










-
Tea products are a popular drink worldwide, due to their rich flavors and important health benefits[1]. Tea products are mostly made using the apical buds and young leaves. The apical buds and young leaves of tea plants are always covered by a high density of trichomes, which has become the sign of tenderness and high-quality of raw materials, since tea trichomes only present on the young tissues, and accumulate most tea characteristic specialized metabolites, particularly volatiles[2−4]. Our previous study has clearly demonstrated that whole sets of biosynthetic genes for tea characteristic metabolites, such as catechins, caffeine, theanine, and terpenoids are expressed in tea trichomes[2]. Therefore, those characteristic compounds detected in tea trichomes were likely synthesized inside the trichome. Those trichomes could contribute to the tea quality, especially the tea aroma, in famous green and white teas, such as Biluochun, Queshe, Xinyangmaojian, which are made from only the apical buds with longer and densely spaced trichomes[2−5].
The trichomes are special type epidermic cells and always appear on the plant buds, leaves, stems, or fruits with diverse structures and functions[6,7]. The tea trichome is unicellular, unbranched, and has non-glandular type of surface hairs[8,9]. The plant trichomes are regarded as the first physical barriers for plant adaption to various environments[10,11]. The trichomes containing more lignin and various types of toxic compounds, such as purine alkaloids, flavonoids and complex terpenoids, are the natural barriers for most herbivores, microbial pathogens, and other biotic stresses[2,12,13]. The trichomes also play crucial roles in responding to drought, high temperature, and cold stresses by reducing transpiration and water loss[14,15]. The presence of densely spaced trichomes on plant organs' surfaces could also provide a layer of obstacle for cold or high temperature transmission from the environment to plant epidermal cells[14,15]. The trichomes also highly expressed many light-inducible genes, providing photoprotection for the plants[16,17]. For instance, trichomes not only physically reflect high-light or UV-B radiation to reduce these light-induced cell damage, but also highly express light- responsive and signaling genes, such as UV RESISTANCE LOCUS 8 (UVR8), ELONGATED HYPOCOTYL5 (HY5), and early light-induced protein 2 (ELIP2), and antioxidant flavonoid biosynthesis genes[8−12, 14]. As the unicellular trichomes, tea trichomes appeared very differently to other unicellular trichomes, such as Arabidopsis and cotton trichomes[2]. Tea trichomes are white and soft with no branches, and appear on the abaxial side of the young leaves[2]. Previous transcriptome analysis has generally indicated the defense roles of trichomes in tea plants[2,4]. However, not much is known about the exact roles of trichomes in protecting the apical buds and young leaves in adverse environments, such as high light, UV-B radiation, cold stress, insect attack, microbial pathogen infection, and other abiotic or biotic stresses, as well as the regulation of tea trichome initiation.
The initiation of unicellular trichomes, such as Arabidopsis trichomes, was finely regulated by positive and negative transcription factors (TFs)[8]. The R2R3-type MYB GLABRA1 (GL1) is the central TF, and positively regulates the trichome formation, via forming a MYB-bHLH-WD40 (MBW) complex with other helix-loop-helix and WD40 repeat (WDR) proteins[18,19]. The MBW complex directly activates the downstream HD-Zip TF GLABRA2 (GL2) to initiate trichome formation[18,20]. In tea plants, some TFs, such as CsMYB1, CsWD40, and CsGL3, have been identified to be involved in trichome initiation by forming MBW complex[21−24]. The MBW complex also activates many repressors, such as R3 MYB TFs CAPRICE (CPC) and TRIPTYCHON (TRY), those repressors are then transferred into neighboring cells to repress the MBW activation on the GLABRA2 (GL2) and trichome initiation[18]. In tea plants, at least six CPC-like genes were identified in the tea genome[25]. One CPC genes have been well characterised in trichome formation, which could competitively bind to CsWD40 and CsGL3 to repress CsGL2 expression and tea trichome formation[21].
Our previous study has demonstrated that the trichomes contribute to tea quality via producing and accumulating tea characteristic metabolites[2]. Here we further conducted in silica analysis on the transcriptome of trichomes and trichome-removed leaves to obtain in-depth the data related to tea plant defense functions and performed wet experiments to further understand the exact physiological roles of trichomes in tea plant resistance against insect attack, high-temperature and UV-B radiation stresses. This study provides fresh insights into the tea trichome functions in defense against multiple abiotic and biotic stresses.
-
The developmental leaves were isolated from Anhui #9 cultivar on June 2021, and then imaged using a camera. The different tissues of C. sinensis (L.) Kuntze 'Fudingdabai' (trichome enriched variety) grown at Guohe Tea Plant Garden (Anhui Agricultural University, Anhui, China) were harvested for transcriptome analysis during the spring season of 2018. Tea plant varieties for trichome phenotype survey and the assay of insect resistance, grown in the Guohe tea plant garden (Anhui Agricultural University), Dongzhi tea plant resource garden (Dongzhi, Anhui, China) during the summer of 2018 to 2019 and Jinhua International Camellia Species Garden of Zhejiang province (Jinhua, Zhejiang, China), were investigated for their trichome phenotypes.
UV-B and high temperature treatments
-
For UV-B radiation treatment, the 1.5-year old tea seedlings grown in a growth chamber were used for UV-B or regular light treatments as described previously[26]. For temperature stress on tea seedlings, the 2.5-year old tea seedlings were treated in a growth chamber set at 35 °C and 22 °C (as control) with regular 8/16 h dark-light period over various time periods.
Bioinformatic analysis
-
The transcriptome data of tea plant tissues and different varieties were as described in our previous report[2]. The trichome phenotype was manually investigated using ImageJ software. The resulting heatmap was structured by pHeatmap R package or Tbtools software[27,28].
Transcriptome data analysis
-
The transcriptome data of trichomes and trichome-removed leaves were generated in our previous study[2]. The shading experiments were described previously, two treatments of tea plants with natural sunlight (control) and tea plants with 90% sunlight blocked by covering black net treatment (shading) under the same temperature and growth conditions[27,28]. The first and second leaves were collected throughout treatments (0 h, 4 h, 8 h, 12 h, 2 d, 4 d, 8 d, and 14 d). For the cold treatment experiments, tea plant leaves were collected during the cold acclimation (CA) process. Control (CK): 25 °C; CA1-6 h: 10 °C for 6 h; CA1-7 d: from 10 °C to 4 °C for 7 d; CA2-7 d: from 4 °C to 0 °C for 7 d; DA-7d: recover under 25 °C to 20 °C for 7 d, as described previously[27,28]. Transcriptomic data from various experimental treatments were retrieved from previous studies and the Tea Plant Information Archive (
http://tpia.teaplant.org/index.html ). Quantitative reverse transcription-polymerase chain reaction (RT-PCR) was performed essentially as previously described[26,27] for validation or examination of expression patterns of key genes, with primers listed in the Supplemental Table S1. -
The young tea leaves are used for processing of high-quality commodity tea, especially the apical buds, which contain more amino acids and less bitter compounds, for high quality green tea, such as Xinyangmaojian, Biluochun, Queshe[2]. The apical buds are the young curled leaves, and always fully covered with signal cell type trichomes (Fig. 1a; Supplemental Fig. S1). It takes approximate 7 d from apical bud to full expansion leaf, and during this time, as the outer tissue, the trichomes are directly exposed to the environment to act as physical barriers, playing crucial roles in protecting the vulnerable apical buds from stresses, especially the high light and excess ultraviolet-B (UVB), until the leaf matures (Fig. 1a). Tea plants grow in high-altitude areas, with high light and UVB always become the main abiotic stress for the young leaves, especially the vulnerable apical buds. Tea trichomes are white and soft with no branches and no chlorophyll, and display strong birefringence under high light due to high cellulose content (Fig. 1b). In Arabidopsis, the trichome birefringence genes (TBR) play crucial roles in the cellulose biosynthesis and trichome birefringence[29]. We identified many CsTBR genes highly expressing in the trichomes (TR) compared with trichome-removed leaves (LE) (Fig. 1c; Supplemental Fig. S2), and cumulative expression analysis also showed that the TBR family genes highly expressed in the TR compared with LE (Fig. 1d). Thus, more TBR gene expression in tea trichomes maybe contributed to reflecting high light further to reduce the harmful light for leaves.
