








-
Fungi are crucial for maintaining the health and sustainability of terrestrial ecosystems, primarily through their roles in nutrient cycling, organic matter decomposition, and symbiotic interactions with nearly all plant species[1−3]. Beyond these fundamental ecological contributions, fungi significantly impact agricultural productivity and forest ecosystem stability[4−6]. Despite their vital ecological importance, fungal biodiversity remains unevenly studied across ecosystems, with particularly limited research focusing on extreme environments such as dry-hot valleys[7−10]. These unique habitats, characterized by arid conditions, severe temperature fluctuations, and limited vegetation[11−13], represent largely unexplored areas where studying fungi could reveal novel insights into biodiversity, ecological roles, and adaptive strategies under conditions typically considered challenging for life[14−16]. Studying fungal diversity in extreme environments is critical, as it deepens our understanding of fungal resilience and adaptability[9,17], revealing how these organisms survive and flourish under severe stress conditions. Such insights are particularly valuable in the context of climate change, providing important lessons on biological adaptation strategies and ecosystem resilience under increasingly challenging conditions. Documenting fungal diversity in dry-hot valleys enhances our global understanding of biodiversity, emphasizing the ecological significance of fungi in environments historically underrepresented in scientific research[14,15].
The majority of mycological research has concentrated on hospitable environments where fungal populations are abundant and diverse[18]. Consequently, dry-hot valleys remain underexplored, despite their potential to harbor fungal species uniquely adapted to extreme aridity and temperature fluctuations[10]. Logistical challenges associated with fieldwork in these remote regions have further contributed to their neglect in biodiversity studies. Addressing this knowledge gap is not only a matter of scientific curiosity but also essential for developing biodiversity conservation strategies encompassing all ecosystem types. Motivated by this necessity, the present study aims to uncover the hidden diversity of microfungi in dry-hot valleys. In southwestern (SW) China, savanna-like vegetation predominantly occurs in deep, hot, and dry valleys formed by several major rivers. Specifically, native seed plants comprising 3,217 species and varieties belonging to 1,038 genera and 163 families have been documented from the savanna-like vegetation in valleys of the three major rivers (Jinshajiang, Yuanjiang, and Nujiang) in SW China[19]. By applying estimation of approximately six fungal species per vascular plant species[20], the fungal diversity in these valleys could potentially exceed 20,000 species. Despite this substantial predicted diversity, studies on microfungi utilizing both morphological and phylogenetic analyses were virtually non-existent in the Yuanjiang (Honghe) valley prior to 2019[21]. However, interest in microfungal research in this region has significantly increased over the last five years, leading to several recent contributions[21−34].
Therefore, this study specifically conducted comprehensive surveys in the dry-hot valleys of the Honghe region, Yunnan Province, to identify fungal taxa using morphological and phylogenetic methods and document their diversity. By illuminating the understudied fungal communities inhabiting these valleys, these findings offer valuable insights into fungal adaptations, enrich global biodiversity inventories, and support conservation strategies. Given global environmental changes, understanding fungal life in these harsh ecosystems has become increasingly pertinent, holding significant implications for the sustainable management and conservation of arid and semi-arid regions worldwide.
Table of contents
Ascomycota R.H. Whittaker
Dothideomycetes O.E. Erikss & Winka
Botryosphaeriales C.L. Schoch, Crous & Shoemaker
Aplosporellaceae Slippers, Boissin & Crous
Aplosporella Speg.
Aplosporella artocarpi Trakun., L. Lombard & Crous
Aplosporella hesperidica Speg.
Dyfrolomycetales K.L. Pang, K.D. Hyde & E.B.G. Jones
Pleurotremataceae Walt. Watson
Longiascospora Wanas., Phookamsak & J.C. Xu, gen. nov.
Longiascospora hongheensis Wanas., L.S. Dissan., Phookamsak & J.C. Xu, sp. nov.
Hysteriales Lindau
Hysteriaceae Chevall.
Rhytidhysteron Speg.
Rhytidhysteron neorufulum Thambug. & K.D. Hyde
Mycosphaerellales (Nannf.) P.F. Cannon
Mucomycosphaerellaceae Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, fam. nov.
Mucomycosphaerella Quaedvl. & Crous
Mucomycosphaerella hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Patellariales D. Hawksw. & O.E. Erikss.
Patellariaceae Corda
Patellaria Fr.
Patellaria microspora Ekanayaka & K.D. Hyde
Pleosporales
Anteagloniaceae K.D. Hyde & A. Mapook
Anteaglonium Mugambi & Huhndorf
Anteaglonium hongheense Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Didymellaceae Gruyter, Aveskamp & Verkley
Gruyteromyces Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, gen. nov.
Gruyteromyces hongheensis Wanas., Phookamsak, & J.C. Xu, sp. nov.
Didymosphaeriaceae Munk
Chromolaenicola Mapook & K.D. Hyde
Chromolaenicola clematidis Phukhams. & K.D. Hyde
Chromolaenicola hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Paraphaeosphaeria O.E. Erikss.
Paraphaeosphaeria jaguarinae Y.P. Tan & R.G. Shivas
Tremateia Kohlm., Volkm.-Kohlm. & O.E. Erikss.
Tremateia guiyangensis J.F. Zhang, J.K. Liu, K.D. Hyde & Z.Y. Liu
Lophiostomataceae Sacc.
Neovaginatispora A. Hashim., K. Hiray. & Kaz. Tanaka
Neovaginatispora fuckelii (Sacc.) A. Hashim., K. Hiray. & Kaz. Tanaka
Pseudocapulatispora Mapook & K.D. Hyde
Pseudocapulatispora hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Pseudocapulatispora Mapook & K.D. Hyde
Pseudocapulatispora longiappendiculata Mapook & K.D. Hyde
Macrodiplodiopsidaceae Voglmayr, Jaklitsch & Crous
Pseudochaetosphaeronema Punith.
Pseudochaetosphaeronema hongheense Wanas., Phookamsak & J.C. Xu, sp. nov.
Pseudochaetosphaeronema punithalingamii Wanas., Phookamsak & J.C. Xu, sp. nov.
Mangifericomitaceae Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, fam. nov.
Mangifericomes E.F. Yang & Tibpromma
Mangifericomes yunnanensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Massarinaceae Munk
Helminthosporium Link
Helminthosporium hongheense Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Parabambusicolaceae Kaz. Tanaka & K. Hiray.
Neomultiseptospora N. Xie, Phookamsak & Hongsanan
Neomultiseptospora hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Neomultiseptospora yunnanensis Phookamsak, Hongsanan & N. Xie
Phaeosphaeriaceae M.E. Barr
Leptospora Rabenh.
Leptospora phraeana Mapook & K.D. Hyde
Murichromolaenicola Mapook & K.D. Hyde
Murichromolaenicola thailandensis Htet, Mapook & K.D. Hyde
Pseudostaurosphaeria Mapook & K.D. Hyde
Pseudostaurosphaeria hongheensis Wanas., Phookamsak & J.C. Xu, sp. nov.
Roussoellaceae J.K. Liu, Phookamsak, D.Q. Dai & K.D. Hyde
Neoroussoella Jian K. Liu, Phookamsak & K.D. Hyde
Neoroussoella bambusae Phookamsak, J.K. Liu & K.D. Hyde
Xenoroussoella Mapook & K.D. Hyde
Xenoroussoella triseptata Mapook & K.D. Hyde
Sulcatisporaceae Kaz. Tanaka & K. Hiray.
Anthosulcatispora Phukhams. & K.D. Hyde
Anthosulcatispora hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Teichosporaceae M.E. Barr
Magnibotryascoma Thambug. & K.D. Hyde
Magnibotryascoma hongheense Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Paulkirkia Wijayaw., Wanas., Tangthir., Camporesi & K.D. Hyde
Paulkirkia hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov.
Torulaceae Corda
Dendryphion Wallr.
Dendryphion hydei J.F. Li, Phookamsak & Jeewon
-
Fresh fungal materials were collected from dead woody litter and twigs of deciduous plants in Honghe (Yunnan Province, China) during both the dry season (March, April, December) and the rainy season (June, August). The local environment was characterized by poor eroded soils, steep valleys (Fig. 1), and a subtropical monsoon climate. The specimens were examined using standard protocols described in Dissanayake et al.[29], Gao et al.[34], and Xu et al.[35]. Single-spore isolations were performed following the approaches of Li et al.[36,37] and Mapook et al.[38], with slight modifications, including overnight incubation at 16–25 °C in darkness. Germinated spores were examined under a Motic SMZ 168 Stereo Zoom microscope, and then transferred to potato dextrose agar [PDA; potato extract 4 g/L (equivalent to 200g of infusion from potatoes), glucose 20 g/L, agar 15 g/L] or malt extract agar (MEA; malt extract 30 g/L, mycological peptone 5 g/L, agar 15 g/L) for further analysis, such as DNA extraction, growth rate measurements and cultural characterization. Isolates with the prefix KUNCC were preserved at the Culture Collection of the Kunming Institute of Botany, Chinese Academy of Sciences. Corresponding voucher specimens were deposited in the Cryptogamic Herbarium of the same institute (KUN-HKAS), Kunming, China. Nomenclatural data for fungal novelties were deposited in the
https://gmsmicrofungi.org [39] and MycoBank[40].
Figure 1.
The local environment showing steep valleys and eroded nutrient-poor soils that characterize the study area.
Morphological classification
-
Digital images of reproductive structures were obtained using a Canon 450D camera mounted on a Nikon ECLIPSE 80i compound microscope. To examine micro-morphological features, squash mounts and hand-cut sections of fruiting bodies were prepared, allowing observation of ascomata/conidiomata and peridial structure. Morphological measurements were taken at the widest points of each structure. Dimensions were recorded using the Tarosoft® Image Framework software, and image plates were compiled using Adobe Photoshop CS3 Extended (version 10.0; Adobe Systems, USA). Each isolate was cultured in triplicate on PDA to assess colony morphology under light conditions at 16–20 °C. Colony dimensions and pigmentation were evaluated following the colour charts of Rayner[41], and mycelial zonation patterns were documented after a three-week incubation period. Descriptions of new taxa conform to the guidelines proposed by Jeewon & Hyde[42] and Maharachchikumbura et al.[43].
DNA extraction, PCR, and sequencing
-
DNA extraction, PCR amplification, and sequencing were carried out using the protocols outlined by Wanasinghe et al.[44], Wijesinghe et al.[45] and Htet et al.[46]. In cases where fungal isolates could not be obtained in culture, DNA was isolated directly from ascomycetous fruiting structures. Approximately 15–20 fruiting bodies (diameter > 500 µm; 10 per sample) were carefully detached from the host material using a sterilized needle and transferred into a drop of sterile water in a 1.5 mL sterile Eppendorf tube under aseptic conditions. Genomic DNA was then extracted using the E.Z.N.A.® Forensic DNA Kit (D3591-01, Omega Bio-Tek) in accordance with the manufacturer's guidelines.
Phylogenetic analyses
-
Sequences generated from this study were compared with homologous sequences retrieved from GenBank. Reference sequences were selected based on BLAST results and recently published studies. Multiple sequence alignments were carried out using MAFFT v.7 via the online platform[47,48], and the alignments were manually adjusted where necessary using BioEdit v.7.0.9[49]. Phylogenetic relationships were inferred from both single-locus and concatenated datasets using Maximum Likelihood (ML) and Bayesian Inference (BI) approaches. For both ML and Bayesian analyses, the best-fitting substitution models for each gene region were selected using MrModeltest v.2.3[50] under the Akaike Information Criterion (AIC), as implemented in PAUP v.4.0b10[51]. Phylogenetic analyses using both Maximum Likelihood (ML) and Bayesian Inference (BI) approaches were conducted via the CIPRES Science Gateway[52]. For ML analyses, RAxML-HPC2 on XSEDE v.8.2.10[53] was employed with default settings and 1,000 bootstrap replicates to assess branch support. Bayesian analyses were carried out using MrBayes on ACCESS[54], implementing the GTR+I+G substitution model. The MCMC algorithm was run for two to five million generations, with trees sampled every 1,000 generations. The run terminated automatically once the average standard deviation of split frequencies fell below 0.01, and the first 25% of trees were discarded as burn-in. ML bootstrap values (MLBS) ≥ 70% and Bayesian posterior probabilities (BYPP) ≥ 0.95 are reported above the corresponding branches in the phylogenetic trees. Final trees were visualized using FigTree v.1.4.0[55] and annotated using either Microsoft PowerPoint (2007) or Adobe Illustrator CS5 (Version 15.0.0, Adobe Systems, San Jose, CA). Accession numbers for all newly generated sequences are provided after "Material examined" section.
-
Comprehensive phylogenetic analyses were conducted across 18 families. Based on the phylogenetic evidence, coupled with morphological support, two new families, two new genera, and 15 new species were introduced and additional information was provided for 15 existing species. Phylogenetic results for each analysis are provided in the figure legends. Detailed discussions of the topologies related to our new strains are presented in the note sections of each respective taxonomic entry below.
Taxonomy
-
Ascomycota R.H. Whittaker
Dothideomycetes O.E. Erikss & Winka
Botryosphaeriales C.L. Schoch, Crous & Shoemaker
Aplosporellaceae Slippers, Boissin & Crous
Aplosporella Speg.
Notes: Aplosporella is a heterogeneous genus comprising over 275 species, typically associated with dead branches and twigs across diverse hosts and habitats[56−59]. Some species within Aplosporella are recognized as pathogens, causing branch and twig blight, canker, or dieback diseases in numerous plant hosts[60−63]. Recent taxonomic revision by Phillips et al.[64] formally synonymized Bagnisiella under Aplosporella. Although Hyde et al.[59] accepted approximately 275 species, phylogenetic positions based on molecular data have been confirmed for only 13 species. In this study, two new host records were reported for Aplosporella artocarpi and A. hesperidica.
Aplosporella artocarpi Trakun., L. Lombard & Crous, Persoonia 34: 91 (Figs 2 and 3).
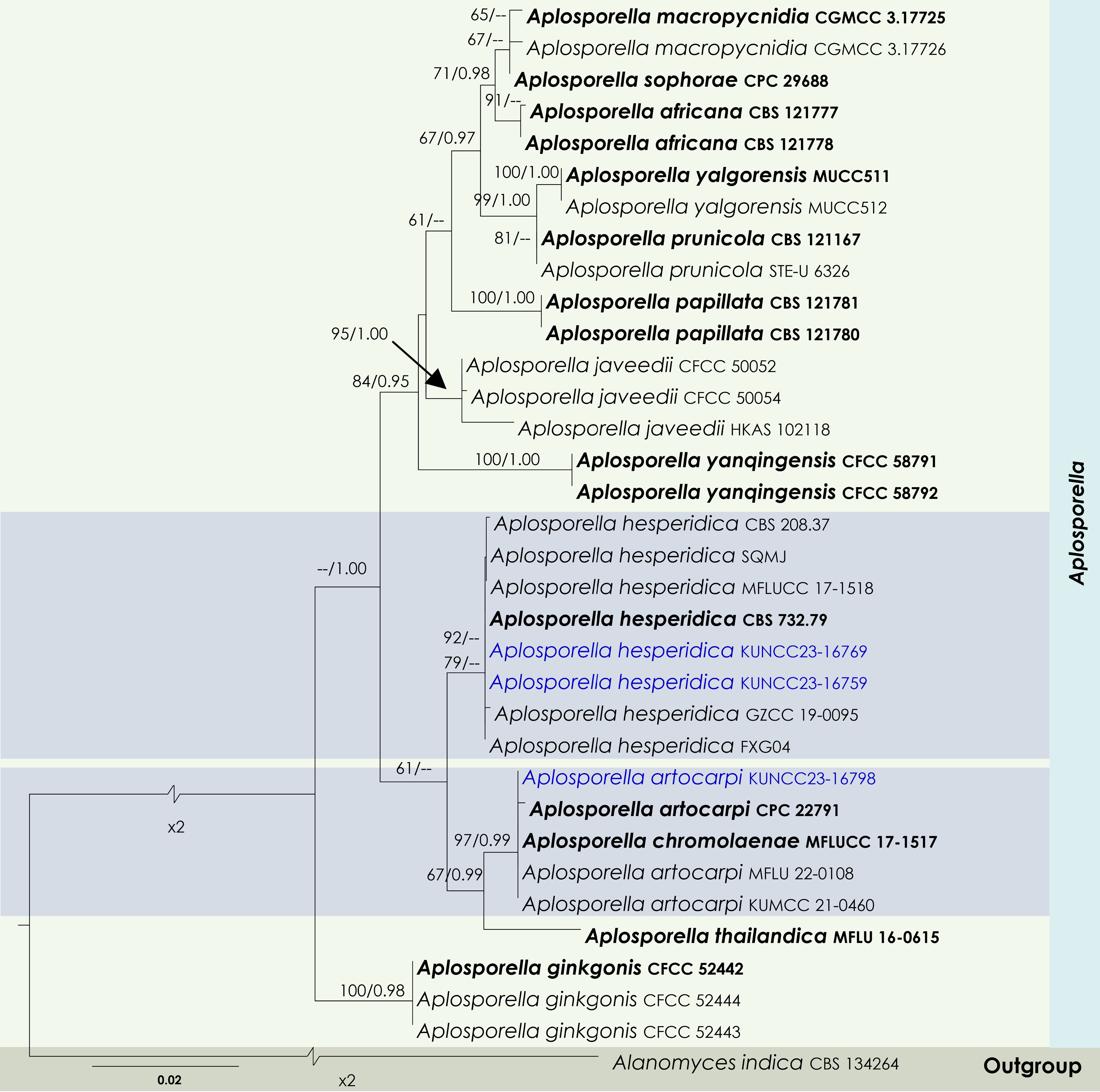
Figure 2.
Maximum Likelihood tree inferred from the concatenated dataset of partial LSU, ITS, and tef1-α sequences representing members of Aplosporellaceae. The phylogeny is rooted with Alanomyces indica (CBS 134264). The final likelihood value is –4,051.526624. The final alignment included 303 unique site patterns, with approximately 25.96% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.230562, C = 0.245552, G = 0.282555, T = 0.241332. The substitution model yielded the following relative rates: AC = 5.894305, AG = 6.724706, AT = 4.532426, CG = 4.107660, CT = 15.165187 and GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.7274, and the gamma distribution shape parameter (α) was 0.6476. Bayesian inference reached convergence after 623,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009918). A total of 3,116 trees were sampled, and 2,337 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 304 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
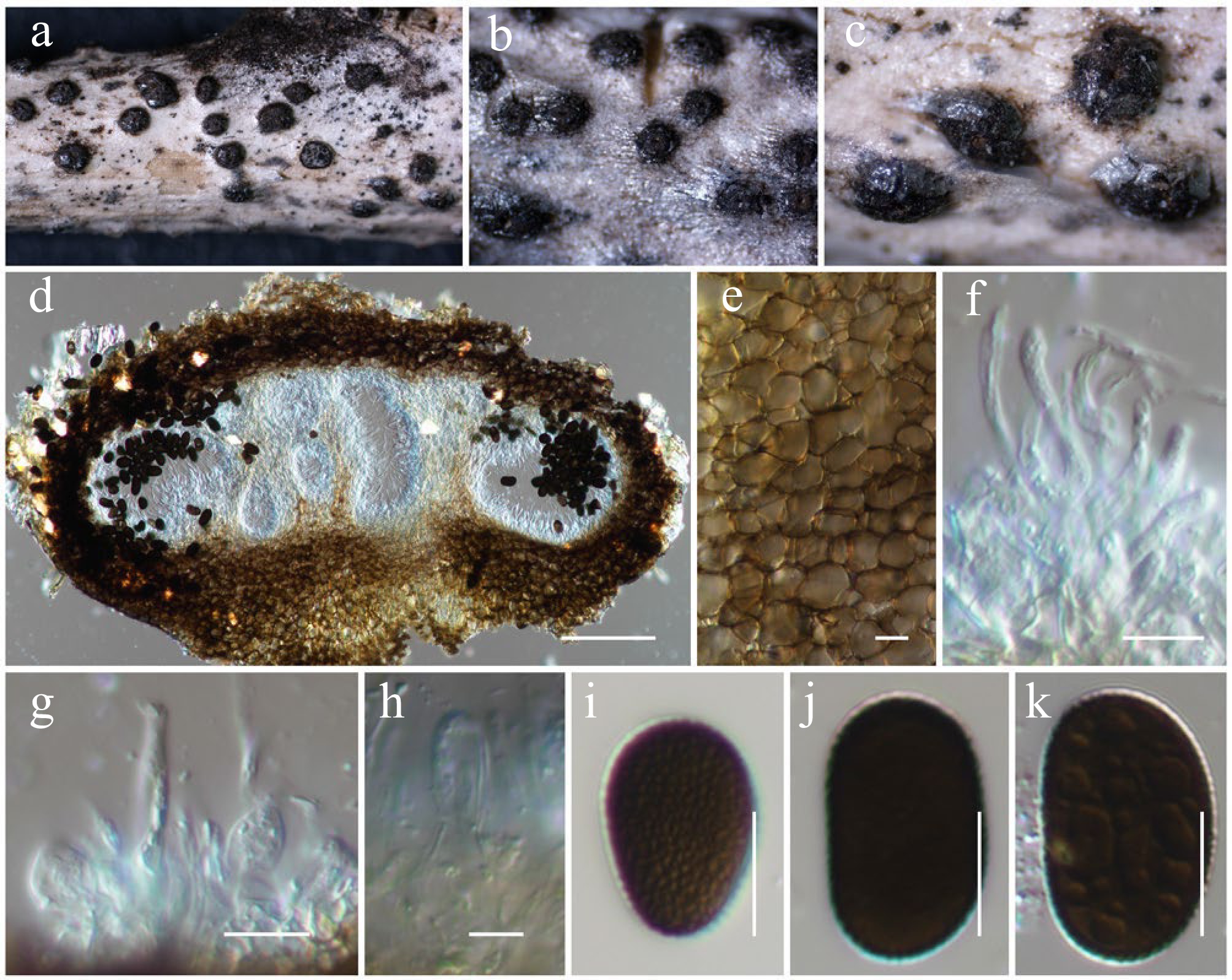
Figure 3.
Aplosporella artocarpi (HKAS146009). (a)–(c) Conidiostromata on host. (d) Cross-section of the conidiostroma. (e) Pycnidial wall. (f) Paraphyses. (g), (h) Conidiogenous cells and developing conidia. (i)–(k) Conidia. Scale bars: (d) = 100 µm, (e)–(k) = 10 µm.
MycoBank No: 810167
Saprobic on a dead twig of Bougainvillea spectabilis. Sexual morph: Undetermined. Asexual morph: Conidiostromata 300–350 μm high, 450–750 μm in diam., pycnidial, semi-immersed, erumpent through host epidermis, multilocular. Disc blackish brown, circular to ovoid, with one central ostiole per disc. Ostioles are inconspicuous, minute papillate, or with pore-like openings. Locules 180–220 μm high, 150–275 μm diam., multi-loculate, globose to ovoid, or irregular in shape, irregularly arranged, with three to five subdivided chambers separated by invaginations with common walls. Pycnidial walls 30–50 μm wide, composed of several cell layers, of large, broad, thick-walled, brown to dark brown pseudoparenchymatous cells, arranged in a textura angularis; interstitial walls of a locule composed of several cell layers of flattened, light brown to brown pseudoparenchymatous cells, arranged in textura prismatica or palisade-like cells. Paraphyses 20–30 × 2–3 μm (
$\overline {\rm x} $ $\overline {\rm x} $ $\overline {\rm x} $ Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421524° N, 102.227973° E, 730 m, on dead twigs of Bougainvillea spectabilis (Nyctaginaceae), 10 June 2021, D.N. Wanasinghe, HH22-07 (HKAS146009), living culture, KUNCC23-16798.
GenBank numbers: KUNCC23-16798: ITS = PV742884, LSU = PV742940, tef1-α = PV738667.
Known hosts and distribution: Asymptomatic twigs of Artocarpus heterophyllus[65], dead stems of Chromolaena odorata[66], and dead woody twigs of Hevea brasiliensis[67] in Thailand; on asymptomatic leaves of Stoechospermum marginatum and Caulerpa taxifolia in India[68]; on dead branches of Mangifera indica[26] and Bougainvillea spectabilis in Yunnan, China (this study).
Notes: Phylogenetic analyses of a combined LSU, ITS and tef1-α DNA sequence demonstrated that the new strain KUNCC23-16798 shares the same branch length with the type strains of Aplosporella artocarpi (CPC 22791) and A. chromolaenae (MFLUCC 17-1517) and other two representative strains of A. artocarpi (MFLU 22-0108 and KUMCC 21-0460) with 97% MLBS and 0.99 BYPP supports (Fig. 2). Morphologically, the new isolate is similar to the type of A. artocarpi (CPC 22791) in having similar size range of conidiostromata, and conidia that are ellipsoid to ovoid, brown to dark brown, reticulate rough-walled when mature[65]. Additionally, the new isolate is also similar to A. chromolaenae (MFLUCC 17-1517) but slightly differs in having smaller conidiostromata (450–750 μm diam. vs 685–780 μm diam.) and larger conidia (18–22 × 10–14 μm vs 13–20 × 8.5–12 μm)[69]. Therefore, the new isolate is identified as Aplosporella artocarpi based on morphological resemblance and phylogenetic evidence. The species is reported on Bougainvillea spectabilis in Yunnan, China, for the first time. Notably, the conspecific status of A. artocarpi and A. chromolaenae is needed to be solved.
Aplosporella hesperidica Speg., Anales de la Sociedad Científica Argentina 13 (1): 18 (Figs 2 and 4).
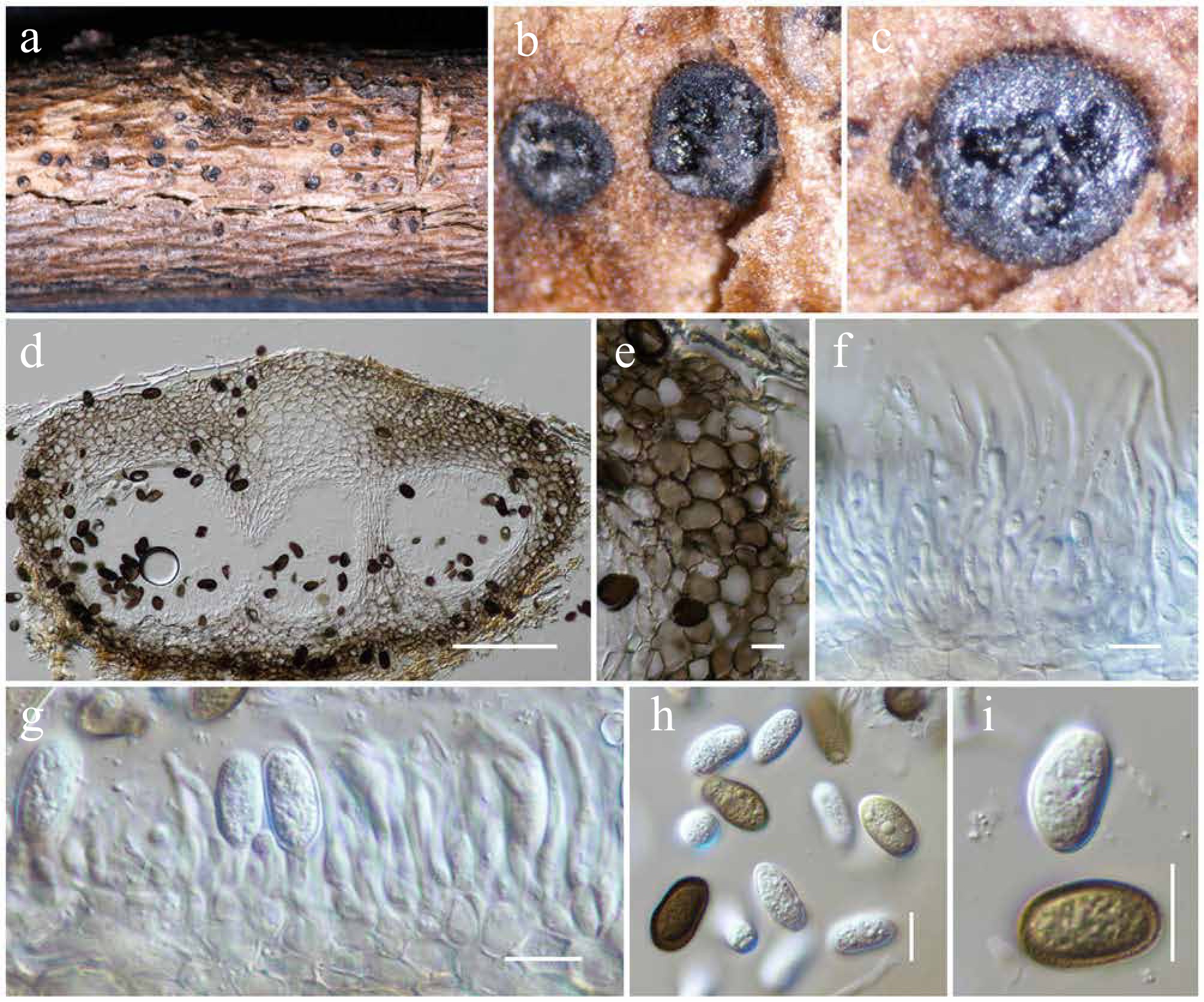
Figure 4.
Aplosporella hesperidica (HKAS146011). (a) Conidiostromata on host. (b)–(d) Cross sections of the conidiostromata. (e) Pycnidial wall. (f) Paraphyses. (g) Conidiogenous cells and developing conidia. (h), (i) Conidia. Scale bars: (d) = 100 µm, (e)–(i) = 10 µm.
MycoBank No: 218239
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Undetermined. Asexual morph: Conidiostromata 400–600 μm in diam., pycnidial, semi-immersed, erumpent through host epidermis, raised, becoming superficial, multilocular, with 3–5 locules per disc. Disc blackish brown, circular to ovoid, with one central ostiole per disc. Ostioles inconspicuous pore-like opening at the same level of the disc surface. Locules 120–180 μm high, 160–240 μm diam., multiple, globose to ovoid, irregularly arranged, with two to three subdivided chambers separated by invaginations with common walls. Pycnidial walls 30–50 μm wide, of unequal thickness, slightly thick at the apex, composed of several cell layers, of large, broad, thick-walled, brown to dark brown pseudoparenchymatous cells, arranged in a textura angularis to textura globulosa; interstitial walls of a locule composed of several cell layers of flattened, hyaline to light brown pseudoparenchymatous cells, arranged in textura prismatica or palisade-like cells. Paraphyses 28–44 × 2–3 μm (
$\overline {\rm x} $ $\overline {\rm x} $ $\overline {\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH13-02 (HKAS146011), living culture, KUNCC23-16769; ibid., 23 April 2020, DWHH08-01 (HKAS146010), living culture, KUNCC23-16759.
GenBank numbers: KUNCC23-16769: ITS = PV742885, LSU = PV742941, SSU = PV742996, tef1-α = PV738668; KUNCC23-16759: ITS = PV742886, LSU = PV742942, SSU = PV742997, tef1-α = PV738669.
Known hosts and distribution (based on molecular data): isolated from soil in Cuba[70]; associated with early stem-end rot on Citrus sinensis in Zimbabwe[71]; on dead stems of Chromolaena odorata in Thailand[69]; associated with cowpea root rot disease in India[72]; on decaying woody host in Guizhou, China[73]; on branches of Euonymus japonicus in Beijing, China[62].
