









-
The finger lime [Microcitrus australasica (F. Muell.) Swingle] is a shrub to small tree of the Rutaceae family, bearing edible fruit[1]. It is native to the subtropical rainforest of eastern Australia, mainly concentrated in small areas of northern New South Wales and southeastern Queensland rainforests. A few countries, including France, the United States, and Japan, introduced finger limes into plantations for cultivation in the early- to mid-20th century[2]. Subsequently, the species was introduced into China from the US in 1977. It was sown in the spring of 1978 and cultivated carefully for many years[3]. Later, it was successfully introduced and cultivated in Guangxi, and began to be cultivated in sporadic plantations. During this time, a few cultivation and planting studies were conducted in Yunnan, Guangdong, Hainan, Hubei and other southern regions of China[4]. Since 2011, Hainan Province has been the center of finger lime introduction and cultivation. After 10 years of effort, nine varieties of excellent commercial varieties suitable for domestic cultivation have been selected and cultivated, and more than 300 acres have been industrially planted in Qionghai. This now forms the largest center of finger lime production in Asia.
Finger lime flesh is made up of small round pearls, which impart the fruit's characteristic taste and visual appeal. Finger limes have been adopted and promoted by celebrity chefs around the world for their unique aroma, caviar-like appearance, and refreshing taste. It exists in a variety of colors, including yellow, red, pink, purple, black, blue, and green, among others, making it very versatile and popular for food decoration purposes. Finger limes enjoy a high price in the domestic and international markets, usually fetching between 600 and 700 CNY per kg (US
${\$} $ However, despite the overall success of finger lime cultivation in China, some challenges still stand in the way of the industry's expansion. These include a lack of information on the composition and acidity genes of the fresh fruit, the classification criteria for different varieties of finger lemons are vague, and the need for more knowledge about ideal cultivation techniques for different varieties. Phylogenetic studies can provide evidence of the relationship between different varieties of finger limes. The results of this study can provide a theoretical basis and tool for the study of genetic diversity conservation and phylogenetic relationships among finger limes. Addressing these issues requires a comprehensive analysis of the quality and genetic resources available in current varieties, to ensure the ongoing sustainability of finger lime production in China.
-
As shown in Table 1 and Fig. 1, there was a wide range of variation in tree height, trunk thickness, and branch thickness among the nine finger lime varieties. The highest average tree height was seen for Hongbaoshi (297 cm) and the lowest for Xiaolubi (91.6 cm), with a coefficient of variation (CV) of 33.2% between all varieties. There was less variation in trunk thickness, which varied from 15.8 cm (in Hongbaoshi) to 32.6 cm (in Lvbaoshi). The coefficient of variation for trunk thickness was 24.7%. Branch thickness showed a similar variation, with a CV of 24.4%, and a range from 4.9 mm (Lvbaoshi) to 8.7 mm (Hongbaoshi). While taller varieties tended to have thicker trunks and branches, this correlation was only significant for tree height and trunk thickness (R7 = 0.735, p < 0.05).
Table 1. Comparison of morphological characteristics of finger lime plants of different varieties.
Variety Tree height (cm) Trunk thickness (cm) Branch thickness (mm) Distance between thorns (mm) No. of thorns per branch Spine length (mm) Lvbaoshi 126 15.8 4.9 18.5 11-18 12.1 Xiangbinhong 161.2 24.6 5.1 20.9 12-20 6.9 Wokepu 200 20.2 6.8 14.5 15-20 10.3 Yaoji 186.2 29.6 5.7 14.7 16-22 7.7 Meiweilv 211.8 21.8 6.0 15.3 18-24 11.3 Xuemeng 146.8 21.6 8.2 21.8 9-14 12.0 Xiaolubi 91.6 20.2 6.4 16.8 12-18 6.4 Hongbaoshi 297 32.6 8.7 20.5 12-17 9.4 Lvshuijing 144 20.2 6.2 17.1 15-19 11.1 
Figure 1.
Appearance of the nine finger lime varieties: (a) Hongbaoshi, (b) Lvbaoshi, (c) Lvshuijing, (d) Meiweilv, (e) Wokepu, (f) Xiangbinhong, (g) Xiaolubi, (h) Xuemeng, (i) Yaoji.
Another notable morphological characteristic of C. australasica is the numerous thorns borne along the branches, although varying considerably in the size and distribution of the thorns. All nine finger lime varieties investigated in this study had cone-shaped thorns, although their average length ranged from 6.4 mm in Lvbaoshi to 12.1 mm in Hongbaoshi, with a CV of 24.1%. The highest number thorn density was seen in Meiweilv (18−24 thorns per branch), followed by Yaoji (16−22 thorns). Most other varieties generally bore 12−20 thorns per branch. Notably, the Xuemeng variety only bore 9−14 thorns per branch.
The average distance between thorns ranged from 21.8 mm in Xuemeng to 14.5 mm in Wokepu, with a CV of 17.6% between varieties. There was a negative correlation between the number of thorns per branch and the average distance between thorns, but no correlation with the average spine length of the thorns.
Leaf morphology of the finger lime varieties
-
Most finger lime varieties had obovate shaped leaves, although some tended toward elliptic in shape (Fig. 2). The leaves of three varieties (Wokepu, Yaoji, and Xuemeng) were borne on petioles, while the leaves of the other varieties did not show any significant petioles.

Figure 2.
Comparison of leaf shape characteristics of different finger lime varieties (left to right: Meiweilv, Xiangbinhong, Yaoji, Wokepu, Hongbaoshi, Lvbaoshi, Xuemeng, Lvshuijing, Xiaolubi).
The leaf size also showed considerable differences between varieties, ranging from an average length of 26.2 mm and width of 15.2 mm in Xiangbinhong, to 7.6 mm with and 13.5 mm length in Xiaolubi (Table 2). The typical length for most other varieties was 21−23 mm, with a width of 11−13 mm. Consequently, while the leaf size may be useful in distinguishing between some varieties, it cannot be the main standard used to discriminate between finger lime varieties.
Table 2. Comparison of the leaf morphology of different finger lime varieties.
Variety Leaf width
(mm)Leaf length
(mm)Presence
of petioleLvbaoshi 14.6 22.3 Absent Xiangbinhong 15.2 26.6 Absent Wokepu 12.7 22.0 Present Yaoji 12.8 20.6 Present Meiweilv 12.4 22.9 Absent Xuemeng 11.3 25.9 Present Xiaolubi 7.6 13.5 Absent Hongbaoshi 13.5 23.2 Absent Lvshuijing 10.5 22.5 Absent Comparison of flower morphology of different finger lime varieties
-
There was a small variation in the flower bud size between varieties, as shown in Table 3. Again, Xiangbinhong had the largest sized flower buds (6.3 mm diameter, on average), while Xiaolubi had the smallest (5.3 mm), and most other varieties were approximately 6 mm in diameter.
Table 3. Comparison of morphological characteristics of finger lime flowers of different varieties.
Variety Bud
length
(mm)Petal
length
(mm)Style
length
(mm)Position relationship between anther and stigma (mm) Lvbaoshi 5.5 8.5 8.1 0.13−0.41 Xiangbinhong 6.3 8.3 8.3 0.11−0.26 Wokepu 5.7 8.3 8.1 0.23−0.42 Yaoji 6.1 8.4 8.4 0.12−0.21 Meiweilv 6.1 8.4 8.1 0.23−0.48 Xuemeng 5.9 8.0 7.8 −0.21−0.24 Xiaolubi 5.3 7.7 7.4 0.28−0.42 Hongbaoshi 6.1 8.5 8.2 0.36−0.58 Lvshuijing 6.0 8.3 8.3 0.12−0.23 The position relationship between anther and stigma is the minimum and maximum value, and other values are the average values of 30 samples randomly selected from five fruit trees. The flower bud size was not directly correlated with the petal length, as Hongbaoshi and Lvbaoshi showed the longest petals (8.5 mm) and Xiaolubi the shortest (7.7 mm). Xiaolubi also had the shortest style (7.4 mm average length), while the longest style belonged to Yaoji (8.1 mm). The coefficients of variation for petal and style length were 8.6% and 4.2%, respectively; indicative of minimal varietal differences in flower size.