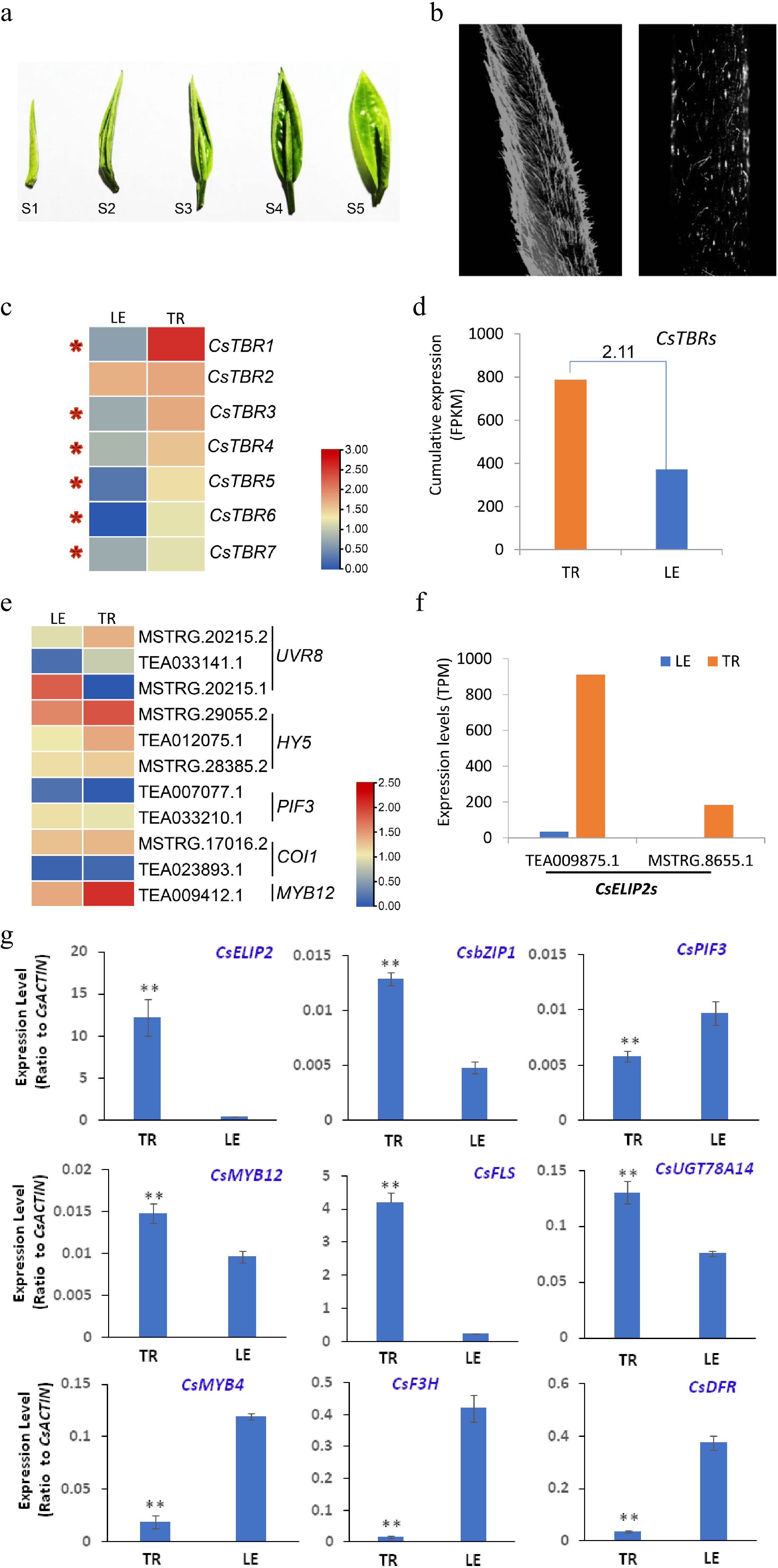
Figure 1.
Tea leaf trichomes play crucial roles in the photoprotection process. (a) Morphology of developing tea leaves. (b) Tea leaf (left) and stem (right) trichomes displaying strong birefringence under high light. (c) The expression pattern of trichome birefringence (TBR) in trichomes (TR) and LE (leaf with trichomes removed). The expression levels (log10) of representative genes were displayed using a heat map. (d) Cumulative expression analysis of the TBR gene family in TR and LE. The fold change (TR/LE) of gene family expression level is also listed on the bar. (e) The expression pattern of genes related to high light or UVB response in TR and LE. The expression levels (log10) of representative genes were displayed using a heat map. (f) The expression levels of two CsELIP2 transcripts in TR and LE. (g) qRT-PCR validation of expression patterns of genes in defense response and secondary metabolism. Data were from three experiments (n = 3) and expressed as means ± s.d. and the significant differences between comparisons were analyzed by using Student’s t-test in a two-tailed comparison (* P < 0.05 and ** P < 0.01). Gene expression data were retrieved from transcriptome data and expressed as FPKM or TPM. Heatmaps for gene expression were determined with these transcriptome data sets. qRT-PCR were conducted to verify the expression patterns of key genes involved in light signaling and defense response, as well as flavonoid biosynthesis.
Our previous study has showed that in tea plants, the UVR8- HY5 work module also plays crucial roles in the response to UVB exposure[26]. UVR8 is the photoreceptor that initiates photomorphogenic responses underlying acclimation and UVB tolerance in plants, which could be activated by UVB, and subsequently activates the expression of down central TF HY5[30,31]. Three UVR8 isoforms generated by alternative splicing events were identified in our RNA-seq data. Interestingly, two isoforms (MSTRG.20215.2 and TEA033141.1) highly expressed in the TR, and another (MSTRG.20215.1) highly expressed in the LE, suggesting the TR and LE responded to the UVB through different UVR8 isoforms (Fig. 1e). HY5 is a central photomorphogenesis-promoting factor and directly regulates thousands of downstream genes involved in development and metabolism processes[32]. Three HY5 transcripts were identified in the transcriptome data, which were all highly expressed in TR compared with the LE (Fig. 1e). HY5 can directly activate MYB12 expression to regulate flavonol biosynthetic genes to produce more flavonol compounds in tea plants[26]. Consistently, we also found the flavonol regulator CsMYB12 highly expressing in the TR compared with LE (Fig. 1e). PHYTOCHROME INTERACTING FACTOR3 (PIF3) and CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1) were two repressors of HY5 to oppositely regulate plant development and metabolism[26,33,34]. Two PIF3 and two COP1 transcripts were respectively identified in our transcriptome data. The two PIF3 transcripts showed equal expression levels in TR and LE, as did COP1 (Fig. 1e). Two transcripts homology to Arabidopsis chloroplastic ELIP2, CsELIP2 (TEA009875.1) was expressed more than 30 times higher in the TR than in LE, likely acting as a photo-protectant to reduce the damaging effects of high light and UV-B (Fig. 1f)[35,36].
Validation of transcriptome data with qRT-PCR was conducted with several representative genes involved in light signaling and high-light stress response, as well as flavonoid biosynthesis and regulation. The high-light response gene CsELIP2, flavanol biosynthesis genes CsFLS and CsUGT78A14, were expressed in TR at much higher levels than in LE, which is consistent with transcriptome data and the protective role of trichomes on high-light stress response, as well as higher flavanol levels in trichomes than in leaves per dry mass[2]. However, CsPIF3, and CsMYB4 were expressed at higher levels in LE than in TR (Fig. 1g; Supplemental Fig. S2). They are involved in light-regulation of flavonoid biosynthesis in tea plants with either positive roles or negative feedback effects on flavanol biosynthesis genes[26]. Also consistent with metabolite profiling, catechins are accumulated at much higher levels in LE than in TR[2], CsF3H and CsDFR were expressed at higher levels in LE than in TR (Fig. 1g; Supplemental Fig. S2).
