









-
Abiotic stresses such as drought, salinity, and oxidative significantly impact plant growth, development, and productivity, particularly in staple crops grown in challenging environments[1]. Drought stress, in particular, leads to a cascade of physiological and molecular changes, including the accumulation of ROS, which cause cellular damage and impair plant growth[2,3]. Plants have evolved complex mechanisms to counteract these effects, including the modulation of antioxidant enzyme activities and the regulation of ROS homeostasis through the expression of stress-responsive genes[4,5]. Under stress conditions, Fe2+ can activate H2O2 to form hydroxyl radicals (·OH), which possess extremely strong redox potential capable of damaging cellular structures. This disruption can impair normal cellular functions and, in severe cases, lead to cell apoptosis and death[6]. Among the antioxidant-related proteins, ferritins are iron storage proteins that play a pivotal role in maintaining cellular iron homeostasis and regulating oxidative stress responses. These proteins mitigate the toxic effects of excess iron by sequestering it in a biologically inert form, thereby preventing the generation of hydroxyl radicals via the Fenton reaction (Fe2+ + H2O2→Fe3+ + OH− + ·OH)[7]. In addition to their well-established role in iron storage, ferritins have been implicated in the regulation of abiotic stress responses in plants. The TaFER5D-1 gene was found to be significantly up-regulated under drought and salt stresses in wheat, over-expression of TaFER5D-1 in Arabidopsis conferred greater tolerance to drought and salt stress[8]. Plant ferritins are localized mainly in chloroplasts, but they can also be targeted to mitochondria, ATFER4 whose role in counteracting the environmental or developmental oxidative conditions in Arabidopsis is ancillary to that of the other isoforms, regardless of its mitochondrial localization[9]. Our experimental results demonstrate that cassava MeFER4 is predominantly localized in chloroplasts, while functional annotation analyses suggest its potential involvement in regulating iron storage and homeostasis. Previous studies confirmed that MeFER4 was significantly associated with drought-related marker-traits[10], which could affect the balance of ROS under stress conditions. Despite these indications, the specific role of MeFER4 in stress tolerance, particularly in response to drought and oxidative stress, remains unclear.
Cassava, a major tropical staple crop, is highly susceptible to seasonal droughts, which significantly impact its productivity and yield[11]. Understanding the molecular mechanisms underlying cassava stress responses is crucial for developing more resilient cultivars[10,12]. Due to the specific characteristics of cassava-growing regions, the crop frequently encounters various abiotic stresses, including drought, flooding, salinity, and extreme temperatures. These stresses often lead to abnormal accumulation of ROS in plants, disrupting cassava growth and development, reducing yields, and causing significant economic losses[13−17]. R2R3-MYB transcription factor MeMYB2 responds to drought, low temperature, and ABA treatment[18]. Furthermore, RNAi-mediated suppression of MeMYB2 enhances cold tolerance in transgenic cassava by promoting anthocyanin accumulation, which effectively scavenges ROS[14]. A CC-type glutaredoxin of MeGRXC3 interacted with two catalases of cassava, MeCAT1, and MeCAT2, and regulated their activity in vivo, thereby affecting cassava drought tolerance[2]. MePP2C1 negatively regulated thermotolerance and redox homeostasis by dephosphorylating MeCAT1S112 and MeAPX2S160[19]. Hence, regulating ROS homeostasis thereby enhances cassava stress tolerance, making it an effective pathway for developing stress-tolerant cassava germplasm.
Genome-wide association analysis (GWAS) linked MeFER4 with physiological traits such as malondialdehyde (MDA), proline, and antioxidant enzymes (e.g., catalase, CAT; superoxide dismutase, SOD) across different years[10]. We hypothesize that MeFER4 plays a significant role in regulating ROS homeostasis and enhancing plant tolerance to drought and oxidative stress. To test this hypothesis, we conducted gene mapping, expression analysis, and overexpression experiments in Arabidopsis. Our primary objective was to decipher how MeFER4 modulates plant stress adaptation mechanisms. Specifically, we investigated: (1) its physical/functional interactions with core antioxidant enzymes; (2) its regulatory effects on ROS homeostasis; and (3) its overall impact on plant vitality under a comprehensive set of abiotic stresses: drought, oxidative challenge, salinity, and osmotic imbalance. This study provides new insights into the function of ferritins in plant stress physiology and their potential applications in improving crop resilience.
-
The coding sequence of MeFER4 was isolated from the cassava cultivar SC124 based on its locus in the cassava genome (
https://phytozome.jgi.doe.gov , M. esculenta v8.1). A total of 100 cassava accessions were selected for re-sequencing from germplasm resources collected by our laboratory[10]. Primers were designed to cover the entire candidate coding sequence region (Supplementary Table S1), and the target segments were amplified using polymerase chain reaction (PCR). The resulting sequences were aligned with the MeFER4 coding sequence in the draft cassava genome for SNP identification (Supplementary Table S2). GWAS was performed to analyze the association between SNPs and DTCs following previously described procedures[2,10].Plant materials and growth conditions
-
Arabidopsis thaliana (Col-0) was used as the wild-type plant for all experiments. Transgenic Arabidopsis lines overexpressing MeFER4 (OE-6, OE-7, and OE-10) were generated and confirmed by PCR using eGFP-specific primers (Supplementary Table S1) and Western blot analysis was conducted using anti-GFP and anti-Actin antibodies to verify the expression of the MeFER4-GFP fusion protein in transgenic lines. Plants were grown in a controlled growth chamber under 8 h light/16 h dark photoperiod, 22 °C temperature, and 60% relative humidity.
Gene cloning and vector construction
-
The MeFER4 gene was amplified from SC124 cassava cDNA using specific primers (Supplementary Table S1). The coding sequence was cloned into the pCAMBIA1300 vector with an eGFP tag under the control of the CaMV35S promoter. The resulting construct was transformed into Agrobacterium tumefaciens strain LBA4404 and used for Arabidopsis transformation. The coding sequence was cloned into the pNC-GBKT7 (provided by Dr. Yan Pu[20]) with an MYC tag and GAL4 DNA binding domain under the control of the ADH1 promoter. The resulting construct was transformed into the yeast strain Y2H Gold and used for yeast two-hybrid screening. The coding sequence was cloned into the pCAMBIA1300-NLUC vector with an NLUC tag under the control of the CaMV35S promoter. The resulting construct was transformed into Agrobacterium tumefaciens strain LBA4404 and used for split-LUC assays. The coding sequence was cloned into the pSPYNE-35S vector with a YFP-N tag under the control of the CaMV35S promoter. The resulting construct was transformed into Agrobacterium tumefaciens strain LBA4404 and used for Bimolecular Fluorescence Complementation (BiFC) Assay.
Drought stress treatment
-
For drought stress experiments, two different approaches were utilized. First, mature leaves of Arabidopsis plants were excised for a detached leaf water loss assay, where the leaf water loss rate (LWLR) was measured over 24-h. Second, drought stress was directly applied to potted seedlings by withholding water for 23 d, and leaf wilting was observed. Each sample consisted of homogenized mature leaf tissue (from third to fifth leaves) from three plants of each line and was treated as one of three biological replicates.
Abscisic acid (ABA), hydrogen peroxide (H2O2), methyl viologen (MV), salt, and osmotic stress treatments
-
For ABA sensitivity assays, 3-week-old Arabidopsis plants were with 40 μM ABA for 3 d. Leaves were collected for 3,30-diaminobenzidine (DAB) staining to assess ROS accumulation. Three-week-old Arabidopsis plants were sprayed with 3% H2O2 or 50 μM MV. After 24 h, leaves were collected for DAB staining to visualize ROS accumulation. Arabidopsis seedlings were grown on MS medium supplemented with 110 mM NaCl or 300 mM mannitol for 3 d. Phenotypic changes, including chlorosis and necrosis, were documented, and leaves were collected for ROS assessment by DAB staining. Expression levels of stress-responsive and antioxidant-related genes were analyzed by qRT-PCR.
DAB staining
-
DAB was performed as previously described by Guo et al.[14]. To detect the presence and distribution of hydrogen peroxide in plant cells, mature leaves of 3-week-old Arabidopsis plants were incubated at room temperature for 2 h with 1 mg mL−1 DAB staining solution, which was freshly prepared in 10 mmol L−1 phosphate buffer (pH 7.8) and 0.05% (v/v) Tween 20. Leaves were then immersed in a bleaching solution (ethanol : acetic acid : glycerol, 3:1:1). Average optical density of DAB staining was quantified using the IHC-Toolbox plugin in ImageJ software (
https://imagej.nih.gov/ij ).cDNA preparation, reverse transcriptase PCR, and qRT-PCR
-
Total RNA was extracted from Arabidopsis leaves using 100 mg of leaf tissue and an RNAprep Pure Plant Plus Kit (DP441, Tiangen Biotech, Beijing, China). First-strand cDNA was synthesized using a FastKing gDNA Dispelling RT SuperMix (Tiangen) per the manufacturer's instructions. qRT-PCR was performed with gene-specific primers to analyze the expression of ROS-related marker genes, including MPK11, SOD, CAT1, PER5, PER63, PER66, APX1, APX3, and APX5 (Supplementary Tables S1 & S3). The MeActin gene was used as a reference[14]. Relative expression levels were calculated using the ∆∆Ct method[14]. All PCR reactions were performed using a Step-One Plus Real-time PCR system (ABI, Carlsbad, CA, USA) using TB Green Premix Ex Taq II (Tli RNaseH Plus) (RR820A, TaKaRa, Dalian, Liaoning, China).
