
-
Floral transition is a critical developmental mechanism in plants that affects their viability and reproductive success. Multiple gene regulatory mechanisms are involved in the reproductive phase transition in plants. Transcription factors (TFs) are master regulators vital for flowering regulatory pathways. TFs directly bind to cis-elements in the promoter region, their specific target genes, or through protein-protein interactions[1]. Various TFs such as MCM1agamous-deficiens-serum response factor (MADS), WUSCHEL-related homeobox (WOX), TEOSINTE BRANCHED1/CYCLOIDEA/PCF (TCP), Auxin Response Factor (ARF), Ethylene Response Factor (ERF), v-myb myeloblastosis viral oncogene homolog (MYB), SQUAMOSA Promoter Binding Protein-like (SPL), basic Helix-Loop-Helix (bHLH), and Zinc Finger Protein (ZFP) have been identified in plants[2].
Zinc finger proteins are a class of TFs that regulate plant development. The zinc finger protein (ZFP) subfamily BBX (B-box gene family) contains one or two B-box conserved domains with specific tertiary structures that are stabilized by the binding of zinc ions[3]. The BBX gene family is highly conserved among plants. The BBX gene family has been identified in many plant species due to the availability of plant genomes. For example, 32 BBX genes have been identified in Arabidopsis[3,4].Thirty BBX genes in rice (Oryza sativa), 22 in tung tree (Vernicia fordii), 31 in tomato (Solanum lycopersicum), 23 in mung bean (Vigna radiata), 28 in alfalfa (Medicago sativa)[5−9], and 59 BBX genes identified in soybean (Glycine max)[10].
BBX genes participate in various developmental mechanisms, such as floral transition, hypocotyl elongation, shade avoidance, circadian rhythm, thermogenesis, and photomorphogenesis[11−14]. BBX genes act as integrative factors in flowers and regulate the expression of downstream genes or affect the activities of other proteins[3,15]. CO (CONSTANS)/AtBBX1 participates in photoperiod regulation of flowering time in plants[16]. In Arabidopsis, AtBBX28, and AtBBX29 promote flowering under long-day (LD) conditions[17]. AtBBX28 and AtBBX29 protein interacted with CO. CO protein is a central player that transcriptionally regulates FT (FLOWERING LOCUS T) and promotes early flowering in Arabidopsis[17]. The CO gene ortholog OsBBX18/OsHd1 (HEADING DATE 1) stimulates early floral induction in rice under short-day (SD) conditions. It regulates the Rice FT-like 1 (RFT1) and Hd3a (HEADING DATE 3a) genes[18]. Recently, significant progress has been made in BBX genes' significant role in flowering and flower development[3]. This complex regulatory mechanism demonstrates the multifaceted role of the BBX gene in plant development and floral regulation. This review summarizes the transcriptional regulatory role of BBX genes in photoperiod, vernalization, and GA (gibberellins) pathways.
-
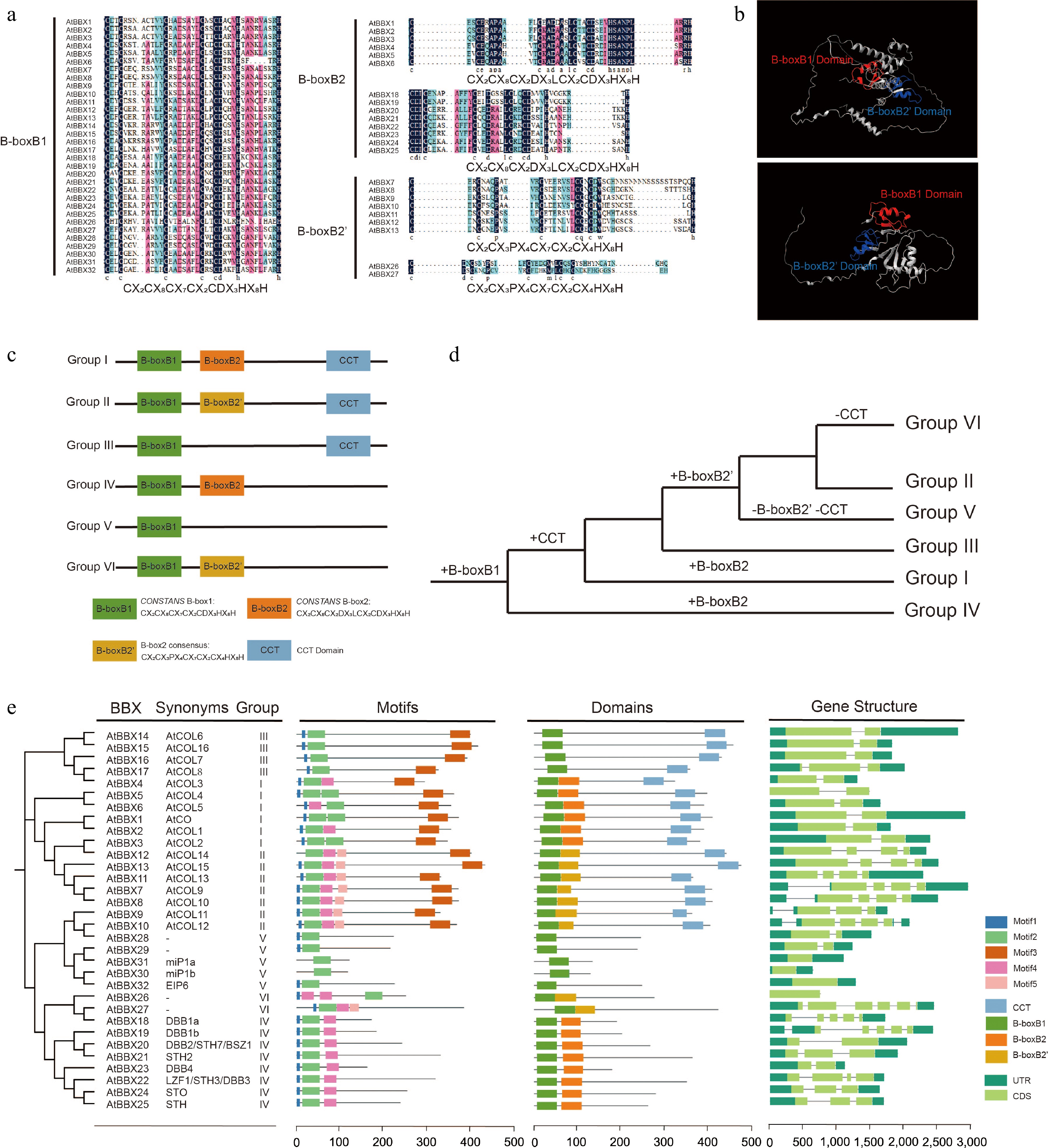
BBX proteins are characterized by the N-terminal inclusion of one or two B-box domains[3]. Arabidopsis BBX proteins are categorized into five groups based on the number of B-box domains and the presence or absence of the CCT (CONSTANS, CO-like, and TOC) domain[4,19]. Group I members are similar to group II in structure, and each member contains one B1(B-boxB1), one B2(B-boxB2/B-boxB2'), and one CCT domain. The B2 structure was slightly different between the group I and group II members (Fig. 1a). Members of group III contained one B1 and one CCT domain. Structural group IV contains one B1, one B2 domain, and no CCT domain. Group V consists of members with only one B1 domain[20]. AtBBX26 and AtBBX27 proteins were previously classified as group V in Arabidopsis, which only contains one B1 domain. In contrast, recent findings have shown that AtBBX26 and AtBBX27 contain one B1 and one B2(B-boxB2') domain, and we categorized them as structural group VI (Fig. 1b, c)[4,6]. Meanwhile, the B2 structures in groups VI and IV differ, the same as in groups I and II (Fig. 1a, c)[20].
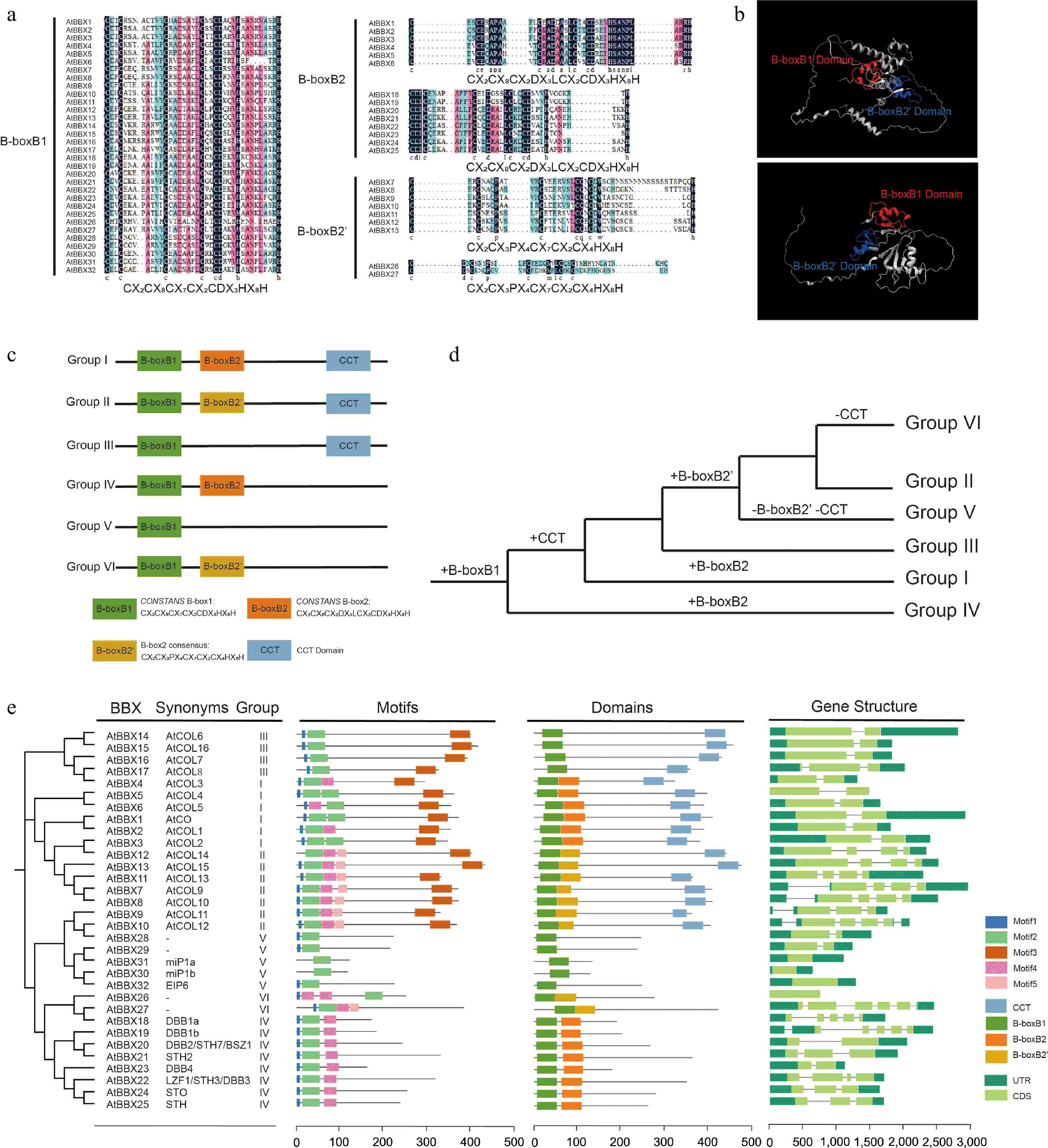
Figure 1.
