









-
Lectins represent a group of non-immune sugar-binding proteins with broad distribution across animals, plants, and microorganisms. These proteins possess a minimum of one non-catalytic domain that exhibits selective, reversible affinity for particular carbohydrate structures and glycoconjugates, and the surface sugar groups of glycolipids without changing the structural basis of sugars. In plants, lectins are widely present in different tissues and organs, each characterized by distinct structures, functions, and sugar-binding specificities. While playing an important role in promoting mitosis[1], cell agglutination, tumor suppression, antiviral, and pest resistance activities[2], lectins also participate in the recognition of rhizobia during symbiosis[3]. Rhizobia can transform into bacteroids in nodules and convert atmospheric nitrogen into ammonia for plant use. Rhizobia surface polysaccharides are essential for nodule formation and function, participating in governing nodule growth and plant immune response. They establish a symbiotic relationship with plants by secreting nodulation factors in response to plant signaling molecules, and subsequently differentiate into nitrogen-fixing bacteria in root hair cells.
The interaction between lectins and rhizobia is an important aspect of plant-microbe interactions, particularly in the process of symbiotic nitrogen fixation (SNF) between leguminous plants and rhizobia, which mainly involves molecular recognition and transduction, nodule formation, nitrogen fixation, and other processes. Nodule formation and nitrogen-fixation capacity are affected by various environmental factors, including nitrate availability, light, temperature, and water conditions, and are also regulated by plant hormones such as auxin, cytokinin, gibberellin, ethylene, and abscisic acid. This process is not only essential for the growth and development of the leguminous plants, but also provides a natural biological nitrogen-fixation mechanism for agricultural production. Over recent decades, substantial advancements have been achieved in elucidating the interactions between lectins and rhizobia, which have been summarized in several reviews; however, the presented knowledge from these reviews is largely from the model legumes alfalfa (Medicago sativa), Lotus japonica, and soybean, which are either non-food or primarily used for grain and oil production.
In contrast, vegetable legumes are characterized by their utilization of immature pods or seeds as the edible portion, which are typically consumed fresh or subjected to light cooking (e.g., steaming, stir-frying). Vegetable legumes play a crucial role in global agriculture, contributing significantly to food security, nutrition, and sustainable farming systems (Fig. 1). They are abundant in protein, fiber, and key micronutrients, making them essential dietary components, particularly in regions where animal protein is scarce and expensive. Among them, common bean (Phaseolus vulgaris) is a staple food for millions worldwide, with America, Morocco, and Mexico being the leading producers[4]. Cowpea (Vigna unguiculata) is widely cultivated in Africa and Asia[5], known for its drought tolerance and high nutritional value. As one of the most ancient domesticated crops, faba bean (Vicia faba), is predominantly produced in China, Ethiopia, and Australia[6]. Pea (Pisum sativum), extensively grown in temperate regions, contributes significantly to vegetable production, particularly in Europe and North America[7]. Sword bean (Canavalia gladiata) is valued in tropical regions for its resilience and potential as both food and fodder[8]. Edamame (Glycine max), the immature form of soybean, has become increasingly popular worldwide owing to its high protein content and health benefits. Hyacinth bean (Lablab purpureus), commonly grown in Africa and South Asia, is prized for its versatility as food, fodder, and soil-improving cover crop[9]. Collectively, these vegetable legumes contribute to global food production, enhance soil fertility through nitrogen fixation, and support sustainable agriculture, making them indispensable to both smallholder and commercial farming systems worldwide. The subsequent sections revisit the basic structure and biological roles of lectins, explain the classification and characteristics of rhizobia, and explore how they recognize each other to achieve the unique nodulation and nitrogen fixation in vegetable legumes whenever possible.
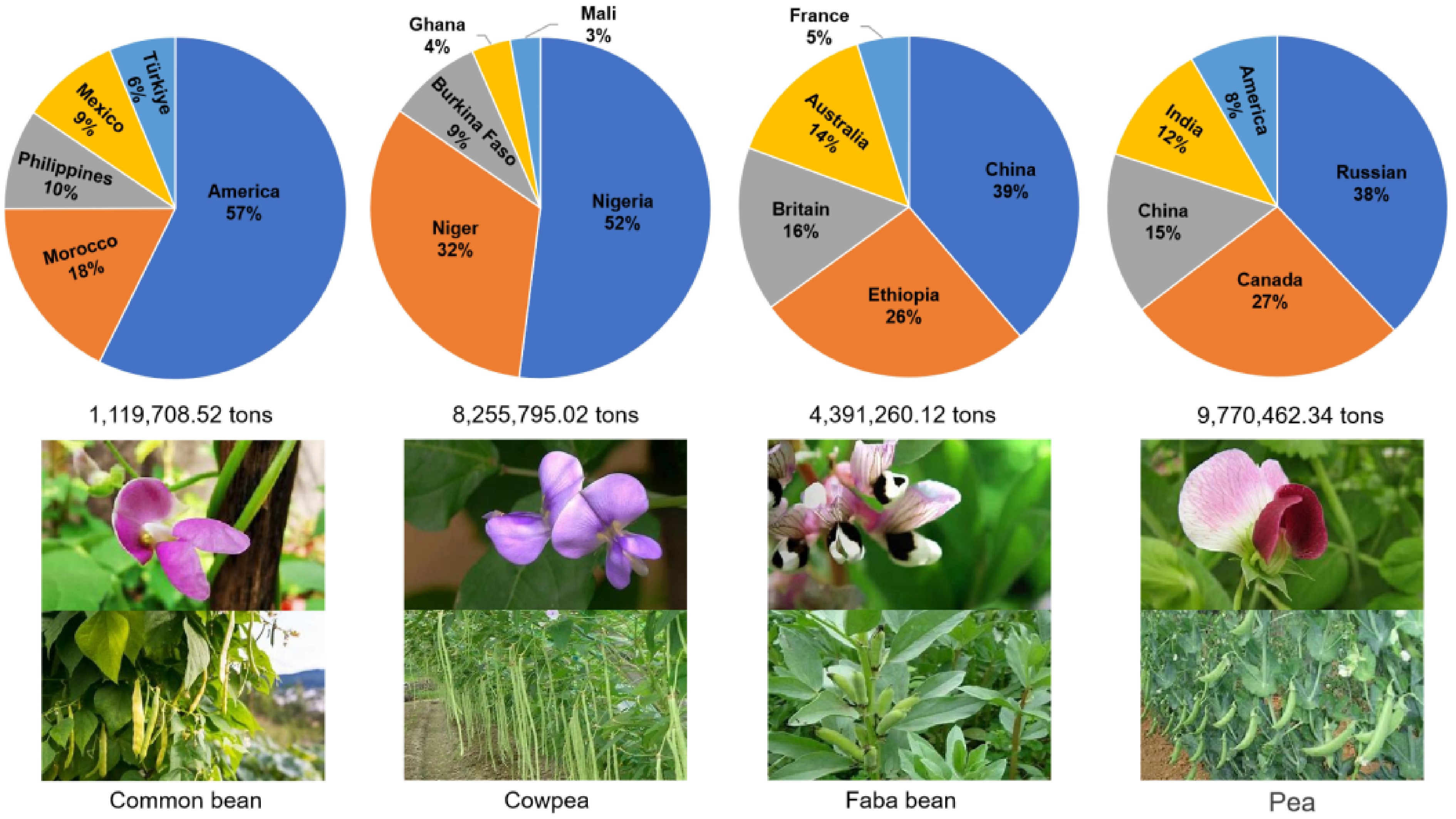
Figure 1.
