









-
Apomixis is a special mode of asexual reproduction whereby asexual seeds are formed, resulting in the production of clonal embryos genetically identical to the mother plant[1]. The introduction or transfer of apomixis into sexual crop plants has great value for breeding for desired agronomic traits and maintaining heterozygosity and heterosis over successive generations[2]. Since John Smith first discovered the apomictic phenomenon in Alchornea ilifolia in 1841, many studies have concentrated on its mechanism in different species from many aspects, such as embryo development, morphogenesis, genetic evolution, physiology and biochemistry, and hybridization breeding. Nonetheless, many problems remain to be explained and solved in terms of apomictic occurrence, including the genetic basis, molecular regulation and evolutionary origin, because of the diversity of apomictic types and the genetic complexity. At present, apomixis can be generally categorized into the following two basic types: sporophytic and gametophytic apomixis. With respect to sporophytic apomixis, the embryo originates from somatic cells in ovules after mitosis, as is found in some members of Malus, Crataegus L. and Zanthoxylum L. in Rosaceae. However, a functional and chromosomally unreduced megaspore is utilized to produce fertile seeds under gametophytic apomixis; here, the embryo is derived from either diploid megaspore mother cells or cells near nucellar cells without fertilization (parthenogenesis) through apomeiosis[3,4]. In addition, in natural apomixis, the formation and development of sexual endosperm by pseudogamy (the fertilization of central cells) is required to nourish the growth of parthenogenetic embryos and ultimately produce viable seeds[5,6]. Otherwise, the embryo will abort. Thus, in addition to the meiosis, fertility and embryogenesis, both the development of endosperm and its sensitivity to parental imprinting could be considered for the transfer of apomixis to experimental or crop species[7].
Some species exhibit obligate and facultative apomixis affected by self-compatibility, chromosome ploidy, hormones and environmental factors over long-term evolutionary histories[3,8]. Most apomictic plants usually exhibit self-incompatibility, which are formed during the process of evolution in woody horticultural species. Different plant species use distinct pathways of apomixis to form seeds successfully[9,10]. There are two mechanisms of apomixis by which asexual seeds are produced through the formation of a reduced megaspore via meiosis (mostly at low frequencies), or the formation of an unreduced megaspore via mitosis[11,12]. However, the generation process of early megaspores, especially unreduced megaporocyte megaspores, varies among different plant species. Although the aposporous development and formation derived from one or more somatic cells in the nucellar tissue, is triggered after abortion of the sexual megaspore among most of the wild populations of Malus hupehensis, M. sieboldii, M. rockii, M. sargentii, and M. xiaojiinensis, these populations exhibit different capacities for apomixis[11]. This phenomenon was first associated with hybridization, polyploidy and female sterility[13−16]. The vast majority of natural apomicts are polyploid[17]. For example, studies on Tripsacum dactyloides indicated that diploid plants showed no apomixis and exhibited sexual reproduction accompanied by regular meiosis, whereas the tetraploid form adopted facultative apomixis by diplospory and pseudogamy. Wet et al. reported that plants above the tetraploid level exhibited obligate apomixis[18]. Pollen also affects megaspore development in woody plants, such as walnut, pear, apple, and cherry, most of which exhibit polyploid apomixis with fertile pollen but are self-incompatible[19−20]. In these polyploid species, some plants show male fertility, but the viability or germination rates of their pollen decrease, whereas some species show abnormal male organ development, limited pollen release, or male organ retardation. These characteristics often lead to a failure in cross-pollination[21]. Thus, apomicts may be closely related to not only the development of the megaspore but also that of the male organs (pollen and anther)[22,23].
In recent years, researchers have identified several genes involved in epigenetic regulation, control of the cell cycle, hormonal pathways and signal transduction processes related to plant apomixis; additionally, DNA- and RNA-binding proteins, as well as non-coding RNAs, are likely to be relevant for the formation of apomixis[24−26]. Moreover, there has been success in the induction of apomixis-like features by mutating genes related to meiosis, fertility or embryogenesis[27]. Studies on Hieracium aurantiacum indicated that LOA1 (loss of apomeiosis 1) is involved in the initiation of apomixis[28]. Oncogene rolB might regulate the reproduction by influencing cellular sensitivity to auxin or altering cell signalling that is induced by changes in ovule morphology[29]. In rice, the MEL1 gene regulates the division of premeiotic germ cells, the modification of meiotic chromosomes, and the proceeding of meiosis and meiosis[30]. The functional losses of the FIE or MEA allele causes the female gametophyte to initiate endosperm development without fertilization[31]. Liu et al found that the MhFIE gene (fertilization-independent endosperm, FIE), a homologue of Arabidopsis FIE, plays an important role in apomixis-related reproductive development in apple[32,33]. In Arabidopsis, the WD-40 domain protein MSI1 is involved in endosperm development and is required for seed development as a component of the MEA/FIE polycomb group complex[32]. Recently, a receptor protein kinase (somatic embryogenesis receptor kinases, SERK) genes have been shown to promote embryogenesis during early apomictic embryo formation and participate in the exchange of information between cells[34], such as AcSERK2 in pineapple[35], SERK1 in triploid Pingyi Tiancha[36], and SERK1 and SERK2 in Brassica napus L[37]. MYB transcription factors are an important class of transcription factors in plants that regulate growth, development, the biosynthesis of secondary metabolites, and biological and abiotic stresses in plants[38−40]. SPL transcription factors classified into the SBP gene family play a wide range of regulatory roles in the growth and development, innate immunity, reproduction and adversity resistance of plants[41]. In the past few years, increasing attention has been focused on the MYB and SPL transcription factors as they are involved in the growth and development of plant reproductive organs. For instance, MYB21, MYB24, and MYB57 promote the growth of the stamen filament[42]; SPL8 regulates the patterning of the gynoecium[43]; and AtMYB32 is necessary for normal pollen development in Arabidopsis thaliana[44]. However, the association between pollen abortion and apomixis has not yet been clarified.
Ornamental crabapples are the hybrid offspring of two Malus species groups, one from East Asia and China that includes the species M. baccata, M. sieversii, M. spectabilis, M. toringoides, M. sieboldii, and M. halliana and the others from North America that include the species M. ioensis and M. coronaria[45]. These hybrids have a dwarf plant architecture and thus are used as special types of apple germplasm resources for the breeding of cultivars and rootstock. In 1931, Sax discovered the apomixis phenomenon in M. hupehensis Rehd., which initiated the research on apomixis in Malus plants[46]. At present, apomixis has been found in ten species in Malus plants, including M. hupehensis Rehd., Malus sargentii Rehd., Malus sieboldii Rehd., Malus sikkimensis Koehne., M. toringoides Hughes., M. platycarpa Rehd., Stator M. rockii Rehd., M. xiaojinensis Cheng et Jiang, M. lancifolia Rehd., and M. coronaria (L.) Mill[47−49]. One cultivar, 'Royalty', exhibits red fruits, leaves, and flowers, which makes it an ideal parent for breeding red apple varieties and dwarf rootstocks[50]. 'Royalty' has been used as a rootstock resource for the apple industry. Thus, determining the apomictic characteristics of this cultivar has the potential to play a significant role in agriculture by creating woody fruit plants with high heterozygosity.
The Malus cultivars 'Royalty' (3x) and 'Flame' (2x) represent the hybrid offspring of M. baccata and M. pumila var. neidzwetzkyana. Malus cultivar 'Flame' (2x) has the same genetic background as 'Royalty', and it has been used for hybridization testing. To reveal the association between pollen development and apomixis, as well as the molecular mechanism of apomixis in Malus crabapples, we took 'Royalty' (3x) and 'Flame' (2x) as the experimental material, we designed different pollination combination treatments and investigated the developmental features of reproduction organs and genetic characteristics in the current study. Based on transcriptome sequencing of 'Royalty', we analysed the expression profiles of genes and transcription factors related to apomictic reproduction. Our results indicated that the triploid cultivar 'Royalty' exhibits both facultative apomixis when no pollen is available and sexual reproduction under the stimulation of foreign pollen. In addition, we observed the developmental features of anthers, pollen, and megaspores during flowering. Our results indicated that both the dysplasia of male organs and the decrease in the germination rate of pollen may have a direct impact on the efficiency of pollination, resulting in the occurrence of apomixis in Malus crabapples. Our study provides new insights into the mechanism of apple apomixis in Malus crabapples from the perspectives of embryology and transcriptional regulation.
-
The methods to study apomixis mainly include morphological and cytoembryological observation and molecular biological detection or identification. Pollination and fertilization experiments in the flowering period are of great significance for the study of apomixis of ornamental crabapples. To investigate whether apomixis exists in Malus crabapples, we conducted eight different pollination treatments: 'Royalty' EB (bagged 'Royalty' flowers after emasculation); 'Royalty' BA (bagged 'Royalty' flowers alone); 'Royalty' EFB (bagged 'Royalty' flowers received the pollen of 'Flame' after emasculation); 'Royalty' CK (natural flowers of 'Royalty' without any treatment); 'Flame' EB (bagged 'Flame' flowers after emasculation); 'Flame' BA (bagged 'Flame' flowers alone); 'Flame' EFB (bagged 'Flame' flowers that received the pollen from 'Royalty' after emasculation); and 'Flame' CK (natural flowers of 'Flame' without any treatment).
As shown in Fig. 1 and Table 1, with regard to the CK groups (untreated control), we observed that the number of setting fruits in 'Royalty' was significantly higher than that in 'Flame'. However, there were no significant differences in the fruit setting rate or the average number of seeds per fruit. For the EFB (pollination followed by bagging) treatment, the number of set fruits and the fruit-setting percentage of fruit-setting in 'Royalty' (♀) × 'Flame' (♂) were 589 and 98.44%, respectively. The number of fruit settings and the percentage of fruit settings in 'Flame' (♀) × 'Royalty' (♂) were 21 and 4.62%, respectively. The results show that the former cross was significantly more successful than the latter. This result strongly indicated that 'Royalty' may provide poorer quality pollen than 'Flame' when used as the donor.
