










-
Rice (Oryza sativa L.) is a world staple crop that feeds over half of the world's population[1,2]. In recent years, high-temperature events are becoming more frequent and intensive as a result of global warming, which can severely affect rice grain yield and quality[3,4]. During flowering and grain filling stages, high-temperature stress can result in a significant reduction in seed setting rate and influence amylose content, starch fine structure, functional properties and chalkiness degree of rice[5−7]. Transcriptome and proteome analysis in rice endosperm have also been used to demonstrate the differences in high-temperature environments at gene and protein expression levels[8−10]. In addition, as an important post-translational modification, protein phosphorylation has proven to be involved in the regulation of starch metabolism in response to high-temperature stress[11]. However, little is known about whether protein ubiquitination regulates seed development under high-temperature stress.
Ubiquitination is another form of post-translational modification that plays key roles in diverse cellular processes[12]. Several reports have described the functions of ubiquitination in rice defense responses based on ubiquitome analysis. Liu et al.[13] investigated relationships between ubiquitination and salt-stress responses in rice seedlings using a gel-based shotgun proteomic analysis and revealed the potential important role of protein ubiquitination in salt tolerance in rice. Xie et al.[14] identified 861 peptides with ubiquitinated lysines in 464 proteins in rice leaf cells by combining highly sensitive immune affinity purification and high resolution liquid chromatography-tandem mass spectrometry (LC-MS/MS). These ubiquitinated proteins regulated a wide range of processes, including response to stress stimuli. A later study revealed the relationships between ubiquitination status and activation of rice defense responses, and generated an in-depth quantitative proteomic catalog of ubiquitination in rice seedlings after chitin and flg22 treatments, providing useful information for understanding how the ubiquitination system regulates the defense responses upon pathogen attack[15]. Although many studies have shown that ubiquitination plays improtant roles in the heat response of plant[16,17], there has been little systematic discussion on the ubiquitome of rice endosperm in the context of global climate change.
In this study, we examine the high-temperature induced ubiquitination change in two rice varieties with different starch qualities, through a label-free quantitative ubiquitome analysis. This study provides a comprehensive view of the function of ubiquitination in high-temperature response of rice developing seed, which will shed new light on the improvement of rice grain quality under heat stress.
-
Two indica rice varieties with different starch quality, 9311 and Guangluai4 (GLA4), were used as materials. 9311 is a heat-sensitive variety, which displays low amylose content with good starch quality; while GLA4 is known to be the parental variety of HT54, an indica breeding line with heat tolerance, and thus GLA4 is possibly heat tolerant, which shows high amylose content with poor starch quality[18,19]. Rice growth conditions, sample treatment and collection were conducted as previously described[11].
Protein extraction
-
Husk, pericarp and embryo were detached from immature rice grains on ice[20]. Rice endosperm was then ground with liquid nitrogen, and the cell powder was sonicated 10 rounds of 10 s sonication and 15 s off-sonication on ice in lysis buffer (6 M Guanidine hydrochloride, pH 7.8−8.0, 0.1% Protease Inhibitor Cocktail) using a high intensity ultrasonic processor. Following lysis, the suspension was centrifuged at 14,000 g for 40 min at 4 °C to remove the debris. The supernatant was collected, and the protein concentration was estimated using BCA assay (Pierce BCA Protein assay kit, Thermo Fisher Scientific, Waltham, MA, USA) before further analysis.
Trypsin digestion
-
The protein mixture was reduced by DTT with the final concentration of 10 mM at 37 °C for 1 h, alkylated by iodoacetamide with a final concentration of 50 mM at room temperature in the dark for 0.5 h, and digested by trypsin (1:50) at 37 °C for 16 h. Then the sample was diluted by adding trifluoroacetic acid (TFA) to the final concentration of 0.1%. The enzymatic peptides were desalted on a Sep-Pak C18 cartridge (Waters, Milford, MA, USA), concentrated by lyophilization and reconstituted in precooled IAP buffer (50 mM MOPS-NaOH PH 7.2, 10 mM Na2HPO4, 50 mM NaCl) for further analysis.
Affinity enrichment of ubiquitinated peptides
-
The peptides solution was incubated with prewashed K-ε-GG antibody beads (PTMScan Ubiquitin Remnant Motif (K-ε-GG) Kit), and gently shaken at 4 °C for 1.5 h. The suspension was centrifuged at 2,000 g for 30 s, and the supernatant was removed. The Anti-K-ε-GG antibody beads were washed with IAP Buffer three times and with ddH2O three times. The peptides were eluted from the beads with 0.15% trifluoroacetic acid (TFA). Finally, the eluted fractions were combined and desalted with C18 Stage Tips.
LC-MS/MS and data analysis
-
LC-MS/MS analysis were performed using the methods of Pang et al.[11]. Raw mass spectrometric data were analyzed with MaxQuant software (version 1.3.0.5) and were compared with the indica rice protein sequence database (Oryza sativa subsp. indica-ASM465v1). Parameters were set according to Pang et al.[11]. All measurements were obtained from three separate biological replicates.
Bioinformatics methods
-
Quantification of the modified peptides was performed using the label-free quantification (LFQ) algorithm[11]. Differentially ubiquitinated sites (proteins) in response to high-temperature were identified by Student's t-test (p < 0.05, log2(fold-change) > 1) with at least two valid values in any condition or the ubiquitination sites that exhibited valid values in one condition (at least two of three replicates) and none in the other.
Subcellular localization was performed using CELLO database (
http://cello.life.nctu.edu.tw ). Gene Ontology (GO) annotation proteome was derived from the AgriGO (http://bioinfo.cau.edu.cn/agriGO/ ). The differential metabolic profiles were visualized with MapMan software (version 3.6.0RC1). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway annotation was performed by using KEGG Automatic Annotation Server (KAAS) software. A p-value of < 0.05 was used as the threshold of significant enrichment. SWISS-MODEL was used to generate the tertiary structure of GBSSI (SWISS-MODEL,http://swissmodel.expasy.org/ ). The figures were annotated with Adobe Illustrator (Adobe Systems, San Jose, CA, USA). -
To elucidate how high-temperature stress influences rice developing endosperm at the ubiquitination level, a label-free analysis was performed to quantify ubiquitome from two indica rice varieties under normal (9311-C and GLA4-C) and high-temperature conditions (9311-H and GLA4-H). The distribution of mass error of all the identified peptides was near zero and most of them (71.5%) were between -1 and 1 ppm, suggesting that the mass accuracy of the MS data fits the requirement (Fig. 1a). Meanwhile, the length of most peptides distributed between 8 and 42, ensuring that sample preparation reached standard conditions (Fig. 1b).

Figure 1.
Characteristics of the ubiquitinated proteome of rice endosperm and QC validation of MS data. (a) Mass error distribution of all identified ubiquitinated peptides. (b) Peptide length distribution. (c) Frequency distribution of ubiquitinated proteins according to the number of ubiquitination sites identified.