Yeast two-hybrid (Y2H) assay
-
Y2H was performed as previously described by Guo et al.[14]. The full-length MeFER4 coding sequence was cloned into the pNC-GBKT7 vector (bait vector) and confirmed by sequencing. The specific primers are listed in Supplementary Table S1. The bait vector and Arabidopsis yeast library plasmid DNA were co-transformed into competent yeast cells and positive clones were selected on a selective synthetic dropout (SD) medium at 30 °C using the Matchmaker Gold Yeast-Two-Hybrid Library Screening System (Clontech-TaKaRa). Positive clones were identified on SD/-Trp/-Leu/-His/-Ade media and sequenced. Candidate interactors were further analyzed. BD-p53 + AD-T and BD-lam + AD-T served as positive and negative controls, respectively.
BiFC assay
-
For the BiFC assay, MeFER4-NYFP, APX1-CYFP, and MeAPX3-CYFP were constructed according to the protocol of Guo et al.[2]. The coding sequences of MeFER4, APX1, and APX3 were cloned into pSPYNE-35S and pSPYCE-35S vectors, respectively. Agrobacterium tumefaciens strain LBA4404 harboring the BiFC constructs was cultured in LB medium supplemented with 50 mg/L rifampicin and 50 mg/L kanamycin at 28 °C overnight. Then, Agrobacterium carrying the vectors were infiltrated into Nicotiana benthamiana leaves and cultured in a greenhouse for 3 d at 28 °C with 16 h light/8 h dark. After 3 d, the fluorescence from the evaluated protein-YFP fusions and the nuclear marker (MebHLH122, a bHLH transcript factor) signal were detected by confocal microscopy (Leica TCS-SP8, Germany).
Split-luciferase (split-LUC) complementation assay
-
The split-LUC assay was performed as previously described by Guo et al.[21]. To validate the interaction between MeFER4 and its target proteins, we performed a split-LUC complementation assay using a tobacco transient expression system. Recombinant plasmids harboring MeFER4-nLUC/cLUC-fusion constructs (cloned into the pCAMBIA vector) were co-infiltrated into young leaves of Nicotiana benthamiana via Agrobacterium tumefaciens (strain LBA4404)-mediated transformation. At 48 h post-infiltration (hpi), a 1 mM D-luciferin potassium salt solution (Solarbio, China) containing 0.01% Triton X-100 was injected into the abaxial leaf surface. Leaves were dark-adapted for 10 min to facilitate substrate permeabilization. Luminescence signals were captured using a cryogenic CCD imaging system (Night SHADE LB985, Berthold Technologies, Germany) at 22 °C with a 5-min exposure time. Photon flux quantification was conducted using Indigo 2.0 software. To minimize background noise, empty vector-infiltrated leaves were included as negative controls. The experiment included three independent biological replicates, each containing five technical replicates.
Protein structure prediction and docking analysis of MeFER4, APX1, and APX3
-
The protein sequences of MeFER4, APX1, and APX3 were retrieved from the phytozome database (
https://phytozome.jgi.doe.gov , M. esculenta v8.1 and TAIR 10) in FASTA format and preprocessed to ensure sequence quality by removing signal peptides or transmembrane regions.Protein structures were predicted using AlphaFold3 (
https://golgi.sandbox.google.com ), where single-chain models were first generated for each protein, followed by docking models for MeFER4 with APX1 and APX3 through multi-sequence alignment. The AlphaFold3 pipeline was run with de novo prediction settings (use_templates = False, and num_recycles = 3, model quality was assessed using pLDDT and PAE scores. The resulting structures were saved as individual and complex cif files for further analysis.The docking models were visualized in PyMOL (
https://pymol.org ) to identify interaction interfaces, with residues within 5 Å proximity of the binding partner highlighted. Key interaction residues were analyzed for hydrogen bonds, salt bridges, and hydrophobic interactions using PyMOL and Discovery Studio. Visualization included secondary structure representation and residue annotation, with MeFER4, APX1, and APX3 colored green, blue, and interacting residues magenta. Outputs included predicted structures, annotated interaction residues, high-resolution images.Subcellular localization of proteins
-
Subcellular localization analysis was performed as previously described by Guo et al.[14]. A full-length coding sequence of MeFER4 without a stop codon was amplified from SC124 cassava leaf cDNA using gene-specific primers. The fragment was cloned into the pG1300-GFP vector and confirmed by sequencing. The OsPHT4-mCherry plasmid, localized to the chloroplast, was used as a positive control for subcellular localization experiments[22]. Leaves from 6-week-old tobacco (Nicotiana benthamiana) plants were transformed by infiltration, using a 1 mL syringe without a needle, of Agrobacterium cells (OD600 = 1.0) harboring OsPHT4-mCherry plasmid, MeFER4-GFP plasmid, mCherry vector, and 35S:pG1300 vector, were mixed to 1:1, respectively. After 2 d, infiltrated tobacco leaves were examined for GFP and RFP fluorescence and imaged with a confocal laser scanning microscope (FluoView FV1100, Olympus, Japan).
Statistical analysis
-
All experiments were performed with at least three biological replicates. Data were analyzed using one-way ANOVA followed by Tukey's test for multiple comparisons. Differences were considered statistically significant at p < 0.05.
-
In this study, the MeFER4 gene was mapped to Chromosome 8, spanning a region from 25,031,300 to 25,033,800 bp (Fig. 1a). Structural analysis revealed conserved ferritin domains, including Euk-ferritin and Ftn, which are essential for iron storage and homeostasis (Fig. 1a). GWAS analysis linked MeFER4 to DTCs, including MDA, proline, and antioxidant enzymes (CAT, SOD, and peroxidase (POD)) across two years (Fig. 1b). Expression analysis demonstrated that MeFER4 was upregulated under drought stress in the cassava cultivar SC124 (Supplementary Fig. S1a) and in mature leaves of several cassava cultivars with different genotypes (Supplementary Fig. S1b). Subcellular localization confirmed that MeFER4 is localized in the chloroplast (Supplementary Fig. S1b). To further investigate its stress resistance function, we generated MeFER4 overexpression transgenic Arabidopsis lines. PCR results showed the presence of a specific 720 bp (eGFP) band in the OE-6, OE-7, and OE-10 lines (Supplementary Fig. S1c), and Western blotting detected the MeFER4-GFP fusion proteins (~50 kDa), with anti-Actin used as a loading control (Supplementary Fig. S1d). These findings confirm that MeFER4 is responsive to drought stress and that the overexpression lines have been successfully established.
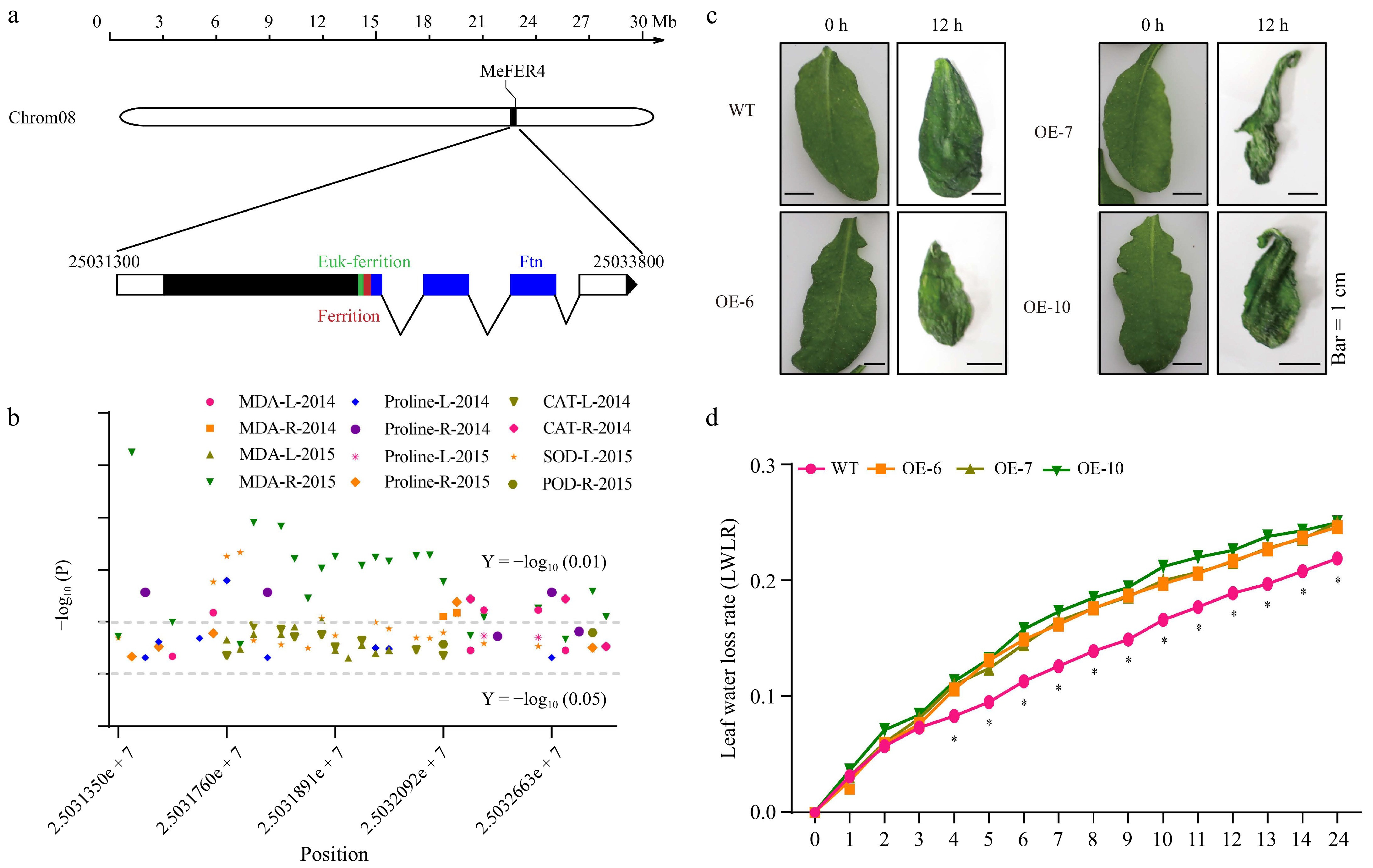
Figure 1.
