









-
During their life cycle, plants often experience undesirable abiotic stresses, such as salinization, heat, waterlogging, and drought. It has been reported that approximately one-third of the cultivated area around the world suffers from a constantly inadequate supply of water[1,2]. Continuous drought can cause economic and financial problems for both agricultural producers and consumers. The drought in America damaged many of the crops including specialty crops, causing price increases of crop products[3]. The Western United States, including California, has experienced extreme drought stress in recent years. For example, in 2013, the 12-month accumulated precipitation was less than one-third of the average precipitation from 1895−2013 in California, which greatly impacted agricultural irrigation due to low rainfall and water deficiency[4]. Therefore, increasing plants' ability to survive under drought stress conditions is of great importance to the horticultural industry.
Most of the previous studies aiming to improve plants' drought tolerance focus on the biosynthetic and signaling pathways of abscisic acid (ABA). ABA accumulates under osmotic, drought and salt stress conditions and plays crucial roles in the tolerance and response to drought, salinity, and cold stresses[5,6]. Recent research also demonstrated that the absence of the phytohormone gibberellic acid (GA) or a lack of GA sensitivity increased plants' drought resistance[7,8]. GA is a plant hormone that plays important roles in the regulation of plant growth and development including seed development and germination, and diverse organ (e.g. leaves, stem, flower and fruit) elongation and expansion[9,10]. Studies on a number of the GA-deficient mutants in various plants, such as Arabidopsis, maize, rice, pea, and tomato, suggested that decreased levels of GA in the mutants consequently caused typical GA-deficient dwarf phenotypes and the enhancement of stress tolerance[8,11].
In the GA signaling pathway, GA first binds to the soluble GA receptor GA INSENSITIVE DWARF 1 (GID1), which leads to its interaction with DELLA proteins[12]. Then, GID1–DELLA interaction triggers the DELLA proteins to bind to an E3 ubiquitin ligase through specific F-box proteins (GID2 ⁄ SLY). This step leads to the polyubiquitination and the 26S proteasome-mediated degradation of the DELLA proteins, relieving DELLA's inhibitory effect on GA signaling[13]. This whole process then stimulates GA responses[14]. Research has found that DELLA deficient mutants, such as rice slr1[15] and Arabidopsis gai-t6 and rga-24[16], are taller and flower earlier than wild type plants. In contrast, DELLA sufficient mutants or transgenic plants are dwarf and flower late[17]. Such a mutant, the DELLA protein mutant gai-1 (gibberellic acid insensitive-1) from Arabidopsis[18] has been shown to reduce plant height and alter GA response in petunia[17], transgenic rice[19], tobacco[20], chrysanthemum[21] and apple[22]. It has been suggested that the DELLA mutant gai-1 protein from Arabidopsis has a 17-amino acid deletion in the conserved DELLA domain of GAI[18]. When GID and the mutant gai-1 protein form a complex together, they cannot be degraded by the 26S proteasome, which inhibits GA signaling, stops cell division and expansion, and results in an undesirable dwarf phenotype with improved drought resistance[17,18,20].
However, over-expression of gai-1 using the constitutively active 35S promoter results in completely dwarf plants that cannot ensure product quality in terms of flower or fruit size[17,19,22]. In order to generate normal plants that can have similar products as wild type plants, and can also survive drought stress, we proposed to use stress-inducible promoters to drive the expression of the gai-1 mutant gene. The RD29A promoter from Arabidopsis has been demonstrated that its activity is quickly and strongly induced by abiotic stresses such as drought, heat and salinity[23]. Studies indicated that within the RD29A promoter region, there are a number of cis-acting elements involved in the dehydration-induced activation, including the drought response element (DRE), and ABA response element (ABRE)[24,25]. Therefore, in this study, we chose to use a stress-inducible RD29A promoter to drive the expression of the gai-1 gene, which will allow us to initiate the expression of gai-1 at specific times or under drought stress conditions. We hypothesize that the temporary inhibition of cell growth, caused by inducible expression of the gai-1 gene, would lead to enhancement of drought stress tolerance and better crop productivity without an undesirable dwarf phenotype when the plants experience drought conditions.