Notes: Phylogenetic analyses of a combined LSU, ITS, and tef1-α DNA sequence (Fig. 2) demonstrated that two new strains (KUNCC23-16769 and KUNCC23-16759) group within the subclade of Aplosporella hesperidica with 79% MLBS support. Morphologically, the new isolates are typical of A. hesperidica in having multilocular, blackish brown, circular to ovoid conidiostromata and aseptate, ellipsoid to broad oblong, brown, rough-walled, reticulate conidia. However, the new isolates differ from the type of A. hesperidica in having smaller conidia (12–18 × 7–9 μm vs 22–25 × 9–11 μm)[74] and the number of locules (3–5 locules vs 3–7 locules of the type)[74]. Based on phylogenetic evidence, the study therefore identified the new isolates as Aplosporella hesperidica herein, and the species is reported from Yunnan, China, for the first time.
Dyfrolomycetales K.L. Pang, K.D. Hyde & E.B.G. Jones, Fungal Diversity 63 (1): 7.
Pleurotremataceae Walt. Watson, New Phytologist 28: 113.
Notes: The family Pleurotremataceae currently comprises three genera, viz. Dyfrolomyces, Melomastia, and Pleurotrema[59]. In this study, a fourth genus is introduced to this family: Longiascospora. Phylogenetically, Longiascospora is nested as the basal lineage within Pleurotremataceae (Fig. 5).
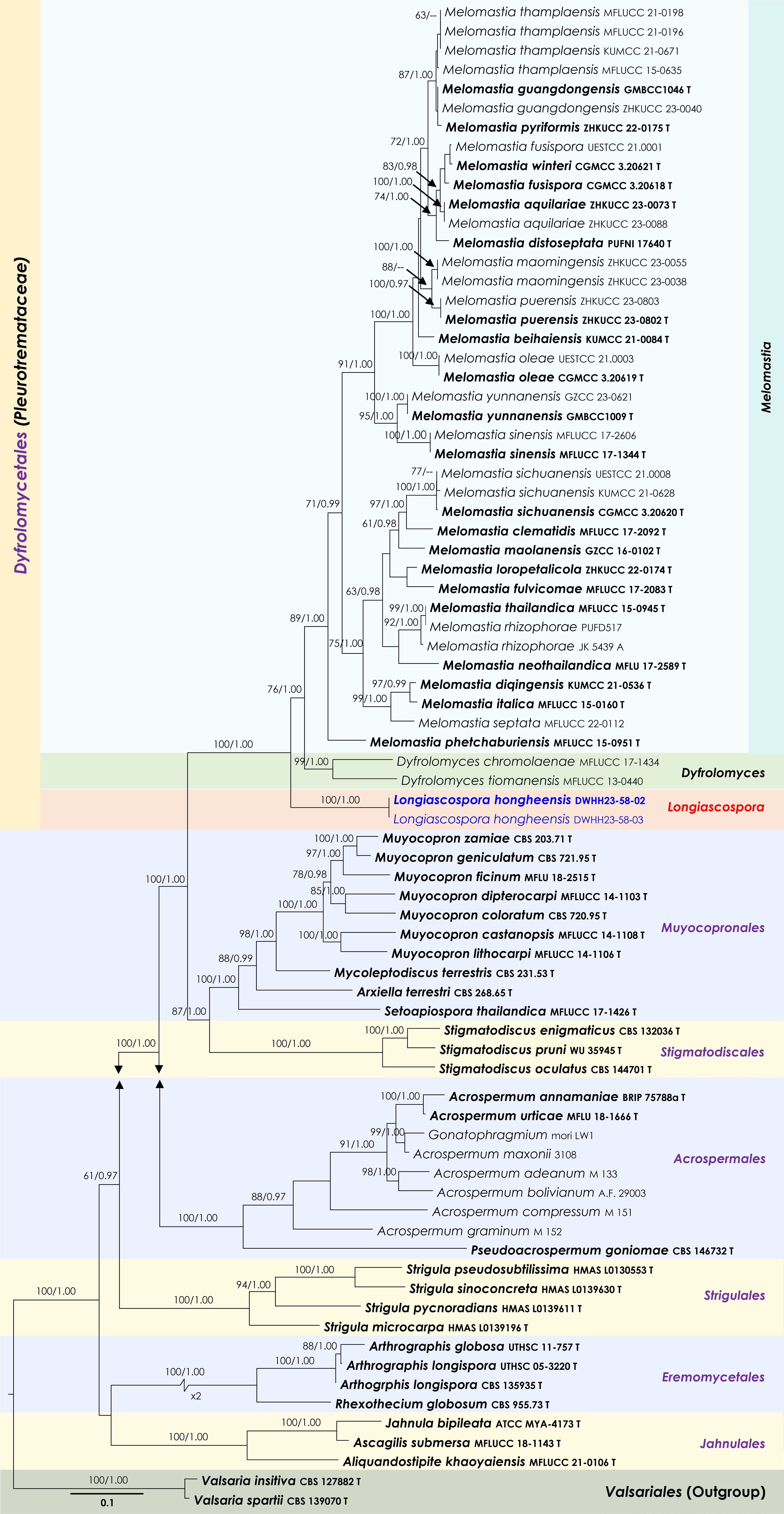
Figure 5.
Maximum Likelihood tree inferred from the concatenated dataset of partial LSU, ITS and rpb2 sequences. The phylogeny is rooted with Valsaria insitiva (CBS 126882) and V. spartii (CBS 139070). The final likelihood value is –48,977.732994. The final alignment included 2,750 unique site patterns, with approximately 44.21% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.236572, C = 0.265016, G = 0.288031, T = 0.21038. The substitution model yielded the following relative rates: AC = 1.574678, AG = 3.286228, AT = 1.515809, CG = 1.482881, CT = 7.546116, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.329292 and the gamma distribution shape parameter (α) was 0.554771. Bayesian inference reached convergence after 336,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.00998). A total of 1,681 trees were sampled, and 1,261 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 2,751 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
Longiascospora Wanas., Phookamsak & J.C. Xu, gen. nov.
MycoBank No: 859350
Etymology: The generic epithet referring to the long ascospores.
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Ascomata solitary, semi-immersed, black, subglobose to ampulliform, glabrous, papillate, ostiolate. Ostiolar neck clypeate, central, elongated, papillate, with periphyses, central, made up of pigmented, pseudoparenchymatous cells. Peridium thick-walled, composed of several celled layers of small, pigmented, pseudoparenchymatous cells of textura angularis, outer layers composed of brown to dark brown cells, fused with host tissues, lighter towards the inside of ascomata. Hamathecium comprises numerous filamentous, unbranched, aseptate, trabeculate pseudoparaphyses observed between and above the asci, and embedded in a gelatinous matrix. Asci 8-spored, bitunicate, cylindrical, short-pedicellate, with rounded apex. Ascospores multi-seriate, tightly fasciculate, straight or slightly curved, filiform, multi-septate, without constrictions at the septa, rounded at both ends, apical end broader than the basal end, hyaline, smooth-walled, guttulate in most cells. Asexual morph: Undetermined.
Type species: Longiascospora hongheensis Wanas., Phookamsak & J.C. Xu
Notes: Longiascospora is introduced as a monotypic genus herein to accommodate Longiascospora hongheensis sp. nov. This new genus can be distinguished from the other three genera in Pleurotremataceae by its spore arrangement in asci and filiform ascospores. Whereas Dyfrolomyces, Melomastia, and Pleurotrema have ellipsoid to fusiform, or elongate fusiform ascospores[75−77]. Phylogenetic analyses of a combined LSU, ITS, and rpb2 DNA sequence demonstrated that the new genus formed an independent monophyletic clade basal to Dyfrolomyces and Melomastia with 100% MLBS and 1.00 BYPP support values (Fig. 5). Therefore, the new genus is introduced based on morphological distinctiveness and phylogenetic evidence.
Longiascospora hongheensis Wanas., L.S. Dissan., Phookamsak & J.C. Xu, sp. nov. (Figs 5 and 6)
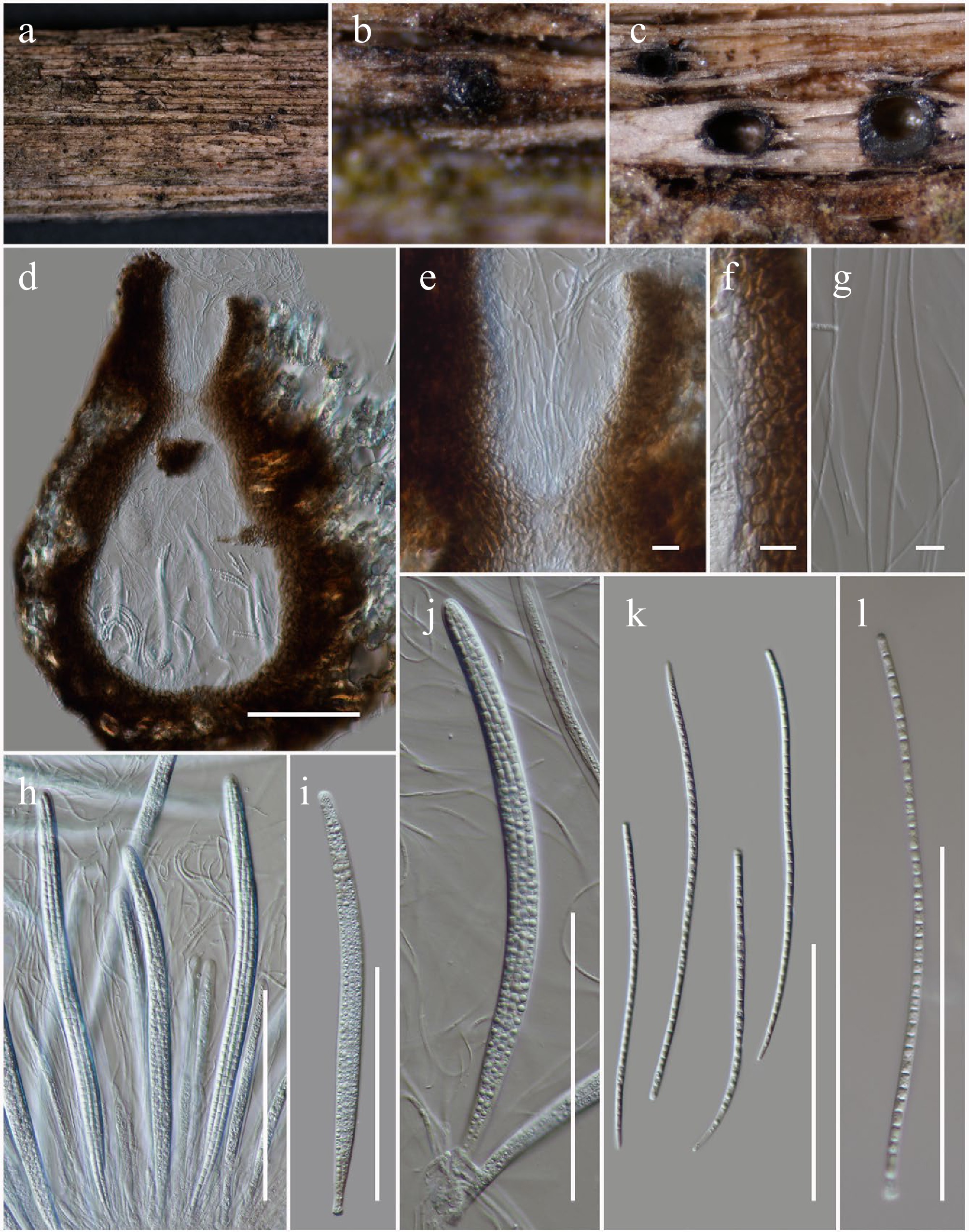
Figure 6.
Longiascospora hongheensis (HKAS146012, holotype). (a), (b) Ascomata on dead woody twigs. (c), (d) Sections of ascomata. (e) Close up of ostiole. (f) Peridium. (g) Pseudoparaphyses. (h)–(j) Asci. (k), (l) Ascospores. Scale bars: (d), (h)–(l) = 100 µm, (e)–(g) = 10 µm.
MycoBank No: 859351
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146012
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Ascomata 370–520 μm high × 290–400 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068 N, 102.229128 E, 735 m, on dead twigs of an unknown deciduous host, 22 April 2020, D.N. Wanasinghe, DWHH23-58-02 (holotype, HKAS146012); ibid., 23 April 2020, DWHH23-58-03 (HKAS146013).
GenBank numbers: HKAS146012: ITS = PV742887, LSU = PV742943, SSU = PV742998, tef1-α = PV700626, rpb2 = PV700675; HKAS146013: ITS = PV742888, LSU = PV742944, SSU = PV742999, tef1-α = PV700627, rpb2 = PV700676.
Notes: The ascomata, peridium, pseudoparaphyses, and asci characteristics of this species resemble those found in Pleurotremataceae. However, the ascospore features are unique, as they are filiform. Phylogenetically, this species constitutes a basal lineage within the family, forming a distinct clade, which warrants its placement in a new genus.
Hysteriales Lindau, Die Natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren Arten 1 (1): 265.
Hysteriaceae Chevall., Flore Générale des Environs de Paris 1: 432.
Rhytidhysteron Speg., Anales de la Sociedad Científica Argentina 12 (4): 188.
Notes: Rhytidhysteron is commonly known as a saprobic fungi that occur on a wide range of hosts in terrestrial and marine habitats[78−81]. However, some species have been reported as endophytes and weak pathogens on woody plants as well as a human pathogen causing subcutaneous phaeohyphomycosis in immunocompetent patients in tropical regions[79−86]. The genus was introduced by Spegazzini[87], and later Rhytidhysteron brasiliense was designated as the type species by Clements and Shear[88]. Currently, 43 species epithets are listed in Index Fungorum[89]; however, Senwanna et al.[81]synonymized R. erioi under R. bruguierae and R. mengziense under R. ligustrum. Therefore, only 41 species are accepted in Rhytidhysteron, of which only about 24 species could be clarified in their phylogenetic placement in Rhytidhysteron. In the present study, the new host record for R. neorufulum was reported on dead twigs of an unknown deciduous host in Yunnan, China.
Rhytidhysteron neorufulum Thambug. & K.D. Hyde, Cryptog. Mycol. 37 (1): 110 (Figs 7 and 8).
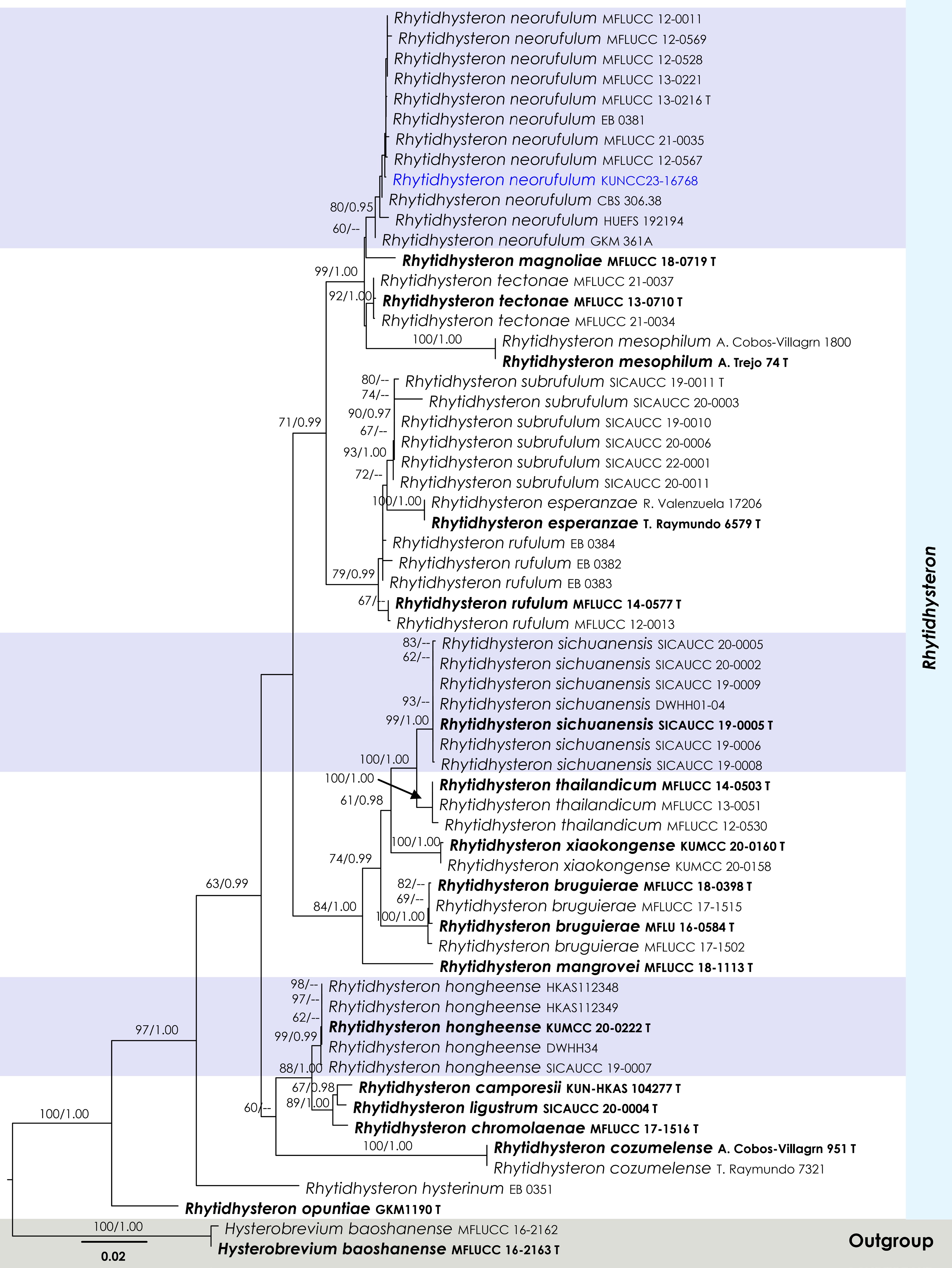
Figure 7.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, and tef1-α sequences. The phylogeny is rooted with Hysterobrevium baoshanense (MFLUCC 16-2162, MFLUCC 16-2163). The final likelihood value is –10,916.669025. The final alignment included 813 unique site patterns, with approximately 23.31% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.240255, C = 0.247157, G = 0.276188, T = 0.236399. The substitution model yielded the following relative rates: AC = 1.376740, AG = 2.437005, AT = 1.147691, CG = 0.904034, CT = 5.839891, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.618399, and the gamma distribution shape parameter (α) was 0.641873. Bayesian inference reached convergence after 761,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009903). A total of 7,611 trees were sampled, and 5,709 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 814 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
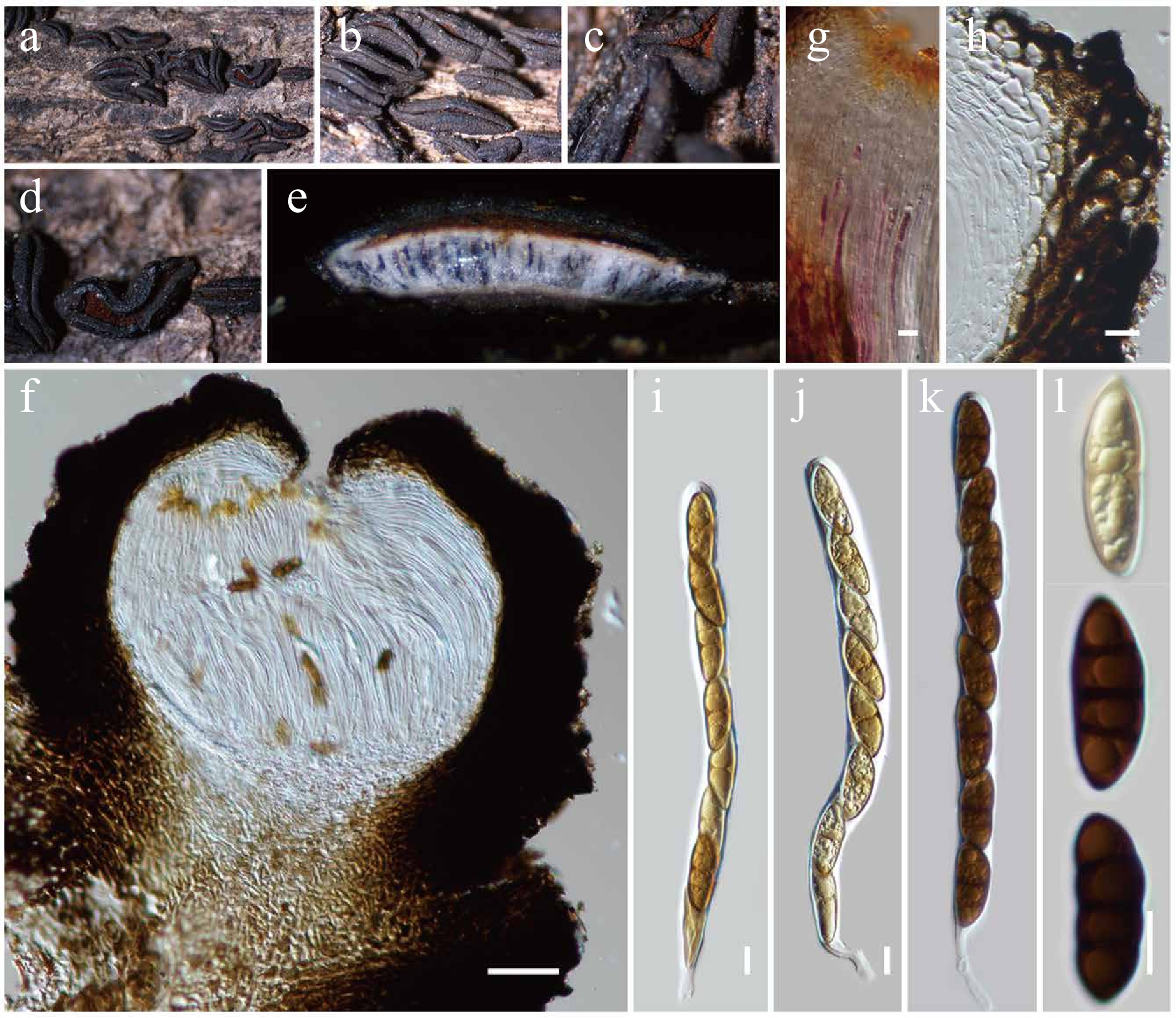
Figure 8.
Rhytidhysteron neorufulum (HKAS146014). (a)–(d) Appearance of hysterothecia on the dead woody twigs. (e) Horizontal section of hysteriothecium. (f) Cross-section of an ascoma. (g) Pseudoparaphyses. (h) Excipulum. (i)–(k) Asci. (l) Ascospores. Scale bars: (f) = 100 µm, (g)–(l) = 10 µm.
MycoBank No: 551865
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Ascomata 1,000–2,000 µm long × 300–400 high × 350–450 µm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, Dayangjiexiang, 23.390025° N, 102.225942° E, 1,186 m, on dead twigs of an unknown deciduous host, 13 March 2023, D.N. Wanasinghe, DWHH11 (HKAS146014), living culture, KUNCC23-16768.
GenBank numbers: KUNCC23-16768: ITS = PV742889, LSU = PV742945, SSU = PV743000, tef1-α = PV700628.
Known hosts and distribution: Saprobic on undetermined dead stem[83]; Hevea brasiliensis[90,91], Tectona grandis[92], Magnolia sp.[93], and Rosa damascena[81] in Thailand; on undetermined hosts in Brazil[94], Europe, and Ghana[95]; on Bursera sp. in Mexico[96]; on decaying wood of Elaeagnus sarmentosa[80], Magnolia sp.[93], and an unknown deciduous host (this study) in Yunnan, China.
Notes: Multigene phylogenetic analyses demonstrated that the new strain KUNCC23-16768 clustered within the monophyletic subclade of Rhytidhysteron neorufulum with 80% MLBS and 0.95 BYPP support (Fig. 7) and is closely related to R. neorufulum (MFLUCC 12-0567). The new isolate (KUNCC23-16768) resembles the type of R. neorufulum in having hysterothecial, navicular to lenticular or irregular in shape, with longitudinally slit, cylindrical asci and brown to dark brown, 3-septate ascospores. However, the new isolate has slightly longer ascomata (1,000–2,000 µm long vs 660–1,415 μm long), larger asci (190–230 × 12.5–16 μm vs 185–220 × 9.5–13 µm), and larger ascospores (30–36 × 10–13 μm vs 27–34 × (6.5–)7–10.6(–12.5))[83].
Mycosphaerellales (Nannf.) P.F. Cannon, Ainsworth & Bisby's Dictionary of the fungi, Ed. 9: X.
Mucomycosphaerellaceae Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, fam. nov.
MycoBank No: 859352
Etymology: Referring to the name of the type genus
Saprobic on leaves and twigs. Sexual morph: Ascomata pseudothecial, solitary or in clusters of two or more joined together, immersed to erumpent, brown to dark brown, globose, ellipsoid to greatly elongated, or depressed ellipsoidal uni- to multi-locular, ostiolate, lacking periphyses. Peridium thin- to thick-walled of unequal thickness, thicker at the side towards the apex, composed of several layers of brown cells of textura angularis, to textura globulosa or textura epidermoidea, poorly developed at the base. Hamathecium lacking pseudoparaphyses; if present, composed of sparse, branched, and anastomosing septate pseudoparaphyses. Asci 8-spored, bitunicate, fissitunicate, fasciculate, ellipsoidal to ovoid, or ampulliform, apedicellate, or with a short pedicel, apically rounded with distinct ocular chamber. Ascospores overlapping multi-seriate, hyaline, oblong to narrowly fusiform, or elongate ellipsoidal, sometimes inequilateral, rounded apex and acute at the bottom, septate, constricted at the septa, deeply constricted at the central septum, rough-walled, guttulate, surrounded by a thick mucilaginous sheath. Asexual morph: Undetermined.
Type genus: Mucomycosphaerella Quaedvl. & Crous.
Notes: Quaedvlieg et al.[97] introduced Mucomycosphaerella to accommodate M. eurypotami (≡Mycosphaerella eurypotami). In this study, Mucomycosphaerellaceae is proposed as a novel family within Mycosphaerellales to accommodate Mucomycosphaerella. The genus Mucomycosphaerella is distinguished from Mycosphaerella sensu stricto by the presence of persistent mucoid sheaths surrounding the ascospores and the absence of Ramularia asexual states. Mucomycosphaerella ascomata are typically depressed with a pale, thin-walled lower half and the hamathecium consists of loosely branched, anastomosing hyphae embedded in a hymenial gel. Ascospores are hyaline. Mucomycosphaerella eurypotami was provisionally included in Mycosphaerella pending a formal genus revision, a process initiated with the epitypification of the type species[98] and subsequent segregation of allied genera and families[99,100]. Quaedvlieg et al.[97] suggested Mucomycosphaerella represents a distinct family within Capnodiales, closely related to Schizothyriaceae. However, with the addition of another species to Mucomycosphaerella, phylogenetic analyses have now placed it outside Capnodiales and instead within Mycosphaerellales, allied closely with Aeminiaceae, Cystocoleaceae, Extremaceae, and Neodevriesiaceae. Aeminiaceae, established by Trovão et al.[101], includes the monotypic genus Aeminium, known only from its hyphomycetous asexual morph characterized by dark brown, thick-walled, septate conidia forming in meristematic chains. Cystocoleaceae accommodates the lichenized genus Cystocoleus, distinguished by its dense filamentous thallus with Trentepohlia photobionts and a unique hyphal sheath composed of jigsaw puzzle-shaped cells[102]. Extremaceae was introduced by Quaedvlieg et al.[97] to accommodate lichenicolous or yeast-like taxa, with Extremus designated as the type genus. The family is characterized by solitary to sporodochial conidiophores that proliferate sympodially or possess a terminal rachis that may be subdenticulate; conidia are brown, solitary or in short, mostly unbranched chains, subcylindrical to narrowly fusoid-ellipsoidal or obclavate, rarely with 1–2 transverse septa, often surrounded by a mucoid sheath, and exhibit non- to slightly darkened hila. Neodevriesiaceae was also introduced by Quaedvlieg et al.[97], with Neodevriesia as the type genus. This family is known for having both sexual and asexual morphs and is characterized in the sexual morph by black, immersed, substromatic ascomata; 8-spored, bitunicate, subsessile, obovoid to broadly ellipsoid asci that are aparaphysate; and hyaline, thick-walled, fusoid-ellipsoidal, 1-septate ascospores. The asexual morph features pigmented, sympodially proliferating conidiophores, and brown, solitary or short unbranched chains of subcylindrical to narrowly fusoid-ellipsoidal or obclavate conidia, which are rarely septate. Morphologically, Mucomycosphaerella resembles taxa in Neodevriesiaceae in having solitary or stromatic ascomata, ellipsoidal to ovoid asci, and hyaline, fusoid-ellipsoidal, septate ascospores. However, the ascospores of Mucomycosphaerella are often covered by a thick mucilaginous sheath, a feature absent in Neodevriesiaceae. Previously, Mucomycosphaerella was treated as a genus incertae sedis within Mycosphaerellales[59]. Rather than leaving it orphaned, Mucomycosphaerellaceae fam. nov. was formally introduced to accommodate Mucomycosphaerella. Phylogenetic analyses confirm Mucomycosphaerellaceae as a monophyletic lineage basal to Aeminiaceae (Fig. 9).
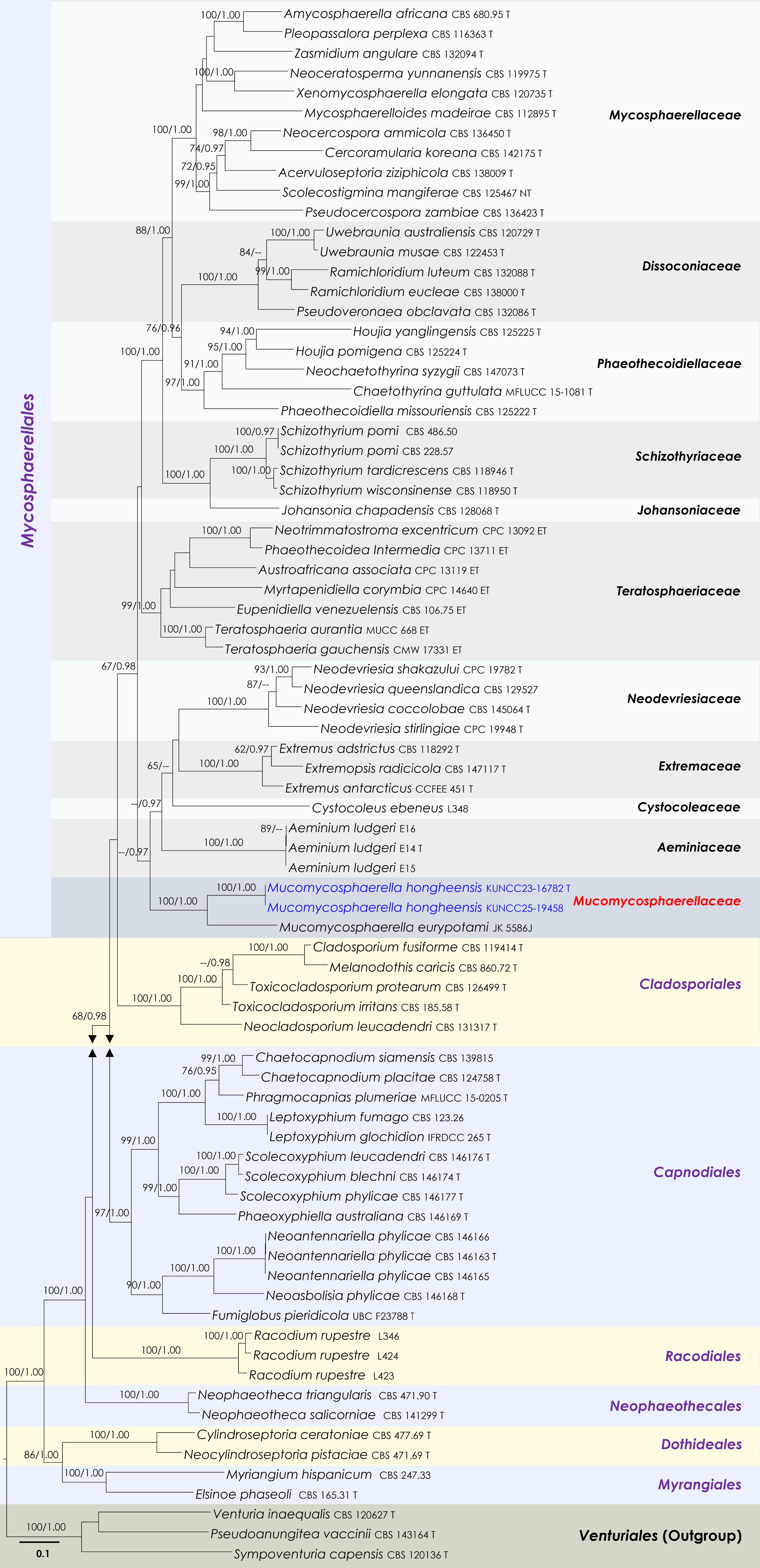
Figure 9.
Maximum Likelihood tree inferred from the concatenated dataset of partial LSU, ITS and rpb2 sequences. The phylogeny is rooted with Pseudoanungitea vaccinii (CBS 143164), Sympoventuria capensis (CBS 120136) and Venturia inaequalis (CBS 120627). The final likelihood value is –46,883.414087. The final alignment included 1,712 unique site patterns, with approximately 28.99% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.244142, C = 0.249865, G = 0.288498, T = 0.217496. The substitution model yielded the following relative rates: AC = 2.074748, AG = 3.272618, AT = 1.73051, CG = 1.485905, CT = 7.286184, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.36136 and the gamma distribution shape parameter (α) was 0.702452. Bayesian inference reached convergence after 467,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009979). A total of 2,336 trees were sampled, and 1,752 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,714 distinct informative sites. Newly generated sequences are shown in blue. Ex-type and type strains are indicated with a 'T' at the end of the strain number.
Mucomycosphaerella Quaedvl. & Crous, Persoonia 33: 22.
Notes: Mucomycosphaerella has been a monotypic genus for the past decade, and in this study, the second species is introduced to the genus from Yunnan, China. Phylogenetically, the generic clade forms an independent lineage within Mycosphaerellales, suggesting that it warrants recognition as a separate family.
Mucomycosphaerella hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 9 and 10)
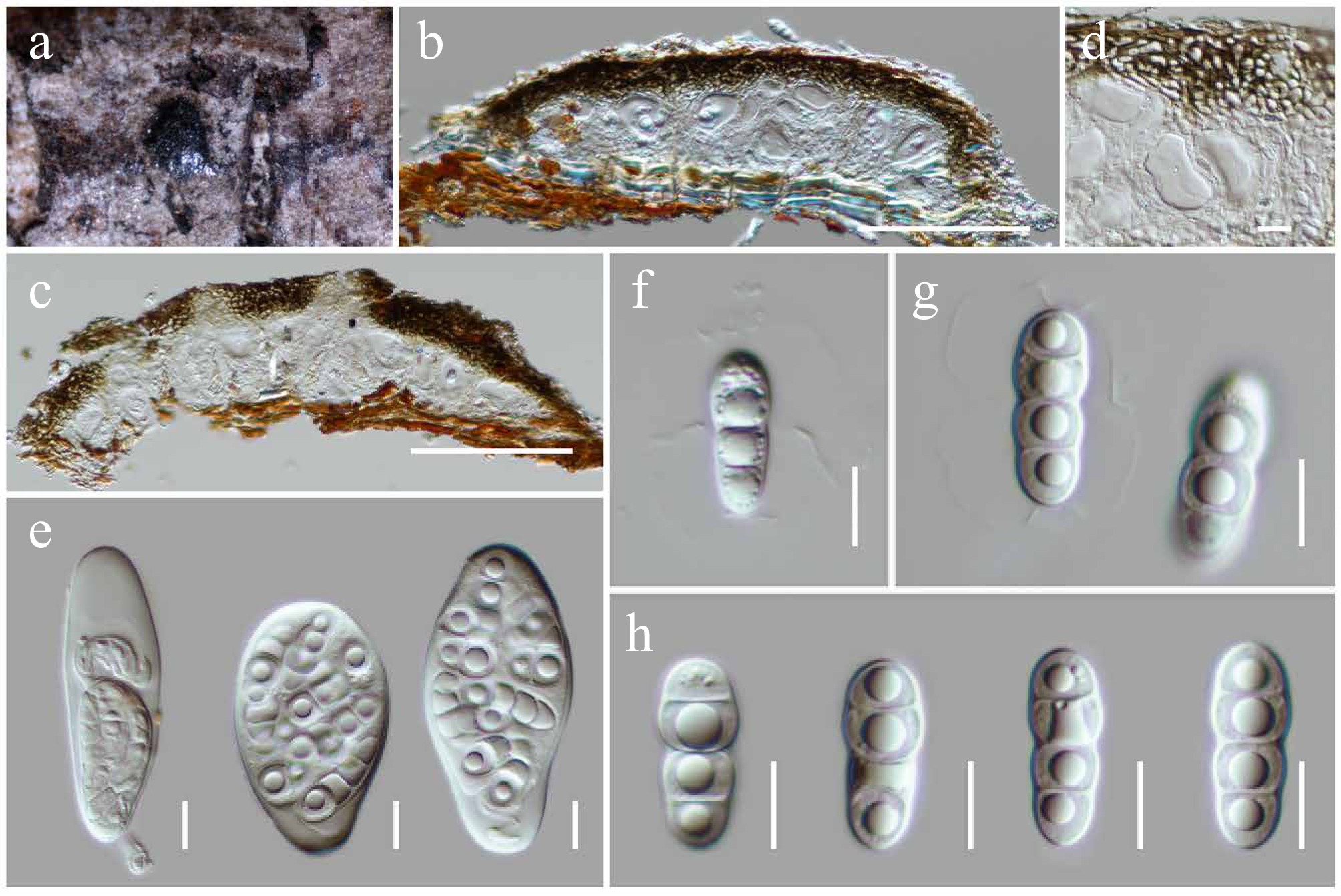
Figure 10.
Mucomycosphaerella hongheensis (HKAS146015, holotype). (a) Ascomata on the dead woody twigs. (b), (c) Cross-section of ascomata. (d) Peridium. (e) Asci. (f)–(h) Ascospores. Scale bars: (b), (c) = 100 µm, (d)–(h) = 10 µm.
MycoBank No: 859353
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146015
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Ascomata 300–400 μm high × 70–100 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies growing fast on PDA at room temperature (25 °C) under normal light, reaching 55 mm diam. after two weeks. Colonies dense at the center, sparse at the margin, circular, flat, slightly raised, with convex edge separating the margin, surface smooth, with edge entire, cottony at the center, feathery at the margin, smooth aspect, colonies white from above and below, do not produce pigmentation in PDA.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead twigs of an unknown deciduous host, 22 April 2020, D.N. Wanasinghe, DWHH19-02 (holotype, HKAS146015), ex-type, KUNCC23-16782; ibid., 23 April 2020, DWHH19-02-01 (paratype, HKAS146016), ex-paratype, KUNCC25-19458.
GenBank numbers: KUNCC23-16782: ITS = PV742890, LSU = PV742946, rpb2 = PV738665, ACT = PV738670, CAL = PV738674; HKAS146016: ITS = PV742891, LSU = PV742947, rpb2 = PV738666, ACT = PV738671, CAL = PV738675.
Notes: Phylogenetic analyses of a combined LSU, ITS and rpb2 sequence data demonstrated that two strains of Mucomycosphaerella hongheensis (KUNCC23-16782 and KUNCC25-19458) formed a robust subclade and clustered with M. eurypotami (JK 5586J) with 100% MLBS and 1.00 BYPP support (Fig. 9). Mucomycosphaerella hongheensis can be distinguished from M. eurypotami in having larger ascomata (300–400 μm high × 70–100 μm diam. vs 115–130 μm high, 190–300 μm diam.), lacking of pseudoparaphyses, and shorter and broader ascospores (18–25 μm × 6.5–9.5 μm vs 23–29 × 5.5–6.5 μm)[103] with (2–)3-euseptate, covered by irregular mucilaginous sheath. In contrast, ascospores of M. eurypotami are 1-euseptate, with one additional pseudoseptum in each cell, surrounded by a rounded mucilaginous sheath. Therefore, the new species, M. hongheensis, is introduced herein based on morphological distinctiveness and phylogenetic evidence.
Patellariales D. Hawksw. & O.E. Erikss., Systema Ascomycetum 5: 181.
Patellariaceae Corda, Icones fungorum hucusque cognitorum 2: 37.
Patellaria Fr., Systema Mycologicum 2 (1): 158.
Notes: Patellaria is commonly known as sabprobe on decaying wood, stems, or bark in terrestrial and marine habitats[79,104]. The genus was introduced by Fries[105] with P. atrata as the type species and is characterized by apothecial, superficial, black, circular, flattened, sessile ascomata, with a carbonaceous rim, exposing the dark hymenium at the center; exciple composed of blue-black (hypothecium) or green-blue to colourless pseudoparenchymatous cells; 8-spored, bitunicate, fissitunicate, cylindrical to clavate, short pedicelate asci, embedded in septate, branched paraphyses, with slightly swollen and rounded apex, forming a dark and thick epithecium over the asci; and clavate to fusiform, or allantoid, hyaline, septate ascospores[75,104,106]. Presently, more than 1,400 species epithets are listed in Index Fungorum[89]. However, only about 50 morpho-species were accepted by Hyde et al.[59], of which 15 species have molecular data available in GenBank[104]. In this study, Patellaria microspora is reported on dead twigs of an unknown deciduous host in Yunnan, China, for the first time.
Patellaria microspora Ekanayaka & K.D. Hyde, Fungal Diversity 105: 131 (2020) (Figs 11 and 12).
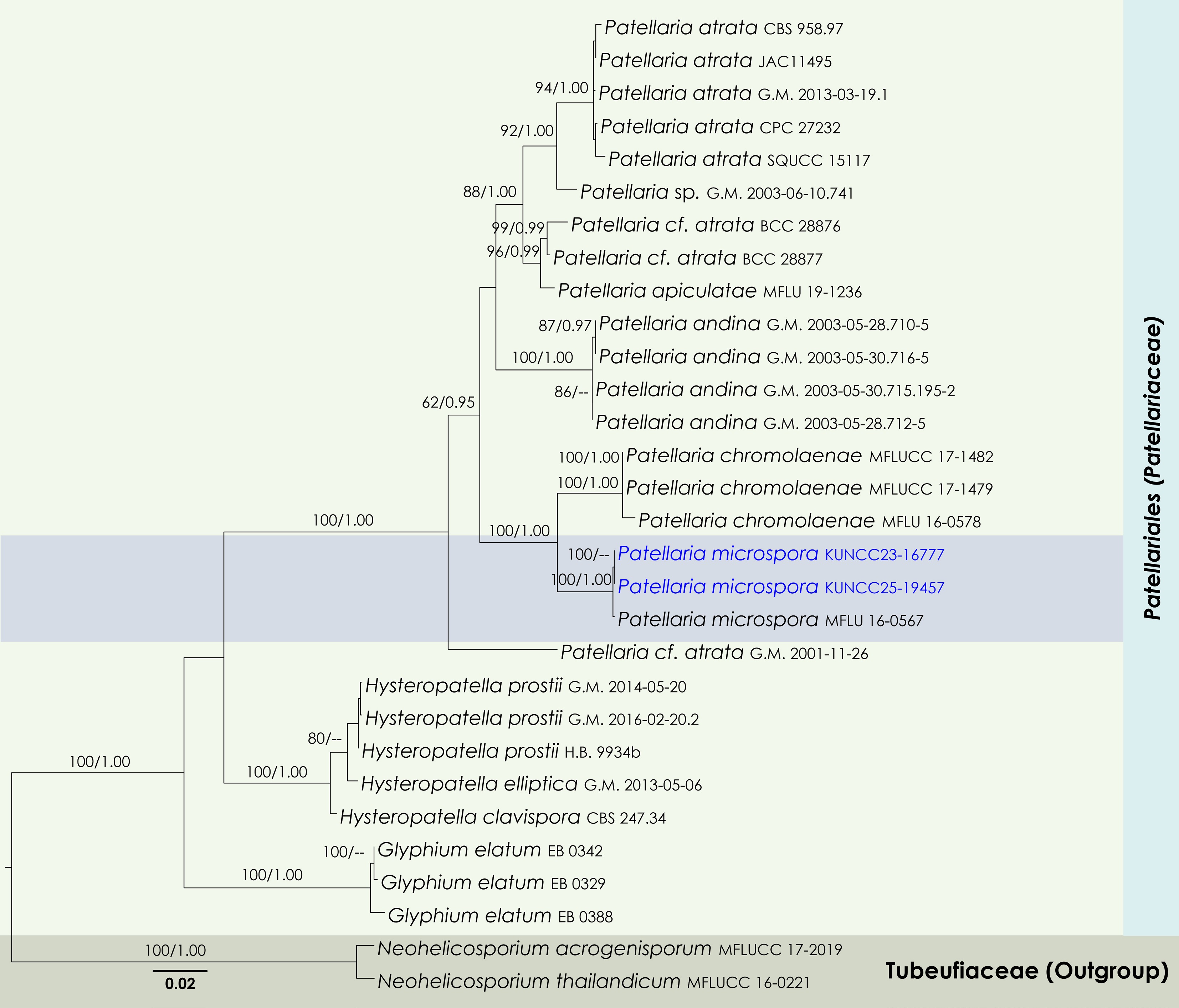
Figure 11.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, and tef1-α sequences. The phylogeny is rooted with Neohelicosporium acrogenisporum (MFLUCC 17-2019) and N. thailandicum (MFLUCC 16-0221). The final likelihood value is –9911.771441. The final alignment included 749 unique site patterns, with approximately 42.67% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.245832, C = 0.243333, G = 0.273432, T = 0.237402. The substitution model yielded the following relative rates: AC = 2.866237, AG = 3.728289, AT = 2.673312, CG = 1.344539, CT = 15.427322, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.580820, and the gamma distribution shape parameter (α) was 0.704216. Bayesian inference reached convergence after 176,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009992). A total of 881 trees were sampled, and 661 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 754 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue.