Global yield percentages of the major vegetable legumes by country. The value under each pie chart represents the total production of this legume across the top five countries. Values in each pie chart represent the production percentage of these five countries. The global production data of common bean, cowpea, faba bean, and pea were sourced from FAOSTAT 2023 (
www.fao.org/faostat/en/#data/QCL ). -
The investigation of lectins can be traced back to 1888, when Russian scientist Herman Stillmark inadvertently identified a protein capable of agglutinating red blood cells while studying castor (Ricinus communis) seeds. This protein, later named ricin, was recognized for its potent cytotoxicity. Since then, substantial advancements have been achieved in the study of phytolectins, as these proteins are pivotal in plant growth, development, reproduction, and defense mechanisms.
Plant lectins can be categorized according to the structural characteristics of their subunits, including merolectins, which contain only a single sugar-binding domain; hololectins, which have at least two identical or structurally similar sugar-binding domains and represent the predominant class of plant lectins; superlectins, which consist of two or more sugar-binding domains; and chimerolectins, which include one or more domains with enzymatic or other biological activity in addition to sugar-binding domains[10]. According to their binding specificity to carbohydrates, they can be classified into N-acetylglucosamine lectins, D-mannose/D-glucose lectins, D-galactose lectins, L-fucose lectins, and sialic acid lectins. In addition, lectins can also be categorized according to their sequence similarity and their evolutionary relationship. Notable examples include legume lectins, chitin-binding lectins, jacalin-related lectins, etc. These classifications highlight the interrelationships among different lectin families and suggest their functional diversity throughout evolutionary history. Based on sequence homology of carbohydrate recognition domains (CRDs), lectins are phylogenetically classified into twelve major families: the legume lectins, the Agaricus bisporus agglutinin (ABA) family, the amaranthins, the chitinase-related agglutinin (CRA), the jacalin-related lectins (JRL), the cyanovirin family, the Galanthus nivalis agglutinin (GNA) family, the Euonymus europaeus agglutinin (EEA) family, the hevein family, the LysM domain lectin family, the ricin-B lectin family, and the Nictaba-like lectins[11].
Sugar-binding properties of lectins
-
The sugar-binding properties of plant lectins are one of their most prominent features. These proteins can specifically identify and attach to specific monosaccharides or oligosaccharides, with this binding typically being reversible and leaving the covalent bond structure of the sugar molecules intact. The sugar-binding sites of lectins usually consist of specific amino acid residues that interact with sugar molecules via multiple non-covalent bonds, including hydrogen bonds, van der Waals contacts, and electrostatic interactions. These interactions are essential for stabilizing the lectin-sugar binding affinity.
The specific sugar-binding ability allows lectins to distinguish between different cell surface sugar chains and thus participate in processes such as cell recognition, signal transduction, and immune response. For example, soybean agglutinin (SBA) can specifically bind to D-galactose and N-acetylgalactosamine (GalNAc), thus participating in symbiotic nitrogen fixation and having anti-nutritional effects[12] (Table 1). Common bean lectin (PHA) binds to N-glycans on T cells, especially β 1,6-branched complex N-glycans (Table 1). This interaction activates downstream tyrosine kinases Syk and ZAP-70, triggering T cell proliferation and IFN-γ secretion, thereby enhancing Th17-mediated antifungal immunity[13]. Faba bean produces lectins capable of recognizing and binding to specific polysaccharides on the surface of compatible Rhizobium leguminosarum (R. leguminosarum) strains. This binding initiates a signaling cascade that promotes the expression of nodulation-related genes, ultimately leading to successful symbiosis. Lectins extracted from cowpea exhibit strong antifungal properties through their binding to fungal cell wall components, including chitin and β-glucans[10]. Although individual lectins may recognize only specific sugar structures, plant lectins collectively have the ability to recognize a broad spectrum of sugar types, such as complex glycoconjugates.
Table 1. Classification and distribution of major plant lectins.
Lectin Mainly distribution Sugar-binding specificity Function Ref. Phaseolus vulgaris agglutinin (PHA) Seeds N-glycans Insect-resistance, antibacterial, antitumor [10,14] Conconvalin A (ConA) Seeds D-Glucose, D-Mannose Antitumor, promote angiogenesis [15,16] Soybean lectin (SBA) Seeds and roots D-Galactose, N-Acetylgalactosamine (GalNAc) Agglutinate red blood cells, promote lymphocyte division [12,17] Peanut agglutinin (PNA) Seeds and roots D-Galactose, GalNAc Antitumor, antibacterial [17,18] Pisum sativum agglutinin (PSA) Seeds D-Mannose, methyl-α-D-glucoside Antitumor [19,20] Lectins in the model and vegetable legumes
-
Lectins represent a varied family of proteins in numerous organisms, which are best characterized in legumes. They are often named after the plants from which they are extracted, such as conconvalin A (ConA), Pisum sativum agglutinin (PSA), lentil lectin (LCA), peanut agglutinin (PNA), and phytohemagglutinin (PHA) (Table 1), which all belong to the class of legume lectins.
In the soybean genome, up to 359 potential lectin genes were identified and categorized into nine structurally distinct families according to their CRD. The GNA family represents the most abundant group, comprising 166 members (46%)[21]. In conjunction with genome annotation, 32 highly confident legume lectins were identified. SBA is the most representative lectin in soybeans. This tetrameric glycoprotein is assembled from four identical 30 kDa subunits. Structural studies reveal that each subunit contains a conserved N-linked oligosaccharide (Man9GlcNAc2) and a GalNAc-specific binding pocket formed by conserved aromatic residues[12] (Table 1).
In the common bean genome, 52 lectin genes were identified and segregated into three evolutionarily distinct clades through phylogenetic analysis[22], among which 18 belong to the legume lectin family. Transcriptomic analysis revealed that five lectin genes (Pvul-BLEC-1 and Pvul-LLEC-15/16/18/27) exhibited constitutive high expression patterns, maintaining top-quartile expression levels in all vegetative and symbiotic tissues analyzed[22]. PHA, the most well-known common bean lectin, was discovered in the 1940s for its ability to agglutinate red cells, leading to its designation. PHA combines red blood cell (E) subunits and lymphocyte (L) subunits to form five different homoglectins, namely L4, L3E1, L2E2, L1E3, and E4. PHA-E is mainly involved in erythrocyte agglutination and has strong hemagglutination activity but weak mitogenic activity, while PHA-L has high mitogenic activity and is often used in immunological research (Table 1).
The composition of the entire pea lectin family has not been reported so far. PSA is a mitogenic globular protein extracted from pea seeds. It is classified as a non-immunoglobulin lectin. PSA forms a non-covalent homodimer at neutral pH, comprising two α chains (7 kDa) and two β chains (17 kDa). The protein contains two distinct CRDs that specifically bind to D-mannose and methyl-α-D-glucoside, respectively[19] (Table 1). Studies have shown that PSA binding to ligands depends on the presence of bivalent metal ions, especially Ca2+, and in the absence of these ions, PSA cannot effectively bind to ligands.