However, there was a large variation in the relative position of the anthers and stigma, with values ranging from −0.21 to 0.58 mm and a coefficient of variation of 60.25%. Based on this parameter, the varieties could be divided into three groups: those where the stigma is lower than the anther, where the stigma is level with the anther, and where the stigma is higher than the anther. Hongbaoshi fell into the last group, with the stigma almost 0.6 mm above the anther, whereas the stigma of Xuemeng was 0.21 mm lower than the anther. Other varieties such as Yaoji and Xiangbinhong had stigmas and anthers of similar lengths.
The Xuemeng variety had four petals per flower; all other varieties had three petals. Other aspects of the bud and flower morphology were similar between varieties. The flower buds were shaped like a flattened sphere; with a rounded apex (Fig. 3).

Figure 3.
Appearance of the finger lime flower during budding (left) and flowering (right).
Comparison of appearance characteristics of finger lime fruits of different varieties
-
Most varieties bore green-skinned fruit, with Meiweilv the closest to a standard green color (see Table 4 & Fig. 4). The skin of Lvbaoshi, Wokepu, and Xiaolubi tended towards a dark green colour, while Lvshuijing, Hongbaoshi, and Xuemeng were light green. A few other varieties had black to red-black skin.
Table 4. Comparison of morphological characteristics of finger lime fruit from different varieties.
Variety Fruit color Fruit shape Whether the pericarp oil cell is obvious Whether
the tail
protrudesLvbaoshi Dark green Cylindrical Not obvious No Xiangbinhong Black red Elongated bar Not obvious Yes Wokepu Dark green Elongated bar Obvious Yes Yaoji Black Oval Obvious No Meiweilv Green Shallow arched Obvious Yes Xuemeng Emerald green Spherical Obvious No Xiaolubi Dark green Oval Obvious No Hongbaoshi Emerald green Oval Obvious No Lvshuijing Emerald green Elongated bar Obvious Yes 
Figure 4.
Comparison of fruit shape characteristics of different finger lime varieties (left to right: Meiweilv, Xiangbinhong, Yaoji, Wokepu, Hongbaoshi, Lvbaoshi, Xuemeng, Lvshuijing, Xiaolubi).
The shape of the fruit also varied; ranging from cylindrical (Lvbaoshi), oval (Yaoji, Hongbaoshi, and Xiaolubi), elongated (Xiangbinhong, Wokepu, and Lvshuijing), shallow arched (Meiweilv), and almost spherical (Xuemeng).
The shape was also closely linked to the presence of the 'tail'-like protrusion at the end of the fruit. The fruit which had a shallow bow or elongated shape tended to bear a protrusion, while the other fruit did not. The final morphological characteristic of the fruit was the visibility of the oil glands on the fruit skin. The Lvbaoshi and Xiangbinhong varieties did not show any obvious oil glands, while these could be seen clearly in all other varieties.
Comparison of phenological performance of finger limes among different varieties
Comparison of reproduction methods of different finger lime varieties
-
Finger limes can be propagated from cuttings from the branches, or through grafting of branches for cultivation. Cutting methods are more commonly used in northern China, as most limes are cultivated as potted plants. However, due to the abundant sunlight, slightly acidic soil, and good rainfall in Hainan, the species is usually propagated by grafting or grafted seedlings are directly purchased for cultivation. If grafted onto sturdy rootstock, the trees will generally bear fruit within 2−3 years.
At Hainan Shengda Modern Agricultural Development Co., Ltd., finger limes are cultivated using grafting propagation. Fructus aurantii is used as the grafting stock and is propagated from seed between mid-September and November.
Comparison of the phenology of different finger lime varieties
-
Finger limes bloom 3-5 times per year; with each flowering episode lasting for around 7 d. The main blooming event occurs in spring, although the varieties began budding at different stages throughout this period (Fig. 5). In general, darker-skinned finger lime varieties appeared to commence flowering earlier, while lighter-colored varieties only began to bud later. Every year by April 20, most varieties begin to sprout, with the number of buds on Yaoji and Xiaolubi reaching their maxima in the following week (April 27th). The peak budding of light-colored varieties did not occur until later (May 4th to May 18th). Most varieties had a flowering period of about 55 d, with Hongbaoshi and Lvshuijing having the shortest flowering period of only about 40 d, and Yaoji having the longest flowering period of up to 60 d.
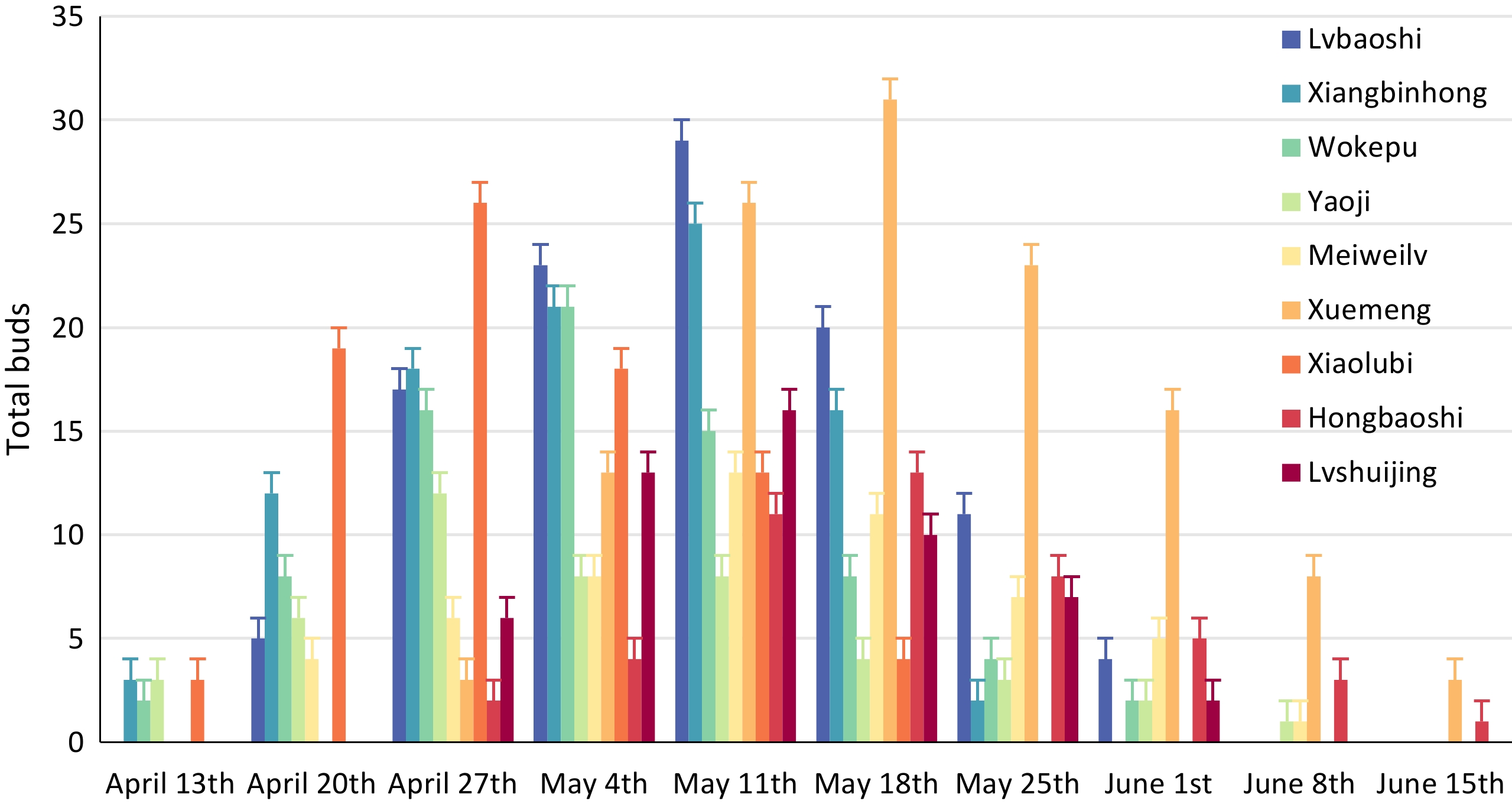
Figure 5.