Figure 12.
Patellaria microspora (HKAS146017). (a), (b) Apothecia on host substrate. (c) Cross-section of an apothecial ascoma. (d) Exciple. (e) Paraphyses. (f) Branched paraphyses, with swollen and rounded at the apex. (g), (h) Asci. (i)–(l) Ascospores. Scale bars: (c) = 100 µm, (d)–(h) = 10 µm.
MycoBank No: 557819
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Ascomata 160–220 high × 550–700 diam. μm (
$\overline {\rm x} $ $\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Colonies grow slowly on PDA at room temperature (25 °C) under normal light, reaching 30–35 mm diam. after four weeks. Colonies dense, circular, raised, slightly umbonate, surface smooth, with edge entire, cottony, smooth aspect, do not produce pigmentation in PDA. Colonies from above grey to greenish grey, with darker greenish grey radiating near the margin, separated the margin from the center; colonies from below dark grey to black at the margin and paler greenish black towards the center.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH18-01 (HKAS146017), living culture, KUNCC23-16777; ibid., DWHH18-01-2 (HKAS146018), living culture, KUNCC25-19457.
GenBank numbers: KUNCC23-16777: ITS = PV742892, LSU = PV742948, SSU = PV743001, tef1-α = PV700629; HKAS146018: ITS = PV742893, LSU = PV742949, SSU = PV743002, tef1-α = PV700630.
Known hosts and distribution: Saprobic on dead stems of unidentified plants in the UK[75]; on dead twigs of an unknown deciduous host in Yunnan, China (this study).
Notes: Phylogenetically, two new strains (KUNCC23-16777 and KUNCC25-19457) formed a subclade (100% MLBS, 1.00 BYPP; Fig. 11) with the type of Patellaria microspora (MFLU 16-0567) and clustered with P. chromolaenae (MFLUCC 17-1482, MFLUCC 17-1479 and MFLU 16-0567) with 100% MLBS and 1.00 BYPP support values (Fig. 11). Morphologically, the new isolates are similar to the type of P. microspora (MFLU 16-0567) in having apothecial, sessile ascomata with turbinate, convex disc of receptacle, cylindric-clavate asci and hyaline, cylindric-clavate, smooth-walled, 7–9-septate ascospores. However, there are some dimensional differences observed in the apothecia (550–700 μm diam. vs 396–402 μm diam.) and ascospores (35–50 × 9–11 μm vs 19.2–20.3 × 8.7–9.5 μm)[75]. These variations may have arisen as a result of the species adapting to distinctly different ecological zones. It is also noteworthy that the ascus measurements provided by Hongsanan et al.[75] appear to be inconsistent with the scale bars shown in their Fig. 65, suggesting a potential error in the original description. Nucleotide comparison between Patellaria microspora (MFLU 16-0567) and our new strains revealed only a two-base pair difference in the ITS region, while other regions were identical.
Pleosporales Luttr. ex M.E. Barr, Prodromus to class Loculoascomycetes: 67
Anteagloniaceae K.D. Hyde & A. Mapook, Fungal Diversity 63 (1): 33
Anteaglonium Mugambi & Huhndorf, Systematics and Biodiversity 7 (4): 460
Notes: Anteaglonium is commonly known as saprobes and endophytes on barks, branches, leaves, twigs, and wood of a wide range of shrubs and tree plants, as well as inhabiting rock[107−112]. Anteaglonium was introduced by Mugambi & Huhndorf[113], and is typified by A. abbreviatum. The genus is known for both sexual and asexual morphs that are characterized by hysterothecial, superficial, carbonaceous ascomata, with slit-like opening ostiolar, 8-spored, bitunicate, fissitunicate, elongate, cylindric-clavate to cylindrical asci, with short pedicellate, and hyaline, fusiform to oblong, 1-septate ascospores[110]. Coelomycetous asexual morph of Anteaglonium is characterized by pycnidial, subglobose to globose, superficial to subperidermal, uni- to multi-loculate conidiomata, cylindrical, unbranched conidiophores, phialidic, globose, smooth-walled conidiogenous cells, and hyaline, oblong to ellipsoidal or oval, aseptate, smooth-walled conidia[110,112]. Presently, there are 12 species accepted in this genus. In this study, the new species, Anteaglonium hongheense, is introduced on dead twigs of Quercus sp. in Yunnan, China based on morphological characteristics and phylogenetic evidence.
Anteaglonium hongheense Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 13 and 14).
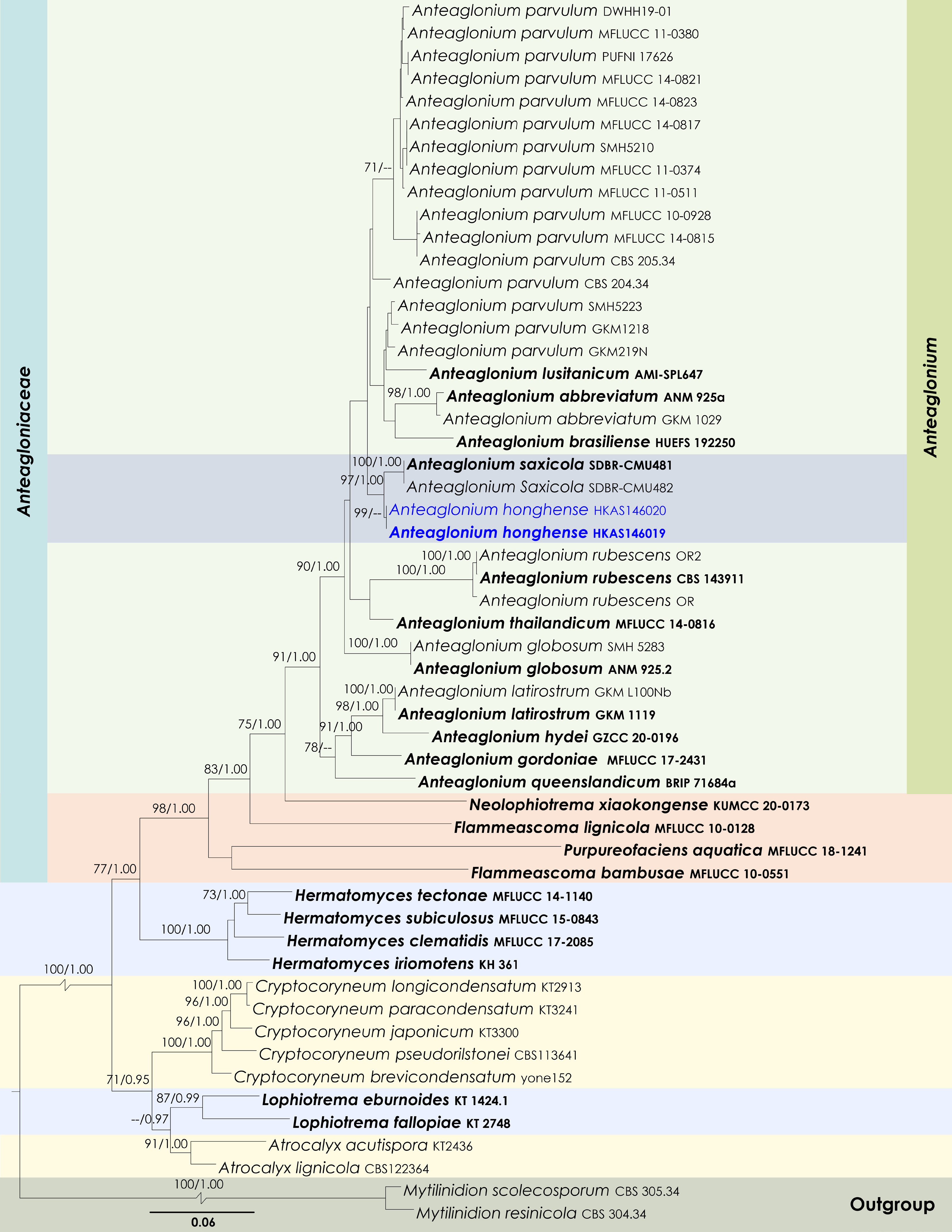
Figure 13.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Mytilinidion resinicola (CBS 304.34) and M. scolecosporum (CBS 305.34). The final likelihood value is –24165.625703. The final alignment included 1,579 unique site patterns, with approximately 41.67% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.244857, C = 0.256513, G = 0.274249, T = 0.224381. The substitution model yielded the following relative rates: AC = 1.294483, AG = 3.792940, AT = 1.360675, CG = 1.200289, CT = 9.762669, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.455208, and the gamma distribution shape parameter (α) was 0.434309. Bayesian inference reached convergence after 2,141,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009965). A total of 21,411 trees were sampled, and 16,059 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,580 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.

Figure 14.
Anteaglonium hongheense (HKAS146019, holotype). (a), (b) Hysterothecia on substrate. (c) Cross-section through hysterothecia. (d) Peridium. (e) Asci with pseudoparaphyses. (f), (g) Ascospores. Scale bars: (d), (e) = 10 µm, (f), (g) = 5 µm.
MycoBank No: 859354
Etymology: The specific epithet 'hongheense' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146019
Saprobic on dead twigs of Quercus sp. Sexual morph: Hysterothecia 350–500 μm long × 150–180 μm high × 170–220 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Jianshui County, Puxiongxiang 23.521432° N, 103.039049° E, 1,917 m, on dead twigs of Quercus sp., 15 March 2019, D.N. Wanasinghe, DW0528-09 (holotype, HKAS146019); ibid., DW1176-02 (HKAS146020).
GenBank numbers: HKAS146019: ITS = PV742894, LSU = PV742950, SSU = PV743003; HKAS146020: ITS = PV742895, LSU = PV742951, SSU = PV743004.
Notes: Multigene phylogenetic analyses revealed that Anteaglonium hongheense (HKAS146019 and HKAS146020) formed a sister subclade with A. saxicola (SDBR-CMU481 and SDBR-CMU482) (97% MLBS and 1.00 BYPP; Fig. 13). However, Anteaglonium hongheense is not morphologically comparable to A. saxicola, as the two species exhibit distinct morphs. Anteaglonium saxicola forms a coelomycetous asexual morph, characterized by pycnidial, dark brown to black, stromatic, globose to subglobose, ostiolate conidiomata; hyaline, ampulliform to subcylindrical, aseptate conidiogenous cells; and hyaline, subglobose to broadly ellipsoid, aseptate, smooth-walled conidia[112]. In contrast, Anteaglonium hongheense develops a sexual morph typical of Anteaglonium, with black, carbonaceous, hysterothecial ascomata and hyaline, fusiform, 1-septate ascospores[110,113]. Based on its distinct phylogenetic placement and morphological features, Anteaglonium hongheense is introduced here as a new species.
Didymellaceae Gruyter, Aveskamp & Verkley, Mycological Research 113 (4): 516.
Notes: Didymellaceae is one of the largest families in Pleosporales (Dothideomycetes), comprising approximately 50 genera and over 850 accepted species[59]. Didymellaceae species are cosmopolitan and inhabit a wide range of hosts and ecological niches, including terrestrial and aquatic environments[110,114−119]. Many taxa within this family have been documented as plant pathogens, endophytes, fungicolous fungi, lichenicolous fungi, or saprobes[110,114,115,118,120,121]. The family Didymellaceae was formally introduced by de Gruyter et al.[122], with Didymella designated as the type genus and Didymella exigua as the type species. In the present study, the most recent taxonomic treatments of Didymellaceae were followed followed Hongsanan et al.[110] and Hou et al.[118,119]. Based on a combination of multigene phylogenetic analyses and detailed morphological observations, the study introduces Gruyteromyces as a novel monophyletic genus within Didymellaceae to accommodate G. hongheensis sp. nov.
Gruyteromyces Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, gen. nov.
MycoBank No: 859355
Etymology: The generic epithet was given in honour of Johannes De Gruyter.
Saprobic on dead twigs of an unknown host. Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata pycnidial, black, solitary, scattered, immersed to slightly erumpent, subglobose, uni-loculate, visible as small black dots on host surface, glabrous, with inconspicuous ostioles. Pycnidial wall composed of brown to dark brown pseudoparenchymatous cells of textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells monoblastic, hyaline, smooth, phalidic, ampulliform, lining the conidiomatal cavity. Conidia ellipsoidal to oblong, hyaline, thin-walled, smooth to slightly rough, aseptate.
Type species: Gruyteromyces hongheensis Wanas. Phookamsak, & J.C. Xu
Notes: Multigene phylogenetic analyses revealed that Gruyteromyces forms a well-supported, monophyletic clade, representing an independent lineage within Didymellaceae. The genus is phylogenetically closely related to Ectodidymella, Ectophoma, and Longididymella (Fig. 15). Morphologically, Gruyteromyces exhibits characteristics typical of Didymellaceae, forming phoma-like asexual morphs similar to those observed in Ectodidymella, Ectophoma, and Longididymella[118]. However, Gruyteromyces can be distinguished from Ectodidymella by its small, subglobose conidiomata with inconspicuous ostioles, and ellipsoidal to oblong, hyaline, aseptate conidia that are smooth to slightly rough-walled. In contrast, Ectodidymella produces larger, globose to depressed-globose conidiomata, typically with a conspicuous neck, and oblong to subcylindrical conidia[118]. Ectophoma differs in having solitary to confluent conidiomata, each bearing one or more short ostiolar necks[116]. Longididymella is characterized by solitary to confluent, flask-shaped or irregular conidiomata with elongated ostiolar necks, variable conidiogenous cell morphology, and ellipsoidal to allantoid, smooth- and thin-walled, 0–1-septate conidia. As Gruyteromyces constitutes an independent lineage that is not strongly affiliated with any existing genus within Didymellaceae, Gruyteromyces gen. nov. is introduced in this study to accommodate this distinct lineage.

Figure 15.
Maximum Likelihood tree inferred from the concatenated dataset of partial LSU, ITS, rpb2 and tub2 sequences. The phylogeny is rooted with Leptosphaeria conoidea (CBS 616.75) and L. doliolum (CBS 505.75). The final likelihood value is –30,111.669487. The final alignment included 1,051 unique site patterns, with approximately 16.06% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.240399, C = 0.243000, G = 0.274422, T = 0.242180. The substitution model yielded the following relative rates: AC = 2.034753, AG = 7.468658, AT = 2.243260, CG = 1.095539, CT = 14.634190, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.633611, and the gamma distribution shape parameter (α) was 0.621330. Bayesian inference reached convergence after 1,476,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009988). A total of 14,761 trees were sampled, and 11,071 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,053 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated with a 'T' at the end of the strain number.
Gruyteromyces hongheensis Wanas. Phookamsak, & J.C. Xu, sp. nov. (Figs 15 and 16).
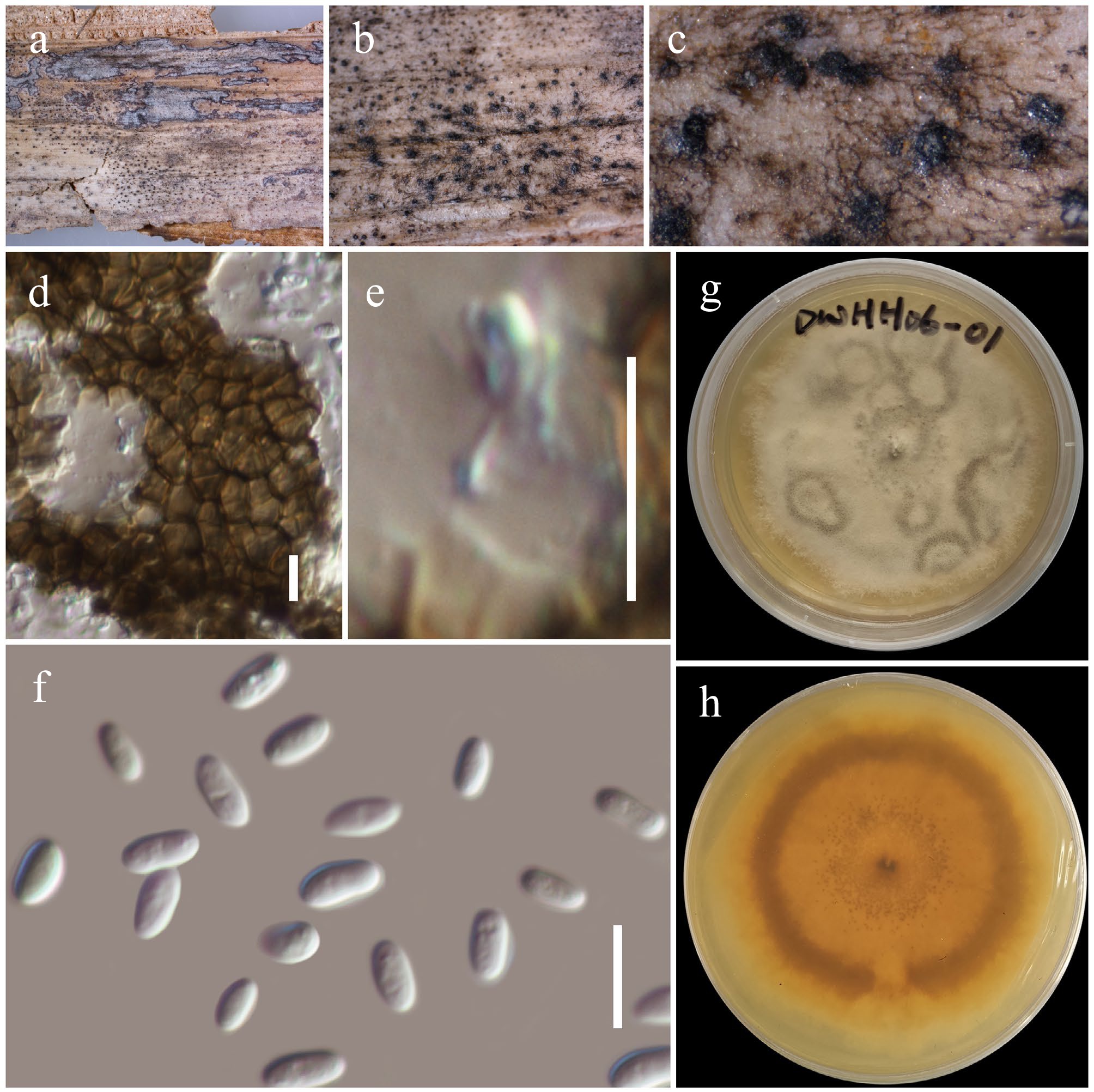
Figure 16.
Gruyteromyces hongheensis (HKAS146022, holotype). (a)–(c) Conidiomata on host. (d) Pycnidial wall. (e) Conidiogenous cell. (f) Conidia. (g), (h) Colony on PDA (h from the bottom). Scale bars: (d)–(f) = 10 µm.
MycoBank No: 859356
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146022
Saprobic on the dead sheath of a Poaceae host. Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 130–170 µm high × 220–280 µm diam. (
$\overline {\rm x} $ $\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Colonies on PDA, 25–30 mm diam after one week, margin irregular, covered by velvety, sparse, flat mycelia, pinkish white, iron-grey ring patterns are visible near the centre and at peripheral areas. Sporulation has started after 45 d under light conditions at 25 °C. Reverse; orangish white, notable iron-grey ring was visible at the periphery.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead sheath of a Poaceae sp., 03 December 2020, D.N. Wanasinghe, DWHH06-01 (holotype, HKAS146022), ex-type, KUNCC23-16758; ibid., DWHH06-02 (paratype, HKAS146023), ex-paratype KUNCC25-19455.
GenBank numbers: KUNCC23-16758: ITS = PV742896, LSU = PV742952, SSU = PV743005, tef1-α = PV700631, rpb2 = PV700677, tub2 = PV738672; HKAS146023: ITS = PV742897, LSU = PV742953, SSU = PV743006, tef1-α = PV700632, rpb2 = PV700678, tub2 = PV738673.
Notes: Multigene phylogenetic analyses demonstrated that two new strains (KUNCC23-16758 and KUNCC25-19455) of Gruyteromyces hongheensis formed a distinct subclade (60% MLBS; Fig. 15) basal to Ectodidymella nigrificans (CBS 100190). Gruyteromyces hongheensis can be distinguished from E. nigrificans in having subglobose conidiomata with an inconspicuous ostiole and slightly smaller conidia (5–8 × 2.5–4 µm vs 6–9 × 2–3 μm). On the other hand, E. nigrificans has globose to depressed globose conidiomata, usually with a conspicuous neck[118].
Didymosphaeriaceae Munk, Dansk botanisk Arkiv 15 (2): 128 (1953)
Chromolaenicola Mapook & K.D. Hyde, Fungal Diversity 101: 20 (2020)
Notes: Chromolaenicola is a well-defined genus in Didymosphaeriaceae, characterized by both sexual and asexual morphs, and supported by morphological features and multigene phylogenetic analyses. The genus was established by Mapook et al.[69] to accommodate four new species and one new combination, with Chromolaenicola nanensis designated as the type species. Most species of Chromolaenicola are saprobic, commonly found on hosts such as Ananas comosus, Bidens pilosa, Chromolaena odorata, Clematis subumbellata, and Leucaena sp. in Thailand[46,69,108,123,124]. To date, only Chromolaenicola sapindi has been recorded outside Thailand, collected from dead woody twigs of Sapindus rarak in Yunnan, China[125]. Currently, eight species are listed under Chromolaenicola in Index Fungorum[89]. The sexual morph of Chromolaenicola is characterized by immersed to semi-immersed, solitary, coriaceous, globose to subglobose ascomata with a protruding ostiolar neck; 6–8-spored, bitunicate, cylindrical asci; and reddish brown to brown, ellipsoid to broadly fusiform, muriform ascospores[69]. The coelomycetous asexual morph produces solitary, pycnidial, globose to obpyriform conidiomata with a papillate ostiole, and hyaline, unbranched, broadly filiform to ampulliform conidiogenous cells. Conidia are oblong to ellipsoid, globose to subglobose, hyaline to brown, 0–1-septate, and finely verruculose[69]. Notably, some species of Chromolaenicola have demonstrated antimicrobial activity against Mucor plumbeus and Bacillus subtilis[46,69]. In the present study, Chromolaenicola hongheensis sp. nov., a saprobe isolated from Citrus australasica, was introduced, marking the first report of this genus from this host and the second record of Chromolaenicola from Yunnan Province, China. In addition, C. clematidis was reported from dead twigs of an unidentified deciduous host in Yunnan, China, for the first time.
Chromolaenicola clematidis Phukhams. & K.D. Hyde, Fungal Diversity 102: 31 (Figs 17 and 18).
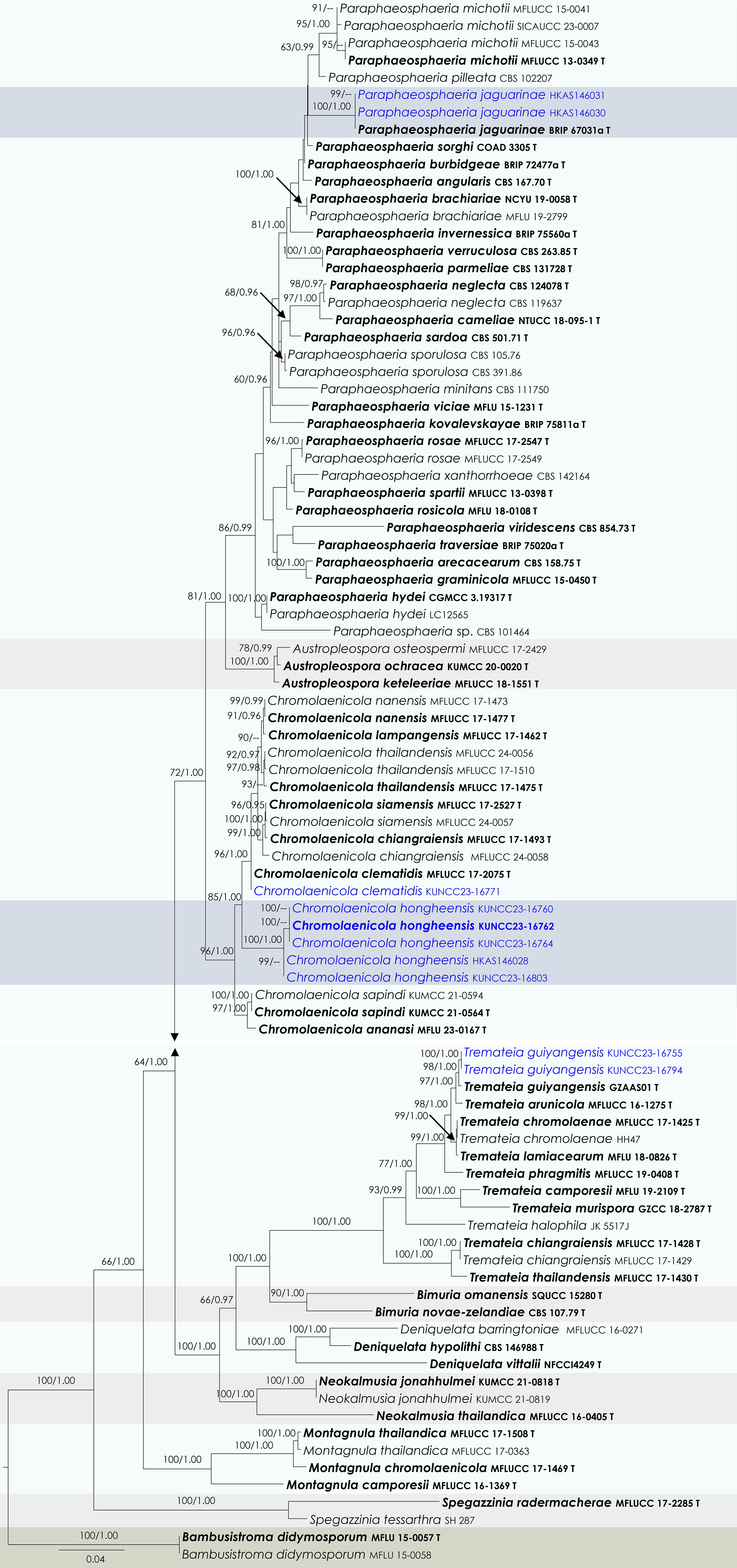
Figure 17.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α and rpb2 sequences. The phylogeny is rooted with Bambusistroma didymosporum (MFLU 15-0057, MFLU 15-0058). The final likelihood value is –29,800.793962. The final alignment included 1,978 unique site patterns, with approximately 36.43% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.238939, C = 0.255983, G = 0.270197, T = 0.234881. The substitution model yielded the following relative rates: AC = 1.677295, AG = 3.846144, AT = 1.480569, CG = 1.417310, CT = 8.251750, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.419546, and the gamma distribution shape parameter (α) was 0.473863. Bayesian inference reached convergence after 2,194,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009957). A total of 21,941 trees were sampled, and 16,456 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,981 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.