PNA is a 110 kDa peanut seed lectin composed of four identical polypeptide chains (236 aa each), exhibiting the typical quaternary structure of galactose-binding plant lectins. While its tertiary structure resembles that of other legume lectins, it lacks D2 and C4 symmetry, exhibiting a unique 'open' quaternary structure among known tetrameric proteins. PNA exhibits dual specificity for D-galactose (D-Gal) and GalNAc binding[17] (Table 1). Currently, there is a lack of reported characterization of lectin proteins in vegetable legumes such as soybean, pea, faba bean, mung bean, and cowpea. To address this, legume lectin sequences, which are the lectin subclass involved in symbiosis, were retrieved from available reference genome resources for these species. These sequences were then aligned, and a phylogenetic tree was constructed (Fig. 2).
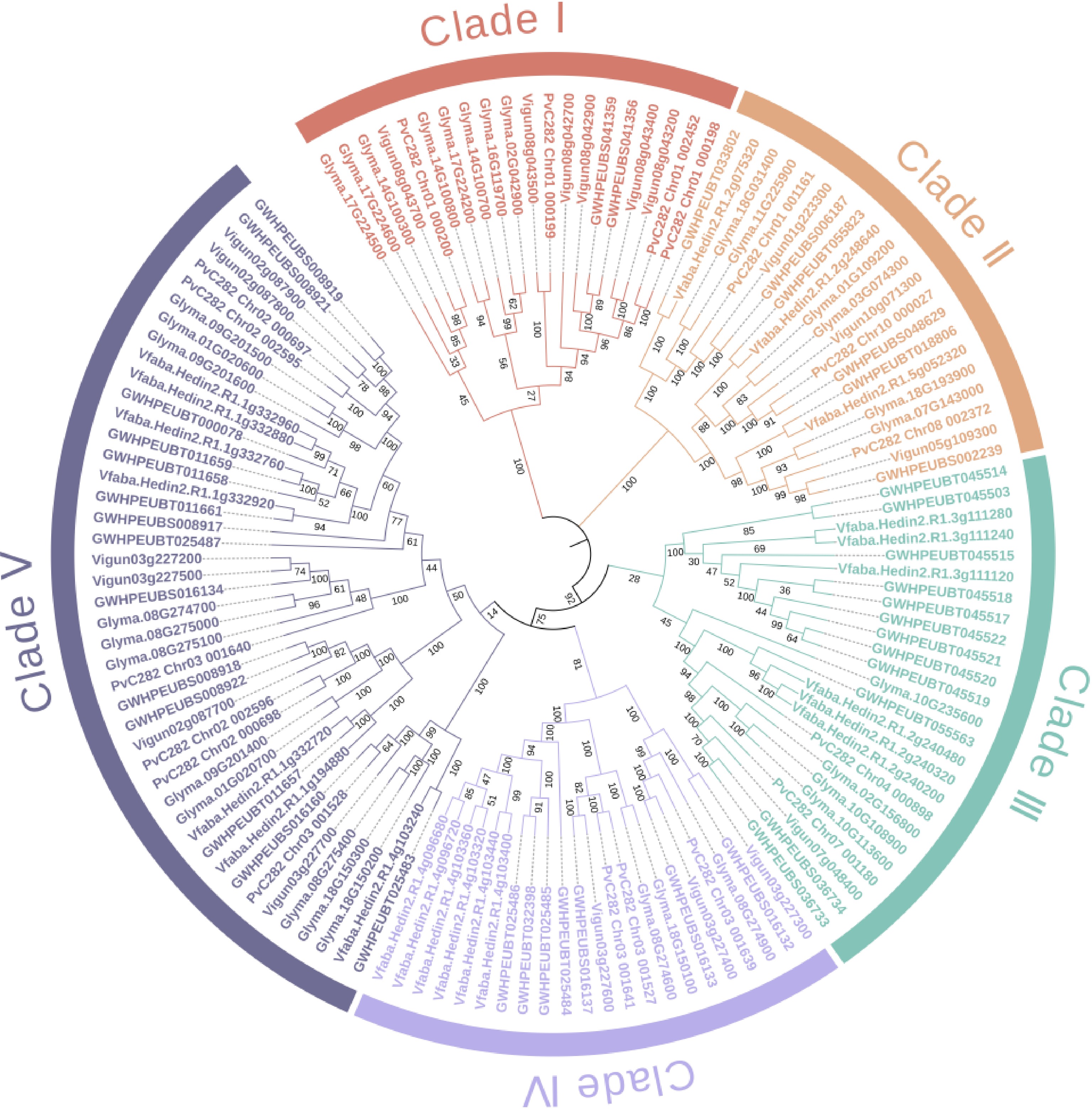
Figure 2.
Neighbor-joining tree of the legume-type lectin proteins in six vegetable legume crops. The identifier names of soybean, pea, faba bean, common bean, mung bean, and cowpea start with 'Glyma', 'GWHPEUBT', 'Vfaba', 'PvC282', 'GWHPEUBS', and 'Vigun', respectively. The lectin sequences were retrieved from the reference genomes: Glycine max Wm82.a2.v1 (soybean), Pisum sativum Pea_genome_GWHEUBT00000000.1 (pea), Vicia faba v1.1 (faba bean), common bean (in-house unpublished, available upon request), Vigna radiata Mungbean_genome_GWHEUBS00000000.1 (mung bean) and Vigna unguiculata v1.1 (cowpea), which are deposited in Phytozome (
https://phytozome-next.jgi.doe.gov/ ) or China National Center for Bioinformation (www.cncb.ac.cn ). In total, 132 legume lectin genes were identified across the six species, with the following distribution: soybean (32), pea (24), faba bean (22), common bean (18), mung bean (17), and cowpea (19). Their corresponding protein sequences are listed in Supplementary Table S1. A neighbor-joining phylogenetic tree was constructed based on these sequences (Fig. 2), resolving them into five major clades (Clades I−V), comprising 20, 21, 25, 22, and 44 genes, respectively.Recent research has also made remarkable progress in exploring the potential functions of vegetable legume lectin in plant growth promotion and pest control. Since lectins constitute 1%–10% of the total storage protein content in legume seeds, their decomposition during seed germination provides essential amino acids for plant growth and development. Lectins demonstrate antimicrobial activity through specific recognition of microbial surface glycoproteins, interfering with the synthesis of cell walls and thereby affecting their normal metabolism. For example, lectins of faba bean, lentil, and pea exhibit antibacterial activity against Klebsiella pneumoniae, Staphylococcus aureus, Streptococcus mutans, and Pseudomonas aeruginosa[23]. Additionally, legume lectins possess the ability to inhibit viral growth in vitro. Studies have found that ConA, faba bean lectins, and pea lectins can inhibit the binding of H9 cells to human immunodeficiency virus (HIV-1)[24]. Plant root lectins also participate in the recognition and binding of soil rhizobia, helping biological nitrogen fixation and reducing the dependence on fertilizer nitrogen. In addition, lectins are also involved in plant cell signaling, cell tissue and embryonic morphogenesis, cell wall growth, induction of mitosis, and pollen recognition[25] (Table 1).
