








-
Liriomyza trifolii belongs to the genus Liriomyza of Agromyzidae, Diptera. It is one of the important quarantine pests in China[1]. It invaded into the Taiwan Province in 1988. In only 2 years, damage occurred on a variety of vegetable crops and ornamental horticultural plants[2]. It was discovered in many areas of Hainan Province in 2006, and it gradually spread to Guangxi, Fujian, Jiangsu, and Shandong[3,4]. L. trifolii is a polyvorous pest that can harm more than 300 species of plants in 25 families, including Asteraceae, legumes, Cruciferae, Solanaceae, and cucurbit[5]. The adult feed and lays eggs on the front of leaves and form nearly round puncture holes about the size of needle tips on the surface. Larvae feed on the upper layer of leaves and form irregular snaking worm channels, which changes from thin to coarse with the growth of larvae; they can seriously damage chloroplast cells in the leaves, reduce plant photosynthesis, and even cause plants to fall leaves and eventually die[1].
Hainan has a tropical monsoon climate, and its unique climatic conditions are not only conducive to the growth and development of crops and the accumulation of nutrients but also provide a favorable growth and reproduction environment for L. trifolii[6]. The explosive occurrence of L. trifolii has seriously affected the yield and quality of cowpeas in Hainan Province thereby hindering the healthy development of the vegetable economy industry. At present, chemical control is mainly used in the field control of L. trifolii, in which indoxacarb, pyrethroids, and other pesticides are widely used[7−9]. Given that L. trifolii is prone to develop resistance to insecticides, and its resistance is higher than that of other species, it has replaced other species as the local dominant species in Hainan[5,10] Therefore, the problem of drug resistance has to be considered in the field control of L. trifolii. The biological characteristics of the clover spot loon must be further analyzed, chemical, physical, and biological control methods should be combined to delay its resistance to insecticides to achieve safe and effective field prevention and control[9].
L. trifolii has strong reproduction ability and serious overlapping of generations. Hainan Province is located in the southern tip of China, and the suitable temperature in the region promotes the occurrence and spread of L. trifolii, which is becoming an increasingly serious concern. Whether the biological characteristics of L. trifolii change with the alterations in surrounding environmental factors also need to be further discovered[11]. In this study, the resistance levels of two detoxifying enzymes and acetylcholinesterase in the population of L. trifolii in a field in Sanya in 2022 were measured. On this basis, the activity levels of two detoxifying enzymes and their correlation with resistance were analyzed. The growth and development of L. trifolii at different temperatures were investigated, and the starting point temperature and effective accumulated temperature were calculated according to the effective accumulated temperature law. Finally, by feeding the samples with different foods, we studied the changes in the development period of L. trifolii under various temperatures. The results of this study will provide a scientific theoretical basis for predicting and forecasting the occurrence and damage of L. trifolii, strengthening quarantine control, and formulating green prevention and control strategies.
-
The sensitive strain was donated by Professor Du of Yangzhou University and has not been exposed to any chemical agents in the room for nearly 15 years. The field strain was collected from Yacheng Town, Yazhou District, Sanya City, Hainan Province (China) (18.373° N, 109.167° E) in 2022. The field and sensitive strains identified as L. trifolii were collected from the field and fed in the laboratory with live Vigna pea plants (Holland Green Stick Four Seasons Green. Hongliang Seed Co., Ltd., Shouguang City, Shandong Province, China), which had not been exposed to any insecticides for a long time, and then propagated and expanded for the experiment.
Identification of L. trifolii
-
The adults were collected from the field, placed on 75% alcohol absorbent cotton with the back side up, and placed on a slide. The head, abdomen, and wings were observed and photographed with Olympus microscope SZ61 (Olympus Corporation, Japan). Four 1–3 days of L. trifolii after pupation were randomly placed into a 1.5 mL centrifuge tube, and KAPA DNA rapid extraction kit (Beijing Pukerui Biotechnology Co., LTD.) was used to extract their genomic DNA according to the manufacturer's instructions. The DNA barcode fragment of the COI gene was amplified using universal primers LCO1490 (5'-GGTCAACAAATCATAAAGATATTGG-3') and HCO2198 (5'-TAAACTTCAGGGTGACCAAAAAATCA-3'); PCR (Phanta® Max Super-Fidelity DNA Polymerase, Nanjing Nuovizan Biotechnology Co, LTD) was performed thereafter[12]. PCR was conducted as follows: predenaturation at 95 °C for 3 min, denaturation at 95 °C for 15 s, annealing at 56 °C for 15 s, and extension at 72 °C for 45 s for a total of 35 cycles; and elongation at 72 °C for 8 min. PCR purification and recovery kit (American Omega BioTek) were used for purification and recovery, and the products were sent to Haikou Nanshan Gene Biotechnology Co for sequencing. The resulting sequences were obtained using a Basic Local Alignment Search tool in the NCBI database (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ) to compare the homology in the COI gene sequences of the clover spot hidden fly. DNAstar8.1 software (DNASTAR, USA) was used for sequence analysis.Bioassay
-
The bioassay method of L. trifolii was slightly modified according to Yu et al.[13]. About 95% of the original indoxacarb was purchased from Beijing Yingtaijiahe Biotechnology Co., LTD. The original drug was diluted into 100,000 mg/L mother liquor with acetone and treated with 0.1, 1, 10, 100, and 1,000 mg/L field strains by 0.1% Triton H2O. Subsequently, 0.1% Triton H2O was diluted into 0.01, 0.1, 1, 10, and 100 mg/L, and 0.1% Triton H2O was set as the control group in both treatments. The leaves of cowpea with the second-stage larvae of L. trifolii were selected, immersed in different concentrations of insecticide, removed for 5 s, and left to dry naturally at room temperature. The petiole was moistened with absorbent cotton soaked in sterile water, placed in a petri dish (90 mm × 20 mm), and covered with wet filter paper to prevent the escape of L. trifolii. The total number of larvae treated with each concentration was 10−20, and the treatment was repeated three times. If the pupae did not normalize at 72 h, then it was considered dead.
