








HTML
-
Shellfish, including mollusks (e.g., oysters and mussels) and crustaceans (e.g., shrimp and crab), have been a cornerstone of healthy dietary recommendations[1]. There were 17.7 million tons of edible mollusks and 9.4 million tons of crustaceans in aquaculture worldwide in 2018[2]. In China, the total output of oyster products was 5.14 million tons[2], and yield and consumption of oyster have been increasing each year. Increasing shellfish consumption to achieve adequate nutrient intake can also increase the threat of food allergies, which may be sufficiently high to be of concern to life[3].
Allergy to shellfish is a long-lasting disorder and effective clinical management focuses on its avoidance. Social impacts of food avoidance and dietary limitation often have negative effects on the quality of life of the sensitized subjects[4]. Many papers subsequently reported that processing procedures may modulate the allergenic properties of foods[5,6]. Food processing has therefore gradually become the new hope for allergic individuals. Maillard reaction (MR) is a non-enzymatic glycation reaction that takes place between reducing sugars and proteins, which is common in food processing for it can form desirable flavors and attractive color compounds in food products[7,8]. It has been reported that MR could reduce the allergenicity of many crustacean allergens[9,10], while the effects of MR on oyster allergen has not been revealed, which has limited the improvement and further practical application of MR-mediated oyster allergen immunoreactivity elimination.
Oyster allergy is caused by allergens, including tropomyosin[11] and sarco-plasmic-calcium-binding protein (SCP)[12]. A novel allergen in Portuguese oyster (Crassostrea angulata) was reported in 2020 by our team and named as Cra a 4 by the WHO/IUIS (
http://allergen.org/ ). The previous research was mainly in the physi-co-chemical properties, allergenicity, cross-reactivity and epitopes of Cra a 4[12,13]. Meanwhile, the low yield and complex purification processing of natural Cra a 4 are restricting further research for hypo-immunoreactivity processing[12]. A standardized recombinant Cra a 4 is urgent in the food processing industry.The allergenicity of food allergens are related with digestion, structure and epitopes. MR could reduce the allergenicity of Scy p 4 (SCP in Scylla paramamosain) by changing the structure[10], which is the same family as Cra a 4[14]. MR could reduce the allergenicity of tropomyosin (TM) in scallop by modifying the amino acid (AA) residues[15]. Liu et al reported that the heat/digested stable epitope peptides of Scy p 4 remain as an allergen in processed crab[16]. It is worth noting that Cra a 4 showed obvious heat and digestion stability[12], its structure and immunological characteristics do not change significantly even if heated at 100 °C. Hence, common thermal processing could not reduce its immunobinding activity. There is doubt regarding the relationship of MR and the immunoreactivity for Cra a 4, and the mechanism is still unknown.
The present study investigated the allergen Cra a 4 in C. angulata, and the aim was to explore the influence of MR on Cra a 4 and to confirm the reason for the hypo-immunoreactivity processing. Firstly, Cra a 4 was cloned, expressed, verified and obtained. Subsequently, the reaction time of MR were optimized between Cra a 4 with xylose and glucose, respectively. Furthermore, the digestibility and immunoreactivity of the glycoconjugates were evaluated. Then the spatial structure was analyzed in silicon using circular dichroism (CD) spectroscopy and surface hydrophobicity. BThe AA residues in the primary structure were analyzed to explain the mechanism of MR. The results of the present study provide new insight into the desensitization of oyster-induced allergy.
-
As shown in Fig. 1a, the positive clone after PCR showed 100% identification with the gene of Cra a 4 by GENEDOC, which is a highly integrated software for molecular biology. Sequence alignment indicated that the present cloning was suitable and effective in Fig. 1a. Subsequently, the induced time was considering for the optimization of expression conditions. Compared with the other induced times, there were greater amounts and intensity of strips in Lane 1 (induced time was 4 h) shown in Fig. 1b. Meanwhile, protein in Lane 1 presented higher concentrations than the other lanes using a Bradford assay kit and protein in Lane 1 showed stronger IgG binding activity with rabbit anti Cra a 4 pAb (Fig. 1c), which indicated that the suitable time was 4 h for the present study.

Figure 1.
Expression, purification and identification of Cra a 4. (a) Base sequence alignment analysis of sequencing and template. (b) Optimization of the expression conditions for Cra a 4 after induced by isoprophyl-β-D-thiogalactoside using SDS-PAGE. (c) Optimization of the expression conditions for Cra a 4 after induced by isoprophyl-β-D-thiogalactoside using Western blotting. Lane M, protein marker; Lane 1, the supernatant of cell lysate after being induced for 4 h; Lane 2, the precipitate of cell lysate after being induced 4 h; Lane 3, the supernatant of cell lysate after being induced for 6 h; Lane 4, the precipitate of cell lysate after being induced for 6 h; Lane 5, the supernatant of cell lysate after being induced for 8 h; Lane 6, the precipitate of cell lysate after being induced for 8 h; Lane 7, pET-22b vector. (d) Purification of Cra a 4 by Ni2+-NTA resin. Lane M, protein marker; Lane sample, the super-natant of cell lysate after being induced 4 h; The other numbers 4, 8, 22, 23, 24, 25, 26 and 29 on the top of the lanes correspond to the fraction number. (e) SDS-PAGE of purified Cra a 4. (f) Western blotting verification of purified Cra a 4 by rabbit anti Cra a 4 pAb. (g) IgE binding activity of Cra a 4 by ELISA with sera. NC-1 and NC-2: the sera of non-atopic individuals were used as the negative control. S1−S12 were the sera of oyster sensitive individuals.