MeFER4 is associated with drought-related physiological traits in cassava cultivars under drought stress. (a) Schematic diagram of the MeFER4 locus in the cassava genome (version 08). (b) SNPs in the MeFER4 Coding Sequence (CDS) region are correlated with drought-related trait coefficients, including MAD (MDA-L-2014, MDA-R-2014, MDA-L-2015, MDA-R-2015), proline (Proline-L-2014, Proline-R-2014, Proline-L-2015, Proline-R-2015), catalase (CAT-L-2014, CAT-R-2014), superoxide dismutase (SOD-L-2015), and peroxidase (POD-R-2015) in 100 cultivated cassava germplasms under drought stress. Horizontal lines represent significance thresholds at Y = −log10(0.01) and Y = −log10(0.05). (c) MeFER4 affects the water retention capacity of Arabidopsis leaves. (d) Water loss rate of detached leaves within 24 h. Asterisks indicate significant differences from wild-type plants, determined by Student's t-test (* p < 0.05).
Overexpression of MeFER4 heightens the sensitivity to drought stress in Arabidopsis thaliana
-
In the potted seedling drought stress experiment, the transgenic lines exhibited more severe wilting and senescence under drought conditions compared to the wild-type plants (Supplementary Fig. S2a). After 23 d of drought stress, their survival rate was significantly lower than that of the wild-type plants (Supplementary Fig. S2b). DAB staining analysis revealed significantly higher ROS accumulation in the transgenic lines compared to the wild-type plants under drought stress (Supplementary Fig. S2c−d). The transgenic lines enhanced sensitivity to drought stress, as evidenced by severe leaf wilting after just 12 h of dehydration, in contrast to the wild-type plants (Fig. 1c). LWLR analysis showed significantly higher LWLR in MeFER4-overexpressing lines (OE-6, OE-7, OE-10) compared to the wild-type plants, indicating impaired water retention under drought conditions (Fig. 1d). These findings suggest that MeFER4 plays a role in drought stress responses and water retention, potentially through its involvement in regulating oxidative stress.
Overexpression of MeFER4 in Arabidopsis enhances sensitivity to ABA
-
Transgenic lines (OE-6, OE-7, OE-10) exhibited pronounced leaf yellowing compared to wild-type plants under ABA application (Fig. 2a). Moreover, DAB staining indicated higher ROS accumulation in transgenic lines under ABA treatment (Fig. 2a). Quantitative analysis of average optical density confirmed significantly elevated ROS levels in transgenic lines (Fig. 2b).
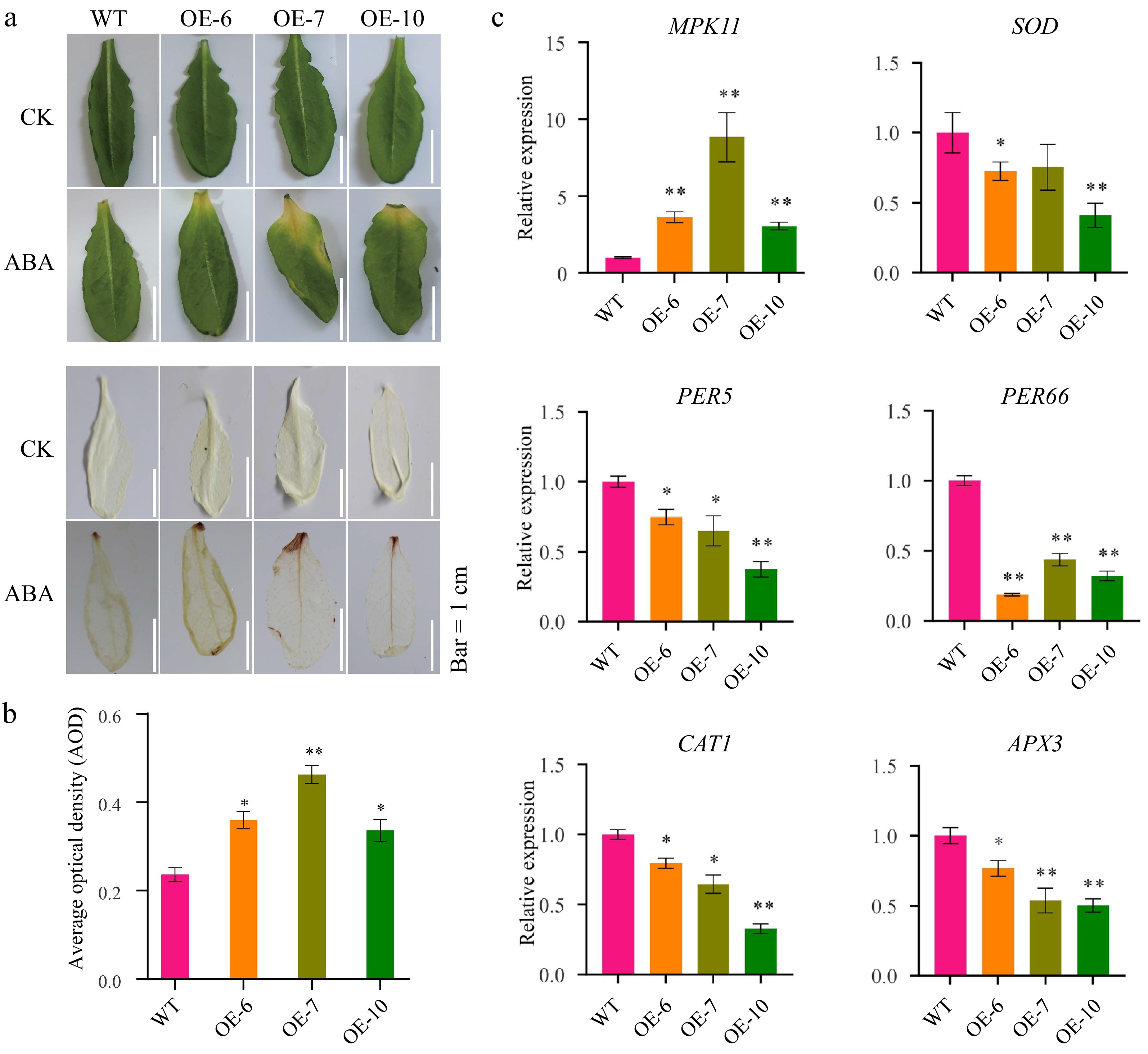
Figure 2.
MeFEE4 transcription level enhances ABA sensitivity and regulates the expression of stress-responsive genes in Arabidopsis. (a) Phenotypic differences in wild-type and MeFER4-overexpressing Arabidopsis lines (OE-6, OE-7, OE-10) under control (CK), and ABA treatment conditions. Top: Leaf coloration and phenotype under CK and ABA treatment. Bottom: DAB staining for ROS accumulation in wild-type and transgenic lines, indicating oxidative stress under ABA treatment. Scale bar = 1 cm. (b) Quantification of ROS accumulation by average optical density in DAB-stained leaves under ABA treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, ** p < 0.01). (c) Relative expression levels of stress-responsive genes, including MPK11, SOD, CAT1, PER5, PER66, and APX3, in wild-type and MeFER4-overexpressing lines under ABA treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, ** p < 0.01).
To investigate the molecular basis of ABA hypersensitivity in transgenic lines, we analyzed key stress-responsive genes in WT and transgenic lines via qRT-PCR (Fig. 2c). MPK11 was significantly upregulated in transgenic lines, with OE-7 showing the highest induction. Conversely, antioxidant genes were suppressed, as SOD, CAT1, and APX3 showed progressive downregulation, with OE-10 exhibiting the most severe reduction. Similarly, PER5 and PER66 were repressed in a line-dependent manner. These findings suggest that MeFER4 overexpression alters ABA signaling and ROS homeostasis under stress.