Tea plant trichomes highly expressed cold-related genes
-
The functions of tea trichomes in the cold stress response is not fully characterized. Our previous study has comprehensively explored the transcriptome variation of tea plants under cold stimulation (Fig. 2a). Like other plants, cold treatment could acutely trigger the CBF gene expression in tea plants, suggesting that the CBF pathway also plays an important role in cold resistance for tea plants (Fig. 2b). Unexpectedly, we found the expression of HY5 and downstream TF MYB12 was significantly up-regulated by cold stress (Fig. 2c). Subsequently, the expression of FLS regulated by MYB12 and HY5 was also strongly induced (Fig. 2c). However, the expression of UVR8 was not significantly up-regulated, suggesting that HY5 expression was activated by cold stress through other factors (Fig. 2c). Meanwhile, the expression of COP1 and PIF4, two HY5 repressors, was slightly up-regulated by cold stress (Fig. 2c). Also, the expression of CsELIP2 was strongly induced by cold stress (Fig. 2c). Glycosyltransferases (UGTs) play critical roles in cold stress in tea plants via promoting flavonoids glucoside or nerolidol glucoside accumulation. CsUGT78A14 could increase flavonols accumulation and ROS scavenging capacity to enhance cold resistance for tea plants[37]. In our RNA-seq data, the expression of CsUGT78A14 was also strongly induced by cold stress and was more than six times higher in the TR than the LE (Fig. 2c−d), which was consistent with the high expression of flavonol regulator CsMYB12 genes in TR (Fig. 1b). Some volatiles such as nerolidol glucoside, were shown to involved in the response to cold stress as well as in triggering plant-plant communication in response to cold stress[38]. Tea UGT91Q2 could catalyze nerolidol to form nerolidol glucoside with higher capacity of ROS scavenging, promoting the enhanced tea plant cold stress tolerance [38]. In our RNA-seq data, the expression of CsUGT91Q2-1 and CsUGT91Q2-2 was reliably strongly induced by cold stress (Fig. 2c). In the TR, both CsUGT91Q2-1 and CsUGT91Q2-2 genes were highly expressed compared with LE (Fig. 2e). Those data demonstrated that the tea trichomes also play important roles in cold stress tolerance via highly expressing cold stress related compounds and genes (Fig. 2f).
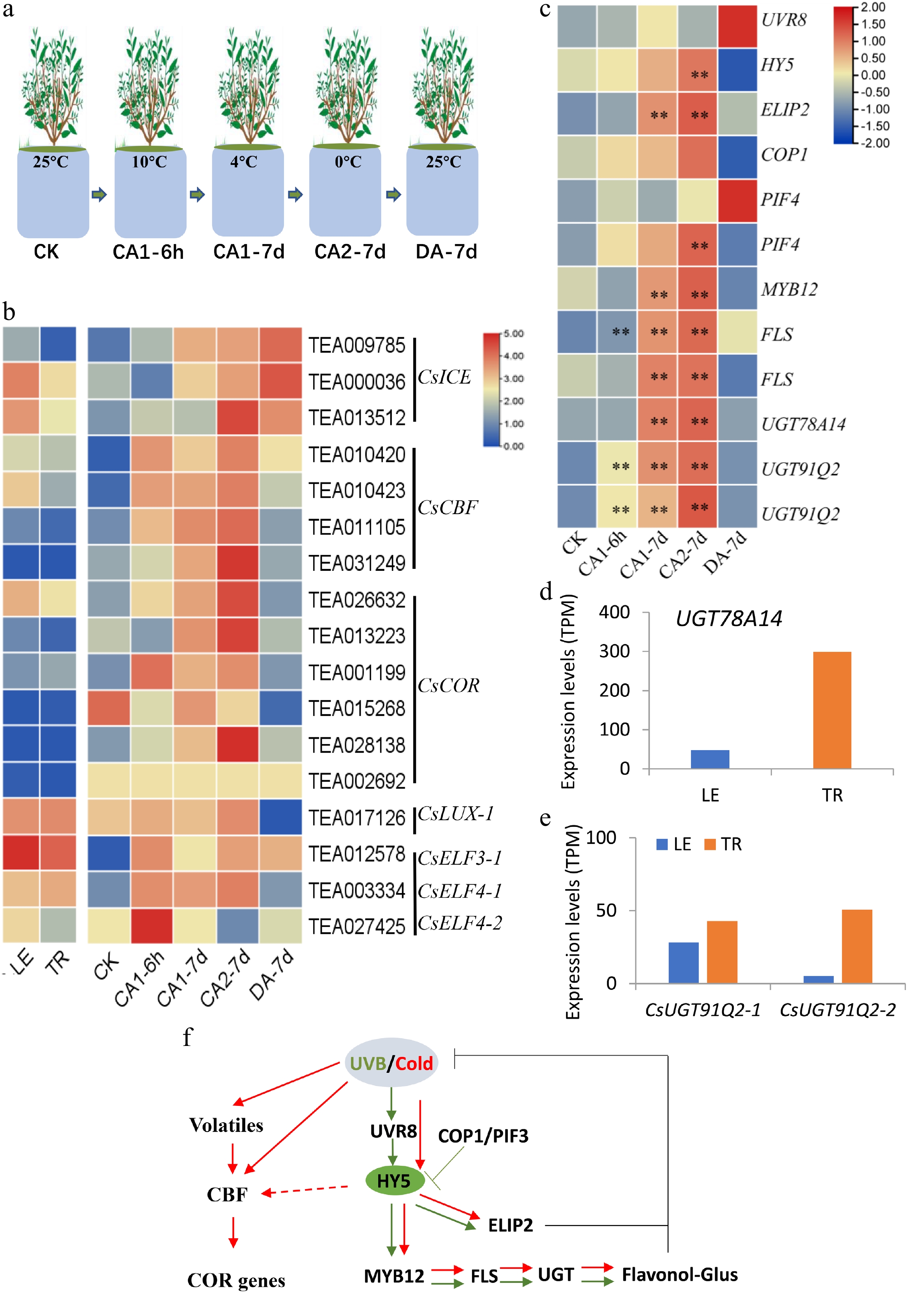
Figure 2.
Tea leaf trichomes play crucial roles in cold stress. (a) Images for tea plants under cold stress treatment. (b) CBF genes were activated by cold stress in tea plants. (c) The expression of light- or cold- response, and circadian genes up-regulated by cold stress. The expression levels (log10) of representative genes were redisplayed using a heat map. The expression levels of (d) UGT78A14, and (e) two CsUGT91Q2 genes in the TR and LE. (f) Proposed working model of the conservative light and cold signal transduction pathway in tea plants
Tea plant trichomes specifically and extensively expressed many defense-related genes
-
In-depth analyses of global gene expression in trichomes (TR) and trichome-removed leaves (LE) showed that tea plant trichomes highly express many anti-insect, anti-microbial, and defensive genes (Fig. 3a−b; Supplemental Fig. S3). More than 17 cysteine proteinase inhibitors (PINs) and other types of PIN genes were highly expressed in TR; particularly the major cysteine PIN genes, such as MSTRG.23424.1, TEA019284.1, and TEA024427.1, were expressed 2-10-fold higher in trichome than in LE (Fig. 3a−b; Supplemental Fig. S3). More than 20 genes encoding peroxidases (PODs) and 10 encoding laccases were expressed several times higher in TR than in LE (Fig. 3a−b; Supplemental Fig. S3), likely oxidizing phenolics to highly reactive quinone compounds with strong activities in defense against insects and pathogens[39,40]. The majority of glutathione-S-transferase (GST) and metallothionein-like protein (MT) genes are expressed significantly higher in trichomes than in leaves (Fig. 3a−b; Supplemental Fig. S3). GST and metallothionein proteins are both highly inducible in responses to toxins and heavy metal stress in trichomes, similar to the trichomes of Vicia faba and Arabidopsis[14,41,42]. All major tea plant chitinase genes and heat shock protein (HSP) were also expressed significantly higher in trichomes than in leaves (Fig. 3a−b; Supplemental Fig. S3), likely to enhance plant immunity, as well as plant responses to abiotic stresses such as hot, salt, cold, and wounding[14,43].
