









-
Chinese cabbage (Brassica rapa ssp. Pekinensis) is a globally significant leafy vegetable, widely cultivated and available year-round. Petiole spot is a significant physiological disorder in Chinese cabbage, typically observed in head leaves. It is characterized by numerous sesame-like spots, black or dark brown in color, primarily localized in the epidermal cells of the petiole[1,2]. These spots generally range from 1 to 2 mm in diameter, which significantly affects appearance and marketability, causing notable economic losses[1,3]. Over the past four decades, research on petiole spot has predominantly centered on field observations and physiological characterization[4−7]. To date, no pathogens have been identified as being closely associated with petiole spot in Chinese cabbage, and it is therefore widely regarded as a physiological disorder[1,3]. Excessive nitrogen fertilization is considered a major contributing factor, with fertilizer trials demonstrating that elevated nitrogen levels significantly exacerbate symptom severity by increasing the number and density of petiole spots[8]. Moreover, the form of nitrogen supplied influences the development of petiole spot: ammonium nitrogen has a stronger promotive effect than either nitrate or amide nitrogen[4]. A higher proportion of ammonium nitrogen can suppress nitrate reductase (NR) activity in the petiole, thereby affecting nitrogen assimilation and contributing to petiole spot formation[8]. In addition to nitrogen, other contributing factors include high copper (Cu), low boron (B) levels[8], and alkaline soil conditions with pH 8.0, particularly conducive to symptom development[1]. Environmental stresses such as suboptimal temperatures, and low light intensity have also been implicated[9]. While existing studies have predominantly focused on factors contributing to petiole spot development[10], the underlying mechanisms governing its occurrence remain poorly elucidated. Moreover, genotype significantly influences the occurrence of petiole spot[2]. Investigations of multiple Chinese cabbage inbred lines, as well as domestic and international cultivars, have revealed substantial varietal differences in symptom incidence, indicating a genotype-dependent response[2]. However, the genetic basis underlying this disorder remains unclear. Overall, petiole spot results from the interaction between genetic and environmental factors, and its underlying mechanisms warrant further investigation.
Through microscopic and scanning electron microscopy (SEM) observations, petiole spots have been identified as resulting from cellular necrosis[11]. Previous studies revealed that resistant cultivars exhibit enhanced ammonium (NH4+) assimilation capacity, whereas susceptible lines accumulate excess NH4+, triggering bursts of reactive oxygen species (ROS)[11]. Microscopic analysis showed that lesions initiate in the intercellular spaces and gradually spread inward, leading to cell death[11]. This spatiotemporal progression suggests that intracellular ROS accumulation is a key driver of symptom development. Simultaneously, as signaling molecules—particularly hydrogen peroxide (H2O2)—ROS play a central role in redox signaling networks that regulate plant stress responses[12−14]. This dual function highlights ROS as a key molecular link between nitrogen metabolic imbalance and disease occurrence.
Chinese cabbage is commonly stored postharvest. Petiole spots manifest not only during head formation but also emerge in storage. Symptom-free heads frequently develop lesions during storage and transport, with existing spots progressing severely: lesions enlarge, causing epidermal cracking or decay[8]. Previous studies focused on environmental and nutritional factors driving spot development at the heading stage[4−7,15], whereas mechanisms underlying symptom exacerbation during storage remain elusive. In this study, it was found that petiole spot symptoms in stored Chinese cabbage intensified from outer to inner leaves during storage, indicating a clear spatial and temporal progression, which is closely linked to ABA signaling and ROS accumulation. Transcriptomic, hormonal, and pharmacological analyses demonstrated elevated ABA levels and upregulation of ABA-related genes in lesion tissues, with expression patterns consistent with the spatial distribution of petiole spots. ROS was identified as a key downstream effector in lesion development. Exogenous H2O2 could mimic ROS-induced symptom formation, while ROS biosynthesis inhibition was suppressed. Notably, ABA and ROS acted synergistically in petiole spot formation. These findings offer a mechanistic foundation for developing novel postharvest intervention strategies for Chinese cabbage.
-
Three Chinese cabbage cultivars—Han Chunwa 4-2-3-4 (spring Chinese cabbage), Line 163 (summer Chinese cabbage), and Zaoshu No. 8 (autumn Chinese cabbage)—were selected for this study based on their susceptibility to petiole spot (Supplementary Fig. S1). These cultivars were bred and provided by the Vegetable Institute of the Zhejiang Academy of Agricultural Sciences (Hangzhou, China). Seeds were germinated on half-strength Murashige and Skoog (1/2 MS) medium and grown under greenhouse conditions with a 14 h photoperiod at 25 °C and a 10 h dark period at 16 °C for 15 d. The seedlings were then transplanted into a controlled-environment growth chamber and cultivated for approximately three months until head formation. At maturity, plants were transferred to storage at 4 °C.
After one week of storage, petiole epidermal tissues without visible lesions were designated as control (CK), and those exhibiting characteristic petiole spot symptoms were labeled as affected samples (PS). Three biological replicates were collected for each condition and cultivar. Each biological replicate was obtained by pooling tissues from three individual plants of the same cultivar. Samples were carefully collected from both the upper and lower epidermal layers of the leaf petioles, with mesophyll tissue excluded. Petiole spots were typically observed on the outer six leaf layers (counted from the outermost to the innermost leaves), exhibiting a clear spatial distribution pattern, with the first leaf showing high spot density and the sixth leaf showing low or no occurrence (Fig. 1). Accordingly, PS samples were collected from the epidermal layers of the first leaves with high spot density, while CK samples were taken from the sixth leaves that were free of visible petiole spots.
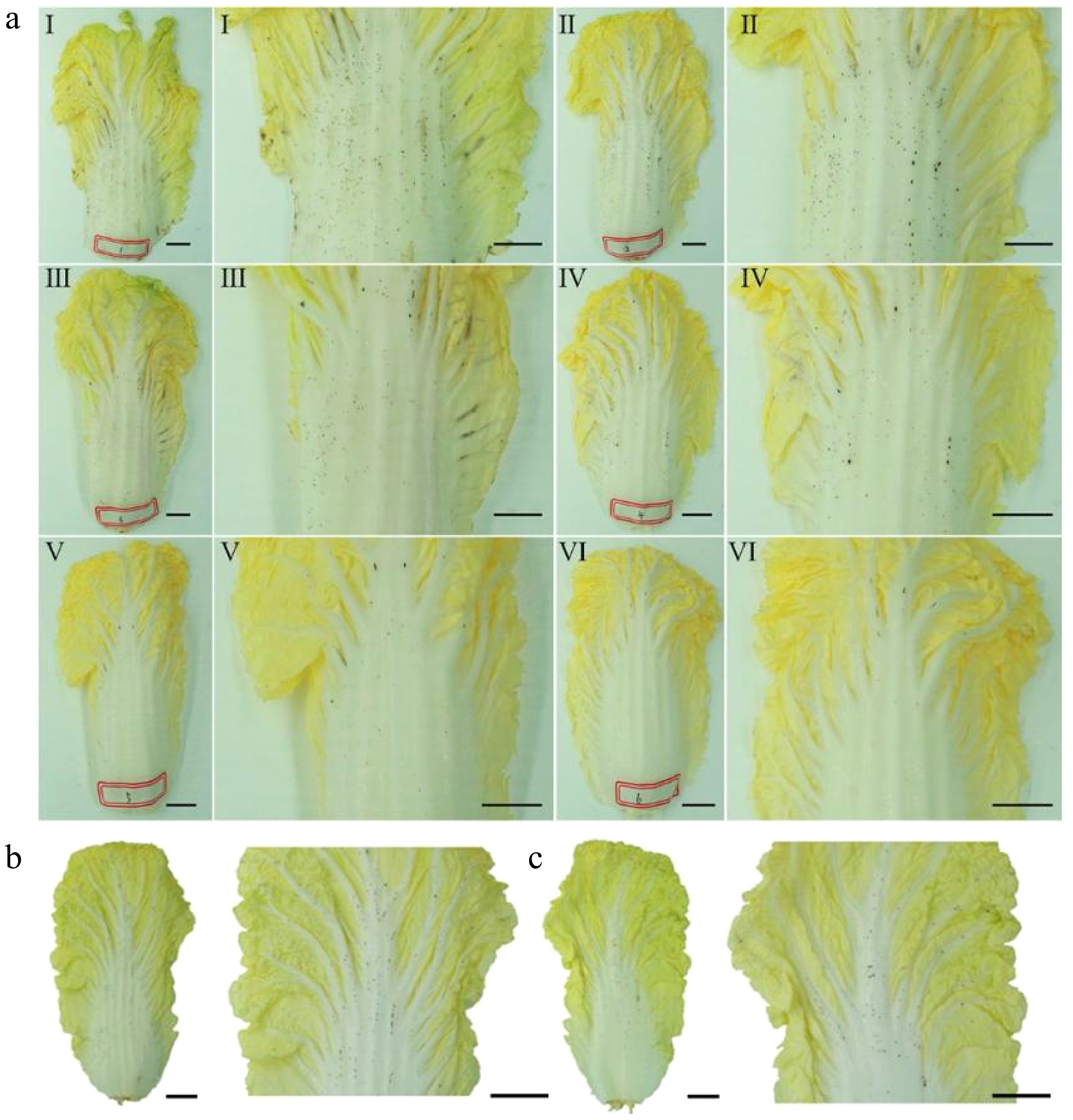
Figure 1.