Figure 18.
Chromolaenicola clematidis (HKAS146029). (a), (b) Conidiomata on host. (c), (d) Sections of conidiomata. (e) Conidiogenous cells. (f), (g) Conidia. (h), (i) Colony on MEA (i from the bottom). Scale bars: (d) = 50 µm, (e)–(g) = 10 µm.
MycoBank No: 557133
Saprobic on dead wood of an unknown deciduous host. Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 75–110 × 100–140 µm (
$\overline {\rm x} $ $\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Colonies on PDA, 35 mm diam. after one week, the margin is irregular, covered by velvety, puffy mycelium that sometimes condenses in the middle as patches, and the mouse gray. Reverse: grayish brown at the middle, grayish white at the periphery.
Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013 N, 102.227243 E, 517 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH14-04 (HKAS146029), living culture, KUNCC23-16771.
GenBank numbers: KUNCC23-16771: ITS = PV742898, LSU = PV742954, SSU = PV743007, tef1-α = PV700633, rpb2 = PV700679.
Known hosts and distribution: Saprobic on dead stems of Clematis subumbellata in Thailand[123]; on dead twigs of an unknown deciduous host in Yunnan, China (this study).
Notes: Phylogenetically, the new strain (KUNCC23-16771) shared the same branch with the type of Chromolaenicola clematidis (MFLUCC 17-2075) and clustered with C. nanensis, C. lampangensis, C. thailandensis, C. siamensis, and C. chiangraiensis with 96% MLBS and 1.00 BYPP support values (Fig. 17). The new strain KUNCC23-16771 shares similar morphological characteristics with the type strain of C. clematidis in having black, subglobose to globose, uni-loculate conidiomata, and brown, 0–1-septate conidia. However, our collection (HKAS146029) has broader conidiomata (75–110 × 100–140 µm vs 76–145 × 107–128 μm), with inconspicuous ostioles, broader (7.5–9.5 × 6.5–7.7 µm vs 7–10 × 4.5–7 μm), globose to subglobose, and smooth-walled conidia. On the other hand, the type of C. clematidis has ostiolate, papillate conidiomata, and ellipsoidal to subglobose, verrucose conidia[123]. Despite these slight dimensional differences, based on phylogenetic results and other morphological similarities, the new isolate is identified as Chromolaenicola clematidis and is reported from Yunnan, China, for the first time.
Chromolaenicola hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 17 and 19).
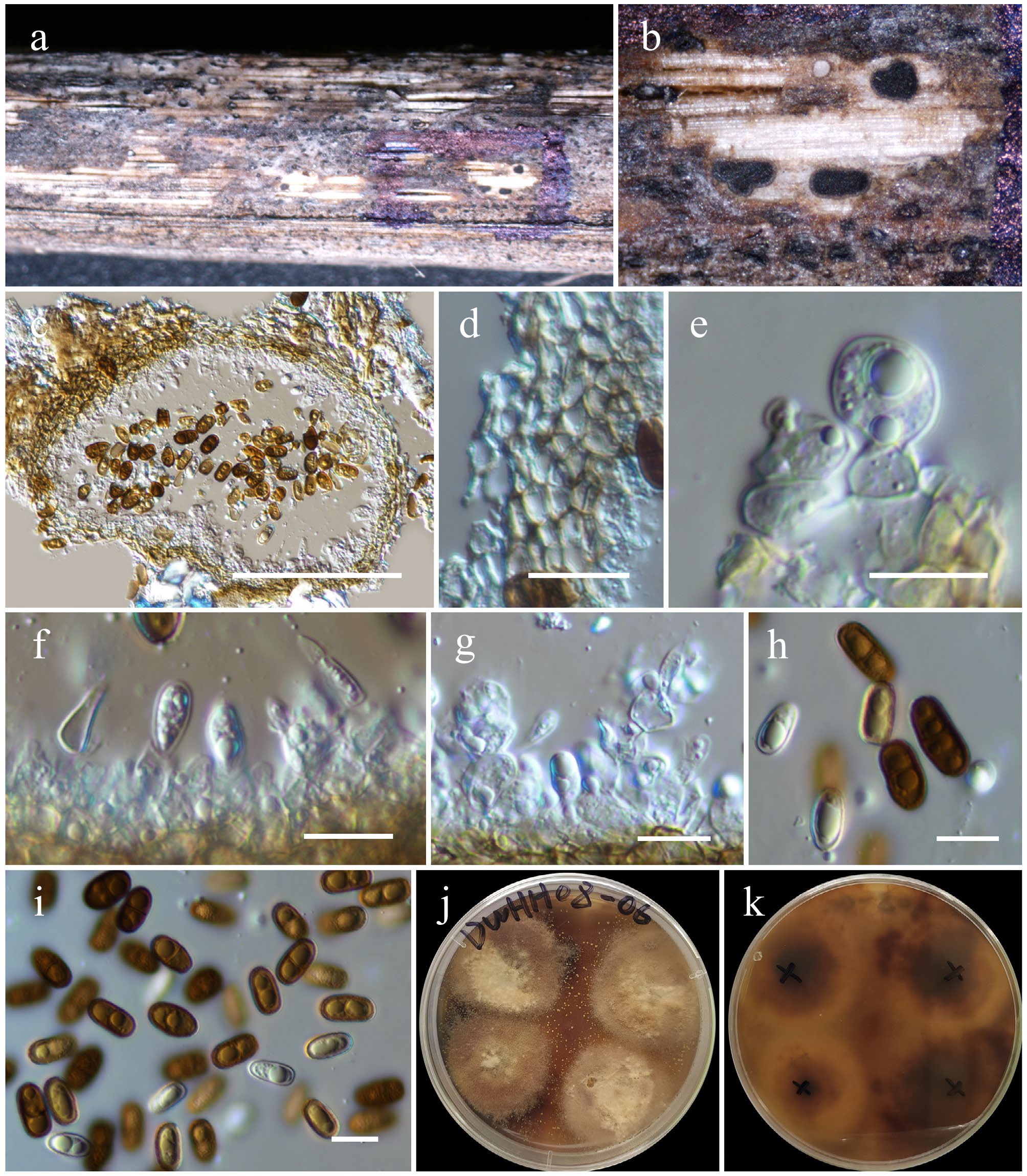
Figure 19.
Chromolaenicola hongheensis (HKAS146024, holotype). (a) Conidiomata on host. (b), (c) Sections of conidiomata. (d) Pycnidial wall. (e)–(g) Conidiogenous cells. (h), (i) Conidia. (j), (k) Colony on PDA (k from the bottom). Scale bars: (c) = 100 µm, (d)–(i) = 10 µm.
MycoBank No: 859357
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146024
Saprobic on dead twigs of Citrus australasica. Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 150–200 µm high × 120–250 µm diam. (
$\overline {\rm x} $ $\overline {\rm x} $ $\overline {\rm x} $ Culture characteristics: Colonies on PDA, 30 mm diam. after one week, margin irregular, covered by velvety, puffy mycelium that sometimes condenses in the middle as patches, and the mouse is gray. Reverse: grayish brown at the middle, grayish white at the periphery.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead twigs of Citrus australasica, 14 August 2022, D.N. Wanasinghe, DWHH08-06 (holotype, HKAS146024), ex-type, (KUNCC23-16762); ibid., DWHH08-05 (HKAS146025), living culture (KUNCC23-16760); ibid., DWHH08-08 (HKAS146026), living culture (KUNCC23-16764); ibid., DWHH22-18-01 (HKAS146027), living culture (KUNCC23-16803); ibid., DWHH22-18-02 (HKAS146028).
GenBank numbers: KUNCC23-16762: ITS = PV742899, LSU = PV742955, SSU = PV743008, tef1-α = PV700634, rpb2 = PV700680; KUNCC23-16760: ITS = PV742900, LSU = PV742956, SSU = PV743009, tef1-α = PV700635, rpb2 = PV700681; KUNCC23-16764: ITS = PV742901, LSU = PV742957, SSU = PV743010, tef1-α = PV700636; KUNCC23-16803: ITS = PV742902, LSU = PV742958, SSU = PV743011, tef1-α = PV700637, rpb2 = PV700682; HKAS146028: ITS = PV742903, LSU = PV742959, SSU = PV743012, tef1-α = PV700638, rpb2 = PV700683.
Notes: Multigene phylogenetic analyses based on a concatenated dataset of SSU, LSU, ITS, tef1-α, and rpb2 loci revealed that five strains of Chromolaenicola hongheensis (KUNCC 23-16762, KUNCC 23-16760, KUNCC 23-16764, KUNCC 23-16803, and HKAS 146028) formed a monophyletic subclade (100% MLBS and 1.00 BYPP; Fig. 17), positioned basally to C. nanensis, C. lampangensis, C. thailandensis, C. siamensis, C. chiangraiensis, and C. clematidis. Among these, C. hongheensis shows the closest phylogenetic affinity to C. clematidis (85% MLBS and 1.00 BYPP; Fig. 17). Morphologically, C. hongheensis is comparable to C. clematidis in conidial shape but can be distinguished by its larger conidiomata (150–200 × 120–250 µm vs 76–145 × 107–128 µm) and larger, ellipsoidal to oblong conidia (10–14 × 5–8 µm vs 7–10 × 4.5–7 µm). In contrast, C. clematidis typically produces ellipsoidal to subglobose conidia[123]. These morphological and phylogenetic distinctions support the recognition of C. hongheensis as a novel species.
Paraphaeosphaeria O.E. Erikss., Ark. Bot. Ser. 2. 6 (4–5): 405.
Notes: Paraphaeosphaeria was introduced by Eriksson[126] with P. michotii as the type species. The genus was initially established to accommodate four sexual species, including P. castagnei, P. michotii, P. obtusispora, and P. rusci[110]. The genus is characterized by immersed to semi-immersed, globose to subglobose ascomata, with protruding ostiole, or a short beak, 8-spored, bitunicate, fissitunicate, broadly cylindrical asci, and brown to yellowish brown, oblong to cylindrical, tapering towards the lower end cell, septate ascospores[110,127]. Paraphaeosphaeria produces coniothyrium-like asexual morphs and is characterized by eustromatic or pycnidial conidiomata, phialidic or annellidic, discrete or integrated conidiogenous cells, and ellipsoid to obovoid-pyriform, hyaline to brown, 0–1-septate, smooth to verrucose conidia[128]. Presently, 52 epithets under Paraphaeosphaeria are available in Index Fungorum[89]. In this study, the new host record for P. jaguarinae in Yunnan, China, was reported for the first time.
Paraphaeosphaeria jaguarinae Y.P. Tan & R.G. Shivas, Index Austral. Fungi 19: 5 (Figs 17 and 20).

Figure 20.
Paraphaeosphaeria jaguarinae (HKAS146030). (a)–(c) Conidiomata on host. (d) Section of a conidioma. (e) Pycnidial wall. (f) Conidiogenous cells. (g), (h) Conidia. Scale bars: (d)–(h) = 10 µm.
MycoBank No: 901285
Saprobic on dead leaf blades of Poaceae hosts. Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 40–60 μm high × 55–70 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.390025 N, 102.225942 E, 1186 m, on dead leaf blades of Poaceae sp., 13 March 2023, D.N. Wanasinghe, DWHH23-38-1 (HKAS146030); ibid., DWHH23-38-2 (HKAS146031).
GenBank numbers: HKAS146030: ITS = PV742904, LSU = PV742960, SSU = PV743013, tef1-α = PV700639, rpb2 = PV700684; HKAS146031: ITS = PV742905, LSU = PV742961, SSU = PV743014, tef1-α = PV700640, rpb2 = PV700685.
Known hosts and distribution: Associated with leaf spot of Arundienella sp. in Queensland, Australia[129]; saprobic on dead leaf blades of Poaceae spp. in Yunnan, China (this study).
Notes: Multigene phylogenetic analyses based on a combined dataset of SSU, LSU, ITS, tef1-α, and rpb2 sequences revealed that two newly obtained strains (HKAS 146030 and HKAS 146031) clustered within the same branch as the type strain of Paraphaeosphaeria jaguarinae (BRIP 67031a), with 100% MLBS and 1.00 BYPP statistical support (Fig. 17). However, when P. jaguarinae was introduced by Tan & Shivas[129], no morphological data were provided for the type strain, rendering direct morphological comparison with our new isolates impossible. Based on robust phylogenetic evidence and their Poaceae host association, the new isolates were identified as P. jaguarinae. This study provides an updated morphological description and photographic illustrations of the species based on our collections.
Tremateia Kohlm., Volkm.-Kohlm. & O.E. Erikss., Bot. Mar. 38: 165.
Notes: Tremateia was introduced as a monotypic genus by Kohlmeyer et al.[130] and is typified by T. halophila. The genus was initially established to accommodate a facultative marine fungus occurring on senescent culms of Juncus roemerianus in North Carolina (USA). Ariyawansa et al.[127] recircumscribed the genus based on the isotype of the type species T. halophila that is characterized by solitary, scattered, brown to black, globose to subglobose ascomata, 4–8-spored, bitunicate, clavate to broadly clavate asci, embedded in cellular pseudoparaphyses, and ellipsoid to fusiform, light brown to brown, muriform, with phoma-like asexual morph. Subsequently, nine species were included in this genus[69,131−134]. These species were found as saprobes in both marine and terrestrial habitats. In the present study, T. guiyangensis is reported on dead twigs of an unknown deciduous host in Yunnan, China, for the first time.
Tremateia guiyangensis J.F. Zhang, J.K. Liu, K.D. Hyde & Z.Y. Liu, Fungal Diversity 80: 48 (2016) (Figs 17 and 21).
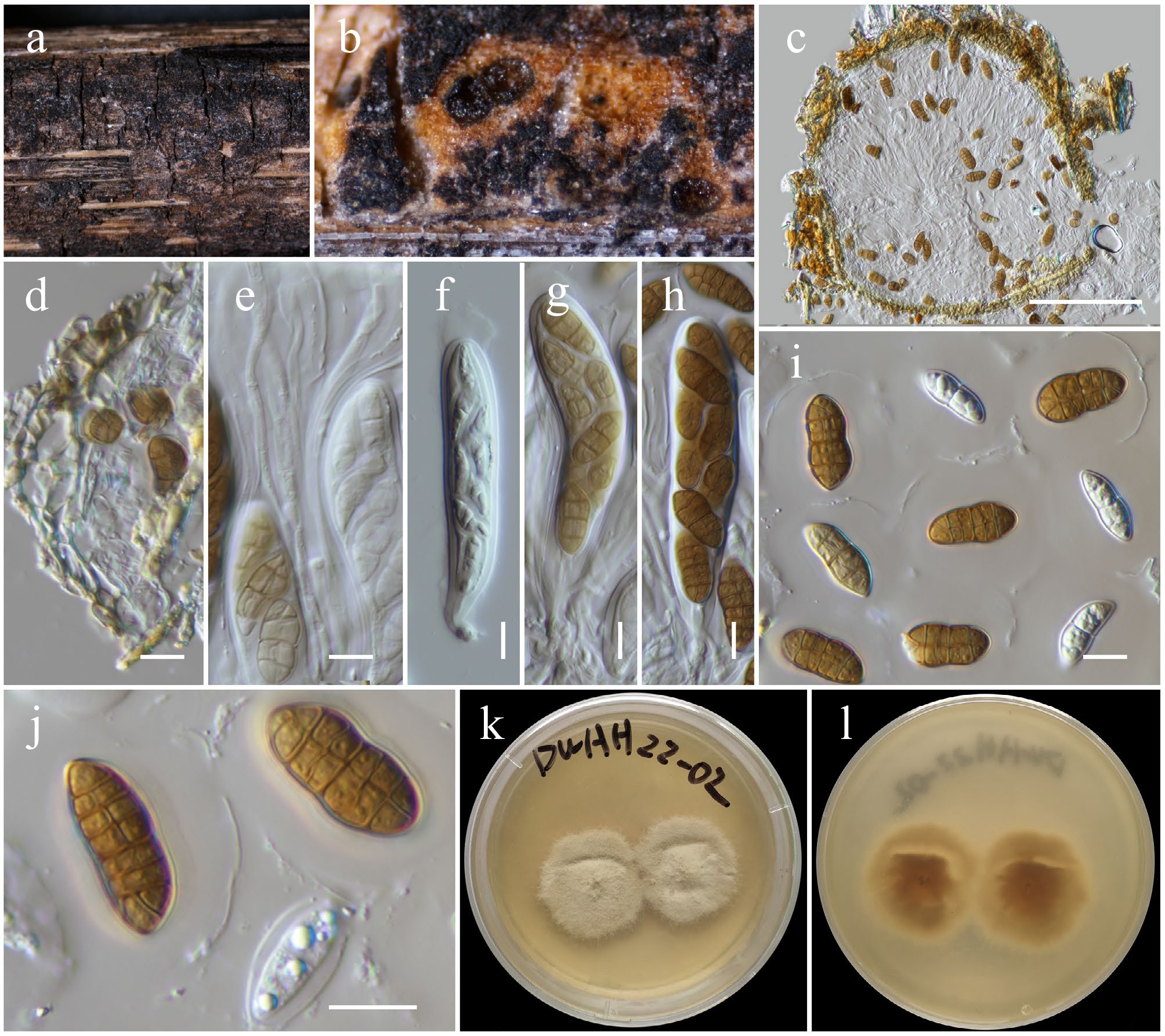
Figure 21.
Tremateia guiyangensis (HKAS146033). (a) Ascomata on the dead woody twigs. (b), (c) Cross-section of ascomata. (d) Peridium. (e) Pseudoparaphyses. (f)–(h) Asci. (i), (j) Ascospores. (k), (l) Colony on PDA (l from the bottom). Scale bars: (c) = 100 µm, (d)–(j) = 10 µm.
MycoBank No: 552160
Saprobic on a dead twig of a deciduous host. Sexual morph: Ascomata 230–270 μm high × 250–280 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA, 20 mm diam. after one week, margin was irregular, covered by pinkish white cottony mycelium, and a notable partial groove was visible. Reverse: pinkish brown.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH22-02 (HKAS146033), living culture, KUNCC23-16794; ibid., DWHH22-03 (HKAS146034), living culture, KUNCC23-16755.
GenBank numbers: KUNCC23-16794: ITS = PV742906, LSU = PV742962, SSU = PV743015, tef1-α = PV700641; KUNCC23-16755: ITS = PV742907, LSU = PV742963, SSU = PV743016, tef1-α = PV700642.
Known hosts and distribution: Saprobic on dead herbaceous stems in Guizhou, China[131]; on dead twigs of an unknown deciduous host in Yunnan, China (this study).
Notes: Phylogenetic analyses based on a combined dataset of SSU, LSU, ITS, tef1-α, and rpb2 sequence data revealed that two new strains (KUNCC 23-16794 and KUNCC 23-16755) clustered with the type strain of Tremateia guiyangensis (GZAAS01) (98% MLBS and 1.00 BYPP; Fig. 17). This clade is sister to T. arunicola (MFLUCC 16-1275, type strain), also with 97% MLBS and 1.00 BYPP statistical support. Morphologically, the new isolates are consistent with T. guiyangensis in possessing coriaceous, subglobose ascomata with minutely papillate ostioles, cylindric-clavate asci and muriform, ellipsoidal to broadly fusiform and golden-brown ascospores. However, they exhibit smaller asci (80–100 × 16–22 μm vs 152–160 × 21–27 μm), and ascospores (19–23 × 9–11 μm vs 20–28 × 9–12 μm). Additionally, the ascospores of the new isolates have 2–7 transverse septa and a single longitudinal septum in each row, and are covered by a hyaline gelatinous sheath. In contrast, the type strain of T. guiyangensis typically has 3–5 transverse septa, one longitudinal septum per row, and lacks a mucilaginous sheath[131]. Based on the strong phylogenetic affinity and general morphological congruence, these two isolates are identified as Tremateia guiyangensis. This represents the first report of the species from dead twigs of an unidentified deciduous host in Yunnan, China.
Lophiostomataceae Sacc., Syll. Fung. 2: 672.
Neovaginatispora A. Hashim., K. Hiray. & Kaz. Tanaka, Stud. Mycol. 90: 188.
Notes: Neovaginatispora was introduced as a monotypic genus by Hashimoto et al.[135] to accommodate the type species, N. fuckelii. The genus is commonly known as saprobic fungi in terrestrial and freshwater habitats[135−138], as well as causing leaf spot on Phoenix canariensis[139]. The genus is characterized by solitary, coriaceous, black, subglobose ascomata, with rounded or slit-like ostioles, 8-spored, bitunicate, fissitunicate, cylindric-clavate asci, embedded in narrow cellular pseudoparaphyses, and fusiform, hyaline, 1–2-septate, smooth-walled ascospores, with appendages at both ends[136]. The coelomycetous asexual morph of Neovaginatispora has been reported for N. aquadulcis by Magaña-Dueñas et al.[140] and is characterized by pycnidial, brown to dark brown, semi-immersed, solitary, globose, ostiolate conidiomata, phialidic, determinate, hyaline, globose, conidiogenous cells, and, aseptate, hyaline, smooth-walled, guttulate, conidia. Chlamydospores are solitary or in chains, abundant, terminal, and intercalary, brown, globose to clavate, aseptate, thick-walled[140]. Presently, only four species are accommodated in this genus, of which the type species, N. fuckelii is cosmopolitan. In this study, N. fuckelii is reported as the new collection on dead twigs of an unknown deciduous host in Yunnan, China.
Neovaginatispora fuckelii (Sacc.) A. Hashim., K. Hiray. & Kaz. Tanaka, Stud. Mycol. 90: 188 (Figs 22 and 23).
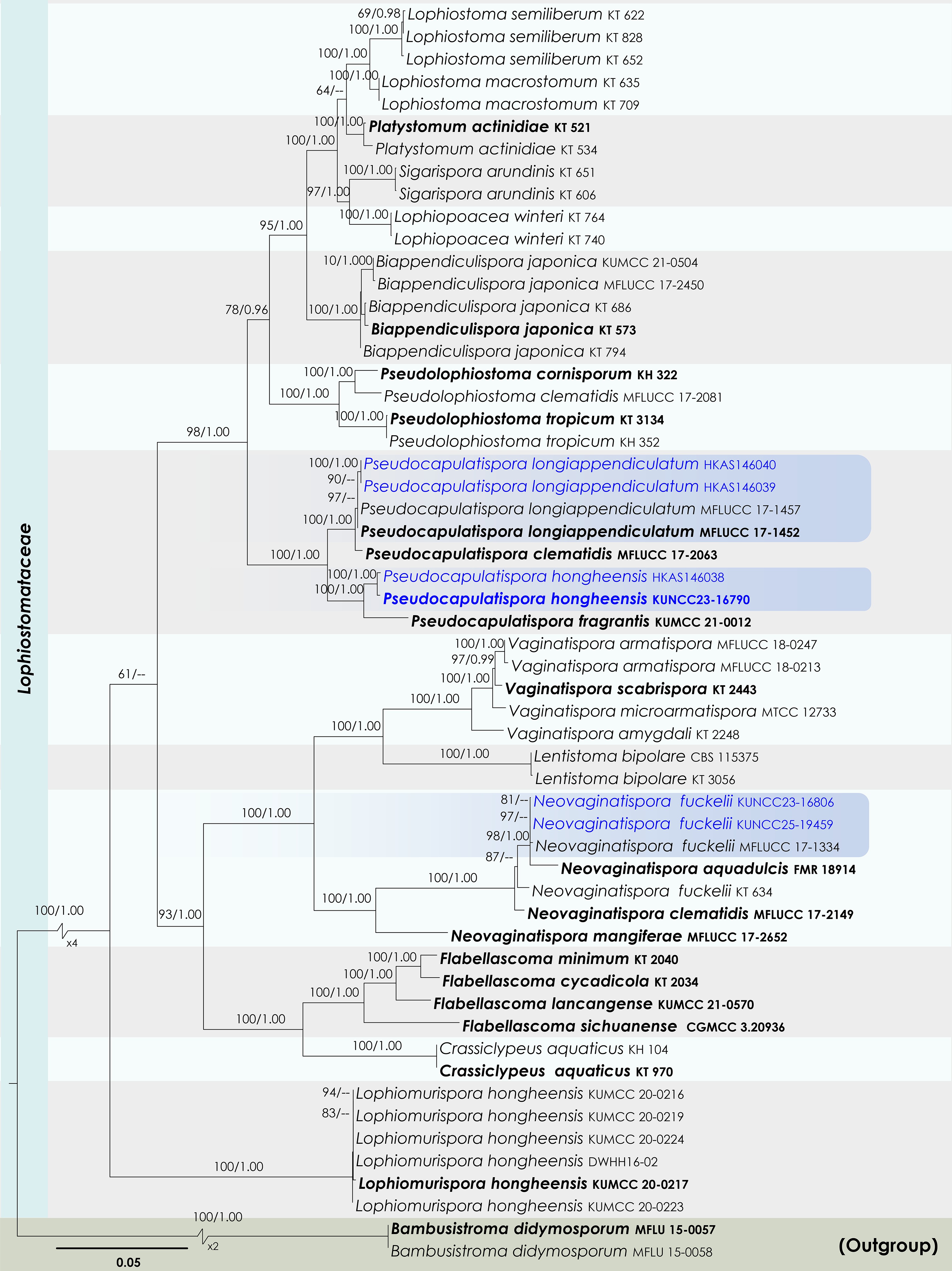
Figure 22.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Bambusistroma didymosporum (MFLU 15-0057, MFLU 15-0058). The final likelihood value is –23,551.161444. The final alignment included 1,560 unique site patterns, with approximately 13.11% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.250154, C = 0.249645, G = 0.265412, T = 0.234790. The substitution model yielded the following relative rates: AC = 1.772887, AG = 4.441914, AT = 1.366252, CG = 1.518065, CT = 9.220171 and GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.527527, and the gamma distribution shape parameter (α) was 0.567921. Bayesian inference reached convergence after 44,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009691). A total of 441 trees were sampled, and 331 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,561 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.

Figure 23.
Neovaginatispora fuckelii (HKAS146035). (a), (b) Ascomata on the dead woody twigs. (c) Transverse section of ascomata. (d), (e) Cross-section of ascomata. (f) Close up of ostiole. (g) Peridium. (h) Pseudoparaphyses. (i)–(k) Asci. (l)–(o) Ascospores. Scale bars: (d), (e) = 100 µm, (h)–(o) = 10 µm.
MycoBank No: 823148
Saprobic on a dead twig of Fabaceae spiny host. Sexual morph: Ascomata 230–270 μm high × 250–280 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA at room temperature (25 °C) under normal light, reaching 28–30 mm diam. after two weeks. Colonies dense, circular, flattened, slightly raised, to low convex, surface smooth, with edge entire, floccose, smooth aspect, do not produce pigmentation in PDA. Colonies sectored, from above grey and white, slightly radiating with concentric ring, pale grey near the margin; colonies from below sectored, white to cream at the margin, dark greenish grey to black near the margin, paler grey to cream toward the center, another white to cream towards the center.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.390025° N, 102.225942° E, 1,186 m, on dead twigs of Fabaceae spiny sp., 13 March 2023, D.N. Wanasinghe, DWHH23-27A (HKAS146035), living culture, KUNCC23-16806; ibid., 23 April 2020, DWHH23-64 (HKAS146036), living culture, KUNCC25-19459.
GenBank numbers: KUNCC23-16806: ITS = PV742908, LSU = PV742964, SSU = PV743017, tef1-α = PV700643, rpb2 = PV700686; HKAS146036: ITS = PV742909, LSU = PV742965, SSU = PV743018, tef1-α = PV700644, rpb2 = PV700687.
Known hosts and distribution: Saprobic on stems of unknown herbaceous plant in Japan[136]; on dead stems of Mangifera indica in Taiwan[141], on decaying wood submerged in a stream in Yunnan[137], on decaying wood of Prunus sp. in Yunnan[132], on a branch of Camellia oleifera, Idesia polycarpa, Juglans nigra and Paeonia suffruticosa in Sichuan[142], pathogen causing leaf spot Phoenix canariensis in Sichuan[139], stem blight on Rosa chinensis in Yunnan, China[143], on dead twigs of Fabaceae sp. (this study).
Notes: Phylogenetic analyses of a combined SSU, LSU, ITS, tef1-α, and rpb2 demonstrated that the new strains (KUNCC23-16806 and KUNCC25-19459) shared the same branch with Neovaginatispora fuckelii (MFLUCC 17-1334) (98% MLBS and 1.00 BYPP; Fig. 22) and formed a subclade with N. aquadulcis (FMR 18914), N. fuckelii (KT 634) and N. clematidis (MFLUCC 17-2149) with 100% MLBS and 1.00 BYPP support (Fig. 22). Morphologically, the new isolates resemble N. fuckelii (MFLUCC 17-1334) in having semi-immersed, coriaceous, black, subglobose ascomata, with periphysate, rounded or slit-like ostiole, cylindrical asci, and hyaline, fusiform, 1-septate ascospores. However, the new isolates slightly differ from N. fuckelii (MFLUCC 17-1334) in having larger asci (100–140 × 14–20 μm vs 47–54 × 6.5–8.5 μm) and larger ascospores (30–38 × 7–8.2 μm vs 12–13 × 3–4 μm). Additionally, the ascospores of new isolates are surrounded by conspicuous hyaline mucilaginous sheath, with tail-like appendages at both ends, and lacking appendages, whereas N. fuckelii (MFLUCC 17-1334) is present with appendages at both ends lacks a mucilaginous sheath covering ascospores[137]. These isolates can be distinguished from N. aquadulcis (FMR 18914) in having smaller, carbonaceous to coriaceous, globose to subglobose ascomata, cylindrical asci, and 1-septate ascospores. At the same time, N. aquadulcis (FMR 18914) has larger (400–550 × 300–325 μm), pyriform ascomata, with conic-truncate necks, cylindrical to cylindrical-clavate asci, and 3-septate ascospores, with papillate to pulvinate appendages at each end[140]. Based on phylogenetic evidence and morphological characteristics, the new isolates are identified as N. fuckelii herein.
Pseudocapulatispora Mapook & K.D. Hyde, Fungal Diversity 101: 47.
Notes: Pseudocapulatispora was introduced by Mapook et al.[69] as a monotypic genus to accommodate the type species, P. longiappendiculata. The genus is characterized by immersed, coriaceous, solitary, ovoid, light brown to brown ascomata, with long, carbonaceous, crest-like, ostiolar necks, 8-spored, bitunicate, fissitunicate, cylindric-clavate asci, and hyaline, broadly fusiform, 1-septate ascospores, with narrow sheaths drawn out to form polar appendages at both ends. Andreasen et al.[144] synonymized this genus under Lophiostoma. Their treatment has not been followed by recent publications[145]. Pseudocapulatispora was also treated as a distinct genus from Lophiostoma, as it exhibits morphological differences from the latter, such as immersed, coriaceous ascomata, with long, carbonaceous, crest-like, ostiolar necks and hyaline, broad fusiform, 1-septate ascospores, with a long sheath drawn out to form polar appendages[69,123,145]. Whereas, Lophiostoma has immersed to semi-immersed, coriaceous to carbonaceous ascomata, with short, pore-like, or crest-like, papillate ostioles, and hyaline to yellowish brown, uni- to multi-septate ascospores, without or with terminal appendages, and lacking a mucilaginous sheath[136]. Currently, three species are accounted in this genus, viz. Pseudocapulatispora clematidis, P. fragrantis, and P. longiappendiculata. However, Pseudocapulatispora fragrantis is considered Nom. inval., Art. 40.7 (Shenzhen) in Index Fungorum[89]. In the present study, the new species P. hongheensis is introduced based on morphological characteristics and phylogenetic evidence.
Pseudocapulatispora hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 22 and 24).