Comparison of the budding phenology of the finger lime varieties.
In addition to their flowering phenology, the finger lime varieties also varied in their flowering rates. Yaoji had the highest flowering rate of 95.7% (Table 5), while others such as Xiangbinhong, Hongbaoshi, and Lvshuijing all had flowering rates above 90%. At the other end of the scale, Lvbaoshi and Xuemeng had relatively low flowering rates, of approximately 78% and 71.5%, respectively. Nevertheless, these varieties tended to have a higher absolute number of buds and flowers, whereas varieties such as Yaoji had fewer buds and flowers.
Table 5. Flowering of different finger lime varieties.
Variety Total buds Total flowers Flowering rate Lvbaoshi 109 85 78.0% Xiangbinhong 97 89 91.8% Wokepu 76 64 84.2% Yaoji 47 45 95.7% Meiweilv 55 46 83.6% Xuemeng 123 88 71.5% Xiaolubi 83 76 91.6% Hongbaoshi 47 43 91.5% Lvshuijing 54 49 90.7% Comparison of fruiting stages of different finger lime varieties
-
The timing of fruiting among the finger lime varieties closely mirrored the budding and flowering phenology. Xiangbinhong, Wokepu, Yaoji and Hongbaoshi were the first to fruit, starting from April 20th (Fig. 6). The other varieties commenced fruiting at the end of April or early May, in the general order of: Lvbaoshi > Meiweilv > Lvshuijing > Xuemeng > Hongbaoshi. Finger lime varieties with longer flowering periods also had longer fruiting periods. The longest fruiting period belonged to Yaoji, although the trees only produced an average of 33 fruit throughout this period; lower than most other varieties. In contrast, Hongbaoshi had a shorter fruiting period, but produced an average of 69 fruit per plant.
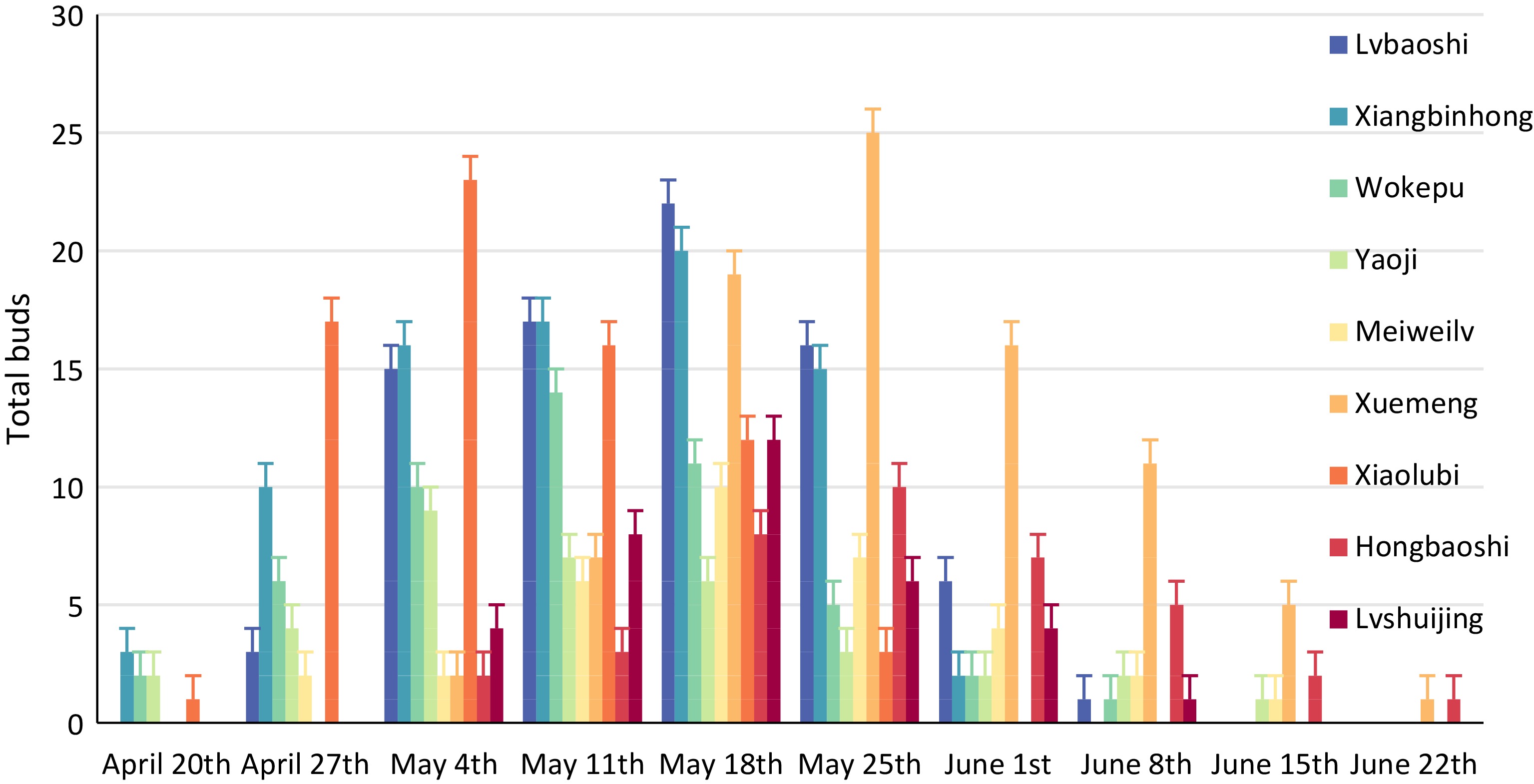
Figure 6.
Phenology of fruiting in the nine finger lime varieties.
The highest rates of fruit set were seen in Xuemeng, Hongbaoshi, Lvbaoshi, and Xiangbinhong, with rates of 97.7%, 94.7%, 94.1%, and 93.3%, respectively (Table 6). Hongbaoshi, Yaoji and Wokepu had set rates of around 80%, while the poorest fruit set was seen in Meiweilv (73.9%) and Lvshuijing (71.4%). However, once the fruit were set, most varieties showed a very high proportion of fruit ripening, from 89.5% in Hongbaoshi to 96.3% in Lvbaoshi. Xuemeng bore the highest average number of fruit per plant (82 limes), followed by Xiangbinhong and Lvbaoshi.
Table 6. Set rates, ripening rates, and total fruit production of the finger lime varieties.
Variety Fruit set rate Fruit ripening
rateNumber of fruit
per plantLvbaoshi 94.1% 96.3% 77 Xiangbinhong 93.3% 95.2% 79 Wokepu 79.7% 90.2% 46 Yaoji 80.0% 91.7% 33 Meiweilv 73.9% 94.1% 32 Xuemeng 97.7% 95.3% 82 Xiaolubi 94.7% 95.8% 69 Hongbaoshi 88.4% 89.5% 34 Lvshuijing 71.4% 91.4% 32 Overall, it appeared that dark-skinned finger lime varieties entered flowering and fruiting stages earlier than light-skinned varieties. Although Yaoji showed very early flowering and fruiting, it produced relatively few fruit per plant, making it less suitable for commercial production. On the other hand, Xiangbinhong performed the best in both flowering and fruiting rates among all the varieties tested, making it a suitable variety to cultivate for fruit production.
Evaluation of different varieties of finger lime fruits
Quality evaluation of different varieties of finger lime fruits
-
The heaviest fruits were produced by Hongbaoshi, which reached an average mass of 29.3 g (Table 7), with a length of 106.8 mm and diameter of 22.6 mm. Most other finger lime varieties had a fruit weight of around 10 g, while the smallest fruit were produced by Xiaolubi (4.7 g per fruit, with a length of 48.6 mm and a diameter of 11.3 mm). Xuemeng, which had been noted for its almost-spherical shape, had the largest diameter (25.7 mm).