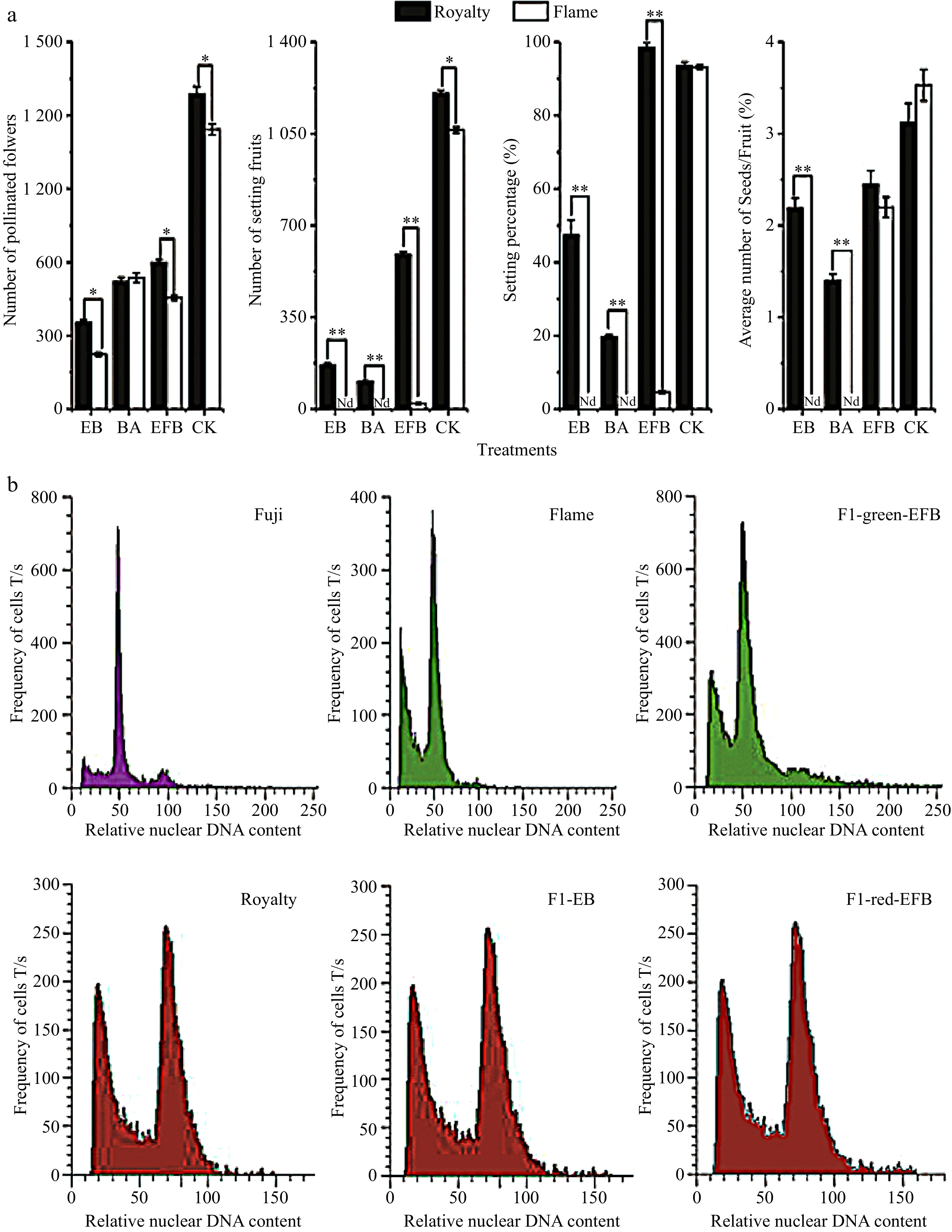
Figure 1. Fruit set and number of seeds across pollination treatments between Malus 'Royalty' and 'Flame' (a) and the fluorescence intensity distribution of nuclear DNA in the young leaf cells of the Malus cultivars (b). (a) The fruit setting rates and the average number of seeds/per fruit in Malus cultivars under different treatments. EB: bagging after emasculation; BA: bagging alone; EFB: pollination followed by bagging; and CK: untreated. All experiments were conducted with labelling at the large bud stage. The values represent the means from 2014 and 2015. The data (mean ± SD) are the sum of three biological repetitions for each treatment. (b) Fluorescence intensity distribution of nuclear DNA in the young leaf cells of the Malus cultivars 'Fuji', 'Flame', and 'Royalty', as well as their offspring. 'Fuji', a diploid Malus cultivar, was used as a reference to detect the ploidy of Malus crabapple 'Flame' and 'Royalty' by using flow cytometry. F1-EB: the F1 seedlings of 'Royalty' EB; F1-red-EFB: the F1 seedlings with a red phenotype from 'Royalty' EFB ('Royalty' (♀) × 'Flame' (♂)); and F1-green-EFB: the F1 seedlings with a green phenotype from 'Royalty' EFB ('Royalty' (♀) × 'Flame' (♂)).
Table 1. Statistical results of the pollination test for Malus 'Royalty' and 'Flame'.
Pollination type Crossing groups Number of pollinated flowers Number of
fruitsSetting percentage (%) Average number of seeds Bagging after emasculation (EB) 'Royalty' (♀) 354 168 47.46 2.19 Bagging after emasculation (EB) 'Flame' (♀) 224 0 0.00 0.00 Bagging alone ('Royalty' BA)) 'Royalty'(♀) × 'Royalty' (♂) 521 104 19.70 1.40 Bagging alone (Flame' BA) 'Flame'(♀) × 'Flame' (♂) 537 0 0.00 0.00 Pollination followed by bagging after emasculation (EFB) 'Royalty' (♀) × 'Flame' (♂) 589 589 98.44 2.45 Pollination followed by bagging after emasculation (EFB) 'Flame' (♀) × 'Royalty' (♂) 455 21 4.62 2.20 Untreated (CK) 'Royalty' (♀) × '?'(♂) 1,287 1,204 93.55 3.10 Untreated (CK) 'Flame' (♀) × '?'(♂) 1,143 1,065 93.18 3.50 Notes: Average number of seeds is average seed number per fruit. At least three biological replications were performed. The fruit setting rate in the 'Royalty' EFB group was 98.44%, which was much higher than the fruit setting rate of 19.7% in the 'Royalty' BA group. This result suggested that 'Royalty' tends to prefer sexual reproduction and has good sexual reproduction ability under the stimulation of foreign pollen. Nevertheless, without available stimulation from foreign pollen, 'Royalty' initiated an apomictic reproduction mode, implying that the pollination and fertilization in 'Royalty' BA may face barriers. Moreover, the fruit setting rate in the 'Royalty' EB group was 47.46%, which was more than twice the fruit setting rate of 19.7% in the 'Royalty' BA group. This result suggested that 'Royalty' exhibits high facultative apomictic reproduction, which suggested that the pollen of 'Royalty' may have developmental defects or exert adverse effects on the reproductive process. The fruit setting rate in the 'Flame' EFB group was 4.62%, which is significantly lower than the fruit setting rate of 93.18% in 'Flame' CK, which also strongly indicated that 'Royalty' provides poor quality pollen when used as the donor. However, with regard to the 'Flame', no fruit setting occurred in 'Flame' EB and 'Flame' BA, indicating that 'Flame' did not exhibit apomixis.
To determine whether the seed vigour differed after different treatments on 'Royalty' and 'Flame' showing normal vigour, we examined the seed germination rates and seedling emergence rates. As suggested in Supplemental Fig. S1 and Supplemental Table S1, the seed germination rates and seedling emergence rates of 'Royalty' from EB and EFB were maintained at a certain level and the potted plants exhibited good growth vigour, although these rates were lower than those of CK, indicating that the seeds of 'Royalty' EB possessed normal vigour. Additionally, the F1 seedlings of 'Royalty' EB showed red leaves and stems, similar to its maternal plant, indicating the characteristics of maternal dependence in apomixis. However, the seedlings of 'Royalty' EFB displayed a certain segregation of phenotypic characteristics. Together, these observations demonstrated that the cultivar 'Royalty' showed facultative apomixis, which may be related to pollen stimulation. That is, when there is external pollen stimulation in the flowering period, 'Royalty' accepts foreign pollen for sexual reproduction and produces sexual seeds. However, when there is no foreign pollen stimulation, this cultivar initiates apomixis and generates apomictic progenies to maintain its genetic stability. On the other hand, the fruit setting rates of BA were lower than those of EB, which may be related to the viability and germination of the donor pollen and 'Royalty'.
Chromosome ploidy analysis of the F1 generation
-
The DNA content of the nucleus in G1 phase reflects the chromosome ploidy of a cell. Based on the proportional relationship between DNA content and fluorescence signal intensity, flow cytometry can be used to accurately determine the content of intracellular DNA and chromosome ploidy. To verify the presence of apomixis in the cultivar 'Royalty', we further analysed the chromosome ploidy in the F1 seedlings of 'Royalty' EB and 'Royalty' (♀) × 'Flame' (♂) EFB using a flow cytometer. As indicated in Fig. 1b and Table 2, we observed that the mean G1 and relative nuclear DNA content exhibited significant differences between the tested cultivars and between their offspring. The F1 seedlings of 'Royalty' EB were triploids (3x), similar to 'Royalty', indicating that they were derived from the apomixis of 'Royalty'. The F1 seedlings of 'Royalty' (♀) × 'Flame' (♂) EFB with green leaves and stems, displayed diploidy (2x), similar to 'Flame', indicating that they were derived from sexual reproduction of 'Royalty' (♀) and 'Flame' (♂). However, the F1 seedlings from 'Royalty' (♀) × 'Flame' (♂) with red leaves and stems were triploids (3x), implying that they may be derived from either apomixis of 'Royalty' (♀) or sexual reproduction of 'Royalty' (♀) and 'Flame' (♂) due to an uneven distribution of chromosomes during meiosis in 'Royalty'. Taken together, these results illustrated the presence of apomixis in the Malus cultivar 'Royalty'.