Structural features of the AtBBX gene family. (a) Multiple sequence alignment analysis of AtBBX gene family member domains. B-boxB1 of groups I−VI, B-boxB2 of groups I and IV, and B-boxB2' of groups II and VI are indicated. The black color indicates 100% similarity. Red and blue indicate 75% and 50% similarity, respectively. (b) Tertiary structures of AtBBX26 and AtBBX27 (Group VI) proteins were retrieved from observations by Cheng & Wang[69]. Homology modeling of two proteins was constructed using the AlphaFold (
https://alphafold.ebi.ac.uk ) online server. Visualization with Chimera X. B-boxB1 and B-boxB2' domains are shown with red and blue colors, respectively. (c) Classification of AtBBX gene family structural groups (Groups I−VI). (d) Evolutionary hypothesis of BBX protein domains. (e) Characterization of Arabidopsis AtBBX genes. Neighbor-joining trees, synonyms, conserved motifs, domains, and exon-intron structures are indicated. '−' signifies that it is not available. Five color boxes indicate different motifs. Four color boxes indicate different domains. The dark green icon means UTR (untranslated region), the light green icon means CDS (coding sequence), and the line between UTR and CDS means intron (a non-coding part of a gene or mRNA molecule).The green plant ancestor (Viridiplantaee) possesses the protein containing two B-box domains that originated from branches of groups IV and I/II/III/V/VI (Fig. 1d)[6]. In addition, it was found that AtBBX26 and AtBBX27 lost their CCT domains during recent evolution, thus separating them from group II but retaining their structural group characteristics of B1 and B2[6]. OsBBX25 and OsBBX27 proteins in rice both contain one B1 and one CCT domain, and these two proteins evolved through internal deletion of the B2 domain, similar to the evolutionary pattern observed in Arabidopsis[5]. In this study, 32 AtBBX proteins were analyzed and found to consist of various sequence-specific binding sites, which may indicate their multifaceted role in plant development (Fig. 1e)[4].
-
Structural characterization of the BBX protein was based on its B-box and CCT domains (Figs 2, 3). Some members also have VP and PF (V/L) FL motifs, which play significant roles in transcriptional regulation and mediating protein-protein interactions[21]. In pear (Pyrus pyrifolia Nakai), PpBBX18 forms a heterodimer through two B-box domains with the bZIP domain at the C-terminal end of PpHY5 (ELONGATED HYPOCOTYL 5) protein in which PpHY5 binds to the G-box motifs of PpMYB10, and PpBBX18 provides the transactivation site, which activates the transcription of PpMYB10 and induces anthocyanin accumulation in the light[22]. In cabbage (Brassica rapa), the BrBBX32 protein N-terminus interacts with BrAGL24 (AGAMOUS-LIKE 24) and promotes flowering[23]. The B-box domain can participate in BBX protein interactions among different species. The B-box domain of AtBBX32 in soybean mediates its binding to GmBBX62. Deletion of the first 17 amino acids in the N-terminal B-box motif of AtBBX32 reduced its binding to GmBBX62 by 80%. However, deleting outside the B-box region does not affect its binding to GmBBX62[24]. Cysteine mutations in the N-terminal B-box domain of AtBBX28 significantly impair heterodimerization between AtBBX28 and CO[25]. Replacing the conserved Cys-25 in the Arabidopsis AtBBX19 Box1 motif with Ser eliminated the interaction of AtBBX19 with CO protein and the inhibitory effect of AtBBX19 on flowering. In contrast, the C76S replacement in the Box2 motif is not helpful[26].
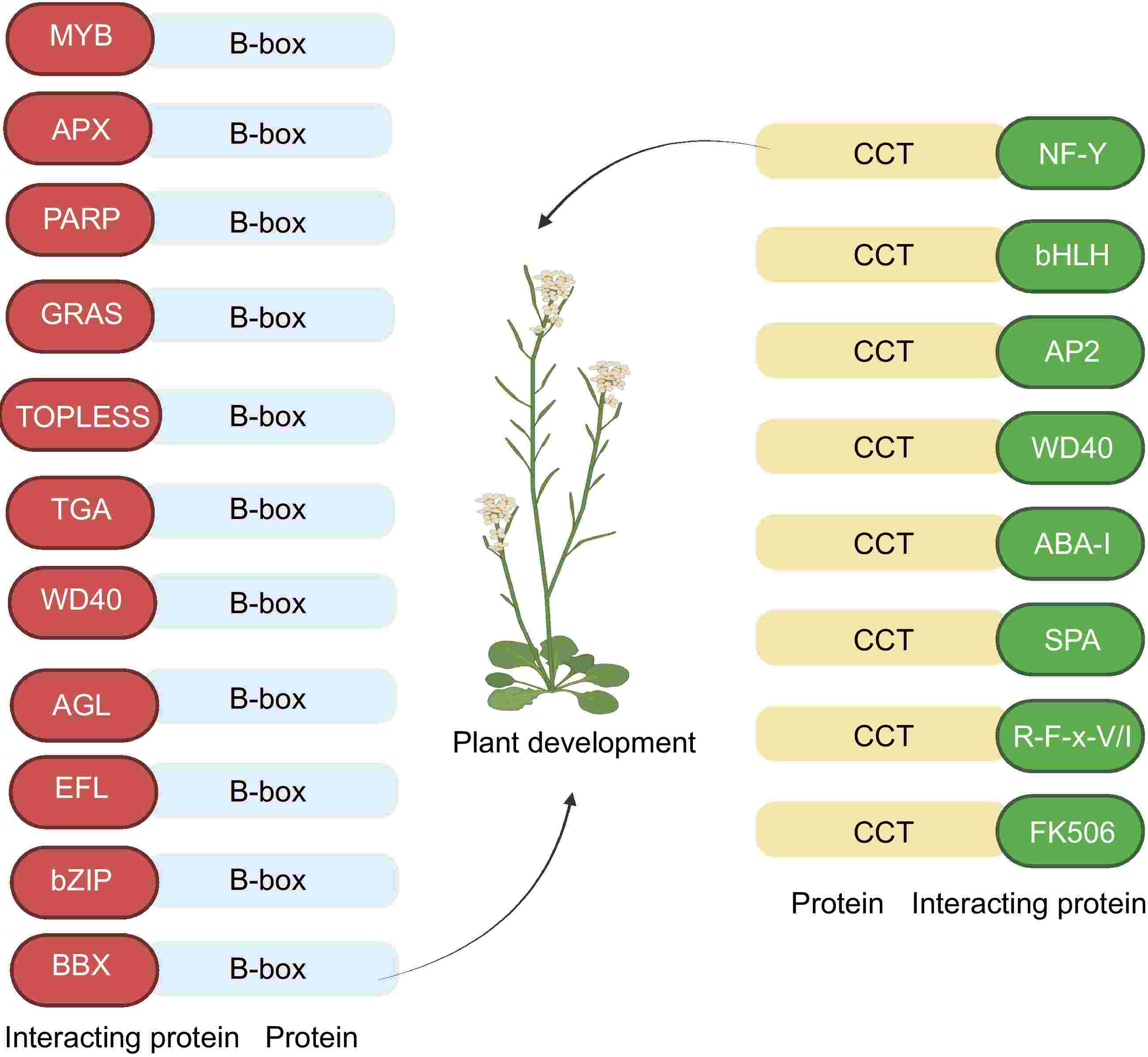
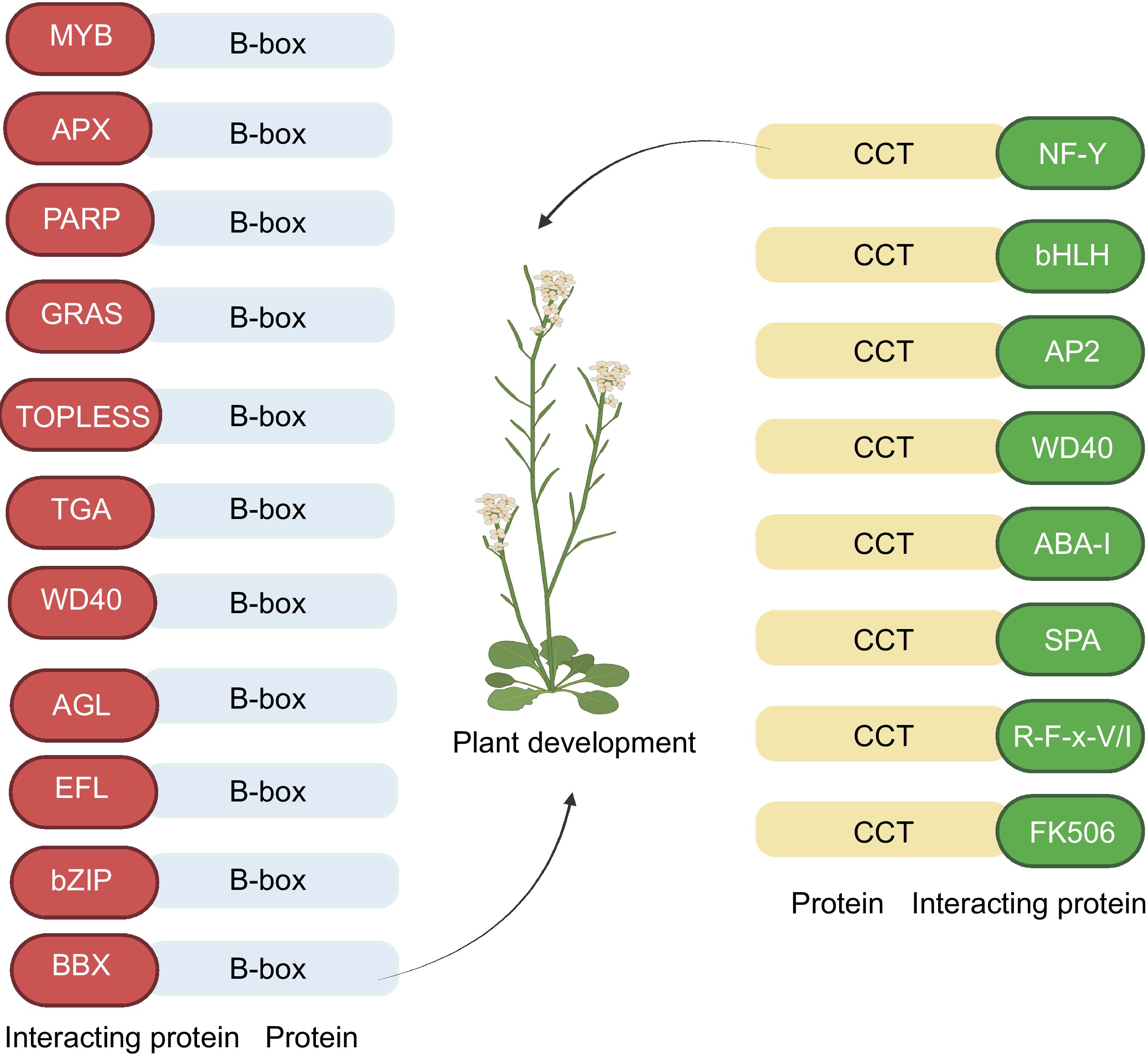
Figure 2.
