









-
Pummelo (C. grandis) is a prevalent plant of the family Rutaceae belonging to evergreen subtropical citrus trees. The 'Guanximiyou' pummelo variety has been widely cultivated in China for more than 400 years, and is known for being rich in carbohydrates, β-carotene, vitamin B1, vitamin B2, vitamin C, calcium, potassium, phosphorous, and other health-promoting compounds[1−4]. 'Guanximiyou' pummelo and its bud mutants 'Hongroumiyou' [Citrus maxima (Burm.) Merr. 'Hongroumiyou'] and 'Sanhongmiyou' [Citrus maxima (Burm.) Merr. 'Sanhongmiyou'] varities are highly affected by juice sac granulation[3].
Citrus fruits are prone to a variety of physiological disorders during the harvest and storage periods. Granulation is an undesirable condition affecting juice sacs of citrus fruits, which become dry, enlarged, grayish, hardened, and less detachable[1]. Granulation was first reported in navel orange in 1934 by Bartholomew et al.[2] in California, being later reported in many fruits, such as pummelo, grapefruit, lemon, and lime[3, 4]. Granulation is often accompanied by enlarged, dried, stiffened, and inflated juice sacs[4−6]. Granulation eventually leads to decreased nutritional and commodity value, which represents significant economic loss[6, 7].
In our previous works, granulated juice sacs showed lower contents of citrate and isocitrate, and consequently lower acidity, which could be attributed to increased juice sac degradation. Moreover, granulation was also associated with increased accumulation of mineral elements [i.e., phosphorus (P), copper (Cu), magnesium (Mg), sulphur (S), and zinc (Zn)] in juice sacs, which might be involved in the occurrence of the granulation phenomenon in pummelo[8]. In fact, previous studies suggested that accumulation of mineral elements in juice sacs may be one of the causes leading to granulation in citrus fruits[9, 10]. For instance, Xie et al. found that high levels of P in juice sacs were associated with higher incidence of granulation in C. grandis[6], an observation that was consistent with alterations described in 'Dancy' tangerine[9] and 'Valencia' orange fruits[10] in other studies. In addition, long-term utilization of phosphatic fertilizer in orchards might induce accumulation of P in fruits. In our previous work, Cu concentration was shown to be higher in granulated juice sacs than in normal ones[8], which is in agreement with previous findings that accumulation of Cu in leaves occurred as granulation progressed in two sweet orange cultivars[11]. Collectively, evidence suggests that granulation is likely associated with increased accumulation of mineral elements (especially P, Cu, Mg, S, and Zn) in pummelo juice sacs.
A variety of complex factors contribute to the occurrence of granulation, such as higher application rate of nitrogen or phosphatic fertilizers, higher irrigation frequency, delayed fruit harvest, and abundant growth after heavy pruning or fertilization[4, 12, 13]. Wu et al. found that abnormal accumulation of lignin in juice sacs was associated with the occurrence of juice sac granulation in pummelo (C. grandis)[3]. Moreover, key genes involved in main lignin synthetic pathways were found to be expressed exclusively in granulated juice sacs[14]. Furthermore, Awasthi & Nauriyal reported that activity of peroxidase and superoxide dismutase was associated with higher incidence rate of granulation[15]. Sharma et al. also found that the activity of enzymes related to antioxidants, phenyl ammonia-lyase, and total phenolic compounds has a strong negative correlation in granulated juice sacs[16]. In contrast, senescence-related enzymes, such as pectin methyl esterase, lipoxygenase, as well as respiration rates or ethylene production were shown to have a strong positive relationship with the occurrence of granulation in 'Kinnow' mandarin[13, 16]. Collectively, previous studies suggest that granulation is a complex and recurrent phenomenon whose underlying molecular mechanisms are largely unknown. Therefore, it is highly important to elucidate the basis of granulation in citrus fruits.
In this study, cDNA-amplified fragment length polymorphism (cDNA-AFLP) was applied to differentiate normal and granulated C. grandis juice sacs in order to understand differences in gene expression during pummelo juice sac granulation.
-
Using a total of 256 primer combinations, differentially expressed TDFs were identified in normal and granulated C. grandis juice sacs (Fig. 1). Supplemental Table S1 shows cDNA-AFLP profiles using one EcoR I selective primer and eight Mes I selective primers. As shown in Table 1, 4,424 clear and legible TDFs were obtained in pummelo juice sacs. Interestingly, 116 granulation-associated genes showed significant homology to genes encoding known or putative proteins. Among these, 41 TDFs were detected in normal juice sacs, 61 TDFs were detected in granulated juice sacs, and seven TDFs were upregulated and seven TDFs were downregulated in granulated juice sacs. According to functional analysis, these TDFs were assigned to different biological processes, such as hormone and biological regulation (12 TDFs, 10.08%), carbohydrate and energy metabolism (16 TDFs, 13.45%), protein and nucleic acid metabolism (48 TDFs, 40.34%), lipid metabolism (five TDFs, 4.2%), stress response and defense (13 TDFs, 10.92%), cell metabolism (12 TDFs, 10.08%), and unknown biological processes (13 TDFs, 10.92%) (Fig. 2).
