









-
The plant cell wall is a unique and complex extracellular matrix that defines cell shape, ensuring structural integrity, growth, and environmental sensing. This sophisticated framework primarily consists of cellulose microfibrils embedded in a matrix of hemicelluloses, pectins, and structural proteins. Unlike the rigid secondary walls of mature tissues, the primary walls of meristematic and growing cells exhibit remarkable viscoelastic properties that permit controlled cell expansion while resisting turgor pressure[1,2]. Pectins, the most complex family of cell wall polysaccharides, are highly hydrated, gel-forming components of this matrix[3]. A central regulator of wall mechanics is pectin methylesterification, which acts as a biochemical rheostat[4]. Newly synthesized pectin is deposited in a highly methylesterified form, which is subsequently modified by pectin methylesterases (PMEs) that remove methyl groups from homogalacturonan chains[5], allowing for two divergent fates: it can enable Ca2+ cross-linking to form stiff 'egg-box' structures or facilitate polygalacturonase-mediated degradation[6]. This post-synthetic modification system, regulated by pH gradients, and PME inhibitors (PMEIs), defines regional mechanical domains that influence wall flexibility for cell expansion and morphogenesis[7].
This principle is exquisitely illustrated at the single-cell level. In the rapidly growing pollen tube, a steep biochemical gradient of pectin methylesterification is closely associated with a corresponding mechanical heterogeneity[8]. At the tip, highly esterified and thus extensible and soft pectins facilitate tip extension under turgor pressure[9]. Over a short distance behind the apex, the accumulated demethylesterified pectins facilitated by PMEs are cross-linked by Ca2+, reinforcing the wall of the cylindrical shank[10]. Finite element models predict that this precise compartmentalization—a soft apex transitioning sharply to a stiff shank—is essential for maintaining the cylindrical shape of the growing tube and directed growth[11]. Consequently, this precise control by localized pectin chemistry dictates local wall mechanics, enabling rapid tip extension while maintaining structural integrity.
At the tissue level, pectin methylesterification patterns play an instructive role in organ initiation and morphogenetic patterning[12]. In the shoot apical meristem (SAM), the emergence of a new organ primordium is preceded by localized pectin demethylesterification, which is associated with a mechanical softening of the cell wall[13,14]. Perturbing this pattern, either by overexpressing PME or PMEI in the meristem or by locally applying PME, is sufficient to alter primordium outgrowth and disrupt phyllotactic patterning[13,14]. Similar pectin-based asymmetries have also been implicated in other developmental contexts, including lateral root initiation[15], and apical hook bending[16]. Collectively, coordinated pectin remodelling across cell populations helps define tissue-level morphogenesis by establishing mechanical contexts that physically permit or restrict growth in specific domains. An important remaining question is how such spatial specificity is established with sufficient precision in proliferating tissues, where new cell walls are continuously generated during cell division and must rapidly acquire distinct pectin states.
A recent breakthrough by Zhu and colleagues uncovered a novel, mechanically distinct pectin pattern within the SAM itself, one that spatially regulates stem cell dynamics[17]. Through systematic cell wall immunolabelling and super-resolution imaging, they showed that mature cell walls of the shoot meristem cells exhibit high methylesterification, whereas demethylesterified pectins are specifically deposited at new cross walls formed during cell division. This spatial heterogeneity is established through an unexpected mechanism: nuclear sequestration of PECTIN METHYLESTERASE5 (PME5) mRNA. The transcription of PME5 is activated by the R1R2R3-MYB transcription factor MYB3R4 at the onset of mitosis[18,19]. However, rather than being immediately exported for translation, the PME5 transcripts are strictly retained within the nucleus. This retention ensures that PME5 enzyme activity is precisely synchronized with cell division. Only upon nuclear envelope breakdown are the PME5 mRNAs released for translation, which enables localized pectin demethylesterification specifically at division planes while preserving the high methylesterification of mature walls essential for stem cell maintenance (Fig. 1).
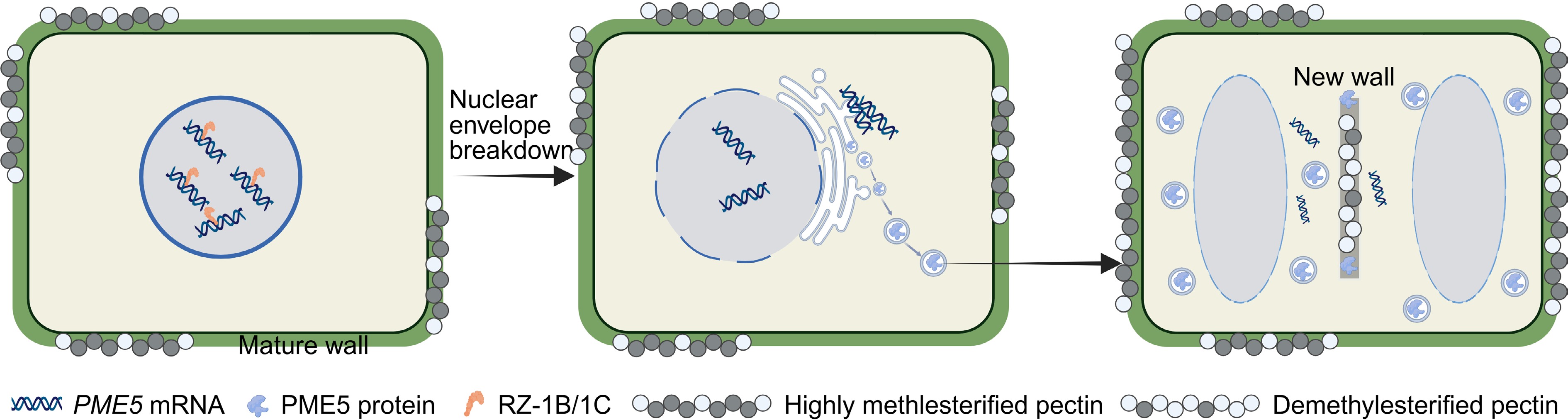
Figure 1.
