









-
Pepper (Capsicum annuum) is a Solanaceae plant that is widely cultivated worldwide, which is commonly used as a food source due to its unique flavor and rich nutritional value. Although the seedlings of Solanaceae plants such as tomato (Solanum lycopersicum), cherry tomato (Lycopersicon esculentum), and Chinese lantern (Physalis alkekengi) can grow upright at the start, the increase in leaves and emergence of flowers and fruits cause the plants to bend as their stems exhibit low degrees of lignification and mechanical strength. Thus, scaffolds must be set up and branches lifted to promote their growth, which cost substantial human and economic resources. However, most pepper varieties can grow upright during production due to their highly lignified stems. Therefore, the mechanical strength of the pepper stem could be studied to provide guidance for the production of more vegetable crops.
Lignin is one of the main chemical components of plant stem. The accumulation of lignin in the cell wall will affect the mechanical strength of the plant stem[1]. As one of the vital secondary metabolites produced by the phenylpropane metabolic pathway, lignin is deposited in the secondary cell wall of all vascular plants. There are several types of monomers which could make up lignin, the three main types being mustard alcohol, coumarin alcohol, and coniferol, to produce p-hydroxyphenyl (H) lignin, guaiacyl (G) lignin, and syringyl (S) lignin, respectively[2]. Lignin mainly contains g-units and a small amount of H-lignin in gymnosperms, whereas G-lignin and S-lignin are the main parts in angiosperms[3]. Lignin monomers are produced in the cytoplasm and transported to the ectoplast, before being moved to the cell wall through an ATP-binding cassette transporter to complete polymerization[4]. The cell wall rigidity depends on the proportion of these three monophenol units, and the structure of lignin in different plants is not necessarily the same. In recent years, some genes related to lignin biosynthesis have been identified. The total amount of lignin synthesis is closely related to the production and activity level of phenylalanine ammonia lyase (PAL)[5], cinnamate-4-hydroxylase (C4H)[6], 4-coumarate: CoA ligase (4CL)[7], ferulate 5-hydroxylase (F5H)[8], caffeic acid/5-hydroxyferulate O-methyltransferase (COMT)[9,10], caffeoyl CoA/5-hydroxyferulate CoA-o-methyltransferase (CCoA-OMT)[10], cinnamyl alcohol dehydrogenase (CAD)[11], p-coumaric acid 3-hydroxylase (C3H)[12]/hydroxycinnamoyl transferase (HCT)[13], and cinnamoyl CoA reductase (CCR)[14]. All the genes listed are crucial for the synthesis of monomers and lignin biosynthesis. The fusion of lignin with cellulose and hemicellulose improves the mechanical strength and lodging resistance of plant cells. Hu et al.[15] reported that lignin content and lignin biosynthetic enzyme activity were crucial in lodging resistance in common buckwheat (Fagopyrum esculentum). Studies on cut flowers such as herbaceous peony exhibited that the development of stem increased the degree of lignification, distribution of lignified cell wall, and stem mechanical strength of varieties with a relatively high degree of lignification[16]. Additionally, the fusion of substances such as lignin and cellulose in the cell wall skeleton creates an effective barrier to prevent the invasion of various pests and pathogens, and improve the cellular defense ability of plants[17].
Arabinogalactan proteins (AGPs) are a high molecular weight glycoprotein commonly existing in the plant kingdom, which are composed of a protein core skeleton and glycosyl side chain[18,19]. AGP responds to abiotic stress factors[20] and participates in the regulation of various hormones such as cytokinin[21], abscisic acid[22], and gibberellin[23]. The protein skeleton mainly contains substantial hydroxyproline/proline, alanine, serine, and threonine, while the different additional domain could divided AGPs into classical AGPs, lysine rich AGPs, Ag peptides, hybrid AGPs, and chimeric AGPs[24]. Among them, Xylogen-like arabinogalactan protein (XYLP) is a kind of chimeric AGP, comprising an additional lipid transfer protein (nsLTP) domain[25]. Previous studies exhibited the crucial role of XYLPs during the development of vascular system in Arabidopsis[25], rice (Oryza sativa L.)[26], wine grape (Vitis vinifera L.)[27], poplar (Pinus taeda L.)[28]and zinnia (Zinnia elegans L.)[29]. Especially the notable function of XYP1 and XYP2 in Arabidopsis, with the xyp1xyp2 double mutant displaying discontinuous veins in roots, cotyledons and leaves caused by the obvious morphological defects in vascular development[25]. Osxylp7 mutants exhibit shorter stems and panicle stem lengths in rice, and OsXYLP7 could be up-regulated not only under abiotic stresses such as drought and salt stress, but also under NAA, IAA and 6-BA treatments[26]. In grape, VvXYLP02 was found to be induced by JA upon the pathogen infection, and participate in plant resistance against gray mold disease[30]. PtXYLP1 and PtXYLP2 are transcriptionally regulated by NAA, 6-BA and GA during Arabidopsis cotyledon development[28]. These results indicate that XYLPs could play important roles in plant growth and under stress. Therefore, analysis of XYLP could provide theoretical significance for understanding the mechanism of plant vascular system.
-
Pepper cultivars 'ST-8' and 'HP' were used in this study, with all of the pepper plants being cultivated at 80−100 μmol photons m−2·s−1, a relative humidity of 70%, and under a 16/8 h day/night photoperiod in the greenhouse. The pepper cultivar 'ST-8' were grown in the greenhouse at 25 °C. 'ST-8' plants were observed from 2 weeks after germination (W) every two weeks until 12 weeks. Stems at each stage at the position between the pair of cotyledons and first true leaves were sampled for observation or left in liquid nitrogen and stored at −80 °C until use. Three independent biological replicates were performed each time. The pepper cultivar 'HP' was used in VIGS (virus-induced gene silencing) to observe the phenotype of TRV2:CanXYLP1 and TRV2:CanXYLP2[31]. 'HP' was grown in the greenhouse at 25 °C before 2 weeks after germination, and then transferred to 17 ± 1 °C after 2 weeks for VIGS. Regarding the stress treatment, 2 weeks after VIGS treatment, when the parallel TRV2:PDS control started bleaching, plants were put under 10 °C for cold stress, or left with watering reduced by half for drought stress for 1 month. The photographs of whole plants were taken using a digital camera (Nikon, D7500), and the stem images were taken under the microscope (Leica, M205FA).
Lignin staining and extraction
-
To stain the lignified area in the stem cross section, we placed cut pepper stems on glass slides, and added 50−100 μl of hydrochloric acid (Baiaolaibo, Beijing, GL2679). After the stem was saturated, we added the same amount of phloroglucinol (Baiaolaibo, Beijing, China GL2679). The lignified cell wall was then stained. After taking the images under a microscope (Leica, M205FA), we used Image J software (
https://imagej.nih.gov/ij/ ) to quantify the stained area. The data are presented as the means ± SD; 20−30 independent biological replicates.To extract and test the lignin content, we followed the method established by Cuzens & Miller[32]. Firstly, we took an appropriate amount of pepper stems, dried them at 40 °C overnight and ground them into a powder. We then extracted ground powders using a benzene-ethanol solution and hydrolyzed them with 67.0% H2SO4. Klason lignin was separated as insoluble matter. We then placed the residues after filtration and ashed them in the Muffle furnace (Daoxu, Zhejiang, China SX2-2.5-10) at 575 ± 25 °C for 4 h. Finally, we calculated the Klason lignin content by comparing the weight before and after. The data are presented as the means ± SD; of three independent biological replicates.
