. S5 Assay of oil and protein content in seeds of the Bnsnrk2;2/Bnsnrk2;2c mutant.
- Supplemental Fig. S6 Bimolecular fluorescence complementation (BiFC) assay of the interaction of BnLEC1 and BnSnRK2,2, BnSnRK2,3, BnSnRK2,6 in leaf lower epidermal cells of tobacco (Nicotiana benthamiana).
- Supplemental Fig. S7 Phosphorylation of Thr177 residue in BnSnRK2;2 is necessary for its kinase activity in phosphorylating BnLEC1 protein.
- Supplemental Fig. S8 Agronomic traits of BnLEC1, BnLEC1S17A and BnLEC1S17D overexpression transgenic Brassica napus.
- Supplemental Dataset S3
- Supplemental Fig. S9 Sequence alignment of AtLEC1, AtL1L and BnLEC1 proteins.
- Supplemental Fig. S10 Bimolecular fluorescence complementation (BiFC) assay of the interaction of BnNF-YCs with 7 versions of BnLEC1 in leaf lower epidermal cells of tobacco (Nicotiana benthamiana).
- Supplemental Fig. S11 Subcellular localization of BnLEC1, BnLEC1S17A and BnLEC1S17D in leaf lower epidermal cells of tobacco (N. benthamiana).
- Supplemental Fig. S12 RT-qPCR analysis of BnLEC1 expression in 25 DAP seeds of BnSnRK2;2, BnSnRK2;2T177A and BnSnRK2;2T177A overexpression transgenic Brassica napus.
- Supplemental Dataset S4
- Supplemental Fig. S13 RT-qPCR analysis of time-course expressions of BnNCED1, BnNCED2, BnNCED3, BnNCED5 and BnNCED6 in seeds of Brassica napus.
- Supplemental Fig. S14 RT-qPCR analysis of expressions of BnLEC1 and BnSnRK2;2 in 25 DAP seeds of Brassica napus with ABA treatment.
- Supplemental Fig. S15 Expression of BnABA2 in seeds of the BnABA2 RNAi transgenic Brassica napus plants.
- Supplemental Fig. S16 RT-qPCR analysis of expressions of BnSnRK2;2, BnLEC1 and ABA synthesis genes in 25 DAP seeds of Brassica napus after 100 mM mannitol, glucose and sucrose treatment for 12 h.
- Supplemental Date S1 Phosphopeptides detected in seeds of Brassica napus by phosphoproteomic analysis (repeat 1, Re 1)
- Supplemental Date S2 The significantly down-regulated phosphopeptides in seeds of Brassica napus (HFA vs LFA).
- Supplemental Date S3 The proteins identified in 25 DAP seeds of Brassica napus by coimmunoprecipitation assay of BnSnRK2;2 protein.
- Supplemental Date S4 The proteins identified in 25 DAP seeds of Brassica napus by coimmunoprecipitation assay of BnLEC1 protein.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Huang KL, Li Y, Wang H, Tian J, Fu YF, et al. 2024. Phosphorylation of BnLEC1 by BnSnRK2;2 is crucial for modulating lipid synthesis in seeds of Brassica napus. Seed Biology 3: e009 doi: 10.48130/seedbio-0024-0009 

Phosphorylation of BnLEC1 by BnSnRK2;2 is crucial for modulating lipid synthesis in seeds of Brassica napus
- Received: 06 February 2024
- Revised: 19 April 2024
- Accepted: 06 May 2024
- Published online: 05 June 2024
Abstract: The intricate machinery responsible for oil synthesis is space/time-strictly controlled in plants, especially in oil crops. Here, we revealed that ABA-dependent phosphorylation of BnLEC1 by BnSnRK2;2 plays a vital role in modulating oil synthesis in seeds of Brassica napus. ABA enhanced BnSnRK2;2 kinase activity for promoting phosphorylation of BnLEC1. BnLEC1-[BnNF-YC1/4/9A/9B] heterodimer was assembled only when Ser153 and Ser158 in BnLEC1 were phosphorylated and translocated into the cell nucleus when Ser17 in BnLEC1 was phosphorylated. Subsequently, the phosphorylated BnLEC1 promoted oil biosynthesis by regulating the expressions of the genes involved in glycolysis and lipid biosynthesis. On the other hand, nonphosphorylated BnLEC1 was rapidly degraded in cells. Furthermore, overexpressions of BnSnRK2;2 and BnLEC1, or adding exogenous ABA resulted in the increased oil content and very long chain unsaturated fatty acids (VLCUFAs) ratio, while restraining ABA biosynthesis led to the decreased oil content and VLCUFAs ratio in seeds of Brassica napus. Additionally, exogenous glucose promoted the expressions of ABA biosynthesis-related genes, leading to the enhanced phosphorylation of BnLEC1. Taken the data together, we uncovered a homeostatic mechanism that ABA-BnSnRK2;2-BnLEC1-[BnNF-YCs] signaling cascade promotes the flow of carbon source to lipid biosynthesis in response to glucose, revealing new insights into oil crop breeding.