Working model for RZ-1B/1C–PME5-mediated precise pectin modification in stem cell dynamics and meristem homeostasis. During formation and maintenance of the shoot apical meristem, PME5 transcripts (encoding a cell-wall pectin methylesterase) are not immediately exported to the cytoplasm after transcription but are bound by the nuclear RNA-binding proteins RZ-1B and RZ-1C, and retained within the nucleus, thereby limiting PME5 protein production, and maintaining a highly methyl-esterified pectin state in pre-existing cell walls. Upon entry into mitosis and nuclear-envelope breakdown, the previously sequestered PME5 mRNAs are rapidly released into the cytoplasm and are efficiently translated into PME5 protein. Zhu et al.[17] propose that newly synthesized PME5 is transported through the secretory pathway to specific domains of the nascent cell wall, where it catalyzes local pectin demethylesterification. This spatially and temporally restricted remodeling of pectin architecture is proposed to fine-tune cell-wall mechanical properties and the local extracellular microenvironment, thereby contributing to the coordination of stem-cell behaviour with shoot apical meristem homeostasis. Created in BioRender:
https://BioRender.com/6edfu6t .The nuclear sequestration of PME5 mRNA represents an unconventional regulatory strategy for gene expression control. Zhu et al. demonstrated that disrupting this mechanism by engineering a truncated PME5 mRNA, PME5Δ1–102, that escapes the nucleus led to ectopic PME5 production, widespread pectin demethylesterification, and severe developmental defects, including reduced cell division and meristem termination[17]. These findings elegantly establish nuclear mRNA compartmentalization as a critical post-transcriptional mechanism that spatially restrict enzyme activity, thereby maintaining distinct pectin modification states in neighbouring cellular domains and ensuring proper stem cell homeostasis.
The mechanistic link between RNA processing and cell wall dynamics was uncovered by the identification of RZ-1B and RZ-1C[20] as the proteins mediating PME5 mRNA nuclear retention. These glycine-rich RNA-binding proteins with zinc finger domains localize to nuclear speckles and function redundantly to retain PME5 transcripts in the nucleus. In vitro pull-down assays, and in vivo immunoprecipitation showed that both proteins directly bind PME5 mRNA. In rz-1b rz-1c double mutants, PME5 mRNA sequestration was abolished and precociously exported for translation, leading to pectin demethylesterification throughout the cells. These mutants recapitulated the phenotypes observed in PME5Δ1–102 overexpression lines, confirming their essential role in maintaining stem cell homeostasis.
Further investigation revealed that nuclear sequestration is not unique to PME5 but represents a more widespread regulatory strategy among PME family members. Moreover, comparative immunofluorescence analysis in soybean, tomato, and maize revealed a conserved accumulation of demethylesterified pectin at newly formed cross walls in dividing cells, indicating evolutionary conservation of this spatiotemporal regulatory mechanism despite the phylogenetic divergence of these species. This conservation underscores the fundamental importance of precise pectin modification control in plant development.
In conclusion, the work by Zhu et al. represents a major conceptual advance in plant developmental biology, revealing how the integration of transcriptional control, mRNA localization, and protein-RNA interactions enables precise spatiotemporal regulation of cell wall remodelling to maintain stem cell homeostasis[17]. This study also highlights several intriguing questions for future research. First, how exactly do RZ-1B and RZ-1C recognize and retain specific PME mRNAs, and what structural features determine the target specificity? Second, to what extent does this mechanism contribute to the regulation of other cell wall-modifying enzymes beyond PMEs? Third, how is the release of sequestered mRNAs coordinated with cell cycle progression? Given that shoot apical meristem stem cells generate all aerial organs, including the reproductive structures that give rise to seeds, manipulating pectic cell wall modification could offer a new approach to optimizing stem cell activity, seed yield, and crop quality.
HTML
This work was supported by the National Key R&D Program of China (2025YFE0200400), National Natural Science Foundation of China (32425009, 32170343), CAS Project for Young Scientists in Basic Research (Grant No. YSBR-078), and the National Key Research and Development Program of China (2022YFF1003500).
-
The authors confirm contributions to the paper as follows: draft manuscript preparation: Liu HR; manuscript revision: Li HJ. Both authors reviewed the results and approved the final version of the manuscript.
-
Data sharing is not applicable to this article, as no new data were created or analyzed in this study.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Liu HR, Li HJ. 2026. The stem cell mRNA gatekeeper: from nuclear sequestration to cell wall remodelling. Seed Biology 5: e002 doi: 10.48130/seedbio-0026-0001


|