Protein interactions of B-box and CCT domain in plants. Letters in the ellipses represent acronyms for the type of proteins interacting with the domain. B-box interacting proteins: BBX (AtBBX32-GmBBX62)[24], bZIP (PpBBX18-PpHY5)[13], EFL (AtBBX19-EFL3)[38], AGL (BrBBX32-BrAGL24)[23], WD40 (AtBBX19-COP1)[38], TGA (CO-TGA4)[39], TOPLESS (CO-TPL/TPR)[35], GRAS (AtBBX4-DELLA)[40], PARP (CmBBX8-CmRCD1)[41], APX (AtBBX18-APX1)[42] MYB (CO-AS1)[43]. Interacting proteins of CCT: FK506 (CO-FKBP12)[44], R-F-x-V/I (WNK-OSR1)[45], SPA (CO-SPA1)[34], ABA-I (CO-ABI3)[46], WD40 (CO-COP1)[47], AP2 (CO-TOE1/2)[48], bHLH (CO-CIB1)[49], NF-Y (CO-NF-YB/YC)[28].
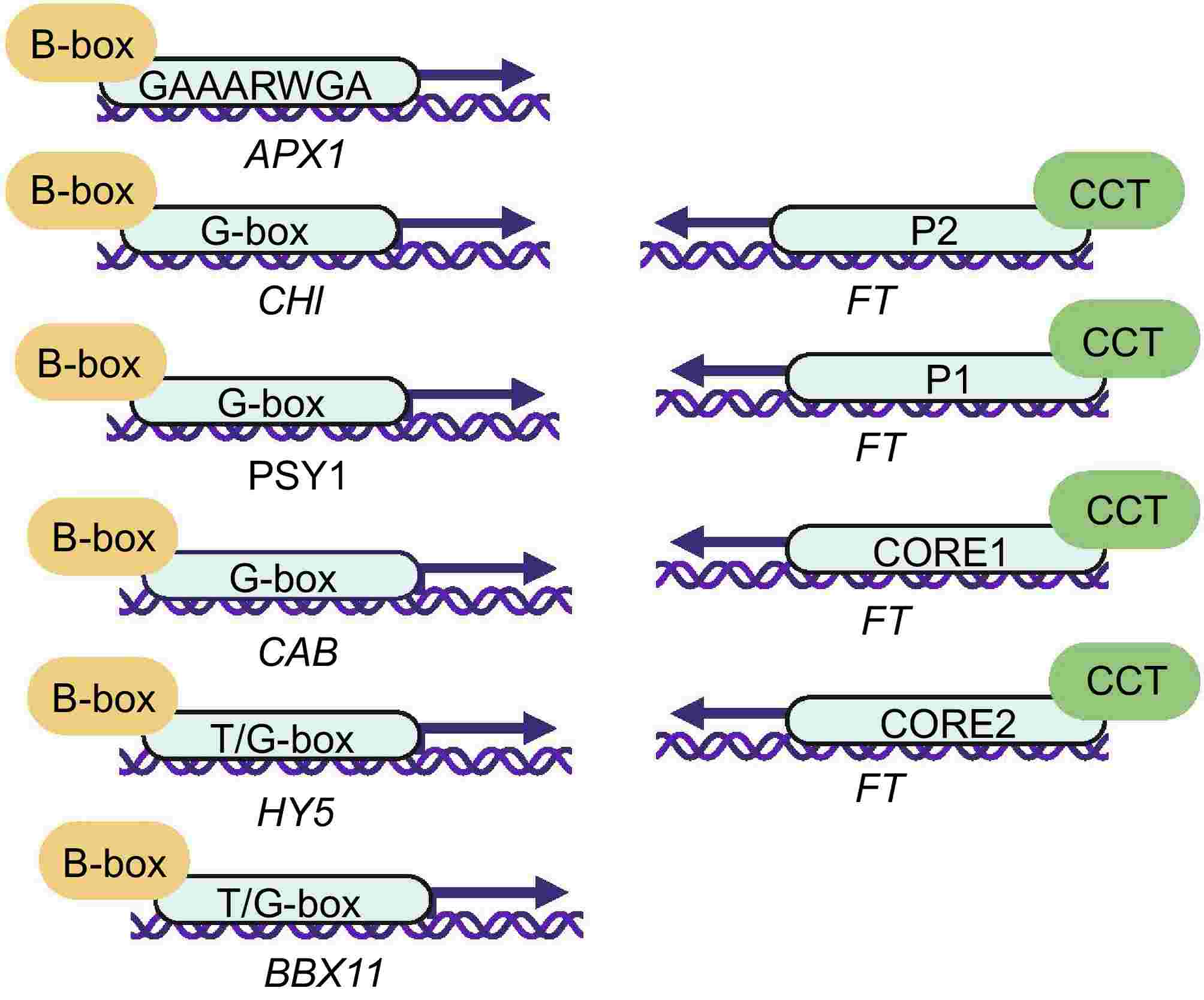
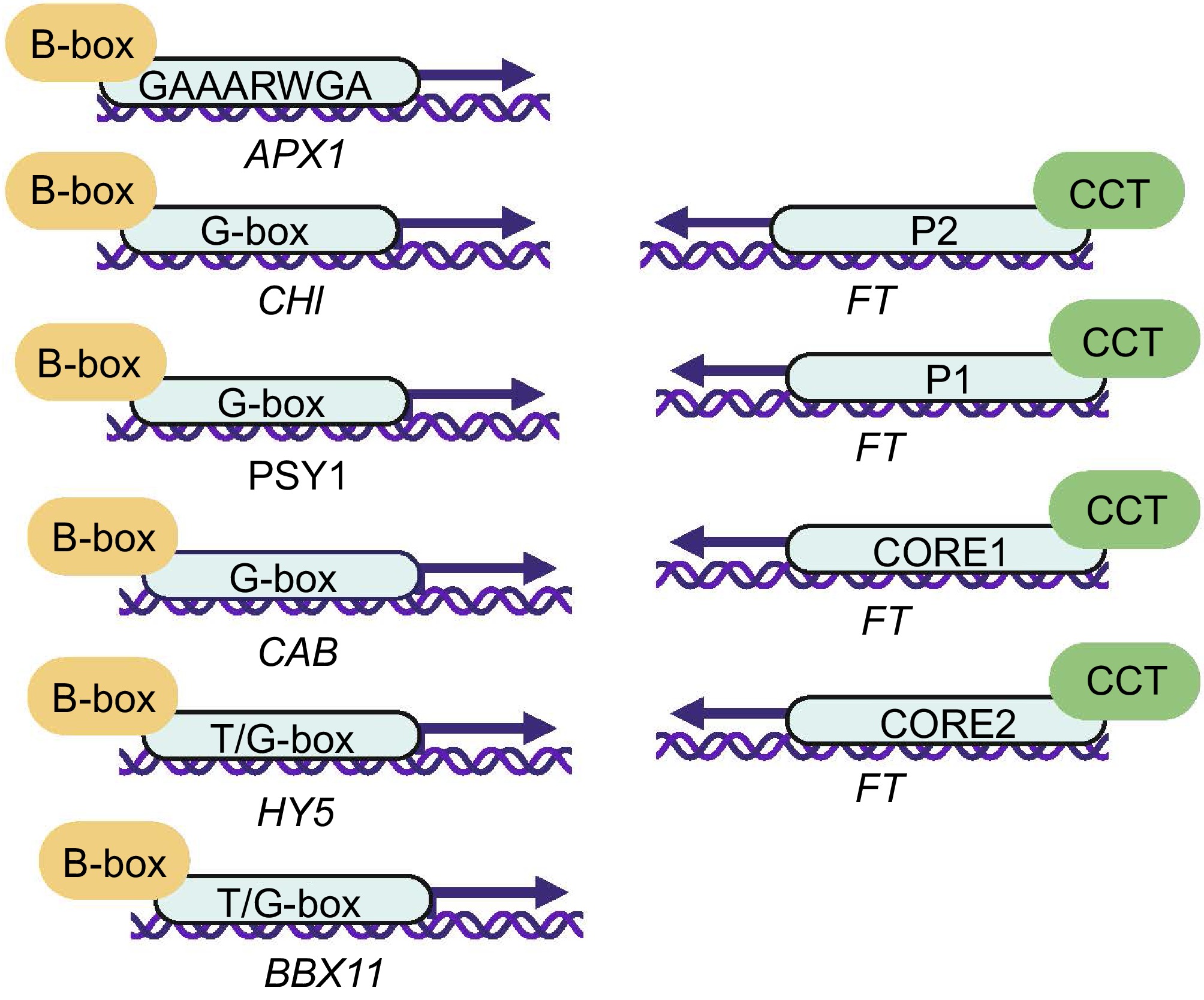
Figure 3.
Transcriptional regulation of B-box and CCT domain in plants. Letters in ellipses with arrows represent promoter binding site abbreviations. Transcriptional regulation of B-box: GAAARWGA (AtBBX18-APX1)[42], G-box (AtBBX21/22-CHI)[50], G-box (SlBBX20-PSY1)[51], G-box (AtBBX22-CAB)[50], T/G-box (AtBBX21-HY5)[52], T/G-box (AtBBX21-AtBBX11)[53]. Transcriptional regulation of CCT: P1/P2/CORE1/CORE2 (CO-FT)[29,54].