-
Petunia (Petunia × hybrida cv. Mitchell diploid) seeds were sterilized and germinated on solid Murashige and Skoog (MS) medium at 22 °C with light intensity of 40 μmol/m2/s (12 h light per day). Seedlings with two true leaves were transferred into pots containing the UC Mix (1/3 Peat, 1/3 Sand, 1/3 redwood compost) and were grown in a greenhouse at the University of California Davis at 25−35 °C under natural conditions without supplemental light. Plants were watered with 1% (v/v) Hoagland solution every 3 d[6]. Seeds were collected from self-pollinated transgenic plants. T1 seedlings containing the transgene were selected on MS Medium containing 100 mg/L kanamycin. For drought and re-water treatments, plants were moved to a growth room in Mann Lab East Bay, where the temperature is 22 °C day and night. For long-term drought treatment, plants were grown in the greenhouse during summer for heat and intermittent drought treatment (watered every 4 d), then plants were moved to Mann Lab East Bay for final drought and re-water treatment.
Vector construction and plant transformation
-
A 1602 bp DNA fragment of the gai mutant gene (gai-1, GenBank accession number NM_101361) was cloned from the Arabidopsis gai-1 mutant plant using primers Atgai_BamHI_F: 5'-ATGGATCCATGAAGAGAGAT-3' and Atgai_SpeI_R: 5'-ATACTAGTCTAATTGGTGGAGAG-3'[17]. For cloning of the RD29A promoter, a 1,626 bp fragment of the RD29A promoter (GenBank accession No. CS191722.1) in the 5' non-coding region was amplified from wild type Arabidopsis genomic DNA using the primers RD29A_BamHI_F 5'-ATGGATCCGCTTGGTTGCTATGGTAG-3' and RD29A_BamHI_R 5'-ACGGATCCTTTCCAATAGAAGTAATC-3'[6]. The PCR products were cloned into a modified pGEM-T easy vector (pDAH11) (Promega, Madison, WI, USA) to generate the pDAH11:RD29A construct for sequence confirmation provided by the DNA sequencing service of the College of Biological Sciences at the University of California, Davis.
After sequence confirmation, we generated a plasmid construct by replacing the glucuronidase (GUS) gene with the gai-1 gene in an existing 35S expression vector (CD3-455, Arabidopsis Biological Resource Center). In addition, the 35S promoter in the upstream of the gai-1 gene was replaced by the RD29A promoter in the expression vector. To do that, restriction enzymes BglII with 5'-AGATCT-3' cutting site and BamH1 with 5'GGATCC-3' cutting site were used to digest the original 35S vector containing the gai-1 gene. According to the sequence analysis, there is a BglII cutting site in the RD29A promoter sequence, therefore, BamHl restriction enzyme was used to digest the pDAH11:RD29A as Bglll and BamHl have the same sticky end. The expression construct (pRD29A:gai) containing the kanamycin resistance gene for selection was used for plant transformation.
Plant transformation
-
Petunia (Petunia × hybrida, 'Mitchell Diploid') was transformed via Agrobacterium tumefaciens (LBA4404) with the pRD29A:gai construct. Agrobacteria were cultured overnight at 28 °C in LB medium (10 g/L Bacto-peptone, 5 g/L Bacto-yeast extract, 10 g/L NaCl, pH 7.2) (Difco, Detroit, MI) containing 50 mg/L kanamycin (Sigma, USA). The agrobacteria culture for inoculation of explants was centrifuged and was diluted to 1:200 (v:v) in Murashige and Skoog (MS) salts and vitamins medium (4.4 g/L) (Sigma, USA). When the plants were 10–15 cm high, leaves were collected, sterilized and infected with the agrobacteria cell suspension. The infected leaf disks were placed on co-cultivation medium at 25 °C for 2–3 d. After co-cultivation, these explants were transferred to a fresh regeneration medium (4.4 g/L MS, 30 g/L sucrose, 2 mg/L 6-BAP, 0.01 mg/l NAA, pH 5.8)[17,26].