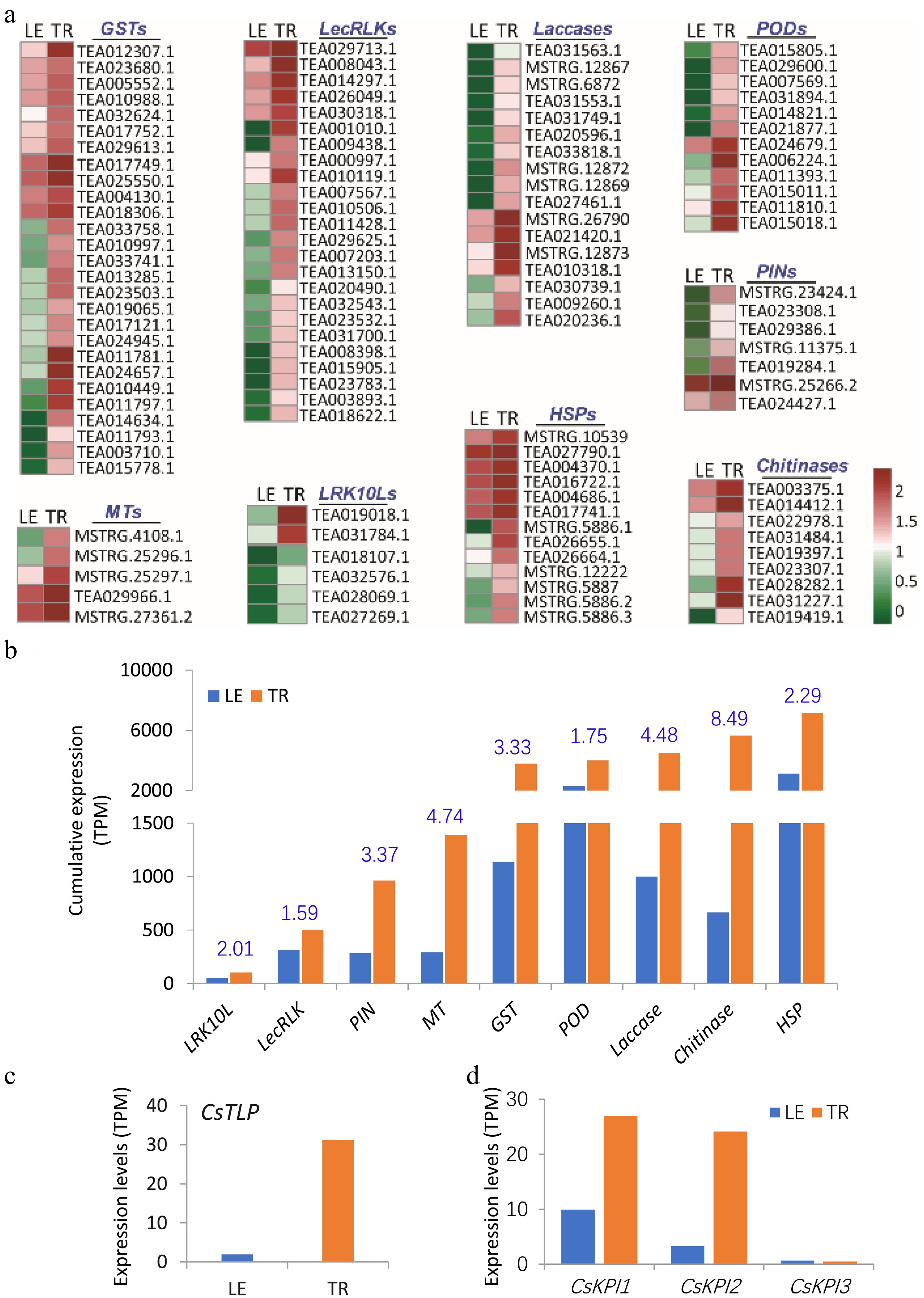
Figure 3.
Enhanced expression of defense-related genes in tea plant trichomes. (a) Several major gene families involved in anti-insect, anti-microbial, and general defense displayed higher expression levels in trichomes (TR) than in trichome-removed leaves (LE). The expression levels (log10) of representative genes were represented using a heat map. (b) Cumulative expression of some gene families in TR and LE. The fold changes (TR/LE) in gene expression are shown in blue on the bar chart. The expression level of (c) Thaumatin-like protein (TLP), and (d) three Kunitz protease inhibitor genes (KPI) in the TR and LE. GST, glutathione-S-transferase; LRK10L, leaf rust 10 disease-resistance locus receptor-like protein kinase; POD, peroxidase; HSP, heat shock protein; MT, metallothionein protein; LecRLK, G-type lectin S-receptor-like serine/threonine-protein kinase; PIN, protease inhibitor protein.
Many disease resistance proteins, such as various receptor-like protein kinases, play important roles in plant defense against various microbial pathogens or abiotic stresses[44−48]. Six genes encoding leaf rust 10 disease-resistance locus receptor-like protein kinase (LRK10L) family were more highly expressed in trichomes than in leaves (Fig. 3a−b; Supplemental Fig. S3). Trichomes also specifically expressed a group of more than 20 G-type lectin S-receptor-like serine/threonine protein kinase (LecRLK) family genes at higher levels than leaves (Fig. 3a−b; Supplemental Fig. S3). Leucine-Rich Repeat Receptor-like kinases (LRR-RLKs) are involved in signal transduction regulating plant growth, morphogenesis, and disease resistance[45,49−52].
In tea plants, some genes have been identified to be involved in defense against insects and pathogens, and cold stress. Gray blight (GB) is one of the most destructive diseases of tea plants. Thaumatin-like protein (TLP) is a crucial positive regulator in the response of tea plants to GB[53]. In our RNA-seq data, the expression of TLP was higher in the TR than the LE by more than 15 times (Fig. 3c). We also found many other TLP genes in the tea genome with higher expression levels in the TR, such as TEA014254, TEA014256 (Supplemental Fig. S4). The TLP genes highly expressed in the trichomes, play critical roles in the response of tea plants to GB stress. Kunitz protease inhibitors (KPIs) act as crucial compounds in defense responses against insect attack and pathogen infection in tea plants[54]. Three tandem-repeated KPI genes have been identified in tea plants and involved in jasmonic acid or salicylic acid mediated resistance[54]. In the TR, both CsKPI1 and CsKPI2 genes were highly expressed compared with LE (Fig. 3d). However, CsKPI3 showed low expression in both TR and LE (Fig. 3d).
Extensive expression in TF gene expression in trichomes
-
To gain further understanding of tea plant trichome initiation and development, as well as regulation of the characteristic secondary metabolism in tea plants, we further investigated the transcriptome data for various TF genes. Different types of TFs, such as MYB, WD40, bHLH, HD-ZIP, and AP2, play crucial roles in the regulation of trichome initiation and development, and the biosynthesis of secondary metabolites[8,55]. Global expression genes in trichome and leaves also include extensive types and abundance of TF genes in trichomes as compared with the trichome-removed leaves. Interestingly, most genes encoding different types of TFs were differentially expressed in trichomes and leaves (Fig. 4; Supplemental Fig. S4). Several MYB TFs, such as TEA005413, TEA012515, TEA030065, TEA002005, and TEA021737 were primarily expressed in trichomes, but almost not expressed in the trichome-removed leaves (Fig. 4). Exceptionally, TEA009412, which was annotated an AtMYB12 homologous gene, was expressed higher in TR than in LE (Fig. 1e; Fig. 4a), consistent with the higher expression levels of FLSs in TR. Our previous study[26] has shown that in the TR most lignin-related biosynthetic genes showed higher expression levels compared with LE, and here we also found some specific MYB lignin regulators highly expressed in the TR. In plants, MYB46 and MYB83 were direct targets of NAC TFs and acted as second-layer master switches in secondary cell wall formation. The homologues genes of AtMYB46/83 in tea plants, including TEA026962.1 and TEA033570.1, highly expressed in the TR compared with LE, which is consistent with the high composition of lignin in the TR (Supplemental Fig. S5a). We also found some MYB genes specifically expressed in the TR, such as TEA017098.1 and TEA007100.1, which were annotated as the trichome differentiation protein GLABROUS1 (GL1) like (Supplemental Fig. S5b). Those trichome specific MYB genes maybe play crucial roles in tea trichome formation, and further functional research is needed to explore the mechanism of tea trichome initiation and differentiation. However, the most genes encoding WD40 TFs displayed equal expression levels in both TR and LE, such as TEA030222, TEA010556, TEA030256, TEA013402, and TEA000080 (Supplemental Fig. S4). Another WD40 TFs, TEA006261 was found to be significantly highly expressed in TR than in LE (Fig. 4d). Moreover, two AP2 TFs (TEA019792, TEA026595) were also identified to be significantly highly expressing in TR, and the transcripts of several AP2 TFs, such as TEA029858, TEA016182, TEA014653, TEA017527, TEA022729, TEA032107, were almost not detected in the trichome-removed leaves (Fig. 4f; Supplemental Fig. S4). bHLH TF, TEA009147, was the most highly expressed in all bHLH family genes, showing almost no expression difference in TR and LE (Supplemental Fig. S4). Another two bHLH TFs, TEA004897 and TEA021786, were found to be significantly highly expressed in TR than in LE. Additionally, three HD-ZIP TFs, including TEA014446, TEA0051790, and TEA006200, were also more highly expressed in TR than in LE (Fig. 4i). The cumulative expression analysis of these TF family genes also indicated the abundance of those TF family genes in TR and LE (Fig. 4j).