Table 1. Summary of transcript-derived fragments (TDFs) in normal and granulated juice sacs of C. grandis.
Found only in normal Found only in granulated Found in both juice sacs Total Total TDFs detected 536 874 3,014 4,424 Total differentially expressed TDFs detected 68 89 26 183 TDFs produced useable sequence data 41 61 14 116 TDFs encoding known or putative proteins 38 57 11 106 TDFs encoding predicted, uncharacterized 8 9 2 19 TDFs without matches in the database 11 26 4 41 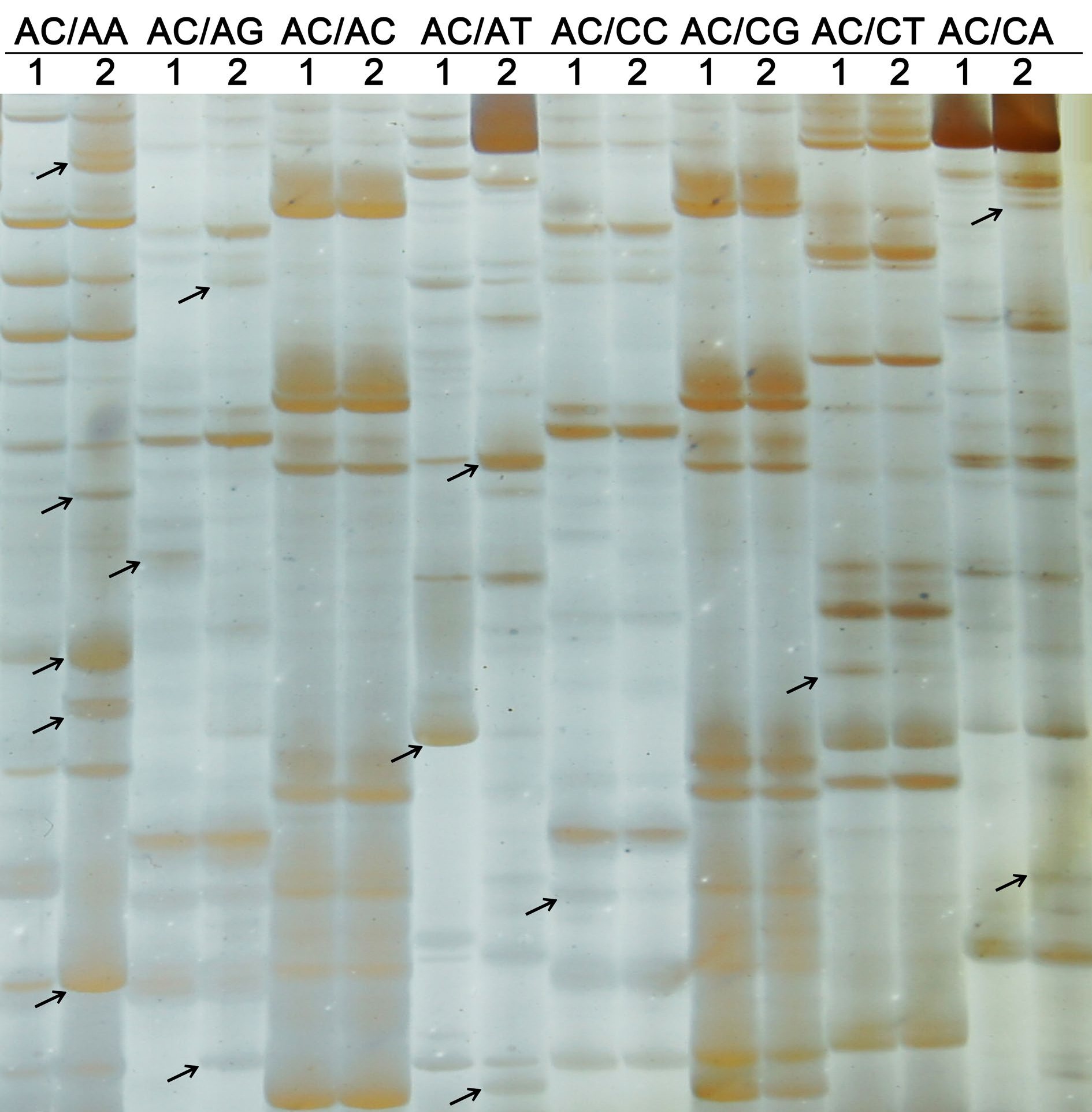
Figure 1.
cDNA-AFLP profiles using one EcoR I selective primer and eight Mes I selective primers. One EcoR I selective primer: EcoR I-AC; Eight Mes I selective primers: Mes I-AA, AG, AC, AT, CC, CG, CT, and CA; Lane 1: Normal juice sacs of C. grandis; Lane 2: Granulated juice sacs of C. grandis; Arrows indicate differentially expressed transcript-derived fragments.