Phenotypic observation of petiole spot occurrence on Chinese cabbage leaves during storage. (a) During storage, petiole spot severity in Chinese cabbage decreases from outer (I) to inner leaves (VI). No significant difference in petiole spot occurrence was found between the (b) adaxial, and (c) abaxial epidermis of Chinese cabbage leaves.
RNA sequencing and differentially expressed gene analysis
-
Transcriptome analysis was performed on three Chinese cabbage cultivars under two treatments: control (CK) and petiole spot (PS). The sample designations were as follows: Zaoshu No. 8 (CK-1: spot-free, PS-1: spot-affected), Han Chunwa 4-2-3-4 (CK-2: spot-free, PS-2: spot-affected), and Line 163 (CK-3: spot-free, PS-3: spot-affected). Each treatment included three biological replicates, yielding 18 samples in total for sequencing (three cultivars × two treatments × three replicates) (Supplementary Fig. S2). Each replicate was generated by pooling tissues from three plants of the same cultivar (nine plants per treatment group). Sample collection was performed as described in Section “Plant materials”. Data presented in Fig. 2 (CK1–CK3, PS1–PS3) represent the mean of three biological replicates.
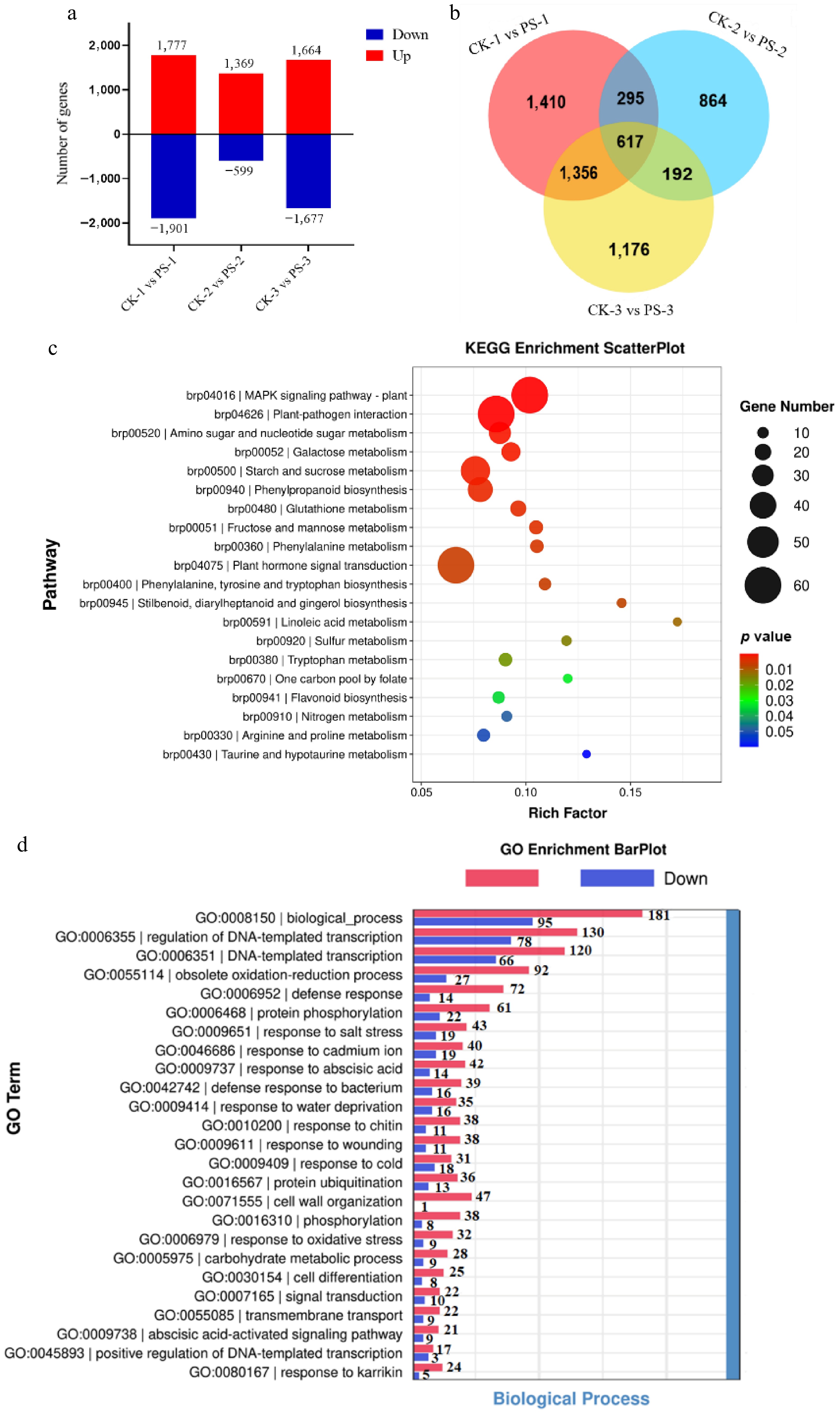
Figure 2.
Transcriptome analysis on leaves with and without petiole spots. (a) Identification of differentially expressed genes (DEGs) in the comparisons of CK-1 vs PS-1, CK-2 vs PS-2, and CK-3 vs PS-3. All the data represent the average of three independent biological replicates. (b) Venn diagram depicting the shared and unique DEGs across the three comparison groups. (c) KEGG pathway enrichment analysis of DEGs between Chinese cabbage leaves without petiole spot symptoms (CK) and those exhibiting petiole spot symptoms (PS). Each bubble represents a metabolic pathway, with bubble size indicating the pathway's impact factor and bubble color intensity reflecting the degree of enrichment. (d) Gene Ontology (GO) enrichment analysis of DEGs between the comparison groups. CK: Chinese cabbage leaves without petiole spot symptoms; PS: Chinese cabbage leaves exhibiting petiole spot symptoms.
Total RNA was extracted from Chinese cabbage petiole tissues using the Spin Column Plant Total RNA Purification Kit (Sangon Biotech, Shanghai, China), following the manufacturer's instructions. RNA quality and concentration were assessed using a spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Double-stranded cDNA libraries were constructed using the EzyNGS DNA Library Construction Kit (Sangon Biotech, Shanghai, China) according to the manufacturer's protocol. Sequencing was performed on an Illumina platform by Biomarker Technologies (Shunyi, Beijing, China). Raw sequencing reads were quality-filtered using Trimmomatic (v3.90) to remove adapter sequences, reads with more than 10% unknown bases (N), and reads with > 50% of bases having a quality score (Q) ≤ 10. Clean reads were aligned to the Brassica rapa reference genome (Brara_chiifu_V3.0) using HISAT2[16], and transcript assembly was performed using StringTie (accessed on January 9, 2023)[17]. Differentially expressed genes (DEGs) were identified using DESeq with the thresholds of adjusted p-value < 0.05 and |log2(fold change)| ≥ 1. Functional enrichment analysis of DEGs was performed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) databases (accessed on January 9, 2023). Gene expression heatmaps were generated using the heatmap function of the TBtools software.
Quantification of endogenous hormones using high-performance liquid chromatography (HPLC)
-
Sample preparation: Approximately 1–2 g of petiole epidermal tissue was ground into a slurry in an ice bath and extracted with 20 mL of pre-chilled 80% methanol. The homogenate was sealed with plastic film and incubated at 4 °C overnight. The extract was filtered, and the mortar was rinsed twice with 10 mL of methanol, which was combined with the initial extract. The combined solution was evaporated under reduced pressure at 40 °C until no methanol remained. The remaining aqueous phase was completely transferred into a conical flask. For pigment removal, the aqueous phase was extracted twice with 30 mL of petroleum ether, and the ether phase was discarded. Then, 0.01 g of polyvinylpolypyrrolidone (PVPP) was added, and the mixture was subjected to ultrasonic treatment for 30 min. After filtration, the aqueous phase was extracted three times with 30 mL of ethyl acetate. The combined ethyl acetate phases were evaporated to dryness under reduced pressure at 40 °C. The residue was dissolved in chromatographic-grade methanol and brought to a final volume of 2 mL. The solution was filtered through a 0.45 µm microporous membrane and stored at 4 °C for HPLC analysis.