The CCT domain consists of 42−43 amino acid residues and is highly conserved among different plants[15]. Several BBX proteins with CCT domains have been reported to play essential roles in transcriptional regulation. The CCT domain of CO has been shown to mediate flowering in Arabidopsis by directly binding to the FT promoter and activating its expression[16]. In addition, the CCT domain of tomato TCOL1 and Arabidopsis CO proteins has been shown to approach target genes in DNA and regulate gene expression by interacting with the HAP5/NF-YC subunit of the DNA-binding protein CBF (C-REPEAT BINDING FACTOR). Mutations in the CCT domain of TCOL causes disruptions in its interaction with tomato THAP5 protein and similar mutations in CO also impair its function and delay flowering in Arabidopsis[16,27]. In subsequent studies, the B-box domain of CO could recruit the B-box domains of three additional CO proteins to form a homotetramer, and each of the four CO proteins, through their CCT domains, formed a complex with NF-YB/YC, which non-specifically recognized DNA along with the CCT domain to bind to the TGTG motif of the FT promoter, and the TGTG motif was shared by P1, P2, CORE1 and CORE2 elements (Figs 2, 3)[27−31].
The VP (Val-Pro) motif consists of six amino acids with a consistent G-I/VV-P-S/T-F sequence. It is located at the C-terminus of BBX proteins[3,21]. The VP motif was previously thought to be specific to group I. In contrast, VP also pairs at the C-terminus in individual members of groups III, IV, and V[15,21]. Sixteen−20 amino acids separate the VP motif from the CCT domain. They are essential for mediating the interactions of some BBX proteins with other proteins[21,32]. The replacement of the VP motif with alanine eliminated the interaction of STH (AtBBX25) and HY5 with COP1 (CONSTITUTIVELY PHOTOMORPHOGENIC 1) proteins[32]. The VP motif and WD40 domain are also crucial for interacting AtBBX4 with COP1 and regulating the degradation of the HY5 protein in plants[32,33]. In addition, the VP motif of CO is essential for its interaction with the SPA1 protein[34]. The PF(V/L)FL (Phe-Val/Leu-Phe-Leu) motif has also been reported to play a crucial role in some BBX proteins, which mediate the interaction of the AtBBX30/AtBBX31 (miP1a/b) microproteins with the TPL/TPR (TOPLESS/TOPLESS-RELATED) proteins as well as formation with the CO trimeric deterrent complex to delay floral transition[35,36].
CO proteins form homodimers with each other and heterodimerize with specific AtBBX proteins[37]. The interaction procedure may require the presence of at least three B-box domains between two proteins[37]. The CCT domain promotes their binding to the FT target site, thereby affecting Arabidopsis flowering[37]. In summary, B-box, and CCT domains are crucial functional domains in plant growth and development (Fig. 3).
-
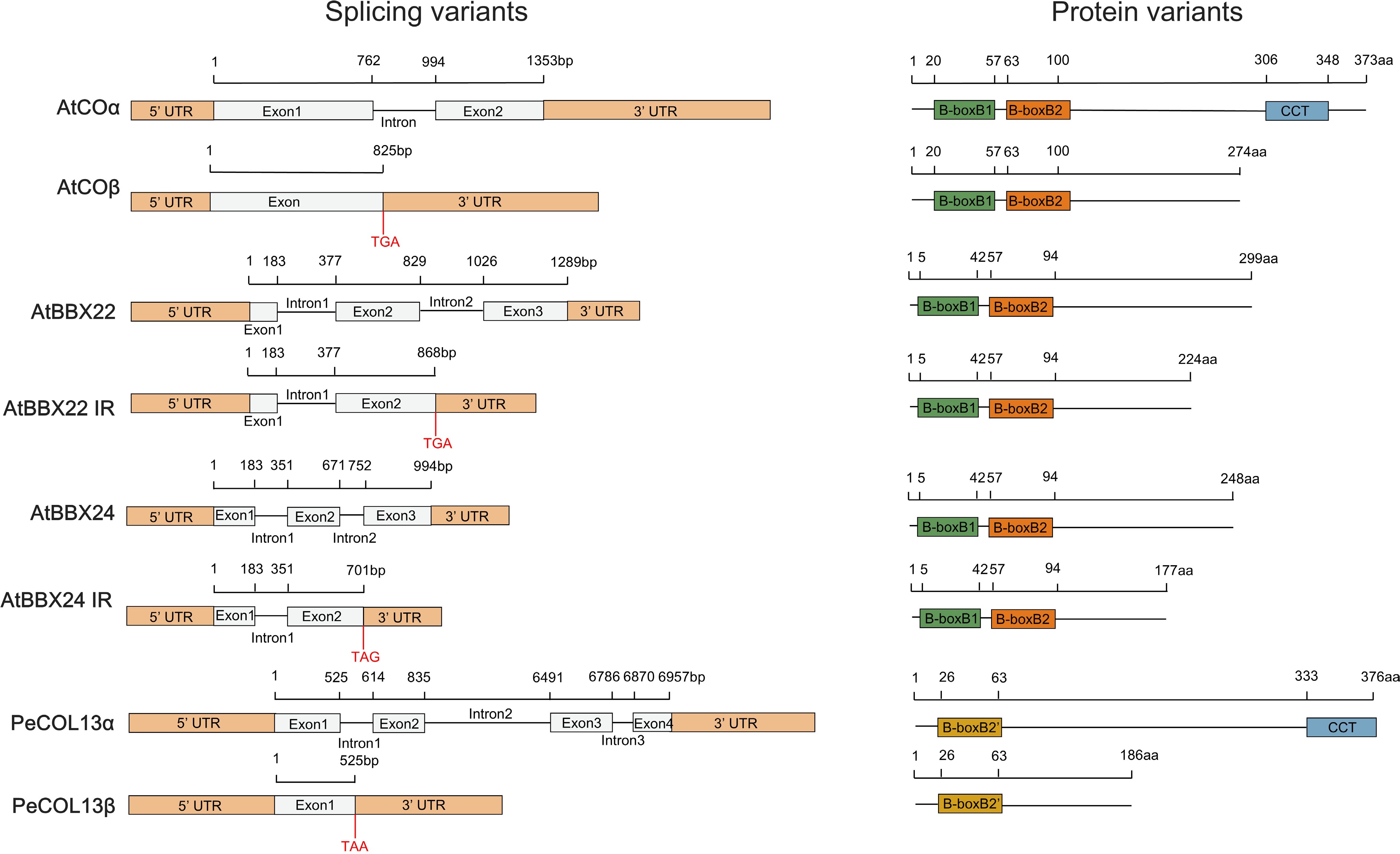
BBX genes can produce multiple transcripts through alternative splicing to regulate plant development[55−57]. The CO gene produces two RNA variants through intron-retentive (IR) splicing: full-length COα with intact functionality and truncated COβ with a C-terminal deletion of the CCT domain. COα gene overexpression induces early flowering, whereas COβ gene overexpression delays flowering. COβ gene reduces the binding of the COα to the FT promoter by interacting with COα protein, thereby inhibiting flowering in COα-COβ co-expression plants[55]. Further studies revealed that COβ protein does not interact with E3 ubiquitin ligases (COP1, HOS1, HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE1, FKS1, FLAVIN-BINDING KELCH REPEAT F-BOX 1), but regulates the interaction between COα and E3 ubiquitin ligases. This regulation destabilizes the COα protein and promotes degradation[55].
PeCOL13 gene(Phyllostachys edulis) produces full-length PeCOL13α and PeCOL13β lacks the CCT domain through IR splicing[56]. In contrast, the PeCOL13β protein diminishes PeCOL13α-repressed flowering by competing with PeCOL13α for binding to the PeFT promoter[56]. While the homologous protein AtCOL13 has two B-box domains (B-boxB1, B-boxB2'), PeCOL13 has only one B-boxB2' domain. It contains distinct amino acid residues from the second B-box domain, potentially owing to their opposing functions (Figs 1e, 4)[56]. IR RNA variant transgenic plants of AtBBX24 and AtBBX22 exhibited photosensitivity and acted as regulators of photomorphogenesis[57]. In summary, the complex splicing pattern of the BBX gene enriches its molecular functional diversity and provides fine regulatory mechanisms for plant development and environmental adaptation.
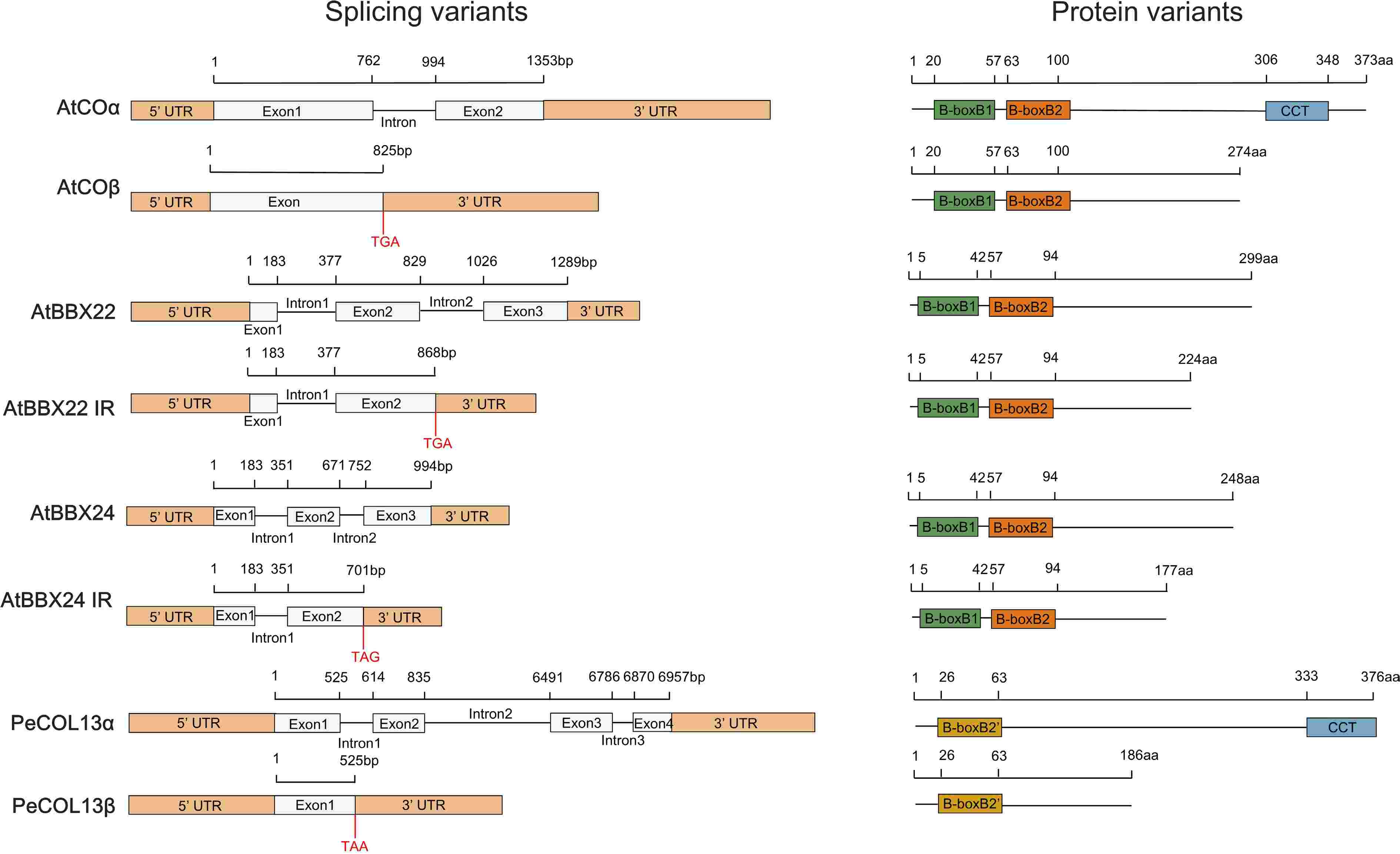
Figure 4.