In all endosperm samples, a total of 437 ubiquitinated peptides were identified from 246 ubiquitinated proteins, covering 488 quantifiable ubiquitinated sites (Supplemental Table S1). Among the ubiquitinated proteins, 60.6% had only one ubiquitinated lysine site, and 18.7%, 8.1%, 5.3%, or 7.3% had two, three, four, or five and more ubiquitinated sites, respectively. In addition, four proteins (1.6%, BGIOSGA004052, BGIOSGA006533, BGIOSGA006780, BGIOSGA022241) were ubiquitinated at 10 or more lysine sites (Fig. 1c, Supplemental Table S1). The proteins BGIOSGA008317 had the most ubiquitination sites with the number of 16. It was noted that besides ubiquitin, two related ubiquitin-like proteins NEDD8 and ISG15 also contain C-terminal di-Gly motifs generated by trypsin cleavage, and the modifications of these three proteins cannot be distinguished by MS[21]. Here, the di-Gly-modified proteome therefore represents a composite of proteins modified by these three proteins. However, the sites from NEDD8 or ISG15 modifications were limited because they mediate only a few reactions in cells[21].
Properties of ubiquitinated proteins and motifs
-
To better understand the lysine ubiquitome changes in rice endosperm induced by high-temperature, we performed a Gene Ontology (GO) functional annotation analysis on all identified ubiquitinated proteins (Fig. 2a). In the biological process GO category, 'metabolic process' and 'cellular process' were mainly enriched, accounting for 75.1% and 74.1% of ubiquitinated proteins, respectively. In addition, 34.6% proteins were associated with 'response to stimulu', emphasizing the regulatory role of ubiquitination modification in response to high-temperature stress. From the cellular component perspective, ubiquitinated proteins were mainly associated with 'cellular anatomical entity' (99.4%), 'intracellular' (84.4%) and 'protein-containing complex' (29.9%). The molecular function category result suggested that these proteins were largely involved in 'binding' (62.7%), 'catalytic activity' (43.4%) and 'structural molecule activity' (16.5%). Furthermore, subcellular location annotation information indicated that 34.7%−39.4% proteins were located in the cytoplasm, and other were mostly located in the nucleus (23.5%−27.7%), plasma membrane (9.4%−11.4%), and chloroplast (9.6%−12.8%) (Fig. 2b). It is noteworthy that the ubiquitinated proteins located in the cytoplasm were decreased in high-temperature environments in both varieties.

Figure 2.
Analysis of ubiquitinated proteins and motifs. (a) Gene ontology (GO) functional characterization of ubiquitinated proteins. (b) Subcellular localization of ubiquitinated proteins. From the inside out, the ring represents 9311-C, 9311-H, GLA4-C and GLA4-H, respectively. (c) Motif enrichment analysis of ubiquitinated proteins.
The following two significantly enriched motifs from all of the identified ubiquitinated sites were identified using MoDL analysis: [A/S]xKub and Kubxx[E/Q/R/V]x[E/G/L/P/Q/R/Y], which covered 84 and 100 sequences, respectively (Fig. 2c). Further analysis showed that the conserved alanine (A) and glutamic acid (E) were included in upstream and downstream of the ubiquitinated lysine sites in rice endosperm. A similar phenomenon also occurred in rice leaf[14], wheat leaf[22], and petunia[23], indicating that alanine (A) and glutamic acid (E) were likely to be the specific amino acids in conserved ubiquitination motifs in plants. Additionally, serine (S) was enriched at the position -2 (upstream) of the ubiquitinated lysine, while various amino acids such as arginine (R), glutamic acid (E), glutamine (Q), valine (V) were found at positions +3 and +5 (downstream).
A temperature-regulated rice endosperm ubiquitome
-
To detect possible changes in rice endosperm ubiquitome attributable to high-temperature stress, we then performed LFQ analysis on all quantifiable ubiquitination sites within our dataset. As shown in Fig. 3a, more ubiquitinated proteins, peptides and sites were detected in the treatment groups (9311-H and GLA4-H), suggesting that exposure to high-temperature stress may increase the ubiquitination events in rice endosperm. Only 282 common ubiquitinated sites in 158 proteins were quantifiable for all sample groups due to reversible ubiquitination induced by high-temperature (Fig. 3b). Principal component analysis (PCA) showed that three repeats of each sample clustered together, and four groups were clearly separated (Fig. 3c). Furthermore, the differentially expression profiles of ubiquitination sites (proteins) in 9311 and GLA4 under high-temperature stress were depicted to further understand the possible changes (Fig. 3d). Where LFQ values were missing, the data were filtered to identify those ubiquitination sites with a consistent presence/absence expression pattern. These analyses yielded 89 ubiquitination sites that were only present in 9311-H and six that were only present in 9311-C (Fig. 3d, Supplemental Table S2). Similarly, 51 differentially expressed ubiquitination sites were present in GLA4-H and 13 ubiquitination sites only occurred in GLA4-C (Fig. 3d & Supplemental Table S3). Beyond that, a total of 113 and 50 significantly changed ubiquitination sites (p < 0.05, log2(fold-change) >1) were screened out in 9311 and GLA4, respectively (Fig. 3d, Supplemental Tables S4 & S5). For subsequent comparative analysis, the ubiquitination expression profiles with consistent presence/absence and ubiquitination sites with significant differences in statistical testing were combined and named as 9311-Up, 9311-Down, GLA4-Up, and GLA4-Down, respectively (Fig. 3d). The number of significantly up-regulated ubiquitination sites was far greater than down-regulated ubiquitination sites in both 9311 and GLA4 varieties. These findings indicated that high temperature not only induced the occurrence of ubiquitination sites, but also significantly upregulated the intensity of ubiquitination. Beyond that, the magnitude of the up-regulation in 9311 was higher than that in GLA4 (Fig. 3b & d), indicating that the ubiquitination modification of heat-sensitive varieties was more active than heat-resistant varieties in response to high-temperature stress.

Figure 3.
A temperature regulated rice endosperm ubiquitome. (a) The number of ubiquitinated proteins, peptides and sites detected in four group samples. (b) Venn diagram of ubiquitination sites (proteins) detected in four group samples. (c) PCA based on ubiquitination intensity across all four sample groups with three biological repetitions. (d) Differentially expression profiles of ubiquitination sites (proteins) in 9311 and GLA4 under high-temperature stress. The expression profiles of selected ubiquitination sites (p < 0.05, log2(fold-change) >1) were normalized using the Z-score and presented in a heatmap.