Figure 24.
Pseudocapulatispora hongheensis (HKAS146037, holotype). (a)–(c) Ascomata on the dead woody twigs. (d) Cross-section of ascoma. (e) Close up of ostiole. (f) Peridium. (g) Pseudoparaphyses. (h)–(j) Asci. (k)–(m) Ascospores. Scale bars: (d) = 100 µm, (e)–(m) = 10 µm.
MycoBank No: 859358
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146037
Saprobic on a dead twig of an unknown deciduous host. Sexual morph: Ascomata 320–380 μm high × 240–290 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA at room temperature (25 °C) under normal light, reaching 18–20 mm diam. after two weeks. Colonies dense, circular, flattened, slightly raised, surface smooth, with curled to undulate edge, floccose to velvety, smooth aspect, slightly radiated, produce yellow pigmentation in PDA. Colonies from above dark greenish to black at the margin, white to pale grey at the center, with yellow droplets; colonies from below dark greenish to black at the margin towards the middle, cream to pale yellowish at the center.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.390025° N, 102.225942° E, 1,186 m, on dead twigs of an unknown deciduous host, 13 March 2023, D.N. Wanasinghe, DWHH23-06 (holotype, HKAS146037), ex-type, KUNCC23-16790: ibid., DWHH23-55 (HKAS146038).
GenBank numbers: KUNCC23-16790: ITS = PV742910, LSU = PV742966, SSU = PV743019, tef1-α = PV700645, rpb2 = PV700688; HKAS146038: ITS = PV742911, LSU = PV742967, SSU = PV743020, tef1-α = PV700646, rpb2 = PV700689.
Notes: Phylogenetic analyses of a combined SSU, LSU, ITS, tef1-α, and rpb2 sequence data demonstrated that two new strains (KUNCC23-16790 and HKAS146038) of Pseudocapulatispora hongheensis sp. nov. formed a robust subclade, sister to P. fragrantis with 100% MLBS and 1.00 BYPP support (Fig. 22). These two species constituted the base of P. clematidis and P. longiappendiculata with 100% MLBS and 1.00 BYPP support (Fig. 22), and are distant from Lophiostoma. Morphologically, P. hongheensis resembles P. fragrantis but can be distinguished by slightly smaller ascomata (320–380 μm high × 240–290 μm diam. vs 220–377 μm high × 162–325 μm diam.), shorter and broader asci (90–120 × 14–20 μm vs 58–127 × 8–15 μm), and ascospores (30–40 × 7–8 μm vs 29–36 × 4–8 μm) with shorter tail-like appendages (10–20 µm long vs 16–46 × 8–14 μm)[145]. Pseudocapulatispora hongheensis produced yellowish pigmentation on PDA, whereas P. fragrantis does not produce pigmentation on PDA. Therefore, Pseudocapulatispora hongheensis sp. nov. is introduced herein as a distinct species.
Pseudocapulatispora longiappendiculata Mapook & K.D. Hyde, Fungal Diversity 101: 48 (Figs 22 and 25).
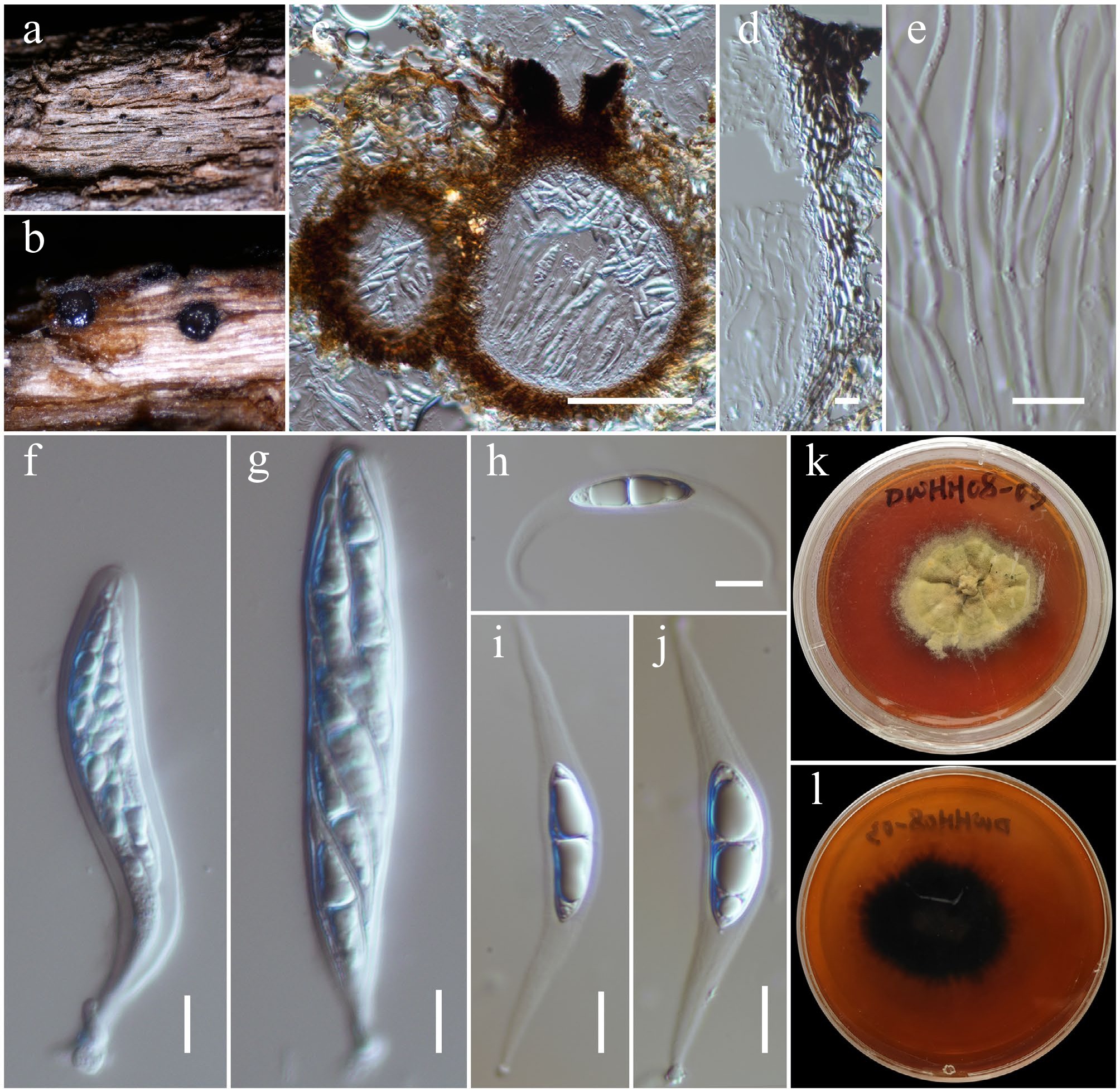
Figure 25.
Pseudocapulatispora longiappendiculata (HKAS146039). (a) Ascomata on the dead woody twigs. (b), (c) Cross-section of ascomata. (d) Peridium. (e) Pseudoparaphyses. (f), (g) Asci. (h)–(j) Ascospores. (k), (l) Colony on MEA (l from the bottom). Scale bars: (c) = 100 µm, (d)–(j) = 10 µm.
MycoBank No: 557286
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Ascomata 220–280 μm high × 140–200 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA at room temperature (25 °C) under normal light, reaching 24–26 mm diam. after two weeks. Colonies dense, irregular, flattened, slightly raised, surface rough, with erose to fimbriate, floccose to velvety, heaped and folded, slightly radiated outwards colony, produce brown pigmentation in PDA. Colonies from above dark olivaceous-brown; colonies from below black.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.390025° N, 102.225942° E, 1,186 m, on dead twigs of an unknown deciduous host, 13 March 2023, D.N. Wanasinghe, DWHH23-02 (HKAS146039); ibid., 23.421013° N, 102.227243° E, 517 m, 03 December 2020, DWHH08-03 (HKAS146040).
GenBank numbers: HKAS146039: ITS = PV742912, LSU = PV742968, SSU = PV743021, tef1-α = PV700647, rpb2 = PV700690; HKAS146040: ITS = PV742913, LSU = PV742969, SSU = PV743022, tef1-α = PV700648, rpb2 = PV700691.
Known hosts and distribution: Saprobic on dead stems of Chromolaena odorata in Thailand[69]; on dead twigs of an unknown deciduous host in Yunnan, China (this study).
Notes: Phylogenetic analyses of a combined SSU, LSU, ITS, tef1-α, and rpb2 sequence data demonstrated that two new strains (HKAS146039 and HKAS146040) clustered with two strains of Pseudocapulatispora longiappendiculata (MFLUCC 17-1457 and MFLUCC 17-1452, type strain) with 97% MLBS support (Fig. 22) and is sister to P. clematidis (MFLUCC 17-2063, type strain) with 100% MLBS and 1.00 BYPP support (Fig. 22). Morphologically, the new isolates resemble the type of P. longiappendiculata but slightly differ in having larger ascomata (220–280 μm high × 140–200 μm diam. vs 195–265 μm high × 105–150 μm diam.), overlapping in ascospore size range (23–29 × 7–9.5 μm vs 24–29 × 6–9 μm). Ascospores of the new isolates turn brown with 1–3-septate at maturity, whereas this character was not described for the type[69]. Furthermore, the new isolates can be distinguished from P. clematidis in having smaller ascomata (220–280 μm high × 140–200 μm diam. vs 197–386 μm high × 160–252 μm diam.), smaller asci (80–110 × 13–16 μm vs 68–117 × 12–22 μm), and slightly narrower ascospores (23–29 × 7–9.5 μm vs 22–28 × 7–12 μm)[123]. Therefore, the new isolates are identified as P. longiappendiculata based on phylogenetic evidence.
Macrodiplodiopsidaceae Voglmayr, Jaklitsch & Crous, IMA Fungus 6 (1): 178 (2015)
Pseudochaetosphaeronema Punith., Nova Hedwigia 31 (1-3): 126 (1979)
Notes: Pseudochaetosphaeronema was introduced by Punithalingam[146], with P. larense designated as the type species. Members of this genus have been reported as human pathogens, endophytes, and saprobes[147−149]. Both sexual and asexual morphs have been documented for species in this genus[93,108,132,142,148−154]. The sexual morph is characterized by immersed, uniloculate ascomata; a peridium composed of textura angularis cells; unbranched, septate pseudoparaphyses; 8-spored, bitunicate, fissitunicate asci with a short, distinct pedicel and rounded apex; and fusiform, 1-septate, guttulate ascospores with pointed ends[148]. The asexual morph features globose conidiomata; monophialidic, cylindrical conidiogenous cells; and hyaline, subglobose to oval, aseptate conidia[93,108]. Currently, 17 species are accepted within the genus[89]. The present study introduces two novel species of Pseudochaetosphaeronema collected from the Honghe region of Yunnan, China (Fig. 26).
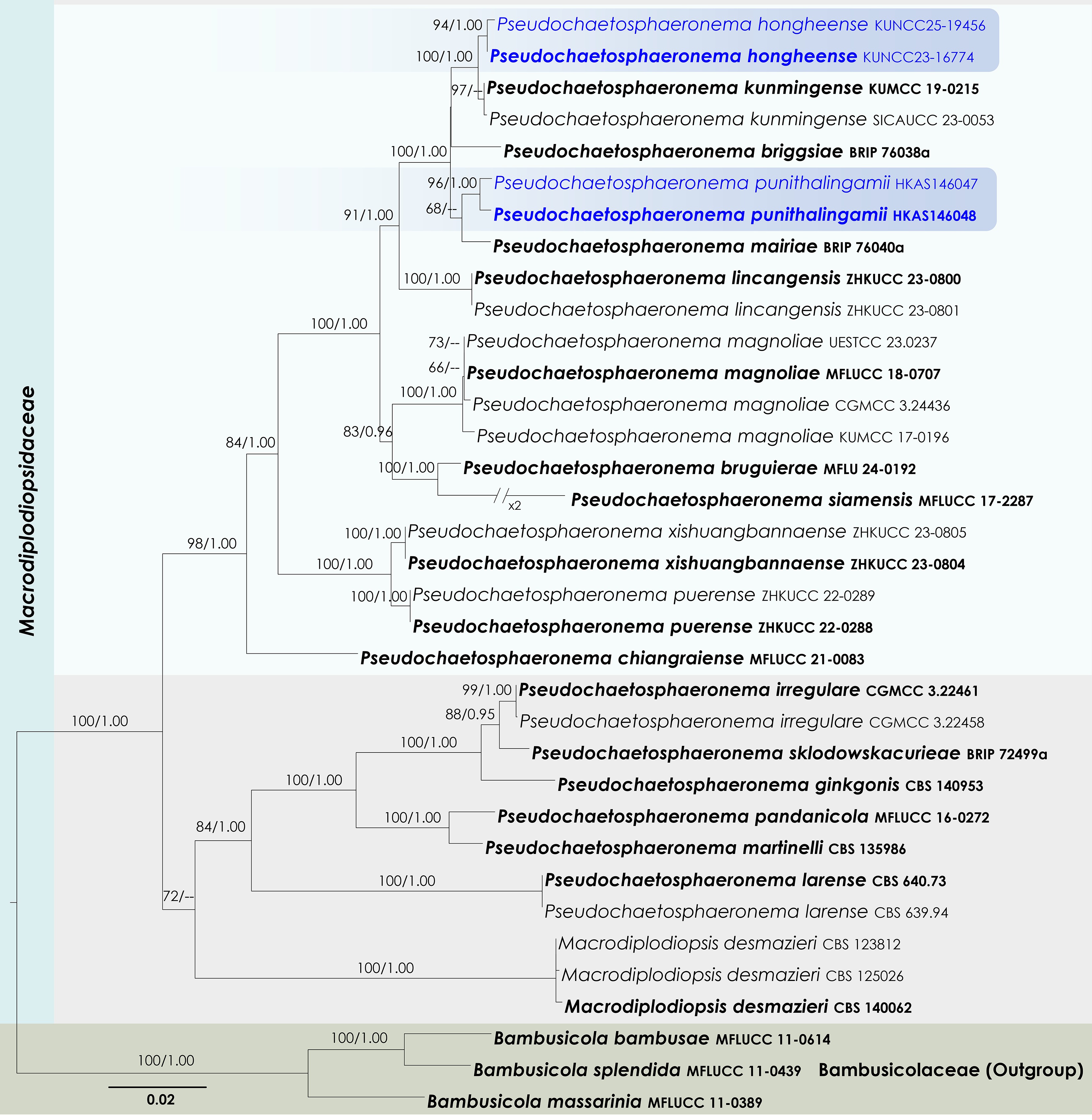
Figure 26.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Bambusicola bambusae (MFLUCC 11-0614), B. massarinia (MFLUCC 11-0389), and B. splendida (MFLUCC 11-0439). The final likelihood value is –14,837.504911. The final alignment included 1,020 unique site patterns, with approximately 30.06% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.238962, C = 0.252036, G = 0.270241, T = 0.238761. The substitution model yielded the following relative rates: AC = 1.527778, AG = 4.223035, AT = 1.623867, CG = 1.375339, CT = 9.995702, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.569470, and the gamma distribution shape parameter (α) was 0.582697. Bayesian inference reached convergence after 34,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009637). A total of 341 trees were sampled, and 256 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,022 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
Pseudochaetosphaeronema hongheense Wanas., Phookamsak & J.C. Xu, sp. nov. (Figs 26 and 27).
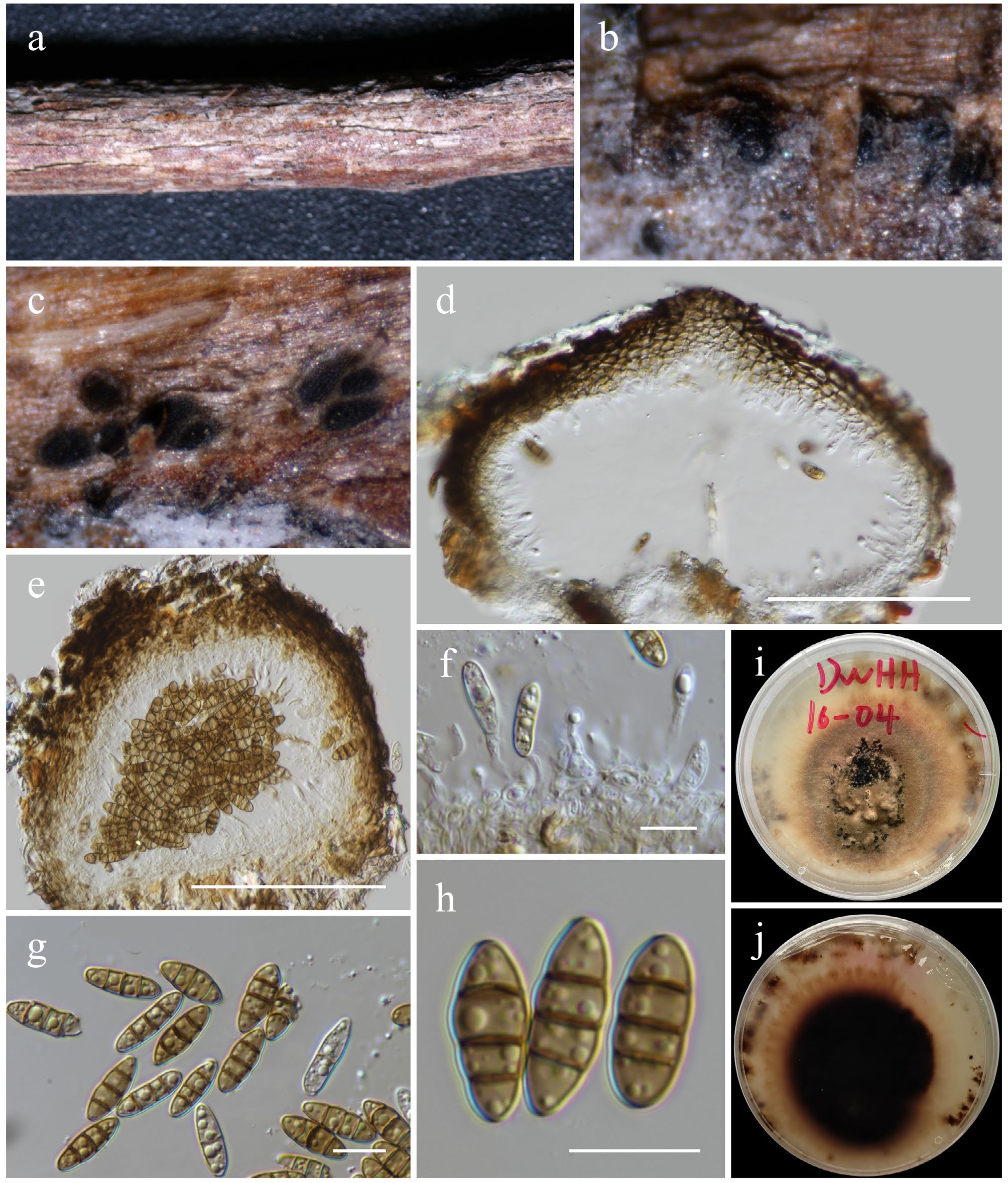
Figure 27.
Pseudochaetosphaeronema hongheense (HKAS146045, holotype). (a), (b) Conidiomata on host. (c)–(e) Sections of conidiomata. (f) Conidiogenous cells. (g), (h) Conidia. (i), (j) Colony on PDA (j from below). Scale bars: (d), (e) = 100 µm, (f)–(h) = 10 µm.
MycoBank No: 859359
Etymology: The specific epithet 'hongheense' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146045
Saprobic on a dead wood of an unknown host. Sexual morph: Undetermined. Asexual morph: Conidiomata 110–150 μm high × 130–200 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA, 25 mm diam. after one week, dense, irregular, flattened, slightly raised at the center, surface rough at the center, smooth towards the margin, with erose to fimbriate edge, fairy fluffy to velvety, covered by pinkish brown and raised, cottony mycelium, profuse growth, with black conidiomata sporulated after 12 weeks; not produce pigmentation in PDA. Colonies from above brown; colonies from below black.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH16-04 (holotype, HKAS146045), ex-type, KUNCC23-16774; ibid., 14 August 2022, DWHH09-01 (HKAS146046), living culture, KUNCC25-19456.
GenBank numbers: HKAS146045: ITS = PV742914, LSU = PV742970, SSU = PV743023, tef1-α = PV700649; HKAS146046: ITS = PV742915, LSU = PV742971, SSU = PV743024, tef1-α = PV700650.
Notes: Multigene phylogenetic analyses based on a combined SSU, LSU, ITS, tef1-α, and rpb2 DNA sequence dataset demonstrated that the two new strains (KUNCC 23-16774 and KUNCC25-19456) formed a subclade (94% MLBS and 1.00 BYPP; Fig. 26), closely related to Pseudochaetosphaeronema kunmingense (100% MLBS and 1.00 BYPP). Morphologically, the new isolates resemble P. kunmingense in possessing black, globose to subglobose conidiomata with apapillate ostioles, monoblastic phialidic conidiogenous cells, and ellipsoidal to fusiform, brown, 3-septate conidia. However, they can be distinguished by having smaller conidiomata (130–200 μm diam. vs 180–250 μm diam.) and slightly larger conidia (13–17 × 5–6.5 μm vs 10–15 × 4–6 μm)[132]. Although the new isolates are morphologically similar to P. kunmingense, their distinct phylogenetic placement supports their recognition as a separate species. Therefore, Pseudochaetosphaeronema hongheense is introduced herein as a new species.
Pseudochaetosphaeronema punithalingamii Wanas., Phookamsak & J.C. Xu, sp. nov. (Figs 26 and 28).

Figure 28.
Pseudochaetosphaeronema punithalingamii (HKAS146048, holotype). (a) Conidiomata on host. (b), (c) Sections of conidiomata. (d)–(f) Conidiogenous cells. (g)–(k) Conidia. Scale bars: (c) = 100 µm, (d)–(k) = 10 µm.
MycoBank No: 859360
Etymology: The species epithet was given in honour of Eliyathamby Punithalingam, who introduced the genus Pseudochaetosphaeronema.
Holotype: HKAS146048
Saprobic on the dead wood of an unknown host. Sexual morph: Undetermined. Asexual morph: Conidiomata 150–230 μm high × 200–300 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.390025° N, 102.225942° E, 1,186 m, on dead twigs of an unknown deciduous host, 13 March 2023, D.N. Wanasinghe, DWHH23-60 (holotype, HKAS146048); ibid., DWHH23-61 (HKAS146047).
GenBank numbers: HKAS146048: ITS = PV742916, LSU = PV742972, SSU = PV743025, tef1-α = PV700651; HKAS146047: ITS = PV742917, LSU = PV742973, SSU = PV743026, tef1-α = PV700652.
Notes: Multigene phylogenetic analyses based on a combined SSU, LSU, ITS, tef1-α, and rpb2 DNA sequence dataset revealed that two new strains (HKAS 146048 and HKAS 146047) formed a subclade (96% MLBS and 1.00 BYPP; Fig. 26), and clustered as sister to Pseudochaetosphaeronema mairiae (68% MLBS). However, morphological comparison with P. mairiae is not possible, as no morphological description was provided when Tan & Shivas[155] introduced the species as an endophyte associated with Geijera salicifolia in Queensland, Australia. Only ITS and LSU sequence data of this species are currently available in GenBank. Based on phylogenetic evidence, the new species Pseudochaetosphaeronema punithalingamii is introduced herein. Pseudochaetosphaeronema punithalingamii also clusters with P. briggsiae, P. kunmingense, and P. hongheense. However, morphological data for P. briggsiae are also unavailable[155]. In contrast, P. punithalingamii, P. kunmingense, and P. hongheense share similar morphological features, particularly ellipsoidal to fusiform, brown, 3-septate conidia. Pseudochaetosphaeronema punithalingamii differs from P. kunmingense and P. hongheense in having larger conidiomata (200–300 μm diam. vs 180–250 μm and 130–200 μm, respectively). Its conidial dimensions (12–16.5 × 5–6.5 µm) are larger than those of P. kunmingense (10–15 × 4–6 μm), but overlap with P. hongheense (13–17 × 5–6.5 µm). Despite some morphological similarities, the phylogenetic distinctiveness of these taxa clearly supports their recognition as separate species.
Mangifericomitaceae Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, fam. nov.
MycoBank No: 859361
Etymology: Referring to the name of the type genus, Mangifericomes
Saprobic on dead or decaying branches, leaves, or wood in terrestrial habitats. Sexual morph: Ascomata immersed to semi-immersed, solitary to gregarious, scattered to clustered, dark brown to black, globose to subglobose, uni-loculate to multi-loculate, emerging in a unique small epidermal clypeus, glabrous, with or without ostiolate, penetrating the host. Peridium thin- to thick-walled of equal thickness, composed of several cell layers of light brown to dark brown, pseudoparanchymatous cells of textura angularis to textura globulosa, or textura prismatica. Hamathecium numerous, composed of filiform, filamentous, hyaline, septate, branched, anastomosed, broad cellular pseudoparaphyses. Asci 8-spored, bitunicate, broadly cylindrical to cylindrical-clavate, pedicellate, apically rounded with an ocular chamber. Ascospores muriform, ellipsoid to broad fusoid, or clavate, brown to dark brown, asymmetrical, multi-septate, sectored, slightly constricted at the central septum, surrounded by a thick, distinct mucilagenous sheath. Asexual morph: Undetermined.
Type genus: Mangifericomes E.F. Yang & Tibpromma.
Notes: The new family Mangifericomitaceae is introduced herein to accommodate the monotypic genus Mangifericomes, with M. hongheensis designated as the type species. Previously, Mangifericomes was treated as a genus incertae sedis within Pleosporales[59,156,157]. In earlier phylogenetic analyses, Mangifericomes formed an unstable lineage: clustering with Brunneoclavispora bambusae (Halotthiaceae) in Yang et al.[156], and forming a separate lineage basal to Pleomonodictydaceae in Du et al.[157]. Although Hyde et al.[59] noted morphological similarities between Mangifericomes and botryosphaeria-like genera, they emphasized their distinction based on muriform ascospores. However, the tree topology in Yang et al.[156] has limitations, particularly due to the unstable placement of Brunneoclavispora bambusae, which appeared distinct from Halotthiaceae, warranting further phylogenetic reassessment[59]. In the present study, phylogenetic analyses based on a concatenated dataset of SSU, LSU, 5.8s, tef1-α, and rpb2 sequences confirmed that Mangifericomes forms a monophyletic clade, distinct from both Halotthiaceae and Pleomonodictydaceae, and clusters as a sister lineage to Corynesporascaceae. Rather than retaining Mangifericomes as an incertae sedis genus within Pleosporales, Mangifericomitaceae fam. nov. is also formally introduced to accommodate this taxon. Morphologically, Mangifericomitaceae differs from Corynesporascaceae in having immersed to semi-immersed, uni- to multi-loculate ascomata emerging beneath a distinct small epidermal clypeus, broadly cylindrical to cylindric-clavate asci embedded in broad cellular pseudoparaphyses, and muriform, sectored, multi-septate ascospores occurring in terrestrial habitats. In contrast, Corynesporascaceae features cleistothecioid, spherical ascomata; obovoid asci surrounded by deliquescent paraphysoids; and 1–3-septate ascospores, or spores with indistinct distosepta, typically found as pathogens or saprobes on leaves[110]. Halotthiaceae is distinguished by its semi-immersed to superficial ascomata, occasionally formed beneath a pseudoclypeus, cylindrical to clavate or subclavate asci embedded in narrowly cellular pseudoparaphyses, and predominantly didymosporous or phragmosporous, thick-walled, septate ascospores. An exception is Brunneoclavispora, which has muriform ascospores with 7–8 transverse and 1–4 longitudinal septa. Members of Halotthiaceae are found in terrestrial, freshwater, and marine habitats as saprobes or pathogens[110,158]. Pleomonodictydaceae, on the other hand, is known solely from hyphomycetous asexual morphs characterized by micro- to semi-macronematous conidiophores (sometimes reduced to mono- or polyblastic conidiogenous cells) and solitary or short-chained, dark brown to black, muriform, verrucose to tuberculate conidia[110]. Therefore, the molecular and morphological evidence supports the establishment of Mangifericomitaceae as a distinct family within Pleosporales.
Mangifericomes E.F. Yang & Tibpromma, Journal of Fungi 8 (2, no. 152): 10.
Notes: Mangifericomes was introduced by Yang et al.[156] as a monotypic genus to accommodate a saprobic fungus occurring on mango (Mangifera indica) in Yunnan, China, and is typified by M. hongheensis. The genus is characterized by immersed or semi-immersed, dark brown to black, globose to subglobose ascomata, sometimes visible apical black neck effuses to host surface, with or without ostiolate, 8-spored, bitunicate, cylindrical-clavate asci, and muriform, ellipsoid, asymmetrical, sectored, multi-transverse and longitudinal septa ascospores, surrounded by mucilaginous sheath[156]. The asexual morph of this genus has not yet been described. Presently, two species are accommodated in this genus[156,157]. In this study, the new species, M. yunnanensis, is introduced as the third species of this genus, of which was collected from different hosts in Yunnan, China.
Mangifericomes yunnanensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 29 and 30).
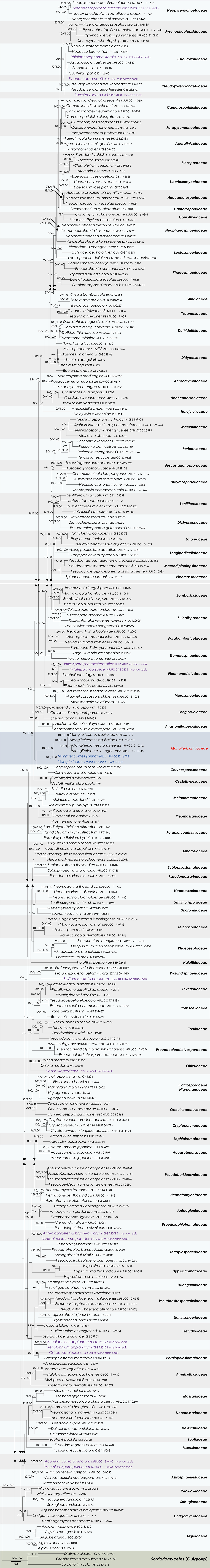
Figure 29.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, 5.8s, tef1-α and rpb2 sequences. The phylogeny is rooted with Diatrype disciformis (AFTOL-ID 927), Graphostroma platystoma (CBS 270.87) and Sordaria fimicola (AFTOL-ID 216). The final likelihood value is −150,266.016088. The final alignment included 2,812 unique site patterns, with approximately 19.07% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.250162, C = 0.244801, G = 0.270528, T = 0.234509. The substitution model yielded the following relative rates: AC = 1.409236, AG = 4.442359, AT = 1.464844, CG = 1.164667, CT = 9.104968, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.409621 and the gamma distribution shape parameter (α) was 0.569317. Bayesian inference reached convergence after 5M generations, when the average standard deviation of split frequencies dropped below 0.01. A total of 50,001 trees were sampled, and 37,501 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 2,814 distinct informative sites. In the resulting phylograms, newly generated sequences are shown in blue and Pleosporales genera incertae sedis are in purple. The new family shown in red.
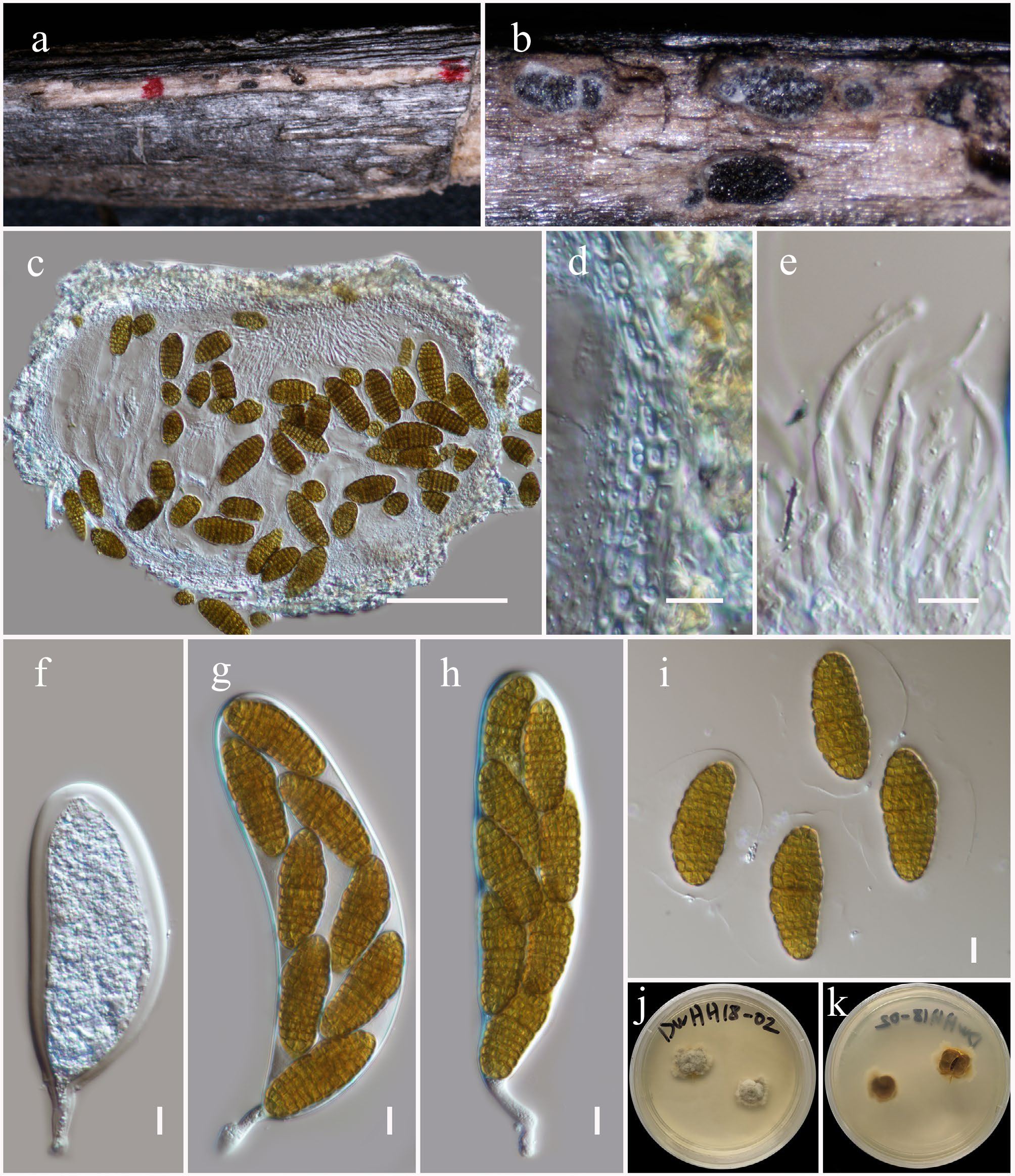
Figure 30.
Mangifericomes yunnanensis (HKAS146058, holotype). (a) Ascomata on the dead woody twigs. (b), (c) Cross-section of ascomata. (d) Peridium. (e) Pseudoparaphyses. (f)–(h) Asci. (i) Ascospores. (j), (k) Colony on MEA (k from the bottom). Scale bars: (c) = 100 µm, (d)–(i) = 10 µm.
MycoBank No: 859362
Etymology: The specific epithet 'yunnanensis' refers to Yunnan Province, where the holotype was collected.
Holotype: HKAS146058
Saprobic on dead twigs of an unknown deciduous host. Sexual morph: Ascomata 250–430 μm high, 390–650 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA at room temperature (25 °C) under normal light, reaching 22–25 mm diam. after two weeks. Colonies dense, circular, flattened, slightly raised, surface rough, with erose to fimbriate, fluffy, with hyphal tufts, produced small, black droplets, did not produce pigmentation in PDA. Colonies from above are white, with brown hyphae at the edge; colonies from below are cream at the margin, and brown to dark brown towards the center.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH18-02 (holotype, HKAS146058), ex-type, KUNCC23-16778; ibid., DWHH18-02-2 (HKAS146059).
GenBank numbers: KUNCC23-16778: ITS = PV742918, LSU = PV742974, SSU = PV743027, tef1-α = PV700653, rpb2 = PV700692; HKAS146059: ITS = PV742919, LSU = PV742975, SSU = PV743028, tef1-α = PV700654, rpb2 = PV700693.
Notes: Phylogenetically, two new strains (KUNCC 23-16778 and HKAS 146059) formed a subclade (100% MLBS and 1.00 BYPP; Fig. 29), positioned basal to Mangifericomes aquilariae and M. hongheensis. Morphologically, the new isolates resemble M. aquilariae and M. hongheensis in producing muriform, multi-septate ascospores. However, they can be clearly distinguished from both species based on differences in the dimensions of ascomata, asci, and ascospores, as well as peridial structure and ascospore septation patterns. The new isolates differ from M. aquilariae in having larger ascomata (250–430 µm high × 390–650 µm diam. vs 280–460 µm high × 250–510 µm diam.), non-ostiolate ascomata with a hyaline to pale brown peridium composed of thick-walled cells of textura angularis to textura prismatica or textura globulosa, smaller asci (160–220 × 34–50 µm vs 170–265 × 32–50 µm), and ascospores with 11–12 transverse septa and 2–5 longitudinal septa. In contrast, M. aquilariae has ostiolate ascomata and ascospores with 10–13 transverse septa and 3–6 longitudinal septa[157]. Compared to M. hongheensis, the new isolates have larger ascomata (250–430 × 390–650 µm vs 160–230 × 220–300 µm), non-ostiolate and pale peridia (vs ostiolate, dark brown to black), larger asci (160–220 × 34–50 µm vs 90–120 × 8–12 µm), and larger ascospores (45–57 × 18–27 µm vs 14–17 × 4–8 µm), with distinct septation (11–12 transverse and 2–5 longitudinal septa vs 7–11 transverse and 5–8 longitudinal septa)[156]. Based on phylogenetic evidence and clear morphological distinctiveness, Mangifericomes yunnanensis is introduced as a novel species in the present study.
Massarinaceae Munk, Friesia 5 (3-5): 305.
Helminthosporium Link, Mag. Neuesten Entdeck. Gesammten Naturk. Ges. Naturf. Freunde Berlin 3 (1): 10.
Notes: Helminthosporium was established by Link[159], with H. velutinum designated as the type species. The genus is characterized by immersed ascomata, an inconspicuous ostiole that does not protrude above the cortical surface, clavate to fusoid asci, and large, hyaline to dark brown, fusoid, subellipsoid to obovoid ascospores surrounded by a thick gelatinous sheath. The asexual morph is distinguished by effuse to punctiform colonies and subhyaline to brown, obclavate, obpyriform to lageniform conidia[159]. Helminthosporium species have been reported as saprobes and plant pathogens on various hosts and are widely distributed worldwide[148,160−167]. Currently, 225 species are accepted under Helminthosporium (out of 521 species epithets), although most lack sequence data[168]. In the present study, a new species of Helminthosporium is introduced.
Helminthosporium hongheense Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 31 and 32).

Figure 31.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α and rpb2 sequence analyses. The tree is rooted to Cyclothyriella rubronotata (CBS 121892, CBS 141486). The final likelihood value is –35,556.858868. The final alignment included 2,182 unique site patterns, with approximately 38.1% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.237453, C = 0.256825, G = 0.269094, T = 0.236628. The substitution model yielded the following relative rates: AC = 1.452161, AG = 3.722496, AT = 1.468538, CG = 0.961651, CT = 7.489299 and GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.518695 and the gamma distribution shape parameter (α) was 0.562000. Bayesian inference reached convergence after 1,923,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009981). A total of 19,231 trees were sampled, and 14,424 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 2,183 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.

Figure 32.
Helminthosporium hongheense (HKAS146049, holotype). (a)–(d) Dark brown or black colonies on natural substrate. (e)–(g) Conidiophores. (h)–(k) Conidia. Scale bars: (f) = 100 µm, (e), (g)–(k) = 10 µm.
MycoBank No: 859363
Etymology: The specific epithet 'hongheense' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146049
Saprobic on a dead wood of a Fagaceae hosts. Sexual morph: Undertermined. Asexual morph: Hyphomycetous. Colonies on the host, black, effuse, velvety, appear as clusters. Mycelium is composed of branched, septate, smooth, and thick-walled hyphae. Conidiophores 220–330 × 8–14 µm (
$\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Jianshui County, Chenguan, 23.500871° N, 102.883429° E, 2,062 m, on dead twigs of Fagaceae sp., 15 March 2019, D.N. Wanasinghe, DW0527-15 (holotype, HKAS146049); ibid., Puxiongxiang 23.521432° N, 103.039049° E, 1,917 m, on dead twigs of Quercus sp., DW0528-07 (HKAS146050).
GenBank numbers: HKAS146049: ITS = PV742920, LSU = PV742976, SSU = PV743029, tef1-α = PV700655; HKAS146050: ITS = PV742921, LSU = PV742977, SSU = PV743030, tef1-α = PV700655.
Notes: In multigene phylogenetic analyses based on a combined SSU, LSU, ITS, tef1-α, and rpb2 dataset, two newly collected strains (HKAS 146049 and HKAS 146050) formed a monophyletic clade (100% MLBS and 1.00 BYPP; Fig. 31) sister to the type strain of Helminthosporium syzygii (CPC 35312). Morphologically, Helminthosporium syzygii and our new collections are similar in colony characteristics, conidiophore structure, conidiogenous cells and conidial shape, with overlapping measurements[169]. The study initially aimed to clarify whether these strains could be considered conspecific with H. syzygii. However, multiple lines of evidence argue against this. The comparison of the ITS region revealed a 5% nucleotide difference between our strains and H. syzygii (29/572 bp, including three gaps). Helminthosporium syzygii was isolated from bark cankers on Syzygium sp. (Myrtaceae) in the Eastern Cape Province of South Africa, while our collections were obtained as saprobes on Quercus sp. (Fagaceae) hosts in Yunnan, China. These differences in host association, lifestyle, geographic origin, and molecular divergence suggest that our collections represent a distinct species. Pending additional morphological evidence (particularly from sexual morphs), our two collections are designated as Helminthosporium hongheense sp. nov.
Parabambusicolaceae Kaz. Tanaka & K. Hiray., Studies in Mycology 82: 115.
Neomultiseptospora N. Xie, Phookamsak & Hongsanan, Journal of Fungi 8 (2, no. 109): 8.
Notes: Xie et al.[25] introduced Neomultiseptospora to accommodate a single species, N. yunnanensis, collected from dead branches of bamboo in Yunnan Province, China. Multigene phylogenetic analyses demonstrated that Neomultiseptospora clustered with the genera Multiseptospora and Scolecohyalosporium. Morphologically, Neomultiseptospora resembles Multiseptospora in possessing phragmosporous, hyaline, multi-septate ascospores but differs in forming raised, hemispherical to subconical ascomata, whereas Multiseptospora develops globose to subglobose ascomata covered with dark, hair-like hyphae and embedded within the host tissue[170]. Although Neomultiseptospora is phylogenetically close to Scolecohyalosporium, it can be distinguished by having phragmosporous ascospores, compared to the scolecosporous ascospores of Scolecohyalosporium[25,67,171]. Morphologically, Neomultiseptospora also closely resembles Parabambusicola, sharing features such as hemispherical to subconical ascomata and hyaline, fusiform, 5-septate ascospores, and occurrence on bamboo[25,172,173]. However, phylogenetically, the two genera are distinct. In the present study, Neomultiseptospora hongheensis is introduced as the second species of the genus, along with an additional collection representing N. yunnanensis.
Neomultiseptospora hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 33 and 34).