Sequence structures of mRNA and protein isoforms of four BBX genes. The black horizontal line indicates the number of base pairs and amino acids. Yellow boxes indicate 5'/3' UTRs, grey boxes indicate exons, and black lines indicate excised introns. Four colour boxes indicate different domains.
-
Flowering is a complex biological process that involves multiple regulatory pathways, such as vernalization, photoperiod, autonomous, gibberellins (GA), and age-dependent pathways[58]. These regulatory pathways comprise a complex network of genes and transcription factors. Arabidopsis AtBBX genes play essential roles in floral regulation and development (Table 1, Fig. 5). However, AtBBX gene orthologues in other plant species are conserved in functions, such as CmBBX8 (Chrysanthemum morifolium), FaCO (Fragaria × ananassa), and BvBBX19 (Beta vulgaris) promote plant flowering. On the other hand, OsCOL4, GmCOL1a (Glycine max), and AcBBX5 (Ananas comosus) inhibited flowering. OsHd1 acts as a floral promoter and repressor (Table 2). BBX genes and their role in development are highly expressed in plant flower organs[59−61]. AcBBX5 was expressed explicitly in the stamens and petals of pineapple (Ananas comosus), and the transgenic Arabidopsis plant's petal and sepal widths were enlarged[59]. BBX genes regulate the miRNA-lncRNA-TF module and trigger floral organ development[62]. For example, lch-lnc3939-AtBBX19 and lch-lnc6617-AtBBX24 are involved in floral organ development[26,62,63]. OsBBX11 and OsBBX19 protein interaction regulates the expression of OsBTB97 (OsBBX11) and controls the growth of flower organs in rice[64].
Table 1. Functions of AtBBX genes in inducing floral transition in Arabidopsis.
Species BBX Gene ID Function Ref. Arabidopsis thaliana AtBBX1 Q39057 CO is involved in the photoperiodic regulation of flowering under LD conditions by binding to the FT promoter and activating its expression over a longer time;
Phytochrome B delays flowering in plants under SD conditions by directly reducing CO protein activity or indirectly antagonizing its effect on flowering time;
CO can inhibit FT induction of flowering by affecting TFL1 expression under SD conditions[16,65] AtBBX4 Q9SK53 Interaction of AtBBX32 with AtCOL3 under LD conditions enables AtCOL3 protein to bind the FT promoter and repress its transcription [66] AtBBX5 Q940T9 AtCOL4 interacts with CO proteins to inhibit flowering under LD and SD conditions in plants [67] AtBBX6 Q9FHH8 AtCOL5 overexpression plants flower early under SD conditions, and AtCOL5 mutant plants do not affect flowering [68] AtBBX7 Q9SSE5 AtCOL9 regulates flowering time under LD conditions by repressing CO expression while reducing FT expression and delaying the floral transition [69] AtBBX10 Q9LJ44 AtCOL12 physically interacts with CO in vivo to inhibit flowering under LD conditions by inhibiting CO protein function while reducing FT expression [70] AtBBX13 Q9C7E8 AtCOL15 interacts with CO protein and represses CO-mediated transcriptional activation of FT in LD conditions;
AtCOL15 can also compete with CO protein and directly bind to the CORE2 motif on the FT promoter to repress flowering in Arabidopsis[71] AtBBX14 Q8LG76 AtBBX14 interacts with CO in the nucleus and disrupts CO binding to the FT promoter, preventing the ability of CO to activate FT and inhibiting flowering under LD conditions [72] AtBBX15 Q8RWD0 AtBBX15 interacts with CO in the nucleus and disrupts CO binding to the FT promoter, preventing the ability of CO to activate FT and inhibiting flowering under LD conditions [72] AtBBX16 Q8RWD0 AtBBX16 interacts with CO in the nucleus and disrupts CO binding to the FT promoter, preventing the ability of CO to activate FT and inhibiting flowering under LD conditions [72] AtBBX17 Q9M9B3 Overexpression of AtCOL8 causes late flowering under LD conditions and AtBBX17 protein inhibits FT transcription by interacting with CO [73] AtBBX19 C0SVM5 AtBBX19 inhibits flowering by consuming CO and ultimately inhibiting FT as the primary pathway for regulating SOC1 [26] AtBBX24 Q96288 Overexpression of AtBBX24 accelerated flowering under LD and SD conditions, whereas mutant AtBBX24 delayed flowering only under SD conditions
AtBBX24 not only repressed FLC expression but also competed with FLC to regulate FT and SOC1 expression, thereby promoting flowering[63] AtBBX28 NP_194461 The interaction of AtBBX28 with CO reduced the binding of CO to FT promoter and inhibited flowering under LD conditions;
AtBBX28-AtBBX29 double mutant plants have reduced transcriptional activation activity of CO to FT promoter, and the interaction of AtBBX28 with CO reduces CO binding to the FT promoter and represses flowering under LD and low-temperature conditions[17,25] AtBBX29 NP_200258 AtBBX28-AtBBX29 double mutant plants have reduced transcriptional activation activity of CO to FT promoter, and the interaction of AtBBX29 with CO reduces CO binding to the FT promoter and represses flowering under LD and low-temperature conditions [17] AtBBX30 Q1G3I2 The miP1b interacts with TPL and forms a trimeric deterrent complex with CO to delay floral transition under LD conditions [35,36] AtBBX31 Q9LRM4 The miP1a interacts with TPL and forms a trimeric deterrent complex with CO to delay floral transition under LD conditions [35,36] AtBBX32 Q9LJB7 Interaction of AtBBX32 with COL3 under LD conditions enables AtCOL3 to bind the FT promoter and repress its transcription [66] 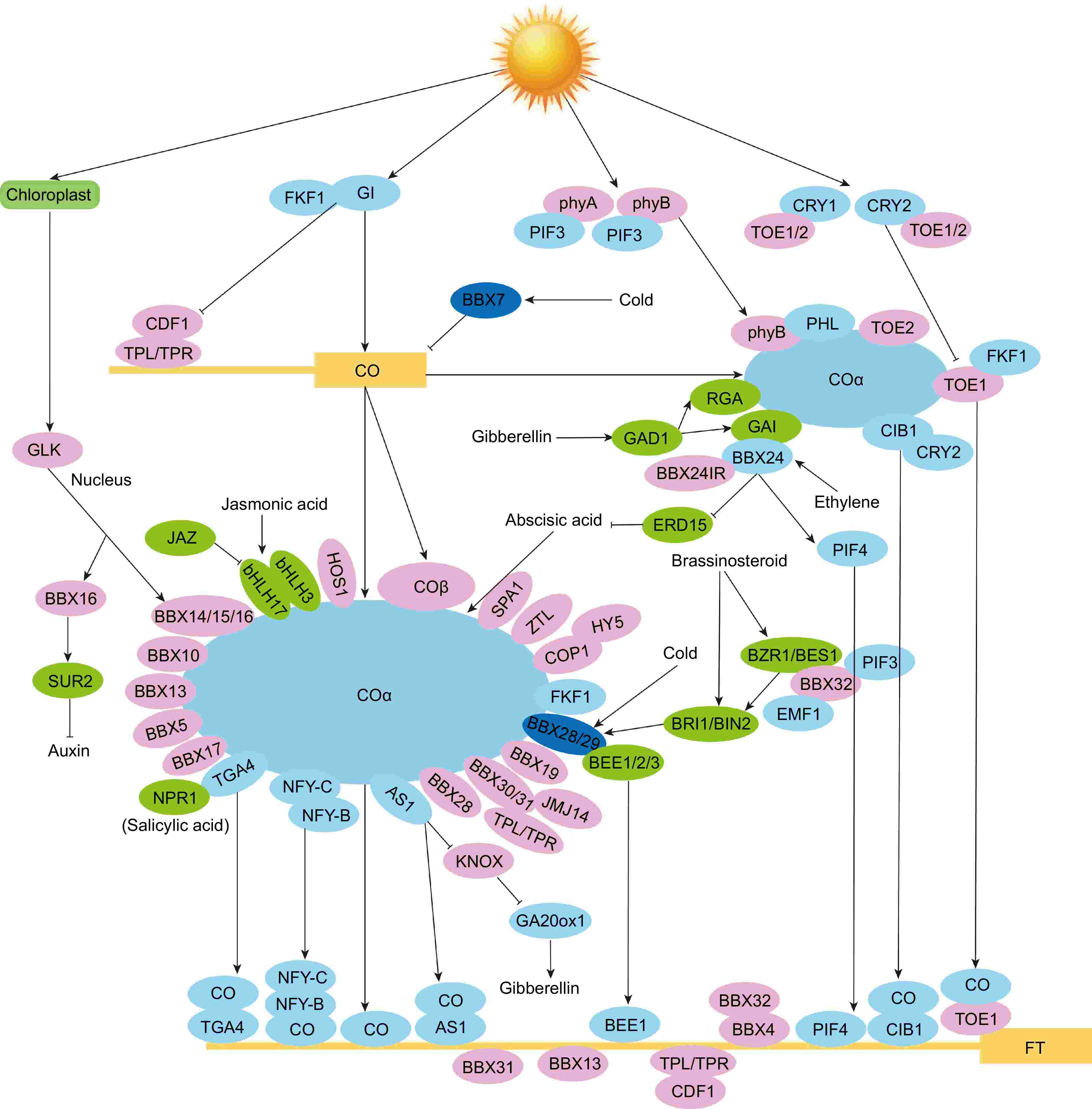
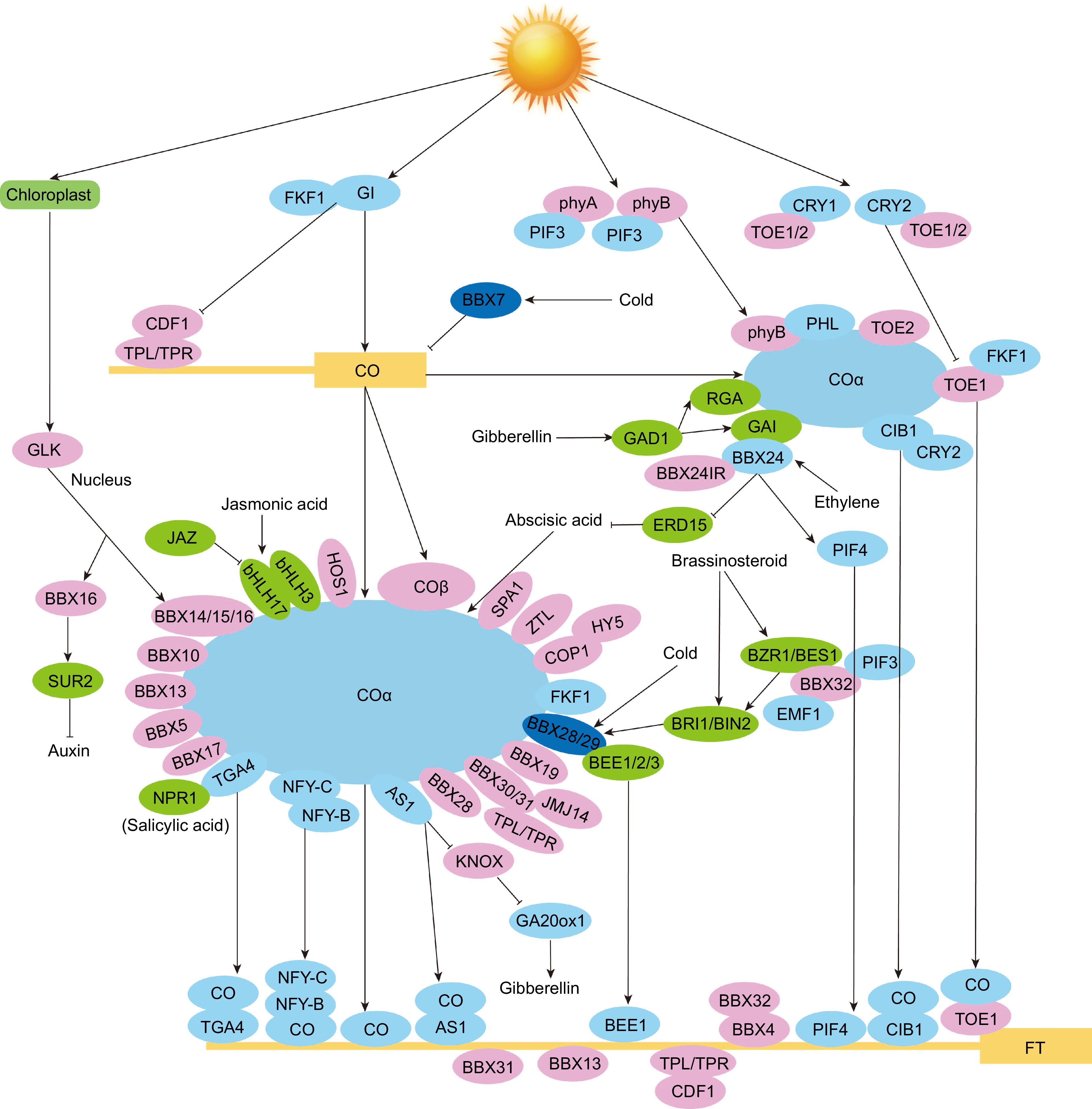
Figure 5.
Mechanism of AtBBX genes in reproductive phase transition in Arabidopsis. The letters in the boxes represent short names of genes, and the letters in the ellipses represent short names of proteins. Positive regulators are shown as light blue circles. Negative regulators are in pink, and hormone-responsive proteins are in green. Cold-responsive proteins are shown in dark blue. Solid lines with arrows represent facilitation, and solid lines with flat ends represent inhibition[1,3,12,14,15,21,30,31,40,48,49,58,63,113−118].
Table 2. Functions of BBX genes in inducing floral transition in different plant species.