Overexpression of MeFER4 in Arabidopsis increases sensitivity of transgenic Arabidopsis to oxidative stress
-
Overexpression of MeFER4 in Arabidopsis increased the sensitivity to H2O2 treatment, as evidenced by more pronounced leaf yellowing in transgenic lines (OE-6, OE-7, OE-10) compared to wild-type plants following H2O2 treatment, indicating heightened susceptibility to oxidative stress (Fig. 3a). Elevated ROS levels in transgenic lines were corroborated by stronger DAB staining signals (Fig. 3a). Quantitative analysis of the average optical density of DAB staining further confirmed significantly higher ROS levels in the transgenic lines (Fig. 3b).
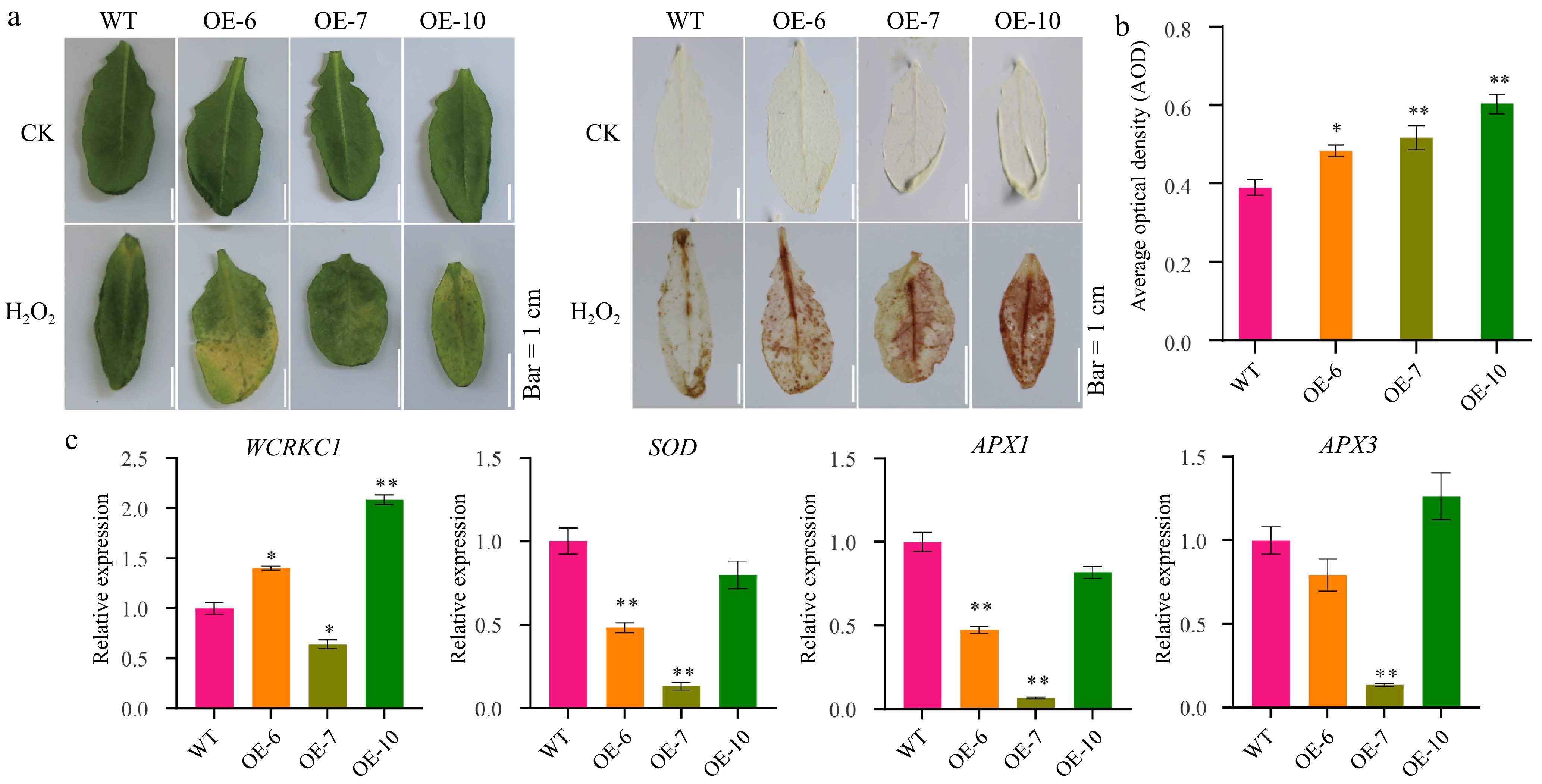
Figure 3.
MeFEE4 transcription level enhances H2O2 sensitivity and regulates the expression of stress-responsive genes in Arabidopsis. (a) Phenotypic differences in wild-type and MeFER4-overexpressing Arabidopsis lines (OE-6, OE-7, OE-10) under control (CK), and H2O2 treatment conditions. Top: Leaf coloration and phenotype under CK and ABA treatment. Bottom: DAB staining for ROS accumulation in wild-type and transgenic lines, indicating oxidative stress under H2O2 treatment. Scale bar = 1 cm. (b) Quantification of ROS accumulation by average optical density in DAB-stained leaves under H2O2 treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, **p < 0.01). (c) Relative expression levels of stress-responsive genes, including WCRKC1, SOD, APX1, and APX3, in wild-type and MeFER4-overexpressing lines under H2O2 treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, ** p < 0.01).
Transcriptional profiling of MeFER4-overexpressing lines revealed a dual regulatory mechanism influencing ROS homeostasis under stress conditions. Quantitative RT-PCR analysis demonstrated significant upregulation of WCRKC1, a kinase associated with oxidative signaling, across all transgenic lines, suggesting enhanced pro-oxidant signaling. Conversely, key antioxidant genes exhibited line-dependent suppression: SOD and APX1 expression decreased markedly in OE-6/OE-7, while APX3 was drastically repressed in OE-6/OE-7, but unaffected in OE-10 (Fig. 3c). This transcriptional reprogramming—elevated kinase activity coupled with compromised ROS scavenging—aligns with observed ROS accumulation patterns (Fig. 3a, b) and suggests that MeFER4 overexpression disrupts redox homeostasis by amplifying oxidative signals while attenuating antioxidant defenses. Notably, the differential suppression of APX3 between OE-10 and other lines may reflect isoform-specific regulatory compensation, highlighting the complexity of ROS metabolic networks.
Transgenic lines exhibited more severe leaf chlorosis and elevated ROS levels compared to wild-type plants (Fig. 4a). This was demonstrated by intensified DAB staining, indicating increased ROS accumulation. Quantitative analysis of average optical density from DAB-stained leaves further confirmed significantly higher ROS levels in the MeFER4-overexpressing lines (Fig. 4b). Expression analysis revealed notable alterations in stress-responsive genes; for instance, genes such as SOD and APX were upregulated, while others like WRKY33 and DREB1B were downregulated under stress conditions (Fig. 4c).
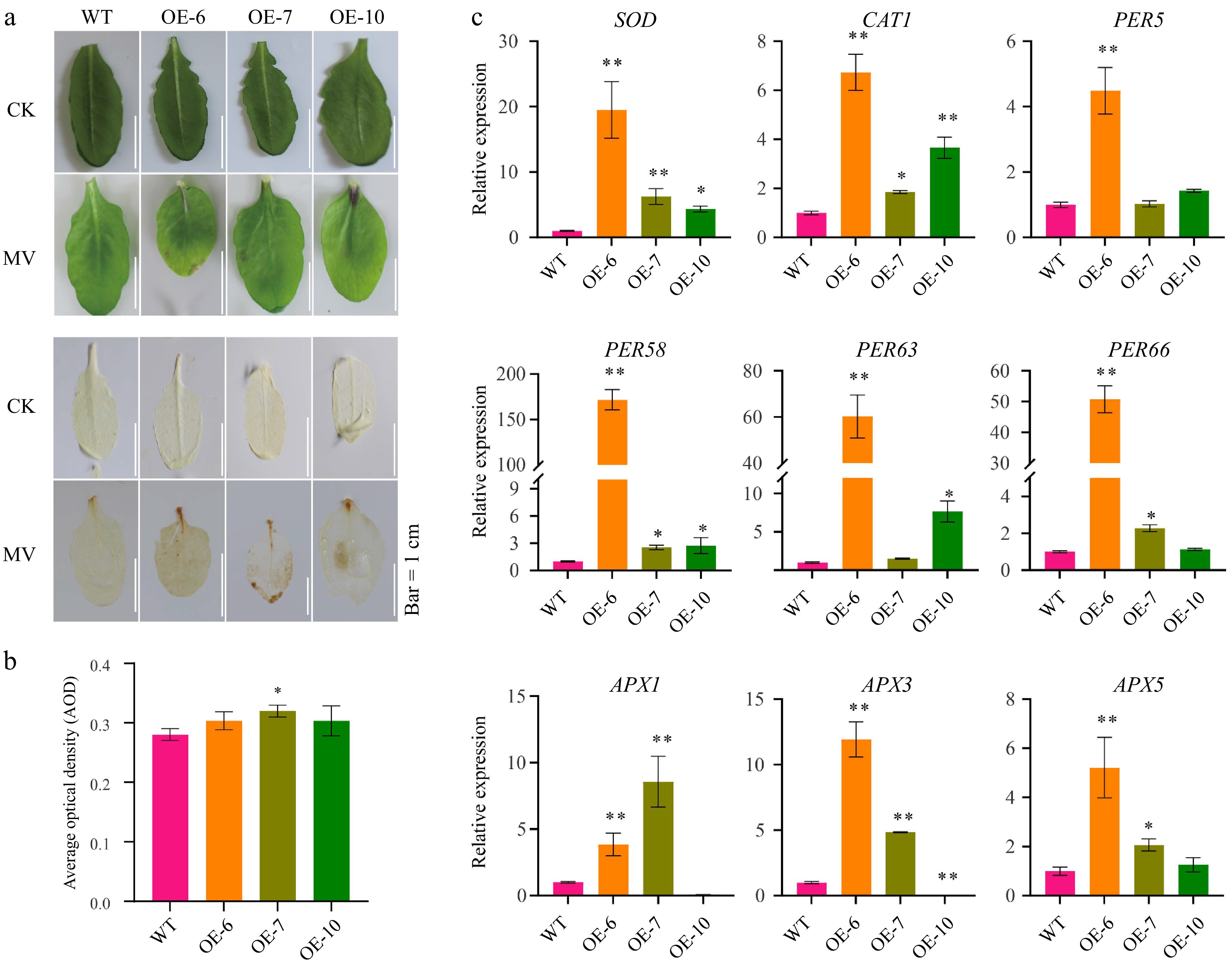
Figure 4.