Cra a 4 was the expressed as a soluble protein in E. coli BL 21 and purified by a Ni2+-NTA resin in Fig. 1d. An obvious peak was present and a single strip with a molecular weight of 24 kDa at 25 and 26 tubes on the elution curve. The tubes were then collected and analysed by SDS-PAGE and Western blotting as shown in Fig. 1e & f. As expected, the protein band near 24 kDa showed obvious IgG binding activity with Cra a 4 pAb, while the negative control-BSA did not react with Cra a 4 pAb, which means that the purified protein near 24 kDa was confirmed to be Cra a 4.
Importantly, the purified protein was verified with oyster sensitized sera shown in Fig. 1g. The purified Cra a 4 showed obvious IgE-binding activity with 12 sensitized sera and did not react with the two nonallergenic sera.
Optimization of MR reactions of Cra a 4
-
The types of reducing sugar, including arabinose, xylose, galactose, mannose and glucose, were optimized to evaluate the effect of MR on the allergen (data not shown). Compared with Cra a 4, the bands of all samples after thermal processing were dispersed. During the same conditions, the production of xylose glycation and glucose glycation showed weaker IgG-binding activity than the others. Xylose and glucose were then chosen as the reducing sugars in MR for the following research.
Subsequently, the reaction time was optimized (Fig. 2). The main bands of gly-coconjugates were degraded gradually as the reaction progressed and persisted, meanwhile, more strips with a molecular mass less than 24 kDa appeared (Fig. 2a & b). The degraded bands showed IgG binding activity with Cra a 4 pAb (Fig. 2c−f), while the strength of binding activity was gradually decreased. Quantitative analysis of IgE-binding activity of MR products showed that the immunoreactivity of MR products decreased with the extension of processing time. For the production of glucose glycation, the IgE-binding activity did not decrease significantly when the processing time reached 60 min. Similarly, when the processing time reached 30 min, the IgE-binding activity of production of xylose glycation did not decrease significantly (Fig. 2g & h).
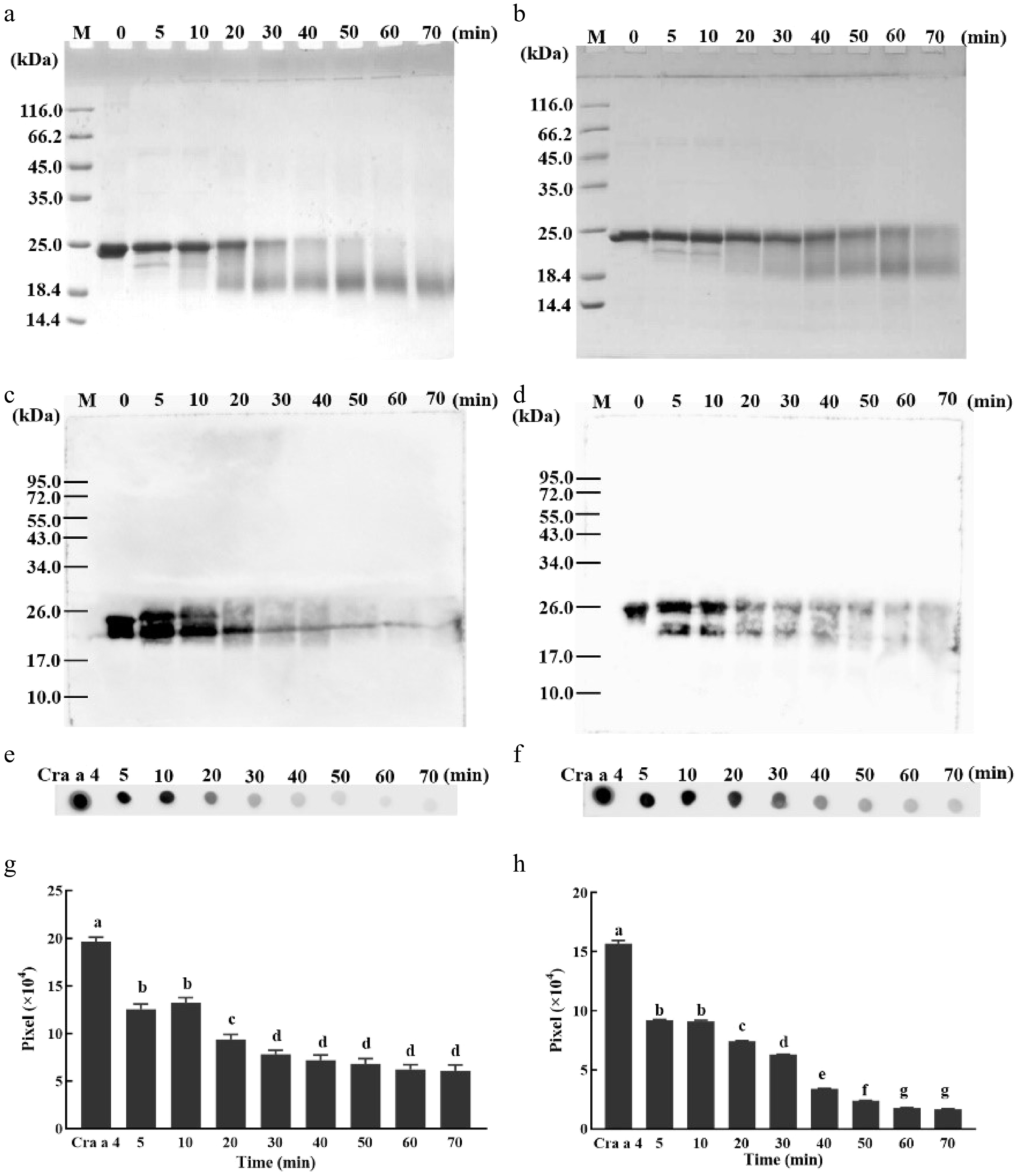
Figure 2.