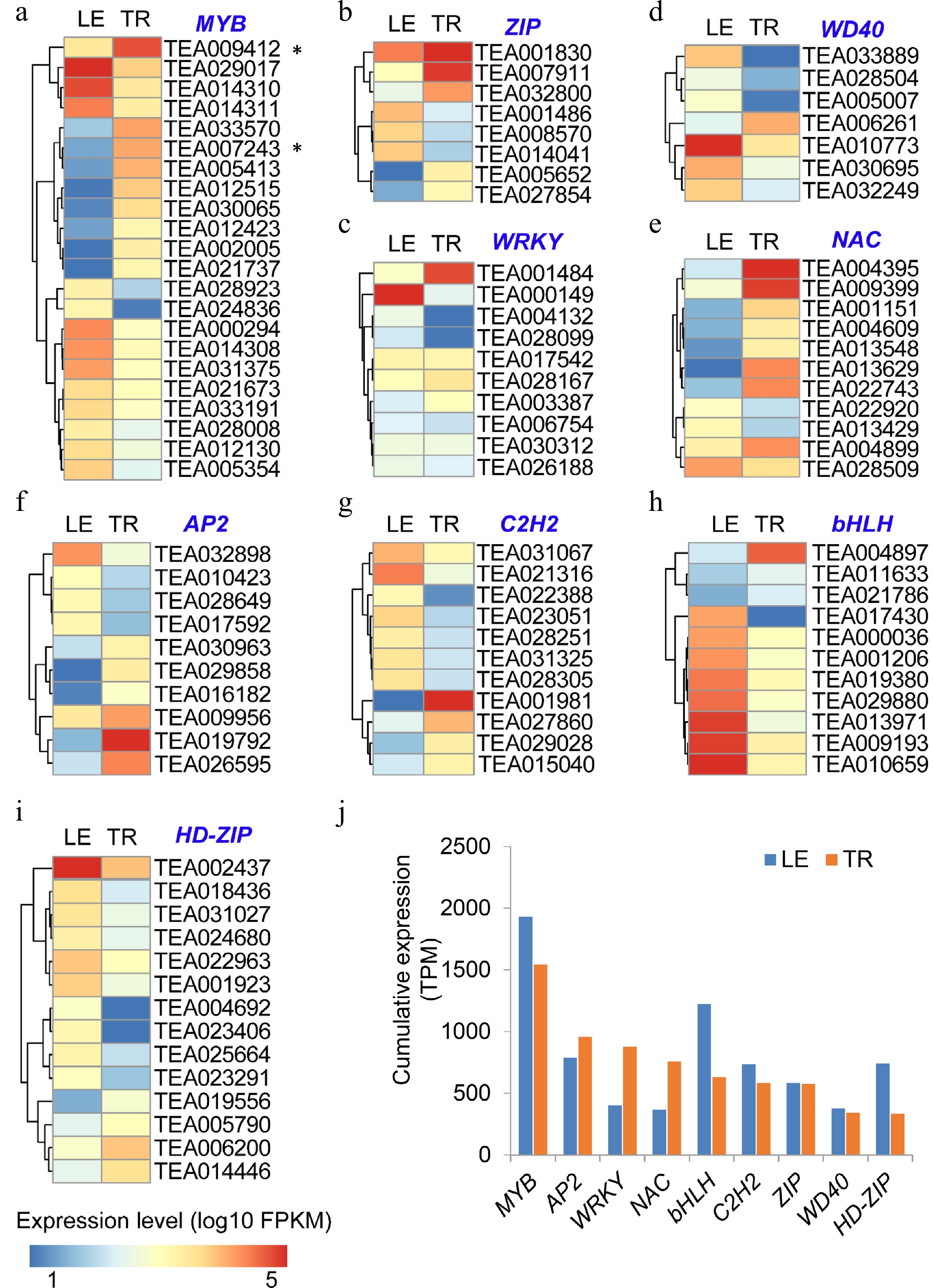
Figure 4.
Abundant expression of transcription factors in tea trichomes (TR) and trichome-removed leaves (LE). (a)-(i) The expression of representative TF genes in comparison with TR and LE, including MYB, ZIP, WRKY, WD40, NAC, AP2, C2H2, bHLH, and HD-ZIP. The relative expression levels (log10) of representative TFs are represented using a heat map. (j) Cumulative expression of each of TF families in TR and LE.
Tea trichomes contribute to insect resistance
-
We then further explored the function of trichomes on the insect resistance in tea plants. Effects of the longer and densely spaced trichomes on buds and young leaves on insect resistance against primarily Toxoptera aurantii Boyer and Empoasca pirisuga Matumura were observed in tea gardens with FDDB, DJBH, HJG, and LJ (Fig. 5). In a comparison of two years of experiments, it was seen that the presence of longer and more densely spaced trichomes on tea plant varieties made their apical buds and young leaves less damaged and better able to repel insects and larvae (Fig. 5).
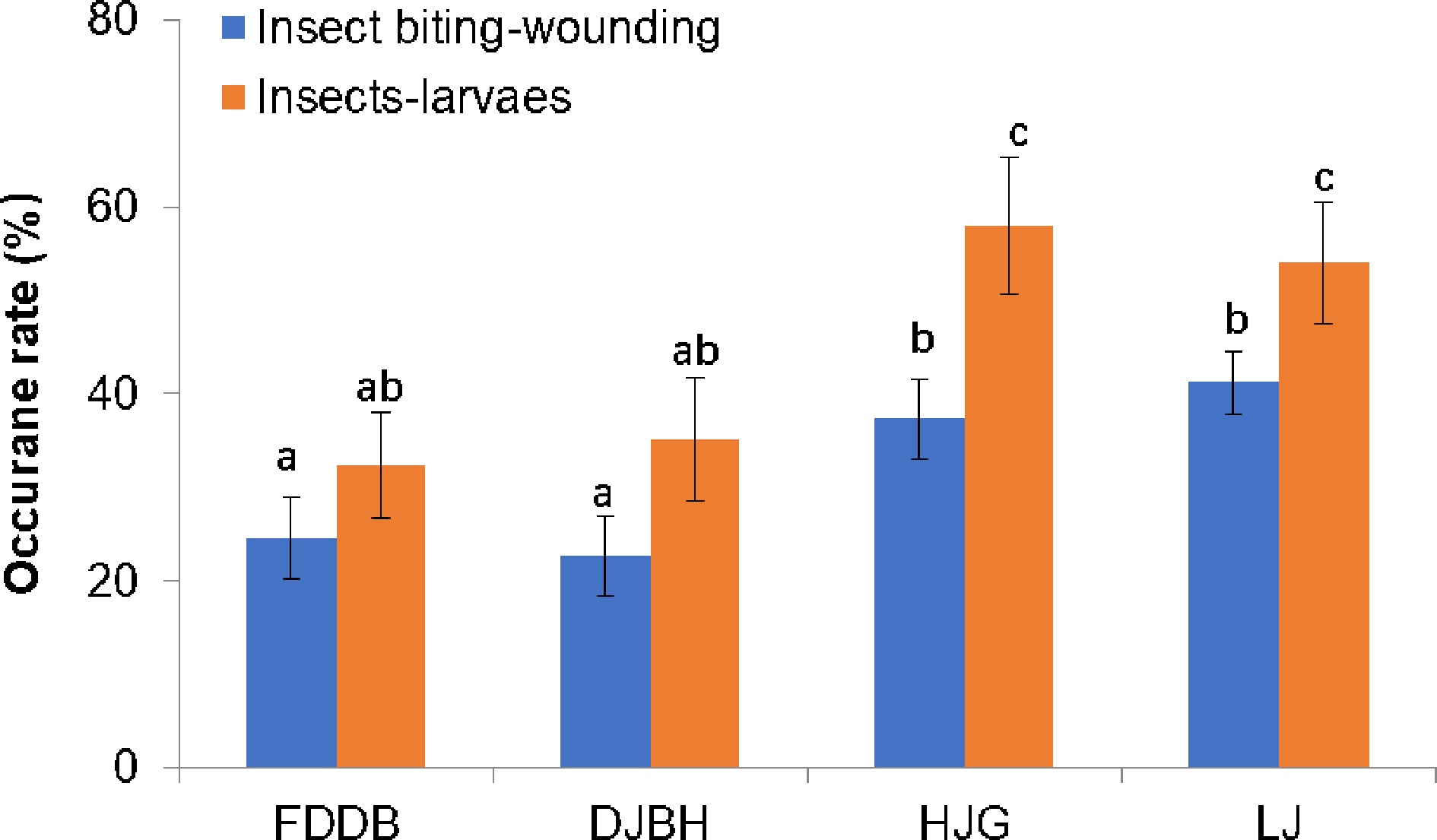
Figure 5.