Figure 33.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α and rpb2 sequences. The phylogeny is rooted with Paradendryphiella salina (CBS 142.60), Pleospora tarda (CBS 714.68) and Stemphylium vesicarium (CBS 191.86). The final likelihood value is –25,430.003747. The final alignment included 1,492 unique site patterns, with approximately 18.07% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.234317, C = 0.258507, G = 0.272066, T = 0.235111. The substitution model yielded the following relative rates: AC = 1.103972, AG = 2.858045, AT = 1.446677, CG = 1.147080, CT = 5.894779, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.7274, and the gamma distribution shape parameter (α) was 0.569941. Bayesian inference reached convergence after 761,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009992). A total of 7,611 trees were sampled, and 5,709 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,493 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
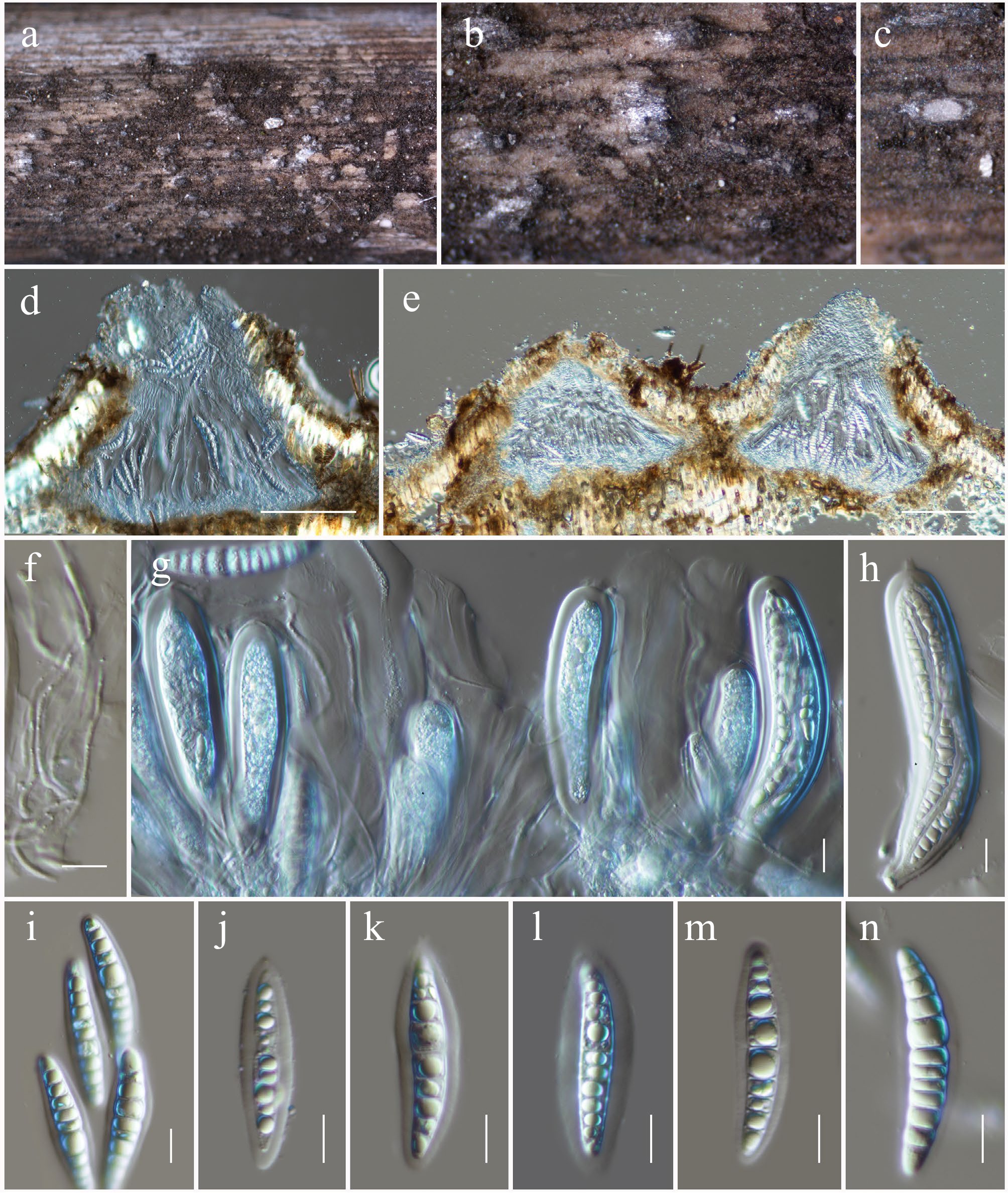
Figure 34.
Neomultiseptospora hongheensis (HKAS146053, holotype). (a), (b) Ascomata on the dead woody twigs. (c)–(e) Cross-section of ascomata. (f) Pseudoparaphyses. (g), (h) Asci. (i)–(n) Ascospores. Scale bars: (d), (e) = 100 µm, (f)–(n) = 10 µm.
MycoBank No: 859364
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146053
Saprobic on dead sheath of a Poacea host. Sexual morph: Ascomata 190–250 µm high × 250–320 µm diam. µm (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead sheath of a Poacea sp., 13 March 2023, D.N. Wanasinghe, DWHH23-80 (holotype, HKAS146053); ibid., DWHH23-58-03 (isotype, HKAS146054); ibid., 23.421013° N, 102.227243° E, 517 m, 03 December 2020, DWHH17-02 (HKAS146056); ibid., DWHH17-02-1 (HKAS146055).
GenBank numbers: HKAS146053: ITS = PV742922, LSU = PV742978, SSU = PV743031, tef1-α = PV700657; HKAS146054: ITS = PV742923, LSU = PV742979, SSU = PV743032, tef1-α = PV700658; HKAS146056: ITS = PV742924, LSU = PV742980, SSU = PV743033, tef1-α = PV700659; HKAS146055: ITS = PV742925, LSU = PV742981, SSU = PV743034, tef1-α = PV700660.
Notes: Four strains of Neomultiseptospora hongheensis formed a monophyletic clade (100% MLBS; Fig. 33), sister to N. yunnanensis. Morphologically, Neomultiseptospora hongheensis fits well within the generic circumscription of Neomultiseptospora, sharing similar characteristics of the ascomata, peridium, asci, and ascospores. However, the two species can be distinguished based on ascospore size and septation. Neomultiseptospora hongheensis possesses comparatively larger ascospores (40–52 × 7.5–10 μm) with 10–11 septa, whereas N. yunnanensis has smaller ascospores (22–27 × 5–8.5 μm) with (4–)5 septa[25].
Neomultiseptospora yunnanensis Phookamsak, Hongsanan & N. Xie, Journal of Fungi 8 (2, no. 109): 8 (Figs 33 and 35).
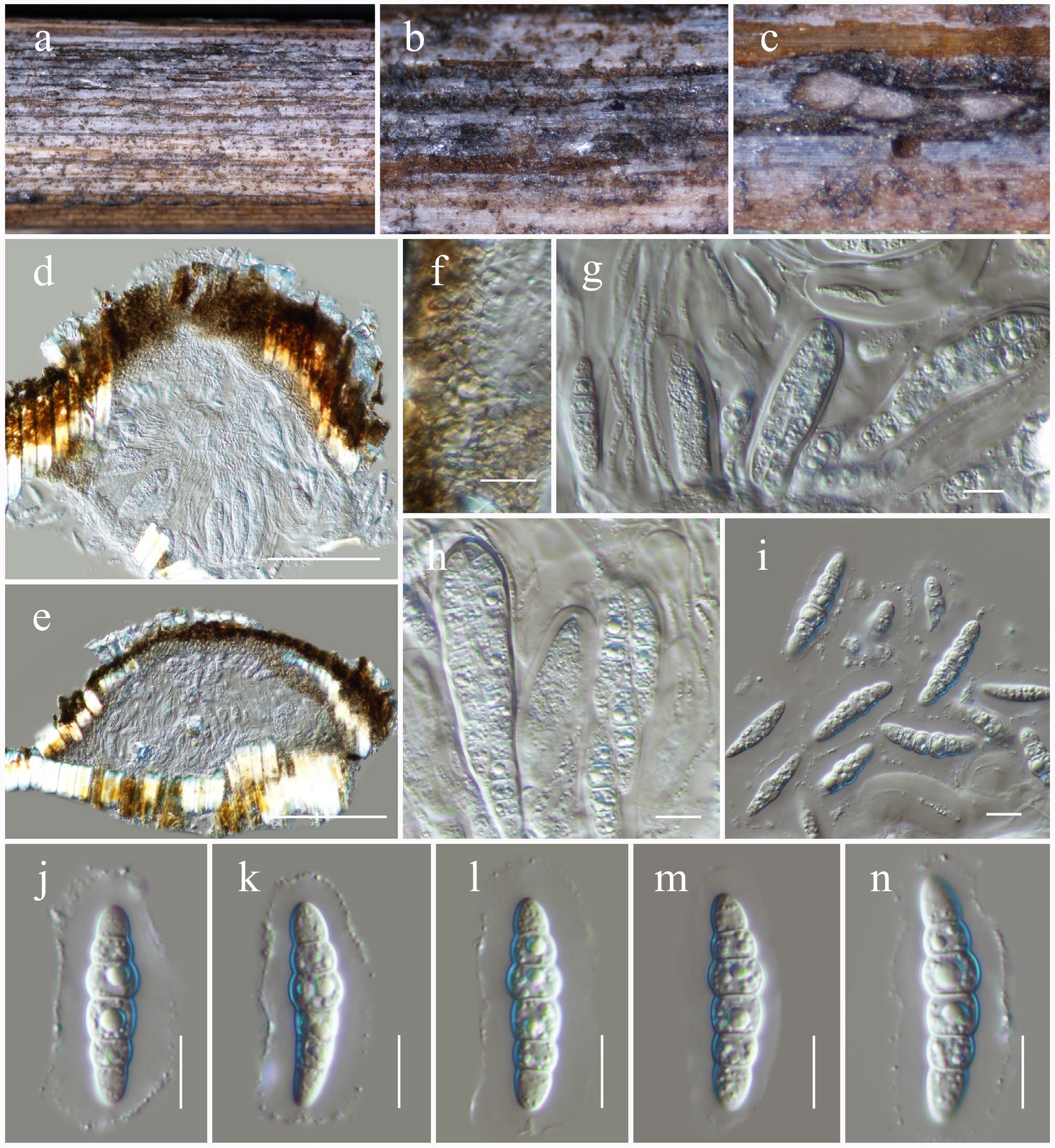
Figure 35.
Neomultiseptospora yunnanensis (HKAS146057). (a), (b) Ascomata on the dead woody twigs. (c)–(e) Cross-section of ascomata. (f) Peridium. (g), (h) Asci and pseudoparaphyses. (i)–(n) Ascospores. Scale bars: (d), (e) = 100 µm, (f)–(n) = 10 µm.
MycoBank No: 559390
Saprobic on dead culms of a Poaceae sp. Sexual morph: Ascomata 100–150 µm high × 260–320 µm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead culms of a Poaceae sp., 13 March 2023, D.N. Wanasinghe, DWHH23-67 (HKAS146057).
GenBank numbers: HKAS146057: ITS = PV742926, LSU = PV742982, SSU = PV743035, tef1-α = PV700661.
Known hosts and distribution: Saprobic on a dead stem of bamboo from a terrestrial environment in Yunnan, China[25]; on dead culms of a Poaceae sp. in Yunnan, China (this study).
Notes: Phylogenetic analyses supported our isolate (HKAS146057) as forming a monophyletic clade with the type strain of Neomultiseptospora yunnanensis. Given the overlapping morphological characteristics[25] and strong phylogenetic affinity (100% MLBS; Fig. 33), our collection is recognized as another collection of Neomultiseptospora yunnanensis from Honghe, Yunnan.
Phaeosphaeriaceae M.E. Barr, Mycologia 71: 948.
Leptospora Rabenh., Hedwigia 1 (18): 116.
Notes: Leptospora is characterized by large, flask-shaped ascomata with long, cylindrical asci and phragmosporous to scolecosporous, fusiform, cylindric-clavate to filiform ascospores. Members of this genus have also been reported to produce a reddish-purple pigment that stains the host tissue around the ostiole[131,174,175]. Hyde et al.[131] designated a reference specimen for the type, L. rubella, and confirmed the placement of Leptospora in Phaeosphaeriaceae based on phylogenetic analyses of ITS, LSU, and SSU sequence data. Species of this genus have been increasingly reported in recent years, mostly from terrestrial habitats[69,123,167,176,177]. Currently, 18 species are accepted in Index Fungorum[89], but sequence data are available in GenBank for only nine species.
Leptospora phraeana Mapook & K.D. Hyde, Fungal Diversity 101: 65 (Figs 36 and 37).
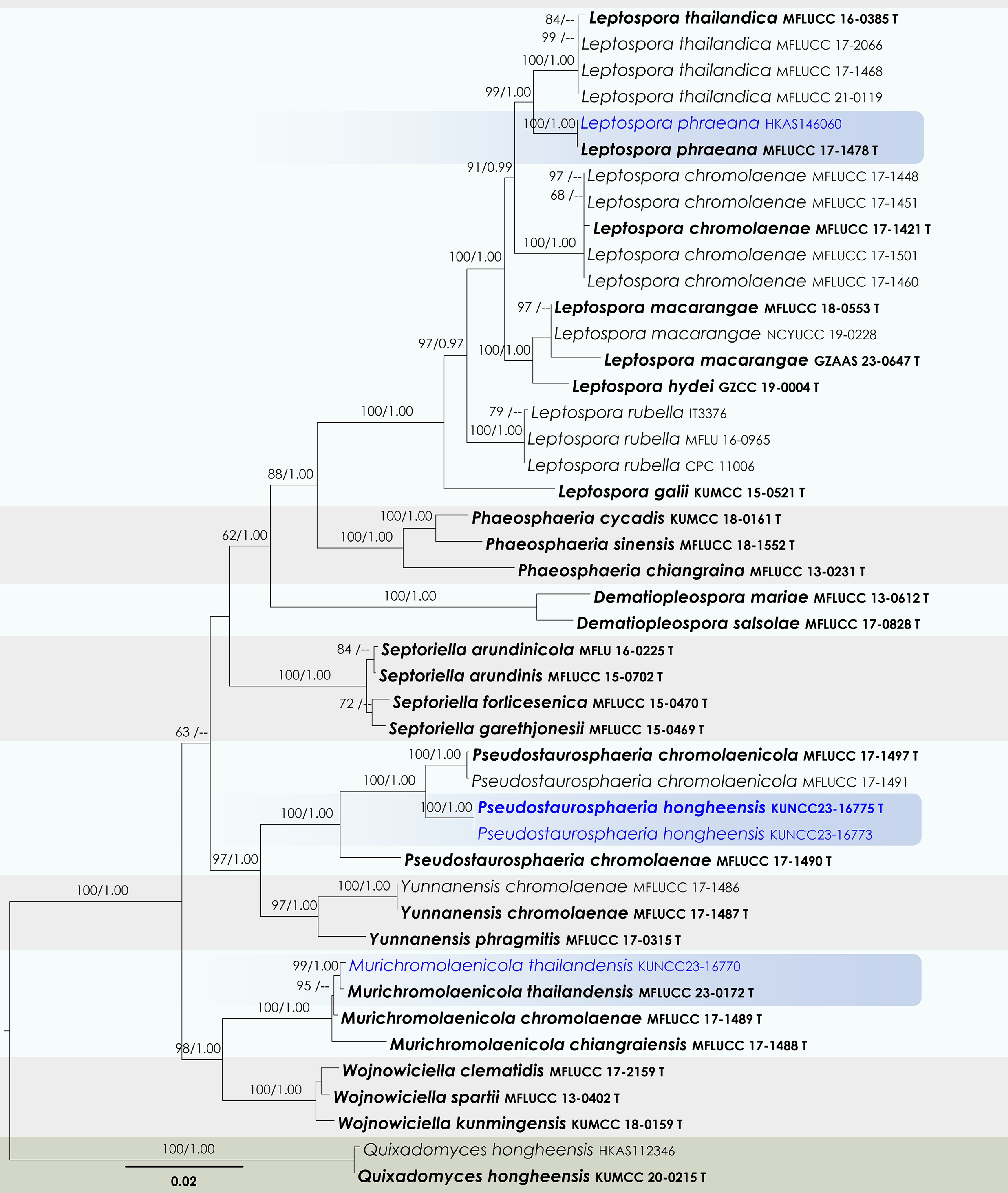
Figure 36.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Quixadomyces hongheensis (HKAS112346, KUMCC 20-0215). The final likelihood value is –11,594.436322. The final alignment included 658 unique site patterns, with approximately 9.44% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.243939, C = 0.239658, G = 0.264036, T = 0.252368. The substitution model yielded the following relative rates: AC = 0.848781, AG = 2.550385, AT = 2.306846, CG = 0.490775, CT = 5.900303 and GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.719615, and the gamma distribution shape parameter (α) was 0.686992. Bayesian inference reached convergence after 221,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009792). A total of 2,211 trees were sampled, and 1,659 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 659 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
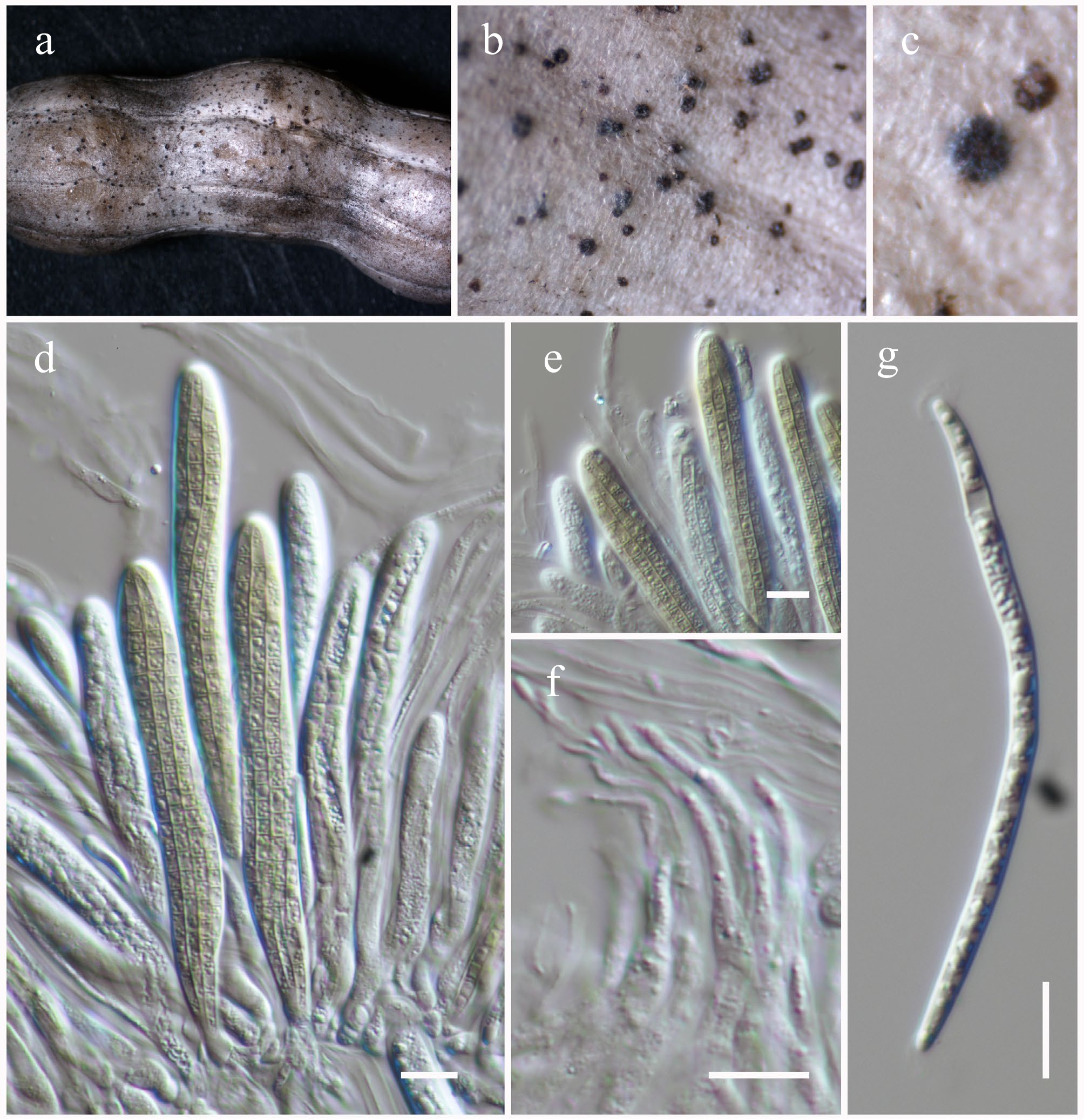
Figure 37.
Leptospora phraeana (HKAS146060). (a)–(c) Ascomata on the dead woody twigs. (d), (e) Asci. (f) Pseudoparaphyses. (i) Ascospore. Scale bars: (d)–(g) = 10 µm.
MycoBank No: 557291
Saprobic on dead pods of a Fabaceae sp. Sexual morph: Ascomata immersed, causing raised regions on the host surface, scattered to gregarious, globose to subglobose, non-ostiolate. Peridium 3–4-layered, comprising thick-walled, hyaline cells of textura angularis to textura globosa. Pseudoparaphyses 1.5–2.5 μm wide, hyaline, septate, branched, cellular. Asci 80–95 × 9–12 μm (
$\overline{\rm x} $ $\overline{\rm x} $ Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead pods of a Fabaceae sp., 14 August 2022, D.N. Wanasinghe, DWHH22-05 (HKAS146060).
GenBank numbers: HKAS146060: ITS = PV742927, LSU = PV742983, SSU = PV743036, tef1-α = PV700662.
Known hosts and distribution: Saprobic on dead stems of Chromolaena odorata in Phrae Province, Thailand[69]; on dead pods of a Fabaceae sp. in Yunnan, China (this study).
Notes: Leptospora phraeana was introduced by Mapook et al.[69] based on a collection from Phrae Province, Thailand, on dead stems of Chromolaena odorata (Asteraceae). This species is characterized by semi-immersed, globose to obpyriform ascomata; hyaline periphysis-like cells near the ostiolar neck; a peridium composed of textura angularis cells; cylindrical, septate, and branched pseudoparaphyses; cylindric-clavate asci with a short pedicel and a prominent ocular chamber; and fasciculate, scolecosporous, pale brown to yellowish-brown, cylindrical to broadly filiform, multi-septate ascospores with polar appendages. During our survey in the Honghe region of Yunnan, China, a fungus exhibiting morphological features consistent with Leptospora phraeana was collected. Phylogenetic analyses further supported our isolate as monophyletic with the type strain of Leptospora phraeana. Given the overlapping morphological characteristics and strong phylogenetic affinity (100% MLBS and 1.00 BYPP; Fig. 36), our collection is recognized as a new record of Leptospora phraeana. Notably, our specimen was isolated from dead pods of a Fabaceae species, thereby expanding the known host range and geographic distribution of this species.
Murichromolaenicola Mapook & K.D. Hyde, Fungal Diversity 101: 71.
Notes: Murichromolaenicola was introduced by Mapook et al.[69] with two species, M. chiangraiensis and M. chromolaenae, isolated from dead stems of Chromolaena odorata in Thailand. These taxa are characterized by semi-immersed to superficial, solitary or scattered ascomata; fissitunicate, cylindric-clavate asci and overlapping, 1–2-seriate, initially hyaline to golden-yellow, uniseptate ascospores[69]. The asexual morphs are defined by pycnidial, uniloculate, globose conidiomata; phialidic, ampulliform to cylindrical, hyaline, unbranched conidiogenous cells and ellipsoid to broadly fusiform, muriform conidia that are 5–7-transversely septate with 1–2 vertical septa, brown in color, and bearing polar appendages[69]. Subsequently, Htet et al.[178] described Murichromolaenicola thailandensis from the same host in Thailand, while Sun et al.[167] reported M. dendrobii on twigs of Dendrobium nobile, also from Thailand. In the present study, Murichromolaenicola thailandensis is reported from Yunnan, China, representing a new geographical record for both the genus and species.
Murichromolaenicola thailandensis Htet, Mapook & K.D. Hyde, Phytotaxa 618 (2): 126 (2023) (Figs 36 and 38).
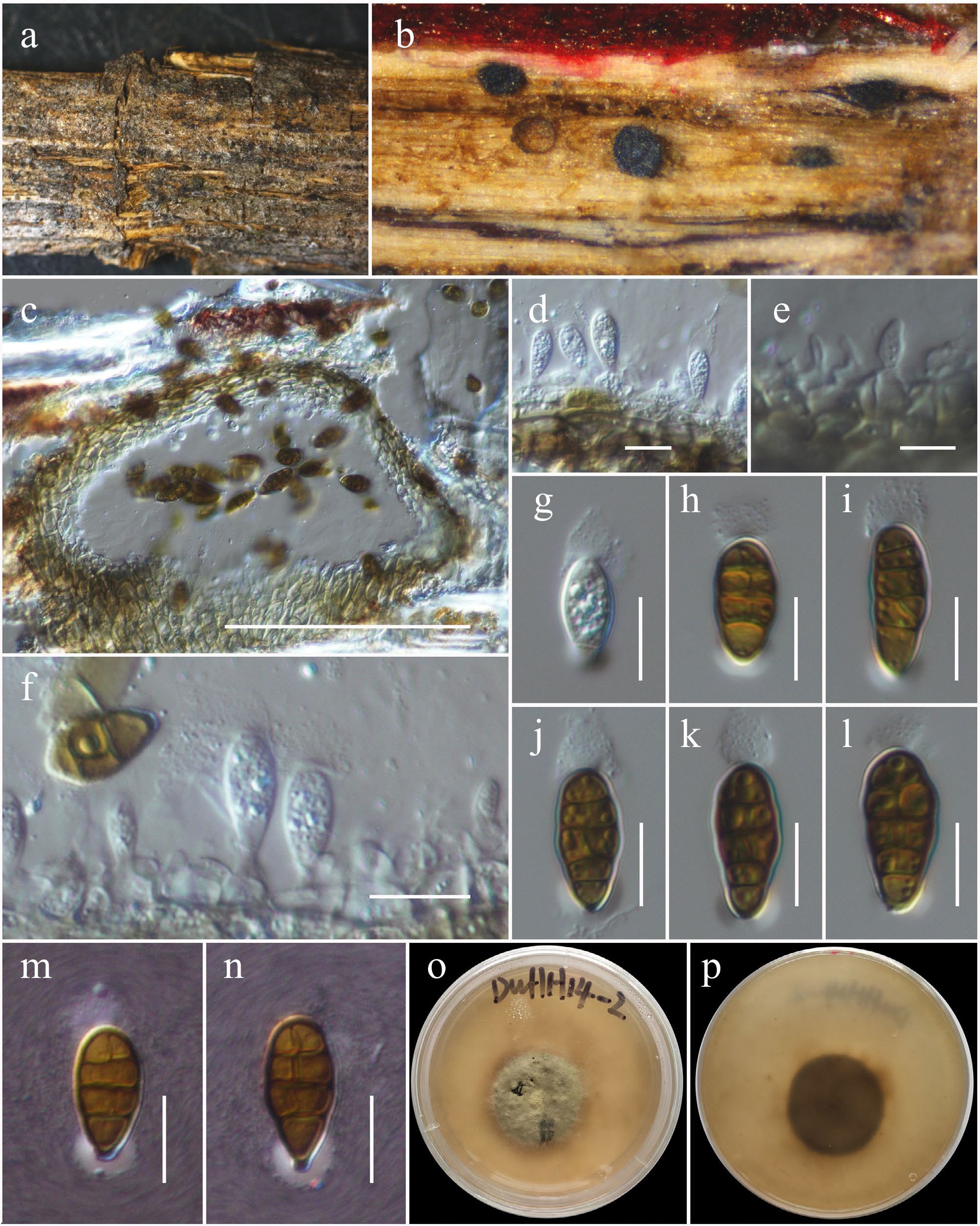
Figure 38.
Murichromolaenicola thailandensis (HKAS146061). (a), (b) Conidiomata on host. (c) Section of a conidioma. (d)–(f) Conidiogenous cells. (g)–(n) Conidia (m and n with the Indian Ink). (o), (p) Colony on MEA (p from the bottom). Scale bars: (c) = 50 µm, (d)–(f) = 10 µm, (g)–(n) = 5 µm.
MycoBank No: 849500
Saprobic on dead twigs of an unidentified host. Asexual morph: Conidiomata 100–120 µm high × 150–190 µm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA at room temperature (25 °C) under normal light, reaching 18–20 mm diam. after two weeks. Colonies dense, circular, slightly raised to low convex, surface smooth to slightly rough, with edge entire, floccose, do not produce pigmentation in PDA. Colonies from above yellow-grey to yellow-pale green, with brown, hyphae at edge; colonies from below black at the margin, brown to dark brown towards the center, with brown, sparse mycelia at the margin.
Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH14-02 (HKAS146061), living culture KUNCC23-16770.
GenBank numbers: KUNCC23-16770: ITS = PV742928, LSU = PV742984, SSU = PV743037, tef1-α = PV700663.
Known hosts and distribution: Saprobic on dead stems of Chromolaena odorata in Chiang Rai Province, Thailand[178]; on dead twigs of an unknown deciduous host in Yunnan, China (this study).
Notes: Based on morphological comparisons, Murichromolaenicola thailandensis closely resembles M. chromolaenae in having immersed conidiomata, yellowish brown to brown cells of textura angularis, and yellowish brown to brown ascospores with both transverse and longitudinal septa[178]. However, Murichromolaenicola thailandensis differs from M. chromolaenae in comparatively smaller conidiomata (130–150 × 140–160 µm vs 200–235 × 195–230 µm) and smaller conidia with a gelatinous cap (10–20 × 5–10 µm vs 14–25 × 6.5–11 µm). The conidia of Murichromolaenicola chromolaenae are ellipsoid to broadly fusiform, with 5–7 transverse septa and polar appendages at both ends, whereas M. thailandensis produces oblong to oval or obovoid conidia, rounded at both ends, with 3–5 transverse septa and a gelatinous cap at one end. The morphology of the conidiomata, conidiogenous cells, and conidia in our new collection (HKAS146061) corresponds well with the original description of Murichromolaenicola thailandensis, further supported by its placement in a monophyletic clade (99% MLBS and 1.00 BYPP; Fig. 36) alongside other reported strains of the species. Therefore, our isolate is recognized as a new record of Murichromolaenicola thailandensis, extending its known geographical distribution to Yunnan, China.
Pseudostaurosphaeria Mapook & K.D. Hyde, Fungal Diversity 101: 81.
Notes: Pseudostaurosphaeria was established by Mapook et al.[69] to accommodate P. chromolaenae and P. chromolaenicola. The genus is characterized by solitary, immersed to semi-immersed or superficial, uniloculate conidiomata; holoblastic, ampulliform to cylindrical or oblong, hyaline conidiogenous cells, and globose, oblong to obovoid conidia with 1–2 transverse septa[69]. The asexual morph of Pseudostaurosphaeria resembles that of Staurosphaeria, particularly in producing brown conidia with a transverse septum and vertical septa that eventually divide the conidium into four sections. However, the two genera are not closely related phylogenetically[44]. Pseudostaurosphaeria is closely related to Yunnanensis in our phylogenetic analysis (Fig. 36), though the latter differs by producing muriform, ellipsoidal to obovoid conidia with three transverse septa and a single vertical septum. In contrast, Pseudostaurosphaeria conidia are globose to obovoid with only 1–2 transverse septa[69,179]. Both previously known species in the genus were reported from dead stems of Chromolaena odorata in Chiang Rai, Thailand. In this study, a third species is introduced: Pseudostaurosphaeria hongheensis, collected from Yunnan, China, thereby extending the known diversity and distribution of the genus.
Pseudostaurosphaeria hongheensis Wanas., Phookamsak & J.C. Xu, sp. nov. (Figs 36 and 39).
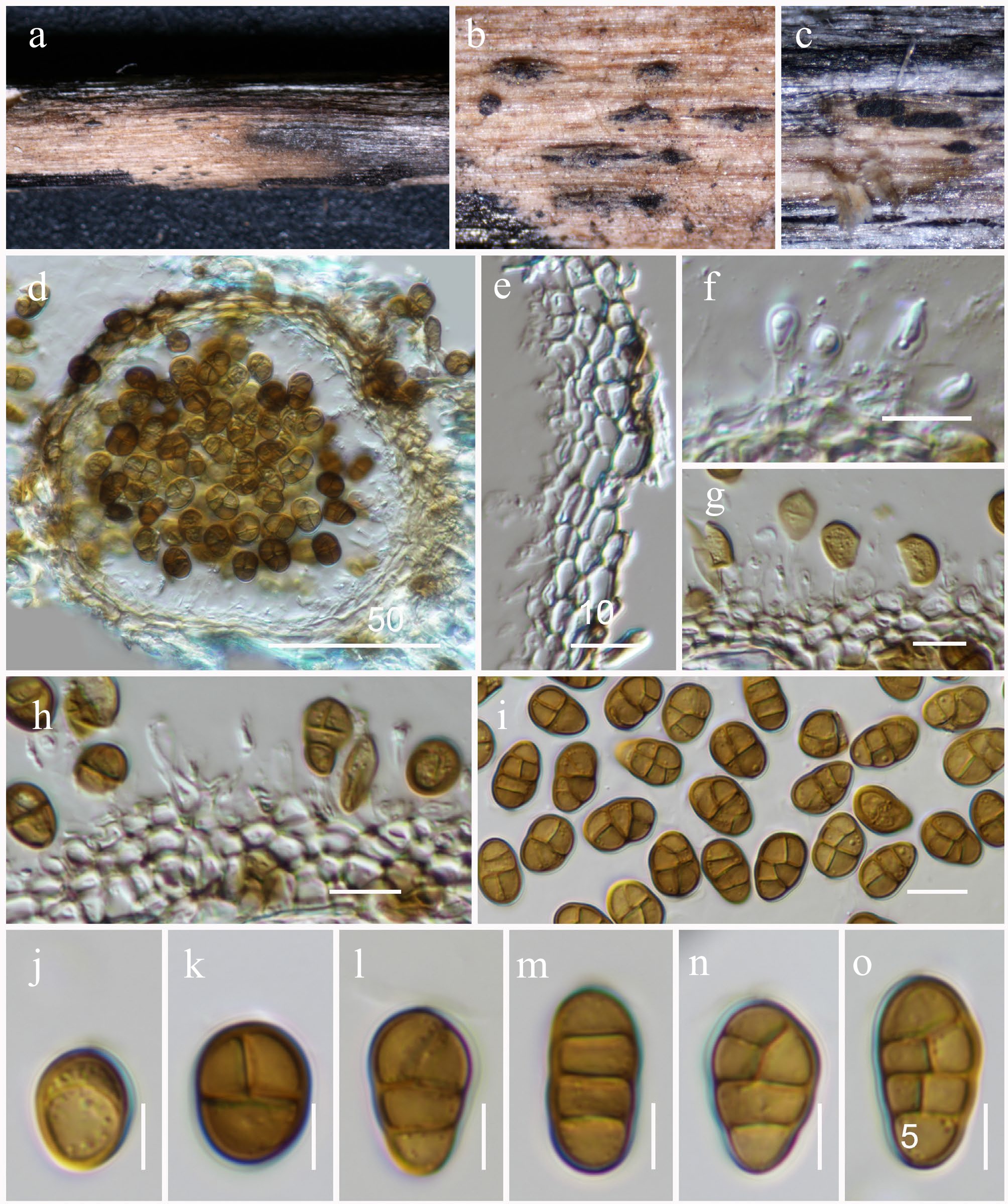
Figure 39.
Pseudostaurosphaeria hongheensis (HKAS146062, holotype ). (a)–(c) Conidiomata on host. (d) Section of a conidioma. (e) Pycnidial wall. (f)–(h) Conidiogenous cells. (i)–(o) Conidia. Scale bars: (d) = 50 µm, (e)–(h) = 10 µm, (i)–(o) = 5 µm.
MycoBank No: 859365
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146062
Saprobic on dead twigs of an unidentified host. Asexual morph: Conidiomata 90–110 µm high × 100–130 µm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Culture characteristics: Colonies on PDA at room temperature (25 °C) under normal light, reaching 20–25 mm diam. after two weeks. Colonies circular, dense, and floccose with a cottony to velvety texture, whitish to creamy coloration across the surface, with a slightly raised central zone and a lighter, slightly wrinkled area in the middle. The colony margin is regular and well-defined; colonies from below are pigmented, showing a dark brown center that diffuses outward into lighter brown zones toward the margin. The pigment production is intense and localized at the colony center.
Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead twigs of an unknown deciduous host, 03 December 2020, D.N. Wanasinghe, DWHH17-01 (holotype, HKAS146062), ex-type, KUNCC23-16775; ibid., DWHH16-03 (HKAS146063), living culture KUNCC23-16773.
GenBank numbers: KUNCC23-16775: ITS = PV742929, LSU = PV742985, SSU = PV743038, tef1-α = PV700664; KUNCC23-16773: ITS = PV742930, LSU = PV742986, SSU = PV743039, tef1-α = PV700665.
Notes: Morphologically, Pseudostaurosphaeria hongheensis conforms well to the generic circumscription of Pseudostaurosphaeria in terms of its conidiomata, conidiogenous cells and conidial features. In our multigene phylogenetic analysis, Pseudostaurosphaeria hongheensis forms a sister relationship with P. chromolaenicola within the genus (100% MLBS and 1.00 BYPP; Fig. 36). While both species exhibit similar conidiomatal structure and conidiogenous cell morphology, their conidia differ in septation and size. Pseudostaurosphaeria chromolaenicola produces conidia with a single transverse septum and one vertical septum at maturity, whereas P. hongheensis has conidia with three transverse septa and 1–2 vertical septa. The conidia of Pseudostaurosphaeria hongheensis (10–13 × 7–9 μm) are notably longer and wider than those of P. chromolaenicola (6.5–9 × 5–6.5 μm).
Roussoellaceae J.K. Liu, Phookamsak, D.Q. Dai & K.D. Hyde, Phytotaxa 181 (1): 7 (2014)
Neoroussoella Jian K. Liu, Phookamsak & K.D. Hyde, Phytotaxa 181 (1): 21 (2014)
Notes: Neoroussoella was introduced by Liu et al.[180] to accommodate N. bambusae, which was collected from dead branches of Bambusa species in Thailand. Species of Neoroussoella are morphologically distinct from those of Roussoella by possessing unilocular ascomata and a coelomycetous asexual morph that produces hyaline to pale brown, smooth-walled conidia[180]. Currently, there are 18 accepted species accommodated in this genus[93,108,123,180−186].
Neoroussoella bambusae Phookamsak, J.K. Liu & K.D. Hyde, Phytotaxa 181 (1): 23 (2014) (Figs 40 and 41).
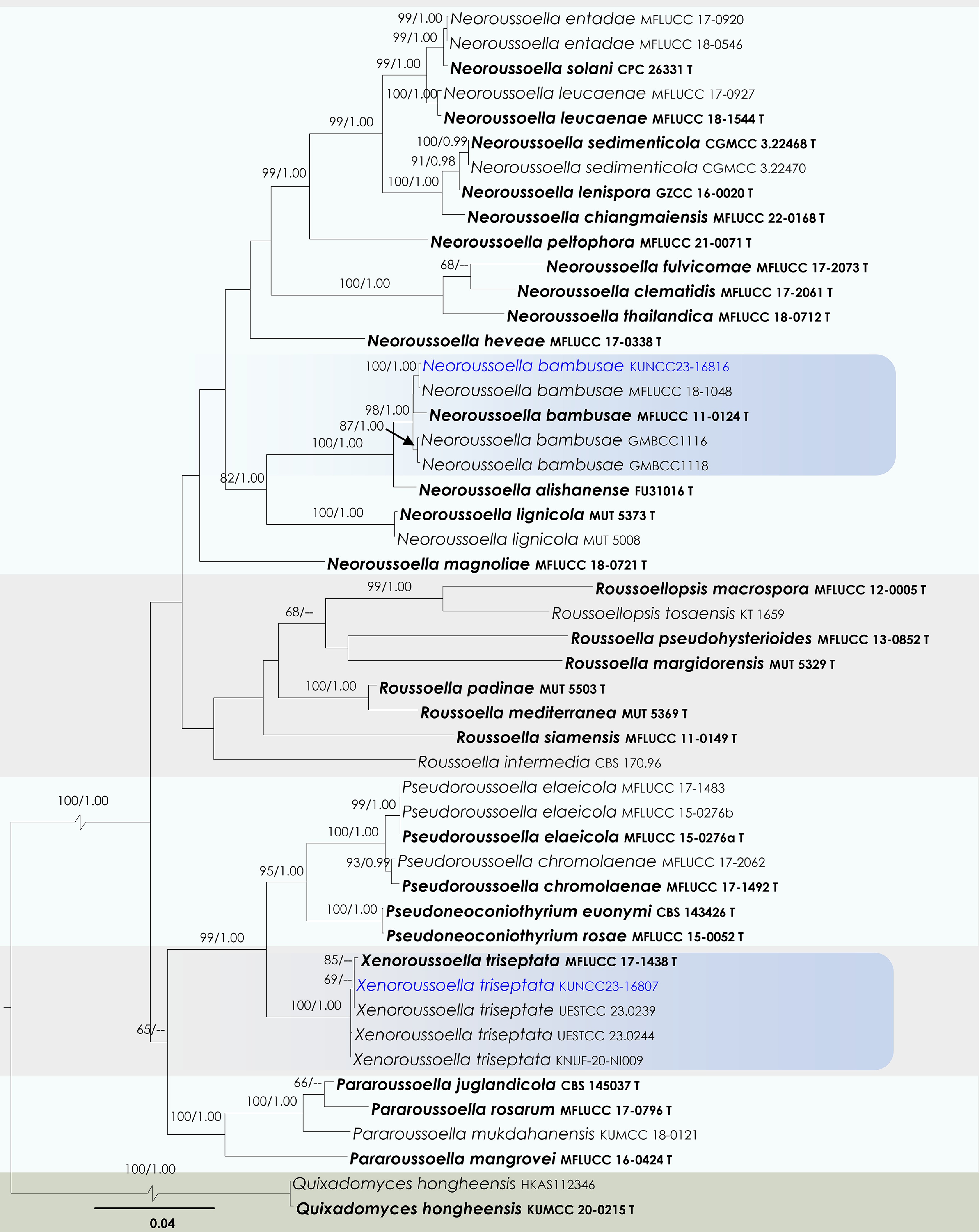
Figure 40.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Quixadomyces hongheensis (HKAS112346, KUMCC 20-0215). The final likelihood value is –24,601.887530. The final alignment included 1,510 unique site patterns, with approximately 21.76% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.246109, C = 0.256427, G = 0.266572, T = 0.230892. The substitution model yielded the following relative rates: AC = 1.941609, AG = 5.780010, AT = 2.469287, CG = 1.378238, CT = 11.805823, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.570250, and the gamma distribution shape parameter (α) was 0.468549. Bayesian inference reached convergence after 1,104,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009906). A total of 11,041 trees were sampled, and 8,281 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,511 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
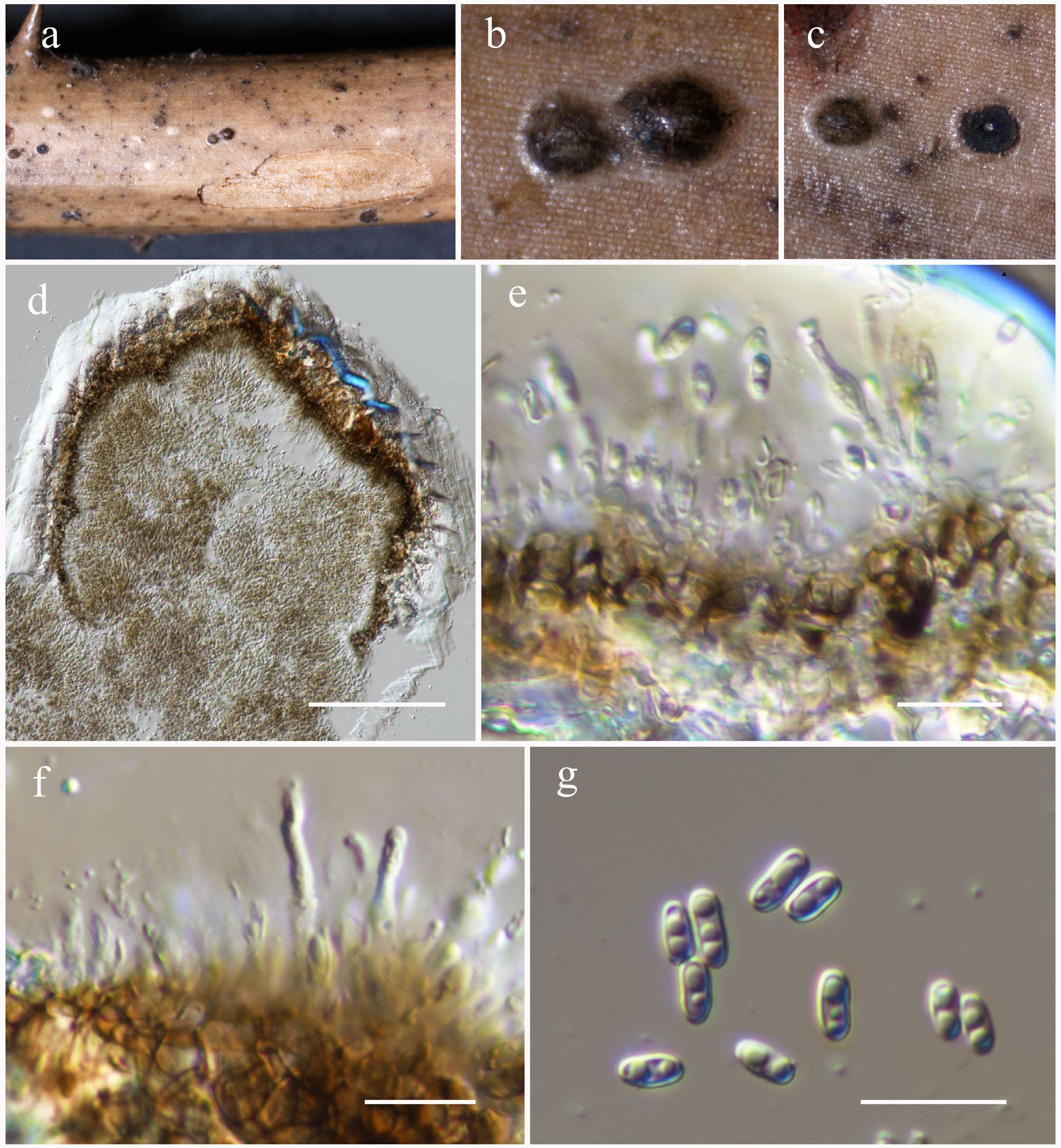
Figure 41.
Neoroussoella bambusae (HKAS146064). (a)–(c) Conidiomata on host. (d) Section of a conidioma. (e), (f) Conidiogenous cells. (g) Conidia. Scale bars: (d) = 100 µm, (e)–(g) = 10 µm.
MycoBank No: 550669
Saprobic on dead branches of Bougainvillea sp. Sexual morph: Undetermined. Asexual morph: Conidiomata 170–220 µm high × 200–300 µm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead branches of Bougainvillea sp., 23 April 2020, D.N. Wanasinghe, DWHH23-50 (HKAS146064), living culture, KUNCC23-16816.
GenBank numbers: KUNCC23-16816: ITS = PV742931, LSU = PV742987, SSU = PV743040, tef1-α = PV700666, rpb2 = PV700694.
Known hosts and distribution: Saprobic on dead bamboo culms in a terrestrial habitat in Thailand[180]; on submerged wood in a stream in Yunnan, China[138]; on dead bamboo culms in Yunnan, China[187]; on dead branches of Bougainvillea sp. in Yunnan, China (this study).
Notes: Neoroussoella bambusae was originally described from dead bamboo culms in a terrestrial habitat in Thailand[180]. It is characterized by semi-immersed to immersed, solitary ascostromata, cylindrical asci, and relatively small, brown, longitudinally ribbed, two-celled ascospores. It is an asexual morph, produced in culture, forms small, hyaline, smooth-walled conidia[180]. Subsequent records include reports from Yunnan, China, on submerged wood in a stream[138] and on dead bamboo culms in a bamboo garden[187]. In our phylogenetic analysis of Roussoellaceae, the new strain (KUNCC23-16816) isolated from Bougainvillea sp. formed a monophyletic clade with previously identified strains of Neoroussoella bambusae (98% MLBS and 1.00 BYPP; Fig. 40). Morphologically, our collection is consistent with the original description of the holotype[180]. Based on both phylogenetic and morphological evidence, Neoroussoella bambusae from Bougainvillea sp. is reported as a new host association for this species.
Xenoroussoella Mapook & K.D. Hyde, Fungal Diversity 101: 93.
Notes: Xenoroussoella was introduced by Mapook et al.[69] as a monotypic genus to accommodate the roussoella-like taxa and is typified by Xenoroussoella triseptata. The genus is initially known only its sexual morph, characterizing by immersed, solitary or scattered, globose to subglobose, brown to dark brown, coriaceous ascomata with protruding ostiolar neck, 8-spored, bitunicate, cylindric-clavate to clavate asci, embeded in oblong to cylindrical, septate, branched pseudoparaphyses, and hyaline to pale brown, or dark brown at maturity, ellipsoid to obovoid, 1–3-septate ascospores, with irregular longitudinal striations[69]. Xenoroussoella is morphologically similar to Roussoella and Pseudoroussoella but phylogenetically distant[69]. Subsequently, de Silva et al.[93], Ryu et al.[188], Tian et al.[189], and Guo et al.[186] reported the coelomycetous asexual morph of Xenoroussoella triseptata that is characterized by pycnidial, solitary to gregarious, immersed to erumpent, uni- to multi-loculate, dark brown to black conidiomata, with an inconspicuous ostiolar, phialidic, ampulliform to cylindrical, hyaline, smooth-walled conidiogenous cells and hyaline to pale brown, oblong to ovoid, aseptate, smooth-walled conidia. Presently, only a single species, X. triseptata, is accommodated in this genus.
Xenoroussoella triseptata Mapook & K.D. Hyde, Fungal Diversity 101: 95 (Figs 40 and 42).
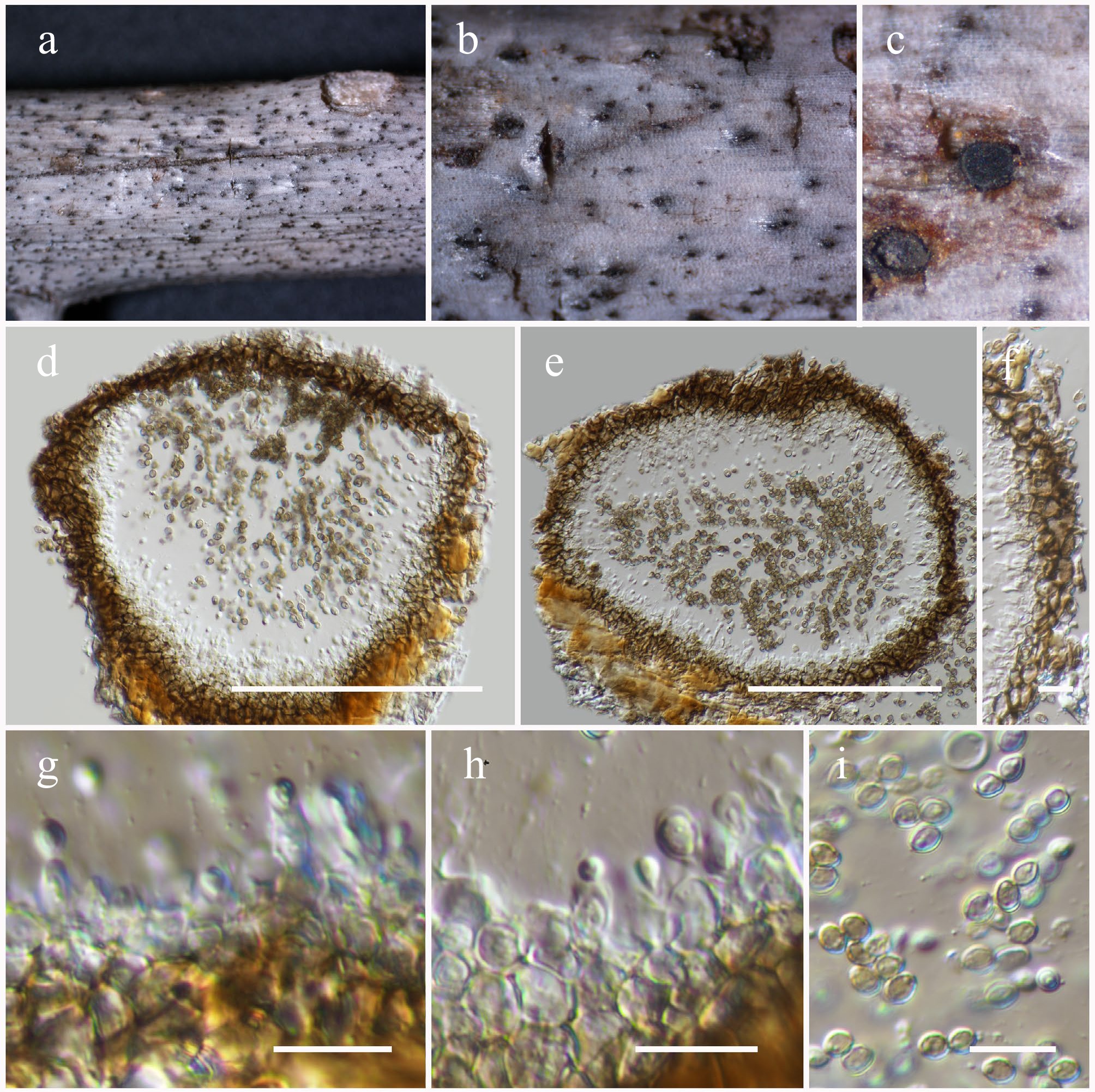
Figure 42.
Xenoroussoella triseptata (HKAS146065). (a)–(c) Conidiomata on host. (d), (e) Section of a conidioma. (f) Pycnidial wall. (g), (h) Conidiogenous cells. (i) Conidia. Scale bars: (d), (e) = 100 µm, (f)–(i) = 10 µm.
MycoBank No: 557368
Saprobic on Bougainvillea sp. Asexual morph: Conidiomata 140–180 µm high × 160–220 µm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead branches of Bougainvillea sp., 23 April 2020, D.N. Wanasinghe, DWHH23-29 (HKAS146065), living culture, KUNCC23-16807.
GenBank numbers: KUNCC23-16807: ITS = PV742932, LSU = PV742988, SSU = PV743041, tef1-α = PV700667, rpb2 = PV700695.
Known hosts and distribution: Saprobic on dead stems of Chromolaena odorata in Thailand[69]; on dead twigs of Anomianthus dulcis and Desmos chinensis in Thailand[93]; on dead stems of Cycas revoluta in China[189]; on dead wood culms in Guizhou Province, China[186]; on dead branches of Bougainvillea sp. in Yunnan, China (this study).
Notes: In our study, phylogenetic analysis based on a combined dataset of SSU, LSU, ITS, tef1-α, and rpb2 gene regions placed one of our new isolates, KUNCC23-16807, within the same clade as the type and previously reported strains of Xenoroussoella triseptata (100% MLBS and 1.00 BYPP; Fig. 40). The morphological characteristics observed in our collection are consistent with those described in earlier studies[93,186,189]. Therefore, this isolate is reported as a new host and geographical record of Xenoroussoella triseptata from Bougainvillea in Yunnan Province, China.
Sulcatisporaceae Kaz. Tanaka & K. Hiray., Studies in Mycology 82: 119.
Anthosulcatispora Phukhams. & K.D. Hyde, Fungal Diversity 102: 117.
Notes: Anthosulcatispora was introduced by Phukhamsakda et al.[123] to accommodate A. brunnea (= Neobambusicola brunnea) and A. subglobosa. The sexual morph was originally described from Neobambusicola brunnea[182]. Within Sulcatisporaceae, Anthosulcatispora is distinct in producing brown ascospores, in contrast to the hyaline, broadly fusiform ascospores with entire sheaths found in Parasulcatispora, Kazuakitanaka and Sulcatispora[24,123,172]. The asexual morph of Anthosulcatispora resembles that of Neobambusicola and Pseudobambusicola, sharing solitary, unilocular pycnidia, phialidic conidiogenesis, and hyaline conidia. However, it differs by having subglobose conidiomata, elongate, cylindrical to truncate conidiogenous cells, and oblong, aseptate conidia[179]. Phylogenetically, Anthosulcatispora forms a distinct lineage within Sulcatisporaceae (Fig. 43).
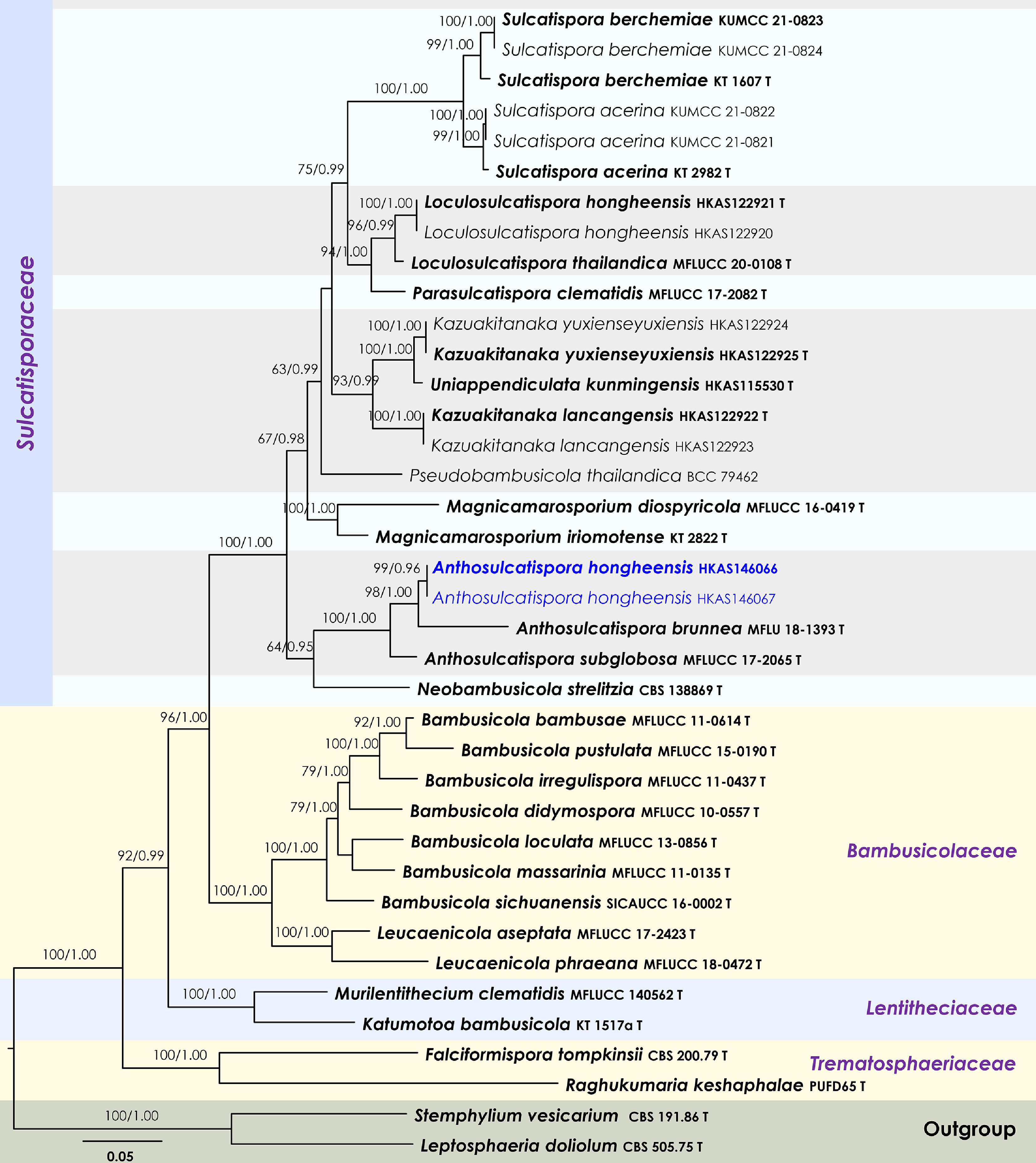
Figure 43.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Leptosphaeria doliolum (CBS 505.75) and Stemphylium vesicarium (CBS 191.86). The final likelihood value is –29,929.948064. The final alignment included 1,836 unique site patterns, with approximately 21.67% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.239264, C = 0.259016, G = 0.269652, T = 0.232068. The substitution model yielded the following relative rates: AC = 1.148753, AG = 2.643315, AT = 1.114998, CG = 0.875753, CT = 6.054602, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.469606, and the gamma distribution shape parameter (α) was 0.537534. Bayesian inference reached convergence after 11,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.007950). A total of 111 trees were sampled, and 84 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,838 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
Anthosulcatispora hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 43 and 44).