Optimization of the reaction time for MR. (a) SDS-PAGE profile of Cra a 4 incubated with xylose for different time periods. Lane M, protein marker; The other numbers 0, 5, 10, 20, 30, 40, 50, 60 and 70 on the top of the lanes represent the different reaction times. (b) SDS-PAGE profile of Cra a 4 incubated with glucose for different time periods. Lane M, protein marker; The other numbers 0, 5, 10, 20, 30, 40, 50, 60 and 70 on the top of the lanes represent the different reaction times. (c) Western blotting of Cra a 4 incubated with xylose for different time periods. Lane M, protein marker; The other numbers 0, 5, 10, 20, 30, 40, 50, 60 and 70 on the top of the lanes represent the different reaction times. (d) Western blotting of Cra a 4 incubated with glucose for different time periods. Lane M, protein marker; The other numbers 0, 5, 10, 20, 30, 40, 50, 60 and 70 on the top of the lanes represent the different reaction times. (e) Dot blotting of Cra a 4 incubated with xylose for different time periods by rabbit anti Cra a 4 pAb. Lane M, protein marker; The other numbers 0, 5, 10, 20, 30, 40, 50, 60 and 70 on the top of the lanes represent the different reaction times. (f) Dot blotting of Cra a 4 incubated with glucose for different time periods by rabbit anti Cra a 4 pAb. Lane M, protein marker; The other numbers 0, 5, 10, 20, 30, 40, 50, 60 and 70 on the top of the lanes represent the different reaction times. (g) The intensity of the dots shown in part (e). The quantification of grayscale dots was analyzed using ImageJ software. (h) The intensity of the dots shown in part (f). The quantification of grayscale dots was analyzed using ImageJ software.
The optimized MR conditions were heated at 100 °C for 60 min (pH 8.5) with glucose and at 100 °C for 30 min (pH 8.5) with xylose.
Digestibility in the gastric passage
-
It is a general consensus that digestibility plays an important role in the allergenicity for food allergens[17]. During the same digestion conditions, Cra a 4 were less stable than glycoconjugates as shown in Fig. 3. The main band of Cra a 4 were degraded gradually as the digestion time increased and it disappeared after digestion at 30 min (Fig. 3a). Interestingly, the bands in the glycoconjugates showed different results. The glycoconjugates of xylose were more stable than of glucose and the main band did not show an obvious degradation tendency, even after digestion for 60 min (Fig. 3b). However, compared with glycoconjugates of xylose, the glycoconjugates of glucose were more easily digested (Fig. 3c). Most bands disappeared at 30 min and the main band near 20 kDa was stable at 60 min.