Identification of XYLPs and bioinformatics analysis
-
Following the method established by Ma & Zhao[33], CanXYLP, SlyXYLP and StuXYLP proteins were identifed respectively (see Supplemental Method for details). An unrooted phylogenetic tree was constructed using MEGA7[34] with the neighbor-joining statistical method and bootstrap analysis (1,000 replicates). The software MCScan X[35] was used to identify the XYLP orthologous genes between pepper and tomato. The possible regulatory elements of the CanXYLP1 and 2 gene promoter sequence were analyzed through the promoter regulatory element prediction website PlantCARE (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )[36].Pepper VIGS
-
The pepper VIGS approach was carried out as per the method of Zhou et al.[31]. The fragments of CanXYLP1 and CanXYLP2 with a length of about 250 bp were inserted into the TRV2-C2b vector at the SmaI site to construct two vectors, TRV2:CanXYLP1 and TRV2:CanXYLP2, as well as the CanPDS gene was inserted into TRV2 empty vecort to construct a positive control TRV2:CanPDS. TRV1 and TRV2:CanXYLP1/TRV2:CanXYLP2/TRV2:CanXYLP1 + TRV2:CanXYLP2/TRV2:CanPDS (resuspended in induction medium at a 1:1 ratio, OD600 = 0.004), were co-infiltrated into the cotyledon of 'HP' pepper at 2 weeks, and then culti
vated at 17 ± 1 °C under 16/8 h day/night photoperiod. At 10 weeks, phenotypic observation and sampling of plants were carried out for RNA isolation, RNA-seq and quantitative real-time PCR. Three independent biological replicates for RNA isolation and quantitative real-time PCR were taken, two of which were used for RNA-seq. RNA isolation, cDNA synthesis and quantitative real-time PCR
-
Total RNA was isolated using TransZol (TransGen Biotech, Beijing, China). Approximately 1 μg of total RNA was used for cDNA synthesis through reverse transcription enzyme (Applied Biological Materials Inc., Richmond, Canada). The relative gene expression was calculated using the 2−ΔΔCᴛ method. A pepper Beta-tubulin gene (CA04g21850) was used as the internal controls. The primers used for qRT-PCR are listed in Supplemental Table S1. In each case, three technical replications were performed for each of the three independent biological replicates.
RNA-seq data analysis
-
Comparing RNA-seq reads with Hisat2 (
http://daehwankimlab.github.io/hisat2/ ) to convert the sam file, and then using Samtools (http://samtools.sourceforge.net/) to bam and sort it, next using FeatureCounts (http://subread.sourceforge.net/ ) to get the count value and finally calculate the FPKM (Fragments per Kilobase Million) value. Differentially expressed gene analysis was conducted using the R package limma (https://bioconductor.org/packages/release/bioc/html/limma.html ) and edgeR (https://bioconductor.org/packages/release/bioc/html/edgeR.html ). Genes that exhibited a difference of at least twofold with a corrected P value ≤ 0.05 were regarded as significantly differentially expressed. Online enrichment analysis of differential genes used KEGG (www.kegg.jp ). Venn diagrams were drawn by DeepVenn (www.deepvenn.com ) online. Volcano plots and heat maps were drawn by Tbtools[37].Data analysis and software
-
All the statistical analyses in this study were performed using IBM SPSS Statistics for Windows, version 23.0 (IBM Corp., Armonk, NY, USA). Variance and significant difference tests were carried out to identify differences among means by one-way analysis of variance (ANOVA) with Duncan's multiple range test. The histograms were generated via GraphPad Prism 8.0.2 (GraphPad Software, San Diego, CA, USA), Excel 2019 (Microsoft, Redmond, WA, USA) and OriginPro 2021 (OriginLab, Northampton, MA, USA) software to draw charts.
-
The present study systemically investigates pepper ST-8's stems at different growth stages from 2 weeks after germination (W) to 12 weeks to determine the lignin content during pepper stem development (Fig. 1a). The transverse growth of the stem is obvious during pepper plant development (Fig. 1ai & ii), whereas the xylem area and lignin content increases gradually (Fig. 1aiii & iv, arrows) (Fig. 1b). The lignin content in pepper stems was rarely detected at 2 weeks, whereas it reached the peak at 10 weeks. The present study observed the lignin area in stem by specific staining using phloroglucinol hydrochloric acid regent to further study the soft stem at 4 weeks and hard stem at 10 weeks. The cross-section of the 4 week pepper stem exhibited four regular and small red spots (Fig. 1c), whereas the cross-section of the 10 week pepper stem exhibited a dark red and circle area filled with lignin (Fig. 1d). These staining areas were also quantified and exhibited a significant difference (Fig. 1e). Therefore, we concluded that the lignin content in the pepper stem increased gradually during growth to provide the pepper stem the essential mechanical strength.
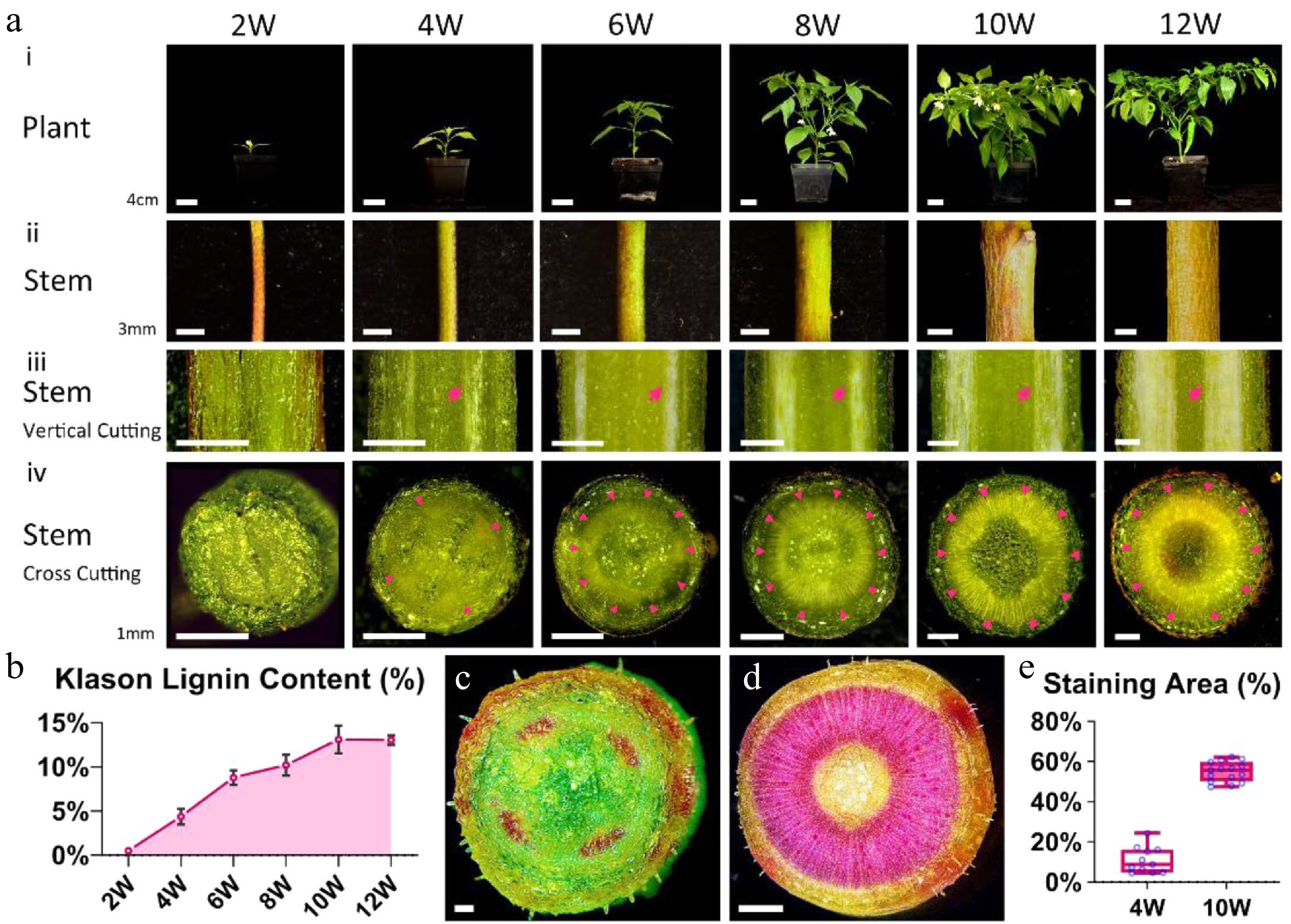
Figure 1.