HPLC system preparation: The chromatography system was initialized by turning on the computer and all module power supplies. After launching the instrument control software, the flushing valve was manually opened. The pump flow rate was set to 5 mL·min−1 to purge the system until all bubbles were removed from the tubing (approximately 5–10 min). The solvent lines for all channels were flushed sequentially until bubble-free. The flushing valve was then manually closed, and the flow rate was reset to 0 mL·min−1. A new method was created in the software. The mobile phase was configured according to analytical requirements, and the flow rate was set to 1.0 mL·min−1. The detector wavelength was set to 254 nm. After stabilization of the baseline (approximately 10 min), samples were ready for injection and detection.
Standard curve and quantification: Standard solutions of ABA at different concentrations were prepared in chromatographic-grade methanol. Each standard was injected (10–30 µL) manually or via an autosampler, and peak areas were recorded. Calibration curves were established based on the linear relationship between hormone concentration and peak area within a defined concentration range. Sample analysis: Test samples were injected using the same volume, and chromatographic peaks were recorded. After complete elution, the system was flushed for 30 min before the next sample was analyzed. Hormones were identified based on the retention time of known standards. Quantification: The concentrations of ABA in each sample were calculated using the standard curves and the corresponding peak areas of the identified hormone peaks.
Hormone and chemical treatments on Chinese cabbage petiole tissues
-
Detached Chinese cabbage leaves were treated with various compounds to assess their effects on petiole spot formation. On each leaf, three layers of filter paper were placed symmetrically on both sides of the midrib. Then, 20 mM ABA, 50 μM fluridone (FLD), 20 mM hydrogen peroxide (H2O2), 50 μM diphenyleneiodonium (DPI), and a mock control (ddH2O) were each applied dropwise onto the filter papers.
A small amount of water was sprayed onto the leaves to maintain humidity, and the treated leaves were covered with plastic wrap to prevent desiccation and evaporation of reagents. The samples were then placed in a 4 °C refrigerator for 14 d. After the treatment period, the number of petiole spots was recorded for statistical analysis.
Quantitative real-time PCR (qRT-PCR) analysis
-
Total RNA was extracted from the petiole epidermis of nine Chinese cabbage samples using the Spin Column Plant Total RNA Purification Kit (Sangon Biotech, Shanghai, China) following the manufacturer's instructions. First-strand cDNA was synthesized from the extracted RNA using the PrimeScript™ 1st Strand cDNA Synthesis Kit (Takara, Japan). Quantitative real-time PCR was performed on a Roche LightCycler® 96 system using SYBR Green chemistry.
Each reaction was carried out in triplicate to ensure reproducibility. Relative gene expression levels were calculated using the 2−ΔCᴛ method when different tissue parts were compared, with Actin used as the internal reference gene for normalization. When CK was included as the reference, the 2−ΔΔCᴛ method was applied. All primers used in this study were synthesized by Beijing Tsingke Biotechnology Co., Ltd, and their sequences are listed in Supplementary Table S1.
Detection of intracellular ROS by H2DCFDA fluorescent staining
-
For the detection of the intracellular ROS level, ROS-sensitive probe H2DCFDA (MCE, Princeton, NJ, USA) was used[18]. The epidermis of Chinese cabbage leaves was peeled using tweezers, then immersed in an MES-KCl buffer (MES 10 mM, KCl 5 mM, CaCl2 50 mM, pH 6.15) for 30 min. Subsequently, dark staining was performed with 50 mM H2DCFDA for 2 h. After rinsing three times with PBS (MCE, Princeton, NJ, USA), the ROS staining was observed using a fluorescence microscope (Leica DM2500 LED, Wetzlar, Germany) with GFP fluorescence.
Statistical analysis
-
All experiments in this study were conducted with three biological replicates. KEGG annotation and enrichment analyses were performed to assess the statistical overrepresentation of differentially expressed genes (DEGs) in specific KEGG pathways. Statistical significance was evaluated using Student's t-test, and Pearson correlation coefficients were calculated using SPSS software (version 24.0; SPSS Inc., Chicago, IL, USA). Differences were considered statistically significant at p ≤ 0.05.
-
During storage, petiole spots develop de novo on initially symptom-free tissues, appearing as dense black lesions (1–2 mm in diameter) on the petiole. As all three varieties exhibited similar petiole spot symptoms (Supplementary Fig. S1), Zaoshu No. 8 was selected as the representative to illustrate the phenotype in Fig. 1. Notably, the number of petiole spots gradually decreases from the outer leaves to the inner leaves. Near the soft leaf region, the spots are concentrated on the veins, while the soft leaves show no obvious spots (Fig. 1a). In addition, small black spots are present on both the inner and outer epidermis (Fig. 1b, c). Collectively, the observations reveal a distinct spatial pattern of petiole spots in stored Chinese cabbage.
Spot lesion-specific transcriptomes in postharvest Chinese cabbage
-
To elucidate the molecular mechanisms underlying petiole spot formation during storage, RNA-seq analysis was performed on petiole tissues with and without petiole spots from three Chinese cabbage varieties susceptible to the disorder. Samples without visible petiole spots were designated as CK-1, CK-2, and CK-3, while those exhibiting petiole spots were labeled as PS-1, PS-2, and PS-3. RNA libraries were constructed and sequenced on the Illumina NovaSeq 6000 platform, generating 150 bp paired-end reads. A total of 34.73 GB of clean data was obtained across the six samples, with each sample yielding approximately 5.12–6.15 GB of valid bases. The number of raw reads per sample ranged from 35.4 to 41.0 million, and valid reads ranged from 34.1 to 40.0 million, with a valid ratio between 96.09% and 97.59%. The Q20 percentages for all samples exceeded 99.69%, and the Q30 percentages ranged from 97.93% to 98.29%, indicating high sequencing accuracy. Clean reads were aligned to the Brassica rapa reference genome (v3.0) using HISAT2 (v2.2.1), and transcript assembly and expression quantification were performed using StringTie (v2.2.1). A principal component analysis (PCA) was conducted based on FPKM values to evaluate the consistency among biological replicates and to distinguish transcriptomic differences between the spot and non-spot groups (Supplementary Fig. S2) Differential expression analysis revealed 1,777, 1,369, and 1,664 up-regulated genes, along with 1,901, 599, and 1,677 down-regulated genes in the CK-1 vs PS-1, CK-2 vs PS-2, and CK-3 vs PS-3 comparisons, respectively (Fig. 2a). The Venn diagram reveals that 617 differentially expressed genes are common across the comparisons CK-1 vs PS-1, CK-2 vs PS-2, and CK-3 vs PS-3 (Fig. 2b).
KEGG analysis was conducted on the commonly differentially expressed genes across sample groups. The results revealed that these genes were predominantly enriched in the 'Plant hormone signal transduction' pathway, followed by the 'MAPK signaling pathway' and 'Plant-pathogen interaction' pathway (Fig. 2c). Interestingly, GO enrichment analysis revealed that 42 upregulated and 14 downregulated genes were associated with the 'response to abscisic acid' category, while 21 upregulated and nine downregulated genes were linked to the 'abscisic acid-activated signaling pathway' (Fig. 2d). In summary, genes associated with plant hormone signal transduction displayed distinct expression patterns in the epidermal cells of Chinese cabbage with and without petiole spot, suggesting that hormonal regulation may contribute to the phenotypic variations observed in petiole spot development during storage.
Analysis of the ABA biosynthesis and conjugation pathway in stored Chinese cabbage
-
To further investigate the role of ABA in petiole spot formation during storage, differential gene expression was performed, and 19 genes that were highly correlated with ABA at the transcriptomic level (Pearson correlation coefficient > 0.8, p-value < 0.05) were identified. These genes include two zeaxanthin epoxidase (ZEP) genes (BraA07g016880.3C, BraA09g009220.3C), six 9-cis-epoxycarotenoid dioxygenase (NCED) genes (BraA05g032610.3C, BraA03g036470.3C, BraA01g010560.3C, BraA01g037270.3C, BraA02g024670.3C, BraA03g048210.3C), one abscisic aldehyde oxidase (AAO3) gene (BraA04g019480.3C), two β-glucosidase (BG1) genes (BraA06g002000.3C, BraA05g017770.3C), three β-glucosidase (BG2) genes (BraA04g003380.3C, BraA09g0047910.3C, BraA04g003370.3C), and one UDP-glucosyltransferase (UGT71CS) gene (BraA10g005910.3C). These genes are involved in ABA biosynthesis and conjugation, playing crucial roles in its metabolic regulation. Gene function analysis revealed significant differences in the expression profiles of genes associated with ABA biosynthesis and conjugation between petiole spot-affected and unaffected Chinese cabbage tissues (Fig. 3).