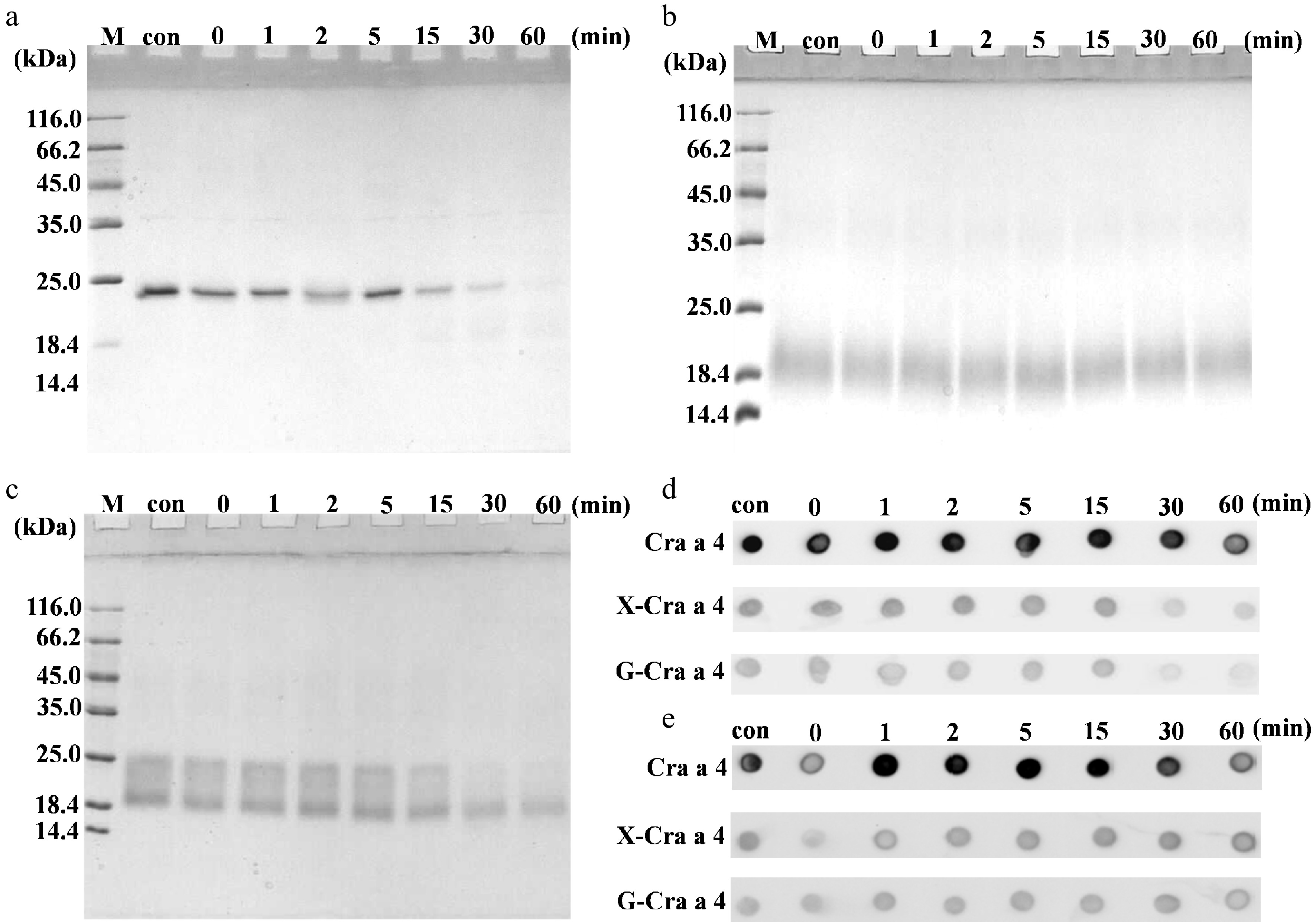
Figure 3.
Protein profiles of the glycoconjugates after MR during the simulated gastric digestion in vitro. (a) SDS-PAGE of the purified Cra a 4. (b) SDS-PAGE of the glycoconjugates between Cra a 4 and xylose. (c) SDS-PAGE of the glycoconjugates between Cra a 4 and glucose. (d) IgG-immunoblot assay of the digested samples by rabbit anti Cra a 4 pAb. (e) IgE-immunoblot assay of the digested samples by the specific sensitized sera pool. Lane M, protein marker. Lane con, the samples after processing and before pepsin digestion. The other numbers 0, 1, 2, 5, 15, 30 and 67 on the top of the lanes represents the different digestion times. X-Cra a 4, the glycoconjugates between Cra a 4 and xylose. G-Cra a 4, the glycoconjugates between Cra a 4 and glucose.
To gain more information about the immunoreactivity after MR, Dot blotting with Cra a 4 pAb and sensitized sera pool was carried out to follow the changes during the digestion process. Compared with Cra a 4, the intensity of dots decreased significantly for the glycoconjugates before and after digestion (Fig. 3d & e). This means that MR could reduce significantly the IgG/IgE-binding activity of Cra a 4.
Secondary structure, surface hydrophobicity and sites of amino acid (AA) analysis
-
Protein structure is crucial for the allergenicity for food protein, while the crystal structure of shellfish SCP-family has not been reported. The secondary and tertiary structure of Cra a 4 and glycoconjugates were researched and shown in Fig. 4. Cra a 4 showed a positive peak and two negative peaks at less than 225 nm, showing typical features of α-helical proteins (Fig. 4a). Interestingly, α-helix content was significantly reduced in glycoconjugates with transformations in the peak values accompanied by flattening of the spectrum. As shown in Fig. 4b, the height of glycoconjugates fluorescence absorption peaks was obviously higher than that in Cra a 4. The glycoconjugates between Cra a 4 and xylose has a strong absorption peak, indicating that its surface hydrophobicity is high. Furthermore, the content of α-helix up to 64.25% and the exposed region up to 59.78% in Cra a 4 in silicon (Fig. 4c).