The contents of lignin increase during pepper stem development. (a) Pepper plant and stem from 2 weeks after germination 2W to 12W: (i) the whole plant morphology of pepper. Bars = 4 cm. (ii) Magnified images of pepper stems. Bars = 3 mm. (iii) Vertical Cutting of pepper stems under stereomicroscope. Red arrows indicate the position of the xylem. Bars = 1 mm. (iv) Cross Cutting of pepper stems under stereomicroscope. Red arrowheads indicate the position of the xylem. Bars = 1 mm. (b) Klason lignin contents in pepper stems from 2W to 12W. (c), (d) Cross section of pepper stem stained with phloroglucinol hydrochloric acid staining solution at 4W (c, bar = 0.2 mm) and 10W (d, bar = 1 mm) under stereomicroscope. (e) The ratio of the area stained by the phloroglucinol hydrochloric acid staining solution to the entire cross-section at 4W and 10W.
Identification and characterization of XYLP family members in pepper
-
BLASTP searches were performed across pepper protein databases (zunla2.0)[38] using ZeXYP1, AtXYP1, and AtXYP2 protein sequences as queries to identify XYLPs in pepper[25]. After confirming the presence of nsLTP-like domains, AGP-like regions, and AG-type glycomodules, and removing redundant sequences, 10 CanXYLPs were identified in pepper (Supplemental Table S2 and S3). Compared with ZeXYP, AtXYP1, and AtXYP2 protein sequences, all 10 CanXYLPs were predicted to possess an N-terminal signal peptide for targeting the endoplasmic reticulum and a C-terminal GPI anchor protein (Fig. 2a and Supplemental Table S2). Additionally, putative AG glycomodules in all CanXYLPs were distributed in the PAST-rich region before and after the nsLTP-like domain. N-glycosylation sites in most of the CanXYLPs were located in the nsLTP-like domain and the PAST-rich region (Fig. 2a and Supplemental Table S3). The existence of signal peptides and AG glycomodules suggested that the 10 CanXYLPs may be chimeric AGPs. To further study the XYLP protein specific feature, the nsLTP-like domains of the above XYLPs were aligned. The distribution of eight cysteine (C) residues was highly conserved with the formation of a C-X-C-X-CC-X-CXC-X-C-X-C region, whereas the hydrophobicity of the residue between 5th C and 6th C was also conserved with leucine (L), except CanXYLP2 and tyrosine (Y) (Fig. 2a and Supplemental Fig. S1). The conserved nsLTP-like domain was involved in the formation of a three-dimensional structure that could firmly bind lipids, implying their crucial ability to bind or transfer various types of hydrophobic molecules such as fatty acids, fatty acyl-CoA, phospholipids, glycolipids, and cutin monomers in vitro[38].
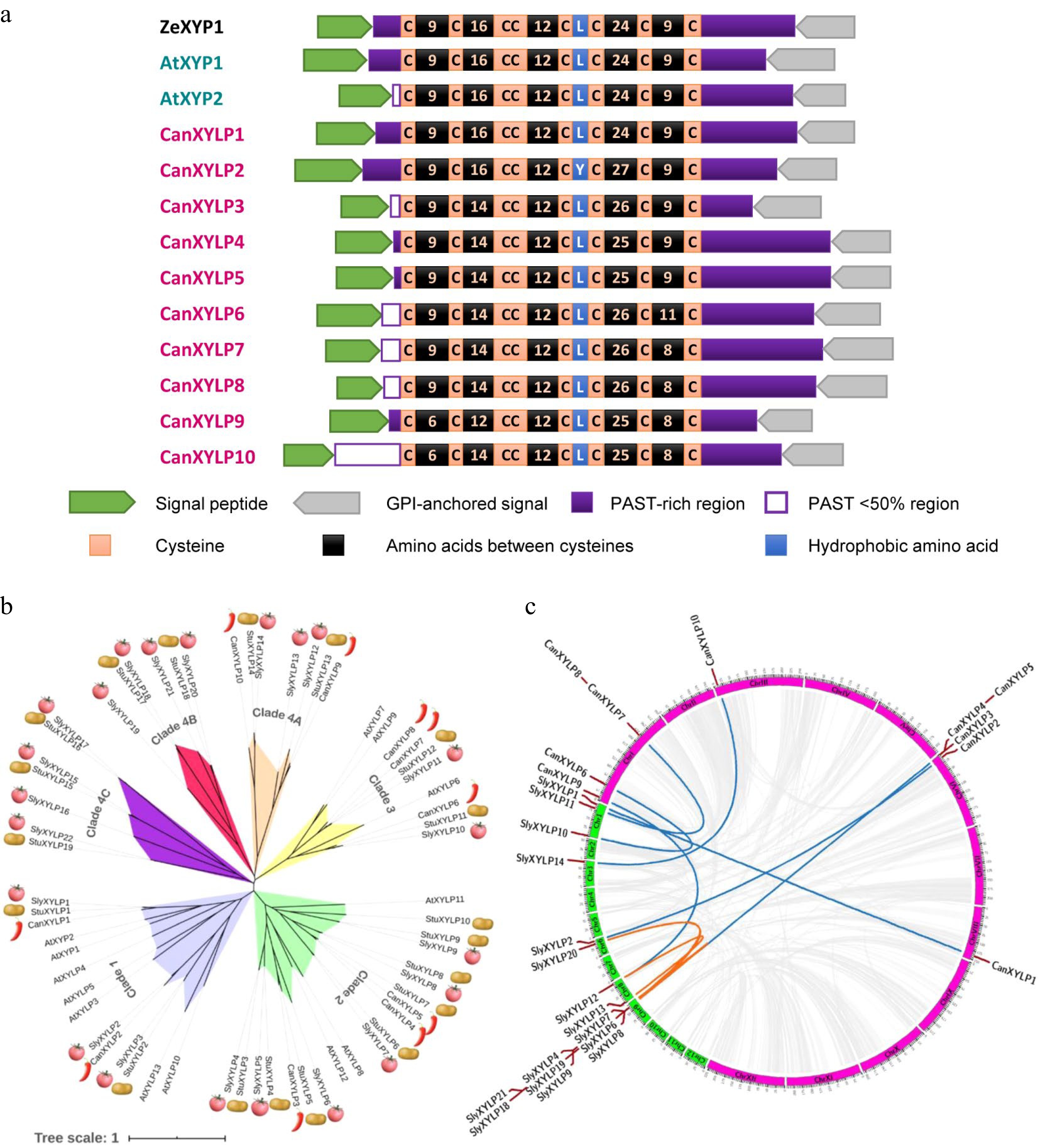
Figure 2.