Determination of enzyme activity
-
The second-instar larvae of the indoor sensitive strain and the Sanya field strain were 0.01 g (about 50 larvae). The LC50 of the sensitive strain and the field population was used as the respective treatment concentration, and the leaves were soaked for 5 s and left for 30 min. An enzyme marker (Hangzhou Aosheng Instrument Co., Ltd, Hangzhou, China) was used to measure the absorption at 340, 450, and 412 nm, as described in the manual of the AChE activity detection kit (Beijing Solaibao Technology Co., Ltd, Beijing, China) for the detection of glutathione S-transferase (GST), carboxylesterase enzyme (CarE), and acetylcholinesterase enzyme (AChE). The enzyme activity before and after treatment with indoxacarb was calculated.
Effects of temperature on the development period and survival rate of L. trifolii
-
In the intelligent artificial climate incubator (Ningbo Yanghui Instrument Co., Ltd, Ningbo, China), the constant temperature was set to 16, 25, and 34 °C; the photoperiod was 16-light/8-dark; and the humidity was 60% ± 5%. Male and female adults of the newly emerged Sanya field strains were paired (1:1), fed into a cage with cowpea seedlings (50 cm long, width, and height), and allowed to mate and lay eggs. After 1 h, the cowpea seedlings with eggs were moved to incubate at constant temperatures of 16, 25, and 34 °C. The developmental periods and survival of eggs and larvae at different temperatures were directly observed on the plant, and the developmental periods and survival of pupae and adults were observed in finger tubes (2.4 cm in diameter and 9.5 cm in height). The length and weight of pupae during the development were recorded. Survival rate, developmental starting point temperature (T0), and effective accumulated temperature (K) were calculated at each developmental stage[14]. At least 30 insects were recorded under each temperature treatment, and each treatment group was independently repeated three times, where survival rate = number of surviving insects/total number of insects × 100%.
Effects of food and temperature on the developmental period of male and female adults
-
The male and female adults of the one-day-old L. trifolii were transferred into individual finger-shaped tubes. Ten groups were established, with each tube being treated and sealed with a breathable cotton plug. Sampling capillary tubes (φ = 0.09 cm, h = 11 cm; sold in Shanghai Great Wall Scientific Instrument Mall) dipped in sterile water or 10% honey water (1 g of honey +10 mL of sterile water) were fed in finger tubes, and adult insects were prevented from escaping. L. trifolii fed with different foods was placed in three constant-temperature intelligent artificial climate incubators set at 16, 25, and 34 °C, with a photoperiod of 16-light/8-dark and humidity of 60% ± 5%. Food was added once every 24 h, and the development of male and female adults was observed and recorded. This experiment was independently repeated three times.
Data analysis
-
Virulence was determined using POLO Plus software (LeOra Software Company, USA) to calculate LC50, resistance multiple, 95% confidence interval, slope, and chi-square value[15]. Resistance ratio = LC50 of field strain / LC50 of sensitive strain. Microsoft Excel 2010 (Microsoft, USA), SPSS 21.0 (IBM, USA), and GraphPad Prism 9 (USA) were used for data statistical analysis and correlation analysis, and correlation coefficients were calculated. Univariate ANOVA and Tukey's new complex range method were used to test the significance of differences. According to the developmental period (day) of each state at different temperatures (°C), the starting point temperature and effective accumulated temperature of each state were calculated[14]. The least square method was used in the calculation according to the effective accumulated temperature rule, and the formula was used as per the study by Yang[16].
-
The ratio of the length of the M3+4 terminal segment to the secondary terminal segment was about 3 (Fig. 1a). The inner and outer top bristle areas of the head were yellow (Fig. 1b). As shown in Fig. 1c, the black stripes on the back of the abdomen of L. trifolii were interrupted, and a yellow stripe formed in the middle. A yellow notch could occasionally be observed in the lower middle of the second to the last clear black stripe (Fig. 1d).

Figure 1.
Anatomical morphology male adult L. trifolii. (a) Forewing, (b) head, (c), (d) dorsal side of the abdomen.
The mitochondrial COI gene was amplified in the pupae of L. trifolii, and PCR results showed a single bright band at the expected location (Fig. 2). A COI gene sequence with a length of about 700 bp was obtained, which was blasted in the NCBI database. The results showed that the sequence was 99.84% consistent with the COI gene of L. trifolii (Gene Bank entry number: ON565812.1). The contents of the A, G, C, and T bases in the measured COI gene sequence were 30.15%, 22.7%, 18.8%, and 28.4%, respectively. The content of A + T was 58.5%, and the content of G+C was 41.5% (Fig. 3). The results revealed an obvious bias toward the A+T base, which was consistent with the characteristics of the base composition of insect mitochondrial genes[17].