MeFEE4 transcription level enhances MV sensitivity and regulates the expression of stress-responsive genes in Arabidopsis. (a) Phenotypic differences in wild-type and MeFER4-overexpressing Arabidopsis lines (OE-6, OE-7, OE-10) under control (CK), and MV treatment conditions. Top: Leaf coloration and phenotype under CK and MV treatment. Bottom: DAB staining for ROS accumulation in wild-type and transgenic lines, indicating oxidative stress under MV treatment. Scale bar = 1 cm. (b) Quantification of ROS accumulation by average optical density in DAB-stained leaves under MV treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05). (c) Relative expression levels of stress-responsive genes, including SOD, CAT1, PER5, PER58, PER63, PER66, APX1, APX3, and APX5, in wild-type and MeFER4-overexpressing lines under MV treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, ** p < 0.01).
Overall, these results highlight the role of MeFER4 in regulating oxidative stress responses and suggest that its overexpression in Arabidopsis enhances sensitivity to oxidative stress.
Overexpression of MeFER4 alters stress responses under salinity and osmotic stress
-
Overexpression of MeFER4 in Arabidopsis heightened sensitivity to NaCl stress. Transgenic lines (OE-6, OE-7, OE-10) exhibited more severe leaf chlorosis and necrosis under NaCl treatment compared to wild-type plants (Fig. 5a), along with elevated ROS levels as indicated by intensified DAB staining (Fig. 5a). Expression analysis revealed significant downregulation of stress-responsive transcriptional regulators, such as MYB73, DREB1B, and WRKY33, while antioxidant defense genes, including SOD and APX, were upregulated (Fig. 5c).
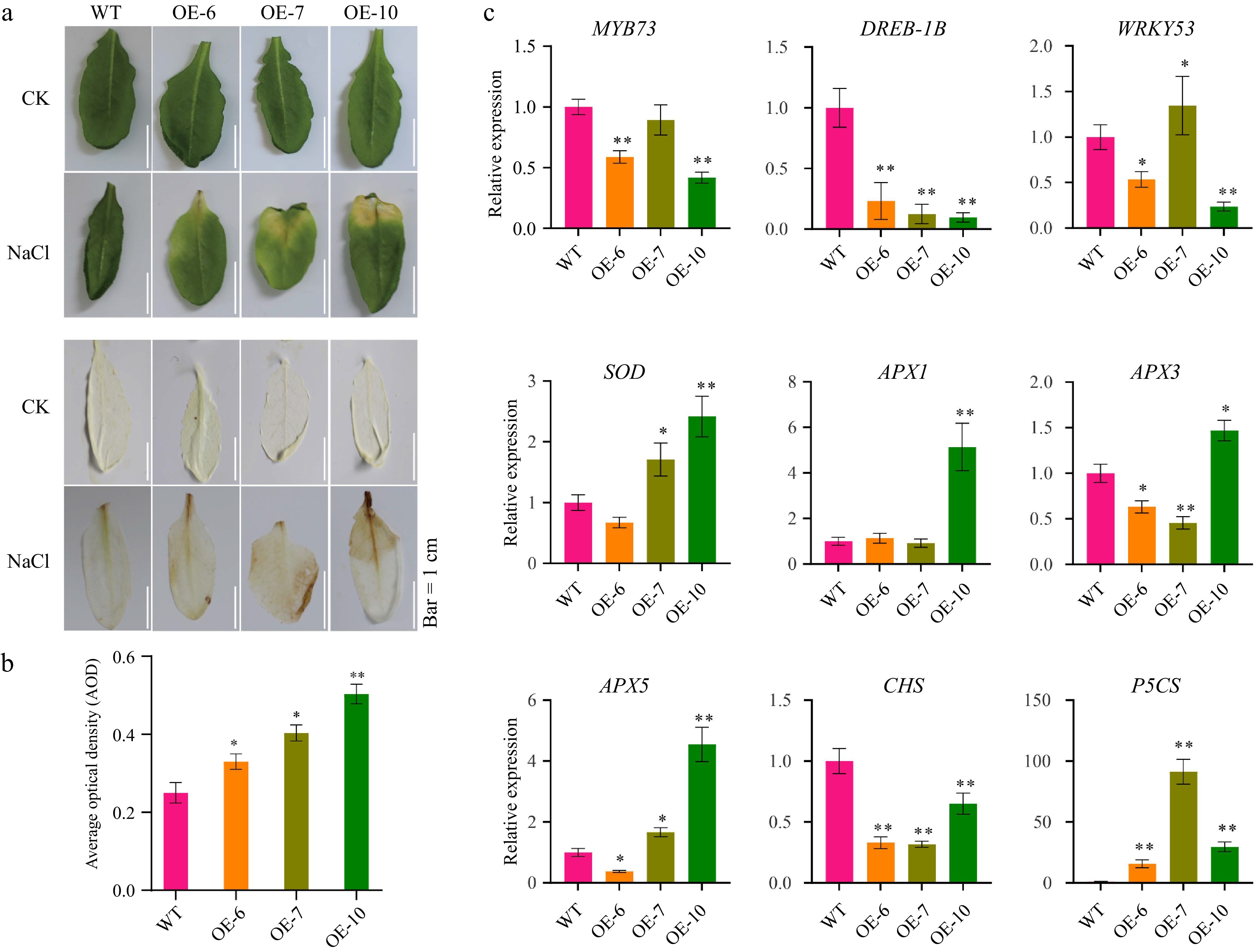
Figure 5.
MeFEE4 transcription level enhances Salinity sensitivity and regulates the expression of stress-responsive genes in Arabidopsis. (a) Phenotypic differences in wild-type and MeFER4-overexpressing Arabidopsis lines (OE-6, OE-7, OE-10) under control (CK), and NaCl treatment conditions. Top: Leaf coloration and phenotype under CK and NaCl treatment. Bottom: DAB staining for ROS accumulation in wild-type and transgenic lines, indicating oxidative stress under NaCl treatment. Scale bar = 1 cm. (b) Quantification of ROS accumulation by average optical density in DAB-stained leaves under NaCl treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, ** p < 0.01). (c) Relative expression levels of stress-responsive genes, including MYB73, DERB-1B, WRKY53, SOD, APX1, APX3, APX5, CHS, and P5CS, in wild-type and MeFER4-overexpressing lines under NaCl treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, ** p < 0.01).
Under mannitol-induced osmotic stress, MeFER4-overexpressing lines demonstrated increased sensitivity, as evidenced by more pronounced leaf discoloration and damage relative to wild-type plants (Fig. 6a). Elevated ROS levels in the transgenic lines were confirmed by stronger DAB staining signals (Fig. 6b). Gene expression analysis showed downregulation of transcription factors such as WRKY53 and DREB1B, whereas antioxidant defense genes, including members of the APX family and P5CS, were upregulated, indicating an adaptive response to oxidative stress (Fig. 6c).