Species BBX Gene ID Function Ref. Oryza sativa OsCOL4 BAS79707 OsCOL4 inhibits rice flowering under SD and LD conditions [74] OsCOL9 BAS84196 OsCOL9 suppresses Ehd1 expression and inhibits rice flowering under SD and LD conditions [75] OsCOL10 BAS86019 OsCOL10 is a flowering repressor linking Ghd7 and Ehd1 in rice and inhibits flowering in rice under SD and LD conditions [76] OsBBX14 BAS92741 OsBBX14 acts as a floral repressor by promoting OsHd1 expression under LD conditions;
OsBBX14 delayed flowering under SD conditions by acting as a repressor of Ehd1[77] OsCCT19 XP_015642185 Transgenic plants of OsCCT19 with delayed tasseling in LD conditions [78] OsCOL16 BAS97134 OsCOL16 up-regulated the expression of Ghd7, which in turn down-regulated the expression of Ehd1, Hd3a, and RFT1, leading to late tasseling under both LD and SD conditions [79] OsCOL13 BAT03112 OsCOL13 functions as a negative regulator downstream of OsphyB and upstream of Ehd1 in rice, resulting in late tasseling under LD and SD conditions [80] OsHd1 BAS97223 OsHd1 acted as a promoter of tasseling under SD conditions and as a suppressor of tasseling under LD conditions [81] OsCOL15 BAT06449 OsCOL15 inhibited flowering under LD and SD conditions by up-regulating Ghd7 and down-regulating RID1 expression, thereby down-regulating Ehd1, Hd3a, and RFT expression [82] OsCO3 BAT06983 OsCO3 inhibits Hd3a and FTL expression to delay flowering under SD conditions [83] OsDTH2 BAS97360 OsDTH2 is independent of OsHd1 and Ehd1 and directly represses the expression of Hd3a and RFT1 to induce rice tasseling under LD conditions [84] Chrysanthemum
morifoliumCmBBX7 AMO42717 CmBBX7 and CmBBX8 interact with each other to positively regulate CmFTL1 expression by binding to its promoter to accelerate flowering under LD conditions [85] CmBBX5 − CmBBX5 interacts with CmBBX8 to inhibit CmFTL1 regulation of chrysanthemum flowering under LD and SD conditions [86] CmBBX8 AMO42713 CmBBX8 protein accelerates plant flowering under LD conditions by directly targeting CmFTL1 by interacting with CmERF3 or CmRCD1;
CmBBX8 and CmBBX7 interact with each other to positively regulate CmFTL1 expression by binding to its promoter to accelerate flowering under LD conditions[41,85,87] CmBBX13 KP963935 CmBBX13 delays flowering in plants under SD and LD conditions in a photoperiod-independent pathway [88] CmBBX24 KF385866 CmBBX24 inhibited flowering by affecting the photoperiod and GA pathways, and under LD conditions, CmBBX24 regulated flowering mainly by affecting the GA pathway [89] Fragaria × ananassa FaBBXx28c1 QOI16737 Overexpression of FaBBX28c1 showed a late-flowering phenomenon in LD conditions [90] FvCO WBW02120 Overexpression of FvCO plants under LD resulted in slightly early flowering, whereas overexpression under SD induced early flowering [91] Glycine max GmCOL1a Glyma08g28370 Overexpression of GmCOL1a leads to plants flowering late under LD conditions [92] Beta vulgaris BvBBX19 XP_019107108 BvBBX19 and BvBTC1 interact to form a heterodimer and bind the BvFT2 promoter to activate flowering under LD conditions [93,94] BvBTC1 BBH85249 BvBTC1 and BvBBX19 interact to form a heterodimer and bind the BvFT2 promoter to activate flowering under LD conditions [93,94] BvCOL1 ACC95129 Transgenic BvCOL1 plants compensate for the late-flowering phenotype of Athaliana co-2 mutants with a positional/gene-dose effect [95] Hordeum vulgare HvCO1 AF490467 HvCO1 activates HvFT1 to induce flowering under LD and SD conditions [96] HvCO9 AY082965 Overexpression of HvCO9 plants flowering late in LD and SD conditions [97] HvCO2 XP_044950638 Overexpression of HvCO2 promotes flowering induction by Ppd-H1 and HvFT1 expression in LD and SD conditions [98] Vitis vinifera VvCO XP_059589686 VvCO expression is associated with seasonal flowering induction in latent buds [99] Sorghum bicolor SbCO EER88227 SbCO promotes flowering by inducing SbEhd1 and SbFTL genes under LD conditions [100] Zea mays ZmCONZ1 ABW82153 ZmConz1 activates ZmZCN8, which acts as a floral inducer involved in photoperiod sensitivity in maize [101] Ananas comosus AcBBX5 XP_020107577 AcBBX5 protein binds the AcFT promoter and reduces its expression, thereby delaying flowering under LD conditions [59] Rosa rugosa RcCO RcChr2g0164091 Interaction between RcCO and RcCOL4 promotes the binding of RcCO protein to the CORE motif in the RcFT promoter and induces RcFT, accelerating flowering under LD and SD conditions [102] RcCOL4 RcChr6g0299051 Interaction between RcCOL4 and RcCO promotes the binding of RcCO protein to the CORE motif in the RcFT promoter and induces RcFT, accelerating flowering under LD and SD conditions [102] Mangifera indica MiCOL2A WED40957 Overexpression of MiCOL2A in Arabidopsis delays flowering of transgenic plants under LD and SD conditions [103] MiCOL2B WED40958 Overexpression of MiCOL2B in Arabidopsis delays flowering of transgenic plants under LD and SD conditions [103] MiCOL6 WED40966 MiCOL6 promotes early flowering in transgenic plants under LD and SD conditions [104] MiCOL7A WED40967 MiCOL7A inhibits flowering under LD and SD conditions by reducing AtFT and AtSOC1 expression [104] MiCOL7B WED40968 MiCOL7B inhibits flowering under LD and SD conditions by reducing AtFT and AtSOC1 expression [104] Solanum lycopersicum SlBBX4 Solyc08g006530 The SlBBX4 mutant showed delayed flowering in both LD and SD conditions [105] '−' signifies that it is not available. Role of CO in the photoperiodic flowering
-
CO was the first BBX protein identified in plants[16] and plays a central role in photoperiod-mediated flowering. The FT gene receives signals from CO under LD conditions to induce the transition from vegetative to reproductive growth in Arabidopsis. In contrast, it did not affect CO under SD conditions. However, the CO gene can suppress FT-induced flowering under SD conditions by promoting the expression of the FT repressor gene TFL1 (TERMINAL FLOWER1)[16,28,65]. In particular, TFL1 interacts with FD (FLOWERING LOCUS D) protein to negatively regulate FD-dependent target gene transcription and form the TFL1-FD complex to delay flowering. FD gene is vital to the FD-FT protein complex, promoting Arabidopsis flowering[106]. Most AtBBX proteins play a role in flowering and are regulated under the photoperiodic pathway by interacting with CO (Fig. 5). For example, AtBBX14, AtBBX15, AtBBX16, and AtBBX28 interact with CO protein in the nucleus to inhibit flowering by preventing the ability of CO to activate FT and reducing FT expression[25,72].