Table 2. Relative nuclear DNA content and chromosome ploidy level of 'Royalty' and 'Flame' as well as their offspring.
Cultivars Mean G1 Relative nuclear DNA content Ratio Chromosome ploidy level Ratio Fuji 50.01 ± 1.36b 100.00 1.00 2x 1.00 Royalty 76.04 ± 1.07a 152.05 1.52 3x 1.52 Flame 51.21 ± 0.75b 102.04 1.02 2x 1.02 F1-EB 78.48 ± 0.87a 149.01 1.49 3x 1.49 F1-red-FEB 77.38 ± 1.21a 150.66 1.51 3x 1.51 F1-green-FEB 54.34 ± 1.45b 108.66 1.09 2x 1.09 Notes: Mean G1: mean fluorescence intensity; F1-EB: F1 seedlings of 'Royalty' EB; F1-red-EFB: F1 seedlings with the red phenotype of 'Royalty' from 'Royalty' (♀) × 'Flame' (♂) EFB; and F1-green-EFB: F1 seedlings with the green phenotype of 'Royalty' from 'Royalty' (♀) × 'Flame' (♂) EFB. At least three biological replications were performed. Observations of megaspore mother cell development in 'Royalty'
-
The developmental characteristics of reproductive organs based on cytoembryology could provide important clues regarding the induction mechanism of apomictic reproduction. To obtain insight into the reason for the mechanism behind apomixis in 'Royalty' at the cytoembryological level, we observed the characteristics of megaspore mother cells at different developmental stages in the ovules of EB plants.
As shown in Fig. 2a1, we found that the nucellar tissue protruded and that double-layered integument tissues formed in the ovary at approximately 5−8 days before blooming. As shown in Fig. 2a2, some nucellar cells at the chalazal end began to divide and grow actively, while other cells disintegrated and died at 2−3 days before blooming (Fig. 2a3). Two days later, the active nucellar cells underwent division and growth with stronger vigour, and the surrounding cells continued to lyse and die, resulting in the formation of a large number of cavities within the nucellar tissue (Fig. 2a4,5). At the beginning of flowering, brightly stained nucellar cells at the chalazal end began to increase in volume and exhibited characteristics similar to those of primary sporogenous cells (Fig. 2a6). Subsequently, rapid vacuolization and an increase in the volume of sporogenous cells were observed, and one cell developed into unreduced megaspore mother cells after competition (Fig. 2a7-9). At the same time, the tetrad in nucellar tissues of 'Royalty' from EFB began to form (Fig. 2b1) and further developed (Fig. 2b2). One member of the tetrad developed into a functional megaspore mother cell, while the other three cells in the tetrad disintegrated (Fig. 2b3). At the full-bloom stage, the unreduced and reduced functional megaspore mother cells developed into monocytes (Fig. 2c1), binucleated megaspores (Fig. 2c2), and octonucleated megaspores (Fig. 2c3). An immature embryo (Fig. 2c4) was observed at 2−5 days after flowering.
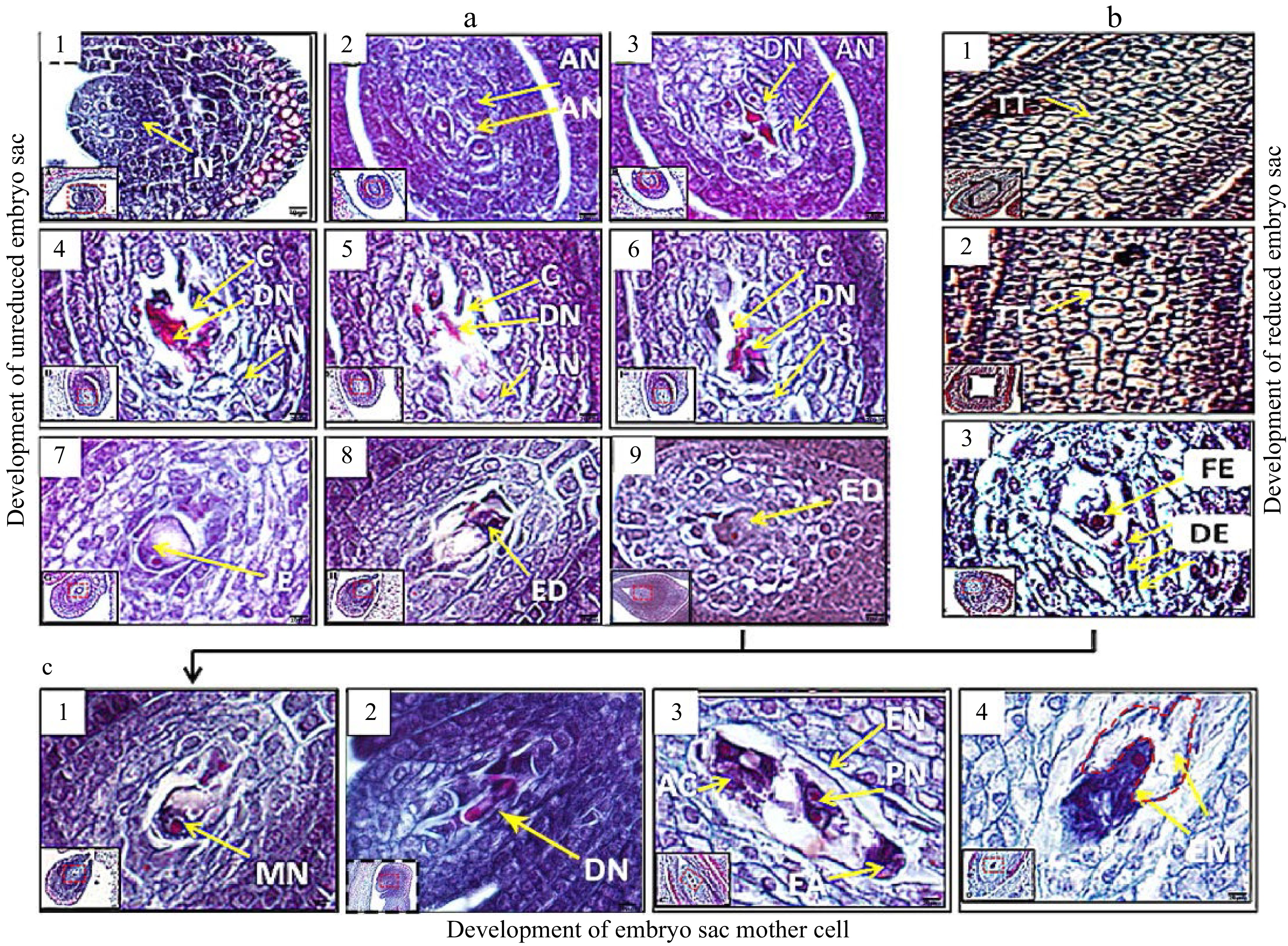
Figure 2. Paraffin sections of the development in the embryo mother sacs of 'Royalty' EB (a and c) and EFB (b) ovules. N: nucellus; AN: active nucellus cells; DN: disintegrated nucellus cells; TT: tetrads via meiosis; C: cavities left after nucellus cells disintegrated and died; S: sporogenous cells; E: embryo sac mother cells; ED: developing embryo sac mother cell; FE: functional embryo sac mother cells; DE: disintegrating embryo sac mother cells; MN: mononuclear embryo sac; DN: double nuclear embryo sac; AC: antipodal cells; EN: eight nuclear embryo sac; PN: polar nuclear; EA: egg apparatus; and EM: embryo. Bar = 5 μm. At least three biological replications were performed.
Observations of anther and pollen activities in 'Royalty'
-
Through the pollination experiments mentioned above, the dehiscence of anthers and specificity of pollen were considered to influence apomixis in 'Royalty' based on the fruit-setting rate. We further observed the pollen ultrastructure via optical microscopy and scanning electron microscopy to explore the specificity of anther dehiscence and pollen morphology in 'Royalty'. When the morphological characteristics of the male flower organs in the two cultivars were observed at early blooming, we found that the two cultivars had a normal appearance but different coloured filaments and anthers (Fig. 3a). Microscopy observations showed that some anthers of 'Royalty' differentiated into distinct structures, such as the epidermal, endothecium, intermediate, and tapetal layers, and produced four pollen sacs during the large bud stage. These anthers were suspected to resemble normal anthers (Fig. 3b1). However, most anthers were stunted; some were empty sacs with few pollen grains (Fig. 3b2); some were solid pollen sacs with undeveloped pollen (Fig. 3b3); and some contained only two or three dissymmetrical pollen sacs (Fig. 3b4). Additionally, with anther development, the tissue between 2 adjacent anthers formed a cavity (Fig. 3b5). The wall of the pollen sacs around the cavity became thin. The anther epidermis and intermediate layer adjacent to the cavity formed a slit through which the pollen could not emerge and scatter or developed only a few seams preventing pollen grains from being released (Fig. 3b6). Fig. 3d shows 15 microscopic view fields of 'Royalty'.