To further investigate the ubiquitination regulatory pattern under high temperature stress in two varieties, four groups with significantly regulated sites were analyzed. There were 37 ubiquitination sites showed the same regulatory trend in 9311 and GLA4, accounting for 17.8% and 32.5% of the total differentially expressed sites in 9311 and GLA4, respectively. Among them, 36 ubiquitination sites were upregulated and one site was downregulated (Fig. 4a). In addition, 159 upregulated ubiquitination sites and three downregulated sites were only present in 9311, while 53 upregulated sites and 15 downregulated sites were only present in GLA4. Moreover, nine ubiquitination sites showed opposite regulatory trends in 9311 and GLA4. A similar regulatory trend of ubiquitination proteins is shown in Fig. 4b. It is noted that some proteins had both upregulated and downregulated ubiquitination sites (Supplemental Tables S6 & S7), indicating that significant differences in ubiquitination were, to some extent, independent of protein abundance.
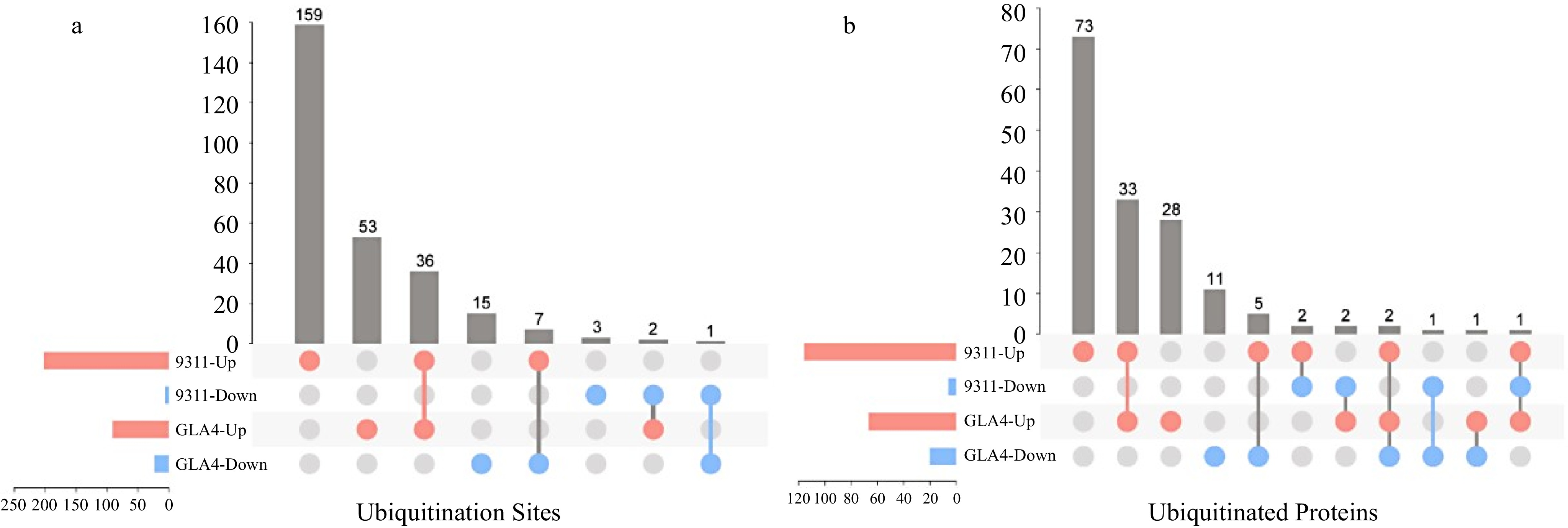
Figure 4.
Comparison of differentially ubiquitinated sites and proteins in 9311 and GLA4 under high-temperature stress.
Functions for differentially expressed ubiquitinated proteins
-
To understand the function of ubiquitination in response to the high-temperature stress of rice endosperm, we conducted GO enrichment-based clustering analysis of the differentially ubiquitinated proteins in 9311 and GLA4 at high temperature, respectively (Fig. 5). In the biological process category of 9311, proteins were relatively enriched in the carbohydrate metabolic process, polysaccharide metabolic process, starch biosynthetic process, cellular macromolecule localization, protein localization, intracellular transport, and phosphorylation (Fig. 5). For the molecular function analysis, we found that the proteins related to kinase activity, nucleotidyltransferase activity, phosphotransferase activity, and nutrient reservoir activity were enriched (Fig. 5). The two principal cellular components were intrinsic component of membrane and integral component of membrane (Fig. 5). There was no significantly enriched GO term in the GLA4 group due to the dataset containing relatively few proteins, and thus, further enrichment analysis was conducted on the proteins that were common to both varieties. The results showed that proteins were over-represented in carbon metabolism, including starch biosynthesis and metabolism, glucan biosynthesis and metabolism, and polysaccharide biosynthesis and metabolism (Fig. 5), indicating the importance of carbohydrate synthesis and metabolism in the ubiquitination regulatory network.

Figure 5.
Enrichment analysis of differentially expressed ubiquitinated proteins based on Gene Ontology (GO) terms.
To identify pathways which were differentially ubiquitinated under high-temperature stress, the KEGG pathway-based clustering analysis was conducted. The results showed that the differentially ubiquitinated proteins in both 9311 and GLA4 were mostly abundant in the pathways of carbohydrate metabolism, starch and sucrose metabolism, folding, sorting and degradation, translation, ribosome, and protein processing in endoplasmic reticulum (Fig. 6a). In the 9311 group, the pathways of carbohydrate metabolism, starch and sucrose metabolism, glycosyltransferases, glycolysis, and energy metabolism were enriched in the differentially ubiquitinated proteins (Fig. 6b); while there was no significantly enriched KEGG pathway in the GLA4 group. We further found the proteins that were common to both varieties were only significantly enriched in the starch and sucrose metabolism pathways (p = 0.04). The ubiquitination proteins involved in the starch and sucrose metabolism mainly include: sucrose hydrolysis (SUS, FK, UGPase), and starch synthesis (AGPase, GBSSI, BEI, BEIIb, PUL, PHO1), which are discussed below.