Molecular characterization of the transformants
-
Putative transgenic lines were transferred into pots containing the UC Mix (1/3 Peat, 1/3 Sand, 1/3 redwood compost)[6] and grown in a greenhouse under natural conditions without supplemental light at 25−35 °C until flowering.
PCR amplification was performed for verification of transgenic lines. Genomic DNA of all the transformants was extracted using a hexadecyl trimethyl ammonium bromide (CTAB) method, as described previously[27]. Primer pairs Atgai_spel_F:5'-ATA CTAGTATGAAGAGAGATCATCAT-3' and Atgai_Sacl_R: 5'-ATGAGCTCCTAATTGGTGGAGAG-3' were used for the PCR reactions. The PCR products were analyzed with 1.0% (w/v) agarose gel electrophoresis using standard protocols[28]. Transformants that did not show positive PCR products were used as a non-transformed control.
In order to collect T0 seeds, the stigma of each flower was artificially pollinated using pollen from the same flower and covered with 3M tape to avoid cross-pollination. T0 seeds were collected, sterilized and sown on MS (Sigma, USA) medium with 100 mg/L kanamycin (Sigma, USA) for selection of transgenic lines. Seeds from wild type and empty-vector control plants were germinated and grown on a solid MS medium without kanamycin. The plates (20 seeds/plate) were kept at room temperature under continuous low fluorescent light (~40 μmol /m2/s) for 21 days[17,26].
Semi-Quantitative Real-Time PCR analysis
-
To determine the expression of the gai-1 gene in the transgenic plants, total RNA was extracted from leaves and flowers using TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) and treated with RNase-free DNase (Ambion, Austin, TX, USA) to remove any genomic DNA contamination. First strand cDNA was synthesized using 2 µg total RNA, oligo d(T) primer, random hexamer, and M-MLV superscript reverse transcriptase (Invitrogen, Carlsbad, CA, USA). This cDNA was normalized by real-time quantitative PCR using 26S ribosomal RNA primers (5'-AGCTCGTTTGATTCTGATTTCCAG-3' and 5'- GATAGGAAGAGCCGACATCGAAGG-3'). The normalized cDNAs were used for quantifying gai-1 gene expression using semi-quantitative PCR. The primers for amplifying gai-1 transcripts were Atgai_Spel_F:5'-ATACTAGTATGAAGAGAGATCATCAT-3' and Atgai_Sacl_R: 5'-ATGAGCTCCTAATTGGTGGAGAG-3'. Transcript levels of the 26S ribosomal RNA were used as a control[29,30]. Semi-quantitative PCR reactions were carried out in 25 µL volumes containing 1 µl reverse transcription reaction products (first strand cDNAs) as templates. The amplification was carried out at 58 °C for 90 seconds each cycle for 40 cycles. PCR products were analyzed by 1% (w/v) agarose gel electrophoresis. Wild type or empty vector plants were used as a negative control.
Fresh and dry weight of flowers
-
To examine effects of drought stress on T0 transgenic lines, T0 plants were grown in the greenhouse (natural light, 25−35 °C) during the early summer. Plants were watered approximately every 4 d. Flowers were selected at the point of fully opening. Fresh flowers were detached from the pedicel, immediately weighed for determining fresh weight and then placed in a 37 °C oven for 5 d until completely dry. The dry weight was subsequently measured. Five flowers from each plant were used for the measurement. The experiments were repeated three times. Flowers from the transgenic Line 1 (dwarf plant) were used as a positive control and wild type flowers were used as a negative control. Fresh and dry weights of the flowers were compared before and after long-term drought and heat treatment.