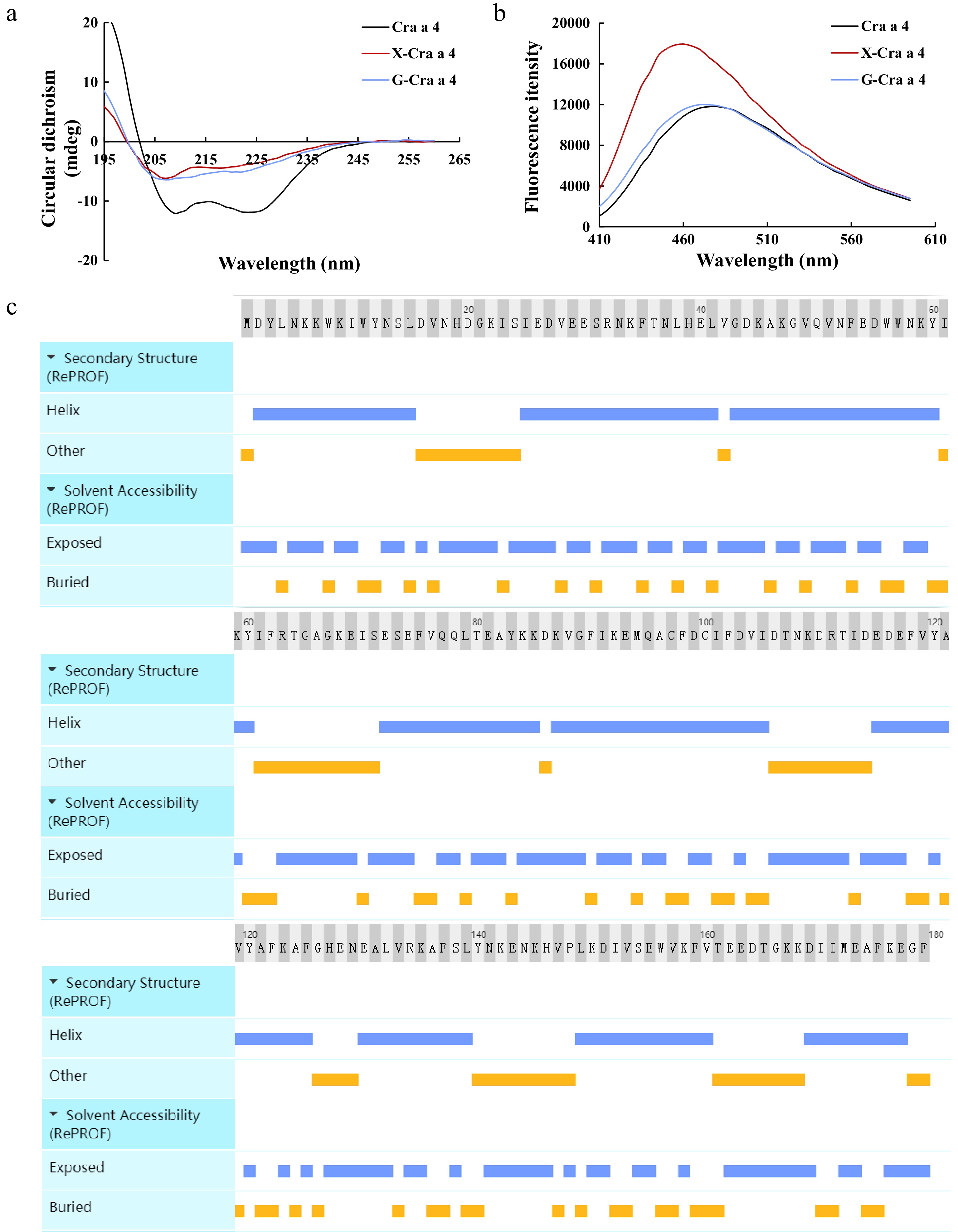
Figure 4.
Analysis of the structure in Cra a 4 and the glycoconjugates. (a) Secondary structural analysis of the glycoconjugates after MR. (b) Surface hydrophobicity analysis of the glycoconjugates after MR. (c) Secondary structure and solvent accessibility of Cra a 4 in silicon. X-Cra a 4, the glycoconjugates between Cra a 4 and xylose. G-Cra a 4, the glycoconjugates between Cra a 4 and glucose.
The results in Fig. 5 could infer that MR could significantly change the protein structure of Cra a 4. During the frequency of AAs (Fig. 5a), lysine (K), glutamic acid (E), aspartic acid (D), phenylalanine (F), valine (V), isoleucine (I) and asparagine (N) performed higher frequency occurrence in the Cra a 4 sequence. Interestingly, there were no potential N-glycosylation sites in the AA sequence of Cra a 4, which was identified by the conserved motif N-X-S/T/C10. It was different to Scy p 4 in crab, the same protein family, and the sequence identification is 17.62% between Cra a 4 and Scy p 4 (Fig. 5b) may be explain the difference. There were 34 possible cleavage sites of Cra a 4 by pepsin (pH 1.3) which were predicted by the program ExPASy peptide cutter. While parts of the pepsin cleavage sites overlapped with the epitopes of Cra a 4 and Scy p 4, the identified epitopes of Cra a 4 and the heat/digestion stable epitopes were mapped as shown in Fig. 5b.

Figure 5.
Analysis of amino acid sequence in Cra a 4. (a) The distribution frequency of amino acids in Cra a 4. (b) Alignment of amino acid sequence of SCP in C. angulate and S. paramamosain. The linear epitopes of Cra a 4 are marked in blue; the heat/digested epitope regions of Scy p 4 are marked in red; the pepsin-cutting sites of Cra a 4 are marked by amino acid residues in blue.