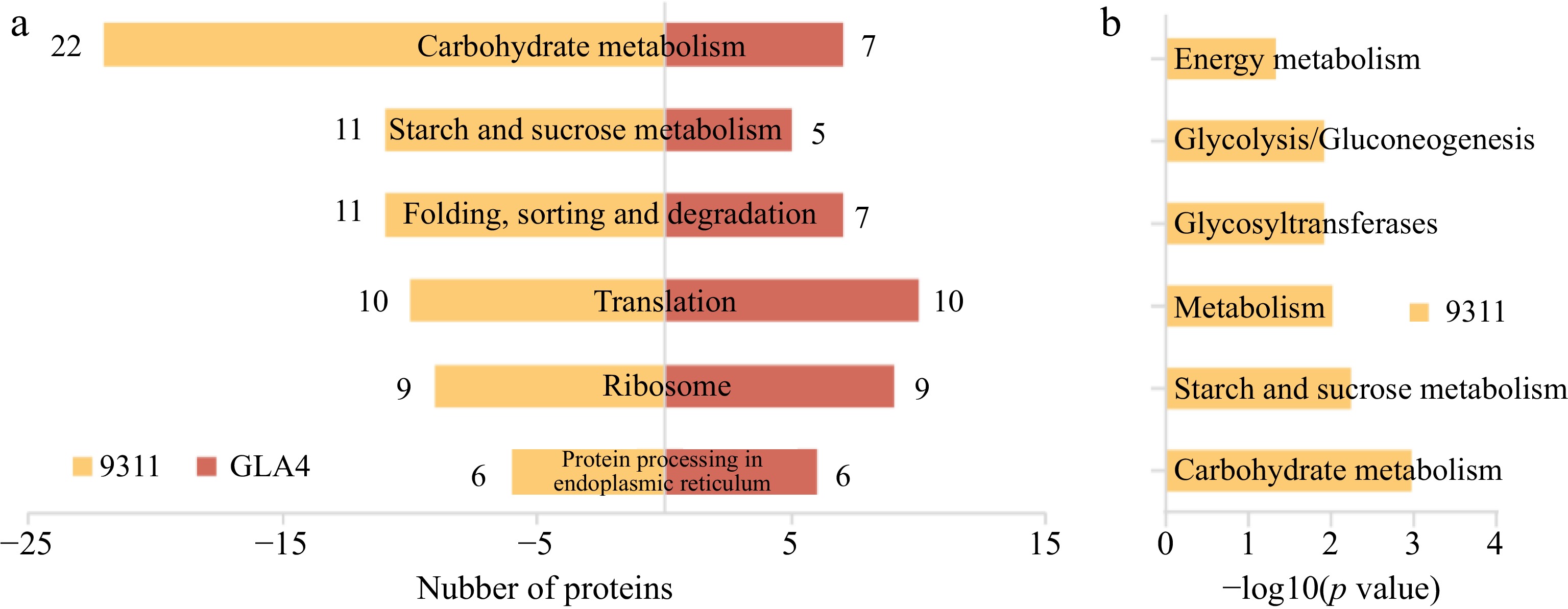
Figure 6.
KEGG classification and enrichment analysis of differentially ubiquitinated proteins. (a) Number of differentially ubiquitinated proteins based on KEGG classification in 9311 and GLA4. (b) KEGG enrichment analysis of differentially ubiquitinated proteins in 9311.
-
Although many reports have described specific examples of ubiquitination in rice defense responses[13,15,16], our knowledge on global changes in the developing endosperm ubiquitome under high-temperature stress is still lacking. In this study, a label-free quantitative proteomic analysis of ubiquitination was applied to examine the high-temperature induced ubiquitination change of two indica rice varieties (9311 and GLA4) with distinct starch quality. We identified many new lysine modification sites on proteins involved in various pathways, highlighting the complexity of the ubiquitination-mediated regulatory system in high-temperature stress responses in rice.
Ubiquitination of heat shock proteins and transcription factors under high-temperature stress
-
Heat shock proteins accumulate under various stresses and play important roles in plant defenses against abiotic stresses[24,25]. Research has shown that a number of heat shock proteins were prominent in the rice ubiquitome network, of which OsHSP71.1, and OsHSP82A showed increased ubiquitination levels under chitin and flg22 treatment[15]. Here, seven lysine residues on five heat shock proteins possessed ubiquitination modification in rice endosperm. Three sites (BGIOSGA011420-K78, BGIOSGA026764-K99, BGIOSGA029594-K106) showed significant up-regulation in 9311 under high-temperature stress, while in GLA4, the ubiquitination level of BGIOSGA011420-K78 was down-regulated. This differential ubiquitination of heat-tolerant and heat-sensitive varieties provided a basis for studying the regulation of post-translational modification of heat shock proteins under high-temperature stress, despite the regulatory role of those heat shock proteins being still unclear.
Transcription factors (TFs) play an essential role in the regulation of gene expression. A total of three transcription factors were identified in the ubiquitination dataset, of which two were NAC family members. As one of the largest plant-specific TF families, NAC is involved in the responses to abiotic and biotic stresses[26]. The ubiquitination modification of K173 in BGIOSGA018048 was specifically expressed in the 9311-H group, which may affect the stress resistance level in high-temperature environments. In addition, two sites K148 and K149 of ERF TF family member BGIOSGA024035 was downregulated in GLA4 under high-temperature stress. This differential ubiquitination was likely to affect the expression of related genes regulated by this transcription factor.
Sucrose and starch metabolism are modified by ubiquitination under high-temperature stress
Ubiquitination affects sucrose hydrolysis
-
The results of GO and KEGG enrichment analysis of differential ubiquitination proteins indicated that the sucrose and starch metabolic pathway was largely affected by ubiquitination regulation under high-temperature stress (Figs 5 & 6). The ubiquitination sites involved in sucrose and starch metabolism are listed in Table 1. To assess how high-temperature stress affects the crucial pathway, the significantly differential ubiquitination sites in 9311 and GLA4 were displayed in the heatmap of specific proteins (Fig. 7).
Table 1. Ubiquitination sites related to sucrose and starch metabolism in rice endosperm.