-
Rhizobia, a group of Gram-negative bacteria, are widely recognized for their capacity to fix atmospheric nitrogen through symbiosis with leguminous plants. They can differentiate into bacteroids in the nodules of leguminous plants and convert atmospheric nitrogen into ammonia nitrogen, which is readily used by plants through biological nitrogen fixation. Initially, rhizobia taxonomy relied on phenotypic traits such as colony morphology, metabolic profiling, and symbiotic specificity. Rhizobia are phylogenetically classified within the domain Bacteria, phylum Pseudomonadota (formerly Proteobacteria). Depending on their host specificity, rhizobia can be categorized into Rhizobium nodulating temperate legumes and Bradyrhizobium nodulating tropical legumes. According to the growth rate of rhizobia, they can be classified into slow-growing rhizobia and fast-growing rhizobia. However, advances in molecular techniques, particularly 16S rRNA gene sequencing, have revealed that rhizobial taxa are primarily distributed across three classes: α-Proteobacteria, β-Proteobacteria, and γ-Proteobacteria. Among these, there are 14 genera of α-Proteobacteria, including Rhizobium, Sinorhizobium, Ensifer, Shinella, Neorhizobium, Pararhizobium, Mesorhizobium, Bradyrhizobium, Phyllobacterium, Methylobacterium, Microvirga, Ocrhobactrum, Azorhizobium, and Devosia. Within β-Proteobacteria, two genera are present: Burkholderia and Cupriavidus. There is only one genus of γ-Proteobacteria, which is Pseudomonas (Table 2).
Table 2. Classification and characteristics of major rhizobial genera involved in symbiotic nitrogen fixation.
Classification Representative strain Host plants Key characteristics Ref. Rhizobium R. leguminosarum Pisum sativum, Vicia faba, Phaseolus vulgaris Fast-growing type, symbiotic with temperate leguminous plants, and high nitrogen-fixing efficiency [26−29] Bradyrhizobium B. japonicum Glycine max, Arachis hypogaea Slow-growing type, acid soil-tolerant, and high nitrogen-fixing efficiency [30,31] Ensifer E. meliloti Medicago sativa, Melilotus officinalis, Phaseolus vulgaris Fast-growing type, strong salt tolerance, and high symbiotic nitrogen-fixing ability [29,32,33] Sinorhizobium S. fredii Glycine max, Vigna radiata, Phaseolus vulgaris Fast-growing type, salt-alkali tolerant, and with a large genome [34] Mesorhizobium M. loti Lotus japonicus, Trifolium repens Moderate growth rate, tolerance to infertile soil, and enhancement of plant resistance to pathogenic bacteria [35,36] Azorhizobium A. caulinodans Sesbania rostrata Moderate growth rate and high nitrogen-fixing efficiency [37] Neorhizobium N. galegae Medicago sativa Fast-growing type, aerobic bacteria, adapted to low-oxygen environments after symbiosis with plants. [38] Pararhizobium P. giardinii Phaseolus vulgari, Lens culinaris Fast-growing type, acid soil-tolerant, and high nitrogen-fixing efficiency [29,39] Allorhizobium A. vitis Vitis vinifera Mainly infect plants as pathogens and do not symbiotically fix nitrogen with leguminous plants [40] Nitrogen-fixing rhizobia in the model and vegetable legumes
-
Nodules result from mutual recognition and communication between rhizobium and legumes. Legume lectins are considered to mediate specific interactions with certain polysaccharides of appropriate rhizobium surface on the root surface of legumes, triggering a series of signal transduction processes between plants and rhizobia, thereby promoting rhizobial infection and the formation of root nodules. Rhizobia harbor symbiotic plasmids or chromosomal symbiotic islands that carry key genes for nodulation and nitrogen fixation, contributing to their ability to interact with diverse host plants. Due to the polyphyletic nature of rhizobia, they have evolved multiple times through horizontal gene transfer, leading to genetic diversity and broad host specificity.
The diversity of the model legume rhizobia associated with Medicago sativa has been well studied in some countries of Central Asia and Europe, the regions where this crop originated. For example, in Morocco, Japan, and Northwestern Iran[41,42], Medicago sativa was mainly associated with Sinorhizobium medicae (S. medicae) in symbiotic nodules. In the Mediterranean, more than 90% of the Medicago sativa rhizobia strains belonged to S. medicae, and only a few belonged to Sinorhizobium meliloti (S. meliloti), but S. meliloti had greater intraspecific genetic diversity and stronger resistance to environmental stress[36] (Table 3).
Table 3. Reported dominant rhizobium strains of legumes in specific areas.
Legume Strain Area Ref. Alfalfa S. meliloti Mexico, Japan,
Northwestern Iran[41,42] S. medicae Mediterranean [36] Soybean B. lingoningense Xinjiang Region [43] S. fredii Xinjiang Region,
Loess Plateau,
Huang-Huai-hai Plain[43,44] B. japonicum Heilongjiang Province [44] Common
beanR. etli Central and South
America, Europe[45] R. leguminosarum Andes, Nepal [46,47] Bradyrhizobium sp. Acidic soil of southern China [48] Pea R. laguerreae Northwestern Spain, Tunisia [49,50] R. leguminosarum and
R. ruizarguesonisTurkey [51] R. anhuiense Shandong Peninsula [52] Peanut B. liaoningense Shandong, Hebei, Jiangsu, Guangdong, and
Guangxi Province[53] B. yuanmingense and the
close relative of B. japonicumHenan and Sichuan
Province[53] B. japonicum and B. elkanii Argentina [54] B. lioningense, B. chanariense, B. aponicum, and B. etae Moroco [55] Cowpea B. elkanii Africa [57] Soybean originates from China. In different ecological regions of this vast country, especially those with varying ecological factors such as climate and soil, the population distribution of soybean rhizobia differs significantly. For instance, in the Xinjiang Region, Bradyrhizobium liaoningense (B. liaoningense) and Sinorhizobium fredii (S. fredii) are the dominant strains[43]. In the Loess Plateau and Huang-Huai-Hai Plain[44], soybean rhizobia exhibit high genetic diversity, with S. fredii being the dominant species, along with a small presence of slow-growing rhizobia. In Heilongjiang Province, most soybean rhizobia are slow-growing, with Bradyrhizobium japonicum (B. japonicum) being the dominant species[44] (Table 3).
The common bean, an important vegetable legume crop, originates from America and is a non-specific host plant that can be infected by various rhizobia. Rhizobium etli (R. etli) is the dominant strain in Central and South America, as well as Europe[45]. R. leguminosarum is the dominant strain in the Andes region of South America and Nepal[46,47], while Bradyrhizobium sp. is the primary rhizobium found in the acidic soils of southern China[48] (Table 3).
Another globally significant vegetable legume crop, pea, is native to the Middle East and Near East. The Rhizobia population capable of symbiosis with peas varies by soil composition and crop history in different geographical regions. For example, Rhizobium laguerreae (R. laguerreae) is dominant in both Northwestern Spain and Tunisia[49,50]. In Turkey, the most common pea microsymbionts are the composite gene species of R. leguminosarum, followed by Rhizobium ruizarguesonis (R. ruizarguesonis)[51]. In the coastal regions of the Shandong Peninsula, Rhizobium anhuiense (R. anhuiense) is the primary rhizobium associated with sweet pea[52] (Table 3).
Peanut sprouts are an emerging vegetable known for their pleasant flavor and high nutritional value. The rhizobia that can establish a symbiotic relationship with peanuts are mainly distributed in the genus Bradyrhizobium. In China, the dominant population in Shandong, Hebei, Jiangsu, Guangdong, and Guangxi Province is Bradyrhizobium lingoningense (B. lingoningense), while in Henan and Sichuan Province, it is Bradyrhizobium yuanmingense (B. yuanmingense) and the close relative of Bradyrhizobium japonicum (B. japonicum), respectively[53]. Outside China, studies on the diversity of peanut Bradyrhizobium spp. mainly focus on major peanut-producing countries such as Argentina and Africa. The phylogenetic status of peanut bradyrhizobium in Argentina is similar to that of B. japonicum and Bradyrhizobium elkanii (B. elkanii)[54]. Phylogenetic analyses reveal distinct geographic patterns among peanut-associated bradyrhizobia. Isolates from Argentina cluster closely with Bradyrhizobium japonicum and B. elkanii[54], while Moroccan strains show higher similarity to B. liaoningense, B. chanariense, B. aponicum, and B. etae[55] (Table 3).