Impacts of tea trichomes on resistance against insect attacks.. Effects of the presence of long and densely spaced trichomes on resistance against insects and larvae of Toxoptera aurantii Boyer and Empoasca pirisuga Matumura were observed in tea gardens. The insect resistance phenotypes of these tea plant varieties were examined during May to August, in the tea gardens in Luan and Dongzhi areas in Anhui province (Anhui, China). Two years of observation data were obtained from at least three different tea gardens. More than 30 plants were examined in each experiment. Data are expressed as means ± s.d. ANOVA were used to examine the significance of differences.
Effects of tea trichome density on tea plant tolerance to UV-B and high temperature stresses
-
To further understand how the presence of trichomes on the apical buds and young leaves could protect them from abiotic stresses, we treated two tea plant varieties, Longjinchaye (LJCY) and Baihaozao (BHZ), which had much lower trichome index and higher trichome indexes, respectively, with UV-B radiation and high temperature. Although after treatment for a certain period of time (more than 48 h), we did not observe any significant morphological difference in shoot tips of these two tea plants, we did detect differently induced genes that usually reflects the physiological status of plant tissues. Under regular white light radiation, both CsCOP1 and CsbZIP1 was up-regulated and expression levels were generally higher in LJCY than in BHZ. Compared with UV-B radiation, their expression pattern slightly changed. CsCOP1 expression became higher in BHZ than in LJCY after 24 h of UV-B treatment, and CsbZIP1 transcript became higher in BHZ than in LJCY after 48 h of UV-B treatment (Fig. 6). Correspondingly, the CsbZIP1 downstream target genes, such as CsMYB12 and CsELIP2, were also expressed at higher levels in LJCY than in BHZ, suggesting that BHZ shoot tips covered with densely spaced trichomes attenuated the UV-B-triggered light signaling and downstream response due to trichome reflection of light (Fig. 6). Both up-regulation of both CsMYB12 and CsELIP2 by bZIP1 homolog protein HY5 in Arabidopsis and tea plants could trigger flavonoid accumulation and defense responses against high-light and UV-B damage (Fig. 6). During high temperature stress, heat shock proteins are often rapidly accumulated to protect plant cells by a variety of means, such as helping protein folding correctly and other biomolecules function properly. The heat stress-induced CsHSP90-1 expression responded more rapidly and significantly in BHZ than in LJCY, but CsHSP90-1 transcripts showed similar patterns. Later both showed higher expression levels in LJCY than in BHZ at 24 h or 48 h after high temperature treatment (Fig. 6). The thermomorphogenesis gene PIF4 homolog gene CsPIF3 were also up-regulated by heat stress, and showed differential induction patterns in BHZ and LJCY shoot tips (Fig. 6). Recently it was found that the Evening Complex, composed of LUX ARRHYTHMO and EARLY FLOWERING 3 (ELF3) and ELF4, is a core component of the plant circadian clock and plays key roles in plant growth under circadian and thermomorphogenesis[56]. ELF4 can act as a transcriptional repressor in a function of exterior temperatures[57]. We also observed that CsELF4-1 and CsELF4-2 were substantially up-regulated by high temperature treatment, and showed differential expression patterns in BHZ and LJCY shoot tips (Fig. 6). BR signaling is also involved in heat stress response in tea plants[58], and BR signaling suppressor BRI1-EMS-SUPPRESSOR 1 (BES1) is an essential signaling component in BR signaling and regulation of various cellular and physiological processes[59]. CsBES1 was also up-regulated by heat stress, with higher levels in BHZ than in LJCY shoot tips after 24 h of treatment (Fig. 6). BRs increase the heat stress resistance of plants and BES1 can interact with heat shock factor HSFA1 to enhance the synthesis of heat shock proteins (HSPs) to induce growth-promoting genes and to accumulate protective natural products[59]. The higher expression levels of CsBES1 and CsPIF3 in BHZ all implied a stronger resistance against heat stress.
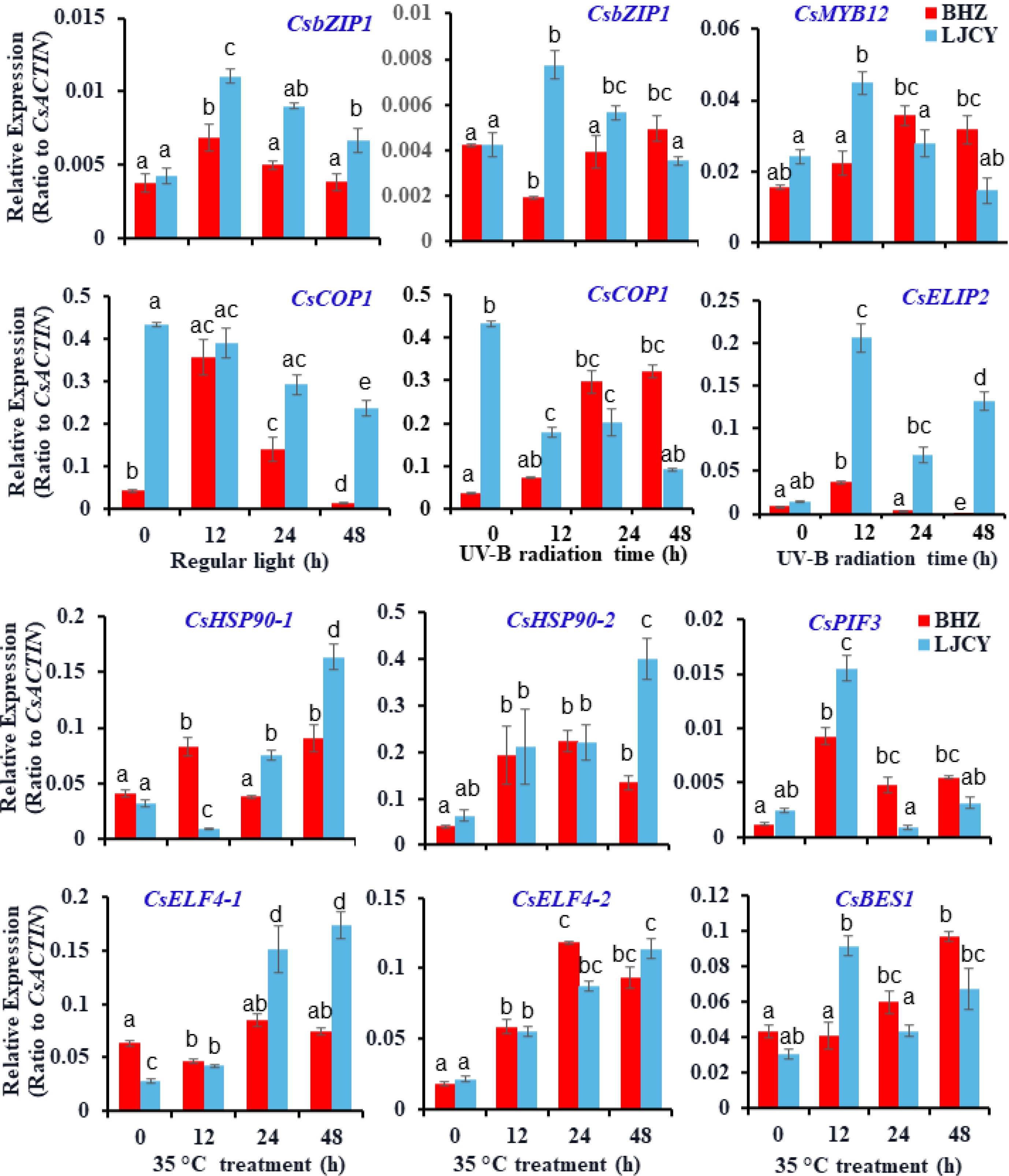
Figure 6.