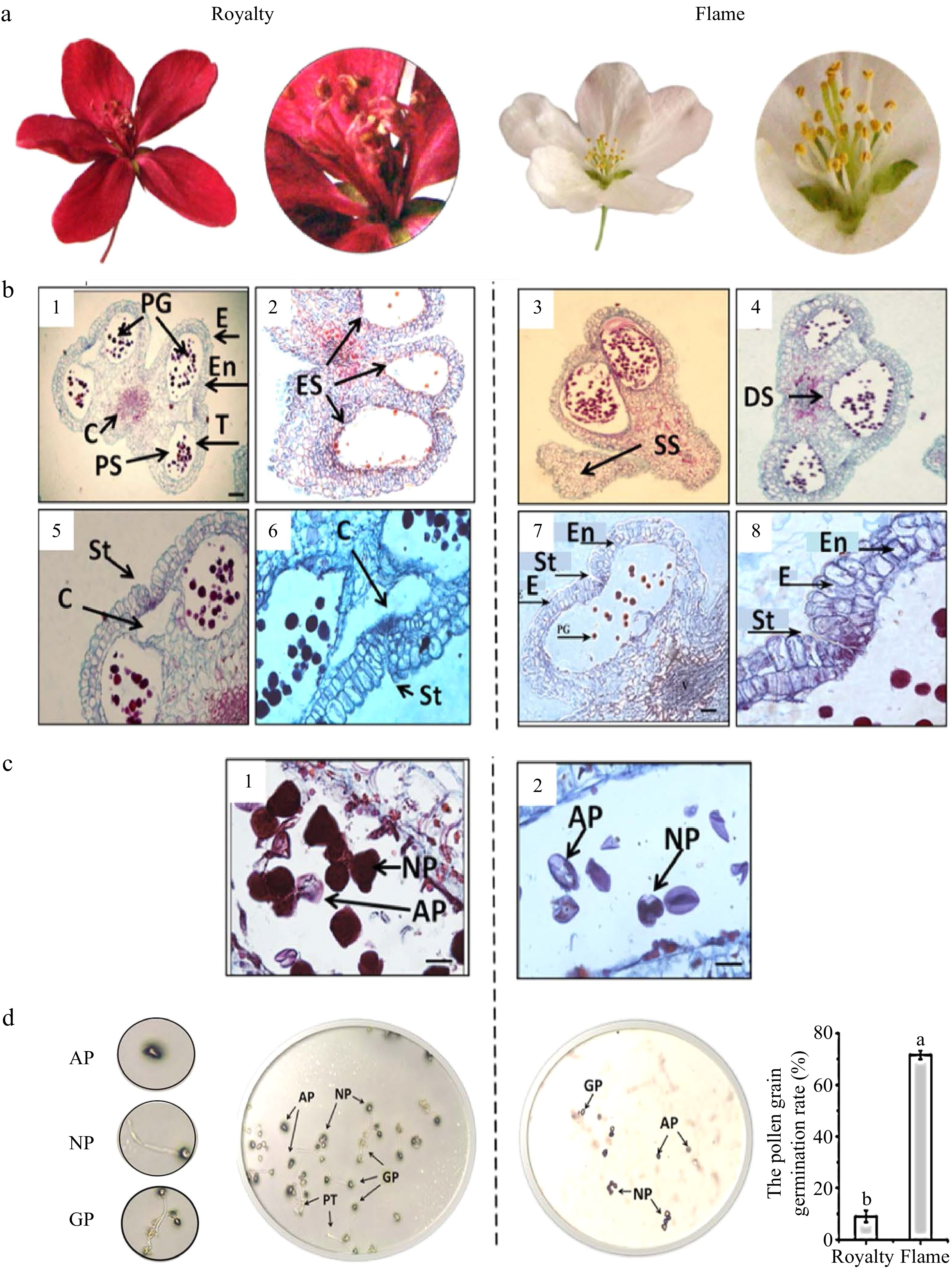
Figure 3. Morphological and anatomical characteristics of anther and pollen grains and pollen grain activity. (a−b) Morphological and anatomical images of anther and pollen grains in 'Royalty' and 'Flame'. (c−d) Pollen grain activity in 'Royalty' and 'Flame'. Note: E: epidermal layer; En: fibrous layer; T: tapetal layer; PG: pollen grains; C: connective tissue; PS: pollen sac; ES: empty sacs with few pollen grains; SS: solid pollen sacs not developed; DS: dissymmetrical pollen sacs without symmetric arrangement; St: stomium in the anther epidermis and intermediate layer adjacent to the cavity; C: cavity between 2 adjacent pollen sacs; NP: normal pollen grains; AP: abortive pollen grains; Gp: germinated pollen grains on culture medium; and PT: pollen tubes. Bar = 100 μm. At least three biological replications were performed.
Scanning electron microscopy revealed a significant difference between the two cultivars in the dehiscence of anthers. At the early flowering stage, the nondehiscent anthers of the two cultivars exhibited an inverted triangle shape and similar morphological features. However, during the full bloom period, most anthers of 'Royalty' were not dehiscent, and a few anthers split only narrowly, while all anthers of 'Flame' were fully dehiscent. At the end of the flowering stage, the anthers of 'Flame' had released their pollen grains, whereas only a few anthers of 'Royalty' had a small number of pollen grains clinging to their surfaces (Fig. 4a). During the statistical analysis of the microscopic fields, we also observed that there were significant differences in the pollen grain number and shape between the two cultivars.
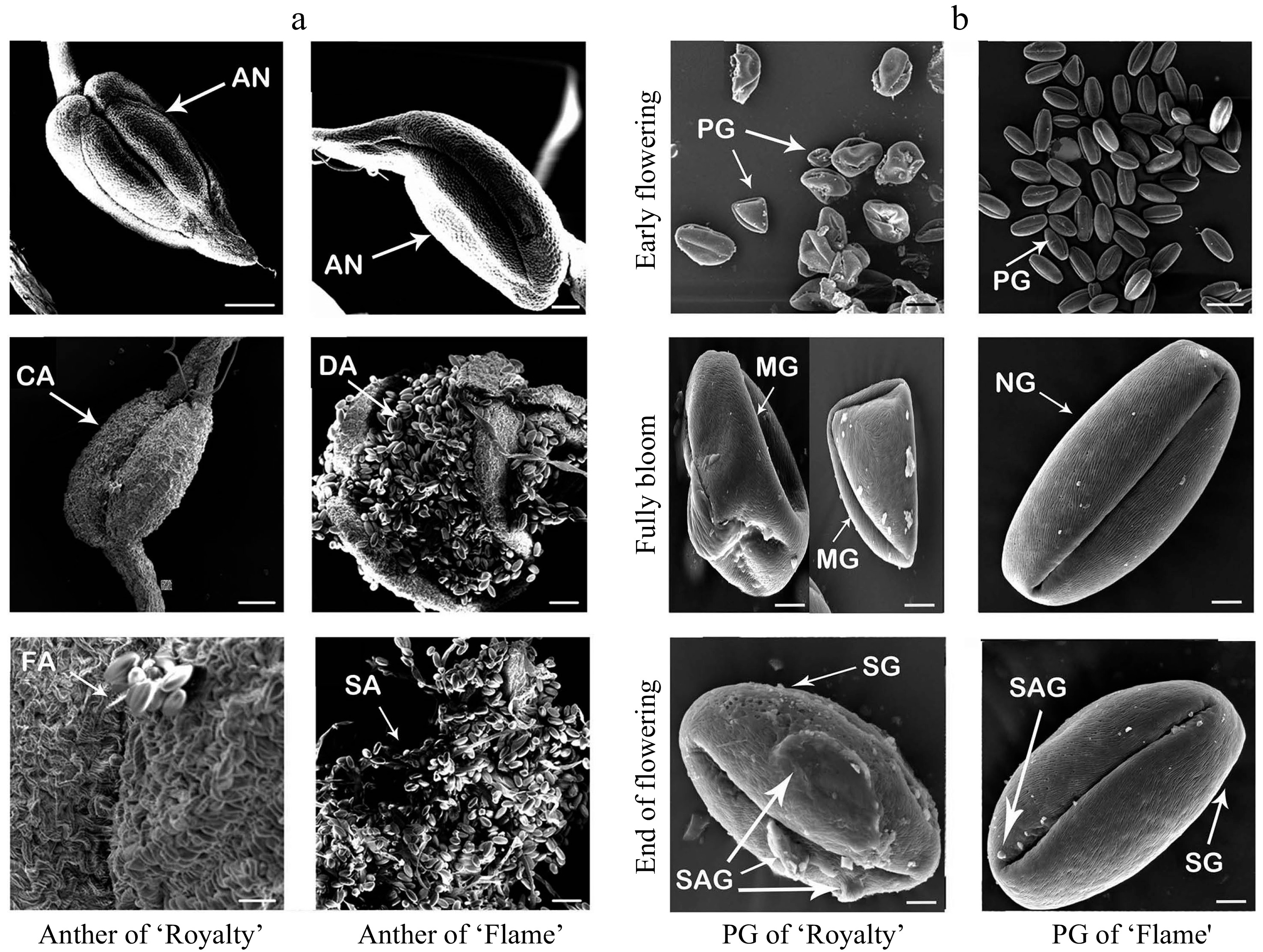
Figure 4. Scanning electron micrographs of anthers at different flowering stages (a) and pollen grains (b) during the blooming period of 'Royalty' and 'Flame'. AN: anther; CA: anthers that cannot split; DA: dehiscent anther and pollens; FA: anthers not fully split; SA: anthers with scattered pollen; PG: pollen grains; MG: malformed pollen grains; NG: normal pollen grains; SG: the surface of pollen grains; and SAG: the surface attachment of pollen grains. At least three biological replications were performed.
The pollen grains of 'Royalty' were significantly less abundant and included more malformed grains than those of 'Flame'. Additionally, the surfaces of pollen grains in 'Royalty' were not smooth as those in 'Flame' because of the numerous protuberance-like attachments on their surfaces, which were not as smooth as the surface of like 'Flame' pollen grains (Fig. 4b).
Expression profiles of genes and transcription factors related to apomixis in the pistil tissue of 'Royalty' during flowering
-
To obtain insight into the mechanism of apomixis in Malus crabapples at the transcriptional level, we performed RNA sequencing of the fruits of 'Royalty' and 'Flame' under the EB treatment at 20 days and 90 days after the blooming period. The results of RNA sequencing (RNA-Seq) identified many differentially expressed genes, including SQUAMOSA promoter-binding protein-like (SPL), MSI, EF-alpha (elongation factor 1 alpha), MYB and SERK genes (Table 3). To further explore the preliminary mechanism of the apomictic phenomenon in 'Royalty', we cloned the following genes associated with plant apomixis: McAPK (adenosine-5'-phosphosulfate kinase), McFIE (fertilization-independent endosperm), McMSI1, and McEF-alpha; genes associated with somatic embryogenesis, such as SERKs; transcription factors essential for anther and pollen development, such as MYB26, MYB32, and MYB103; and transcription factors associated with embryonic development, such as SPLs. Furthermore, we analysed the expression level of these genes in the two cultivars during flower development (at the T1, T2, T3, and T4 stages).