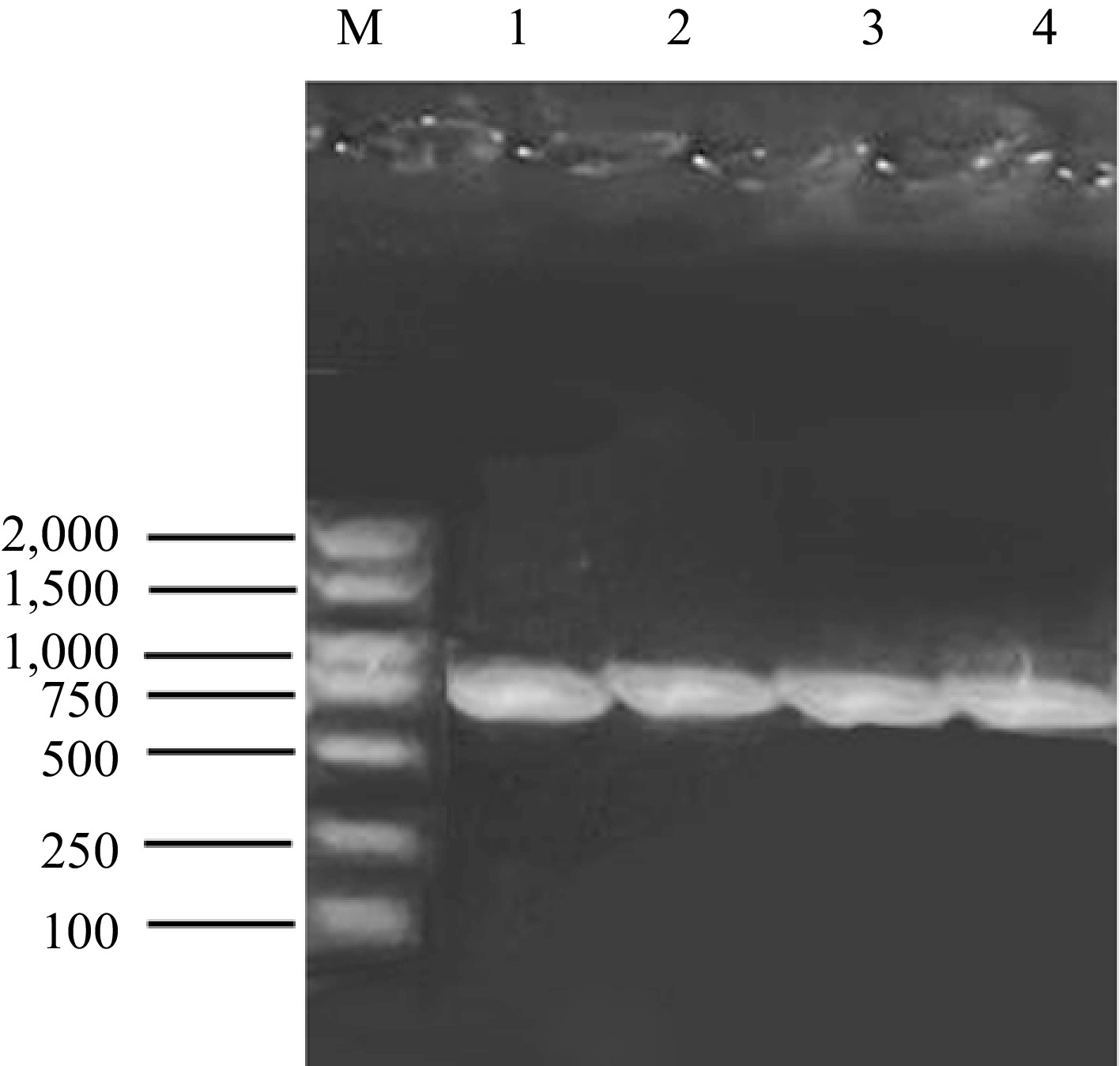
Figure 2.
PCR of the COI gene of L. trifolii. M showed DNA Marker DL2000 (Purchased from TaKaRa, Japan); Lines 1−4 represent four repetitions of PCR.