Table 7. Physical parameters of the finger lime varieties.
Variety Length (mm) Diameter (mm) Fruit weight (g) Peel weight (g) Flesh color Av. no. of seeds Tightness of peel Lvbaoshi 41.0 17.1 8.8 2.1 Green 9.6 Loose Xiangbinhong 43.6 15.8 8.6 2.3 powder 15.0 Tight Wokepu 61.3 17.8 14.0 6.0 White 13.0 Tight Yaoji 48.7 18.2 10.7 3.3 Red 19.4 Loose Meiweilv 58.4 13.4 6.3 2.8 Green 7.4 Tight Xuemeng 32.2 25.7 13.5 6.1 Yellow 3.0 Loose Xiaolubi 48.6 11.3 4.7 2.4 Red 8.4 Tight Hongbaoshi 106.8 22.6 29.3 16.1 powder 14.6 Loose Lvshuijing 58.7 19.4 10.7 5.0 White 11.6 Loose The flesh color of the finger lime varieties fell into three main groups: green, red, and yellow (Fig 7). Among the red-fleshed finger limes, Yaoji had the darkest flesh, followed by Xiaolubi. Xiangbinhong and Hongbaoshi had lighter red flesh, appearing more pink. Among the green-fleshed finger limes, Lvbaoshi was the darkest, while Wokepu and Lvshuijing were the lightest. In these varieties, the flesh color was more transparent and closer to white/cream. Xuemeng was the only variety with a light yellow flesh.

Figure 7.
Comparison of the pulp color of the finger lime varieties. (a) Lvbaoshi, (b) Xiangbinhong, (c) Wokepu, (d) Yaoji, (e) Meiweilv, (f) Xuemeng, (g) Xiaolubi, (h) Hongbaoshi, (i) Lvshuijing.
The skin of Xiangbinhong, Wokepu, Meiweilv, and Hongbaoshi are looser, making them easier to peel, whereas Lvbaoshi, Yaoji, Xuemeng, Hongbaoshi, and Lvshuijing have a tighter skin and are harder to peel. As shown in Table 7, Hongbaoshi had the highest ratio of skin mass to fruit mass (approximately 55%), and followed by Hongbaoshi, (51%), while the lowest ratio was in Lvbaoshi (about 24%).
Most finger lime varieties contained around 10 seeds per fruit, while Xuemeng had only three seeds per fruit, and Yaoji had an average of 19.4 seeds per fruit.
Flavor evaluation of different varieties of finger lime fruits
-
When juicing the finger lime samples prior before the measurement of acidity and sugar content, it was noted that the ease of juicing differed between varieties. Among the dark-skinned varieties, Lvbaoshi, Xiangbinhong, Wokepu, and Yaoji were all relatively easy to juice, while Hongbaoshi was more difficult to juice. Similarly, Meiweilv and Lvshuijing were the easiest to juice among the light-skinned varieties.
The sugar-acid ratio plays a significant role in determining the sweetness and flavor of citrus fruit. As shown in Table 8, Xiaolubi had the highest sugar content of 28.5%. Meiweilv had the second-highest, at 20.7%, while most other varieties had a content around 15%. Lvbaoshi had the lowest sugar content, at only 10.9%. Most finger lime varieties were strongly acidic, with a juice pH between 2.3 and 2.8. However, Xiaolubi had a much higher pH value of 3.95.
Table 8. Fruit flavor evaluation of the finger lime varieties.
Variety Ease of juicing Sugar Juice pH Valve number Edible portion (%) Lvbaoshi Easy 10.9% 2.33 6 73.1% Xiangbinhong Easy 17.4% 2.29 5 69.6% Wokepu Easy 15.7% 2.24 6 54.7% Yaoji Easy 12.7% 2.49 5 66.6% Meiweilv Easy 20.7% 2.84 6 51.1% Xuemeng Difficult 13.1% 2.54 8 52.1% Xiaolubi Difficult 28.5% 3.95 6 42.3% Hongbaoshi Difficult 11.3% 2.71 4 44.5% Lvshuijing Easy 14.5% 2.35 6 51.7% Most varieties had five or six valves in the fruit, although this ranged as low as four (Hongbaoshi) and as high as eight (Xuemeng). This may affect the perceived texture of the fruit.
Chloroplast genome structure and phylogeny
Basic information of chloroplast genome
-
This study conducted an in-depth analysis of the chloroplast genomes of different finger lime varieties, covering nine major varieties: 'Hongbaoshi', 'Lvbaoshi', 'Lvshuijing', 'Meiweilv', 'Wokepu', 'Xiangbinhong', 'Xiaolubi', 'Xuemeng', and 'Yaoji'. The research results show that although the length of chloroplast genome sequences varies slightly among these varieties, the differences are not significant (Fig. 8). Its length ranges from 160,335 bp [Microcitrus australasica (xiaolubi)] to 160,722 bp [Microcitrus australasica (xuemeng)], with a difference of only 387 bp. These chloroplast genomes exhibit typical tetrad structures in angiosperms, including a large single copy region (LSC, 87,676−87,917 bp), a small single copy region (SSC, 18,754−18,789 bp), and a pair of reverse repeat regions (IR, 26,952−27,008 bp). The maximum differences in the LSC, SSC, and IR regions of the finger lime chloroplast genome are 241, 35, and 56 bp, respectively.
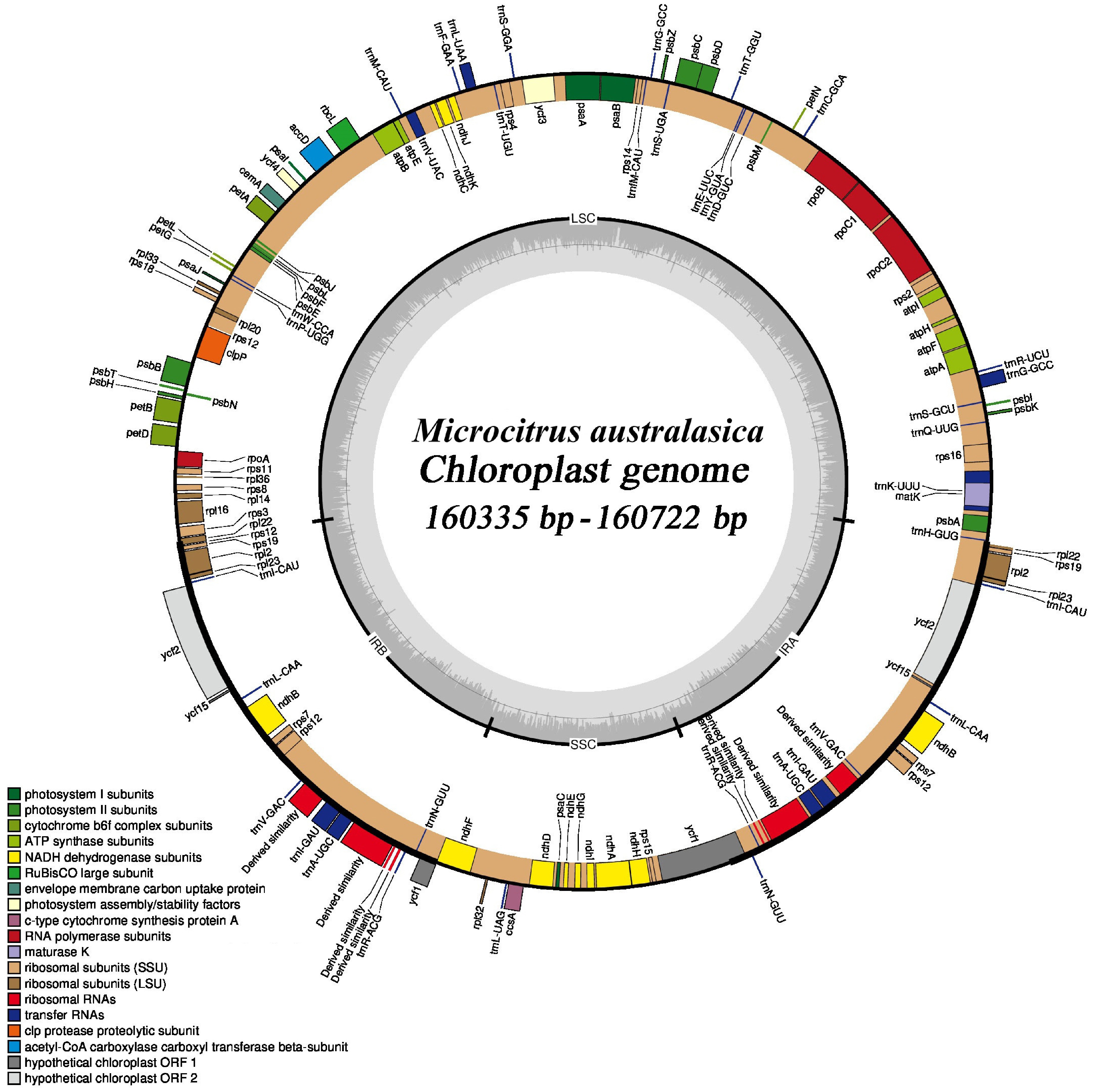
Figure 8.