Table 3. The results of RNA sequencing (RNA-Seq) of 'Royalty' and 'Flame' fruits.
Gene description F/R_20d F/R_90d SPL domain class transcription factor, SBP domain 1.5155** −1.9002 SPL domain class ranscription factor, Mlo family//NUDIX domain//PSK 0.0018** 0.7627 Putative WD-40 repeat protein, MS12, WD domain, G-beta repeat 0.1297** −0.6362 Elongation factor 1 alpha subunit −0.482** −0.0517 MYB-like DNA-binding domain −1.0009** −0.0038 Nodulation protein S //Methyltransferase domain//Thiopurine S-methyltransferase −1.6507 −1.7201 Anther-specific gene, CcmE −2.0335** 0.0536 Putative pollen surface protein, Peptidase//Delta Atracotoxin 2.003* 3.6557 MYB domain class transcription factor −0.2966** 0.5912 MYB transcription factor, Cytotoxic 0.7377 0.4173 Methyltransferase domain//Putative methyltransferase 0.9047* 1.6914 Anther-specific gene, TAP42-1ike family −0.9796** −0.3826 Methylransferase domain//Fructose-bisphosphate aldolase class-1 −0.7489 −0.8379 Probable methyltransferase PMTI-like, Methyltransferase domain −0.8516** −0.0800 MYM-type Zinc finger //Methyltransferase domain 0.8559** −0.2436 Peroxidase//Methyltransferase domain −0.263** 0.2461 Hypothetical methyltransferase//ER lumen protein retaining receptor//Secreted protein acidic and rich in cysteine Ca binding 0.3528** 1.1421 Methyltransferase domain//Methyltransferase small domain −0.5462** 0.3341 Sigma-70, region 4//MYB like DNA-binding domain 0.166** −0.5268 Stromal cell-derived factor 2-like protein MIR domain 0.2252** −0.5918 Somatic embryogenesis receptor kinase 3B precursor, Leucine Rich Repeat//Protein kinase domain −0.9486** 0.4198 Notes: F/R-20d: the related gene expression ratios of 'Flame' to 'Royalty' fruits at 20 days after the blooming period; F/R-90d: the related gene expression ratios of 'Flame' to 'Royalty' fruits at 90 days after the blooming period. At least three biological replications were performed. As shown in Fig. 5, the expression of McMYB26 in 'Royalty' was lower than that in 'Flame' at T3 and T4 but higher at T1 and T2. The expression of McMYB32 in 'Royalty' was lower than that in 'Flame' at T1, T2, and T4 but higher at T3. The expression of McMYB103 in 'Royalty' was also lower than that in 'Flame', except at the T1 stage. These results indicated the potential differences in anther and pollen development between the two cultivars. Moreover, the expression level of McSERK2 in 'Royalty' was higher than that in 'Flame' at all four stages. McSERK3/5 expression was lower in 'Royalty' than in 'Flame' at T1, T2, and T4 but higher at T3. The expression differences in these genes may explain the differences in zygotic embryogenesis between the two cultivars. Among the genes related to embryonic development, the expression of McSPL1, McSPL2, and McSPL4 was lower in 'Royalty' than in 'Flame' at all four stages; McSPL3 expression was lower in 'Royalty' than in 'Flame' except at T3; and McSPL5 expression was higher in 'Royalty' than in 'Flame' except at T3. With respect to the genes related to plant apomixis, McAPK and McFIE expression was lower in 'Royalty' than in 'Flame', except at T1, and McEF-alpha and McMSI expression was higher in 'Royalty' than in 'Flame', except at T3 (Fig. 5).
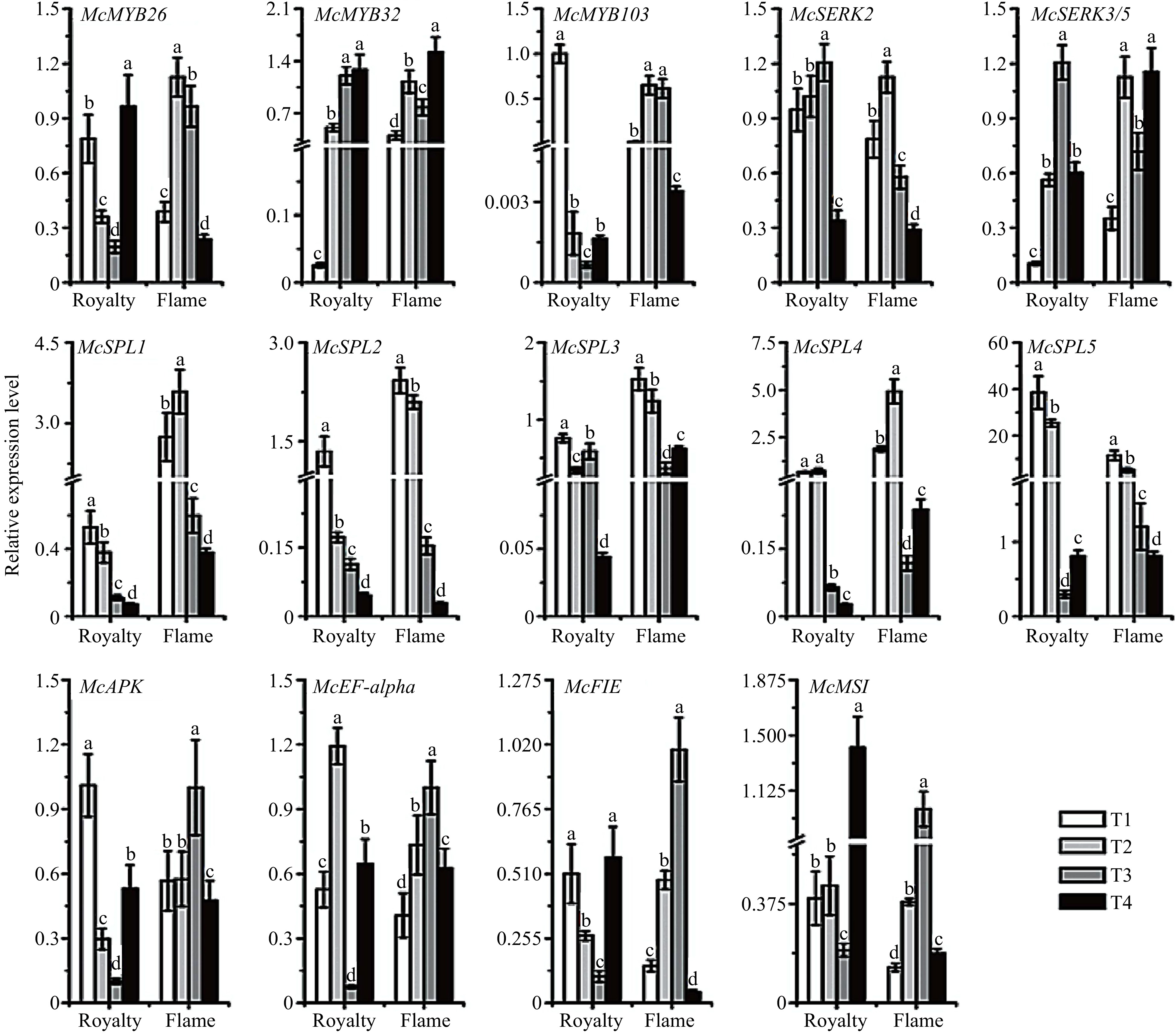
Figure 5. Relative expression profile of the genes related to apomixis in 'Royalty' and 'Flame' during flowering periods. T1-T4: flower organ developmental stages. The data (mean ± SD) are the sum of three biological repetitions for each treatment.
-
Apomixis is an important reproductive mode that has developed in plant species over long-term natural selection and evolution. This asexual reproduction pathway has been found in 400 species of flowering plants, providing clues to the diversity of plant apomictic reproduction[51,52]. In Asteraceae, Poaceae and Rosaceae, approximately 75% of the plants present gametophytic apomixis[53], which is highly beneficial to the fixation of hybrid vigour, application of the potential genotype resources and breeding of the rootstock. To date, a total of ten species of Malus have been found to exhibit apomixis, including M. sieboldii Rehd., M. hupehensis Rehd., M. platycarpa Rehd., M. rockii Rehd., M. sargentii Rehd., M. sikkimensis Koehne., M. coronaria (L.) Mill., M. toringoides Hughes., M. lancifolia Rehd., and M. xiaojinensis[54−56].
As shown in Fig. 2, we observed the developmental profiling of the megaspore of 'Royalty' EB and EFB ovules through paraffin sections. Our observations indicated that the megaspore mother cell of 'Royalty' from EB could develop from apospory initials, where the cavities in the centre of the nucellar provided space for the differentiation and formation of sporogenous cells, resulting in one of the sporogenous cells developing into a functional megaspore mother cell. In addition, tetrad- and functional megaspore mother cells did not formed via meiosis. Thus, it could be deduced that this megaspore mother cell might be an asexual cell formed by mitosis from EB. However, under EFB treatment, we found that some of the functional megaspore mother cells of 'Royalty' were sexual cells that formed via meiosis. These results demonstrated that 'Royalty' had facultative apomixis and could produce megaspores via apomeiosis or mitosis.
The fruit-setting rates from 'Royalty' EB (bagging after emasculation) were remarkably higher than the fruit setting rates from 'Royalty' BA (bagging alone), indicating that the apomixis ability of 'Royalty' was stronger than sexual reproduction under the stimulation of its pollen. Compared with other species of Malus, such as M. hupehensis and M. xiaojinensis, our results indicated that 'Royalty' had a higher capacity for apomixis[57]. In view of the high apomixis capacity and the characteristics of facultative apomixis in 'Royalty', this cultivar could be an interesting model plant for investigating the mechanism of apomixis and the enigmatic origins of apomixis in angiosperms.