Purification and identification of recombinant Cra a 4
-
Food allergies today represent a major public health burden, with a continuous growing prevalence[18]. IgE-mediated food allergy is primarily responsible for fatal anaphylactic reactions[19]. The harsh truth is that there are no preventive medications for IgE-mediated shellfish allergy. The primary treatment option is strict avoidance of the causal food; while accidental ingestion is often inevitable, especially in modern society with highly processed food[20]. In this context, food processing with hypo-immunoreactivity is of great interest for the sensitized individuals and the public[21]. As for the novel allergen in oyster, Cra a 4[12], the morbidity of Cra a 4 allergy is lacking, while its risk cannot be overlooked. The research of food processing with hypo-immunoreactivity for Cra a 4 is urgent and important.
The present research initially purified Cra a 4, the protein near 24 kDa showed obvious immunoreactivity with rabbit anti oyster Cra a 4 pAb and oyster sensitized sera, which was coincides with previous research[12]. Even though the molecular mass of natural Cra a 4 is 20 kDa, the existence of His-tags increased the molecular weight and it did not affect the characters of expressed allergens[22]. Furthermore, there were no obvious differences both in recombinant and natural Cra a 4[12]. It is worth noting that the presence of heat/digestion resistant epitopes is closely associated with the structure of allergens[23]. Based on the sequence of Cra a 4, there were more helix content (64.25%) than the other structure type in silicon, which was consistent with the results by CD spectrum. This means that the structure predicted in silicon is useful for protein, which has been widely used by peers[24,25]. The content of solvent-exposed structure (59.78%) in Cra a 4 indicated that most of the hydrophobic regions were centered on the surface of the protein[26], which increased the surface hydrophobicity and added a level of complexity to Cra a 4. In short, the spatial structure of Cra a 4 was in agreement with previous reports[12].
MR is one of the well-known non-enzymatic chemical reactions between amino acids and reducing sugars[8], which ensues frequently in food following thermal processing or after long-term storage[27]. MR has shown to be promising in mitigating mollusc and crustacean allergenicity[15,27], while MR could be influenced by various factors, including the structural diversity of (poly)saccharides and proteins, temperature and time[27]. The species of protein influences the distribution of a products' molecular weight. Maleki et al.[28] reported the different molecular weight distribution of the glycoconjugate between WPE and glucose, Ara a h 1 and glucose. Similarly, the species of sugars influences the distribution of products' molecular weight. And it has been identified that the molecular weight of the glycoconjugate between Cra a 4 and xylose were lower than the glycoconjugate between Cra a 4 and glucose. In the present study, MR product profiles of the five-carbon sugars (xylose) and Cra a 4 were different with the six-carbon sugars (glucose, mannose, galactose), which was consistent with the other recent research[15], the reactivity of the five-carbon sugars in MR was faster than that of six-carbon sugars. Furthermore, the IgG binding activity reduced significantly in a relatively short timeframe (30 min) in the MR products of five-carbon sugars (xylose) rather than of the six-carbon sugars (glucose, 60 min). And it was speculated that the degree of glycosylation increased as the carbohydrate decreased in size[29]. All the bands after MR appeared as smears on SDS-PAGE, which was consistent with the previous research, and the reason was owning to the cross-linking and non-cross-linking adducts formed in MR[28]. Importantly, extended high temperature treatments of MR might also influence the conformational modifications of the structures of allergens, which would change the binding activity of antibodies from accessing antigenic epitopes and enhance the allergenicity of protein[27,30]. In present research, 30 min of xylose and 60 min of glucose were chosen as the optimal reaction time for MR.
It is well known that resistance to proteolysis in vivo has been proposed to be a prerequisite for a protein to sensitize via the mucosal immune system in food allergy[17]. For ethical reasons in vitro digestibility has been used widely[31]. In the present investigation, Cra a 4 and the glycoconjugates of glucose were digested more easily, although the glycoconjugates of xylose was more stable to digestion. It could be speculated that the saccharides with a smaller molecular size of five-carbon sugars (xylose) had less steric hindrance and greater accessibility for pepsin[15]. Besides an assessment of the pepsin digestibility, the present study has, in addition, evaluated the residual allergenicity of the emerging digestion product after MR. Although Cra a 4 appeared to be more easily digested, the IgG/IgE-binding activity of the small peptides retained a high potential for allergenicity, which was consist with the previous study of Scy p 4[10]. Meanwhile, the possible reason might be the existence of heat/digestion stable epitopes in Cra a 4, which has been proven in Scy p 4[16].
The helical content was decreased and the surface hydrophobicity was increased for the glycoconjugates, which coincides with the results for Scy p 4[10]. The fluorescence intensity of the glycoconjugates by xylose was significantly higher than that of glucose, which was speculated to be due to the fact that glycoconjugates by xylose may expose more hydrophobic regions to the protein surface. Meanwhile, there was a strong negative correlation between pepsin activity and protein surface hydrophobicity[32], which is due to the more stable digestion tendency for xylose (shown in Fig. 3).