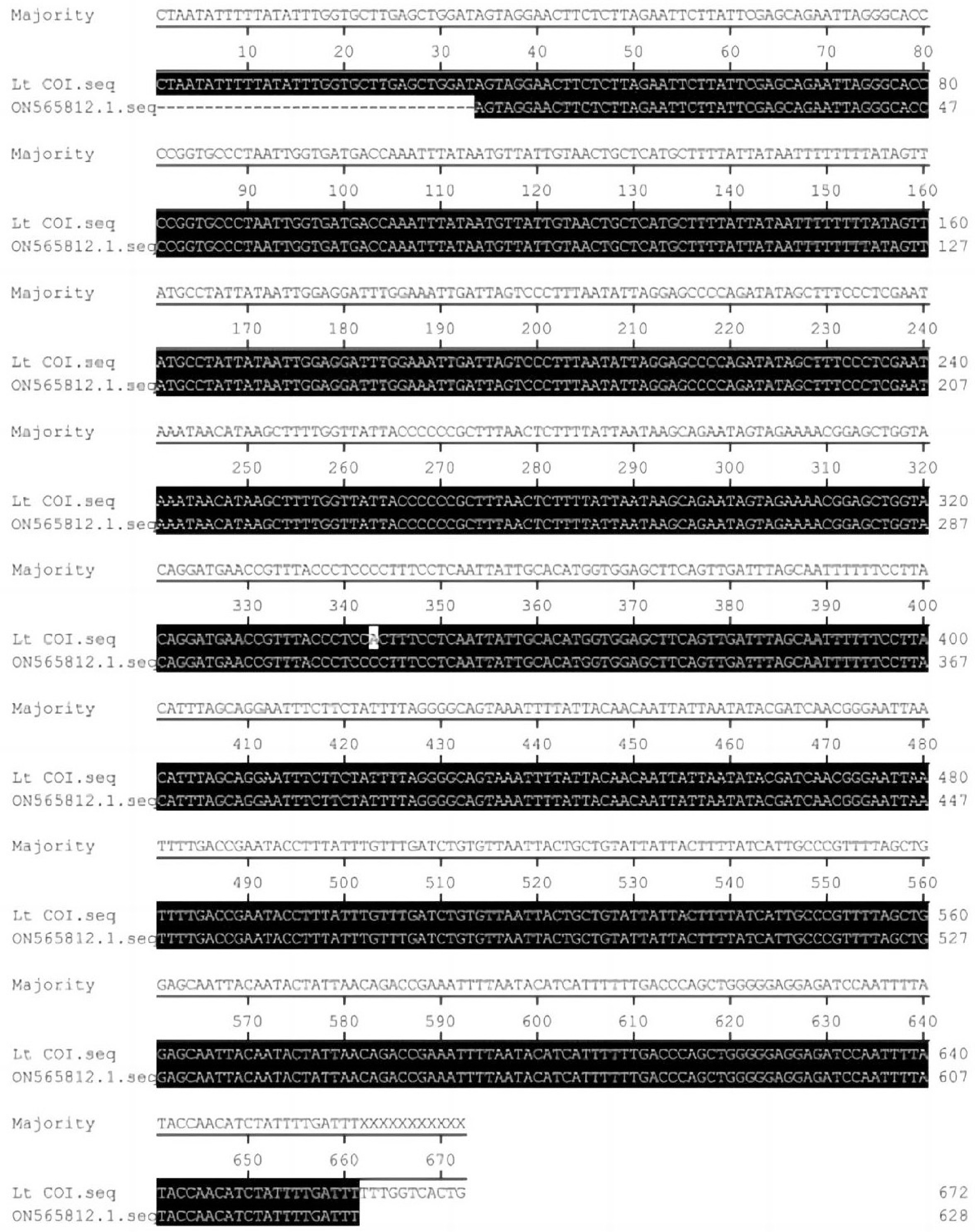
Figure 3.
Sequence comparison between amplified and the COI gene from the NCBI database.
Toxicity of indoxacarb to the second-instar larvae of L. trifolii and its effects on enzyme activities of three enzymes in vivo
-
The LC50 of indoxacarb against the second-instar larvae of the sensitive strain was 0.89 mg/L, the 95% confidence interval was 0.59−1.36, and χ2 (df) was 11.632 (13). The resistance ratio of the second-instar larvae to indoxacarb was 1. The LC50 of indoxacarb against the second-instar larvae of the field strain was 762.59 mg/L with 95% confidence interval of 203.58−16,554.02, and χ2 (df) was 6.17 (13). The resistance ratio of the second-instar larvae to indoxacarb reached 856.84 times. In the 95% confidence interval, the critical value of the chi-square test with 13 degrees of freedom was 22.362, and the virulence measurements were consistent with the probability model (Table 1).
Table 1. Resistance level of indoxacarb resistance of Sanya population of L. trifolii in Hainan province.
Population Year LC50 (95% CL)
(mg/L)Slope ± SE χ2 (df) Resistance
ratioaSensitive strains − 0.90
(0.29−3.15)1.00 ± 0.10 46.68 (10) 1 Sanya 2022 697.00
(241.80−
13,809.27)0.68 ± 0.23 4.76 (13) 776.17 a Resistance ratio = LC50 of field strain / LC50 of sensitive strain. The activities of GST, carboxylesterase, and acetylcholinesterase in the indoor-sensitive and second-instar larvae of the Sanya field strain before and after indoxacarb treatment were measured. As shown in Table 2, the activities of two detoxification enzymes and acetylcholinesterase in the second-instar larvae of the Sanya field strain before and after indoxacarb treatment increased compared with those of the sensitive strain. Before treatment, the activities of GST, carboxylesterase, and acetylcholinesterase in the second-instar larvae of field strain increased by 15.35, 4.51, and 1.49 times compared with those of sensitive strain, respectively. After treatment, the activities of GST, carboxylesterase, and acetylcholinesterase in the second-instar larvae of the field strain increased by 10.09, 9.26, and 2.06 times compared with those of the sensitive strain, respectively. In addition, indoxacarb treatment could increase the activities of GST, CarE, and AChE in both the sensitive and field strains of the second-instar larvae, but only GST activity significantly increased in the sensitive strains of the second-instar larvae (p < 0.05). Only AChE significantly increased in the second-instar larvae of the field strain (p < 0.05).