Chloroplast genome of finger lime. In a circular form, genes inside the outer ring are transcribed clockwise and genes outside the ring are transcribed counterclockwise. The gray area in the inner circle indicates the GC content of the chloroplast genome. Different colors indicate different functional genes.
Further investigation revealed that the chloroplast genome of finger lime encodes a total of 133 or 135 genes, including 115 or 116 unique genes (81 or 82 protein-coding genes, 30 tRNA genes, and 4 rRNA genes) as well as 18−20 genes located in the reverse repeat region. In addition, this study also found that there was no difference in the total GC content between the chloroplast genome sequences of different varieties, both of which were 38.4%. The GC content of large and small single-copy regions and reverse repeat regions were 36.8%, 33.2%, and 43.0%, respectively. These data provide us with a deeper understanding of the structure and function of the finger lime chloroplast genome.
Phylogenetic analysis based on whole chloroplast genome
-
To further reveal the evolutionary relationship of the finger lime family, the chloroplast genome sequences of nine newly determined finger lime varieties were compared with other plants of similar families and genera. Based on the comparison results, maximum likelihood trees and Bayesian trees were constructed by using 55 whole chloroplast genome sequences and three exologous chloroplast genome sequences, respectively (Fig. 9). There is strong support for family relationships and intergenus relationships in the evolutionary tree, but the support for some species branches within citrus is slightly lower. There are some differences among the newly determined finger lime varieties, but the degree of variation is very small, and all varieties are descended from the same branch of evolution, which is closely related to Citrus medica and Citrus indica.
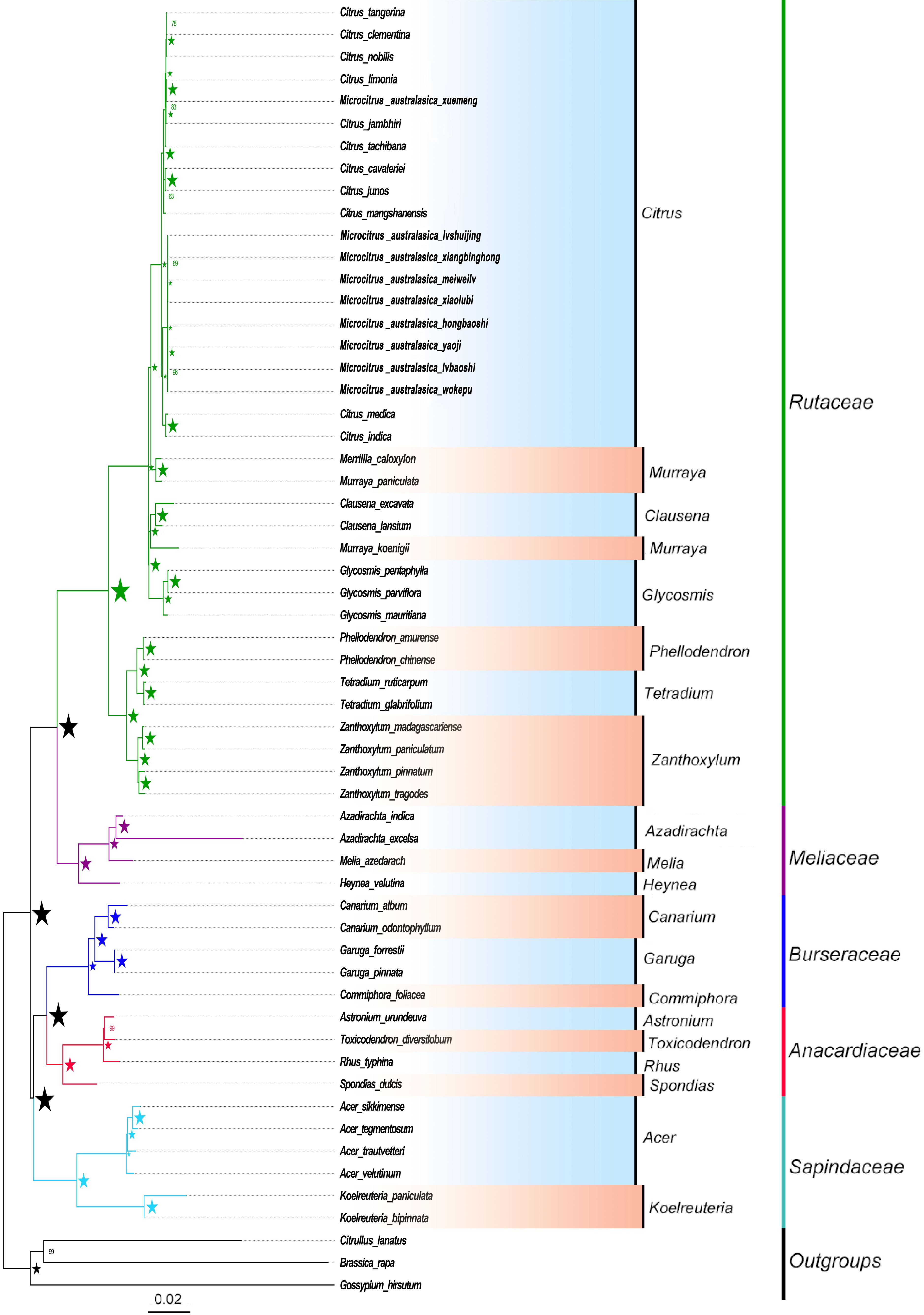
Figure 9.
ML (Maximum Likelihood) and BI (Bayesian Inference) trees constructed based on whole chloroplast genome dataset. ★ Indicates that the support value is 100%/1.0, the number near the node indicates the support value of 60% and 0.6 or more obtained by the analysis, and different colors represent different branches of the family.
SNP marker development based on genome sequencing data
-
There are six types of genomic SNP mutations in nature, including two types of transformation and four types of transmutation. Taking T:A > C:G as an example, this type of SNP mutation includes T > C and A > G. Since sequencing data can be compared to both the positive and negative chains of the reference genome when the T > C type mutation appears on the positive chain of the reference genome, the A > G type mutation is in the same position on the negative chain of the reference genome. Therefore, we divide T > C and A > G into a class[6]. Among these variation sites, the frequency of conversion (A/G, G/A) was 36.1% and 27.8%, and the frequency of transmutation (A/C, A/T, G/C, G/T) was 13.9%, 11.1%, 2.8%, and 8.3%, respectively.