Protein structure, Phylogenetic and syntenic relationship of XYLPs in Solanaceae. (a) Conserved motifs and structure domain of ZeXYP1, AtXYP1, 2 and 10 CanXYLP proteins. (b) Phylogenetic tree of 12 AtXYLPs, 22 SlyXYLPs, 19 StuXYLPs and 10 CanXYLPs. (c) Comparative synteny of the XYLP gene family between pepper and tomato. Blue lines show interspecific collinearity relationship, orange lines show intraspecific collinearity relationship and gray lines show collinearity relationship of all genes.
Phylogenetic and synteny analysis of XYLPs in Solanaceae
-
The present study identified 22 SlyXYLP proteins in tomato (Solanum lycopersicum) and 19 StuXYLP proteins in potato (Solanum tuberosum) to fully understand the XYLPs among nightshade (Supplemental Table S2). The Solanaceae XYLP protein full sequences were then used with 13 well-known AtXYLPs to obtain an unrooted phylogenetic tree exhibiting their phylogenetic relationships (Fig. 2b). According to the sequence homology of proteins, CanXYLPs, SlyXYLPs, StuXYLPs, and AtXYLPs proteins were divided into four independent evolutionary clades (1−4), and the family members with high sequence homology were divided into the same clade. Clades 1, 2, and 3 contained XYLPs from these four species, whereas clade 4 is nightshade specific, comprising three sub-clades (A−C). Clade 4B and 4C mainly contain XYLP protein from tomato and potato. The phylogenetic tree exhibited that the XYLP protein underwent different evolutionary pathways after the differentiation of Arabidopsis and Solanaceae plants. Additionally, the XYLP protein in Clade 4 can only appear in Solanaceae plants, indicating that the XYLP proteins of these subfamilies were isolated later.
Large-scale comparative synteny maps were analyzed at genome-wide levels to reveal the mechanism for expansion and evolution of the XYLP gene family between pepper and tomato (Fig. 2c). Eight pairs of XYLP genes were identified between pepper and tomato (Fig. 2c, blue lines and Supplemental Table S4) exhibited interspecific collinearity relationships, suggesting that most CanXYLPs were orthologous in tomato. Additionally, among the synteny events, six intraspecific collinearity pairs of XYLP genes were observed in tomato (Fig. 2c; orange lines and Supplemental Table S4), which were not observed in pepper, indicating more XYLP duplication in tomato.
Expression patterns of CanXYLP genes during pepper stem development
-
The real-time quantitative reverse-transcription polymerase chain reaction (qRT-PCR) was conducted to measure their relative expression levels at different stages of stem development to elucidate the roles of CanXYLPs in stem development (Fig. 3). The stems of six stages were harvested from 2 weeks to 12 weeks after germination. The expression levels of 10 CanXYLP genes during stem development could be divided into three categories, namely the expression levels of CanXYLP1, 2, and 10, which were gradually increased during stem development and reached the peak at 12 weeks. On the other hand, CanXYLP7 and 8 were highly expressed in the early stage and then gradually decreased with plant growth, suggesting that these genes may be closely related to stem lignification. Both CanXYLP3 and 9 exhibited an expression peak at 4 weeks, whereas CanXYLP4, 5, and 6 groups peaked at 6 weeks. The two pair of tandem replication genes, CanXYLP4 and 5, and CanXYLP7 and 8, exhibited the same expression pattern, indicating that they may have the same biological function. The different expression pattern of these 10 CanXYLP genes during pepper stem growth indicated their different function in this process.
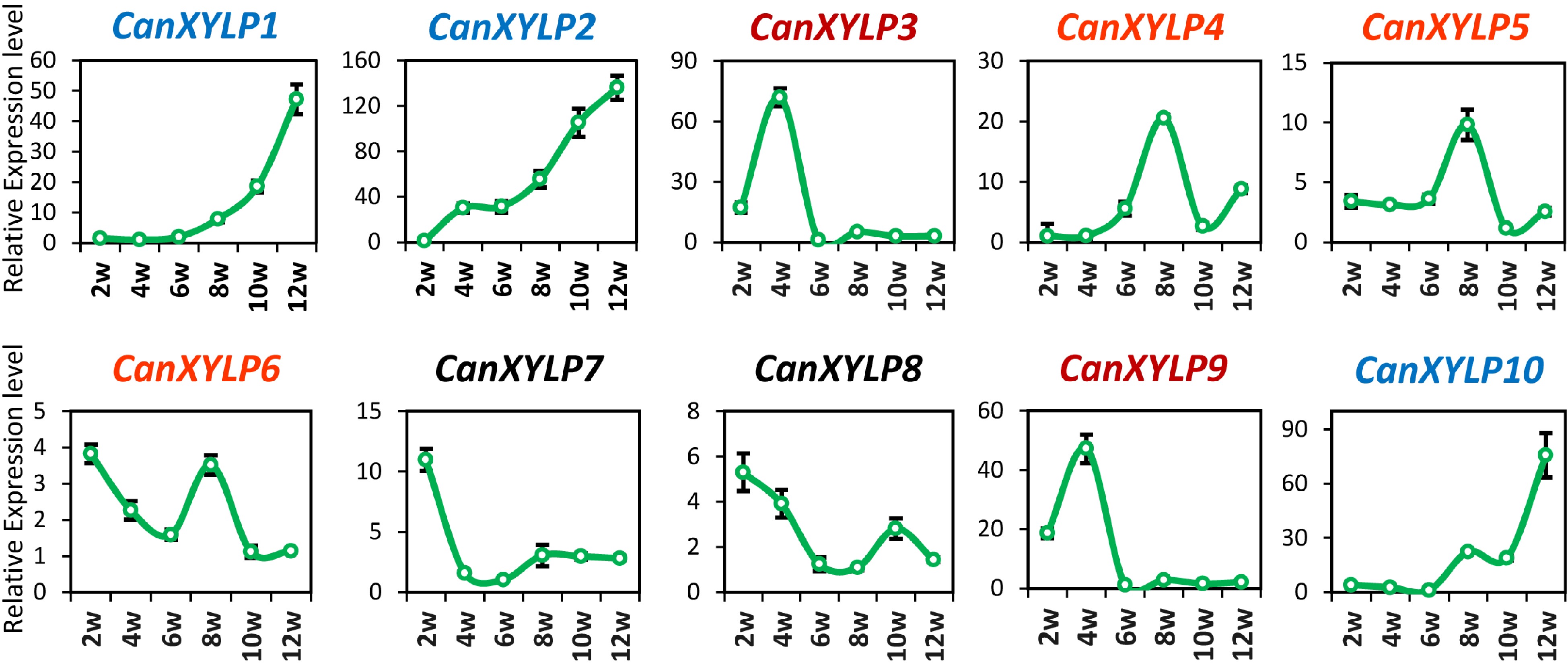
Figure 3.