In general, MR has been believed to decrease the allergenicity of food as it leads to the disruption of allergens or participates in modulation of the food allergy[8]. It is known that basic AA residues (such as lysine and arginine) participate in the MR of food allergens, while the other AA residues have been recently showns to play an important role in MR, including occasionally histidine and cysteine residue[8]. Interestingly, the frequency of K was higher than the other AA residues in Cra a 4 sequence, and most of them were exposed on the surface (see Fig. 4a), which means that MR might be an effective method to modify the AA residues and change the structure/allergenicity[21]. It has been proven that MR could unfold the α-helix, increase the surface hydrophobicity and reduce the immunoreactivity for Cra a 4. Meanwhile, the disulfide bond (C97 and C100) was somewhat beneficial for MR[13]. Besides, most of the pepsin cleavage sites and 2 Ca2+-binding sites (located in AA16-27 and AA106-117) were exposed on the surface of Cra a 4, which helps to reduce the immunoreactivity in the digestion samples.
According to previous reports, allergenicity could not be eliminated completely by MR. In the present research, the MR digestion samples presented weaker IgG/IgE-binding activity by immunological techniques than that of Cra a 4 (shown in Fig. 3), which indicated that the heat/digestion stable linear and conformational epitopes were retained after MR[33]. Compared with the conformational epitopes, the linear epitopes play a more important role in food allergy as most food must be ingested after thermal processing[34]. Han et al identified five linear epitopes of Cra a 4, and the region of epitopes (AA80-90, AA107-116 and AA144-159) was partially overlapped with the heat/digestion stable epitopes of Scy p 4[13]. Though the identity between Cra a 4 and Scy p 4 was low, the existence of the overlapped region indicated that the heat/digestion stable epitopes of Cra a 4 may exist as they are the same protein family. Then the unidentified stable epitopes could explain the reason for the lower immunoreactivity after MR.
-
In conclusion, MR could reduce the immunoreactivity of Cra a 4. The immunoreactivity of glycoconjugates between Cra a 4 and five-carbon sugars (xylose) for 30 min, and the six-carbon sugars (glucose) for 60 min, could be reduced significantly. The changing of spatial structure and the modifying of AA residues exposed on the surface might be the reason for the hypo-immunoreactivity of the MR products. The present study clarified the mechanism of MR decreasing the immunoreactivity of oyster to a certain extent, and provided new methods for the industrial production of hypoallergenic oyster. In the future, the exploration of the heat/digestion stable epitopes in Cra a 4 is important for the seafood processing industry and oyster sensitized individuals.
-
Live oyster (C. angulata) was purchased form Jimei Market in Xiamen, China, and the transportation procedures were similar with the previous report[16]. Briefly, live oysters were transported to the laboratory within approximately 30 min. The oysters were handled by placing in a plastic bucket filled with seawater.
Goat anti-human immunoglobulin (Ig) E horseradish antibody, pepsin (≥ 250 U mg−1), 8-anilino-1-naphthalenesulfonic acid (ANS) were from Sigma Aldrich (Sigma Aldrich, St Louis, USA). Goat anti-rabbit IgG horseradish antibody were purchased from Southern Biotech (Birmingham, Alabama, USA). Arabinose, xylose, galactose, mannose and glucose were obtained from Macklin (Shanghai, China). Rabbit ant Cra a 4 polyclonal antibody (pAb) were prepared in our lab[12]. All other reagents were analytical grade.
Sera of oyster-sensitized individuals
-
All procedures for human sera collection and manipulation were performed in accordance with the Guidelines for Care and Use of Women and Children's Hospital Affiliated with Xiamen University (Xiamen, Fujian, China), and the experiments were approved by the Human Ethics Committee of the review board (approval No. KY-2018-018 and KY-2019-014). All the volunteers or parents of participating children provided informed consent. Oyster-sensitized people were selected based on their clinical history and a positive IgE test to the natural Cra a 4 that was prepared following previous methods[12]. Absorbance > 0.10 were regarded as positive according to Liu et al.[16]. Sera from 12 oyster-sensitized individuals and two nonatopic individuals were collected for the present study (Table 1).
Table 1. Specific IgE levels and symptoms of the oyster-sensitized individuals.
Sera No. Age Sex OD450 a Symptoms NC-1 b 24 M 0.0608 − c NC-2 b 23 F 0.0611 − c S1 12 M 0.1082 Cough S2 8 F 0.1576 Anaphylactic rhinitis S3 9 F 0.1295 Atopic dermatitis S4 9 M 0.1004 Chronic urticaria S5 7 F 0.1051 Chronic urticaria S6 8 M 0.1062 Allergic purpura S7 6 F 0.1013 Bronchitis S8 10 M 0.1089 Acute tonsillitis S9 6 M 0.1040 Atopic dermatitis S10 4 M 0.1491 Urticaria S11 11 M 0.1067 Atopic dermatitis S12 3 F 0.1033 Atopic dermatitis a A serum with specific IgE≥0.10 is defined as positive.
b A nonallergic individual.
c Means no symptoms at the time of the experiment.
M, male; F, female.Cloning, expression and purification of Cra a 4
-
The protocols of gene cloning were similar with the previous study[12], the positive clones were obtained by PCR. The full-length PCR products were purified, collected and sequenced by Borui BIO Co. Ltd. (Xiamen, Fujian, China).