Figure 44.
Anthosulcatispora hongheensis (HKAS146066, holotype). (a)–(c) Ascomata on the dead woody twigs. (d) Cross-section of an ascoma. (e), (f) Peridium. (g) Pseudoparaphyses. (h)–(j) Asci. (k)–(n) Ascospores. Scale bars: (d) = 100 µm, (e)–(n) = 10 µm.
MycoBank No: 859366
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146066
Saprobic on dead branches of an unknown host. Sexual morph: Ascomata 220–300 µm high × 180–250 µm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead branches of an unknown host, 23 April 2020, D.N. Wanasinghe, DWHH23-54 (holotype, HKAS146066); ibid., DWHH23-54-2 (HKAS146067).
GenBank numbers: HKAS146066: ITS = PV742933, LSU = PV742989, SSU = PV743042, tef1-α = PV700668; HKAS146067: ITS = PV742934, LSU = PV742990, SSU = PV743043, tef1-α = PV700669.
Notes: In this study, Anthosulcatispora hongheensis is introduced as the third species within the genus Anthosulcatispora. The new species is currently known only from its sexual morph and resembles members of Sulcatisporaceae in having hyaline, broadly fusiform ascospores with an entire sheath[24,123,172,179]. However, it is different from the sexual morph of Anthosulcatispora, which is distinct in producing brown, oblong to ellipsoidal ascospores with rounded ends[182]. Despite these morphological differences, our phylogenetic analysis based on combined SSU, LSU, ITS, tef1-α, and rpb2 sequence data places A. hongheensis in a monophyletic clade within Anthosulcatispora, with 100% MLBS and 1.00 BYPP support values, showing a close relationship to A. brunnea (Fig. 43). Unfortunately, the asexual morph of Anthosulcatispora hongheensis was not observed, and thus comparisons with asexual characteristics across the genus could not be made. Nevertheless, given its phylogenetic placement and morphological affinities within Sulcatisporaceae, the study proposes the inclusion of this species within Anthosulcatispora. Future studies with additional sexual morphs are warranted to further resolve the phylogenetic framework and clarify morphological variation within the genus.
Teichosporaceae M.E. Barr, Mycotaxon 82: 374 (2002)
Magnibotryascoma Thambug. & K.D. Hyde, Fungal Diversity 74: 249 (2015)
Notes: Thambugala et al.[136] introduced Magnibotryascoma to accommodate M. uniseriatum (= Misturatosphaeria uniseriata) within Floricolaceae. The genus is characterized by solitary or aggregated ascomata, fissitunicate, cylindrical to cylindrical-clavate asci, and partially overlapping, brown to dark brown, fusiform to elliptical, usually 1–3-septate ascospores[136,190]. Jaklitsch et al.[191] synonymized Magnibotryascoma with Teichospora, yet morphological differences are notable: Magnibotryascoma species (M. melanommoides, M. rubriostiolatum, and M. uniseriatum) possess fusiform, elliptical, 1–3-septate ascospores, whereas Teichospora (i.e., T. pusilla and T. trabicola) produces ellipsoid to oblong, muriform ascospores[191]. Furthermore, phylogenetic analyses by Hongsanan et al.[110] positioned Magnibotryascoma as a distinct lineage separate from Teichospora within Teichosporaceae, thereby supporting its recognition as a distinct genus.
The asexual morph of Magnibotryascoma rubriostiolata was described by Phukhamsakda et al.[123] from dead stems of Clematis vitalba in the UK, characterized by pycnidial, uniloculate, scattered, semi-erumpent to superficial, subglobose to globose, dark brown to black conidiomata, multi-layered conidiomatal walls, enteroblastic, phialidic, integrated, truncate to cylindrical, hyaline conidiogenous cells, and hyaline (young) to reddish-brown (mature), aseptate conidia. Subsequently, Mortimer et al.[23] introduced another asexual species, M. kunmingense, isolated from dead twigs of Machilus yunnanensis, exhibiting morphological similarities to previously described Magnibotryascoma species. Tennakoon et al.[192] transferred Curreya acaciae[193] as Magnibotryascoma acaciae to Magnibotryascoma based on molecular phylogenetic evidence. The current study introduces an additional species closely resembling other known asexual morphs within Magnibotryascoma[23,123,192,194].
Magnibotryascoma hongheense Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 45 and 46).
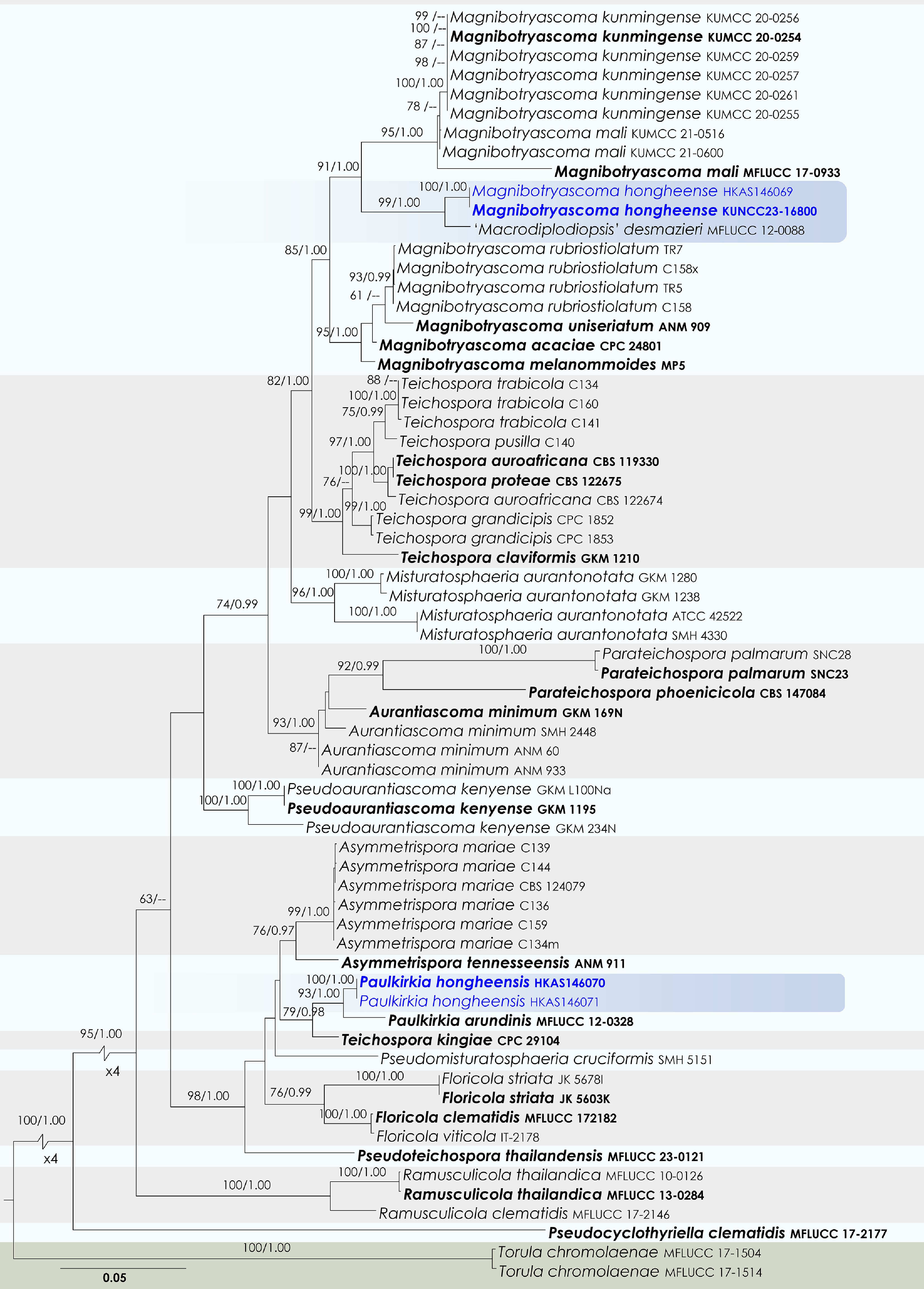
Figure 45.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Torula chromolaenae (MFLUCC 17-1504, MFLUCC 17-1514). The final likelihood value is –21,213.763713. The final alignment included 1,467 unique site patterns, with approximately 43.11% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.243744, C = 0.254910, G = 0.276839, T = 0.224507. The substitution model yielded the following relative rates: AC = 1.338566, AG = 3.975920, AT = 1.726619, CG = 1.303434, CT = 9.924008, GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.496561, and the gamma distribution shape parameter (α) was 0.466889. Bayesian inference reached convergence after 755,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009983). A total of 7,551 trees were sampled, and 5,664 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,468 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.
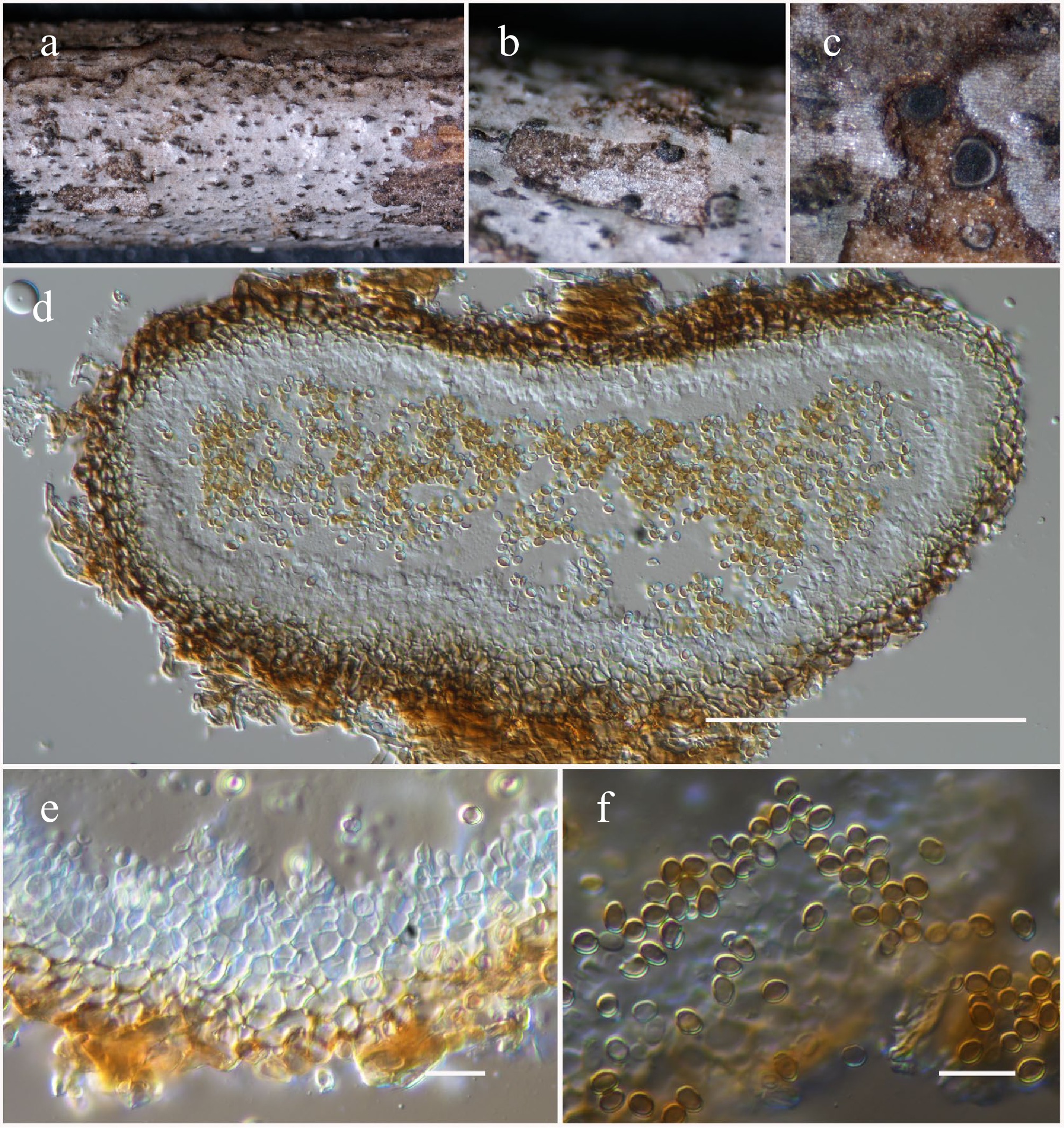
Figure 46.
Magnibotryascoma hongheense (HKAS146068, holotype). (a)–(c) Conidiomata on host. (d) Section of a conidioma. (e) Conidiogenous cells. (f) Conidia. Scale bars: (d) = 100 µm, (e), (f) = 10 µm.
MycoBank No: 859367
Etymology: The specific epithet 'hongheense' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146068
Saprobic on dead twigs of Fagaceae hosts. Sexual morph: Undetermined. Asexual morph: Conidiomata 100–150 μm high, 250–350 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421013° N, 102.227243° E, 517 m, on dead twigs of Fagaceae sp., 14 August 2022, D.N. Wanasinghe, DWHH22-12 (holotype, HKAS146068), ex-type KUNCC23-16800; ibid., DWHH22-12-02 (isotype, HKAS146069).
GenBank numbers: KUNCC23-16800: ITS = PV742935, LSU = PV742991, SSU = PV743044, tef1-α = PV700670, rpb2 = PV700696; HKAS146069: ITS = PV742936, LSU = PV742992, SSU = PV743045, tef1-α = PV700671, rpb2 = PV700697.
Notes: The new species introduced in this study formed a distinct lineage, closely related to Macrodiplodiopsis desmazieri, within the genus Magnibotryascoma (99% MLBS and 1.00 BYPP; Fig. 45). Morphologically, the new species differs notably from Macrodiplodiopsis desmazieri by having unicellular conidia, whereas M. desmazieri produces 3-distoseptate conidia[195]. The asexual morphology of Macrodiplodiopsis desmazieri differs from the generic description of Magnibotryascoma. Macrodiplodiopsis desmazieri is characterized by immersed conidiomata, annellidic conidiogenous cells, and 3-distoseptate conidia[195], whereas Magnibotryascoma exhibits superficial conidiomata, enteroblastic phialidic conidiogenous cells, and unicellular, brown conidia[23,123,192,194]. Therefore, studying the sexual morphology of Macrodiplodiopsis desmazieri is necessary to clarify its taxonomic placement within the genus.
Paulkirkia Wijayaw., Wanas., Tangthir., Camporesi & K.D. Hyde, Fungal Diversity 77: 198 (2016)
Notes: Wijayawardene et al.[196] introduced the genus Paulkirkia to accommodate P. arundinis, isolated as a saprobe on dead stems of Arundo plinii (Poaceae) in Italy. Morphologically, Paulkirkia is characterized by globose to subglobose, unilocular, dark brown to black conidiomata, holoblastic conidiogenous cells, and ellipsoidal or oblong to irregular, 1-septate conidia[196]. This genus shares similarities with Microdiplodia and Verrucoconiothyrium. However, the validity of Microdiplodia remains questionable due to the absence of sequence data from its type species. In recent phylogenetic analyses, Paulkirkia arundinis (MFLU 13-0315) grouped closely with Teichospora kingiae (CPC 29104), both nested within Teichosporaceae (Floricolaceae)[192]. Teichospora kingiae was described by Crous et al.[197] from leaf substrates of Kingia australis in Western Australia. Morphologically, Teichospora kingiae can be distinguished from Paulkirkia arundinis by its verruculose, subcylindrical, 1–3-septate conidia, whereas P. arundinis has ellipsoidal to oblong, 1-septate conidia[196,197]. The sexual-asexual morphological connections between these two species remain unproven. Phylogenetic resolution of these taxa remains ambiguous due to limited sequence availability (LSU and ITS data for T. kingiae and only LSU data for P. arundinis). The present study conducted a robust phylogenetic analysis using combined SSU, LSU, ITS, tef1-α, and rpb2 gene regions across Teichosporaceae. Our results confirm a close phylogenetic relationship between Teichospora kingiae and Paulkirkia arundinis. Additionally, two new isolates obtained in our study clustered with Paulkirkia arundinis with statistical support values of 93% MLBS and 1.00 BYPP (Fig. 45). Morphologically, our isolates fit well within the circumscription of Paulkirkia. However, pending further studies with additional strains and sequence data, it is important to refrain from transferring Teichospora kingiae into Paulkirkia at this time.
Paulkirkia hongheensis Wanas., Phookamsak, L.S. Dissan. & J.C. Xu, sp. nov. (Figs 45 and 47).
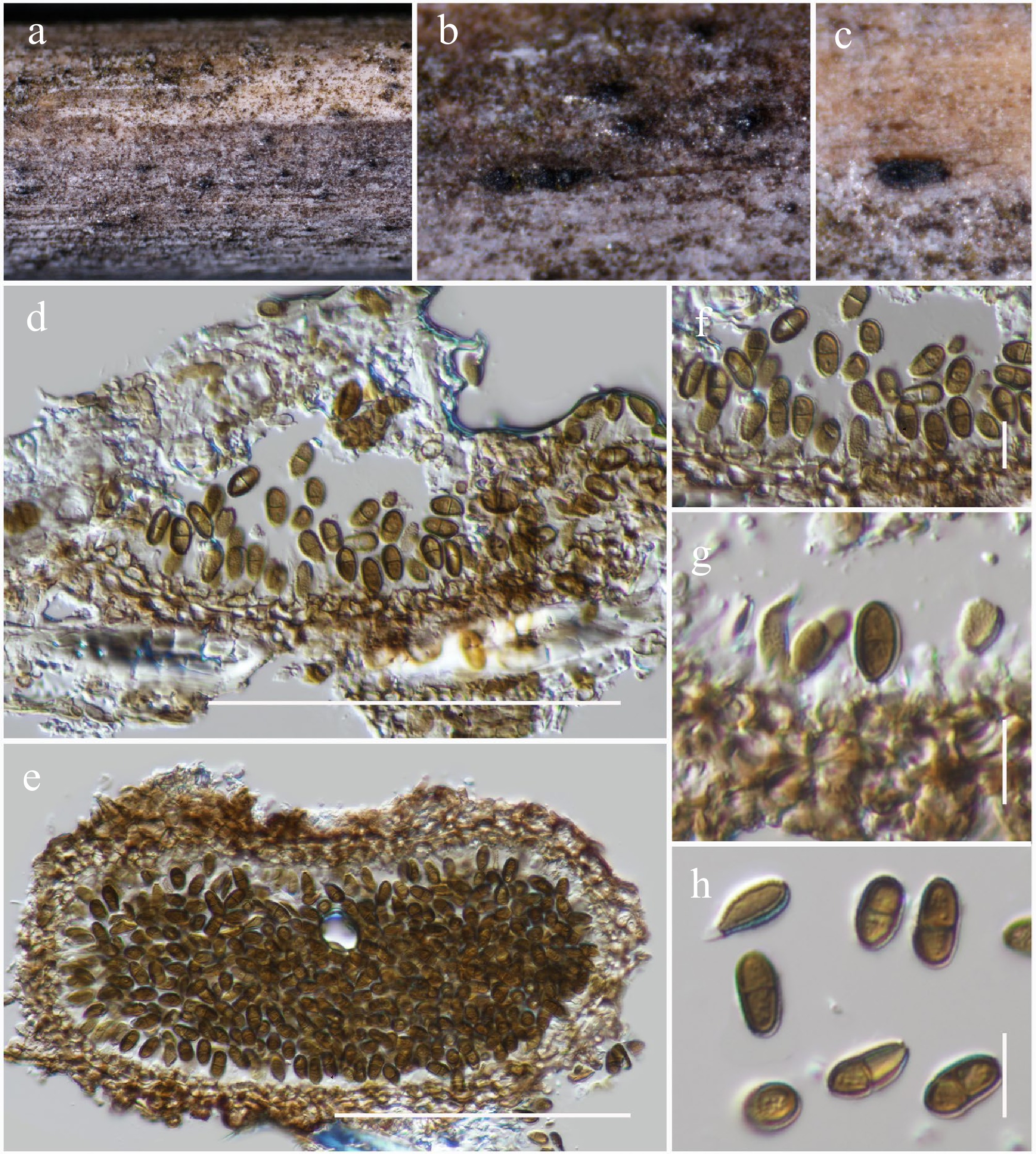
Figure 47.
Paulkirkia hongheensis (HKAS146070, holotype). (a)–(c) Conidiomata on host. (d) Vertical section of a conidioma. (e) Horizontal section of a conidioma. (f), (g) Conidiogenous cells. (h) Conidia. Scale bars: (d), (e) = 100 µm, (f)–(h) = 10 µm.
MycoBank No: 859368
Etymology: The specific epithet 'hongheensis' refers to Honghe, Yunnan Province, where the holotype was collected.
Holotype: HKAS146070
Saprobic on dead stems of a Poaceae host. Sexual morph: Undetermined. Asexual morph: Conidiomata 190–230 μm high, 100–150 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Materials examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead stems of a Poaceae sp., 23 April 2020, D.N. Wanasinghe, DWHH23-56 (holotype, HKAS146070); ibid., DWHH23-56-2 (isotype, HKAS146071).
GenBank numbers: HKAS146070: ITS = PV742937, LSU = PV742993, SSU = PV743046, tef1-α = PV700672, rpb2 = PV700698; HKAS146071: ITS = PV742938, LSU = PV742994, SSU = PV743047, tef1-α = PV700673, rpb2 = PV700699.
Notes: Our newly isolated strains (HKAS146070 and HKAS146071) were collected from dead stems of a Poaceae sp. Morphological examination reveals features consistent with the genus Paulkirkia, including dark, globose to subglobose conidiomata, blastic conidiogenous cells, and ellipsoidal, 1-septate conidia. Phylogenetic analyses using combined sequence data (SSU, LSU, ITS, tef1-α, and rpb2) robustly place our isolates in close relationship with the type species, Paulkirkia arundinis (Fig. 45). Despite morphological variations from the type species, particularly in conidial dimensions (7–10 × 4–6 μm vs 9–12 × 4.5–6 µm), our strains resemble the generic boundaries of Paulkirkia.
Torulaceae Corda, Deutschlands Flora, Abt. III. Die Pilze Deutschlands 2: 71 (1829)
Dendryphion Wallr., Flora Cryptogamica Germaniae 2: 300 (1833)
Notes: Dendryphion was established by Wallroth[198] with D. comosum as the type species to accommodate a group of hyphomycetous fungi. These fungi are typically saprobic, occurring on decaying wood and dead stems of herbaceous plants. The genus is morphologically defined by erect, solitary conidiophores that are branched in the upper region and polytretic. Conidiophores typically possess septate, pigmented, thick-walled stipes with finely roughened surfaces and a well-developed conidiogenous apparatus characterized by prominent scars. The conidia are brown, septate, and produced in simple or branched chains, often displaying didymo- or cheiro-type forms[199]. Species within Dendryphion are reported from both terrestrial and freshwater environments. Currently there are 29 species accepted in the genus Species Fungorum[168], although only eight have been confirmed through DNA-based evidence.
Dendryphion hydei J.F. Li, Phookamsak & Jeewon, PLoS ONE 15 (2): e0228067, 6 (Figs 48 and 49).
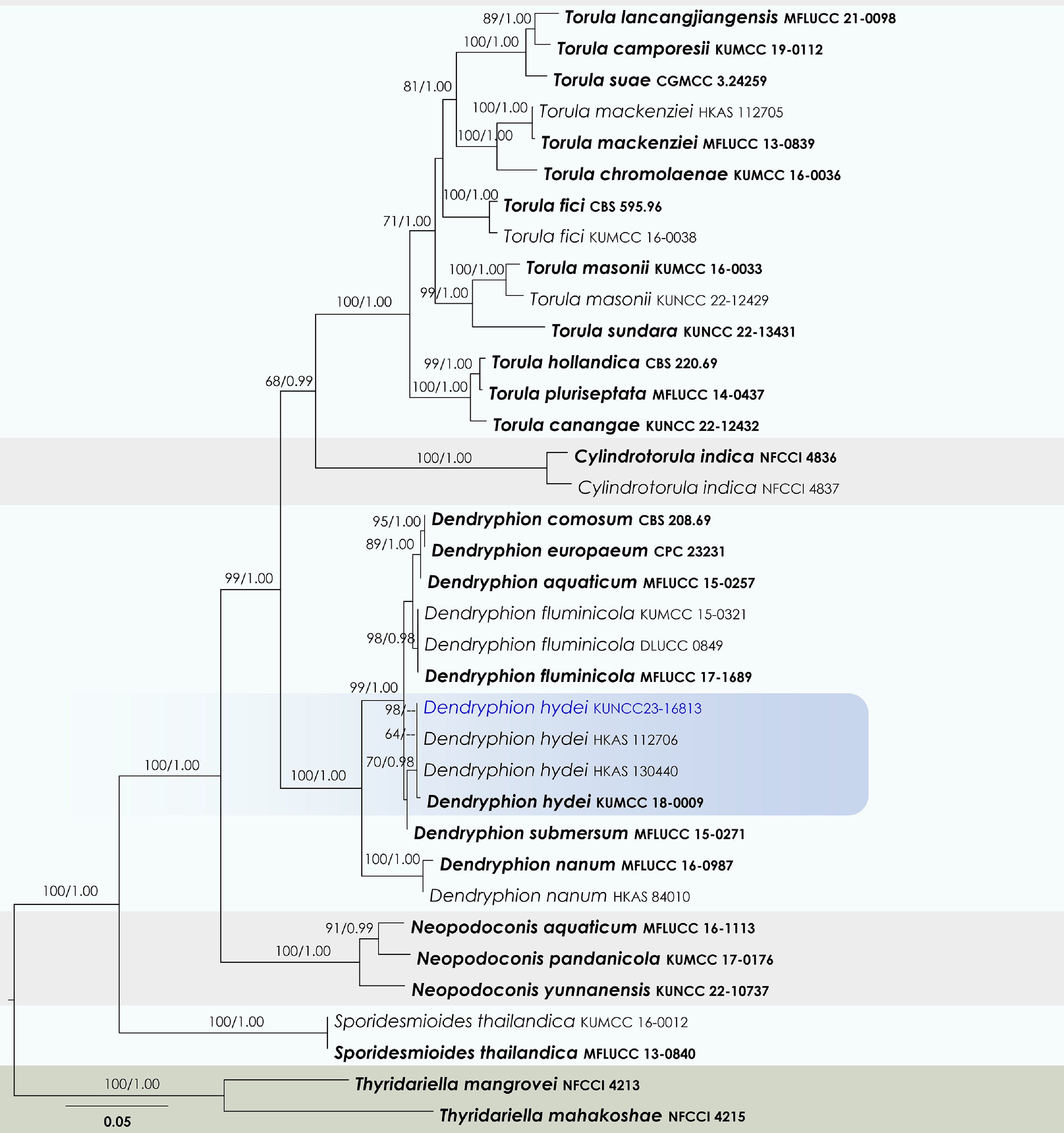
Figure 48.
Maximum Likelihood tree inferred from the concatenated dataset of partial SSU, LSU, ITS, tef1-α, and rpb2 sequences. The phylogeny is rooted with Thyridariella mangrovei (NFCCI 4213), and T. mahakoshae (NFCCI 4215). The final likelihood value is –20,543.293701. The final alignment included 1,363 unique site patterns, with approximately 22.3% of the positions comprising gaps or ambiguous characters. The estimated nucleotide frequencies were as follows: A = 0.245801, C = 0.261172, G = 0.268740, T = 0.224288. The substitution model yielded the following relative rates: AC = 1.969233, AG = 4.496613, AT = 1.690807, CG = 1.174737, CT = 9.891350 and GT = 1.000000. The proportion of invariable sites (I) was estimated at 0.574647, and the gamma distribution shape parameter (α) was 0.522487. Bayesian inference reached convergence after 183,000 generations, when the average standard deviation of split frequencies dropped below 0.01 (observed value: 0.009703). A total of 1,831 trees were sampled, and 1,374 of these were retained for the final analysis after discarding the initial 25% as burn-in. The alignment also revealed 1,364 distinct informative sites. In the resulting phylograms, sequences generated in this study are highlighted in blue, while ex-type or type strains are indicated in boldface.