Expression analysis of CanXYLP genes during pepper stem growth. Relative expression level of 10 CanXYLPs in the stems of pepper at 6 developmental stages from 2W to 12W.
Virus induced gene silening of CanXYLP1/2 leading to reduced lignin contents in pepper stem
-
Previous studies have exhibited the vital role of AtXYP1 and AtXYP2 in vascular development[25]. The present study focused on CanXYLP1 and CanXYLP2 to explore their function in pepper. By using the virus-induced gene silencing (VIGS) approach, we have either CanXYLP1 (TRV2:CanXYLP1) or CanXYLP2 (TRV2:CanXYLP2), as well as both of CanXYLP1 and 2 gene silenced pepper 'HP' plants (TRV2:CanXYLP1+2) (Fig. 4a). The qRT-PCR results exhibited that the CanXYLP1 and CanXYLP2 gene expression decreased in these VIGS plants (Fig. 4d, e). As a technical marker, the PDS (Phytoene desaturases) gene-silenced plant exhibited bleaching of the leaves and stem (Fig. 4b, c), indicating that the VIGS method was effective and could be used for stem development analysis. All of the stems from TRV2:CanXYLP1, TRV2:CanXYLP2, and TRV2:CanXYLP1+2 plants were much thinner than the negative control TRV2:EV (Fig. 4a, g), which might be due to the blocked xylem development. Thus, we next investigated the lignin content in these stems, and a significant decrease was observed in the CanXYLP1/2-silenced plants (Fig. 4a, h). Furthermore, the region with less lignin was demonstrated after phloroglucinol hydrochloric acid staining (Fig. 4a, i), indicating that both CanXYLP1 and 2 are necessary during pepper stem development and lignification, and the loss of either of them will affect lignin biosynthesis.
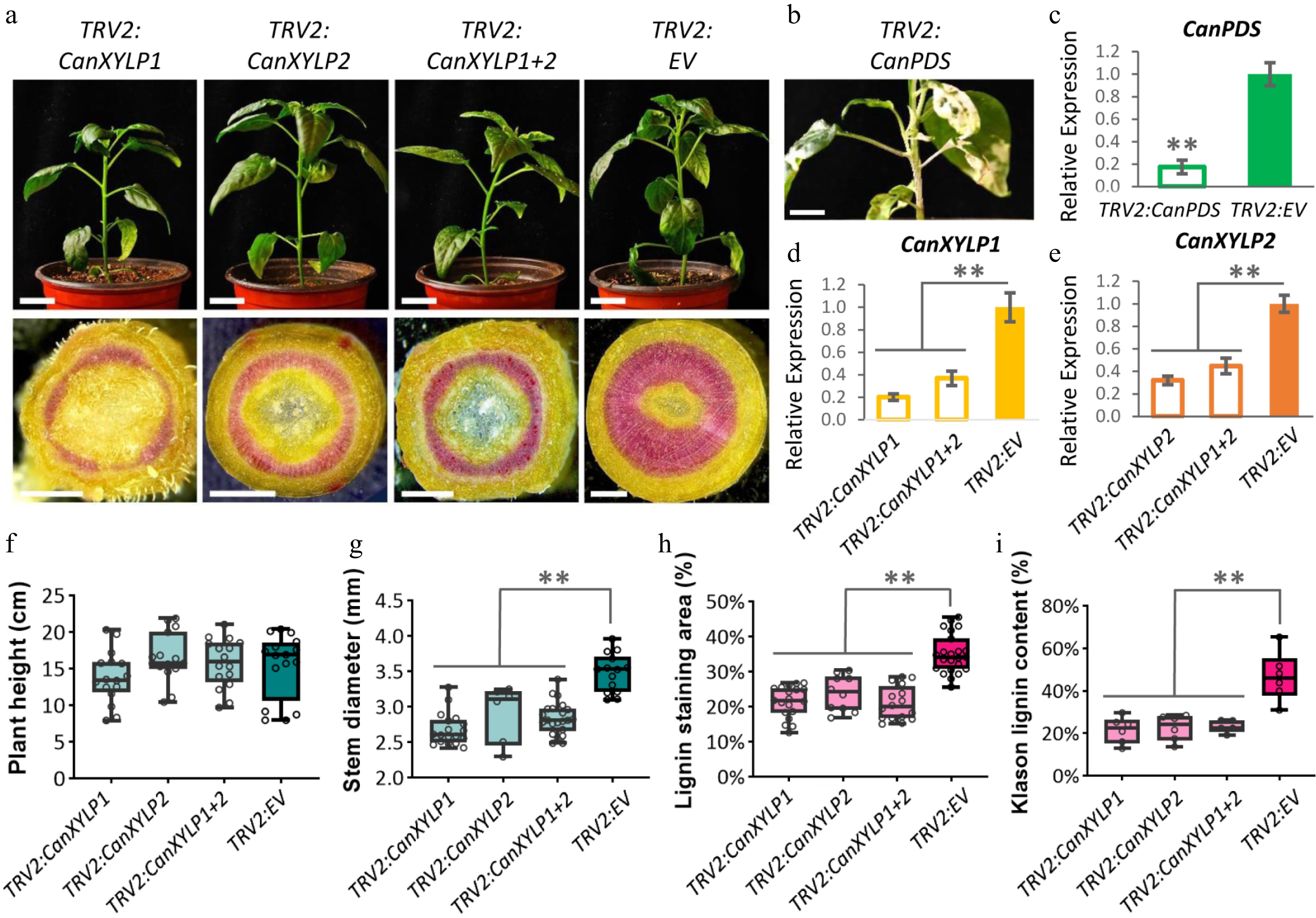
Figure 4.
CanXYLP1 and 2 silencing significantly reduced the lignin contents in pepper stem. (a) Phenotypes of TRV2:CanXYLP1/TRV2:CanXYLP2/TRV2:CanXYLP1+2/TRV2:EV + TRV1 expressed pepper plants (Bars = 2 cm) and the cross sections of their stems (Bars = 1 mm) at 10W stage. (b) Bleached stem of pepper plant expressed TRV2:CaPDS. Bar = 2 cm. (c) Relative expression of CanPDS gene after TRV2:CanPDS/TRV2:EV + TRV1 infiltrated stems at 10W stage. (d) Relative expression of CanXYLP1 in TRV2:CanXYLP1/TRV2:CanXYLP1+2/TRV2:EV + TRV1 infiltrated stems at 10W stage. (e) The relative expression of CanXYLP2 in TRV2:CanXYLP2/TRV2:CanXYLP1+2/TRV2:EV + TRV1 infiltrated stems. (f)-(i) Comparation of plant height (f), stem diameter (g), ratio of area stained by phloroglucinol hydrochloric acid to the entire cross section (h) and Klason lignin content (i) among TRV2:CanXYLP1+TRV1, TRV2:CanXYLP2 + TRV1, TRV2:CanXYLP1+2 + TRV1 and TRV2:EV + TRV1 infiltrated stems at 10W stage (**, P < 0.01).