Gene name Annotation Protein entry Modification site(s) SUS1 Sucrose synthase 1 BGIOSGA010570 K172, K177 SUS2 Sucrose synthase 2 BGIOSGA021739 K160, K165, K176, K804 SUS3 Sucrose synthase 3 BGIOSGA026140 K172, K177, K541, K544, K588 FK Fructokinase BGIOSGA027875 K143 UGPase UDP-glucose pyrophosphorylase BGIOSGA031231 K27, K150, K303, K306 AGPS1 ADP-glucose pyrophosphorylase small subunit 1 BGIOSGA030039 K94, K464, K484 AGPS2 ADP-glucose pyrophosphorylase small subunit 2 BGIOSGA027135 K106, K132, K385, K403, K406, K476, K496 AGPL2 ADP-glucose pyrophosphorylase large subunit 2 BGIOSGA004052 K41, K78, K134, K191, K227, K254, K316, K338, K394, K396, K463, K508, K513 AGPL3 ADP-glucose pyrophosphorylase large subunit 3 BGIOSGA017490 K509 GBSSI Granule bound starch synthase I BGIOSGA022241 K130, K173, K177, K181, K192, K258, K371, K381, K385, K399, K462, K517, K530, K549, K571, K575 BEI Starch branching enzyme I BGIOSGA020506 K103, K108, K122 BEIIb Starch branching enzyme IIb BGIOSGA006344 K134 PUL Starch debranching enzyme:Pullulanase BGIOSGA015875 K230, K330, K431, K736, K884 PHO1 Plastidial phosphorylase BGIOSGA009780 K277, K445, K941 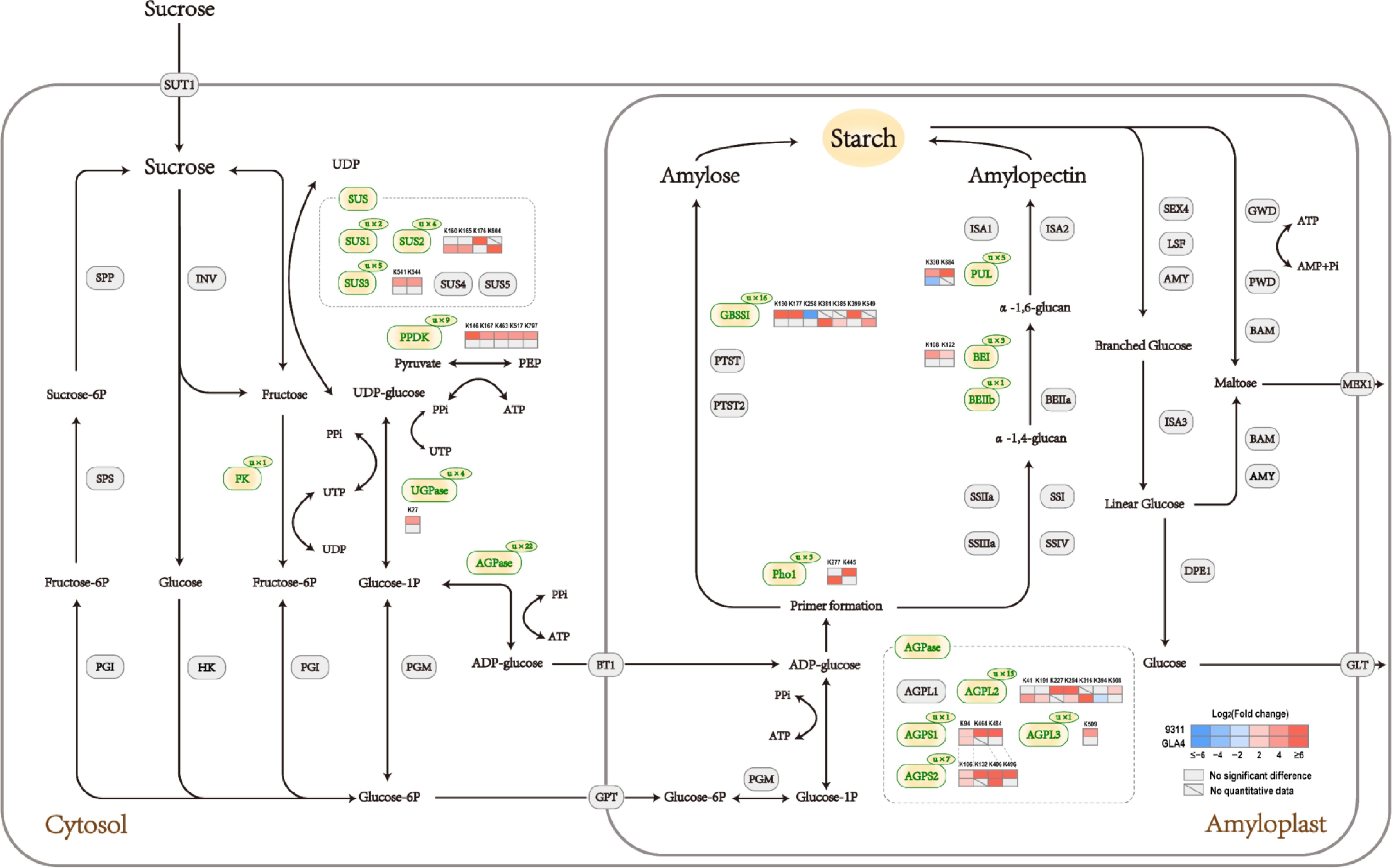
Figure 7.
Sucrose and starch pathway at the ubiquitination levels in rice endosperm under high-temperature stress.
In cereal endosperm, sucrose is the substrate for the biosynthesis of starch. The formation of glucose 1-phosphate (G1P, used in starch synthesis, see below) from sucrose requires a series of enzymes[27]. Here, we found that sucrose synthase 1 (SUS1), SUS2, SUS3, fructokinase (FK), and UDP-glucose pyrophosphorylase (UGPase) were ubiquitinated among all sample groups (Table 1, Fig. 7). In the ubiquitome of seedling and leaf in japonica rice, ubiquitination sites have been found in SUS1, SUS2, UGPase, and FK, which were related to sucrose hydrolysis[14,15]. SUS catalyzed the process of cleaving sucrose into UDP-glucose (UDPG) and fructose. Two ubiquitination sites, K172 and K177, were identified in SUS1 in rice endosperm, which were also found in rice leaves[14]. A total of four ubiquitination sites were identified in SUS2, two of which were also reported in rice seedling and leaf, indicating the conservation of the lysine residues in different rice tissues. It was noted that all four ubiquitination sites in SUS2 were upregulated in high-temperature environments, although the regulated sites of 9311 and GLA4 were different. In 9311, the ubiquitination levels of K160, K174, and K804 were increased, while GLA4 was only upregulated in K176. The ubiquitination sites K541, K544, and K588 in SUS3 were screened from developing rice seeds for the first time. In addition, SUS3 had two completely overlapping sites K172 and K177 with SUS1, and it was difficult to determine which enzymes the two sites belonged to. The ubiquitination levels of SUS3-K541 and SUS3-K544 in 9311 significantly increased in high-temperature environments, while there was no significant difference in the ubiquitination level of SUS3 in GLA4. Overall, the ubiquitination sites of SUS in rice endosperm were located in the functional domain except for SUS2-K804, reflecting the importance of ubiquitination regulation in SUS.
UGPase catalyses the conversion of glucose 1-phosphate and UTP into UDPG[28]. Research has shown that the mutation of UGPase gene lead to chalky endosperm[29]. As shown in Table 1, four ubiquitination sites K27, K150, K303, and K306 were identified in rice endosperm, which were completely inconsistent with the seven ubiquitination sites in rice seedlings and two in leaves[14,15], reflecting the tissue specificity. We speculated that the UGPase with different modification sites may play different regulatory roles in metabolic pathways in different tissues. Under high-temperature stress, the ubiquitination level of UGPase-K27 was 8.1-fold up-regulated. Liao et al.[10] demonstrated that the expression of UDPase was down-regulated in both heat-tolerant and heat-sensitive rice lines under high temperature conditions, which could reasonably explain the significant up-regulation of UGPase-K27 ubiquitination level. The ubiquitination site K143 of FK was also reported in seedling tissues[15].