Cowpea-associated rhizobia are predominantly slow-growing Bradyrhizobium species[56], demonstrating remarkable adaptability to nutrient-deficient acidic soils through efficient nitrogen fixation. A study of Ethiopian isolates revealed 93.3% nodulation efficiency, with significant phenotypic diversity observed among the symbiotic strains[56]. Notably, B. elkanii emerges as the dominant symbiont in African cowpea cultivation systems (e.g., Nigeria, Kenya), exhibiting exceptional tolerance to soil infertility[57] (Table 3). Muindi et al. assessed the symbiotic efficiency and genetic diversity of native rhizobia isolated from root nodules of cowpea genotypes grown in the semi-arid lower Eastern Kenya region. Through 16S rRNA gene sequencing, the isolates were found to be closely related to bacteria in the genera Rhizobium, Paraburkholderia, and non-rhizobial endophytes (Enterobacter, Stenotrophomonas, and Pseudomonas). Notably, this study first reported the presence of an efficient native cowpea-nodulating Beta-Rhizobia (Paraburkholderia phenoliruptrix BR3459a) in the African continent[58].
-
Legume roots release a number of secondary metabolites into the rhizosphere as chemical signals for rhizobia Nod factor induction, which have been identified as flavonoids or closely related compounds (polycyclic aromatic hydrocarbons) (Fig. 3). Up to now, more than 9,000 different compounds have been identified as flavonoids, and most SNF is known to be initiated by the root-sourced flavonoids. Upon perception by rhizobia, flavonoids bind to the bacterial transcriptional regulator NodD, activating the expression of nod genes (Fig. 3). To initiate the symbiotic signaling pathway, Nod factors are recognized by plant receptors, which mediate rhizobia invasion and nodule development. Different flavonoids, such as daidzein, genistein, and luteolin, induce nod gene expression in a host-specific manner. For example, daidzein and genistein, two isoflavonoids primarily found in soybeans, are pivotal in the symbiosis between rhizobia and leguminous plants like soybeans[59]. Luteolin, a flavonoid commonly found in Medicago sativa, induces nodulation by activating the NodD protein of S. meliloti. Similarly, quercetin, another flavonoid found in several legumes like chickpeas and lentils, induces nod gene expression in R. leguminosarum and enhances nodule formation.
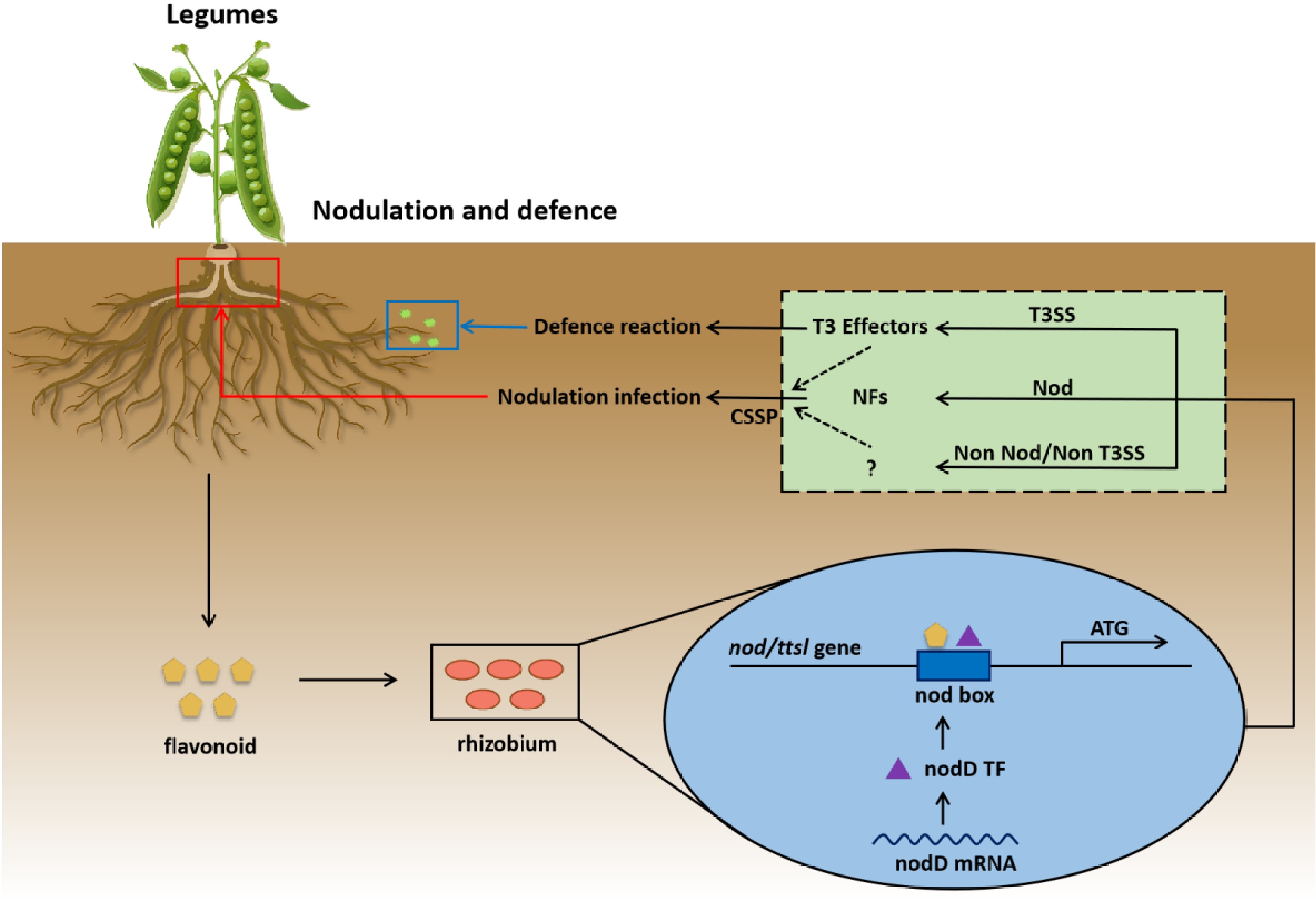
Figure 3.
Schematic diagram of the symbiotic nitrogen fixation mechanism with rhizobia. In this process, flavonoids secreted by legume roots act as initial signals and are recognized by rhizobia. Inside the rhizobia cell, flavonoids induce the expression of the nodD gene. The nodD transcription factor (TF), which binds to the nod box adjacent to the nod/ttsl genes, initiates the production of Nod factors (NFs) related to the Nod operon. NFs, likely in association with cell-secreted signaling proteins (CSSP), drive the nodulation infection of the legume root. Concurrently, a parallel pathway involves the type III secretion system (T3SS) in rhizobia. T3SS secretes T3 effectors, which trigger a defense reaction in the legume. Additionally, a 'Non Nod/Non T3SS' pathway exists, which may represent a regulatory mechanism to fine-tune rhizobia-legume interactions. This mechanism helps prevent excessive nodulation or over-activation of the defense response, thus maintaining a balanced interaction and ensuring the efficient progress of symbiotic nitrogen fixation.