Downregulation of lignin biosynthesis genes in CanXYLP1/2-silenced plants
-
The knockdown of either or both CanXYLP1 and CanXYLP2 exhibited an obviously narrowed stem phenotype and inhibition of lignin production. The expression of some genes associated with lignin biosynthesis might be changed. Therefore, RNA Sequencing (RNA-seq) was conducted to discover the differentially expressed genes (DEGs) from the stem samples of TRV2:CanXYLP1 (TX1), TRV2:CanXYLP2 (TX2), TRV2:CanXYLP1+2 (TX1_2), and TRV2:EV (EV) at 10W. The RNA-seq experiment was repeated with two independent biological replications. The RNA-seq approach produced approximately 10 Gb of data for each sample and approximately 93% of the reads were mapped to the pepper genome (Supplemental Table S5 and Supplemental Fig. S2a), ensuring that the RNA-seq results were relatively credible. Thousands of genes were specifically differently expressed in TX1, TX2, and TX1_2 compared with the EV. There were 3,113 upregulated DEGs and 3,311 downregulated DEGs in the TX1 compared with the EV, 2,839 upregulated DEGs and 2,138 downregulated DEGs in the TX1 compared with the EV, and 1,910 upregulated DEGs and 1,394 downregulated DEGs in the TX1_2 compared with the EV (Fig. 5b). Overlapping data were chosen for further analysis due to data redundancy among TX1/EV, TX2/EV, and TX1_2/EV. Finally, 651 upregulated DEGs and 914 downregulated DEGs were obtained by sequencing (Fig. 5c). Then, the DEG data were used to identify the weakened gene expression profiles involved in the phenylpropanoid metabolic pathway in the TX1, TX2, and TX1_2 samples to explore the relationship between DEGs and lignin biosynthesis by KEGG (Kyoto Encyclopedia of Genes and Genomes) analysis (Supplemental Fig. S2b). Additionally, the transcript levels of the genes sharply decreased in all of TX1, TX2, and TX1_2 samples, including most essential enzymes that make up the lignin biosynthesis pathway (Fig. 5a and Supplemental Table S6). Furthermore, the qRT-PCR results for CanPAL2, CanCCR3, CanPER5, CanCCoA-OMT2, and CanCOMT were consistent with the FPKM (Fragments per Kilobase Million) results (Fig. 5d), indicating that the gene downregulation related to lignin biosynthesis was consistent with the lignin content being dramatically reduced due to loss of function of CanXYLP1/2.
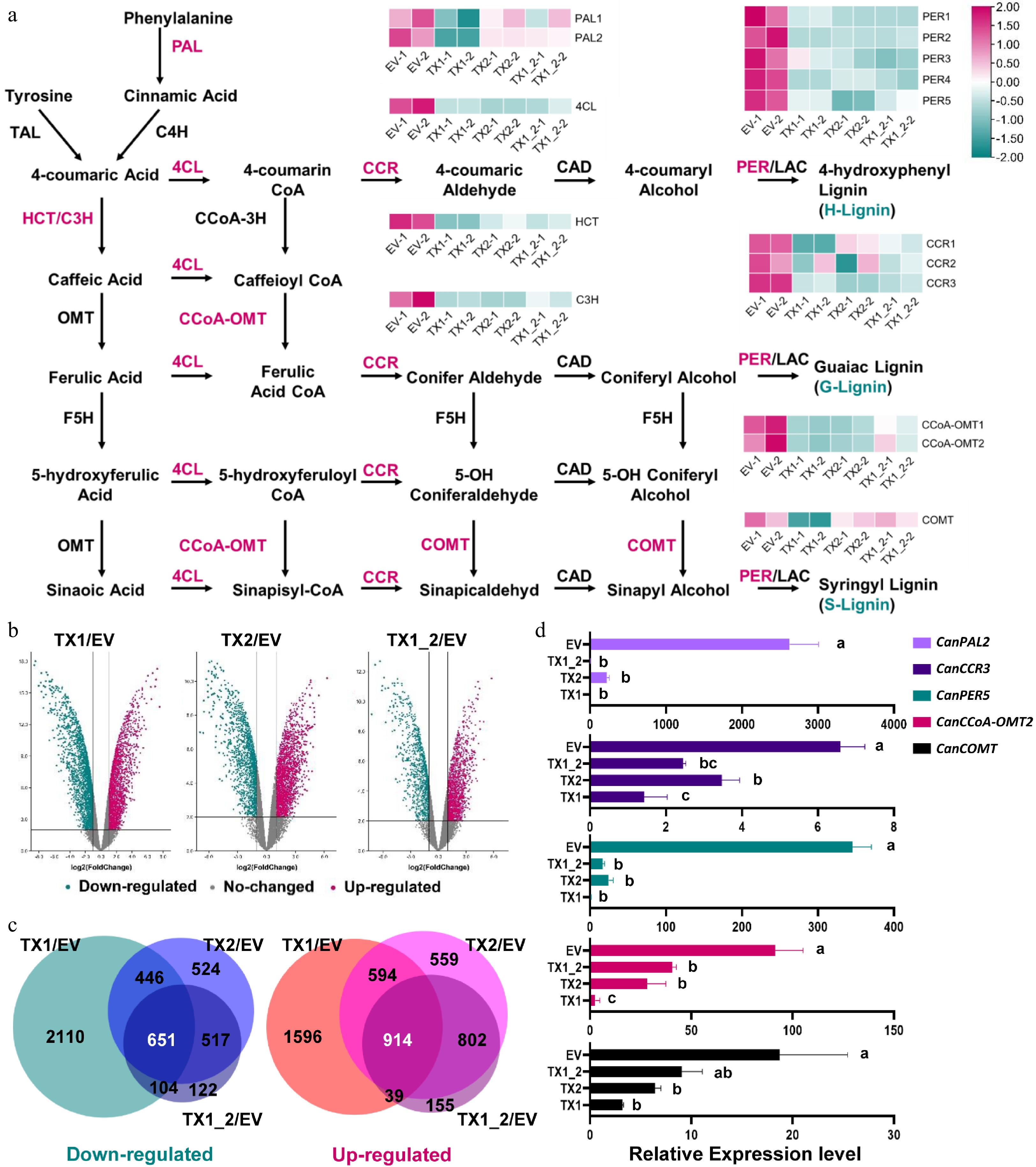
Figure 5.