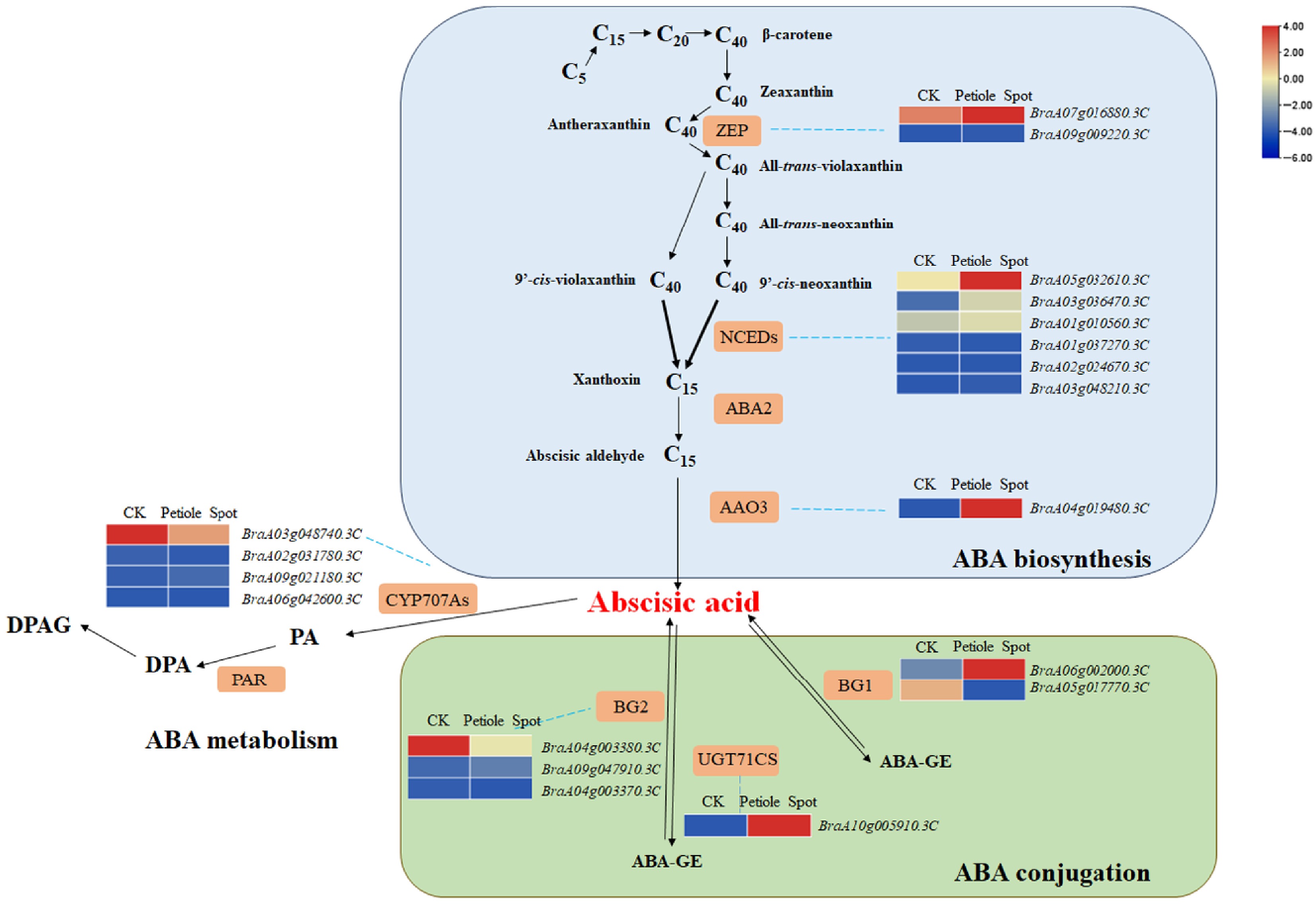
Figure 3.
Heat map illustrating the expression profiles of genes associated with ABA biosynthesis and metabolic pathways.
In the ABA biosynthesis pathway, the expression of BraA07g016880.3C and BraA09g009220.3C (ZEP) showed distinct regulatory patterns between CK and petiole spot-affected tissues. Notably, several NCED genes (BraA05g032610.3C, BraA03g036470.3C, BraA01g010560.3C, BraA01g037270.3C, BraA02g024670.3C, BraA03g048210.3C) exhibited significantly elevated expression in petiole spot-affected tissues, suggesting increased ABA biosynthetic activity. Similarly, the upregulation of BraA04g019480.3C (AAO3) in diseased tissues further supports enhanced ABA biosynthesis during petiole spot formation. Regarding ABA conjugation, genes encoding BG1 (BraA06g002000.3C, BraA05g017770.3C) and BG2 (BraA04g003380.3C, BraA09g0047910.3C, BraA04g003370.3C) displayed distinct expression patterns, indicating their involvement in regulating ABA homeostasis. Furthermore, BraA10g005910.3C (UGT71CS), a key gene responsible for ABA glucosylation, was significantly upregulated in petiole spot-affected tissues compared to CK, suggesting enhanced ABA conjugation in response to petiole spot development. These findings highlight the critical role of ABA biosynthesis and conjugation pathways in the formation of petiole spots during storage. The upregulation of key biosynthetic genes, particularly NCEDs and AAO3, along with the altered expression of conjugation-related genes, underscores the importance of ABA metabolism in the development of petiole spots in Chinese cabbage.
Endogenous hormone elevated in petiole spot lesions of stored Chinese cabbage
-
Considering the potential involvement of plant hormones in the development of petiole spots during storage, the study quantified the levels of various hormones in the leaves of cultivar Zaoshu No. 8, comparing those with and without petiole spot symptoms (CK vs PS). The results revealed a significant accumulation of ABA and ABA-GE in the epidermal cells of petiole spot-affected tissues compared to unaffected tissues (Fig. 4a). qRT-PCR was performed to examine the expression patterns of key genes associated with ABA biosynthesis and conjugation in petiole spot-affected and control Chinese cabbage tissues (Fig. 4b). The results demonstrated a significant upregulation of BraA07g016880.3C (ZEP), BraA01g010560.3C, and BraA03g048740.3C (NCED), as well as BraA04g019480.3C (AAO3) in petiole spot-affected tissues, indicating an enhanced ABA biosynthetic activity. Additionally, genes involved in ABA conjugation, including BraA06g002000.3C (BG1) and BraA10g005910.3C (UGT71CS), also exhibited increased expression in diseased tissues. These findings further support the transcriptome analysis, reinforcing the critical role of ABA metabolism in the development of petiole spot during storage.