Nodule formation involves the participation of numerous genes, among which nod genes (nod, nol, and noe) co-synthesize Nod factors (Fig. 3). Common nod genes such as NodA, NodB, and NodC exist in all rhizobia and are necessary for synthesizing the core structure of Nod factors. They exist as single-copy genes in most rhizobia and control the synthesis of the basic skeleton structure of Nod factors. For instance, in R. leguminosarum (a model rhizobium species for peas), NodC encodes N-acetylglucosamine transferase, responsible for synthesizing the LCO skeleton, which is essential for nodule formation in peas[60]. NodB encodes deacetylase, enabling the deacetylation of N-acetylglucosamine at the non-reducing end of LCOs. NodA encodes acyltransferase, catalyzing the transfer of long-chain acyl groups to the amino groups of glucosamine[61]. Another essential gene for nod factor synthesis is NodD. The NodD protein is a member of the prokaryotic LysR family of transcriptional regulators and typically exists as a tetramer. Conformational changes in this protein enable it to specifically recognize and bind to the conserved sequences within the promoter regions of nodulation genes, thereby directly activating the NodA–NodB–NodC operon. For example, in S. meliloti, NodD is activated by plant-produced flavonoids, initiating the expression of nod genes involved in nodule formation. Additionally, by regulating the expression of other nod genes, NodD further modifies the side-chain structure of Nod factors, thereby determining host specificity. For instance, luteolin in alfalfa and daidzein in soybeans can modulate the specificity of NodD-induced gene expression, influencing which rhizobial strains can successfully form nodules on these plants[59].
During the synergistic nitrogen fixation process between legumes and rhizobia, plant-derived flavonoids, the regulatory protein NodD in rhizobia, and the nod-box promoter sequence are involved in the early recognition of symbiotic signals. Rhizobia recognize plant flavonoids through NodD proteins, thereby triggering the nod gene expression[62]. In the absence of an inducer, the NodD protein can bind to DNA and cause nod-box sequence bending, but it does not activate downstream nod gene expression. When flavonoids are present, the NodD protein binds to the nod-box, enhancing the degree of DNA bending. The NodD protein may change its structure and conformation after recognizing the corresponding flavonoids. The allosteric NodD protein binds to the promoter sequence of downstream nod genes, allowing RNA polymerase to form an active transcriptional open complex downstream of nod genes, thereby activating their expression. During early nodule formation, rhizobia are involved in root hairs and form invasion lines at the site of invasion. Rhizobial surface polysaccharides can interact with host plants.
Lectin and symbiotic nitrogen fixation
-
The historical hypothesis on rhizobium-legume lectin interactions began with Hamblin & Kent's observation that adding a suspension of Rhizobium phaseoli to Phaseolus vulgaris lectin induced agglutination, leading them to propose that lectin binding directed rhizobia to specific root sites for infection[63]. Dazzo & Hubbell later demonstrated that clover root surface lectins could agglutinate infectious rhizobia and proposed the cross-bridge hypothesis, suggesting that clover lectins recognize rhizobial surface antigens to form a molecular interface for specific adsorption[64]. Lerouge et al. extended this idea by proposing that lectins can specifically interact with rhizobial Nod factors, with leguminous lectins possessing glycobinding sites and hydrophobic pockets that bind either the oligomeric chitin moiety of Nod factors or the fatty acid chains of rhizobia[65]. Diaz et al. further reported that host plant lectins play a decisive role in legume-rhizobium symbiosis. However, subsequent studies have shown that while lectin-mediated agglutination and recognition are important in some legume-rhizobium interactions, they may not be universally essential across all legumes (Fig. 4). In many species, the primary symbiotic signals are Nod factors (lipo-chitooligosaccharides, LCO), which are perceived by plant receptor kinases containing LysM domains (e.g., NFR1/NFR5 in Lotus japonicus and LYK3/LYK4 in Medicago truncatula) rather than solely by lectins[66] (Fig. 4). Thus, the early hypotheses that lectins serve as the exclusive 'bridge' between rhizobia and legume roots have evolved into a more nuanced model. Current evidence suggests lectins may act as accessory or fine-tuning elements, enhancing host specificity, attachment, or infection efficiency in certain legume-rhizobium pairs (Fig. 4). Moreover, the discovery of numerous lectin-like receptor kinases in legume genomes indicates additional, potentially overlapping, recognition mechanisms that extend beyond simple agglutination[67]. These insights underscore the complexity of legume-rhizobium interactions and highlight the need for integrated approaches spanning genetics, biochemistry, and structural biology to fully unravel how lectins contribute to symbiotic specificity.
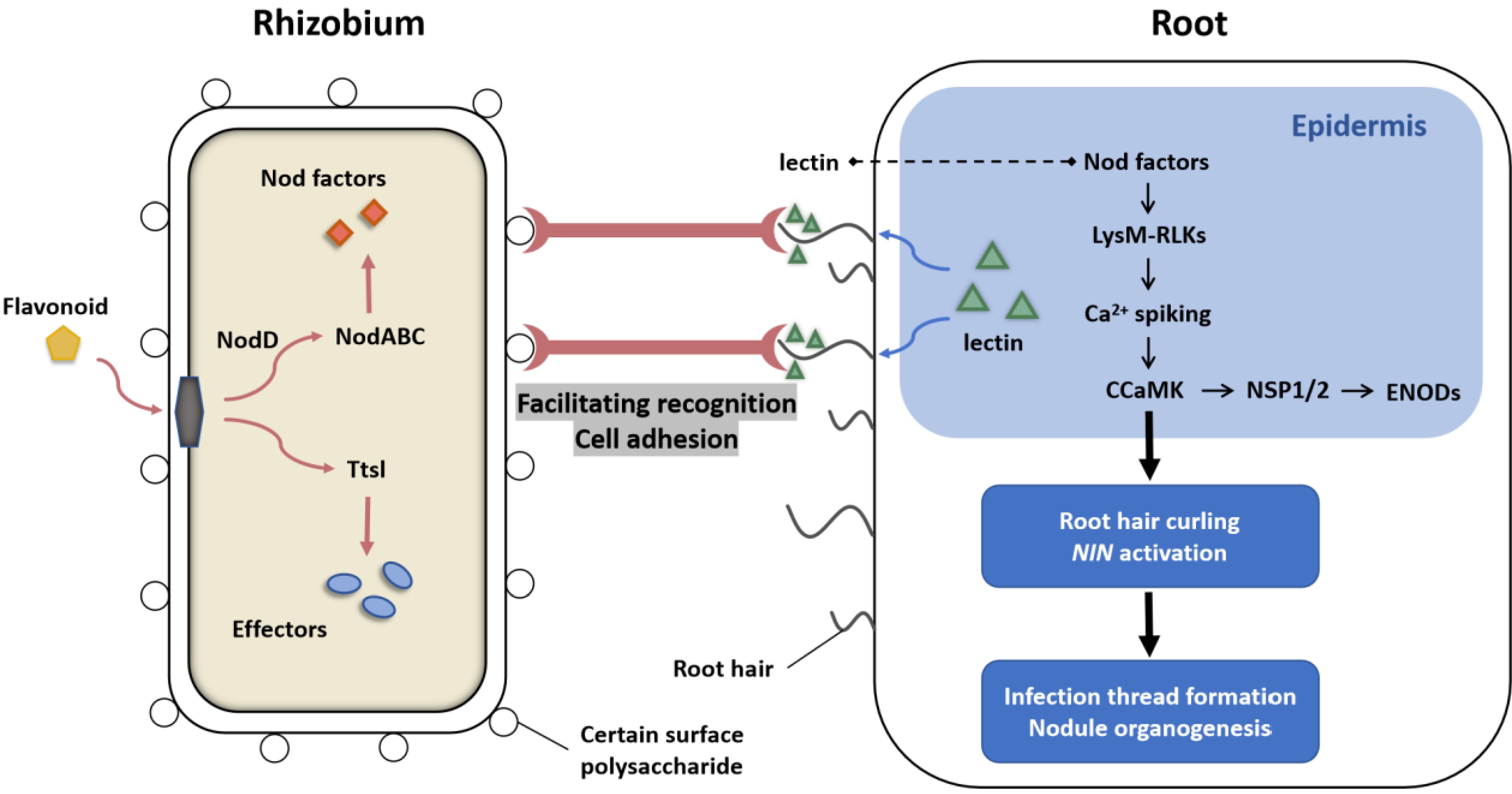
Figure 4.