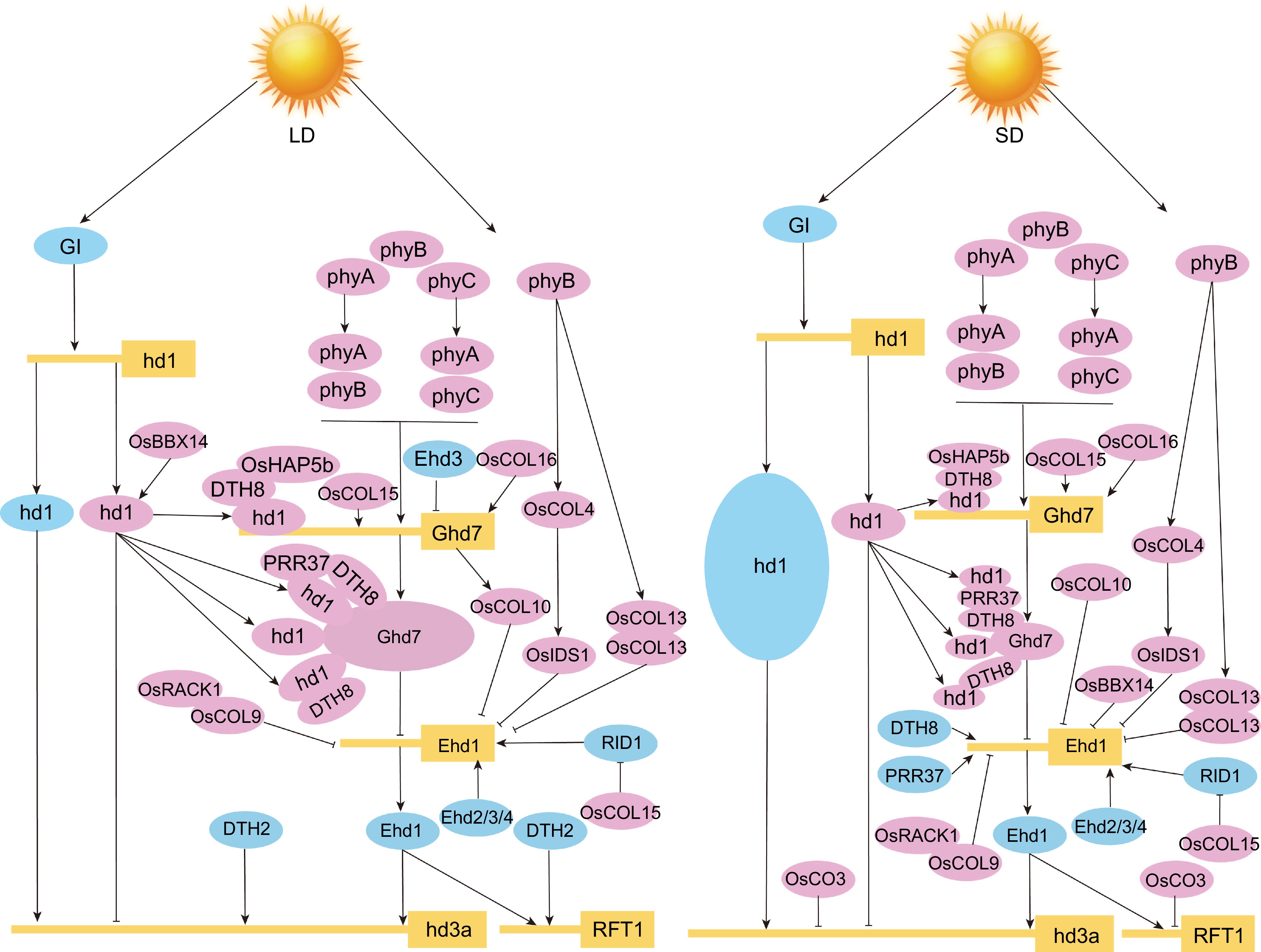
Interestingly, the interaction between AtBBX28 and CO did not affect the transcriptional activation activity of CO protein[25]. Similar to Arabidopsis, one rice CO homolog, OsHd1, promotes flowering under SD conditions and represses flowering under LD conditions[81]. OsHd1 regulation of rice tasseling involves only the enhancement and reduction in the expression of Hd3a (HEADING DATE 3A) and RFT1 (RICE FLOWERING LOCUS T 1), homologs of the FT gene[18]. Recent studies have identified some OsBBX genes that affect rice tasseling through the photoperiodic pathway (Fig. 6). Among these, OsBBX5 (OsCOL4), OsBBX7 (OsCOL9), OsBBX10 (OsCOL10), and OsBBX23 (OsCOL13) inhibit flowering by decreasing the expression of the rice RFT1 gene through Ehd1 (EARLY IN SHORT DAYS 1)[69,74,76,80].
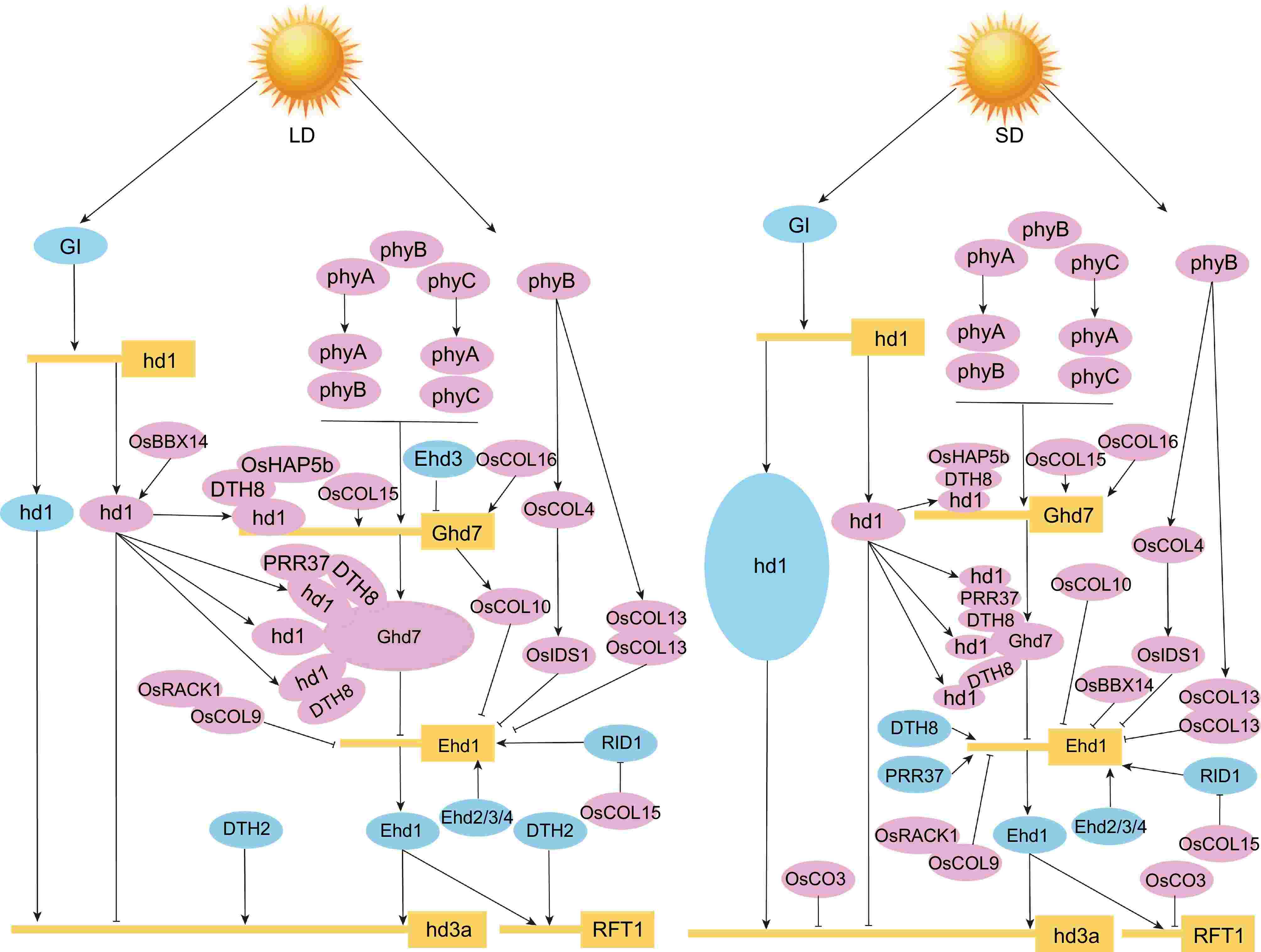
Figure 6.
Rice OsBBX genes regulatory network during photoperiodic flowering. The left plot represents LD conditions, and the right plot represents SD conditions. The letters in the boxes represent short names of genes, and the letters in the ellipses represent short names of proteins. Positive regulators are shown in light blue circles, and negative regulators are shown in pink. Solid lines with arrows represent facilitation, and solid lines with flat ends represent inhibition[74−77,79,80,82−84,119−122].