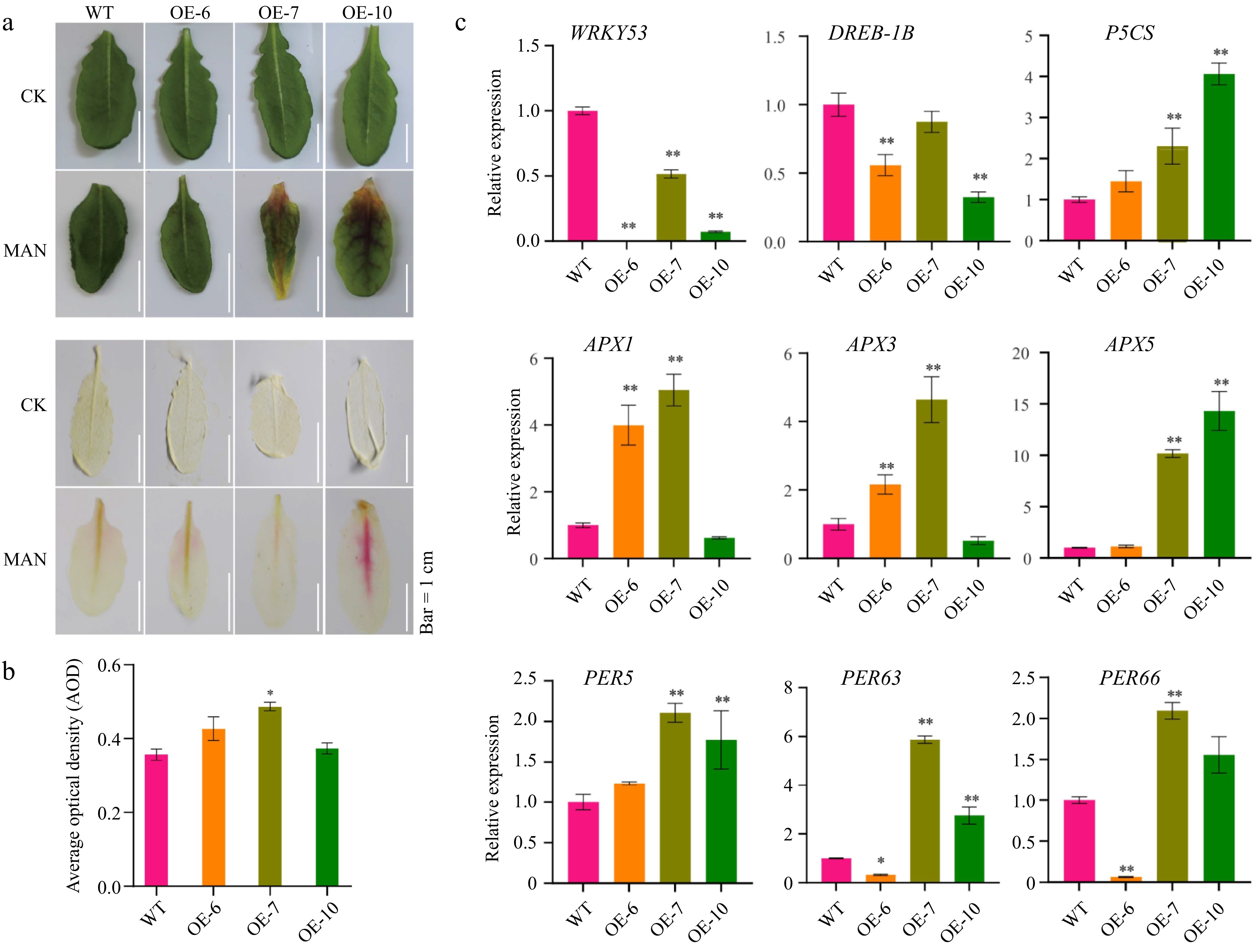
Figure 6.
MeFEE4 transcription level enhances osmotic stress sensitivity and regulates the expression of stress-responsive genes in Arabidopsis. (a) Phenotypic differences in wild-type and MeFER4-overexpressing Arabidopsis lines (OE-6, OE-7, OE-10) under control (CK), and mannitol (MAN) treatment conditions. Top: Leaf coloration and phenotype under CK and MAN treatment. Bottom: DAB staining for ROS accumulation in wild-type and transgenic lines, indicating oxidative stress under MAN treatment. Scale bar = 1 cm. (b) Quantification of ROS accumulation by average optical density in DAB-stained leaves under MAN treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05). (c) Relative expression levels of stress-responsive genes, including SOD, CAT1, PER5, PER58, PER63, PER66, APX1, APX3, and APX5, in wild-type and MeFER4-overexpressing lines under MAN treatment. Data represent means ± SD (n = 3). Asterisks indicate significant differences compared to wild-type plants (* p < 0.05, ** p < 0.01).
In conclusion, these findings suggest that the overexpression of MeFER4 leads to altered stress responses under both salinity and osmotic stress conditions in Arabidopsis.
Interaction of MeFER4 with antioxidant enzymes
-
Yeast two-hybrid (Y2H) screening identified ascorbate peroxidase 1 (APX1) and ascorbate peroxidase 3 (APX3) as interaction partners of MeFER4 (Fig. 7a). To further confirm these interactions, split-LUC assays were conducted. The split-LUC assay results indicated strong fluorescence signals in the presence of both APX1 and APX3, corroborating the Y2H findings and confirming the physical interaction between MeFER4 and these proteins (Fig. 5b).
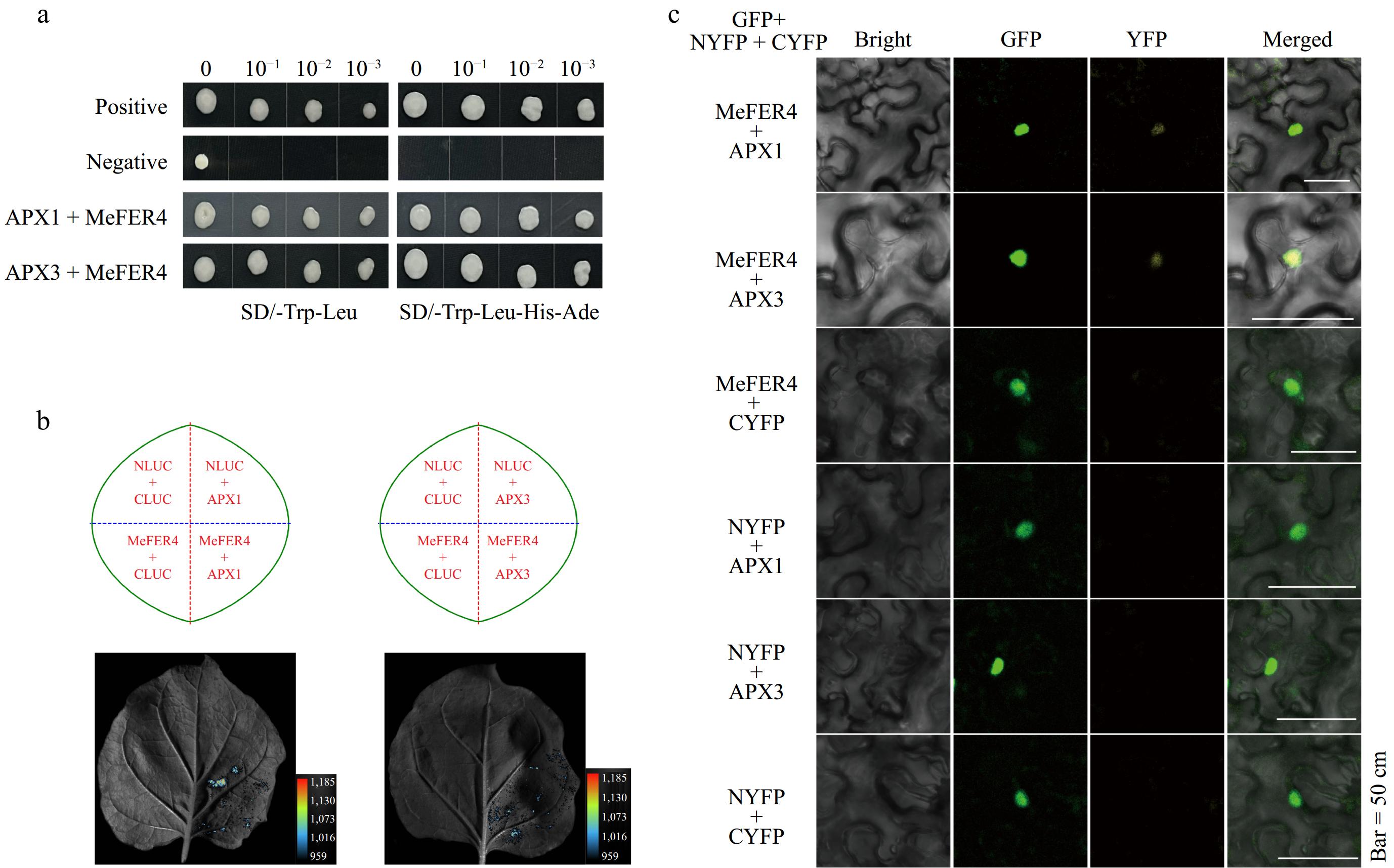
Figure 7.