Effects of trichome density on tea plant resistance against UV-B radiation and high temperature stress. Two tea plant varieties, with distinct trichome indexes were chosen for examination of the protection roles of trichomes on tea plant shoot tips. Two tea plant seedlings were treated with UV-B radiation or high temperature (35 °C) for a variety of times (regular white light in growth chambers or normal temperature (22 °C) as controls). Shoot tips were monitored for any phenotypes or sampled for RNA analysis with qRT-PCR examination of stress-related genes. More than nine plants for each genotype of tea plant varieties were treated in each experiment. Data are expressed as means ±s.d. ANOVA were used to examine the significance of differences.
-
Tea plants grow in both high and low altitudes, the young leaves often face high light radiation or UV-B damage. High light and excessive UV-B could lead to serious damage to tea plants tender tissues, and repress plant growth. Plants have evolved several photoprotection strategies to adapt to the surrounding environment, such as forming a trichome layer on the young tissue exterior for light energy shielding[16,17]. Although the tea trichomes belong to unicellular non-branching, and non-glandular type of trichomes, these soft and spidery surface hairs fully cover the abaxial side of apical buds before leaf opening, providing the first protective physical barriers for reflecting high light and UV-B radiation. Tea trichomes have structural features with strong reflection of high light and UV-B, and could protect the mesophyll cells from most light damage (Fig. 1b). From transcriptome data, we found that many trichome birefringence genes that play crucial roles in trichome birefringence, mostly displayed higher expression levels in trichomes compared to trichome-removed leaves (Fig. 1d), supporting a role of trichomes in protection from high-light damage. In addition, our previous study has demonstrated that the CsUVR8-CsbZIP1 work model is also crucial in tea plant response to high light or UV-B stress[26]. As a central signaling factor in the light-mediated developmental and metabolism processes, the HY5 homolog gene CsbZIP1 was also highly expressed in tea trichomes. All three transcripts of CsbZIP1 showed higher expression levels in trichomes than in the trichome-removed leaves, so were the CsbZIP1 upstream regulator UVR8s (Fig. 1e). Interestingly, the expression of CsCOP1 and CsPIF3, two CsbZIP1 repressors, showed no significant variation in the TR or LE (Fig. 1e), supporting that the tea trichomes are bound to express more positive and earlier light responsive genes than the mesophyll cells. HY5 also responded to high light or UVB stress through activating some early light-induced proteins, such as ELIP2. The expression level of CsELIP2 was up-regulated more than 30 times in the TR compared with the LE (Fig. 1f). ELIP2 is involved in the response to multiple environmental stresses related to photoinhibition, such as UV-B, high light, photooxidative, and cold stress[36], and in tea plants, CsELIP2 and CsbZIP1, with high expression levels in trichomes, seem to build an early warning mechanism of multiple environmental stresses to reduce the damaging effects of mesophyll cells. Our previous study[26] has demonstrated that in tea trichomes, many flavonol compounds were synthesized and accumulated, as well as the flavonol biosynthetic genes with higher expression levels. The flavonol compounds play a crucial role in responding to multiple stresses, particularly UV-B. We have identified the central roles of the CsbZIP1-MYB12 work model in regulating flavonol biosynthesis[26]. Gene expression analysis showed that the CsMYB12, as well as its regulator CsbZIP1, was highly expressed in tea trichomes (Fig. 1e). All these together support that, as the first physical barrier, the trichomes protected the apical buds and young leaves from the high-light and UV-B radiation stress with more comprehensive strategies, through both morphological features, multiple highly expressed light-responsive genes, and synthesis of many protective metabolites, including flavonols and catechins.
Defensive peptides and disease resistance proteins in tea trichomes against insects and microbial pathogens
-
Tea plants are subject to attacks by a wide range of microbial pathogens and herbivores, which cause more than 380 diseases and more tea production loss[60,61]. Our previous study suggested that the tea plant trichomes built up chemical defenses, which can discourage insect predators and protect the leaves from microbial attack[2]. Compared with LE, trichomes expressed relatively equal levels of many biosynthetic genes. Thus, trichomes have built both physical barriers and chemical defenses on tea plant leaves against microbial infection or insect herbivore attacks.
Moreover, tea plant trichomes specifically synthesize many defensive peptides, such as GLPs, PINs, proline-rich peptides, MTs, HSPs, and GST, and reactive enzymes such as laccases and PODs for defense against abiotic and biotic stresses[14]. Tea plant trichomes were also enriched in expression of various receptor-like kinases and chitinase as defense signaling proteins, such as LRK10Ls and LecRLKs. Synthesis of PINs is a well-known defense response for plants against insect herbivore attack. PINs and anti-insect caffeine are induced by herbivore insect attacks, mediated by jasmonate signaling (Fig. 3). The constitutively expressed PIN genes and accumulation of defensive catechins and caffeine in tea plant trichomes strongly suggested a role of trichomes against insects in tea plants. The latest research showed that the combination of more trichomes and exogenously applied chitinases significantly reduced growth of herbivore larvae[62]. Cumulative expression analysis of related gene family members further supported the functions of tea trichomes in the defense against abiotic and biotic stresses. In tea plants or other plants, it is not common that many whole gene families were highly expressed to participate in defense stresses in trichomes.
In tea plants, some genes have been well characterized in defense against insects and pathogens, such as Thaumatin-like protein (TLP) against gray blight, Kunitz protease inhibitor (KPI) against insect attack and pathogen infection[53,54]. Interestingly, those inducible type genes kept low expression levels in trichome-removed leaves, however high expression in trichomes (Fig. 3). The high expression levels of those genes in trichomes clearly demonstrated that the trichomes played crucial roles in protection for mesophyll cells against biotic stresses.
Tea trichomes synthesize specialized metabolites for innate immunity
-
Like unicellular trichomes of Arabidopsis and cotton[12], tea plant trichomes also contained the major specialized metabolites at high levels, such as tea plant-specific secondary metabolites, such as catechins, theanine, caffeine, and volatiles, with well-known functions in anti- microbial pathogens and anti-insects[2]. Our previous study has clearly shown that tea trichomes contained large amounts of volatile compounds, such as decanal, nonanal, and geranylacetone, which played important roles in tea flavor formation and plant defense[2]. It was reported that the volatiles involved in the response to tea plant cold stress are nerolidol and nerolidol glucoside[38]. Tea UGT91Q2 could elevate the nerolidol glucoside formation, promoting the enhancing tea plant cold stress tolerance[38]. Notably, two UGT91Q2 genes, particularly UGT91Q2-2, highly expressed in the trichomes compared with the trichome-removed leaves (Fig. 2e).
In tea plants, CsUGT78A14, which catalyzed flavonol to form flavonol glucoside, is also involved in cold tolerance for tea plants[37]. In tea trichomes, many flavonol compounds were detected, and play crucial roles not only in the high light and UVB stress, but also in cold tolerance. In tea plants, flavonol compound biosynthesis were specially regulated by MYB12 TF[26]. CsUGT78A14 highly expressed in tea trichomes, which is consist with the CsMYB12 and CsbZIP1 expression patterns. HY5 has been reported to positively regulate cold tolerance via the HY5-MYB15-CBF network in tomato[63]. Interestingly, in tea plants, the expression levels of CsbZIP1 and CsMYB12 synergistically were significantly activated by cold stress. Thus, CsUGT78A14 is also likely involved in cold tolerance via the CsbZIP1-CsMYB12 work model. Certainly, HY5 could also directly regulate the C-repeat/DREB binding factor (CBF) pathway to involve cold tolerance, which was supported by the presence of many CsbZIP1 binding motifs, such as G-box and C-box, in the promoter region of tea CBF genes (Supplemental Fig. S6). Conservatively in tea plants, the C-repeat/dehydration responsive element binding factor (CBF/DREB), inducer of CBF expression (ICE) and cold-responsive (COR) genes form a ICE-CBF-COR signaling pathway to activate plant defense responses to survive cold stress [62,63]. ICE1 plays a central role in mediating cold-induction of CBF3, ICE1 is also involved in regulation of stomata development, which were promoted in Arabidopsis in cold conditions [64].