Ubiquitination of ADP-glucose pyrophosphorylase (AGPase)
-
The AGPase reaction represents the first committed step of starch biosynthesis[27]. A total of 22 lysine ubiquitination sites were identified in four AGPase subunits (AGPL2, AGPL3, AGPS1, AGPS2). AGPL2 had 13 ubiquitination sites, of which six were located in NTP_transferase domain, including K254, K338, K191, K134, K227, and K316. High-temperature stress resulted in an increase in the ubiquitination level of K254 in both 9311 and GLA4, and significant upregulation of K508 and K513 in 9311, as well as K191, K227, and K316 in GLA4. In contrast, AGPL2-K394 were significantly downregulated in GLA4. AGPL3 contained one ubiquitination site K509, and the modification level of AGPL3-K509 was up-regulated in high-temperature environments in 9311. AGPS1 had one specific ubiquitination site K464 and another two sites K94 and K484 that completely overlapped with AGPS2-K106 and AGPS2-K496, respectively. The modification levels of K464 and K484 significantly increased in high-temperature environments in 9311, and K94 was significantly up-regulated in both varieties. There were seven ubiquitination sites in AGPS2 in rice endosperm, which were different with the sites found in rice leaves[14]. In addition to the two sites that overlapped with AGPS1, AGPS2 had another two ubiquitination sites (K406 and K132) that upregulated in high-temperature environments.
Ubiquitination regulates amylose biosynthesis
-
Amylose content is one of the key determinants that strongly influence rice grain quality[30]. The biosynthesis of amylose requires the catalytic effect of granule-bound starch synthase I (GBSSI)[30,31]. Here, a total of 16 ubiquitination sites were identified in GBSSI (Table 1, Fig. 8a). Among these ubiquitination sites, six lysine residues (K130, K173, K177, K181, K192, K258) were located in glycosyltransferase 5 (GT5) domain, and three sites (K399, K462, K517) were located in GT1 domain (Fig. 6), indicating the important role of ubiquitination regulation of GBSSI. Under high-temperature stress, the ubiquitination levels of six sites (K130, K177, K399, K381, K385, K549) increased in two indica rice varieties, while one sites (K258) showed downregulation in 9311 (Fig. 8a). Numerous studies had described that the amylose content was reduced under high-temperature stress in rice[5,7], which might be due to the degradation of GBSSI proteins caused by the increased significantly up-regulated ubiquitination sites. These ubiquitination sites identified in rice GBSSI with significant differences under high-temperature stress were expected to become a new breakthrough point for the improvement of starch quality.
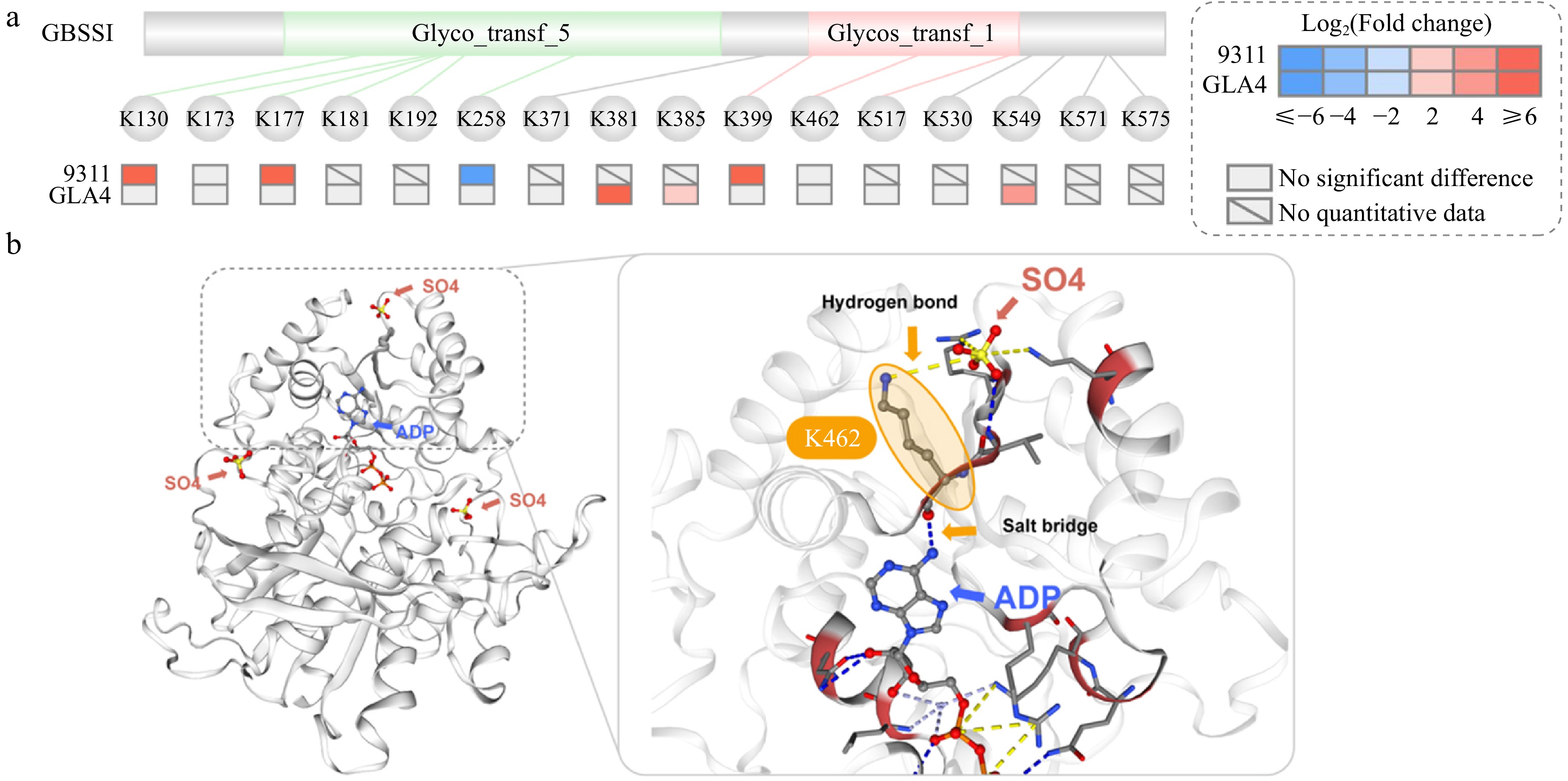
Figure 8.
Structure of GBSSI. (a) Domain structure of GBSSI and ubiquitination sites with significant differences in response to high-temperature stress. (b) 3D model of GBSSI and the relationship between ubiquitination sites K462 and ADP, SO4 (salt bridge or hydrogen bond).
To further determine the regulatory role of the ubiquitination sites in GBSSI, SWISS-MODEL was used to predict 3D structural model. As shown in Fig. 8b, GBSSI had three SO4 (sulfate ions) and one ADP ligand. These ligands interact with GBSSI through hydrogen bonds and salt bridges. Three sites, K447, R458, and K462, were associated with SO4 through salt bridges, while G100, N265, Q412, K462, and Q493 interact with the hydrogen bonds of ADP in GBSSI[32,33]. Based on this finding, it can be reasonably inferred that the K462 site with ubiquitination modification located in the GT1 domain played an important role in the interaction between GBSSI, SO4, and ADP. An in-depth investigation was necessary to gain a more comprehensive understanding of the regulatory function of ubiquitination modification at GBSSI-K462, although there was no significant difference in the ubiquitination level under high-temperature stress.