Cell density
-
Flowers were collected when fully opened and placed in a tube with distilled water to avoid wilting. Abaxial epidermis cells (1 cm2 samples) from the fully opened flower petals were counted in each view area under a microscope with an enlargement factor of 10*40. Three view areas for each flower were counted. Three flowers for each plant were used for biological replicates. The experiments were repeated three times. Transgenic dwarf flowers (Line 1) were used as a positive control and wild type flowers were used as a negative control. Flower cell density was compared before and after long-term drought and heat treatments.
Drought treatment with limited irrigation
-
Non-transformed control and transgenic T1 plants were grown in a greenhouse (25−35 °C with natural light) for one month with limited water. Plants were watered once every 7 d. A volume of 500 mL water was provided in each tray (26 cm × 52 cm, F1020, Hummert International, Earth City, Missouri, USA) with 6 plants per tray. After that, plants were moved to a growth room at 22 °C with light intensity of 40 μmol/m2/s, and subjected to drought stress by withholding irrigation for 18 d. Then, plants were re-watered. The recovery process was photographed using a camera (Nikon P50). In this experiment, four transgenic lines #7, #9, #12 and #13 were used while wild type and non-transformed plants were used as controls with four plants per line.
Data analysis
-
All measurements were repeated at least three times from different individual samples. Data were analyzed by a statistical program R Studio (Version 0.99.491). A 95% confidence interval was constructed using Tukey's method to confirm if the differences between factors are significant after two-way ANOVA analysis.
-
A total of 17 independent transgenic lines were generated and 11 T0 lines were shown to contain the gai-1 transgene (Fig. 1). Among these 11 lines, three lines showed a dwarf phenotype even under non-stressed conditions, including lines 1, 10 and 13. The dwarf phenotype suggests that there might be leaky expression of gai-1 without drought induction (Fig. 2). This is consistent with the result of a previous study by Liang et al.