Interestingly, as Fig. 1 and Table 2 suggested, we found that the F1-green-EFB plants were diploid (2x). We supposed that several possibilities exist for this result. First, and most likely, during the meiosis stage in the embryo sac mother cell of 'Royalty', an unbalanced distribution of chromosome numbers occurred, resulting in haploid egg cells, which performed sexual reproduction with haploid sperm derived from 'Flame' pollen, thus producing diploid fertile seeds and F1-green-EFB plants. Second, another less likely possibility is that male parthenogenesis occurred due to the invasion of diploid microspore mother cells derived from the pollen of 'Flame' entering the embryo sac of 'Royalty', which is male parthenogenesis, a form of gametophytic apomixis[58], thus resulting in diploid fertile seeds.
Apomixis may be a complement and substitute for sexual reproduction in Malus crabapples when encountering adverse reproductive environments
-
With respect to the most apomictic species, the reproduction modes of Malus are mostly facultative apomixis. Partial apomicts retain the capacity to sexually reproduce. Mráz and Zdvořák found that the mode of reproduction in Hieracium and other plants was tightly associated with ploidy. While diploids exclusively produced seeds sexually, polyploids produced seeds by obligate or almost obligate apomixis; certain polyploid plants were still able to reproduce asexually, although the reproductive mode was affected by environmental factors such as temperature and humidity[59−61].
The formation of an asexual megaspore is often triggered after abortion of the sexually produced megaspore. During ovule development in H. praealtum, H. rhamnoides and A. cordifolia plants, when the sexual pathway ceased, development of a functional megaspore mother cell was found to be the origin of a diploid aposporous initial cell, which differentiated, enlarged, underwent mitosis, and formed an aposporous megaspore that displaced sexual structures[62]. Conversely, the sexual reproduction pathway terminates during functional megaspore (FM) differentiation, when somatic aposporous initial (AI) cells undergo mitotic megaspore formation[28,63,64]. In the model plant Hieracium, the loss of the dominant LOA and LOP loci results in the reversion of the apomixis to sexual reproduction; and Juranić et al. proposed that in apomictic Hieracium, the contact between the AI and FM cell may be required for AI mitosis and megaspore degeneration[65]. These studies indicate that apomixis and sexual reproduction may be mutually complemented and perfected.
Consistently, the results of the pollination combination experiments showed that the 'Royalty' exhibited facultative apomixis. Moreover, we found that the offspring of apomixis in 'Royalty' showed weaker viability compared with the descendants from sexual reproductions. Furthermore, some of the offspring seedlings showed growth stagnation and death at a certain development stage. These results demonstrate that apomixis may be a complement and substitute for sexual reproduction in Malus crabapples which could be activated if sexual reproduction is blocked. Due to the repression of anther dehiscence, malformed pollen, and the low germination ability of pollen, 'Royalty' may adopt facultative apomixis to maintain genetic stability during the process of reproductive selection. Together, we proposed that apomixis and sexual reproduction may not be mutually exclusive but are complementary to guarantee genetic stability in plants.
Stimulation of the ovary by pollen may determine whether Malus crabapples adopt apomixis or sexual reproduction
-
In this study, as displayed in Table 1, we observed that when 'Royalty' was treated by bagging after emasculation (EB) and pollination followed by bagging (EFB), the fruit setting rate and seed number per fruit were adequate, and the offspring displayed good growth vigour in the pots compared with the diploid sister cultivar 'Flame'. During flowering, the chalaza nucellar cells exhibited vigorous division and growth, accompanied by the accelerated disintegration of peripheral cells in the ovary of 'Royalty' EB, leading to the formation of cavities in the central nucellus. Subsequently, when the sporogenous cells rapidly increased in volume, one of them developed into an unreduced megaspore mother cell. However, a tetrad developed via miosis was observed in the ovary of the 'Royalty' EFB. Therefore, through the anthesis pollination test and the observations of paraffin sections, we proposed that under no pollen stimulation (as in EB), the megaspore mother cells of the ovule did not undergo meiosis but underwent apomixis, resulting in the offspring retaining all of the maternal genetic information and displaying a phenotype similar to that of the mother plant. However, in the presence of pollen stimulation, the megaspore mother cells developed into diploid spores via miosis towards sexual reproduction. As shown in Fig. 6, we proposed a mechanism for apomixis and sexual reproduction in 'Royalty'.
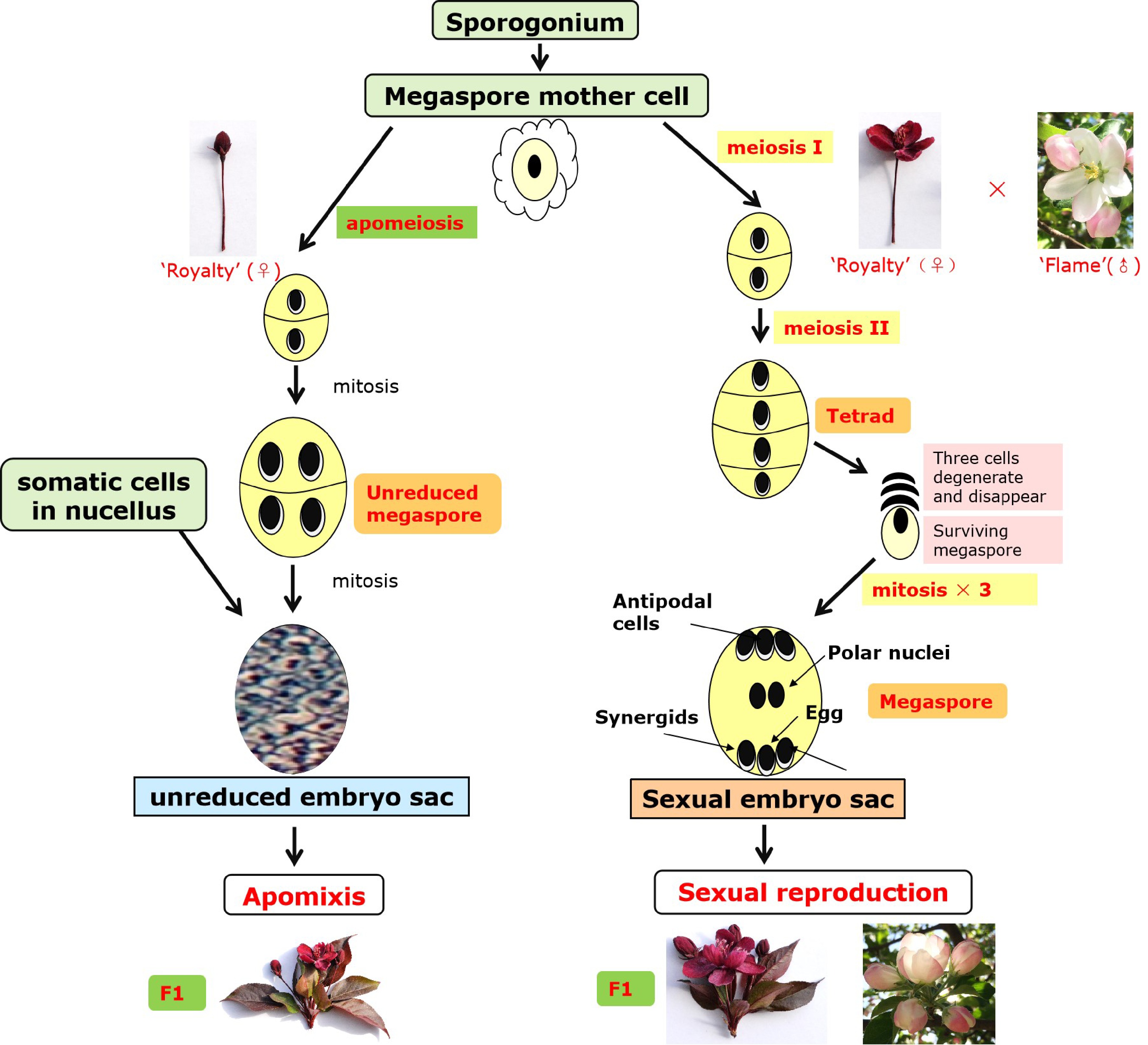
Figure 6. Mechanism of apomixis and sexual reproduction in 'Royalty'.
Most apple cultivars have a cross-compatible reproduction mode to form fruits and seeds, in which abundant anthers and pollen grains exhibit fertility, even in apomicts. However, 'Royalty' anthers during the flowering period were not readily dehiscent, and their pollen grains were small and infertile (Figs 3 and 4). These findings indicated that 'Royalty' exhibits a partial male-sterile pattern that is rare in Malus plants. However, this cultivar could form either a reduced megaspore by meiosis or an unreduced megaspore derived from nucellar cells by mitosis. Moreover, this cultivar could successfully produce fertile seeds. Accordingly, we speculate that the triploid 'Royalty' inherited similar features of facultative apomixis possessed by most members of the apple genus (polyploid), in which long-term self-incompatibility induced malformed anther development and the partial or complete abortion of pollen, which could lead to a trend towards greater obligate apomictic ability, preventing genetic transmission from the male parent. This process may be one of the important reasons why 'Royalty' and its descendants maintained the all-red phenotype.
Apomixis may be induced by the dysplasia of male organs and the low germination rate of pollen in 'Royalty'
-
Self-fertilization and apomixis have often been considered alternative evolutionary strategies of flowering plants that are advantageous for colonization scenarios and bottleneck situations[66]. Apomictic lineages usually evolve from sexually self-incompatible outcrossing plants, but pseudogamous apomicts frequently show a breakdown of self-incompatibility (SI). Additionally, apomictic plants with no viable pollen or with low pollen viability tend not to be visited by pollinators and may produce strictly apomictic fruits. In contrast, pollinators tend to visit apomictic and sexual plants with high pollen viability; in this case, apomictic plants may produce fruits and seeds by sexual and apomictic means[67,68]. These phenomena have often been seen in certain species of fruit plants, such as grape, citrus, and persimmon, as well as a large number of polyploid species of apple, pear, peach, and cherry[69]. Two possible pathways may explain this evolutionary trajectory: polyploidy may not only trigger gametophytic apomixis, but also result in a partial breakdown of SI systems. Alternatively, frequent pseudo-self-compatibility (PSC) via aborted pollen may induce the selfing of pseudogamous apomicts (mentor effects). Self-fertile pseudogamous genotypes will be selected within mixed sexual-apomictic populations because of the avoidance of interploid crosses[70,71].