Table 2. Activities of two detoxification enzymes and acetylcholinesterase enzyme (U/mg pro) in the second-instar larva of L. trifolii after treatment with LC50 (697.00 mg/L) indoxacarb.
Detoxification
enzymeBefore indoxacarb treatment Increased multiplier After indoxacarb treatment Increased multiplier Sensitive strain Field strain in Sanya Sensitive strain Field strain in Sanya GST 0.23 ± 0.27 c 3.53 ± 0.29 a 15.35 0.99 ± 0.47 b 9.99 ± 9.45 a 10.09 AChE 50.74 ± 29.29 c 229.02 ± 50.56 b 4.51 65.77 ± 11.74 c 609.16 ± 22.19 a 9.26 CarE 2.70 ± 0.07 b 4.01 ± 0.90 ab 1.49 3.14 ± 0.33 b 6.46 ± 1.66 a 2.06 GST: Glutathione S-transferase; CarE: Carboxylesterase enzymes; AChE: Acetylcholinesterase enzyme. Data in the table are mean ± SD, and different small letters in the same line mean significant difference (p < 0.05) in the enzymatic activity of sensitive strain and field strain in Sanya between before indoxacarb treatment and after indoxacarb treatment by Tukey's multiple test. Correlation analysis between the resistance to indoxacarb and the activities of three enzymes of the second-instar larvae of the field strain of L. trifolii (Table 3) revealed that the correlation coefficient between the resistance to indoxacarb and the activity of GST of the second-instar larvae before indoxacarb treatment was greater than 0.80, indicating a strong positive correlation. The correlation coefficients between the activity of AChE and CarE and the resistance of field strains to indoxacarb were 0.79 and 0.45, respectively, with moderate positive correlation. After indoxacarb treatment, the correlation coefficients between indoxacarb resistance and the activities of AChE, GST, and CarE were 0.85, 0.73, and 0.64, respectively, indicating strong positive correlations among them.
Table 3. Correlation between enzyme activity and the resistance of the second-instar larva of field L. trifolii in Sanya to LC50 (697.00 mg/L) indoxacarb.
Enzyme Regression equation r Before indoxacarb treatment GST y = 0.2337 + 0.0034x 0.82 AChE y = 46.259 + 0.1895x 0.79 CarE y = 2.1688 + 0.0016x 0.45 After indoxacarb treatment GST y = 0.879 + 0.0116x 0.73 AChE y = 54.349 + 0.615x 0.85 CarE y = 2.5721 + 0.0038x 0.64 GST: Glutathione S-transferase; CarE: Carboxylesterase enzymes; AChE: Acetylcholinesterase enzyme. Effects of temperature and food on the development period of L. trifolii.
-
Under different temperature treatments, the development history of each state of the Sanya field strain is shown in Table 4. From 16 to 34 °C, the developmental period of each species decreased with the increase in temperature. The developmental duration of eggs at 16 °C was 6.67 d, which was significantly higher than that at 25 and 34 °C (F(2,6) = 52.33, p < 0.05). The developmental duration of eggs at 25 and 34 °C was 2.67 and 2.33 days, respectively, with no significant difference (p > 0.05). The developmental duration of larvae was significantly different between 16 and 34 °C (13.67, 7.33, and 4.67 d, respectively; F(2,6) = 130.20, p < 0.05). Pupae stopped developing at 34 °C, and the development duration was significantly lower than that at 16 and 25 °C (F(2,6) = 197.24, p < 0.05).
Table 4. Developmental duration of field Liriomyza trifolii in Sanya under treatments with different temperatures and foods.
Developmental stage Food Temperature (°C) 16 25 34 Egg − 6.67±0.58 a 2.67±0.58 b 2.33±0.58 c Larva − 13.67±0.58 a 7.33±0.58 b 4.67±0.58 b Pupa − 17.11±1.84 a 7.00±0.00 b 0.00±0.00 c Adult 10% Honey water ♀ 22.00±6.93 aB 6.33±7.51 bB 3.33±0.58 cB ♂ 27.25±7.63 aA 5.67±2.31 bB 2.67±0.58 cB Sterile water ♀ 4.33±0.58 aC 1.67±0.58 bB 1.33±0.58 cB ♂ 4.67±1.15 aC 1.67±0.58 bB 1.33±0.58 cB Data in the table are mean ± SD, and different small letters in the same row and different capital letters in the same column mean significant difference (p < 0.05) in the developmental duration between different temperatures and food treatments by Tukey's multiple test. After feeding the samples with different kinds of food, the development time of adult L. trifolii was the longest (22.00 d for females and 27.25 d for males) when fed honey water at 16 °C. The developmental period of female adults fed honey water was significantly longer than those fed sterile water (F(3,8) = 19.42, p < 0.005). At 25 and 34 °C, no significant difference in the developmental period of male and female adults fed the same or different food was found (p > 0.05).
Effects of temperature on the survival of L. trifolii.