[17], which showed that even without dexamethasone (Dex) inducers, the expression of the Arabidopsis gai-1 mutant gene under the control of a GVG inducible system in T1 petunia seedlings caused dramatic growth retardation with short internodes in a few transgenic lines. The leaky expression may be the result of transgene insertion into the vicinity of an endogenous enhancer or promoter because of the random integration of the transgene. Following self-pollination and kanamycin selection, the T1 plants without dwarf phenotypes were subjected to drought stress treatment. These plants exhibited similar growth and developmental changes under non-stressed growth conditions to the control lines (wild type and empty-vector). This phenotype confirmed that the RD29A promoter does not act under non-stressed growth conditions. Nevertheless, under non-stressed conditions, the seeds from transgenic dwarf lines showed lower germination percentage and the plants grew much slower than the wild type control. Karssen et al.[31] suggest that GAs are required for seed germination. Therefore, this is consistent with our results that seeds from transgenic dwarf plants geminated slowly or did not germinate at all due to the deficiency of GA responses.
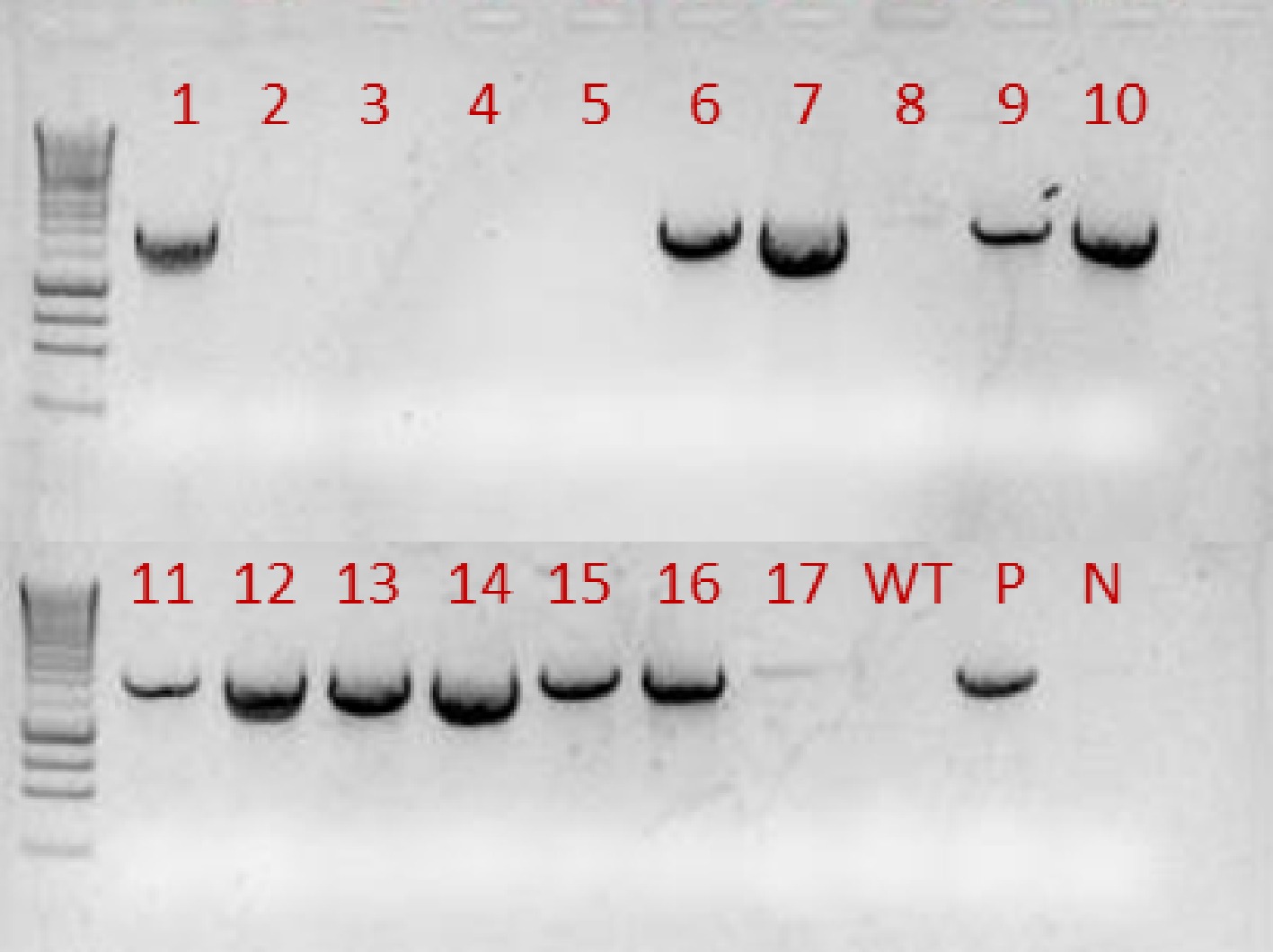
Figure 1. A total of 17 independent putative transgenic T0 lines were generated. Genomic DNA was extracted from leaves of these lines. PCR-based genotyping was performed to confirm the presence of the transgene using Forward Primer: 5’-CTAGAAAACAATCAGGAATAAAGGGT-3’ from RD29A and Reverse Primer: 5’-ATACTAGTCTAATTGGTGGAG AG-3’ from gai-1. Results showed that at least 11 lines contained the transgene. WT: non-transgenic wild-type; P: positive control using the plasmid DNA containing transgene gai; N: negative control.

Figure 2. Transgenic plants show normal growth and development under non-stress conditions. The L1 line with the leaky expression of the transgene displayed a dwarf plant phenotype compared with the transgenic lines L7, L9 and wild type.