Mutual recognition mechanism between lectins and rhizobia. Flavonoids interact with rhizobial components such as nodD and induce the expression of nod genes. Lectins, acting as 'molecular sentinels' on the root surface, accumulate at the tips of root hairs-key sites for rhizobial infection. Their carbohydrate-binding domains specifically recognize glycan modifications on the rhizobial surface, promoting bacterial colonization and initial attachment to the host root. This lectin-mediated accumulation enhances local bacterial density on root hairs, creating a favorable micro-environment for subsequent infection. By stabilizing the interaction between rhizobia and root hairs, lectins may also facilitate intercellular communication and signal exchange among rhizobia, increasing the chances of successful symbiosis. Notably, lectins can also recognize specific sugar structures on NFs, possibly through direct binding with the lectin's carbohydrate-binding site and the sugar chains of NFs. The NF signal is subsequently perceived by LysM receptor kinases, such as NFP and LYK3, in the root epidermis. Upon NF recognition, these receptors trigger root hair deformation and intracellular Ca2+ spiking, initiating downstream signaling cascades. This includes the activation of the CCaMK-NSP1/2-ENOD pathway and NIN, ultimately driving infection thread formation and nodule organogenesis. However, some evidence proves that lectins are not indispensable for root nodule nitrogen fixation, suggesting redundant recognition pathways.
The mutual recognition between Legumiaceae and rhizobia is an affinity-based recognition system. The bacteria must first adsorb to the host root hair, and then invade the host by the infection thread to induce nodules. Common bean seedlings can release root lectins onto the medium[68]. In the primary root meristem, PHA accumulated in vacuoles, while in elongating root cells, PHA was detected exclusively in cell walls, which was suggested to be the source of PHA in the medium. However, it was later found that LysM receptor kinase recognizes Nod factors secreted by rhizobia through its extracellular domain (Fig. 4). Gherbi found that in Medicago sativa, Lotus japonica, and other legumes, LysM receptor kinase NFR1 and NFR5 participate in Nod factor perception, thus activating a series of intracellular signal transduction processes[66]. Tan et al. reported that two LysM receptors, MpaCERK1 and MpaLYR, can recognize chitin oligosaccharides of different lengths in the liverwort Marchantia paleacea and mediate immune and symbiotic reactions[69]. They speculated that LysM receptors may perceive the presence of rhizobia by recognizing chitin structures in Nod factors[69]. The pea Sym37, which encodes a LysM receptor kinase similar to Lotus japonicus NFR1 and Medicago truncatula LYK3, controls infection thread initiation and nodule development[66]. Additionally, studies have revealed that lectins enhance the attachment ability of rhizobia to plant root hairs by recognizing and binding to sugar molecules on the surface of rhizobia (Fig. 4). Bhagwat tracked its binding to rhizobia by labeling lectins, and found that peanut lectins could achieve specific recognition of rhizobia by interacting with two distinct binding sites on the bacterial cell surface, which provided the necessary molecular basis for the colonization of rhizobia and subsequent infection. According to the selective adsorption theory, the selectivity of rhizobia to adsorb root hairs of leguminous and non-host plants is an important reason for nodule specificity. One rhizobia or its specific strain can only specifically infect its source host and a few other hosts and induce nodule formation.
Nod Factors are the major signaling molecules during nodule formation. The structural diversity of Nod factors, including different glycan lengths and lipid chain modifications, determines their specific recognition with host plants. These signaling molecules are able to be recognized by receptors in plant roots, triggering a cascade of downstream signaling processes that ultimately result in nodule formation. It was found that when lectins from leguminous trees were mixed with rhizobia exopolysaccharides (EPS), not all lectins could bind to rhizobia EPS, and not all species with binding reaction could nodulate, indicating that the interaction between Nod factors and lectins is a key step in nodule formation. The lectin-like protein Db-LNP, isolated from Dolichos biflorus, binds various rhizobial Nod factors with high affinity and possesses apyrase activity, hydrolyzing ATP, ADP, and AMP—an enzymatic function important in plant signal regulation[70]. Lectins can recognize specific sugar structures on Nod factors. This recognition may involve a direct interaction between the lectin's carbohydrate-binding site and the sugar chains of the Nod factors. Completion of symbiosis depends on the correct composition of rhizobial surface polysaccharides, a second determinant of rhizobia specificity.
Nodule formation and nitrogen fixation
-
The process of nodule formation, following successful molecular recognition and bacterial infection, includes differentiation of root hair cells and formation of nodule organs. During the initial phase of nodule formation, rhizobia colonize root hairs and initiate the formation of infection threads at the site of invasion. These infection threads guide the bacteria into the root cortex, where they stimulate cortical cells to develop into the nodule primordium. Once released from the infection thread, the bacteria differentiate into bacteroids and gain the capability to fix nitrogen in the microaerobic environment within the nodule. Nitrogenase, an enzyme conserved across most nitrogen-fixing bacteria, catalyzes nitrogen fixation. This nitrogenase is a two-component enzyme complex that includes dinitrogenase MoFe as the catalytic component and dinitrogenase reductase. The nifD, nifK, and nifH genes encode these two metal-containing components, respectively[71]. Moreover, nif genes also encode some regulatory proteins involved in nitrogen fixation. Leghemoglobin, an oxygen-binding protein present in the root nodules of leguminous plants, protects the nitrogenase enzyme from oxygen inactivation and facilitates oxygen supply to nitrogen-fixing bacteria. It was found that leghemoglobin expressed in the infected area and the differentiated area of rhizobia had a lower affinity for oxygen, while leghemoglobin expressed in the mature nitrogen-fixing area had a higher affinity for oxygen. This difference may be related to the gradual induction of the nitrogen-fixing genes of rhizobia, indicating that the expression of leghemoglobin is closely related to the nitrogen-fixing ability of rhizobia. Ren et al. recently identified iron as a crucial driver of nodule formation in leguminous plants and further elucidated the molecular mechanism by which the iron receptor BTS facilitates nodulation and symbiotic nitrogen fixation. Specifically, BTS promotes the monoubiquitination of the key symbiotic signaling transcription factor NSP1, enhancing its protein stability and transcriptional activity[72].