-
The survival rate of the Sanya field strain was closely related to temperature change at each developmental stage (Fig. 4). At 16, 25, and 34 °C, the egg survival rate was above 90%. The larval survival rate reached 100% at 25 °C, and the pupal survival rate was only 48% at 34 °C. The survival rate of pupae was the highest at 25 °C and only 33.0% at 16 °C. The survival rate of pupae at 34 °C was 0, which was significantly lower than that of larvea and eggs (F(2,18) = 22.75, p < 0.01).
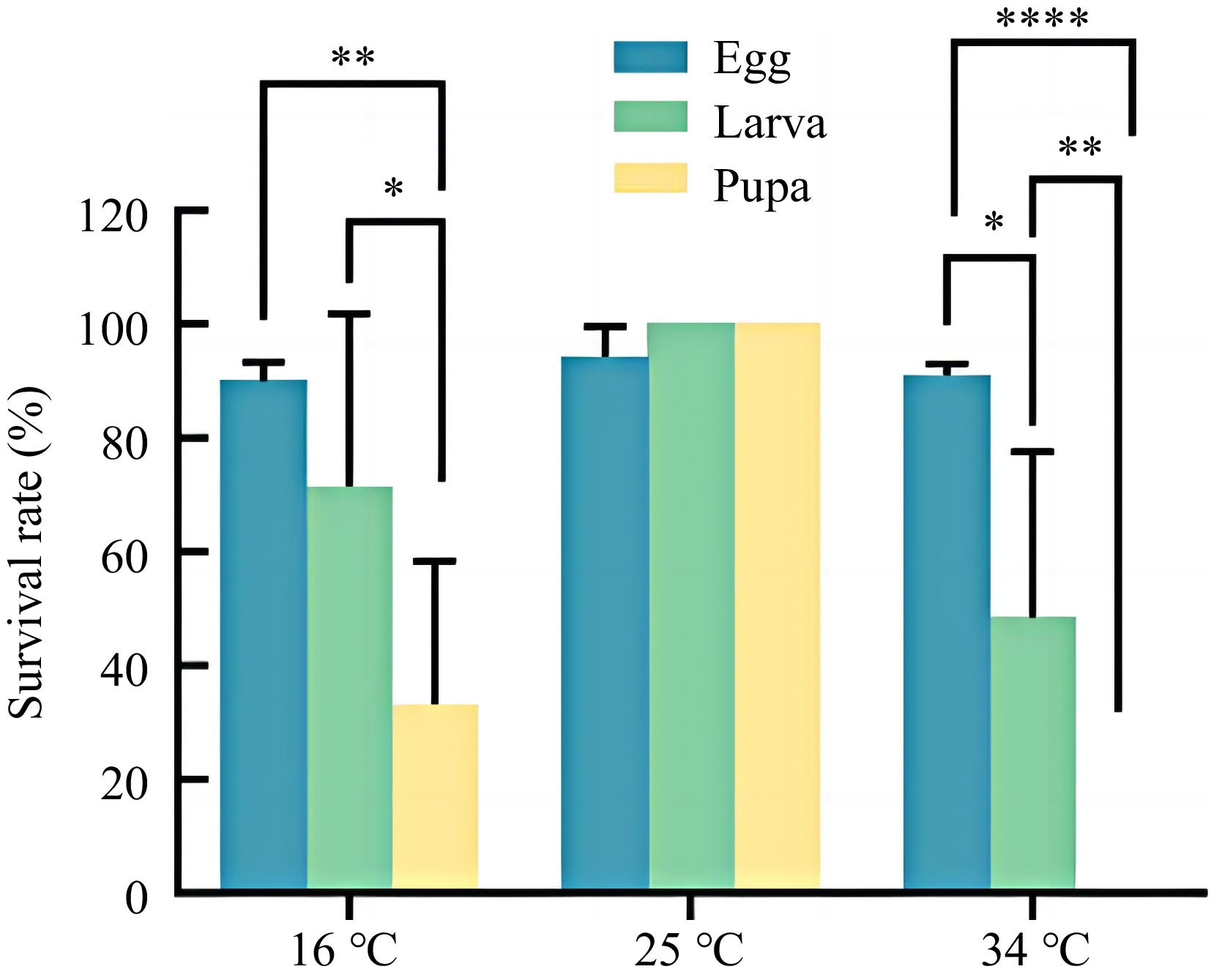
Figure 4.
Survival rate of field L. trifolii in Sanya under different temperatures. Data in the figure are mean ± SD. Asterisks above bars indicates significant difference (p < 0.05) between two groups by Tukey's multiple test.
Effect of temperature on the pupae of L. trifolii
-
As shown in Table 5, the body length of pupa at 16−34 °C was 0.12−0.16 cm. With the growth and development of pupa at different temperatures, its body length and body weight did not change significantly (p > 0.05).
Table 5. Body length and body weight of pupae of field L. trifolii in Sanya under different temperatures.