Expression of the gai-1 transgene was strongly induced after drought treatment in petunia petals and leaves
-
In order to confirm that the gai-1 transgene can be induced by the RD29A inducible promoter under drought, T0 flowers were detached and exposed to open air at room temperature (20 °C) to induce mild dehydration. Expression of the gai-1 transgene was strongly induced in the T0 petals after 5 h of this 'drought' treatment (Line 7, Fig. 3). Transgenic Line 1, the dwarf plant with leaky expression, was used as a positive control. Wild type flowers were used as a negative control. Line 1 flowers had very high gai-1 expression both at 0 h and 5 h while expression was not detected in wild type flowers (Fig. 3a). Line 7 flowers had low level of gai-1 expression at 0 h, which may be due to the summer greenhouse conditions (25−35 °C) having already induced RD29A promoter activity and triggered a slight expression of gai-1. After 5 h, expression of the gai-1 transgene was strongly induced in Line 7 flowers (Fig. 3a). To further examine the induction of gai-1 expression, transgenic T1 whole plants were also treated with drought for 5 d. Leaves from treated plants were sampled and RNA from leaves was extracted and reverse transcribed to cDNA. Semi-quantitative PCR analysis showed that transgenic Line 1 and Line 7 both had a strong gai-1 transgene expression (Fig. 3b).

Figure 3. Semi-quantitative PCR analysis of gai-1 expression in transgenic petunia plants. Expression of the gai-1 transgene was strongly induced after 5 h in the petals (a) and 5 d in the leaves (b) after drought treatment. The RD29A:gai L1 line showed strong leaky gai expression even without drought treatment (a).
Drought treatments resulted in darker green leaves and smaller flowers
-
After the examination of gene expression, we further applied drought treatment on T0 plants. As there was only one plant for each T0 line, flowers and leaves from these lines were used as replicates. Non-transformed wild type and dwarf transgenic (leaky expression) plants were used as controls. Phenotypically normal transgenic and wild type control plants have similar flower cell density (around 17 cells per view area) before drought treatment in the transgenic T0 plants, whereas dwarf plants have higher flower cell density (around 24 cells per view area) than wild type control plants (Fig. 4a). After the drought treatments, phenotypically normal transgenic plants and dwarf plants have higher flower cell density (around 20 cells per view area) compared to wild type control (around 16 cells per area) (Fig. 4b). Leaves of the transgenic plants turned darker green similar to dwarf plant leaves after the drought treatment (Fig. 4c).
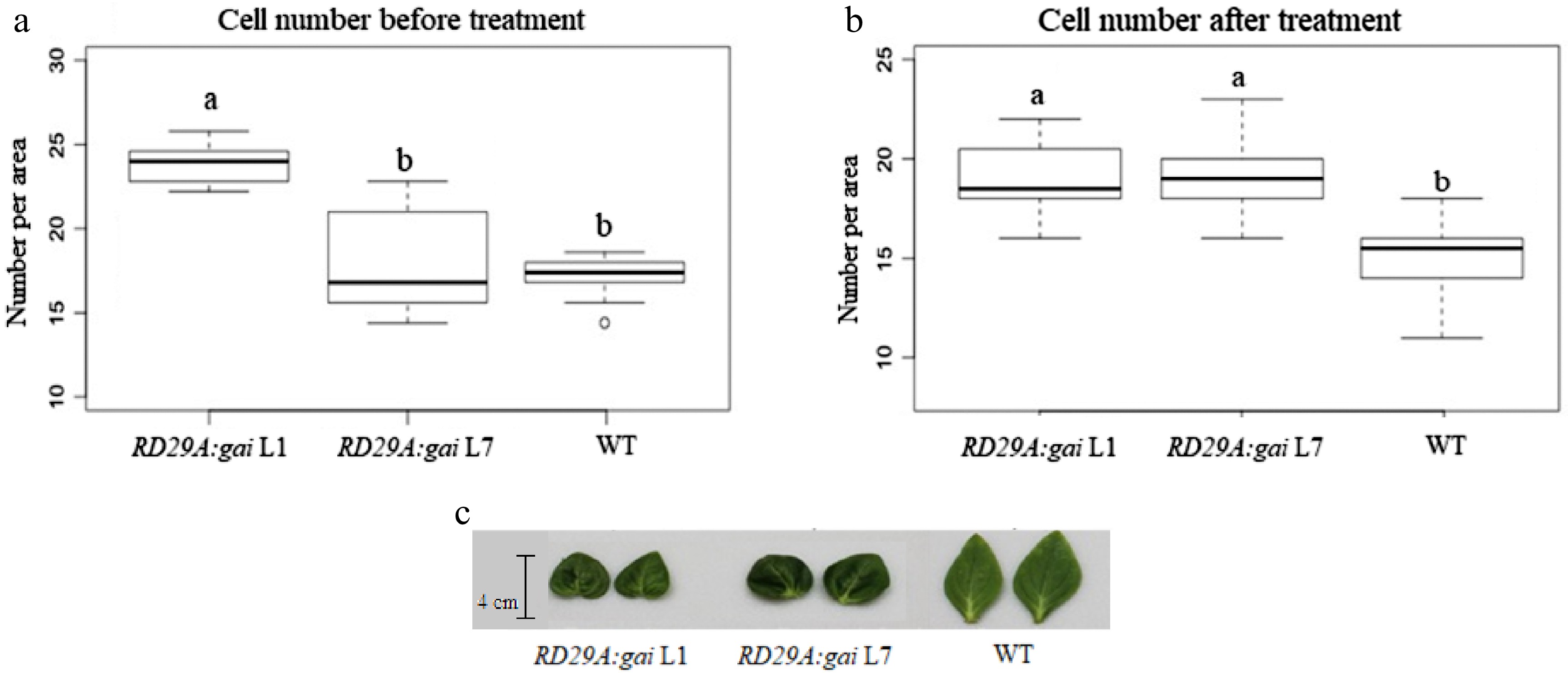