In vegetable legumes, many other genes have been identified as affecting the nodulation and nitrogen fixation. For example, the noncanonical heat shock protein PvNod22 plays a crucial role in the progression of infection during rhizobial endosymbiosis in common bean[73]. MADS-Domain/AGL transcription factors and the microRNA319d/TCP10 node were also found to control the rhizobia nitrogen-fixing symbiosis in common bean[74,75]. In common bean roots, overexpression of PvBI-1a enhances rhizobial infection events and subsequently increases nodule formation, but also leads to premature nodule senescence, thereby reducing the nitrogen fixation efficiency of nodule cells[76]. As technology advances, the gene pool involved in the fine-tuning of SNF between leguminous vegetables and rhizobia is believed to be continuously expanding.
-
As recent advances underscore the pivotal role of legume lectin-rhizobia interactions in SNF, it is both logical and promising to extend this knowledge beyond the legume family. Multiple lines of evidence now suggest that transferring lectin genes from legumes (e.g., peas, beans) into non-legume crops can modify root surface properties and enhance rhizobial adhesion. By doing so, cereals and other non-legume species may acquire the capacity to engage with nitrogen-fixing microbes, thereby reducing their dependence on synthetic fertilizers.
Realizing this vision will require optimizing lectin expression patterns, for instance, by using tissue-specific promoters and engineered microbial strains that respond appropriately to the lectin signals of the transgenic host. Success in these endeavors would mark a significant step toward sustainable agriculture, enabling lower environmental impacts alongside higher productivity. Though still in its early stages, this approach opens an exciting avenue for leveraging the nodulation toolkit of vegetable legumes to establish novel plant-microbe symbioses in staple non-legume crops.
Multifaceted benefits to plants from lectin-rhizobium interactions
-
Beyond their role in promoting nodulation, legume lectins confer diverse advantages that strengthen plant health and resilience. In legumes such as soybeans and beans, these proteins are pivotal not only for recognizing beneficial rhizobia but also for defending roots against pathogens and tolerating environmental stresses. For instance, expression of a soybean lectin in transgenic tobacco results in enhanced resistance to pathogens and pests[77], while red kidney bean lectin inhibits the growth of fungi. Moreover, a lectin derived from kidney beans shows strong inhibitory activity against Rhizoctonia solani and Sclerotium rolfsii. These antifungal properties extend to viral inhibition as well: sword bean, fava bean, and pea lectins can all prevent HIV-1 from binding to host cells in vitro[24]. Such evidence suggests that lectins can act as broad-spectrum defense agents, safeguarding the plant's root environment even as they facilitate symbiotic infection threads for rhizobia.
Additionally, mounting data indicate that lectin-rhizobium interactions may enhance tolerance to abiotic stresses (e.g., drought and salinity). For example, a high density of root hairs can secrete flavonoid metabolites to promote the enrichment of rhizobiaceae microorganisms, thus enhancing the drought resistance of wheat[78]. In soybeans infected by rhizobia, GmNAC181 enhances nodulation and improves the salt tolerance of root nodules through direct regulation of GmNINa gene expression[79]. By fine-tuning stress-responsive hormone levels and bolstering antioxidant enzyme activity, lectins help maintain nodule function and overall plant vigor under challenging conditions. In this sense, vegetable legumes stand as a valuable system for elucidating how lectins integrate multiple signaling pathways to promote nitrogen-fixing symbioses while fortifying the plant's defenses against both biotic and abiotic threats. Together, these multifaceted benefits underscore the remarkable versatility of legume lectins. Future work should focus on pinpointing the structural and regulatory elements behind lectin-mediated pathogen defense and stress tolerance, particularly in economically significant vegetable legumes. Ye et al. underscored the transformative impact of omics technologies in decoding the intricate processes of symbiotic nitrogen fixation, offering promising avenues for agricultural innovation and environmental sustainability[80]. Such insights could drive the development of new biotechnological tools or breeding approaches that leverage lectin functions to enhance both yield stability and sustainability in legume-based cropping systems.
Despite significant progress in understanding lectin-rhizobium interactions, several key questions remain unresolved: The precise molecular mechanisms by which plant lectins recognize and bind specific rhizobial surface molecules, such as Nod factors, lipopolysaccharides, or exopolysaccharides, are still not fully understood. The specificity and affinity of lectin-ligand interactions across different legume-rhizobium pairs also require deeper investigation. Moreover, the downstream signaling pathways activated by lectin binding, and their integration with canonical symbiotic signaling components like NFRs (Nod factor receptors), remain unclear. It is also not well understood how lectins modulate host specificity and contribute to the selection or exclusion of certain rhizobial strains. Finally, the cross-species effects of lectins and their potential applications in enhancing symbiosis in non-legume crops are still largely unexplored. A comprehensive understanding of these unresolved issues will be critical for both advancing the fundamental biology of symbiosis and applying lectins in sustainable agriculture.
This study is supported by the National Natural Science Foundation of China (Grant Nos U24A20419, 32302545, 32201254), Natural Science Foundation of Zhejiang Province (Grant Nos LTGN24C130003, LQ24C060006), and the Shandong Province Key Research and DevelopmentProgram (Grant No. 2023LZGCQY012).
-
The authors confirm their contributions to the paper as follows: conceptualization: Yang S; writing – original draft: Yang S, Wang S; writing – review and editing: Chen H, Gong Y, Lu C, Xu M, Ning K; supervision, resources, funding acquisition: Xu P. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Su Yang, Siyi Wang
- Supplementary Table S1 Legume lectin protein sequences used for phylogenetic reconstruction.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang S, Wang S, Chen H, Gong Y, Lu C, et al. 2025. The lectin-rhizobium interactions underlying symbiotic nodulation: a focal review on vegetable legumes. Vegetable Research 5: e036 doi: 10.48130/vegres-0025-0029 

The lectin-rhizobium interactions underlying symbiotic nodulation: a focal review on vegetable legumes
- Received: 01 April 2025
- Revised: 06 June 2025
- Accepted: 23 July 2025
- Published online: 10 October 2025
Abstract: Lectins are carbohydrate-binding proteins that play key roles in cell recognition, signaling, and plant defense. In leguminous plants, lectins are crucial for symbiotic interactions with rhizobia—nitrogen-fixing bacteria that enhance soil fertility and promote plant growth. Understanding the regulatory networks underlying lectin-rhizobium interactions is essential for advancing agricultural biotechnology and global food security. Although substantial advances have been made in elucidating these interactions and summarized in several reviews, most presented knowledge comes from model legumes such as Medicago sativa, Lotus japonicus, and Glycine max. In contrast, vegetable legumes, characterized by their edible immature pods or seeds, occupy a significant position in global agriculture, yet their lectin-rhizobium interactions remain poorly summarized. To address this gap, this review explores the intricate mechanisms governing lectin-rhizobium interactions, with a particular emphasis on insights derived from vegetable legumes such as common beans, cowpeas, and peas. Following the introduction of lectins and rhizobia, the complete process of symbiotic nitrogen fixation is revisited, spanning from mutual recognition mechanism between lectins and rhizobia to nodule formation and nitrogen fixation. The potential of transferring legume lectin genes into non-leguminous crops to improve nitrogen fixation is also discussed. Finally, the importance of unraveling the molecular mechanisms governing these interactions is highlighted to enhance symbiotic efficiency and promote sustainable crop production.
-
Key words:
- Lectins /
- Rhizobium /
- Symbiotic nitrogen fixation /
- Plant-microbe interactions /
- Model legumes /
- Vegetable legumes