CanXYLP1 and 2 function on lignin synthesis pathway in pepper. (a) Heatmap of transcript levels in FPKM for genes that are involved in the lignin biosynthetic pathway and are distinctly expressed among TX1, TX2, TX1_2 and EV. (b) Volcano plot of DEGs. Red dots indicate up-regulation, blue dots indicate down-regulation, gray dots indicate unchanged. (c) Venn diagram analysis of lower (left) and higher (right) expressed genes in TX1/EV, TX2/EV and TX1_2/EV. (d) qRT-PCR analysis verified the down-regulated relative expression levels of CanPAL2, CanCCR3, CanPER5, CanCCoA-OMT2 and CanCOMT in TX1, TX2, TX1_2 compared with EV (P < 0.05).
CanXYLP1 and 2 gene response to biotic and abiotic stresses
-
The cis-regulatory elements in CanXYLP1 and CanXYLP2 promoter sequences were analyzed to further study the upstream regulation mechanism of CanXYLP1 and CanXYLP2 genes. The promoter regions (1,500 bp of genomic DNA sequence) upstream of the translation starts site were submitted in the PlantCARE database[36]. In addition to the basic TATA and CAAT boxes, a large number of cis-acting elements involved in biotic and abiotic stresses were identified both in the promoter of CanXYLP1 and CanXYLP12 (Fig. 6a), including cold (Fig. 6a, green triangles), drought (Fig. 6a, dark blue triangles), ABA (Fig. 6a, red triangles), GA (Fig. 6a, orange triangles), and JA (Fig. 6a, light blue triangles), suggesting that these two genes may respond to the stresses. Then, the expression pattern of CanXYLP1 and CanXYLP2 in roots was investigated after biotic and abiotic treatment in the Pepperhub database[39]. Thus, the expression level of both CanXYLP1 and CanXYLP2 was activated after treated for 1–6 h (Fig. 6b, c). Additionally, qRT-PCR assay proved that compared with the mock treatment, CanXYLP1 and CanXYLP2 genes exhibited an increased expression under all the five treatments (Fig. 6d, e). Thus, CanXYLP1 and CanXYLP2 genes could be upregulated by biotic and abiotic signals, indicating that lignin could protect plants from various stresses. Additionally, the MYB transcriptional factor binding element was also observed in the promoter region of both CanXYLP1 and CanXYLP2 genes (Fig. 6a, purple triangles), suggesting that CanXYLP1 and CanXYLP2 participate in the lignin biosynthesis in pepper stem through MYB regulation.
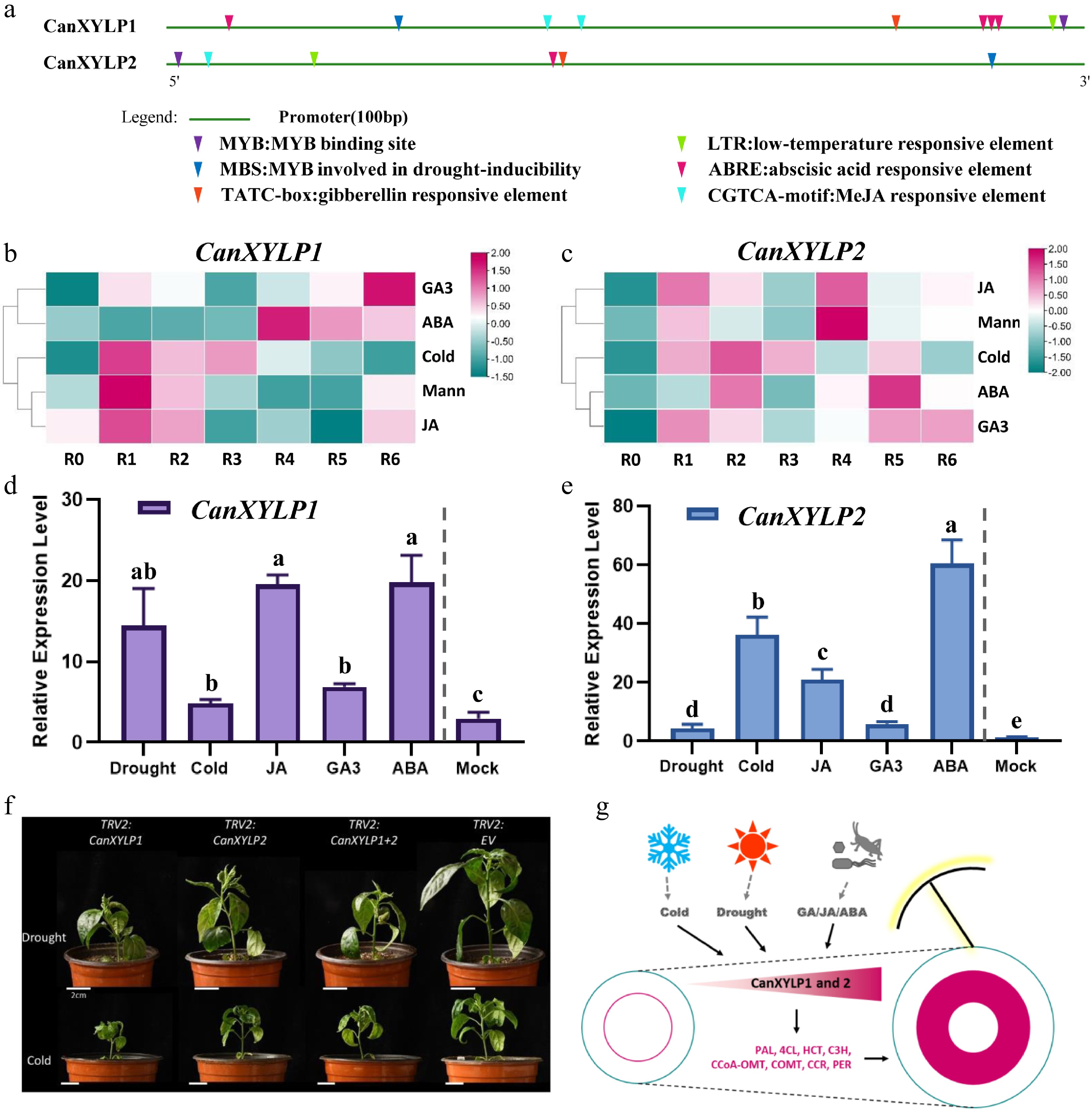
Figure 6.
CanXYLP1 and 2 have positive responses to biotic and abiotic stresses. (a) Promoter cis-regulatory elements analysis of CanXYLP1 and CanXYLP2 promoter sequences. (b), (c) Heatmap showing the expressing level of CanXYLP1(b) and CanXYLP2 (c) gene after biotic (GA3, ABA, JA) and abiotic (Cold, Mannitol) treatment for 1-6 hours. (d, e) qRT-PCR analysis of the relative expression levels of CanXYLP1 (d) and CanXYLP2 (e) under above biotic and abiotic treatment (equally mixed R1-R6) compared with Mock (R0) (P < 0.05). (f) Phenotypes of TRV2:CanXYLP1/TRV2:CanXYLP2/TRV2:CanXYLP1+2/TRV2:EV + TRV1 expressed pepper plants under drought and cold stress. Bars = 2 cm. (g) Proposed model for CanXYLP1 and CanXYLP2 regulation of lignin biosynthesis during pepper stem development.