-
The classification of cultivated plant varieties plays an important role in crop breeding, cultivation, reproduction, and application, which studies the origin and development of the species and varieties of these crops. A complete and proper classification system is required to give them accurate and unified names and to better understand the characteristics of these crops[7−9]. The relevant literature on the classification of different cultivated finger lime varieties is relatively scarce, and the research on the biological characteristics of this plant is also relatively general. Too few taxonomic revisions to the geographic distribution of the entire taxa have been made in recent years, meaning that the classification and naming of these taxa are ad hoc, with many specimens still misnamed, unidentified, and/or not identified for decades. The reasons may be that first, most tropical taxa have not been monographed in the last 50 years. In addition, many other published measurements indicate levels of uncertainty in the classification and naming of flowering plants[10]. Consequently, there is little in the way of detailed statistics and classification on the botany, biology, characteristics, and differences of different varieties. Therefore, this study draws on the survey methods and classification criteria of other plant species for a systematic classification of different varieties of finger lime.
Discussion and analysis of the botanical properties of finger lime
-
The nine finger lime varieties could be divided into two broad categories, according to the differences in their peel color: a dark-skinned group and a light-skinned group. Hongbaoshi is particularly prominent among these cultivated varieties, with a significantly greater height and branch thickness, making it the most easily identified. Xuemeng is also relatively easy to identify from its unique spherical fruit shape. However, most of the other varieties are more difficult to identify from their morphological features.
In addition to the first-tier characteristic of peel color, we propose the presence of oil glands and a protruding fruit 'tail' as secondary-tier classification characters. Finally, the leaf shape (including the presence of a petiole) and the tree shape can be considered as third- and fourth-tier characteristics, respectively. Using these characteristics, the different finger lime varieties in this study can be accurately distinguished.
Discussion and analysis of finger lime phenological performance
-
Air temperature, accumulated temperature and precipitation play a vital role in the growth, development and yield of finger lime[11]. The climate in the Hainan region is very suitable for the growth and development of finger lime. Hainan Island has a tropical monsoon marine climate, with an annual average temperature between 22.5−25.6 °C, between 1,780−2,600 annual sunshine hours, total solar radiation between 4,500−5,800 megajoules / square meter, and annual precipitation between 1,500−2,500 mm. These climatic conditions provide sufficient light and water for finger lime.
According to the statistics of Hainan Meteorological Bureau, the average temperature of Hainan Province in April 2023 was 23.1 °C. This meets the temperature demand of finger lime in its early stage of fertility, but without affecting their fertility[12]. At the same time, the average precipitation in April was 90 mm. Consequently, Hainan Island has relatively low precipitation during the flowering and fruit period of finger lime, which help avoid the problem of finger lime root decay during growth.
There were some phenological differences between the finger lime varieties, although the commencement of their flowering and fruiting stages overlapped for most varieties .
Although the varieties were all cultivated under a consistent management regime in this study, previous research has shown that the phenology of flowering and fruiting can be affected by light, temperature, air, humidity, cultivation method, water availability, fertilization rates, pest control and other external factors[13]; all of which directly or indirectly affect the plant's growth. Consequently, the timing of flowering and fruiting cannot be used as a reliable method for the classification of finger lime varieties.
Discussion and analysis of the fruit quality of finger lime
-
Agricultural scientists around the world continue to work on breeding new finger lime varieties[14−17]. The weight, acidity and sweetness of the fruit are regarded as the basic indicators for breeding quality, as these reflect the taste and nutritional value of finger limes.
The nine finger lime varieties investigated in this study showed significant differences in their single fruit weight. The average fruit weight across all varieties was 11.8 grams, although Hongbaoshi had an average weight of 29.3 grams, more than 6 times that of the smallest fruit (Xiaolubi).
Similarly, the average sugar content of the nine varieties was 17.2%, but this ranged considerably from 10.9% in Lvbaoshi to 38.5% in Xiaolubi. Most varieties were quite acidic, with a pH between 2.3 and 2.8, although Xiaolubi again had a much higher pH (3.95). Coupled with the high sugar content, this suggests that fruit from this variety are likely to have quite a sweet flavor.
In addition, Adhikari et al. observed the new finger lime variety P1f 2-10, and found that the average single fruit weight reached 12.8 g and the maximum single fruit weight was more than 19 g. Its sugar content ranged from 18.0% to 21.0%[18]. These data are similar to the fruit weight, sugar content and acidity content of finger limes observed in this experiment.
Combined with the results of other varieties in previous studies, the nine finger lime varieties investigated in this experiment can enrich the diversity of finger lime varieties in China; providing a significant germplasm resource in terms of fruit weight, sugar content and acidity. Additionally, it provides much-needed reference information for breeders to develop new high-quality finger lime varieties, helping improve the competitiveness of China's finger lime industry and meet consumers' demand for high-quality fruits.
Correlation of finger lime traits
-
Detailed observations on fruit growth revealed that finger limes experience two important growth peaks from flowering to fruit ripening, with the first occurring 8–12 d after pollination, and the second peak occurring from 25 d post-pollination through to fruit harvest. This bimodal growth curve observed in this species[19−21] is similar to the trends in fruit growth seen in litchi, apricot, kiwi, cherry and pitaya[22−26]. During both periods, cell expansion accelerates and the fruit weight increases rapidly, with an ultimate impact on the overall fruit yield. These growth periods are also key formation periods for the fruit quality, impacting the taste, nutritional value, and appearance of the fruit.
To explore the relationship between the size of the finger lime trees and fruit quality, correlation analysis was conducted between 10 quantitative traits. As shown in Fig. 10, there was a highly significant negative correlation between tree height and the number of fruits per plant, while there was a highly significant positive correlation between tree height and branch thickness, fruit longitudinal diameter, soluble solid content, and single fruit weight. This shows that fruit size can be predicted from the tree size and branch thickness. Taller trees tend to produce thicker branches, and larger fruit (in both diameter and weight) with a higher soluble solid content. This observation has important practical implications for informing the cultivation of finger lime. The trees should be properly supplemented with adequate nutrients to allow robust growth, which in turn should promote the subsequent growth of finger lime fruit and improve the fruit yield and quality.
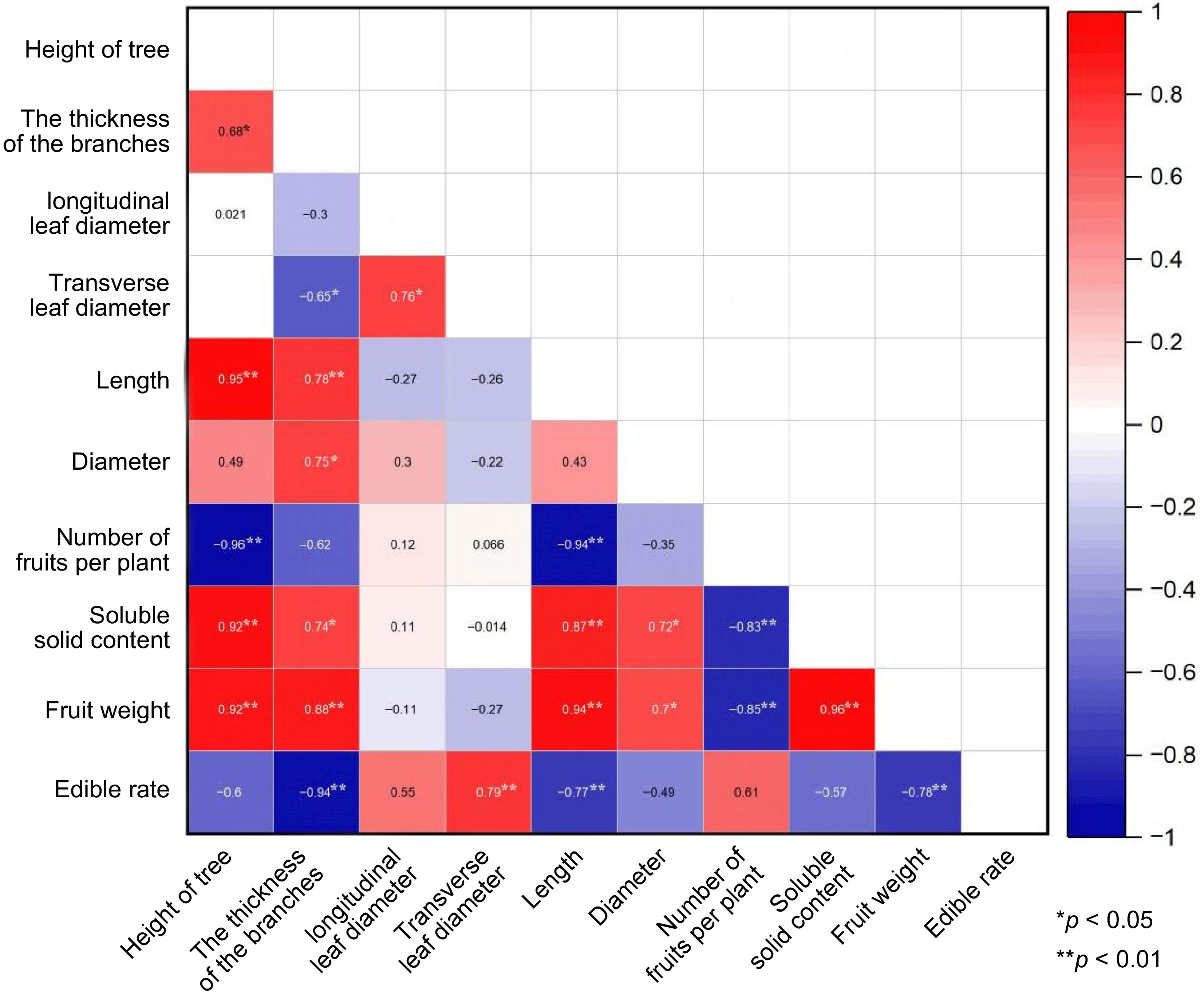
Figure 10.