Ubiquitination regulates amylopectin biosynthesis
-
Amylopectin, the major component of starch, is synthesized by the coordinated action of multiple enzymes including soluble starch synthase (SSs), starch branching enzyme (BEs), starch debranching enzyme (DBEs), and phosphorylases (PHOs or Phos) with ADPG as a substrate. In this study, ubiquitination sites were detected in BEs, DBEs, and Phos.
BEs, covering two isoforms, BEI and BEII, are responsible for catalyzing the formation of α-1,6-glucosidic linkages of amylopectin[34]. There were three ubiquitination sites (K103, K108, and K122) identified in BEI (Fig. 9a). K122 was the first amino acid in the carbohydrate-binding module 48 (CBM48) domain. Sequence alignment analyses of BEs from eight plants revealed K122 was conserved among all plants' BEI (Fig. 9a), suggesting a high probability of the functional effects of ubiquitination modification at this site. In high-temperature environments, ubiquitination levels of K108 and K122 were significantly up-regulated in 9311, while no significantly regulated ubiquitination sites of BEI were observed in GLA4. Only one ubiquitination site, K134, was found in BEIIb (Fig. 9a). The ubiquitination levels showed a slightly upward trend with no significant differences in high-temperature environments in both varieties. These changes could be one of the reasons for increased gelatinization temperature and relative crystallinity of rice starch in response to high-temperature[5].

Figure 9.
Domain structure of (a) BEs, (b) PUL and (c) Pho1 as well as their ubiquitination sites with significant differences in response to heat stress. Residues in red indicate the ubiquitination site. Non-ubiquitinated residues are shown in dark grey.
DBEs consists of isoamylase (ISA) and pullulanase (PUL) with catalytic function for hydrolyzing α-1,6-glucosic linkages[35]. In the present study, we found that only PUL was ubiquitinated in rice endosperm (Fig. 9b). Among five ubiquitination sites (K230, K330, K432, K736, and K884) identified in PUL, K230 was located in the PULN2 domain, while K330 was in the CBM48 domain. Under high-temperature stress, K330 showed completely opposite regulatory trends in two cultivars. In addition, the ubiquitination level of K884, located in the DUF3372 domain, was significantly up-regulated in 9311. Previous study has reported that the expression of PUL was significantly up-regulated in 9311 under high-temperature stress, while GLA4 showed down-regulation in PUL abundance[11]. Consequently, there might be two possible functions of these ubiquitination sites. One possibility is that ubiquitination sites were unrelated to protein degradation; instead, they regulated the biosynthesis of amylopectin by affecting other functions of the protein. Secondly, ubiquitination sites were associated with protein degradation, and the levels of ubiquitination modification were based on protein abundance, resulting in a completely consistent regulation of ubiquitination modification and protein abundance under high-temperature stress.
PHOs, including two types, Pho1/PHO1 and Pho2/PHO2, are responsible for the transfer of glucosyl units from Glc-1-P to the non-reducing end of a-1,4-linked glucan chains[36]. Pho1 is a temperature-dependent enzyme and considered crucial not only during the maturation of amylopectin but also in the initiation process of starch synthesis[37,38]. The three ubiquitination sites (K277, K445, K941) identified in Pho1 were located in two phosphorylase domains. We found that two sites, Pho1-K277 and Pho1-K445, were only ubiquitinated in high-temperature environments in 9311 and GLA4, respectively. Pang et al.[11] has demonstrated that the protein abundance of Pho1 decreased under high-temperature stress, especially in GLA4. Satoh et al.[38] reported that the functional activity of Pho1 was weakened under conditions of high temperature and its function might be complement by one or more other factors. Hence, these ubiquitination modifications that specifically occurred in high-temperature environments might be related to the degradation of Pho1 proteins.
Ubiquitination affects ribosomal systems under high-temperature stress
-
As a factory for protein synthesis in cells, the ribosome is an extremely crucial structure in the cell[39]. It has been proven that multiple ribosomal subunits were abundantly ubiquitinated in Arabidopsis and wheat[22]. In the present study, 57 ubiquitination sites involving 33 ribosome subunits were identified in 40S and 60S ribosome complexes in rice. Under high-temperature stress, the ubiquitination levels of some sites were significantly upregulated or downregulated, implying that ubiquitination of ribosomal proteins is likely to be an important regulatory mechanism in high-temperature response in rice endosperm. The results of GO and KEGG enrichment analysis indicated that the ribosome system was one of the most active systems for ubiquitination regulation under high-temperature stress. We speculated that the ubiquitin-proteasome system might be involved in the removal of subunits or entire ribosomes that were improperly folded in high-temperature environments. As shown in Fig. 10, the S10e, L18Ae, S27Ae, L9e, S3e, S28e, S20e, and S2e subunits were significantly up-regulated in 9311, while L13e subunits showed a completely opposite regulatory trend at the ubiquitination sites K81 and K88. In GLA4, the ubiquitination levels of S10e, S27Ae, L10Ae, L9e, S3e, S2e, and L4e showed a significant increase, while the ubiquitination level of L17e was significantly down-regulated under high-temperature stress. A total of seven ubiquitination sites involving S10e, S27Ae, L9e, S3e, and S2e subunits were jointly up-regulated in both two varieties. These sites might be related to the degradation of improperly folded ribosome subunits under high-temperature stress, while other ubiquitination sites with variety specificity might be associated with ribosomal function.