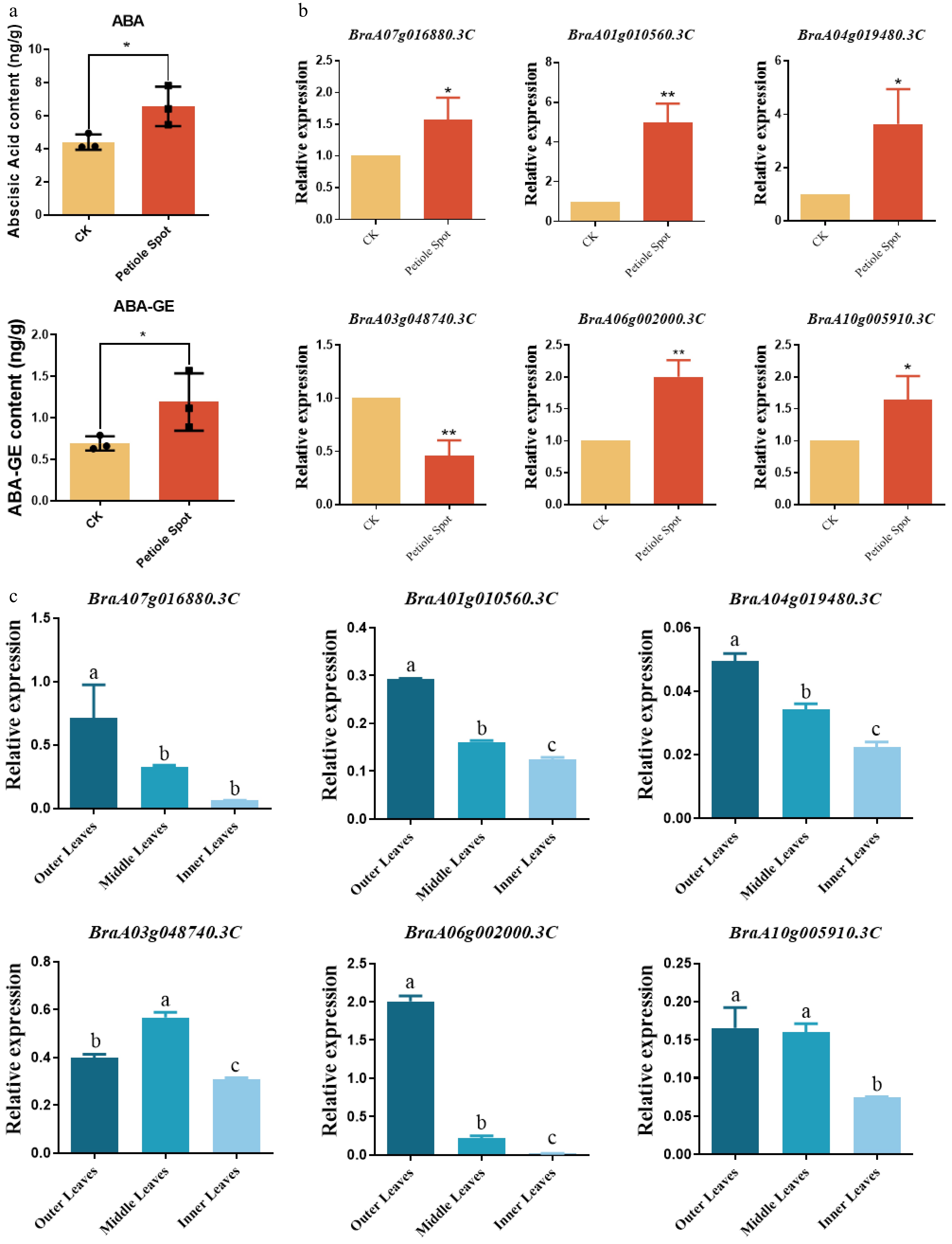
Figure 4.
Expression analysis of ABA-related genes in cultivar Zaoshu No. 8. (a) The concentrations of ABA and its conjugated form GA-ABA were significantly elevated in the epidermis of Chinese cabbage leaves exhibiting petiole spot symptoms compared to symptom-free control leaves (CK). (b) Quantitative PCR (qPCR) analysis of key ABA pathway genes in Chinese cabbage leaves with and without petiole spot symptoms. The relative expression levels analyzed by qRT-PCR and calculated by 2−ΔCᴛ and bars with different lowercase letters are significantly different (* < 0.05, ** < 0.01). (c) qPCR analysis of key ABA pathway genes in the epidermis of outer, middle, and inner leaf layers. The relative expression levels analyzed by qRT-PCR and calculated by 2−ΔΔCᴛ and bars with different lowercase letters are significantly different (p < 0.05).
To further explore the relationship between ABA metabolism and petiole spot formation during storage, qRT-PCR was performed to analyze the expression patterns of key genes involved in ABA biosynthesis and conjugation across different leaf positions (outer, middle, and inner leaves) (Fig. 4c). The results showed that the expression levels of BraA07g016880.3C (ZEP), BraA01g010560.3C, BraA04g019480.3C (AAO3), and BraA03g048740.3C (NCED) exhibited a decreasing trend from the outer to the inner leaves, consistent with the observed reduction in petiole spot incidence. Similarly, BraA06g002000.3C (BG1), a key gene in ABA conjugation, showed significantly higher expression in the outer leaves compared to the middle and inner leaves. Additionally, BraA10g005910.3C (UGT71CS) expression was significantly lower in the inner leaves, further indicating a potential role in ABA homeostasis regulation. Together, the spatial distribution of petiole spot formation is closely associated with the differential regulation of ABA biosynthesis and conjugation, suggesting the elevated ABA metabolic activity observed in the outer leaves may contribute to their heightened susceptibility to petiole spot during storage.
ABA promotes petiole spot development in Chinese cabbage during storage
-
To further validate the impact of ABA on petiole spot development during Chinese cabbage storage, 20 μM ABA and ddH2O patches were applied to both sides of the same leaf, using three layers of filter paper. The results revealed that ABA-treated areas exhibited more pronounced petiole spot formation, supporting the role of ABA in enhancing petiole spot development (Fig. 5a, c). In contrast, treatment with the ABA synthesis inhibitor (FLD) effectively suppressed petiole spot occurrence, as indicated by the reduced symptom severity in FLD-treated areas compared to the mock and ABA-treated ones (Fig. 5b, d). These phenotypic observations provide strong evidence that ABA plays a crucial role in the development of the petiole spot during storage.
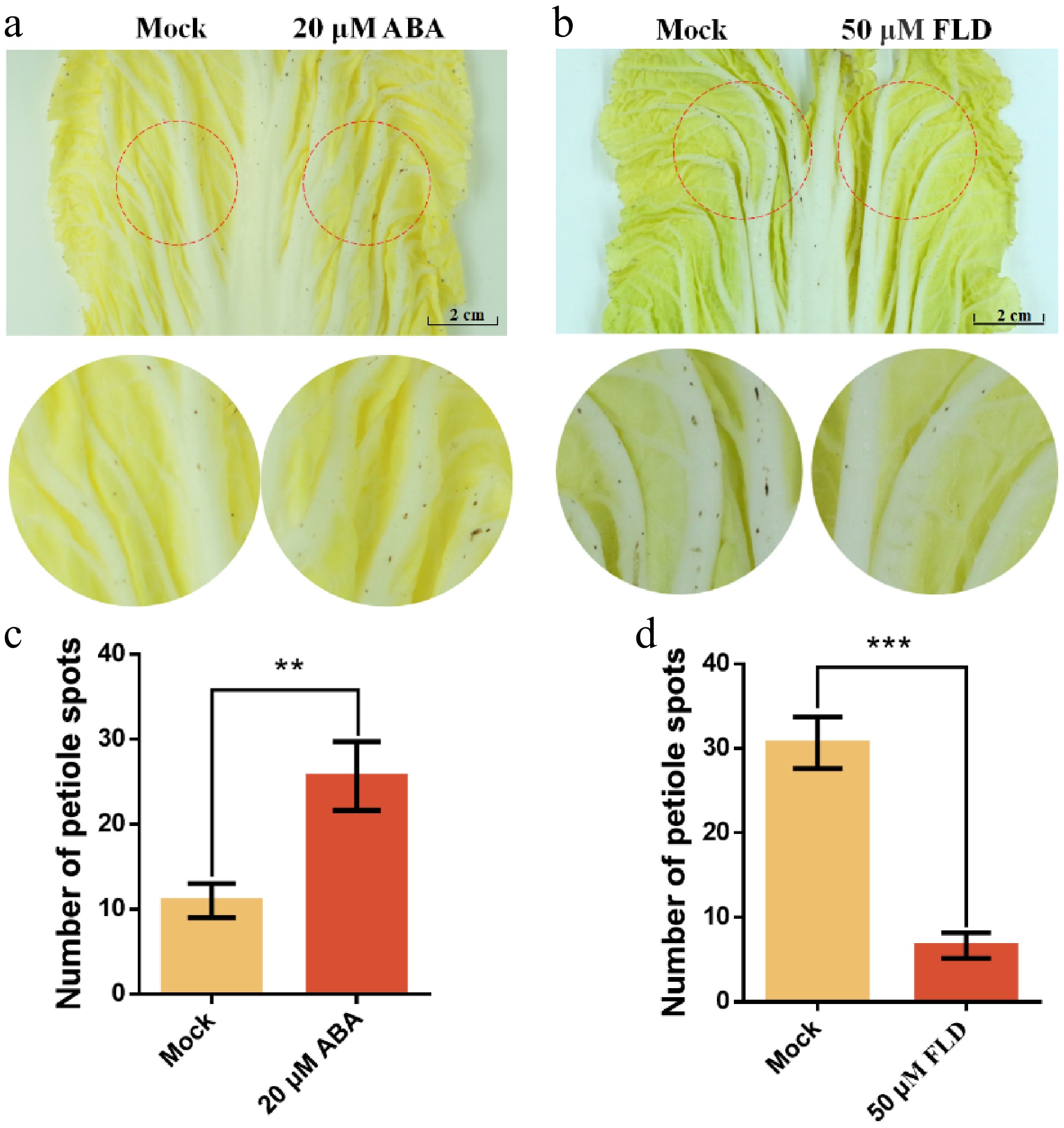
Figure 5.