In addition, the OsBBX10 gene functions downstream of Ghd7 (GRAIN NUMBER, PLANT HEIGHT, and HEADING DATE 7) to delay rice tasseling by reducing the expression of Ehd1[76,82]. Furthermore, Ghd7 expression is regulated by SE5 (PHOTOPERIOD SENSITIVITY 5)[76]. Interestingly, Ghd7 and Ehd1 in rice do not exist as homologous genes in Arabidopsis, and their presence largely changes the GI-CO-FT-mediated photoperiodic pathway in rice[107]. Ghd7 could form multimers with Oshd1, Oshd1-DTH8 (DAYS TO HEADING 8), and Oshd1-DTH8-PRR37 (PSEUDO-RESPONSE REGULATOR 37) protein complex, respectively, and changes their activities to inhibit flowering under LD conditions[108]. Ghd7 gene expression is repressed under SD conditions, resulting in the release of the Oshd1 gene to achieve Oshd1-Hd3a-mediated early flowering[108]. The Ehd1 gene is similarly highly expressed and regulated by multiple BBX genes to achieve early flowering[108]. In addition to rice, CO homologs are also found in other plants, such as SbCO, ZmCONZ1, and HvCO1, which can affect photomorphogenesis to regulate plant flowering through different pathways[96,100,109]. Three members of the sorghum (Sorghum bicolor L.) PEBP (PHOSPHATIDYLETHANOLAMINE-BINDING PROTEIN FAMILY) family, SbCN8, SbCN12, and SbCN15, have been shown to affect flowering through the classical GI-CO-FT regulatory pathway[18,100,101]. The SbCO gene can indirectly activate SbCN8 and SbCN12 to induce flowering by promoting SbEhd1 expression under SD conditions[100]. In addition, SbEhd1 expression is suppressed by SbPRR37 and SbGhd7 under LD conditions[100].
AtBBX32 inhibits the expression of EMF1 (EMBRYONIC FLOWER 1) by interacting with it through the N-terminal B-box domain and delays floral formation in Arabidopsis[110]. Under SD conditions, AtBBX6 can lead to early flowering of plants by promoting the expression of FT and SOC1 (SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1), and AtBBX6-overexpression plants in CO mutants can inhibit the later-flowering effect to a certain extent. In contrast, the time of floral formation was not altered in AtBBX6-deficient mutants, and the molecular mechanism associated with CO has not been clarified[68]. Cotton (Gossypium hirsutum) GhBBX5, GhBBX8, GhBBX23, and GhBBX26 interact with GhFT to regulate cotton flowering[61]. Thus, some BBX proteins can act on flower genes to regulate plant flowering directly.
Many BBX proteins play a role in flowering by affecting the COP1-HY5 module[13]. The ubiquitin ligase COP1/SPA (CONSTITUTIVELY PHOTOMORPHOGENIC 1/SUPPRESSOR OF PHYA-105) in Arabidopsis mediates its degradation under dark conditions by interacting with CO and AtCOL12, which is hypothesized to compete with CO for the binding site of COP1, thus indirectly affecting plant flowering[70,111]. Similar to AtCOL12, AtBBX28 functions in COP1-AtBBX28-HY5 and AtBBX28-CO-FT by interacting with proteins[25,111,112]. AtBBX28 interacts with COP1 and mediates its degradation in the dark through the 26S proteasome, whereas AtBBX28 also interacts with HY5 in a dose-dependent manner, negatively regulating HY5 expression[112]. AtBBX4, AtBBX30, EIP6, AtBBX32, and OsBBX14 proteins participate in floral regulation by affecting the COP1-HY5 and CO-FT modules[13,33,77].
Role of BBX genes in hormone signaling pathway
-
The BBX genes play an essential role in the GA pathway during floral regulation (Fig. 5). CmBBX24 negatively regulates GA biosynthesis by affecting the regular expression of GA biosynthesis genes GA20ox (GA 20-OXIDASE) and GA3ox (GA 3-OXIDASE) and inhibits flowering. It also acts as a flowering repressor by reducing the expression of GI (GIGANTEA), PRR5, CO, FT, FTL, and SOC1 genes in the photoperiodic pathway[89]. The expression pattern of CmBBX24 in the overexpression plants was consistent with that of the florigen-resistant gene CmAFT (ANTI-FLORIGEN TRANSPORTER). In contrast, the expression levels of CmFTL3 and CmSOC1 were significantly increased by silencing CmBBX24[89]. CmBBX24 transgenic plants showed suppression of CmBBX24 expression after treatment with GA4/7. GA may negatively inhibit the regulatory process of CmBBX24 in chrysanthemum flower formation[89]. AtBBX24 can compete with FLC (Flowering Locus C) to regulate FT and SOC1 expression, promote flowering in Arabidopsis, and interact with GAI (GIBBERELLIN INSENSITIVE) protein in the gibberellin pathway[40]. CO protein could also interact with GAI, RGL1 (RGA-LIKE1), and RGA (REPRESSOR OF GA1-3), and RGA inhibited CO interaction with NF-YB2 in vitro[116].
BBX genes also participate in other plant growth regulators, such as ethylene (ETH) and abscisic acid (ABA), promoting flowering. The CmBBX8 gene affects flowering time in response to ETH signaling, and further studies found that CmBBX8 reduces the expression of CmFTL1 by interacting with CmERF3 (ETHYLENE RESPONSIVE TRANSCRIPTION FACTOR) or CmRCD1 (RADICAL-INDUCED CELL DEATH 1) protein[41,123]. CmBBX5 interacts with CmBBX8 protein to inhibit the regulation of CmFTL1 in chrysanthemum flowering under LD and SD conditions via a mechanism different from CmERF3[86]. Similarly, cabbage BcERF070 repressed the expression of downstream BcFT genes by directly binding to the promoter of BcBBX29, resulting in the inhibition of flowering under ETH conditions[87].
The synthesis and accumulation of ABA can induce the transcriptional activation activity of GI and CO, affecting the expression of their downstream FT genes to promote floral transition[124]. CO proteins also interact with TGA4 (TGACG MOTIF-BINDING FACTOR 4), which not only binds to NPR1 (NON-EXPRESSOR OF PR1) to regulate the expression of defense genes mediated by salicylic acid (SA) signaling but also directly binds to the FT promoter and regulates its expression to control the flowering time of the plant[39,125]. In summary, BBX proteins are essential in multiple hormone signaling pathways that affect plant flowering. Thus, BBX genes play a vital role in many aspects of the hormone pathway that regulate plant flowering through transcriptional and post-transcriptional regulation.
BBX genes and vernalization signaling pathway
-
Some BBX proteins act as regulators of low-temperature signaling and participate in the vernalization pathway to control plant floral formation. There is an interaction between beet BvBBX19 and BTC1, and the two BBX proteins interact in vivo to form a heterodimer that acquires a CO-like function and binds to the BvFT2 promoter to activate twitching and F2 plants with mutations in both genes do not twitch even after vernalization[93,94]. BrBBX32 in Chinese cabbage regulates flowering time by interacting with BrAGL24 protein, which governs the interaction between BrAGL24 and BrSOC2. In contrast, Arabidopsis AtAGL24 and SOC1 upregulate the expression of each other and mediate vernalization to promote flowering in an FLC-independent manner[23].
AtBBX28 nor AtBBX29 single mutants possess flowering-related phenotypes, whereas AtBBX28 and AtBBX29 double mutant plants exhibit significantly delayed flowering phenotypes at low temperatures (16 °C)[17]. Low temperature reduced the expression of FT, CO, and TSF (TWIN SISTER OF FT), which may have resulted from the mutation of AtBBX28 and AtBBX29, affecting the transcriptional activation activity of CO on the FT promoter. In contrast, a delayed flowering phenotype appeared in AtBBX28 overexpression plants, which may be related to AtBBX28's response to temperature changes[25]. The cold-induced expression of AtBBX7 and AtBBX8 is positively regulated upstream by CRY2 (CRYPTOCHROME 2) and HY5 but suppressed by COP1. In addition, AtBBX7 and AtBBX8 positively regulate cold resistance by regulating the expression of downstream COR (Cold-Responsive) genes[126]. OsBBX7, OsBBX17, and CmBBX24 have also been shown to be highly expressed under low-temperature conditions while affecting the flowering time of plants. However, a specific association has not been clarified[60,89]. These studies have confirmed the central role of BBX genes in floral induction and floral development in plants.
-
BBX genes are essential regulators of reproductive phase transition in plants. BBX genes trigger the transcriptional activity of downstream target genes and BBX proteins interact with other TFs and regulate flowering regulatory pathways, such as photoperiodic, GA, and vernalization pathways (Figs 5, 6). These regulatory mechanisms interact with each other to synergize and regulate the plant flowering process to ensure that the plants flower well and adaptively. This study explored the interactions, domain specificity, and transcriptional regulation mechanisms between BBX proteins and other regulatory factors to reveal their delicate regulatory mechanisms and potential application values in optimizing floral regulation in plants. Flowering is directly linked to fruit and commercial production. An in-depth investigation of the function of BBX genes in the flowering transition of plants will provide a basis for understanding flowering to meet the needs of agricultural production and obtain desirable agronomic traits.
The project was funded by the National Natural Science Fund of China (32160687, 32360723), the National Key R&D Program of China (2023YFD2300804), the Natural Science Foundation of Hainan Province (322MS013), Hainan graduate innovative research project (Qhyb2022-55) and the Priming Scientific Research Foundation of Hainan University (KYQD(ZR)-20090).
-
The authors confirm contribution to the paper as follows: writing—original draft preparation: Xu Z; data curation and software: Xu Z, Wang C; writing—review and editing: Khan FS; supervision and conceptualization: Khan FS, Zhang H; resources and investigation: Khan FS, Ouyang Y; funding acquisition: Zhang H. All authors have read and agreed to the published version of the manuscript.
-
The data are available on request from the corresponding author.
-
The authors declare that they have no conflict of interest.
-
Received 5 November 2024; Accepted 17 February 2025; Published online 3 April 2025
-
# Authors contributed equally: Ziqiang Xu, Faiza Shafique Khan
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu Z, Khan FS, Ouyang Y, Wang C, Zhang H. 2025. BBX proteins and their multifaceted roles in floral regulation. Tropical Plants 4: e014 doi: 10.48130/tp-0025-0011 

Figure 1.
Structural features of the AtBBX gene family. (a) Multiple sequence alignment analysis of AtBBX gene family member domains. B-boxB1 of groups I−VI, B-boxB2 of groups I and IV, and B-boxB2' of groups II and VI are indicated. The black color indicates 100% similarity. Red and blue indicate 75% and 50% similarity, respectively. (b) Tertiary structures of AtBBX26 and AtBBX27 (Group VI) proteins were retrieved from observations by Cheng & Wang[69]. Homology modeling of two proteins was constructed using the AlphaFold (https://alphafold.ebi.ac.uk) online server. Visualization with Chimera X. B-boxB1 and B-boxB2' domains are shown with red and blue colors, respectively. (c) Classification of AtBBX gene family structural groups (Groups I−VI). (d) Evolutionary hypothesis of BBX protein domains. (e) Characterization of Arabidopsis AtBBX genes. Neighbor-joining trees, synonyms, conserved motifs, domains, and exon-intron structures are indicated. '−' signifies that it is not available. Five color boxes indicate different motifs. Four color boxes indicate different domains. The dark green icon means UTR (untranslated region), the light green icon means CDS (coding sequence), and the line between UTR and CDS means intron (a non-coding part of a gene or mRNA molecule).