Correlation analysis of some observed traits of finger limes. * p < 0.05 is considered significant. ** p < 0.01 is considered highly significant.
The density of single fruits also significantly impacts on the quality of the fruit. An excessive fruit load will not only reduce fruit yield and quality, but also affect the accumulation of tree nutrition[27, 28]. The present study supported this, finding a significant negative correlation between the number of fruit per tree, and the soluble solids and single fruit weight. In other words, as the fruit density increases, the single fruit weight and the soluble solids content will decrease accordingly. Similarly, Li Jinxue observed that thinning flowers and fruits had a positive effect on improving the quality of lemon fruit grown in Dehong prefecture, Yunnan province. Thinning significantly increased the longitudinal diameter, TSS, and TA content of lemon fruit across two seasons[29]. This confirms that appropriate measures of fruit thinning and controlling the quantity of fruit produced are important to improve the yield, quality, and nutrient accumulation of finger lime fruit.
Analysis of the structure of finger lime chloroplast genomes
-
According to the comparative analysis of the chloroplast genome structure (Fig. 9), the finger lime chloroplast genome resembles most other citrus plants[30−35].
This study used whole-chloroplast genome data for ML and BI analysis to parse the phylogenetic analysis of finger limes, showing strong support for every node in the phylogenetic tree, with only slightly lower support for some internal species branches of Citrus. In the interior of the genus, citron (Citrus medica) and Indian orange (Citrus indica) are recently related to the finger limes. Citron fruit are large (10−25 cm diameter), with a yellow/green outer pericarp, thick yellow/white mesocarp (1.5−3.5 cm), scattered oil glands, and 1−2 seeds[36]. These biological properties are somewhat similar to the biological traits of finger lime observed in the study, which may support their relationship.
Although there was minimal variation between the newly determined finger lime varieties; the species had a higher degree of variation and appeared to be closely related to lemon (Citrus limonia). The Xuemeng variety was created by crossing the original finger lime species (C. australasica) and a local Hainan lemon variety, explaining the close relationship between these varieties. The wild form of C. limonia is reportedly a small tree, bearing strong thorns, and oblong leaves with a short petiole. The fruit is small (4 cm diameter), yellow, and semi-spherical, with 8−10 segments and 3−16 seeds per fruit[37]. The main traits of the Xuemeng variety observed in this study share the same characteristics, as it grows in the form of a shrub, with oblong, petioled leaves, long buds, and short stigmas. The fruit is small (5 cm diameter), with eight segments and a small 'tail' which disappears during maturation. These common biological characteristics support the aforementioned phylogenetic relationship between Xuemeng and lemon.
Molecular markers play important roles in breeding; their main applications include molecular mapping and gene mapping, fingerprint library establishment, and molecular marker-assisted selection for desirable traits[38]. Through sequence-level analysis of the germplasm, the genetic properties of the germplasm and their relationships can be assessed more accurately, thus laying the foundation for genetic improvement. Single nucleotide polymorphism (SNP) provides a rich diversity of information while maintaining stability, making it one of the most prominent molecular markers. It is of great significance for documenting the genetic diversity of fruit tree germplasm resources, construction of high-density genetic maps, and allowing mapping of quantitative trait loci (QTL)[39−41].
Single nucleotide polymorphism (SNP), a representative of the third-generation molecular marker technology proposed by Lander, refers to the sequence polymorphism[42,43] formed by a single nucleotide variant of DNA at a specific nucleotide position within the genome. SNP includes four forms, namely transition, transversion, insertion and deletion[44−47]. There are two main strategies currently used for SNP development. The first is direct sequencing of DNA amplified fragments; although it is expensive, it provides a very low false positive rate and allows identification of the haplotype composed of SNP[48]. However, this method is expensive and expensive, so it is suitable for the development and validation of a specific SNP. The second method is the automated identification of polymorphic sites through bioinformatics approaches, which is a simpler and more cost-effective way of obtaining SNP candidates[49].
SNP markers based on transcriptome sequencing have been successfully developed and applied to various foodplants (apple, peach, citrus, grape, apricot, cherry, banana and almond)[50−57]. In this study, we obtained candidate SNP for various finger lime fingerprint features through bioinformatics methods and found that the SNPs were widely distributed throughout the genome, with SNP marker conversion occurring at a frequency of 63.88% and 36.11%. This is consistent with the results reported in apple by Liu[6]. However, across all the mutations reported by Wang in radish, the frequency of A/G was close to C/T and much greater than that of other types of mutations, and the frequency of the other four types of mutations (A/T, A/C, G/T, and C/G) were less different[58]. This may be related to species differences, gene structure properties, sequencing data size, SNP retrieval tools, and search criteria. The SNP marker sequences identified here are widely involved in various metabolic processes of finger limes and affect the formation of various agronomic traits of finger limes. However, the specific functional localization and influence mode of these SNP punctuation points have not been fully clarified, and further studies on functional gene localization and breed fingerprinting are needed in the future.
-
There were nine experimental samples used in the sequencing study. The nine experimental samples were collected from Shengda Modern Agricultural Development Co., Ltd. (19.349320, 110.450803) in Qionghai, Hainan, China; comprising 3−4 leaves from nine different varieties of finger lime fruit trees. All samples were from healthy and strong-growing plants. After rinsing with water, they were dried and stored in a plastic bag, prior to subsequent genome extraction.
Experimental statistical method
-
At the fruit ripening stage, 30 fruit trees were randomly selected to investigate the botanical characteristics of finger lemon varieties. The specific contents are as follows:
(1) Plants: Observe and record the tree height, trunk thickness, branch thickness, and distance between thorns on branches of different varieties of finger lemons, and count the number of thorns on the branches;
(2) Leaves: 30 leaves were randomly selected on randomly selected fruit trees, and the shape characteristics of the leaves were observed and recorded;
(3) Flowers: Randomly select 30 flowers on randomly selected fruit trees, observe and record the appearance characteristics of flower buds;
(4) Fruit: 30 fruits were randomly selected on randomly selected fruit trees, and the appearance characteristics of the fruits were observed and recorded;
(5) The propagation mode and growth time of different varieties of finger lemons were investigated and counted.