Figure 4. Cell number and morphology of leaves in response to drought treatment. (a) Plants were grown in the growth chamber at 22 °C with regular irrigation. (b) Plants were grown in the greenhouse at approximately 35 °C with drought treatment. Abaxial epidermis cells (1 cm2 samples) from the fully opened flower petals were counted in each view area under a microscope with a camera for recording cell images. (c) Representative leaves from transgenic plants under drought and heat treatments showed darker green phenotype and were smaller than the wild type control. Different lower case letters represent groups that are statistically different from each other at p < 0.05.
Flowers from phenotypically normal transgenic and wild type control plants have significantly higher fresh and dry weight (~0.50 and 0.078 g of fresh and dry weight, respectively) compared to the dwarf plant flowers (~0.42 and 0.069 g of fresh and dry weight, respectively) before the drought treatment (Fig. 5a). After the drought treatment, flowers from phenotypically normal transgenic and dwarf plants have significantly lower fresh and dry weights (~0.35 and 0.055 g of fresh and dry weight, respectively) than wild type plant flowers (~0.45 and 0.075 g of fresh and dry weight, respectively) (Fig. 5b).

Figure 5. Flower fresh and dry weights before (a) and after (b) drought treatments. Fully opened flowers are selected and fresh weights were measured. After that, the flowers were placed in a drying-oven at 37 °C for 5 d for measuring the dry weight. Ratios of Dry weight/Fresh weight were calculated. Different lower case letters represent groups that are statistically different from each other at p < 0.05.
These results are a confirmation of the pRD29A:gai inducible system, i.e. that the RD29A promoter was induced by drought stress, which then triggered the expression of gai-1. This in turn prohibited cell expansion resulting in smaller cells in new leaves and flowers.
Drought treatment with limited irrigation
-
To test drought stress tolerance in T1 transgenic plants, a limited water and drought treatment was carried out. Plants were grown under greenhouse conditions for one month with limited irrigation regime once a week with 500 mL water each tray. Wild type, empty-vector control and 4 transgenic lines with 4 plants each were included in a tray. Two trays were set up for replication. After the treatments, plants were subjected to drought stress testing in a growth room for 18 d. All plants including wild type, empty-vector control and transgenic plants have a similar wilting level after the final 18 d drought treatment (Supplemental Fig. S1a). However, transgenic plants recovered significantly faster than the control plants after re-watering (Fig. 6; Supplemental Fig. S1b). The mean recovery time for transgenic plants was 3 d. On the other hand, it took around one week for the wild type and empty-vector control plants to fully recover (Fig. 6). These results provide strong evidence that temporary inhibition of cell growth caused by over-expression of the gai-1 mutant with a drought stress-inducible promoter, leads to better drought resistance when the plants experience drought conditions.