Number of
tested insectsTemperature Body length Body weight 30 16 0.2 ± 0.01 a 0.01 ± 0.004 a 0.2 ± 0.01 a 0.03 ± 0.02 a 0.2 ± 0.01 a 0.02 ± 0.02 a 30 25 0.1 ± 0.02 a 0.02 ± 0.02 a 0.1 ± 0.02 a 0.05 ± 0.04 a 0.1 ± 0.02 a 0.03 ± 0.007 a 30 34 0.1 ± 0.006 a 0.09 ± 0.002 a 0.1 ± 0.004 a 0.02 ± 0.009 a 0.1 ± 0.004 a 0.06 ± 0.0006 a Data in the table are mean ± SD. Different small letters in the same column mean significant difference (p < 0.05) in the body length and body weight between different temperatures by Tukey's multiple test. Starting point temperature and cumulative temperature of each stage of development of L. trifolii
-
As shown in Table 6, the starting temperature of larval development was the lowest at only 2.75 °C, and the effective accumulated temperature was 17.53 day·°C. The starting temperature of pupal development was the highest at 12.30 °C, and the effective accumulated temperature was 110.23 day·°C. The egg had a starting temperature of development of 10.82 °C. The starting temperature of adult development is 7.11 °C. The effective accumulated temperatures of egg and adult are 43.23 and 8.37 day·°C, respectively.
Table 6. Developmental threshold temperature (T0) and effective accumulated temperature (K) of field L. trifolii in Sanya at different stages.
Development stage T0 K Egg 10.82 ± 1.70 43.23 ± 4.77 Larva 2.75 ± 8.72 17.53 ± 6.20 Pupa 12.30 ± 1.70 110.23 ± 12.49 Adult 7.11 ± 6.57 8.37 ± 2.98 -
The growing areas of the inner and outer top bristles on the head of Liriomyza are diverse. Chen[18] found that more than 70% of the inner and outer top bristles are yellow, about 20% of the inner top bristles are yellow, about 10% of the inner top bristles are yellow and the outer top bristles are black. In all the insects collected in this study, the outer top bristle growth area was yellow, and no black growth area was found (Fig. 1). Many species of Liriomyza have been reported; the individual insects are small, and the abdominal pattern is changeable. Through observation and comparison, the abdominal characteristics of L. trifolii are the most obvious and the simplest way to distinguish L. sativae and L. chinensis. The COI gene can be used as a molecular basis for species identification of L. trifolii to distinguish it from other species of L. huidobrensis[18]. In this study, the molecular identification of the collected COI gene was conducted, and the COI gene sequence of the collected COI gene was 99.84% consistent with that of the NCBI database. Aided by morphological characteristics, the collected specimens were identified as L. trifolii.
Since the invasion of L. trifolii, many studies on its biological characteristics and dynamic monitoring of damage have been published, but research on its resistance level, especially on the resistance level of indoxacarb, is limited. Only Li et al. studied the resistance of the Sanya field population, the Wuzhishan population, and Ledong population to indoxacarb in Hainan Province; they found that the resistance level of the Sanya field population to indoxacarb was the highest in 2021, LC50 reached 631.80 mg/L, and the resistance multiple reached 768.61 times[7]. By contrast, the resistance ratio of the Wuzhishan and Ledong populations to indoxacarb was 531.16 and 140.03 times, respectively. In this study, the resistance to indoxacarb of the Sanya field strain in Hainan Province in 2022 was up to 856.84 times, which was higher than the resistance level reported by Li et al. Given that indoxacarb was used, field populations of various insects have developed different levels of resistance to the insecticide. Zhao et al.[19] monitored the resistance of Plutella xylostella population in Hawaii and found that the population of P. xylostella in Hawaii is not sensitive to indoxacarb and has reached a high level of resistance. Sayyed et al.[20] found that Spodoptera litura field population in the Multan area of Pakistan had developed a moderate level of resistance to indoxacarb, with a resistance multiple of 15 times. The results of this study showed that the resistance to indoxacarb in the field population of Sanya was high. By further measuring the activity of detoxification enzyme between the sensitive and field populations, we found that the activity of AChE in the field population was higher than that of the sensitive strains, and the activity of AChE and CarE was significantly induced after treatment with indoxacarb (Table 2). Many studies also showed that detoxification enzymes might be involved in the resistance mechanism of insects to indoxacarb. For example, Shono[21] found that the multifunctional oxidase improves the resistance of Musca domestica to indoxacarb. Different esterases in S. litura all play a role in resistance to indoxacarb to a certain extent[22]. Similarly, Li et al.[23] pointed out that the activation and metabolism of indoxacarb in P. xylostella may be related to esterase. This finding was similar to the results of this study, indicating that esterase plays an important role in the resistance of L. trifolii to indoxacarb (Table 3).
In addition to pesticides and other non-natural factors in the field that can affect insects, natural factors such as temperature have an important impact on the growth and development of insects. The developmental starting point temperature and effective accumulated temperature are the basic parameters and biological characteristics of insect growth and development. At a suitable temperature range, the growth, development, and reproduction rates of insects are significantly accelerated. Studies of the biological characteristics of L. trifolii can be traced back to 2011[11] and even to the 1980s. In Hainan Province, the number of L. huidobrensis has declined sharply and even gradually replaced by L. trifolii, and the difference in its biological characteristics may be one of the reasons for this phenomenon. As a result of the temperature variation range and light conditions in Hainan Province, the field environment is continuously simulated. In this study, three temperatures were selected to explore the development history and survival rate of each state of L. trifolii, and the developmental starting temperature and effective accumulated temperature were calculated. The results showed that temperature influenced the growth, development, and reproduction of L. trifolii. With the increase in temperature, the time required by L. trifolii to complete a stage was gradually shortened (Table 4). The influence of temperature on different insect states varied; the pupa was the most sensitive to temperature, whereas the larva was the least sensitive, which was consistent with the results of Xiao et al.[11]. At 34 °C, the pupae cannot emerge, but 16−34 °C is suitable for the development of the eggs and larvae of L. trifolii (Fig. 2).