Plant material DNA extraction
-
The modified CTAB (Cetyltrimethyl ammonium bromide) method, commonly used for DNA extraction from plant samples[59], was used to extract the DNA in this study.
Chloroplast genomic DNA sequencing and sequence assembly
-
The plant chloroplast genomic data was obtained using shallow genome sequencing technology. Before performing whole genome sequencing (WGS), the Agilent 2100 Bioanalyzer of the UCDAVIS Genome Center (Davis, California, USA) was used to perform quality testing on each plant's DNA sample. Samples with a total DNA content of more than 0.8 μg were selected for subsequent library construction and sequencing. Using the BGISEQ-500 platform of Shenzhen BGI, a paired end-sequencing library with an insertion length between 300−500 bp was constructed and de novo sequenced. The high-quality original data source (raw reads) of each sample can reach about 8 GB. We used SOAPfilter v2.2 developed by BGI, Shenzhen, China to filter and prune the original data source generated by sequencing, including screening for low-quality base readings (> 10% Ns and/or > 40% low-quality bases), removing erroneous readings generated by PCR replication, and adjusting the adapter sequence.
The chloroplast genome sequence was then assembled from the filtered clean data using GetOrganelle v1.7.5.0[60], with the software's default parameters (see online guide
https://github.com/Kinggerm/GetOrganelle ). All chloroplast genome assembly results were visualized using Bandage v0.8.1[61] software to check whether the new assembly map formed a tetrad structure, in order to evaluate the quality of the assembly and the integrity of the final assembly map.Using the sequencing data, the SNPs (single nucleotide polymorphisms) of finger lime genome sequence were retrieved. The SNPs found were screened according to the following criteria to ensure their accuracy: (1) the distance between two SNPs is not less than 2 kb; (2) The number of base-sequencing reads at the mutation site was not less than 10; (3) The sequencing quality values of the two bases at the mutation site were not less than 2,062. Through these screening, we obtained SNPs with high confidence. A relatively large number of SNP sites have been detected in the varieties of "Xuemeng", while only a small number have been detected in other varieties.
Chloroplast genome annotation
-
Before sequence annotation, a global alignment was performed using Mauve v1.1.3 to check the collinearity of the genomic sequence[62]. The annotation of the chloroplast genome sequence was carried out by means of Geneious Prime v2021.2.2 and the already annotated sequence of its close relatives as the reference sequence[63]. After the preliminary annotation was completed, the annotation results were evaluated and manual modifications were made, including the adjustment of the start/stop codon and intron/exon boundaries. To ensure the accuracy of the annotated genes, we performed error detection on the annotated information of the genomic sequence using the software Sequin v.15.10.[64] The tRNAscan-SE software was used to verify the presence of the tRNA gene. In addition, the individual partition boundaries of the annotated sequence were determined by using Unipro UGENE v38.1[65]. We performed the mapping of the chloroplast genome using the online software OrganellarGenomeDRAW (OGDRAW) v1.3.1 (
https://chlorobox.mpimp-golm.mpg.de/OGDraw.html )[66].Construction of phylogenetic relationships
-
The nine newly sequenced chloroplast genome sequences were used to construct a phylogenetic analysis, along with 50 chloroplast genomes downloaded from NCBI and three outgroup chloroplast genome sequences(Supplemental Table S1). When constructing the phylogenetic analysis, this study used the MAFFT plug-in in Geneious Prime v2021.2.2 software to align all sequences, manually check the alignment results, and remove areas with more than 80% missing data. The phylogenetic tree was reconstructed using Maximum likelihood (ML) and Bayesian inference (BI). For ML and BI methods, (Akaike Information Criterion, AIC) and (Bayesian Information Criterion, BIC) respectively used in (Smart Model Selection, SMS) to determine the model that best fits this matrix[61]. The phylogenetic tree was constructed using the ML method, GTR + G was selected as the optimal model, and the Rapid bootstrap algorithm was used to calculate the bootstrap support rate of each branch 1,000 times. Each analysis is performed twice independently, setting a random starting tree for a total of 400,000,000 iterations, and recording the sample parameters and the system tree every 1,000 generations. MrBayes v 3.2[62] was used for BI analysis, GTR + I + G was selected as the optimal model, and the analysis started from a random evolutionary tree, and then 2 million generation operations were carried out on four parallel evolutionary chains. Markov chains were sampled every 100 generations, resulting in a total of 20,000 phylogenetic trees. The first 25% (the initial 5,000 Burn-in trees) were removed from the analysis, and the remaining 15,000 phylogenetic trees were analyzed for consistency (using the 50% majority rule). A 50% majority rule consensus tree is obtained. FigTree v1.4.4 was used to analyze all ML and BI obtained systems. This allows for visual analysis of tree cultivation (
http://tree.bio.ed.ac.uk/software/figtree ). This study was funded by Hainan Province Science and Technology Special Fund (ZDYF2022XDNY190), University level scientific research project of Hainan University (XTCX2022NYB09), the Project of Sanya Yazhou Bay Science and Technology City (SCKJ-JYRC-2022-83) and Hainan Provincial Natural Science Foundation of China (421RC486).
-
The authors confirm contribution to the paper as follows: study conception and design: Lv W, Wang HF; data collection: Zhu MH, Wang Q, Huang YS; analysis and interpretation of results: Lv W, Johnson JB, Zaman QU; draft manuscript preparation: Lv W, Zhang Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0024-0015)
-
Received 22 January 2024; Accepted 7 April 2024; Published online 22 May 2024
-
Finger lime varieties are divided into two groups based on skin color: dark and light varieties.
Under consistent cultivation and management conditions, Xiangbinhong appears to be the most suitable variety for commercial production.The Xuemeng variety is very suitable for use as ornamental shrubs or grafting rootstocks.
The chloroplast genome of lime presents a typical tetrad structure of angiosperms, consisting of a single copy region of the same size and a pair of separated reverse repeat regions.
-
# Authors contributed equally: Wei Lv, You Zhang
- Supplemental Table S1 Overview of the main features of chloroplast genomes of finger lemons and their related varieties.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lv W, Zhang Y, Johnson JB, Zaman QU, Zhu MH, et al. 2024. An investigation of germplasm resources and phylogenetic analysis of finger lime in Hainan Province, China. Tropical Plants 3: e016 doi: 10.48130/tp-0024-0015 

An investigation of germplasm resources and phylogenetic analysis of finger lime in Hainan Province, China
- Received: 22 January 2024
- Revised: 31 March 2024
- Accepted: 07 April 2024
- Published online: 22 May 2024
Abstract: The finger lime [Microcitrus australasica (F. Muell.) Swingle] is a small tree with high economic, culinary, and ornamental value, originating in the rainforests of eastern Australia. This study collected and comprehensively evaluated nine finger lime varieties, characterizing 42 traits across all plant parts (tree, thorns, flowers, fruit). The tested traits include botanical characteristics, phenological period, fruit comparison, and evaluation, to better understand the characteristics and differences between varieties, and provide reference data for the development of production technologies. The main results are summarized in the following points: (1) The nine finger lime varieties were divided into two groups according to the color of the fruit peel: the dark varieties (Lvbaoshi, Wokepu, Xiaolubi, Xiangbinhong, and Yaoji) and the light varieties (Lvshuijing, Meiweilv, Hongbaoshi, and Xuemeng). (2) Under consistent cultivation and management conditions, Xiangbinhong appears to be the most suitable variety for commercial production. This variety displays excellent qualities for fresh fruit production, including its moderate fruit size, thin peel, high sweetness, and delicate flavor. Additionally, the Xuemeng variety produced round, green fruit – albeit of an average flavor – making it quite suitable for use as an ornamental shrub or grafting stock. (3) The chloroplast genome of the finger lime showed a typical angiosperm tetrad structure, consisting of a large and small single-copy region and a separated pair of reverse repeat regions. The other varieties of chloroplast genome appear to be descended from a common ancestor and are most closely related to C. medica and C. indica.
-
Key words:
- Finger lime /
- Breed distinction /
- Chloroplast genome structure /
- Phylogeny