-
Xylogen is a glycoprotein involved in the differentiation of tracheary element (TE) and local intercellular communication. XYLPs are xylogen-like AGPs characterized by an AGP domain and a plant nsLTPs domain[29]. The present study identified 10, 22, and 19 XYLPs in pepper, tomato, and potato, respectively (Fig. 2b). All of the XYLP protein structures were well-characterized with specific AGP and nsLTPs domain (Supplemental Fig. S1, Supplemental Table S2), indicating that the XYLPs were conserved in Solanaceae. To investigate the relationship of XYLPs and enhanced lignification during pepper stem development (Fig. 1), the expression pattern of 10 XYLP genes was also analyzed in pepper (Fig. 3). CanXYLP1 and CanXYLP2 expression was closely related to lignin accumulation in pepper stem growth, which inspired us to continue to explore their functions during pepper stem development. Another study showed that xyp1xyp2 double mutant displayed the phenotype of vascular development defects in Arabidopsis[25]. The present study demonstrated that both CanXYLP1 and CanXYLP2 were crucial for pepper stem development and that either CanXYLP1 or CanXYLP2 gene silencing will cause lignin accumulation block in xylem (Fig. 4). The non-redundant function of XYLP1 and XYLP2 in pepper may cause their similar expression pattern, which differs from the redundant function of XYP1 and XYLP2 with different expression pattern in Arabidopsis[27]. Additionally, compared with Arabidopsis, pepper has evolved a certain number of spare genes to maintain its function normally under extreme conditions.
The present study observed that the transcript level of vital enzymes working in lignin biosynthesis pathway in the downstream of CanXYLP1 and CanXYLP2 had been downregulated in the CanXYLP1/2-silenced pepper stems (Fig. 5) and some abiotic and biotic related cis-elements in the upstream of CanXYLP1 and CanXYLP2, which will influence their expression after stress treatment (Fig. 6) such as drought, low temperature, GA, JA, and ABA. The lignin content and lignification degree were regulated by a variety of biotic and abiotic responses. Under drought stress, the ZmCAD and ZmCOMT expression levels in maize (Zea mays) leaves increased[40], and ZmCCR1/2 in maize root elongation zone was significantly upregulated[41]. Additionally, drought-induced ABA and JA exhibited a positive regulatory effect on CmCAD gene and lignin synthesis in melon (Cucumis melo) stems[42]. Furthermore, low temperature affected the JA expression and lignin biosynthetic pathway-related genes in bamboo (Phyllostachys violascens) shoots after harvest[43]. JA, GA, and ABA are vital hormones in plants response to stresses. Shi et al.[44] exhibited that ABA could accelerate the lignification of juice sacs in pummelo (Citrus grandis) fruit. GA3 regulated the formation of different types of lignin in Coleus Blumei (Plectranthus scutellarioides) stems[45]. As CanXYLP1 and CanXYLP2 were positively regulated by stress, more lignin content will help pepper to survive stresses such as drought and chilling. MYB transcriptional factors will also play essential roles during lignin biosynthesis in horticultural plants. CmMYB15 and CmMYB19 enhanced lignin accumulation and resistance of chrysanthemum (Chrysanthemum morifolium) to aphids[46,47]. MdMYB88 and MdMYB124 were the causes of lignin accumulation in apple under drought stress[48]. MYB binded cis-elements in the promoter region of both CanXYLP1 and CanXYLP2 (Fig. 6a, purple triangle), suggesting that CanXYLP1 and CanXYLP2 might be regulated by MYB too.
-
In the present study, 10 XYLP genes were identified from the pepper genome and classified into four clades according to their evolutionary relationships. Among them, the most interesting CanXYLP1 and CanXYLP2 have an increased expression pattern along with pepper stem development. Further experiments discovered that silencing CanXYLP1 and CanXYLP2 alone or together can significantly reduce lignin deposition and their upstream and downstream expression regulation, indicating that CanXYLP1 and CanXYLP2 genes may be involved in lignin biosynthesis through various stresses regulation, which provide a novel insight into the characteristics of XYLPs in pepper stem lignification.
This study was supported by the National Key Research and Development Program of China (2019YFD1000301), the National Natural Science Foundation of China (32172600, 31972420) and the Fundamental Research Funds for the Central Universities (2662018QD020).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 The detailed information of XYLP proteins characterize in pepper, tomato and potato.
- Supplemental Table S2 Protein backbones of XYLPs in pepper, tomato and potato.
- Supplemental Table S3 Collinearity Pairs of XYLPs between pepper and tomato.
- Supplemental Table S4 General information of transcriptome data.
- Supplemental Table S5 FPKM of lignin biosynthetic genes
- Supplemental Table S6 Primers used in this study.
- Supplemental Fig. S1 Consensus sequence of nsLTP domain among CanXYLP proteins.
- Supplemental Fig. S2 FPKM (Fragments per Kilobase Million) distribution and KEGG (Kyoto Encyclopedia of Genes and Genomes) enrichment analysis.
- Supplemental Method Identification of XYLP proteins.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang M, Zhang Q, Cheng L, Li Q, He X, et al. 2022. Pepper (Capsicum annuum) xylogen-like arabinogalactan protein (XYLP) 1 and XYLP2 promote synthesis of lignin during stem development to cope with stresses. Vegetable Research 2:15 doi: 10.48130/VR-2022-0015 

Pepper (Capsicum annuum) xylogen-like arabinogalactan protein (XYLP) 1 and XYLP2 promote synthesis of lignin during stem development to cope with stresses
- Received: 10 August 2022
- Accepted: 10 October 2022
- Published online: 28 October 2022
Abstract: Pepper stems exhibit a high level of strength and lignin deposition to support plant growth, which direct the cultivation style without binding sticks or scaffolding. However, regulation of lignin synthesis and accumulation in pepper stem has not been extensively studied. Herein, we first investigated the pepper stem developmental process and confirmed that increasing lignin accumulation occurs during stem growth. We then performed genome-wide identification and characterization of xylogen-like arabinogalactan protein (XYLP) family members and obtained 10, 22, and 19 XYLPs in pepper, tomato, and potato respectively. Evaluation of the phylogenetic relationship among the identified XYLPs suggested that these proteins are conserved in Solanaceae. Thereafter, we analyzed the 10 CanXYLP genes and observed that these genes exhibit differential expressing patterns at different stages of pepper stem development. Among these genes, two XYLPs, namely CanXYLP1 and CanXYLP2, exhibited an increased expression pattern and a strong correlation with lignin accumulation in pepper stem. We further found that CanXYLP1 and CanXYLP2 play a role in pepper stem lignification by positively regulating the lignin synthesis pathway genes in pepper, and the CanXYLP1/2-silenced plants displayed a blocked lignification phenotype. Finally, we confirmed that CanXYLP1/2 expression is upregulated in response to some abiotic and biotic signals, suggesting that these two genes enhance the tolerance of pepper stem to unfavorable conditions. These results contribute to our understanding of the molecular mechanism controlling pepper stem lignification, and the relationship between the lignin content of pepper stem and XYLPs.
-
Key words:
- Capsicum annuum /
- Xylogen-like arabinogalactan protein /
- Stem /
- Lignin /
- Stresses