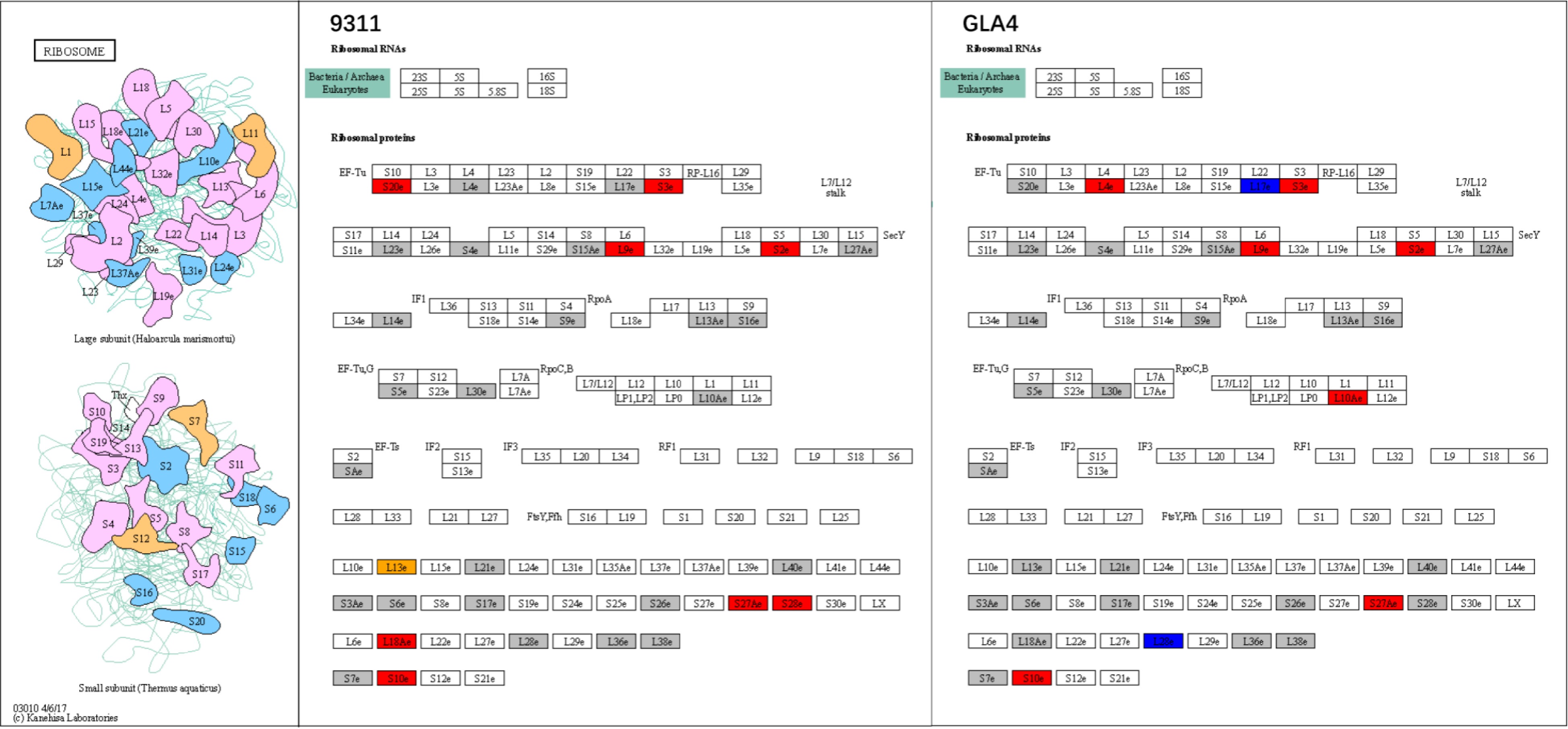
Figure 10.
Ribosome system at the ubiquitination levels in rice endosperm under high-temperature stress. Grey shadings represent ubiquitinated proteins with no significant differences under heat stress. Red and blue shadings indicate up-regulated and down-regulated ubiquitinated proteins, respectively. Orange shading displays a combination of up- and down-regulated ubiquitinated sites in the same ubiquitinated protein.
In conclusion, this study provides the first comprehensive view of the ubiquitome in rice developing endosperm, and demonstrated that ubiquitination has diverse functions in the high-temperature response of rice endosperm by modulating various cellular processes, especially the sucrose and starch metabolism. Comparative analysis of the temperature-induced ubiquitination status revealed some similarities and more interesting differences between 9311 and GLA4. These differences might be the reason for the different qualities formation of the two indica rice varieties, which could provide potential genetic resources for the improvement of the heat resistance in rice. Considering the diversity of ubiquitination modification, it is worthwhile to further validate and explore the function and regulatory mechanism of the key targets and key pathways. The findings provide valuable insights into the role of ubiquitination in response to high-temperature stress and lay a foundation for further functional analysis of lysine ubiquitination in rice.
-
The authors confirm contribution to the paper as follows: study conception and design: Bao J, Pang Y; data collection: Pang Y; analysis and interpretation of results: Pang Y; draft manuscript preparation: Ying Y; Revised manuscript preparation: Ying Y, Pang Y, Bao J. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This work was financially supported by the AgroST Project (NK2022050102) and Zhejiang Provincial Natural Science Foundation (LZ21C130003).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 List of ubiquitination sites identified in rice endosperm.
- Supplemental Table S2 Consistent presence/absence expression profile of 9311 under high-temperature stress.
- Supplemental Table S3 Consistent presence/absence expression profile of GLA4 under high-temperature stress.
- Supplemental Table S4 List of ubiquitination sites significantly changed under high-temperature stress in 9311.
- Supplemental Table S5 List of ubiquitination sites significantly changed under high-temperature stress in GLA-4.
- Supplemental Table S6 Comparison of differentially ubiquitinated sites in 9311 and GLA4 under high-temperature stress.
- Supplemental Table S7 Comparison of differentially ubiquitinated proteins in 9311 and GLA4 under high-temperature stress
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ying Y, Pang Y, Bao J. 2023. Comparative ubiquitome analysis reveals diverse functions of ubiquitination in rice seed development under high-temperature stress. Seed Biology 2:23 doi: 10.48130/seedbio-2023-0023 

Comparative ubiquitome analysis reveals diverse functions of ubiquitination in rice seed development under high-temperature stress
- Received: 16 September 2023
- Accepted: 09 November 2023
- Published online: 05 December 2023
Abstract: Protein ubiquitination plays vital roles in regulation of diverse cellular processes during plant growth and development. However, how protein ubiquitination regulates seed development in high-temperature environments is less understood. Here, a label-free quantification identified 488 lysine modification sites in 246 ubiquitinated proteins in the endosperm of two rice varieties, 9311 and Guangluai4 (GLA4). Under high-temperature stress, the number of significantly up-regulated sites was far greater than down-regulated sites, and 37 ubiquitinated proteins were commonly regulated with the same trend in the two varieties. The sucrose and starch metabolism were greatly over-represented by functional and pathway enrichment analyses. The key functions of ubiquitinated proteins related to starch metabolism are SUS1, SUS2, SUS3, FK and UGPase for sucrose hydrolysis, and AGPL2, AGPL3, AGPS1, AGPS2, GBSSI, BEI, BEIIb, PUL and Pho1 for starch synthesis. Most lysine modification sites were first identified in rice and tended to be up-regulated under heat stress, providing evidence for decreased protein abundance of starch synthesis related enzyme at the ubiquitination level. Predicted 3D models of GBSSI revealed an important role of ubiquitylation sites K462 involved in the interaction between the GBSSI and ligands (SO4 and ADP). Our study provides the first comprehensive view of the ubiquitome in rice seeds, which will provide important insight into the mechanism underlying seed development and grain quality improvement under high-temperature stress.
-
Key words:
- Rice endosperm /
- Ubiquitination /
- High temperature /
- Sucrose and starch metabolism /
- Starch biosynthesis