Interaction analysis of MeFER4 with APX1 and APX3. (a) Yeast two-hybrid assay results showing the interaction between MeFER4 with APX1 and APX3. The positive and negative controls indicate the expected results. Growth on selective media SD/-Trp-Leu and SD/-Trp-Leu-His-Ade demonstrates the interactions between the proteins in APX1 + MeFER4 and APX3 + MeFER4. (b) Split-LUC assay results indicating the interaction between MeFER4 with APX1 and APX3. The left and right panels show the interaction of MeFER4 with APX1 and APX3 in tobacco leaves, respectively. The luminescence signal confirms the protein-protein interaction. (c) BiFC assay results illustrating the interaction between MeFER4 with APX1 and APX3. The GFP, YFP, and merged images demonstrate the fluorescence signal in tobacco leaf cells, further validating the protein interactions. Scale bar = 50 μm.
Furthermore, BiFC assays were employed to validate the interactions in vivo. The results showed clear and distinct fluorescence signals when MeFER4 was co-expressed with either APX1 or APX3, demonstrating that MeFER4 forms complexes with both proteins in living cells (Fig. 5c). These in vivo interaction studies provide robust evidence that MeFER4 physically interacts with APX1 and APX3, suggesting a functional partnership in the regulation of ROS homeostasis.
Collectively, these findings imply that MeFER4 may play a crucial role in modulating ROS levels and stress responses in plants through its interactions with key antioxidant enzymes, APX1 and APX3. The identification and validation of these interactions underscore the potential significance of MeFER4 in maintaining oxidative balance and enhancing plant resilience under stress conditions.
-
In this study, we explored the role of MeFER4, a ferritin-like protein, in regulating oxidative stress responses and its potential involvement in stress tolerance in cassava. Our results suggest that MeFER4 plays a complex role in modulating ROS homeostasis, and its overexpression in Arabidopsis thaliana significantly affects the plant responses to various abiotic stresses, including drought, oxidative stress, salinity, and osmotic stress. These findings provide new insights into the role of ferritins in plant stress physiology and offer potential avenues for improving stress resilience in crops, particularly cassava.
Complex role of MeFER4 in drought tolerance
-
Previous studies have shown that ferritin plays an important role in enhancing iron content in plants, such as rice, pineapple, and banana[23−25]. Ferritin not only stores iron but also helps mitigate oxidative stress-induced damage by regulating intracellular iron levels[6]. Nitric oxide (NO) can induce ferritin accumulation in barley, alleviating seedling damage under salt stress[26]. In maize, the mRNA abundance of ZmFER1 increases in response to H2O2 induction[27]. In white lupin, the ferritin gene FER1 has been shown to participate in antioxidant pathways[28]. Under drought stress, the transcriptional abundance of ferritin2 in Populus is upregulated[29]. Two rice ferritins, OsFER1 and OsFER2, are induced by paraquat, copper, and high iron levels, and it has been found that rice ferritins primarily defend against iron-mediated oxidative stress[30]. In Arabidopsis, ferritin can respond to short-term induction by MV[31,32]. As an important stress-response protein in Arabidopsis, AtFER4 collaborates with other ferritins to counteract oxidative stress caused by environmental stress[33].
In this study, we found that the MeFER4 gene is closely associated with drought tolerance traits in cassava, particularly in terms of MDA, proline accumulation, and antioxidant enzyme activity (Fig. 1a, b). While MeFER4 was upregulated under drought stress in cassava (Supplementary Fig. S1a), which could suggest its role in mitigating oxidative damage by regulating ROS homeostasis, iron storage, and cellular protection. However, overexpression of MeFER4 in Arabidopsis showed heightened sensitivity to drought stress (Fig. 1c). This was evidenced by increased leaf wilting, higher leaf water loss rates, and impaired water retention under dehydration (Fig. 1d). These findings suggest that MeFER4 may play a more complex role in drought responses. Specifically, its overexpression could lead to an imbalance in ROS homeostasis, potentially overloading ROS scavenging mechanisms or interfering with the plant stress response system, thus impairing its ability to cope with drought. Overall, these results highlight the nuanced role of MeFER4 in drought tolerance, where the balance between ROS generation and scavenging is crucial for maintaining plant health under stress.
Role of MeFER4 in oxidative stress sensitivity and interaction with antioxidant enzymes
-
To counteract ROS, plants have evolved enzyme systems, including SOD, CAT, and peroxidase (POD), as well as low molecular weight antioxidants such as ascorbic acid (ASA), carotenoids, and glutathione (GSH)[34]. The expression of ferritin is closely linked to the expression profile of enzymes related to oxidative stress resistance. An increase in ferritin gene expression is often accompanied by enhanced antioxidant enzyme activity. Oxidative stress positively regulates the expression of SOD1, CAT1, and FER1 in Arabidopsis to counteract the accumulation of H2O2. After 24 h of light exposure, the mRNA levels of FER1, SOD1, and CAT1 were elevated, likely due to ROS-induced signaling cascades that triggered ferritin activity[35]. Oxidative stress also induced an increase in the transcript levels of SOD1, CAT1, and FER1 in white lupin[27]. Our findings indicate that MeFER4 plays a significant role in plant responses to oxidative stress and ABA. Overexpression of MeFER4 in Arabidopsis resulted in heightened sensitivity to ABA, as shown by more pronounced leaf yellowing and increased ROS accumulation upon ABA treatment (Fig. 2a). ABA is a key phytohormone that regulates drought and oxidative stress responses, and MeFER4 appears to modulate ABA signaling pathways, potentially disrupting ROS balance. The upregulation of ROS-related genes such as MPK11 and PER63, and downregulation of PER5 and APX5 in transgenic lines, suggests that MeFER4 influences ROS homeostasis in response to ABA (Fig. 2c).
Similarly, overexpression of MeFER4 increased sensitivity to oxidative stress induced by H2O2 and MV, as evidenced by more severe leaf chlorosis and higher ROS levels compared to wild-type plants (Figs 3a & 4a). These results imply that MeFER4 may regulate ROS detoxification but could also disrupt the balance between ROS production and scavenging, contributing to increased oxidative stress sensitivity.
We further explored the impact of MeFER4 under salinity and osmotic stress conditions. Transgenic lines exhibited greater sensitivity to NaCl and mannitol treatments (Figs 5a & 6a), with more severe leaf chlorosis, necrosis, and higher ROS accumulation (Figs 5a, b, & 6a, b). This suggests that MeFER4 may influence a plant's ability to manage osmotic stress and salt tolerance, likely through its interaction with antioxidant defense systems. Despite the increased oxidative stress sensitivity, MeFER4 may still play a role in maintaining cellular integrity. The elevated ROS levels and altered gene expression patterns in transgenic lines indicate that MeFER4 may modulate oxidative signaling pathways (Figs 5c & 6c), but its overexpression could overwhelm the plant's ability to maintain ROS balance, making it more vulnerable to stress-induced oxidative damage.
Additionally, we identified key antioxidant enzymes, APX1 and APX3, as interaction partners of MeFER4 through various assays (Fig. 7). Molecular docking revealed stable interaction interfaces between MeFER4 and APX1/APX3 mediated by hydrogen bonds and salt bridges. Key residues in MeFER4 (e.g., D110, K166, with APX1; E80, E98, with APX3) formed critical bonds with corresponding residues in APX1 (H179, K28) and APX3, ensuring structural stability and functional synergy of the complexes (Supplementary Fig. S3). These interactions suggest that MeFER4 works in concert with APX1 and APX3 to regulate ROS homeostasis, highlighting its role in balancing ROS production and scavenging, which is crucial for plant resilience under stress.
The complexity of ROS homeostasis in MeFER4-OE Arabidopsis under various stress conditions
-
The differential expression patterns of antioxidant enzymes under distinct stress conditions (ABA, H2O2, MV vs salinity/osmotic stress) may reflect the complexity of ROS signaling dynamics and the dual roles of antioxidant systems in stress adaptation. Under ABA and H2O2, the downregulation of key antioxidant genes (e.g., SOD, APX3) (Figs 2c & 3c) in MeFER4-OE lines suggests an overload of ROS scavenging capacity or disrupted coordination between ROS production and detoxification. ABA, as a central regulator of stress responses, enhances ROS production as a signaling molecule, but excessive ROS accumulation under MeFER4 overexpression may overwhelm the antioxidant system, leading to feedback inhibition of genes like SOD and APX3[36,37]. Similarly, MV generates superoxide radicals via electron leakage in chloroplasts, which may directly impair antioxidant enzyme stability or transcription, exacerbating oxidative damage.
In contrast, under salinity and osmotic stress, the upregulation of SOD and APX (Figs 5c & 6c) in transgenic lines implies a compensatory mechanism to counteract stress-specific ROS sources. Salt stress induces ionic imbalance and osmotic stress triggers water deficit, both of which activate calcium signaling and MAPK cascades that stimulate antioxidant gene expression independently of ABA[37]. For instance, the Br14-3-3 gene in Brassica rapa is upregulated under salt and osmotic stress to enhance ROS scavenging, highlighting conserved stress-specific regulatory networks[37]. Additionally, osmotic stress often induces proline accumulation, which stabilizes antioxidant enzymes and mitigates ROS toxicity, potentially buffering the system against MeFER4-induced dysregulation[38].