Exogenous ABA promotes the development of petiole spot symptoms in Chinese cabbage cultivar Zaoshu No. 8. (a) ddH2O (Mock) and 20 μM ABA were applied to symmetrically corresponding regions on the same leaf. (b) ddH2O (Mock) and 50 μM of the ABA biosynthesis inhibitor fluridone (FLD) were applied to symmetrically corresponding regions on the same leaf. (c), (d) The number of petiole spots within the filter paper-covered area was recorded. Each treatment group consisted of three cabbage leaves, and all experiments were independently repeated three times. ** indicates p < 0.01, and *** indicates p < 0.001.
H2O2 could mimic ROS-induced petiole spot occurrence during storage
-
Previous studies have demonstrated increased intracellular ROS levels in petiole spot-affected cells. To elucidate the role of ROS in petiole spot formation during storage, H2DCFDA fluorescence staining was conducted, and the results revealed a significant elevation in green fluorescence intensity within petiole spot-affected regions and their adjacent cells compared to unaffected cells, indicating increased ROS accumulation in petiole spot tissues (Fig. 6a).
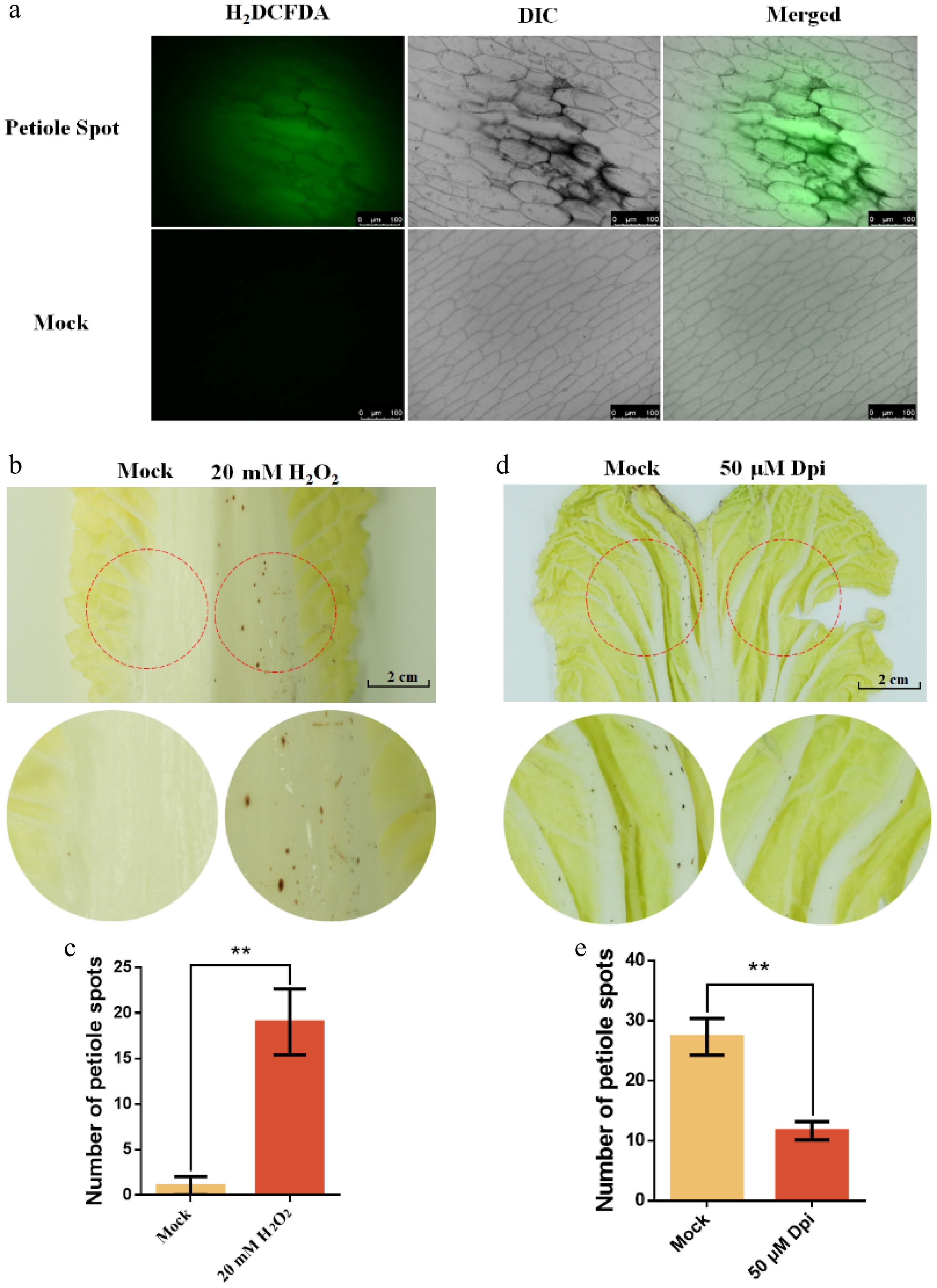
Figure 6.
Exogenous H2O2 mimics ROS-induced petiole spot formation in Chinese cabbage cultivar Zaoshu No. 8. (a) Reactive oxygen species (ROS) accumulation in the epidermis was assessed using H2DCFDA staining. ddH2O (Mock) was used as a negative control, while 20 mM H2O2 served as a positive control. (b), (c) ddH2O (Mock) and 20 mM H2O2 were applied to symmetrically positioned regions on the same leaf, and the number of petiole spots within the filter paper-covered area was quantified. (d), (e) ddH2O (Mock) and 50 μM of the ROS synthesis inhibitor diphenyleneiodonium (Dpi) were applied to symmetrically positioned regions on the same leaf, and the number of petiole spots within the filter paper-covered area was quantified. Each treatment group consisted of three cabbage leaves, and all experiments were independently repeated three times. ** indicates p < 0.01.
To further confirm the involvement of ROS in petiole spot development during storage, H2O2 was applied to mimic ROS induced petiole spot formation. The results showed that 20 mM H2O2-treated regions exhibited more severe petiole spot formation, indicating H2O2 could mimic ROS-induced petiole spot progression (Fig. 6b, c). In contrast, application of the H2O2 synthesis inhibitor DPI effectively mitigated petiole spot occurrence, as evidenced by the reduced symptom severity in DPI-treated regions compared to the mock and H2O2-treated areas (Fig. 6d, e). These findings provide strong evidence that ROS plays a pivotal role in the regulation of petiole spot development during the storage of Chinese cabbage.
ABA modulates ROS-mediated petiole spot development in postharvest Chinese cabbage
-
To further investigate the role of ABA in petiole spot development during storage, 20 mM H2O2, ddH2O, 20 mM ABA, and 50 μM FLD were applied to different regions of the same Chinese cabbage leaf using three layers of filter paper. The results demonstrated that H2O2 treatment significantly promoted petiole spot formation, with more severe symptoms observed in the affected areas compared to the mock treatment. Furthermore, co-application of ABA and H2O2 exacerbated the symptoms, suggesting a synergistic effect of ABA and ROS in promoting petiole spot development (Fig. 7a, c).