In tea plants, more experiments are needed to explore the multiple functions of HY5 regulating cold tolerance, which can be exploited in the future to enhance cold tolerance in tea plants. Based on our data, the HY5-MYB12 work model could integrate the signal of high light, UV-B stress and cold stress to protect tea plants through regulating multiple downstream pathways[26]. Summarily, tea trichomes could synthesize many secondary metabolites and defensive peptides, and express many signal transduction genes to be involved in the biological defense or cold tolerance for tea leaves.
Tea trichomes expressed a wide array of TFs regulating defense and development
-
The phenylpropanoid and flavonoid pathways are tightly regulated at the transcriptional level by MYB, bHLH, WD40-repeat, WRKY, NAC, HD-ZIP, and other TFs. The MBW complexes have been demonstrated as one of the most important regulatory mechanisms that are involved in both regulating phenylpropanoid and flavonoid biosynthesis and plant epidermal cell differentiation, including trichome/root hair formation[64]. The tea plant MBW complex, formed by CsAN1-CsGL3 (CsEGL3)-CsWD40 for regulating anthocyanin production, and by CsAN2 (CsMYB5e)-CsGL3(CsTT8)-CsWD40 possibly for regulating anthocyanin and proanthocyanidin biosynthesis have been reported[24,65,66]. Several tea plant MYB factors, such as CsMYB5a, CsMYB5b, CsMYB5e, CsMYB5-1, and CsMYB5-2, were reported with functions such as regulating anthocyanin and proanthocyanidin accumulation when expressed in other plants[66,67]. TF genes, such as CsNST1 (TEA013629) and a MYB TF gene TEA007243 homology to Arabidopsis AtMYB86 gene involved in cell wall biosynthesis, were expressed higher in trichomes [68,69]. Our RNA-Seq data showed that many MYB, bHLH, WD40, NAC, and WRKY TF genes were expressed at significantly higher levels in trichomes than in leaves with trichomes being removed. These TFs and their complexes could regulate flavonoid synthesis and trichome development. Several key genes homology to TTG1, GL3, GL1, TRY, and TRANSPARENT TESTA GLABRA2 (TTG2) that are involved in trichome initiation and development in Arabidopsis, also showed higher expression levels in tea plant trichomes, indicating that they may similarly be involved in regulating trichome differentiation and growth. On the other hand, several genes that are involved in the negative regulation of trichome development were expressed much lower in trichomes than in leaves, such as TEA023420, an MYB gene homology to Arabidopsis negative trichome development regulator AtMYB82[70,71]. In this study, we found that genes either specifically expressed in trichomes or specifically repressed in trichomes compared to leaves, such as MYB, NAC, and WRKY genes, suggesting complex regulatory networks in differentially regulating multiple metabolic pathways, or developmental, or cell growth processes[72,73].
Tea cultivars with longer and densely spaced trichomes on the apical buds and young leaves usually showed higher insect resistance compared with the cultivars with shorter and thin trichomes. Based on our data, the trichome phenotype seems a domestication trait of tea plants, and most wild Thea section plants have either glabrous leaves or with only short and rare trichomes. The trichome indexes are positively related to the tea quality and tea plant tolerance against adversaries, which is even more pronouncedly observed between the wild tea and current tea cultivars.
-
Tea trichomes not only contribute special flavors and health benefits to tea by synthesizing and accumulating a wide range of secondary metabolites, but also highly express many disease resistance-related and abiotic stresses-related genes. Both toxic chemicals, physical barriers, disease resistant gene expression, and defensive peptide synthesis, enabled tea trichomes to build up defense strategies to protect the apical buds and young leaves from complex biotic and abiotic stresses. Thus, tea plant trichomes confer effective physical and biochemical defenses. Several pieces of experimental evidence were shown to provide new insights into the multiple biological functions of tea trichomes in tea plant adaptation to various environmental cues. The study lays a foundation for the future investigation of the defense mechanisms of trichomes in tea plant resistance against adverse environments.
The work is supported by the National Natural Science Foundation of China (32002089); the Anhui Provincial Natural Science Foundation (201902a05020408); the National Key Research and Development Program of China (2018YFD1000601) and the funding from Anhui Agricultural University and the State Key Laboratory of Tea Plant Biology and Utilization. We thank Dr. Fangdong Li for assistance on the bioinformatics analysis. The authors also thank members in Zhao’s lab for all of the assistance with the experiments and data analyses.
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Trichome appearances on leaves of different tea plant varieties. MSBH, Mingshanbaihao, FDDB, Fudingdabai, MX213, Mingxuan213, SCZ, Shuchazao, LJCY, Longjingchangye, JHC, Juhuachun.
- Supplemental Fig. S2 The expression pattern of trichome birefringence (TBR ) gene family (a) and flavonoid biosynthesis related genes (b) in TR and LE. The expression levels (log10) of representative genes were represented by heat map. The TEA006040.1 (TLP) involved in defense against gray blight is reported by Wang et al., 2021.
- Supplemental Fig. S3 Comparative expression of enriched defense-relate genes in trichome (TR) compared with trichome-removed leaf (LE). The genes which were significantly up-regulated in TR, were remarkable using black line at ID right. Other genes with no marker mean significantly down-regulation in TR.
- Supplemental Fig. S4 The abundant expression of transcription factors in tea trichomes (TR) and trichome-removed leaves (LE). The expression levels (log10) of representative MYB, WD40, bHLH, HD-ZIP, and AP2 type transcription factors were represented by heat map.
- Supplemental Fig. S5 Some MYB transcription factors highly expressed in tea trichomes. (a) The expression levels of two homologous genes of lignin regulators AtMYB46/MYB83 in TR and LE. (b) The expression levels of two homologous genes of trichome differentiation regulator GL1-like in TR and LE.
- Supplemental Fig. S6 Light-related cis-elements in the promoters of tea core binding factor (CBF) genes.
- Supplemental Table S1 Primers used in this study.
- Copyright: 2022 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li P, Zhang Y, Ye Z, Zuo H, Li P, et al. 2022. Roles of trichomes in tea plant resistance against multiple abiotic and biotic stresses. Beverage Plant Research 2:14 doi: 10.48130/BPR-2022-0014 

Roles of trichomes in tea plant resistance against multiple abiotic and biotic stresses
- Received: 30 June 2022
- Accepted: 04 August 2022
- Published online: 23 August 2022
Abstract: Tea trichomes contribute significantly to tea flavors by providing diverse and specific flavor-determining metabolites, including catechins, caffeine, theanine, and volatiles. However, not much is known about the physiological functions of tea trichomes in tea plant adaptation to complex environments, nor the trichome development. Tea trichomes not only build up chemical defenses, but also act as the first physical barrier protecting herbivore attacks, reflecting high light and UV-B radiation, and preventing water loss. Moreover, transcriptome profiling on the tea trichomes compared with the trichome-removed leaves also showed that tea trichomes highly expressed numerous defense-related genes involved in protection from high light and UV-B radiation, cold stress, disease resistance signal transduction, anti-herbivore or anti-abiotic peptide biosynthesis, and other defense responses. Several pieces of experimental evidence supported the notes, highlighting the roles of tea trichomes in plant defenses against both abiotic and biotic stresses. The study provides fresh insights into the multiple protective functions of tea trichome for tea plant adaptation to harsh environments. The new understanding on tea trichomes could benefit the development of better breading strategy for new tea varieties with greater adaption and tolerance to changing environmental challenges.
-
Key words:
- Trichome /
- Photoprotection /
- Tea Quality /
- Insect attack /
- High temperature.