Although the same insect was used in the present experiment, the time difference of 10 years and the space difference of 20,000 km, which led to many generations of variation and adaptation, resulted in obvious differences in their biology. In this study, the starting temperatures of eggs, larvae, pupae, and adults were 10.82, 2.75, 12.30, and 7.11 °C, respectively. According to the study of Xiao et al., the developmental starting point temperatures of eggs, larvae, and adults are 10.80, 6.43, and 8.40 °C, respectively[11]; the developmental starting point temperatures of larvae were significantly different from those in this study. China has a vast land area, and the climate difference between the north and the south is substantial. The climate diversity in China may be the reason for the variation in L. trifolii loon in different places[19,24]. However, different from other similar studies, at least five temperatures were used to calculate the starting point temperature and effective accumulated temperature. In this study, the developmental starting point temperature and effective accumulated temperature of each state were measured at 16, 25, and 34 °C, so the developmental starting point temperature and effective accumulated temperature of each developmental stage measured in this study could be used as a reference (Table 6). In this study, the adult development time of L. trifolii fed honey water significantly increased. Notably, the developmental period of female adults fed honey water at 16 °C was significantly higher than that of female adults fed sterile water (Table 4). Therefore, changing the nutritional state of the adult can change its development period, which also provides ideas for reducing the population density of L. trifolii.
In summary, this study found that indoxacarb was no longer suitable for field control of L. trifolii in Sanya, and other agents such as abamectin and cyromazine should be reduced or mixed. Molecular control methods could be found by studying the physiological and biochemical mechanisms of its detoxification enzyme. Above 34 °C, the survival rate of the pupae decreased, and the survival rate could be reduced by high-temperature confinement and other methods to reduce the insect population density. Honey water was used to trap and kill adult L. trifolii to reduce its egg production and damage degree. This study has guided a significance for the green control of L. trifolii in the field.
-
The authors confirm contribution to the paper as follows: study conception and design: Gong X, Dong W, Li F, Wu S; data collection: Gong X; analysis and interpretation of results: Gong X, Chen Y; draft manuscript preparation: Gong X, Dong W. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
This study was supported by the National Key Research and Development Project of China (2021YFC2600600), the Hainan Major Science and Technology Project (ZDKJ2021016), the Hainan Major Science and Technology Project (ZDKJ2021007), the Project of Sanya Yazhou Bay Science and Technology City (SCKJ-JYRC-2023-15), and the National Key Research and Development Program of China (2022YFD1401200 and 2022YFD1400900).
-
The authors declare that they have no conflict of interest.
-
Received 31 May 2024; Accepted 15 July 2024; Published online 23 August 2024
-
L. trifolii has developed high resistance to indoxacarb in Sanya.
Detoxification enzymes of L. trifolii maybe involved in the detoxification metabolism of indoxacarb.
Temperature and food have an effect on the growth and development of L. trifolii.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gong X, Chen Y, Dong W, Li F, Wu S. 2024. Toxicity of indoxacarb to the population of Liriomyza trifolii (Diptera: Agromyzidae) in Sanya (China), and the effects of temperature and food on its biological characteristics. Tropical Plants 3: e028 doi: 10.48130/tp-0024-0032 

Toxicity of indoxacarb to the population of Liriomyza trifolii (Diptera: Agromyzidae) in Sanya (China), and the effects of temperature and food on its biological characteristics
- Received: 31 May 2024
- Revised: 08 July 2024
- Accepted: 15 July 2024
- Published online: 23 August 2024
Abstract: Liriomyza trifolii is mainly distributed in tropical and subtropical regions, and it is one of the important invasive pests in China, which can damage a variety of plants. L. trifolii has caused serious economic losses to agriculture in China. Morphological and molecular characterization results showed that the collected Sanya field strain was L. trifolii. Bioassay results showed that the sensitivity of the 2nd instar larvae of the Sanya field strain in 2022 to indoxacarb was reduced by 776.17 times. The correlation between the activities of GST and AChE and the resistance of indoxacarb was higher. Temperature affected the developmental period and survival rate of different stages of field strains of L. trifolii, with a greater effect on the survival rate of nymphs, but no effect on the body length and weight of pupae. At 16, 25, and 34 °C, the developmental periods of larvae were shortened with the increase in temperature. The developmental periods of adults fed with honey water were significantly longer than those fed with sterile water. Based of the temperature and developmental period, the development point temperatures of eggs, larvae, pupae, and adults of L. trifolii were 10.82, 2.75, 12.30, and 7.11 °C, respectively. The results of this study may provide important theoretical support for resistance monitoring, management, and control strategies for this insect.
-
Key words:
- Liriomyza trifolii /
- Indoxacarb /
- Enzyme activity /
- Temperature /
- Biological characteristics