Figure 49.
Dendryphion hydei (HKAS146072). (a)–(c) Colonies on host. (d), (e) Conidiophores. (f), (g) Apex of conidiophores with conidial structures. (h), (i) Conidia. Scale bars: (d), (e) = 100 µm, (f)–(i) = 10 µm.
MycoBank No: 556746
Saprobic on dead twigs of an unknown host. Asexual morph: Colonies on substratum superficial, effuse, gregarious, hairy, brown to dark brown. Mycelium composed of branched, septate, brown to dark brown hyphae. Conidiophores 260–330 μm long × 6–14 μm diam. (
$\overline{\rm x} $ $\overline{\rm x} $ $\overline{\rm x} $ Material examined: China, Yunnan, Honghe Hani and Yi Autonomous Prefecture, Honghe County, 23.421068° N, 102.229128° E, 735 m, on dead twigs of an unknown host, 23 April 2020, D.N. Wanasinghe, DWHH23-45B (HKAS146072), living culture, KUNCC23-16813.
GenBank numbers: KUNCC23-16813: ITS = PV742939, LSU = PV742995, SSU = PV743048, tef1-α = PV700674, rpb2 = PV700700.
Known hosts and distribution: Saprobic on branch litter of Bidens pilosa in Chiang Mai Province, Thailand[200]; submerged wood in freshwater habitats in Yunnan, China[148,201]; dead twigs of an unknown host in Yunnan, China (this study).
Notes: Dendryphion hydei was originally described from the branch litter of Bidens pilosa in Chiang Mai Province, Thailand[200]. Subsequently, Boonmee et al.[148] and Wang et al.[201] reported this species from submerged wood in freshwater habitats in Yunnan Province, China. In our phylogenetic analysis, the newly obtained isolate (KUNCC23-16813) formed a monophyletic clade with the ex-type strain (HKAS 97479) with 70% MLBS and 0.98 BYPP support values (Fig. 48) and the aforementioned Chinese collections. Morphologically, our specimen fits well with the holotype; however, the other collections exhibit longer conidia and conidiophores in comparison[148,200,201].
-
This study presents a comprehensive investigation of microfungal diversity associated with terrestrial plant debris in a subtropical savanna-like ecosystem of Honghe, Yunnan Province, China. Field collections were conducted across both dry (March, April, December) and rainy (June, August) seasons, capturing a broad temporal representation of fungal diversity. The majority of samples were collected from an approximately 5 ha area at the Centre for Mountain Futures (CMF), which is situated in a subtropical plateau humid monsoon climate zone at low latitude. The sampling environment is marked by steep valleys and highly eroded, nutrient-poor soils. Vegetation is sparse, composed mainly of perennial grasses interspersed with scattered deciduous trees and shrubs, including xerophytic and thorny species adapted to periodic drought and soil instability. More than 100 isolates were obtained during this investigation, of which only those assignable to Dothideomycetes are treated herein. These taxa were isolated from aerially exposed substrates, including senescent foliar material, decorticated twigs, lignified culms, and woody litter. The preferential colonization of such recalcitrant substrates reflects the saprotrophic ecological guild that typifies Dothideomycetes, particularly in seasonally dry, disturbance-prone environments[21,75,110,179]. Members of this class are documented as primary decomposers capable of enzymatic degradation of lignocellulosic plant components[75,110,202] and are characterized by stress-tolerant reproductive traits[203−205]. Taxonomic classification of the isolates, using both morphological features and multigene phylogenetic analyses, revealed a high diversity. The fungi were placed within six orders of Dothideomycetes viz. Botryosphaeriales, Dyfrolomycetales, Hysteriales, Mycosphaerellales, Patellariales, and Pleosporales. Within these orders, taxa distributed across 18 families and 26 genera were identified. Among the collected samples, 34.9% of isolates represented new species described herein. This high novelty rate (> 1/3 of all Dothideomycetes isolates) emphasizes the neglected fungal diversity harbored in this subtropical savanna-like ecosystem. Totally, 34 microfungal taxa, including two new families, two new genera, 15 new species, and 15 new records, were documented.
Phylogenetic resolution of Pleosporales taxa
-
Our phylogenetic analyses highlight several unresolved taxonomic placements within Pleosporales, emphasizing areas that require further taxonomic clarification (Fig. 29). Notably, strains of Lizonia sexangularis (M179 and M222), are grouped closely with members of Didymellaceae. This placement highlights the need for comprehensive future studies involving morphological reassessment and additional molecular data to clarify the familial status of Lizoniaceae. To further resolve taxonomic uncertainties, Pleosporales genera incertae sedis taxa with available DNA sequence data were included in the family-level phylogeny in Pleosporales. Our findings revealed that several genera previously treated as incertae sedis require reassignment to families where they exhibit strong phylogenetic relationships, contingent upon detailed morphological validation. Among our newly generated sequences, only Mangifericomes displayed clear phylogenetic relationships with them. Consequently, establishing a new family, Mangifericomitaceae, is proposed to accommodate Mangifericomes. Other novel sequences that were generated did not show a clear phylogenetic relationship with existing incertae sedis taxa, highlighting the necessity for further in-depth morphological and molecular studies on these taxa.
Our analyses also supported specific taxonomic transfers and placements. Setophaeosphaeria citricola (CBS 143179) clustered within Neopyrenochaetaceae (97% MLBS and 1.00 BYPP; Fig. 29), indicating that Setophaeosphaeria likely belongs in this family. Similarly, Phialophorophoma litoralis (CBS 129112) was strongly affiliated with Cucurbitariaceae (98% MLBS and 1.00 BYPP; Fig. 29), affirming its probable relationship within this family. Additionally, Pyrenochaeta nobilis (CBS 407.76) formed a distinct lineage, emerging as a sister group to Cucurbitariaceae, suggesting a potential novel familial lineage or the expansion of existing family circumscriptions. Likewise, Parastenospora pini (CPC 40385) showed a strong phylogenetic relationship with Pseudopyrenochaetaceae (91% MLBS and 1.00 BYPP; Fig. 29), indicating its likely close affinity to this family. Inflatispora caryotae (MFLUCC 13-0825) and I. pseudostromatica (IFRD 2013) grouped within Pleomonodictydaceae (99% MLBS and 1.00 BYPP; Fig. 29), warranting their transfer from Pleosporales genera incertae sedis to Pleomonodictydaceae. The phylogenetic results of our study support previous suggestions regarding the placement of Inflatispora, initially considered within the Massarineae genera incertae sedis by Tibpromma et al.[206]. However, later Inflatispora was treated as Pleosporales incertae sedis in subsequent studies[59,75,110,207−210].
Further taxonomic uncertainties were observed with taxa like Hobus wogradensis (CBS 141484), which emerged as sister to Ohleriaceae without strong statistical support, indicating the necessity for additional sequence data and thorough morphological studies to resolve its precise familial affinity. Antealophiotrema brunneosporum (CBS 123095) and A. populicola (CBS 147528), Xenolophium applanatum (CBS 123127, CBS 123123), and Acuminatispora palmarum (MFLUCC 18-0460, MFLUCC 18-0264) each formed well-supported independent monophyletic clades with 100% MLBS and 1.00 BYPP statistical values (Fig. 29). These results suggest that each clade potentially represents separate familial lineages, meriting recognition as distinct families pending further morphological and molecular investigation. Ostropella albocincta (SMH 3536) was affiliated as a sister lineage to Paralophiostoma hysterioides (PUFNI 17617) within Paralophiostomataceae, with 87% MLBS and 1.00 BYPP support (Fig. 29). This grouping indicates that the precise familial placement of these taxa requires additional molecular evidence from broader sampling.
Morphological adaptations to dry-hot environments
-
The diverse assemblage of microfungal taxa documented in this study likely reflects a suite of ecological and morphological adaptations to the extreme environmental conditions prevalent in dry-hot valleys. Among the most conspicuous adaptive traits observed was the fully or semi-immersed development of conidiomata and ascomata, with the majority of species forming fruiting bodies beneath the host epidermis. This subepidermal development likely serves as a protective mechanism against desiccation, facilitating successful sporulation under dry conditions. Only three taxa viz. Anteaglonium hongheense, Patellaria microspora and Rhytidhysteron neorufulum were found to produce superficial fruiting bodies (Figs 8, 12 and 14). These species possessed distinctly carbonaceous outer peridial layers, which may function as a mechanical barrier against environmental stressors, particularly heat and desiccation. Such carbonized structures are associated with drought-tolerant fungi and may indicate convergent adaptations to extreme terrestrial habitats. Hyphomycetous taxa were notably rare in our collections. Only Dendryphion hydei and Helminthosporium hongheense exhibited hyphomycetous morphologies. This suggests a limited representation of this asexual reproductive strategy in the sampled habitat. Instead, a balanced representation of sexual and asexual morphs was observed, highlighting the reproductive versatility of the assemblage. Sexual morphs frequently produced ascospores with terminal appendages or encased in hyaline gelatinous sheaths. They may facilitate hydration retention or delayed germination under harsh conditions. Interestingly, such spore ornamentation was absent in species possessing carbonaceous peridia, suggesting potential functional redundancy or divergence in survival strategies. No other consistent spore traits indicative of specific environmental specialization was observed across taxa. Variation in ascospore, morphology including shape, pigmentation, and septation, was evident but not taxonomically or ecologically uniform. These findings suggest that while certain morphological traits (i.e., immersion of fruiting structures, ascospore sheaths) may confer adaptive advantages under drought conditions, many of the observed taxa rely on a combination of generalized and lineage-specific features to persist in these challenging environments.
Conservation and future perspectives
-
Biodiversity conservation emerges as a critical theme within the context of this study. The rich and novel microfungal diversity documented emphasizes the ecological significance and uniqueness of the subtropical savanna-like ecosystems in Honghe. Given the high novelty rate observed, these ecosystems clearly harbor substantial undocumented fungal biodiversity. However, the dry-hot valleys face threats from climate change, habitat degradation, and anthropogenic disturbances[13,211−214], emphasizing the urgency of conservation efforts. Preserving this fungal diversity contributes to maintaining ecosystem integrity, function, and safeguards ecological resilience against environmental stressors. Effective conservation strategies must therefore incorporate comprehensive biodiversity assessments, habitat protection measures, and continuous ecological monitoring to ensure the long-term preservation of these distinctive microbial communities and their associated ecological processes. Future comparative analyses of fungal diversity observed here with similar extreme or arid ecosystems worldwide may yield valuable insights into global patterns of fungal distribution and adaptation. Such comparative analyses could help explain biogeographic patterns and evolutionary processes shaping fungal diversity. While our methodological approach, integrating morphological and molecular analyses, provided robust taxonomic resolution, there are potential biases from selective sampling and culture-dependent methods. Future research could benefit from incorporating complementary methods such as environmental DNA sequencing[215−217]. Detailed ecological studies, investigations into host-fungus interactions, and long-term monitoring of microfungal community dynamics across seasons represent important avenues for further research. Multidisciplinary collaborations integrating taxonomy, ecology, molecular biology, and environmental sciences will be essential for comprehensively understanding and conserving the fungal biodiversity in these ecologically distinctive regions.
Dhanushka N. Wanasinghe is funded by the Distinguished Scientist Fellowship Program (DSFP), King Saud University, Kingdom of Saudi Arabia. Rungtiwa Phookamsak sincerely acknowledges the Talent Introduction Plan of Kunming Institute of Botany, Chinese Academy of Sciences, Yunnan Revitalization Talent Support Program 'Young Talent' Project (Grant No. YNWR-QNBJ-2020-120), Yunnan Revitalization Talent Support Program: High-end Foreign Expert Project (Grant No. XDYC-GDWZ-2024-0016), Independent Research of Department of Economic Plants and Biotechnology, Yunnan Key Laboratory for Wild Plant Resources, Kunming Institute of Botany, Chinese Academy of Sciences (Grant No. Y537731261), Yunnan Provincial Department of Human Resources and Social Security, Yunnan Province Foreign Expert Project (Grant No. 202505AO120002) and Yunnan Intelligence Union Program for Young Scientists, Yunnan, China (Grant No. 202503AM140005). Lakmali Dissanayake is supported by the State Administration of Foreign Expert Affairs (Grant No. Y20240197). Jianchu Xu thanks the Yunnan Department of Sciences and Technology of China (Grant Nos 202302AE090023, 202303AP140001).
-
The authors confirm their contributions to the paper as follows: conceptualization, formal analysis, writing – original draft: Wanasinghe DN; data curation: Wanasinghe DN, Dissanayake LS; methodology: Wanasinghe DN, Phookamsak R, Dissanayake LS; resources, project administration: Wanasinghe DN, Xu J; supervision: Xu J; writing – review and editing: Phookamsak R, Dissanayake LS. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated for this study can be found in the NCBI, GenBank and MycoBank.
-
The authors declare that they have no conflict of interest. Wanasinghe DN, Phookamsak R, and Xu J are the Editorial Board members of Studies in Fungi who were blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of these Editorial Board members and the research groups.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wanasinghe DN, Phookamsak R, Dissanayake LS, Xu J. 2025. Unexplored microfungal diversity from dead woody litter in the dry-hot valleys of Honghe (Yunnan, China). Studies in Fungi 10: e017 doi: 10.48130/sif-0025-0016 

Unexplored microfungal diversity from dead woody litter in the dry-hot valleys of Honghe (Yunnan, China)
- Received: 30 May 2025
- Revised: 02 July 2025
- Accepted: 07 July 2025
- Published online: 27 August 2025
Abstract: Despite the critical ecological role fungi play across various ecosystems, the mycoflora of dry-hot valleys remains largely underexplored. This study aims to fill this gap by documenting the diversity and taxonomy of microfungal species in this unique environment. Employing a combination of field sampling, morphological analysis, and molecular techniques, the study conducted a comprehensive survey of microfungi in selected dry-hot valleys of the Honghe region in Yunnan (China). The findings reveal a rich and previously undocumented diversity of Dothideomycetes (Ascomycota). Among the collected samples, 34.9% of the Dothideomycetes isolates were identified as new species described in this study. The identified microfungi belong to six orders encompassing 18 families and 26 genera. A total of 34 microfungal taxa are documented, including two new families, two new genera, 15 new species, and 15 existing records. During this study, Mangifericomitaceae (Pleosporales) and Mucomycosphaerellaceae (Mycosphaerellales) are introduced as new families. Gruyteromyces and Longiascospora are introduced as new genera in Didymellaceae (Pleosporales) and Pleurotremataceae (Dyfrolomycetales), respectively. The study also introduces new species: Anteaglonium hongheense, Anthosulcatispora hongheensis, Chromolaenicola hongheensis, Gruyteromyces hongheensis, Helminthosporium hongheense, Longiascospora hongheensis, Magnibotryascoma hongheense, Mangifericomes yunnanensis, Mucomycosphaerella hongheensis, Neomultiseptospora hongheensis, Paulkirkia hongheensis, Pseudocapulatispora hongheensis, Pseudochaetosphaeronema hongheense, P. punithalingamii, and Pseudostaurosphaeria hongheensis. Aplosporella artocarpi, A. hesperidica, Chromolaenicola clematidis, Dendryphion hydei, Leptospora phraeana, Murichromolaenicola thailandensis, Neomultiseptospora yunnanensis, Neoroussoella bambusae, Neovaginatispora fuckelii, Paraphaeosphaeria jaguarinae, Patellaria microspora, Pseudocapulatispora longiappendiculata, Rhytidhysteron neorufulum, Tremateia guiyangensis, and Xenoroussoella triseptata, reported as new records. These results highlight the significance of dry-hot valleys as reservoirs of fungal biodiversity and highlight the importance of including these ecosystems in conservation strategies. This study contributes to the expanding knowledge on fungal diversity in extreme environments and calls attention to the need for further research into the ecological roles and potential applications of these neglected mycoflora.
-
Key words:
- Ascomycota /
- Biodiversity conservation /
- Extreme environments /
- Fungal adaptations /
- Fungal diversity /
- Phylogeny /
- Taxonomy