Our findings suggested that the facultative apomixis capacity was closely associated not only with triploidy but also with the development of whole reproductive organs in flowers, as well as the self-incompatibility formed during the evolution of the species. Mao Baoqin et al. proposed that the loss of pollen was related to apomixis in their study of 20 species of Malus; additionally, other plants did not exhibit this phenomenon[72]. Based on our results, we propose the following explanations. Initially, apomixis might be induced because sexual reproduction is blocked or modified. Both the dysplasia of male organs and the low germination rate of pollen may have a direct impact on the efficiency of pollination, resulting in the blocking of sexual reproduction and the subsequent occurrence of the apomixis mode in 'Royalty'. This would indicate that the male gamete could influence the choice of a female gamete in reproductive patterns. Thus, female gametes may also affect the development of male gametes. Maruyama et al. proposed that each female gamete independently controls the avoidance of second pollen tubes to maximize reproductive fitness in flowering plants[73]. Another possibility is that the induction of apomixis may be the result of long-term adaptation and evolution in response to an adverse external environment. To maintain the good traits of the mother plant in 'Royalty', including the antioxidant ability achieved by abundant anthocyanins and other flavonoids within its red leaves, stems and fruit tissue, 'Royalty' developed the apomixis mode at the cost of the stunted or decreased vitality in male organs. However, the causal relationship and the induction mechanism of apomixis in 'Royalty' still need further study.
Considering the dehisce difficulty in anthers of 'Royalty', this characteristic can be used to develop male sterile lines and introduce into an apomict, thus dramatically accelerating plant breeding schemes. Additionally, mining the key genes regulating the cleavage of anthers would create a new path for preventing the dispersal and germination of pollen, thus leading to the induction or transfer of apomixis and preventing gene flow through pollen.
Related genes regulate apomixis at the transcriptional level
-
Apomixis is related not only to the development of functional megaspore mother cells and anthers as well as pollen grains but also to the genes involved in megaspore development[74,75]. Current opinion tends to support that apomixis develops by mutations or epigenetic modifications[76,77]. In the last few years, dozens of genes and transcription factors associated with apomixis have been identified in many plant species through mapping analysis, BAC sequencing, transcriptome sequencing, or amplified fragment length polymorphism (AFLP) analysis. In cultivated citrus, the expression level of CitRWP is relatively high in ovules of polyembryonic cultivars, and a miniature inverted-repeat transposable element insertion within its promoter region was discovered due to its cosegregation with polyembryony, a form of apomixis[78].
In the current study, we detected the expression level of those genes and transcription factors possibly associated with apomixis, including SPL (SQUAMOSA promoter-binding protein-like), MYB, FIE, MSI, SERK, and APK, which are mainly involved in plant organ development, meiosis and mitosis, megaspore development and seed formation[79,80]. In Arabidopsis, the expression levels of SERKs were relatively high during the early stage of embryo somatic cell development and increased in undeveloped ovules and unfertilized megaspores; once fertilized, their expression levels sharply declined, and the levels in the nucleus cells near the functional megaspore mother cell were also high during the apomictic process[81,82]. FIE and MSI are associated with endosperm development. The Arabidopsis mutant msi, which requires pollination to produce endosperm, forms a complex with FIE, which inhibits endosperm occurrence and development[32]. As suggested in Fig. 5, our results indicated that the expression of genes associated with apomixis is relatively high in the bud stage in 'Royalty', such as McAPK and McFIE. This may be because the plants are facing reproductive selection during the bud stage because the plants are more susceptible and sensitive to the internal and external environment or the lack of pollen stimulation. When flowers bloom, 'Royalty' plants receive pollen stimulation and thus have the requirement of sexual reproduction, and then the plants tend to exhibit a sexual reproduction pattern, resulting in the decreased expression of McAPK and McFIE.
SPLs are cell development-related genes that are expressed in megasporocytes and microsporocytes and participate in regulating male fertility and gynoecium differential patterning[83,43]. MYB transcription factors are closely related to pollen development, anther tapetal layer development, anther rupture, stigma callus formation, and photosynthate transportation during pollen development[42]. Preston et al. found that changes in the expression levels of AtMYB32 and AtMYB4 may influence pollen development by diverting the flux into the phenylpropanoid pathways, thereby affecting the composition of the pollen wall[44]. Previous studies suggested that EgMYB4 possibly restrained lignin biosynthesis in oil palm flowers, allowing enhanced carbon flux into the phenylpropene pathway. Interestingly, McMYB26 exhibited high expression at the bud stage in 'Royalty' and showed an opposite expression trend between 'Royalty' and 'Flame', indicating that McMYB26 may be involved in apomixis in 'Royalty'. This may augment our understanding of the role MYBs play in fine-tuning the metabolic flux along the various branches of the core phenylpropanoid pathway in male and female organ development.
-
Six-year-old Malus cv. 'Royalty' and M. cv. 'Flame' trees grafted onto M. hupehensis (Pamp.) Rheds with similar growth potential and flowering features were collected from the Beijing University of Agriculture crabapple germplasm collection, which is located in the Shunyi District (Beijing, China, latitude 40°00′–40°18′, longitude 116°28′–116°58′). This area has sandy soil and a temperate continental subhumid monsoon climate with four distinct seasons. The annual average temperature is 11.5 °C, the amount of annual sunshine is 2,746 h, the annual relative humidity is 58%, the frost-free period is approximately 195 days, and the annual average rainfall is 610 mm. The flowering time of the two cultivars is approximately April 5th to April 15th. We selected four floral development stages for further analysis: T1 was the budding stage (approximately 6 days before full bloom), T2 was the initial flowering stage (approximately 3 days before full bloom), T3 was the full bloom stage, and T4 was the end of the flowering stage (approximately 3 days after full bloom).
Pollination experiments
-
From 2012 to 2014, we performed our bagging pollination experiments when more than 60% of all flower buds on the trees were about to open at the T2 stage (3 days before full bloom). Eight different pollination treatments were designed as follows: 1) 'Royalty' EB (sulfuric acid paper bags were used for bagging 'Royalty' flowers after artificial emasculation); 2) 'Royalty' BA (sulfuric acid paper bags were used for bagging 'Royalty' flowers to exclude the influence of foreign pollen); 3) 'Royalty' EFB ('Royalty' flowers received the pollen from 'Flame' after emasculation); 4) 'Royalty' CK (natural flowers and natural pollination of 'Royalty' without any treatment); 5) 'Flame' EB (sulfuric acid paper bags were used for bagging 'Flame' flowers after emasculation); 6) 'Flame' BA (sulfuric acid paper bags were used for bagging 'Flame' flowers to exclude the influence of foreign pollen); 7) 'Flame' EFB ('Flame' flowers received the pollen from 'Royalty' after emasculation); and 8) 'Flame' CK (natural flowers and natural pollination of 'Flame' without any treatment).
Anthers were collected at the bud stage (approximately 8−10 days before early flowering) and placed in a sulfate paper box at 20−25 °C. The pollen was collected into a glass bottle to avoid the loss of water. For EB, we removed the whole stamens and then bagged the flower in a sulfate paper bag. For BA, the flowers were bagged with sulfate paper without emasculation. For EFB, stigmas of flowers were pollinated promptly with prepared pollen grains via artificial hand pollination after being emasculated. The bags were replaced after 15 days. The fruit setting rate and seed number per fruit were counted at 90 days after treatment.
Stratification and seeding
-
The fruits obtained from the different pollination treatments were collected and stored at 4 °C for stratification. After three months, the seeds were removed from the fruit. The seeds were washed twice in warm water at 55 °C to remove impurities. The washed seeds were soaked in warm water at 55 °C and removed when the seed coat changed colour and absorbed enough water. Then, the seeds were wrapped in wet gauze and soaked in wet sand with carbendazim in a dark place at room temperature for approximately 7−10 days. Distilled water was sprayed on the sand to keep it moist every day. Afterwards, the budding seeds in the soil were sown with nutritive soil, vermiculite and perlite at a ratio of 2:2:1.
Seed tests
-
The treated fruits were harvested after maturation in autumn. The seeds were removed from the fruits and then dried at room temperature. In February of each year, the treated seeds were selected and stored in a refrigerator at 4 °C for stratification. After the seed coat was broken, the percentage of seed germination was measured. A single seed was directly sown into the pots with a 2:2:1 mixture of vermiculite, perlite and sandy soil at approximately 80% of soil water capacity. When seedlings grew to 5 cm high, the percentage of seedling germination was measured. Whether visible white and transparent germs appeared under visual observation was regarded as the determination criterion for seed germination. The presence of two symmetrical cotyledons off the ground was set as the determination criterion for seed sprouting.
The ploidy of seedlings was determined with flow cytometry according to the instructions of the Partec CyStain UV Precise T Reagent Kit (PARTEC, Cod. 05-5003). First, 50 mg of the young leaves from mature seeds were collected and placed into a capsule with a 5 cm diameter and chopped with a razor blade in 1 mL of nuclei extraction buffer containing 15 mmol/L Tris-HCl, pH = 7.5, 80 mmol/L KCl, 20 mmol/L NaCl, 20 mmol/L EDTA-2Na, 15 mmol mercaptoethanol and 0.05% Triton-100 (v/v), followed by the addition of another 1 mL of nuclei extraction buffer. Subsequently, nuclear suspensions were filtered down to 5 mL in a centrifuge tube through a 300-µm nylon mesh to remove debris and centrifuged at 12,000 rpm/min for 10 min. After the supernatant was discarded, 400 μL of 50 mg/mL staining buffer (3,000 U/mL RNaseA and 10 μg/mL PI (propidium iodide) was added; then, the solution was fully mixed and placed in dark conditions for staining for 15−20 min at 4 °C. Later, ploidy was measured using a 15 mW 488 nm argon-ion laser-based flow cytometer (BD FACSCalibur, USA). At least three different plants for each treatment were measured, and the analysis of each sample analysis was repeated three times.