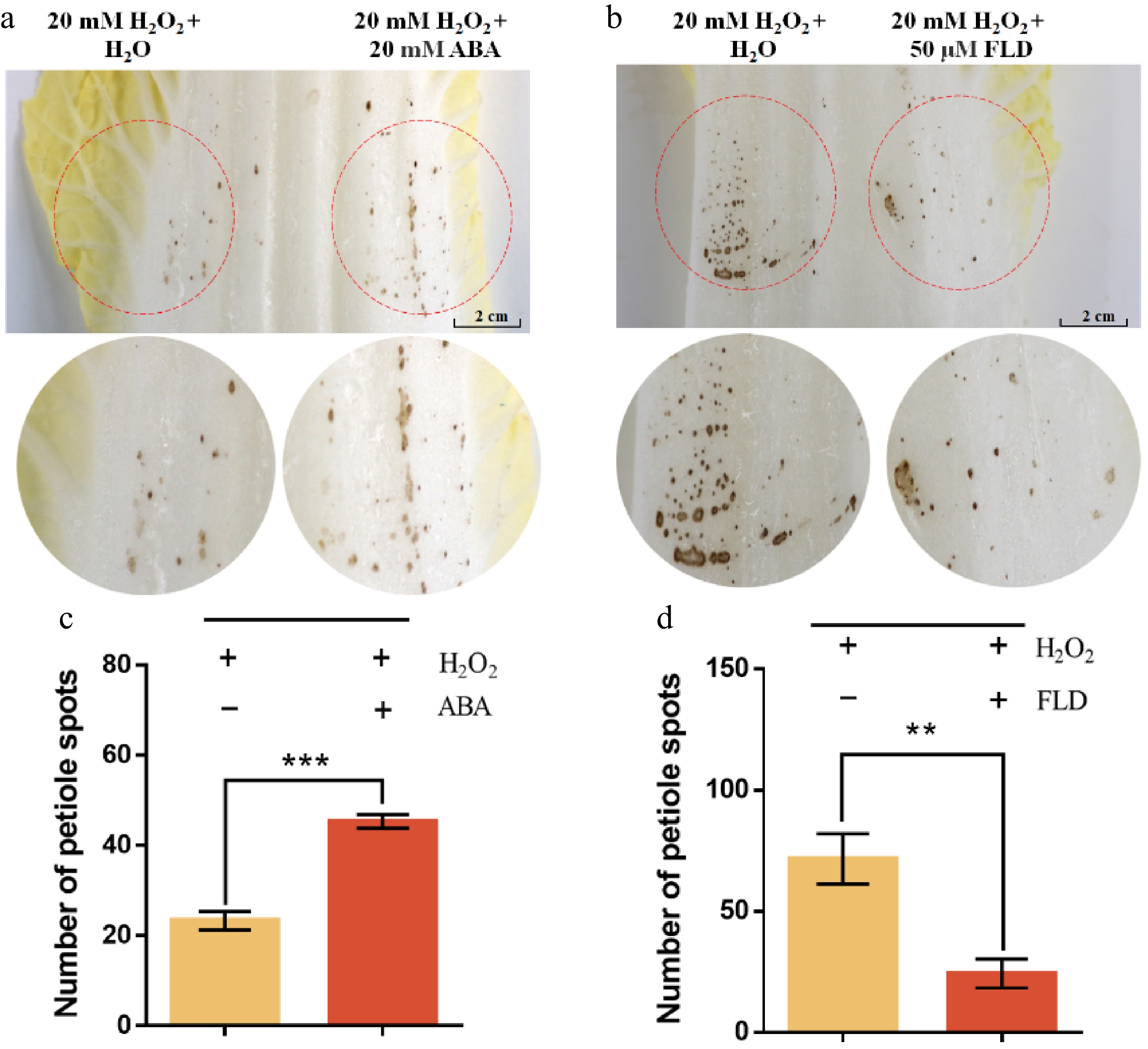
Figure 7.
ABA enhances and FLD suppresses H2O2-mimic ROS-induced petiole spot formation in Chinese cabbage cultivar Zaoshu No. 8 during cold storage. (a) Visual comparison of petiole spot formation after localized application of 20 mM H2O2 alone or in combination with 20 mM abscisic acid (ABA) on Chinese cabbage petioles. Red dashed circles indicate treated areas. (b) Visual comparison of petiole spot formation following co-treatment with 20 mM H2O2 and 50 μM fluridone (FLD, an ABA biosynthesis inhibitor). (b) Quantification of petiole spot numbers from (a), showing that co-treatment with ABA significantly increased the number of spots compared to H2O2 alone (*** p < 0.001). (d) Quantification of petiole spot numbers from (c), demonstrating that FLD significantly reduced H2O2-induced spot formation (** p < 0.01). Data represent means ± SE of three biological replicates. Statistical significance was assessed using Student's t-test.
In contrast, treatment with the ABA synthesis inhibitor FLD effectively suppressed petiole spot formation, as indicated by the reduced symptom severity in FLD-treated regions compared to the H2O2-only treatment. These findings suggest that ROS accumulation plays a crucial role in petiole spot formation, and ABA further enhances its effects, while FLD mitigates ROS-induced petiole spot development during storage (Fig. 7b, d).
-
Chinese cabbage is one of the most widely cultivated leafy vegetables globally and can be supplied all year round[19]. The year-round availability of cabbage is due to the flexibility of its planting seasons and its good storability[20,21]. However, the postharvest storage of Chinese cabbage is often accompanied by the development of petiole spot, which not only progresses severely but also newly emerges in previously asymptomatic tissues[22]. Microscopic analysis reveals that the petiole spot in stored Chinese cabbage initiates as inter- or extracellular lesions before spreading to neighboring cells (Supplementary Fig. S1), which is in line with the characteristics of the petiole spots induced by excessive nitrogen fertilizer[11], suggesting that storage-induced and nitrogen-induced petiole spots may share common cellular mechanisms. The distinct gradient distribution of petiole spot in storage is consistent with observations under nitrogen-excess field conditions, where lesions follow a similar gradient within the head leaves, suggesting that petiole spot formation may be influenced by leaf age. Notably, the outer leaves—located at the head's periphery, and uninvolved in tight head formation—exhibit no petiole spots, even though they are the oldest. This paradox suggests that petiole spot formation is governed by complex factors beyond leaf age. It is well studied that petiole spot development is correlated with nitrogen excess[11], however, this triggering factor is absent in the storage environment. Thus, the mechanisms underlying petiole spot formation in stored Chinese cabbage warrant in-depth investigation.
Compared to pathogen-induced diseases, the underlying mechanisms of physiological disorders in Chinese cabbage remain relatively poorly understood[23−26]. Among these, petiole spots and tipburn are two common physiological disorders, both arising from multiple factors. Petiole spots are primarily associated with excessive nitrogen fertilization, whereas tipburn is generally attributed to calcium (Ca2+) deficiency[27]. Recently, significant progress has been made in elucidating the molecular mechanisms of tipburn development and resistance[27,28]. Genome-wide association studies and linkage analysis have identified a common tipburn-associated locus, within which BrCRT2 was highlighted for its elevated expression during tipburn progression. Functional analyses confirmed BrCRT2 as a strong candidate gene for tipburn resistance in Chinese cabbage[27]. Moreover, gene silencing, overexpression, and heterologous expression studies demonstrated that the cyclic nucleotide-gated ion channel (CNGC) genes BrCNGC12 and BrCNGC16 are key regulators of calcium uptake and translocation, playing pivotal roles in conferring tipburn resistance[28]. In contrast, the genetic basis of petiole spot formation remains largely unexplored. Transcriptome analysis is an indispensable tool that provides fundamental insights into the pathways potentially involved in biological processes, as well as into gene function and interactions, thereby facilitating advancements in scientific research. Transcriptomic analysis of Chinese cabbage grown under normal and calcium-deficient conditions revealed significant enrichment of genes involved in hormone and calcium signaling pathways. Notably, BrCNGC12 and BrCNGC16, key components of the calcium signaling pathway, were selected for further investigation[28]. Similarly, KEGG analysis of the transcriptomes from tissues with and without petiole spots revealed significant enrichment in hormone signaling pathways, while GO analysis showed direct enrichment in abscisic acid (ABA)-related processes. ABA can be stored in the form of glycosylated ABA-GE, a reaction mediated by UDP-glucosyltransferases (UGTs). This conjugated form can be quickly converted back into active ABA through the action of β-glucosidase[29−31]. Subsequent endogenous hormone quantification, exogenous hormone application, and the use of a chemical inhibitor, combined with gene expression analysis together, confirmed that ABA is involved in petiole spots development (Figs. 4−7). ABA is a well-established phytohormone integral to plant stress responses[32,33]. Beyond stress adaptation, ABA regulates plant growth and development, orchestrating key biological processes during seed maturation, including endosperm and embryo development, storage compound accumulation, acquisition of desiccation tolerance, and dormancy establishment[34−37]. In climacteric fruits, ABA coordinates ethylene biosynthesis and signaling to initiate ripening[38−40]. Critically, as a primary signaling molecule induced by water stress, ABA triggers physiological disorders such as husk scald in stored pomegranate[41−43]. Similarly, the experimental data demonstrate that ABA induces the formation of petiole spots, a physiological disorder occurring during the storage of Chinese cabbage, consistent with previously reported mechanisms in postharvest pathology[44]. Although the involvement of the ABA signaling pathway in petiole spot formation has been established, the specific gene within this pathway that plays the most critical regulatory role remains unidentified. Further research is required to pinpoint this key gene, which may serve as a valuable target for future breeding and genetic improvement strategies.