Escherichia coli BL21 harboring pET-22b-Cra a 4 (70 μL) was inoculated in a LB liquid preculture (500 mL) containing ampicillin (500 μL) and incubated overnight at 37 °C at 200 rpm. Subsequently, isoprophyl-β-D-thiogalactoside induced the expression strain for 4 h, 6 h and 8 h at 37 °C. The three samples were then centrifuged at 10,000 ×g for 10 min at 10 °C, the sediment cells were collected and resuspended with 4 mL buffer. Cells were broken up by ultrasound treatment and subjected to centrifugation and filtration (filter membrane size was 0.22 μm) to obtain the supernatant. The substance was resuspended with 2 mL buffer. The supernatant and substance were collected for further analysis, and the pET-22b without cells were used as control.
After the expression conditions were optimized, the target protein was purified using a Ni2+-NTA resin according to Chen et al.[14]. The purified protein was stored at 4 °C prior to analysis. The concentrations of all protein samples were determined using the Bradford assay kit (Bio-Rad Laboratories, Hercules, California, USA).
Optimization of MR conditions
-
Maillard reaction was performed as per the method of Hu et al.[10], with modifications: samples were mixed at the Cra a 4-to-reducing sugars (arabinose, xylose, galactose, mannose, glucose) ratio of 1:4 (w/w) and incubated at 100 °C, pH 8.5 for 30 min. The products were dialyzed against 20 mm PBS (pH 7.4) at 4 °C for 16 h, and then stored at −30 °C until use for the chosen sugar species in MR.
In addition, the reaction time was also optimized. Briefly, the reaction times were set as follows: 0, 5, 10, 20, 30, 40, 50, 60 and 70 min. Other conditions were same as above.
Gastric digestion in vitro
-
Samples after MR were digested according to the published protocols[16]. In brief, 4.5 mg of MR products were dissolved in 12 mL of pre-warmed simulated gastric fluid (pH 2.0). Pepsin was added (enzyme-to-substrate ratio, 1:50 w/w) at pH 2.0, 37 °C with continuous shaking. The digestion tubes were periodically withdrawn at 0, 1, 2, 5, 15, 30, and 60 min and stopped by raising the pH to 7.4 with NaOH. The digestion samples were stored at 4 °C prior to analysis.
Sodium dodecyl sulfate polyacrylamide gel electrophoresis and immunoreactivity analysis
-
All the samples, including protein and MR products, were analyzed by 10-well 12% and 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels[35].
For Western blotting, the rabbit anti Cra a 4 pAb (dilution 1:10,000) as the primary antibody was used as previously described[12]. For Dot blotting, the primary antibody was Cra a 4 pAb (dilution 1:10,000) and human sera pool (the satisfactory sera were mixed in equal volume, dilution 1:5) using the method of Bai et al.[15]. For ELISA, the satisfactory sera (dilution 1:5) was used as the primary antibody[35].
CD spectroscopy
-
The secondary structure of Cra a 4 and MR products was measured by CD spectrum (Applied Photophysics Ltd., Surrey, UK) as per the published method[36]. The final concentration of all samples was adjusted to 0.25 mg/mL.
Surface hydrophobicity analysis
-
The surface hydrophobicity of Cra a 4 and MR products was determined using ANS as the fluorescence probe using the method of Hu et al.[10]. The final concentration of samples was adjusted to 0.25 mg/mL.
Secondary structure and solvent accessibility in silicon
-
RePROF predicts secondary structure elements and solvent accessibility using evolutionary information from multiple sequence alignments and a multi-level system[25]. Secondary structure and solvent accessibility of Cra a 4 were predicted by a system of neural networks with a window size of 17 on the web (
https://predictprotein.org/ ).Amino acid sites in the primary sequence of Cra a 4 analysis
-
The composition and occurrence frequency of AAs in Cra a 4 (GenBank accession number: MN956521) were calculated according to Fu et al.[37].
The pepsin-cutting sites of Cra a 4 were analyzed using the program ExPASy peptide cutter according to the method[38]. Pepsin (pH 1.3) is preferentially chosen and the lowest cleavage probability is 100% for the present study. Alignment of amino acid sequence of SCP in C. angulate and S. paramamosain were used by ClustalW (
www.clustal.org/clustal2 ).Statistical analysis
-
Each experiment was repeated more than three times, with all results expressed as the mean ± standard deviation. Analysis of variance was used to assess differences between related groups using the Statistical Product and Service Solutions statistics software (SPSS for Windows, IBM Corp., New York, USA) and p < 0.05 was considered to be statistically significant.
Materials
This work was supported by a grant from Xiamen Ocean Vocational College (KYG202108), a grant from the young and middle-aged teachers’ education research project of Fujian Province (JAT210801, JAT210803), the National Natural Scientific Foundation of China (32072336, 31871720, 32001695, 31901811), the science and technology program of Fujian Province (2021L3013), the science and technology program of Xiamen City (3502Z20203059), and grants from the National Key R&D Program of China (2019YFD0901703).
-
The authors declare that they have no conflict of interest.
-
These authors contributed equally: Meng Liu, Fei Huan
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
Liu M, Huan F, Zhang J, Huang L, Yu L, et al. 2022. The immunoreactivity of Cra a 4 decreased after Maillard reaction with xylose and glucose. Food Materials Research 2:13 doi: 10.48130/FMR-2022-0013


|