Morphological observations of the anthers, pollen grains and megaspores
-
At each of the four developmental stages, bagged flowers with bags of both Malus cultivars under all treatments were collected. When observing the megaspore, only the ovary was retained. These samples were quickly placed into formalin-acetic acid-alcohol (FAA; 50% ethanol, 5% acetic acid, 3.7% formaldehyde, 41.3% water) and placed in cold storage at 4 °C. Anthers of all treatments were collected and placed in the drying vessel at 20−25 °C. When the CK plants were blooming, anthers and pollen grains were removed and fixed in the above FAA fixative. Plant materials were embedded in paraffin according to standard methods. Thin paraffin sections were cut from the paraffin block with a sharp two-sided blade under an anatomical lens. The thin material sections rolled up when they were cut off. The section rolls were placed on a slide, which was heated to melt the paraffin section roll. When the paraffin melted, the sections of plant material were rolled out according to standard procedures for staining and mounting[84]. The samples were observed and documented by using a light microscope (Olympus AX70TRF, Olympus Optical, Japan) equipped with a U-photo system and a digital camera.
For ultrastructural observation, the collected anthers and pollen grains were analysed by using a Quanta 200 low-vacuum environmental scanning electron microscope (ESEM) (FEI Company, Oregon, USA) equipped with a tungsten filament (FEI). The signal was collected by using a gaseous secondary electron detector (GSED) at 15 kV. All images were observed and provided valuable information about the ultrastructure of the anthers and pollen grains. Additionally, the ESEM images obtained were analysed using the MATLAB (MATrix LABoratory) software program® v.7.1 (Lab valorization and Safety of Food Products, Beni Mellal, Morocco) to determine the surface structure of anthers and pollen grains.
Pollen grain viability and germination rates
-
To evaluate pollen viability, one or more anthers were placed on the centre of the glass slide. Then, a drop of distilled water was added, and the anthers were squeezed gently with tweezers to release the pollen grains. Afterwards, one to two drops of I2-KI solution were added, and the samples were covered with a coverslip. After staining for 1 min at room temperature, we observed the dyed pollen by using a Leica DM2500 microscope (Leica, Wetzlar, Germany) equipped with an ultraviolet filter.
The in vitro pollen germination test was performed as follows. Briefly, the collected pollen grains were placed on pollen germination medium (1 mmol/L CaCl2, 1 mmol/L Ca(NO3)2, 1 mmol/L MgSO4, 0.01% (w/v) H3BO3, and 18% (w/v) sucrose, solidified with 0.5% (w/v) agar, pH 7.0) and cultured in a constant-temperature incubator at 25 °C. After 3 h of incubation, digital images were collected by using a Leica DFC420 CCD camera attached to a Leica M165FC stereoscope (Leica, Wetzlar, Germany). The pollen grain germination percentages and lengths of the pollen tubes were measured using ImageJ software. Protrusion from the aperture was regarded as positive germination.
Differential expression analysis and RNA-Seq library preparation
-
'Royalty' and 'Flame' fruits at 20 days and 90 days after the blooming period were sampled for RNA extraction following the manufacturer's instructions (Total RNA Extraction Kit with Polysaccharide Polyphenol for Plant was purchased from TIANGEN BIOTECH, item no: DP441); 1% agarose gel electrophoresis was used to detect whether the RNA was degraded. A Nano Photometer® spectrophotometer (IMPLEN, CA, USA) and an RNA Nano 6000 Assay Kit with the Bioanalyzer 2100 system (Agilent Technologies, CA, USA) were used to determine RNA integrity and purity, respectively. A Qubit® RNA Assay Kit in a Qubit® 2.0 A fluorometer (Life Technologies, CA, USA) was used to measure the RNA concentration. RNA sequencing (RNA-Seq) was performed using the Illumina HiSeq 2000 platform. The raw reads were filtered to obtain clean reads and exclude those adapter related, low quality, containing N (unable to determine the base information). The de novo assembly of the transcriptome was carried out by Trinity[85], which was used as a reference sequence or unigene for subsequent analysis.The following public protein databases were considered: NCBI nonredundant protein sequences (Nr) database, NCBI nucleotide sequences (Nt) database, Protein family (Pfam) database, Clusters of Orthologous Groups of proteins (KOG/COG) database, Swiss-Prot protein database (Swiss-Prot) database, Kyoto Encyclopedia of Genes and Genomes (KEGG) orthologue database, and Gene Ontology (GO) database. With the transcriptome obtained by Trinity as the reference sequence (ref), the clean reads of each sample were mapped to the reference and the readCount number of each gene was further obtained. RSEM software was used for this process[86]. The unigene expression level was normalized by estimating reads per kilobase per million (RPKM), which takes into account the effects of sequencing depth and gene length on the count of read count[87]. Differential expression analysis of the four groups provided statistical evidence for determining the differences in digital gene expression data; the fold change and P-values were adjusted by using the Benjamini and Hochberg approach for selecting the genes that were considered to be differentially expressed.
Gene cloning and quantitative real-time polymerase chain reaction (PCR)
-
At each of the four developmental stages (T1, T2, T3, and T4), only the ovaries of both Malus cultivars were immediately frozen in liquid nitrogen and stored. Total RNA was extracted according to the manufacturer's instructions (TIANGEN, item no: DP441). Highly pure RNA samples (5 µg) with a ratio of 260/280 nm > 1.9 were DNase treated (TIANGEN) and reverse transcribed into complementary DNA (cDNA) by using the oligo(dT)18 primer and M-MLV reverse transcriptase (TaKaRa) following the manufacturer's protocol. Primers were designed according to the CDS of homologous genes of the apple genome, and the PCR primer sequences are listed in Supplemental Table S2. The full-length cDNAs of candidate genes were amplified, and all PCR products were subcloned into the pGEM T-Easy Vector (Promega, Madison, WI, USA) and transformed into Escherichia coli DH5α cells and sequenced.
The synthesized cDNAs were then diluted for quantitative real-time reverse transcription PCR (qRT-PCR) analysis using qPCR Mix (TaKaRa) in conjunction with a Bio-Rad CFX96 Real-Time PCR System (BIO-RAD). The differences and relative quantification of gene expression were calculated through the 2−ΔΔCTanalysis method[88]. The Malus 18S ribosomal RNA gene was used as the reference gene. The primers used for qRT-PCR are listed in Supplemental Table S2.
Statistical analysis
-
All data were analysed by using one-way ANOVA followed by Duncan's SSR test (shortest significant ranges) test to compare differences among the experimental sites at P < 0.05. At least three biological replications were performed for the quantitative analysis, which included the transcriptomics study, phenotypic measurements, quantitative assays and qPCR expression analysis. Microsoft Excel 2007, OriginPro8 and Data Processing System (DPS) software 7.05 were used for analysis.
- This work was financially supported by grants from the National Natural Science Foundation of China (31901997, 31772263, 31872081), the General Project of the Scientific Research Program of the Beijing Municipal Commission of Education (KM202010020013), the Opening Project of Beijing Key Laboratory of New Technique in Agricultural Application (NYYYXJS201704), the National Key Project of Research and Development Plan (2016YFD0201116), and the Construction of Beijing Science and Technology Innovation and Service Capacity in Top Subjects (CEFFPXM2019_014207_000032). The funding institutions were not involved in conducting this study. We would like to thank the Beijing Nursery Engineering Research Center for Fruit Crops for providing experimental resources and the Beijing Collaborative Innovation Center for Eco-environmental Improvement with Forestry and Fruit Trees.
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Phenotypes of the fruits and seeds of Malus cultivars ‘Royalty’ and ‘Flame’ (Bar=1 cm). (a) Seeds after different treatments; (b) F1 seedling phenotypes (c-d). R X F, ‘Royalty’ pollinated with ‘Flame’ pollen; F X R, ‘Flame’ pollinated with ‘Royalty’ pollen; R-Bag, ‘Royalty’ bagged alone; and R-Emasculation, ‘Royalty’ bagged after emasculation.
- Supplemental Table S1 Oligo primer sequences used for gene amplification and qRT-PCR to determine the gene expression of Malus cultivars ‘Royalty’ and ‘Flame’.
- Supplemental Table S2 Statistics of the seed sprouting rate and seed germination rate under different pollination treatments.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hu Y, Lu Y, Chao L, Wang Z, Bu Y, et al. 2021. Dysplasia of male organs induces apomixis in Malus crabapples. Ornamental Plant Research 1: 12 doi: 10.48130/OPR-2021-0012

Dysplasia of male organs induces apomixis in Malus crabapples
- Received: 19 September 2021
- Accepted: 14 December 2021
- Published online: 27 December 2021
Abstract: Apomixis has high value in fruit crop propagation and breeding applications, particularly for the transgenerational fixation of heterosis. In this study, we found that flowers of the triploid Malus cultivar 'Royalty' that were bagged after emasculation (EB) and pollinated with pollen from the diploid cultivar 'Flame' (EFB) produced high fruit set and increased seed numbers per fruit. The offspring exhibited superior growth vigour compared to those of the diploid sister cultivar 'Flame'. The fruit setting rate for 'Royalty' EFB was 98.44%, which was much higher than the fruit setting rate for 'Royalty' BA (Bagging without emasculation) at 19.7%. The fruit setting rate for 'Royalty' EB was 47.46% compared to 19.7% for 'Royalty' BA. Furthermore, offspring derived from either 'Royalty' EB or EFB exhibited superior growth vigour compared to those of the diploid 'Flame'. These results suggested that 'Royalty' exhibits facultative apomixis and has strong sexual reproductive ability under the stimulation of foreign pollen. Additionally, 'Royalty' anthers did not dehisce after flowering and had malformed pollen grains and a low pollen germination rate. Therefore, we proposed that dysplasia of the male organ could be an important driver factor of apomixis in 'Royalty'. Our study provides new insight into the mechanism of apomixis in Malus crabapples.