ROS, long recognized as mediators of abiotic stress damage, also appear to be intricately linked to petiole spot formation[11,14,45]. It is reported that petiole spots are associated with cell death triggered by ROS bursts resulting from excessive accumulation of NH4+. Similarly, petiole spot formation during storage is accompanied by the accumulation of ROS. Crucially, elevated ABA levels may drive ROS bursts in stored Chinese cabbage. In the H2O2-mimic petiole spots formation assay, ABA exacerbates ROS-induced petiole spots formation, whereas the ABA biosynthesis inhibitor FLD mitigates spot development (Fig. 7). These findings support the hypothesis that ABA-induced ROS bursts mediate petiole spot formation during storage. Notably, ABA plays a critical role in oxidative stress responses by regulating the generation and detoxification of ROS, thus maintaining cellular redox homeostasis[35−37]. Under drought conditions, ABA accumulation triggers stomatal closure and activates antioxidant defense mechanisms, thereby minimizing water loss and mitigating oxidative damage. During salt stress, ABA enhances plant tolerance by regulating ion homeostasis and inducing the expression of genes involved in osmotic adjustment[46]. Extensive research establishes ABA as a key regulator of ROS generation under multiple stress conditions[47,48]. This signaling function manifests in several physiological processes. Under drought stress, ABA triggers stomatal closure through ROS accumulation in guard cells[49,50]. ABA activates respiratory burst oxidases (RBOHs) to produce ROS essential for pollen tube germination. During nitrogen deficiency, elevated leaf ABA enhances carbon translocation from senescing leaves to developing grains[51,52]. In summary, ABA plays a dual role in regulating ROS homeostasis in plants, with the ability to either promote ROS generation or inhibit its accumulation, depending on the environmental context. In addition to ABA, other hormone signaling pathways are also pivotal in the development of physiological disorders. Aberrant expression of genes in the auxin signaling pathway has been shown to induce tipburn symptoms in Brassica oleracea[53]. In Chinese cabbage, calcium deficiency down-regulates genes involved in auxin and brassinosteroid signaling pathways, while upregulating those associated with ABA, jasmonic acid, and salicylic acid signaling, suggesting that multiple hormones may be involved in tipburn development[53]. In contrast, the role of these hormone pathways—either individually or in combination—in the formation of petiole spots remains largely unclear and warrants further in-depth investigation.
Collectively, these results suggest that ABA promotes petiole spot formation not only via classical abiotic stress pathways but also by modulating ROS-related signaling. The co-occurrence of elevated ABA levels and ROS accumulation in lesion tissues supports a synergistic model in which ABA amplifies oxidative stress, accelerating cell death and lesion expansion. Given that ROS production is a common response to both hormonal and environmental cues, ABA may act upstream to potentiate ROS generation and tissue deterioration. This interaction likely constitutes a critical regulatory node in petiole spot pathogenesis. Further supporting this model, co-treatment experiments involving H2O2 with either exogenous ABA or FLD revealed that ABA significantly exacerbates, whereas FLD attenuates, H2O2-induced lesion development. These findings reinforce the hypothesis that ABA and ROS act in concert to drive petiole spot formation and highlight the importance of their crosstalk in the progression of this postharvest disorder. Despite these findings, numerous fundamental questions related to the formation and regulation of petiole spots remain unanswered. Transcriptome enrichment analysis also revealed that the MAPK signaling pathway and the plant–pathogen interaction pathway were significantly enriched among DEGs, suggesting their potential involvement in the development of this physiological disorder. To date, the mechanism by which plants perceive excessive nitrogen and transduce this signal to trigger ROS burst and ultimately PCD, leading to the manifestation of petiole spots, remains largely unknown. Whether the MAPK signaling cascade participates in this process requires further investigation. Although petiole spots are not associated with pathogenic infection, the enrichment of the plant–pathogen interaction pathway raises the intriguing possibility that this disorder may reflect an endogenous immune-like response of Chinese cabbage to abiotic stress, potentially induced by nitrogen overload. This hypothesis warrants detailed functional validation. Moreover, the unpredictable spatial distribution of petiole spots and the possible mechanistic differences between their occurrence during plant growth and postharvest storage remain poorly understood. These aspects merit comprehensive exploration to fully elucidate the underlying regulatory networks. In parallel, genetic dissection of petiole spot susceptibility through population-level mapping approaches is urgently needed to identify key regulatory loci and inform future breeding strategies aimed at improving postharvest quality and stress resilience in Chinese cabbage.
-
In this study, it was found that petiole spot symptoms in stored Chinese cabbage intensified from outer to inner leaves, indicating a clear spatial and temporal progression. Comparative transcriptomic analysis revealed that differentially expressed genes were significantly enriched in hormone signaling genes, especially in ABA biosynthesis and metabolism, which exhibit distinct expression patterns between healthy and lesioned tissues. Notably, the concentration of ABA and its conjugate ABA-GE was significantly higher in affected regions. Further exogenous hormone treatments confirmed that ABA significantly promoted the occurrence of petiole spots during storage. Conversely, an ABA biosynthesis inhibitor effectively suppressed spot formation, demonstrating the crucial involvement of ABA in this physiological disorder in the storage stage. Interestingly, ABA also enhanced H2O2-mimic petiole spot development, whereas FLD mitigated the symptoms, suggesting a synergistic interaction between ABA and ROS in petiole spot formation. Together, these findings demonstrate a central role for ABA–ROS crosstalk in the development of the petiole spot in stored Chinese cabbage and provide new insights for potential postharvest intervention strategies.
This work was supported by the National Key Research and Development Program of China (Grant No. 2023YFD1201504-5), Zhejiang province Major Science and Technology Program (Grant No. 2021C02065-5-2).
-
The authors confirm contributions to the paper as follows: study conception and design: Zhao Y, Li B; data collection: Mei Y, Yue Z, Sun H, Hu Q; analysis and interpretation of results: Mei Y, Tao P, Lei J; draft manuscript preparation: Yue Z, Sun H, Mei Y, Zhao Y. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Zhichen Yue, Haifeng Sun
- Supplementary Table S1 List of primers used for qPCR analysis.
- Supplementary Fig. S1 Phenotypic characterization of three Chinese cabbage cultivars susceptible to petiole spot. Zaoshu No. 8 (right), Line 163 (middle) and Han Chunwa 4-2-3-4 (left) .
- Supplementary Fig. S2 Principal component analysis (PCA) of 18 transcriptome samples from three Chinese cabbage cultivars under CK and PS conditions. Each point represents one biological replicate.
- Supplementary Fig. S3 Microscope image of petiole spots of various sizes on stored Chinese cabbage.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yue Z, Sun H, Hu Q, Mei Y, Lei J, et al. 2025. ABA-mediated ROS burst contributes to petiole spot formation in stored Chinese cabbage. Vegetable Research 5: e047 doi: 10.48130/vegres-0025-0040 

ABA-mediated ROS burst contributes to petiole spot formation in stored Chinese cabbage
- Received: 30 June 2025
- Revised: 08 September 2025
- Accepted: 19 September 2025
- Published online: 01 December 2025
Abstract: Chinese cabbage (Brassica rapa ssp. pekinensis) is one of the most widely consumed vegetables and is commonly stored postharvest. However, a physiological disorder known as petiole spot often develops during storage, significantly impairing the visual quality and marketability of the produce. Despite its economic impact, the underlying mechanisms of petiole spot formation during storage remain poorly understood. In this study, it was observed that petiole spots primarily occur in the outer leaf petioles and gradually diminish toward the inner leaves during storage. Lesions are largely restricted to the petiole, with minimal occurrence in the lamina tissue. Transcriptomic and physiological analyses revealed that genes involved in abscisic acid (ABA) biosynthesis and metabolism exhibit distinct expression patterns between healthy and lesioned tissues. Notably, the concentration of ABA and its conjugate ABA-glucose ester (ABA-GE) was significantly higher in affected regions. Further assays confirmed the role of ABA in symptom development: exogenous ABA application exacerbated lesion formation, while treatment with the ABA biosynthesis inhibitor fluridone (FLD) effectively suppressed symptom severity. Interestingly, ABA also enhanced H2O2-mimic petiole spot development, whereas FLD mitigated the symptoms, suggesting a synergistic interaction between ABA and reactive oxygen species (ROS) in petiole spot formation. Together, these findings demonstrate a central role for ABA–ROS crosstalk in the development of the petiole spot in stored Chinese cabbage and provide new insights for potential post-harvest intervention strategies.
-
Key words:
- Chinese cabbage /
- Physiological disorder /
- Abscisic acid /
- Reactive oxygen species /
- Postharvest