Figure 6. Enhanced drought resistance in the transgenic plants. Plants were grown in the greenhouse in a tray for limited irrigation and watered once a week with 500 mL water per tray for a month. Plants were then subjected to drought treatment by withholding irrigation for 18 d. The phenotypes of plants were photographed one week after re-watering.
Drought resistance is not a single-hormone-induced process but the interaction among different hormones. ABA usually plays a dominate role in protecting plants under drought conditions. A previous study by Estrada-Melo et al.[6] indicated that over-expression of 9-cis-epoxycarotenoid dioxygenase (NCED), a key enzyme in ABA biosynthesis, in petunia plants under the control of a stress-inducible promoter, RD29A can significantly increase drought resistance. Not only did the transgenic plants survive longer than control plants under drought conditions, but transgenic plants also recovered earlier than control plants after stopping watering for the same period of time. It would be interesting if we can create the hybridization of RD29A:gai and RD29A:NCED and test the effect of the cross-talk between GA and ABA on plant drought resistance.
-
The Arabidopsis gibberellic acid insensitive mutant gai-1 can be expressed heterologously in petunia using an inducible system. Transgenic petunia plants showed normal growth and development under non-stressed conditions at 22 ºC day/night temperature. The data confirmed that the RD29A promoter was induced by drought and therefore gai-1 was expressed, which resulted in the reduced cell size when transgenic plants were treated with drought stress. Transgenic plants had better drought resistance than wild type and non-transformed control plants after drought treatment.
- This work was partially supported by the United States Department of Agriculture (USDA) CRIS project 2032-21220-008-00D. Yaping Zhang thanks the China Scholarship Council (CSC) for fellowship support.
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Enhanced drought resistance in the transgenic plants. Plants were grown in the greenhouse in a tray for limited irrigation and watered once a week with 500 mL water per tray for a month. Plants were then subjected to drought treatment by withholding irrigation for 18 days. (a) The phenotype of plants was recorded 18 days after the drought treatment. (b) Plants were re-watered to allow recovery. Photographs of the phenotype were taken three days after re-watering.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang Y, Norris A, Reid MS, Jiang CZ. 2021. Improvement of drought resistance through manipulation of the gibberellic acid pathway. Ornamental Plant Research 1: 11 doi: 10.48130/OPR-2021-0011

Improvement of drought resistance through manipulation of the gibberellic acid pathway
- Received: 08 July 2021
- Accepted: 28 October 2021
- Published online: 17 November 2021
Abstract: Improving plants' ability to survive under drought is of great importance to the horticultural industry. The plant hormone gibberellic acid (GA) mediates diverse aspects of plant growth and development. The Arabidopsis gibberellin acid insensitive mutant gai-1 displays reduced plant height, altered GA response, and enhanced drought resistance. However, over-expression of gai-1 using the constitutive 35S promoter results in dwarf plants with drought resistance. Here, we tested the hypothesis that the temporary inhibition of cell growth caused by inducible expression of the gai-1 gene would lead to better drought resistance and improve crop productivity without an undesirable dwarf phenotype. We generated transgenic plants in which the gai-1 gene was over-expressed in petunia, under a stress-inducible RD29A promoter from Arabidopsis. When these plants were subjected to limited irrigation and drought treatments, transgenic plants showed phenotypes of darker green leaves and compact flowers compared to the wild type plants. Importantly, these transgenic plants recovered sooner than wild type and the empty vector-transformed control plants. This study provides evidence that temporary inhibition of cell growth caused by over-expression of the gai-1 mutant gene with a drought stress-inducible promoter leads to better drought resistance when the plants experience drought conditions.
-
Key words:
- Drought /
- Gibberellic acid (GA) /
- Gibberellic acid insensitive gai-1 /
- Inducible promoter /
- Petunia /
- RD29A