The interaction of MeFER4 with APX1/3 further complicates this balance. While APX1/APX3 are critical for H2O2 detoxification, their physical association with MeFER4 may alter enzyme activity or substrate availability under specific stresses. For example, in wheat roots, exogenous ferulic acid enhances APX activity under boron toxicity but fails to restore glutathione reductase, illustrating how stress context dictates antioxidant responses[38]. Similarly, MeFER4-OE plants may disrupt APX1/APX3 function under ABA or MV stress but permit their activation under salinity/osmotic conditions due to alternative signaling pathways.
Implications for crop improvement
-
In MeFER4-overexpressing transgenic lines, the expression of MeFER4 leads to a significant downregulation of key antioxidant genes such as SOD, APX, CAT, and POD, which are crucial for mitigating oxidative stress. This downregulation disrupts the normal antioxidant defense mechanisms in the plants. Furthermore, MeFER4 interacts with the antioxidant enzymes APX1 and APX3, which exacerbates the imbalance in the oxidative homeostasis of the plants. As a result, this interference promotes the accumulation of ROS in the cells. The increased ROS levels eventually make the transgenic plants more sensitive to abiotic stress factors, such as drought, oxidative stress, salinity, and osmotic stress (Fig. 8).
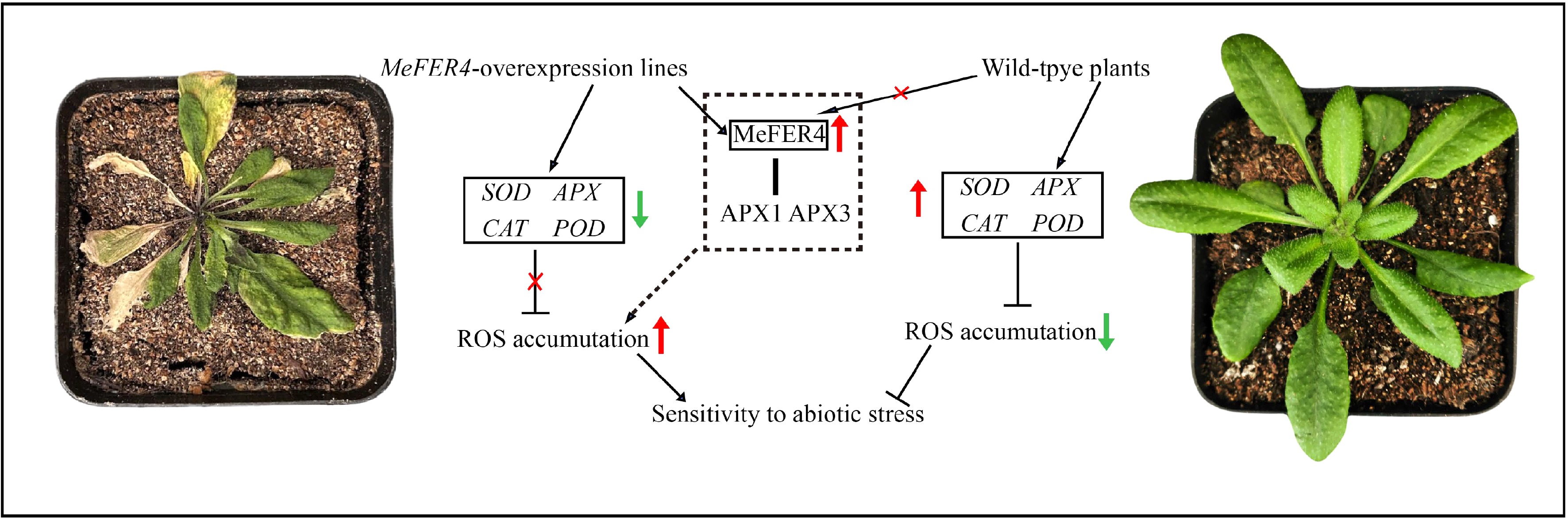
Figure 8.
The mechanism by which MeFER4 promotes ROS accumulation in Arabidopsis under abiotic stress. In MeFER4-overexpressing transgenic lines, the presence of MeFER4 leads to the downregulation of antioxidant genes such as SOD, APX, CAT, and POD, and interacts with APX1 and APX3, ultimately promoting ROS accumulation and increasing sensitivity to abiotic stress. In contrast, in wild-type plants, the absence of MeFER4 allows the normal expression of antioxidant enzyme genes, resulting in higher resistance to abiotic stress.
In contrast, in wild-type plants, the absence of MeFER4 allows the proper expression of antioxidant enzyme genes. This leads to a robust antioxidant defense system, enabling the plants to better cope with abiotic stress. The normal function of these enzymes helps maintain the ROS balance within the cells, contributing to enhanced protection against the damaging effects of abiotic stress. Consequently, wild-type plants exhibit a higher resistance to such stresses compared to the MeFER4-overexpressing lines (Fig. 8).
The findings from this study have important implications for crop improvement strategies, particularly in the context of developing drought-tolerant and oxidative stress-resistant varieties. While MeFER4 appears to play a vital role in regulating stress responses, its overexpression in Arabidopsis resulted in heightened sensitivity to drought and oxidative stress, indicating that a careful balance must be struck between activating stress-related genes and maintaining cellular homeostasis. Further studies are needed to explore the potential of MeFER4 in cassava and other crops for improving drought tolerance and stress resilience. Moreover, genetic manipulation of MeFER4 expression levels in crops could be fine-tuned to optimize its role in stress tolerance without compromising plant growth and productivity.
-
In conclusion, our results demonstrate that MeFER4 is an important regulator of oxidative stress responses in plants. By modulating ROS homeostasis and interacting with key antioxidant enzymes, MeFER4 plays a central role in mediating plant responses to drought, oxidative, and osmotic stress. The findings of this study provide a foundation for further research into the potential applications of MeFER4 in improving stress tolerance in crops, especially in regions where drought and oxidative stress are significant constraints on crop productivity. Further investigation into the molecular mechanisms underlying MeFER4-mediated stress tolerance will be critical for harnessing its full potential in agricultural applications.
We thank Dr. Mengbin Ruan and Dr. Pu Yan for sharing vector plasmids. This research was supported by the Hainan Provincial Natural Science Foundation of China (324MS122), the National Natural Science Foundation of China (32360458), the startup funds for the Hainan University E-class Talents (XJ2300007513), and the China Agriculture Research System (CARS-11-HNCYH), both of which made this research possible.
-
The authors confirm contribution to the paper as follows: study conception and design: Guo X, Wang W, Yu X, Peng M; data collection: Li L, Zheng M, Lin C, Wang B, Yu F, Xie X; analysis and interpretation of results: Guo X, Zhao P, Li J, Zheng M, Wang W, Yu X, Si C, Chen F; draft manuscript preparation: Guo X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0010)
-
Received 9 January 2025; Accepted 12 March 2025; Published online 3 April 2025
-
The MeFER4 gene in cassava was mapped to chromosome 8 and is associated with drought tolerance traits.
Overexpression of MeFER4 in transgenic Arabidopsis increased sensitivity to various stresses, including drought, ABA, H2O2, NaCl, and osmotic stress, with elevated ROS levels and reduced survival rates.
Physical interactions between MeFER4 and antioxidant enzymes APX1 and APX3 were confirmed, suggesting a role of MeFER4 in regulating ROS homeostasis and stress responses.
-
# Authors contributed equally: Mingjuan Zheng, Jianjun Li
- Supplementary Table S1 Primers used in this study.
- Supplementary Table S2 SNPs in MeFER4 genomic DNA.
- Supplementary Table S3 Annotation information of stress-related genes.
- Supplementary Fig. S1 Analysis of MeFER4 expression and subcellular localization, and validation of transgenic Arabidopsis.
- Supplementary Fig. S2 Effects of MeFER4 overexpression on drought stress tolerance in Arabidopsis.
- Supplementary Fig. S3 Molecular docking analysis of MeFER4 with APX1 and APX3.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng M, Li J, Lin C, Wang B, Zhao P, et al. 2025. Functional analysis of MeFER4 in regulating ROS homeostasis during abiotic stress responses in Arabidopsis. Tropical Plants 4: e013 doi: 10.48130/tp-0025-0010 

Functional analysis of MeFER4 in regulating ROS homeostasis during abiotic stress responses in Arabidopsis
- Received: 09 January 2025
- Revised: 04 March 2025
- Accepted: 12 March 2025
- Published online: 03 April 2025
Abstract: The MeFER4 gene in cassava (Manihot esculenta Crantz) encodes a ferritin-like protein involved in stress responses, particularly under drought and oxidative stress. This study explores the role of MeFER4 in regulating oxidative stress in transgenic Arabidopsis. Overexpression of MeFER4 enhanced sensitivity to various stresses, including drought, oxidative, salt, and osmotic stress, with increased levels of reactive oxygen species (ROS) and altered expression of ROS-related genes in Arabidopsis. Furthermore, interactions between MeFER4 and antioxidant enzymes (APX1 and APX3), suggest its role in managing ROS homeostasis. These findings highlight MeFER4 as a key regulator of oxidative stress responses, offering the potential for improving stress tolerance in crops.
-
Key words:
- MeFER4 /
- Regulating /
- ROS homeostasis /
- cassava /
- Abiotic stress