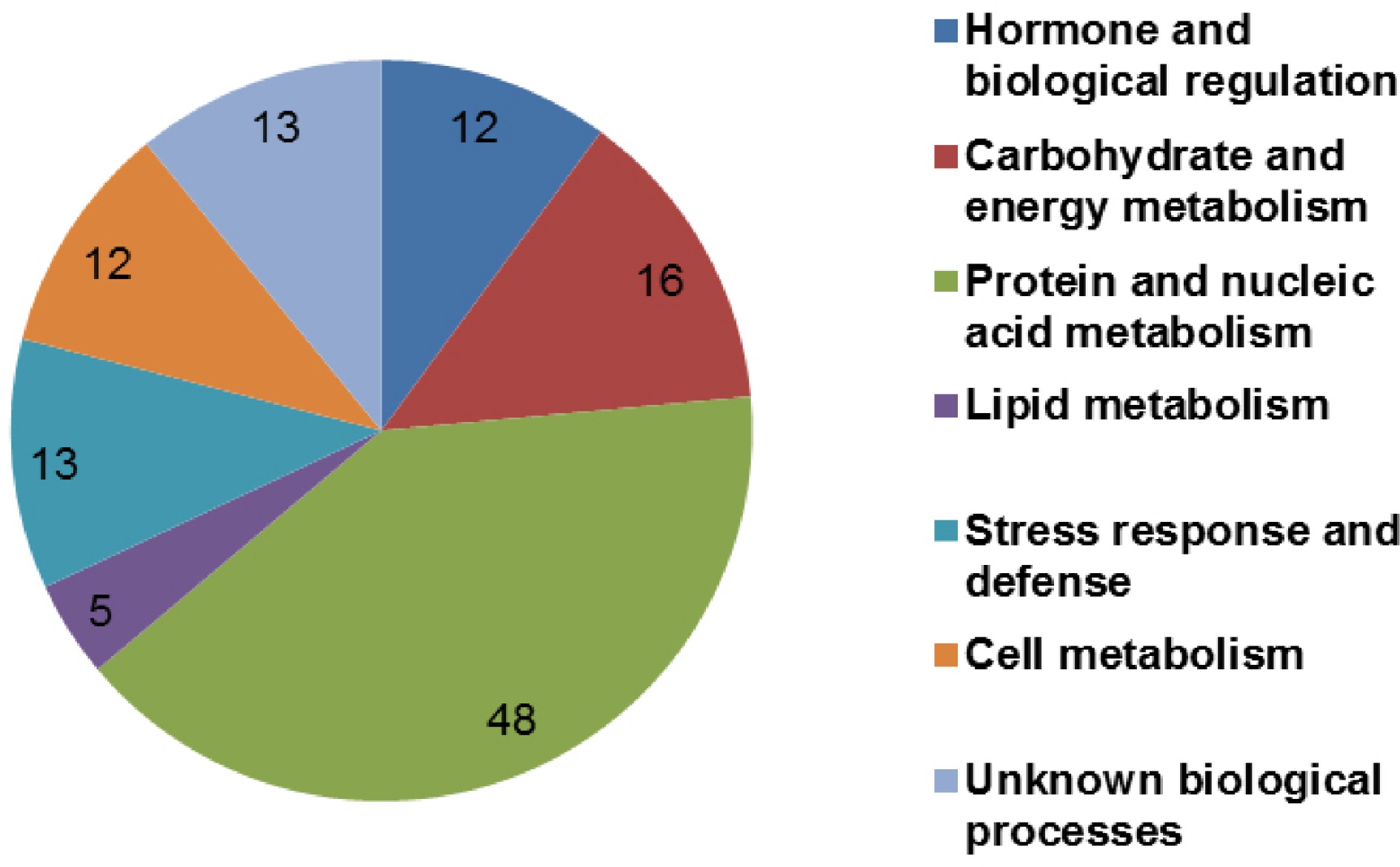
Figure 2.
Functional classification of differentially expressed transcript-derived fragments (TDFs) in normal and granulated juice sacs of C. grandis. Functional classification was performed based on information reported for each sequence in the NCBI database (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ).Validation of expression patterns of differentially expressed TDFs using qRT-PCR
-
As shown in Fig. 3, 20 TDFs were selected for qRT-PCR analysis in order to confirm cDNA-AFLP expression patterns. These TDFs were selected based on significantly different expression patterns in C. grandis granulated juice sacs and a high degree of homology with genes that play very important roles in various metabolic pathways. Expression levels of selected TDFs corroborated cDNA-AFLP findings, except for TDF #246-4 (Fig. 3). This discrepancy might indicate a gene family with complex regulation, which can be identified exclusively by the cDNA-AFLP technique.
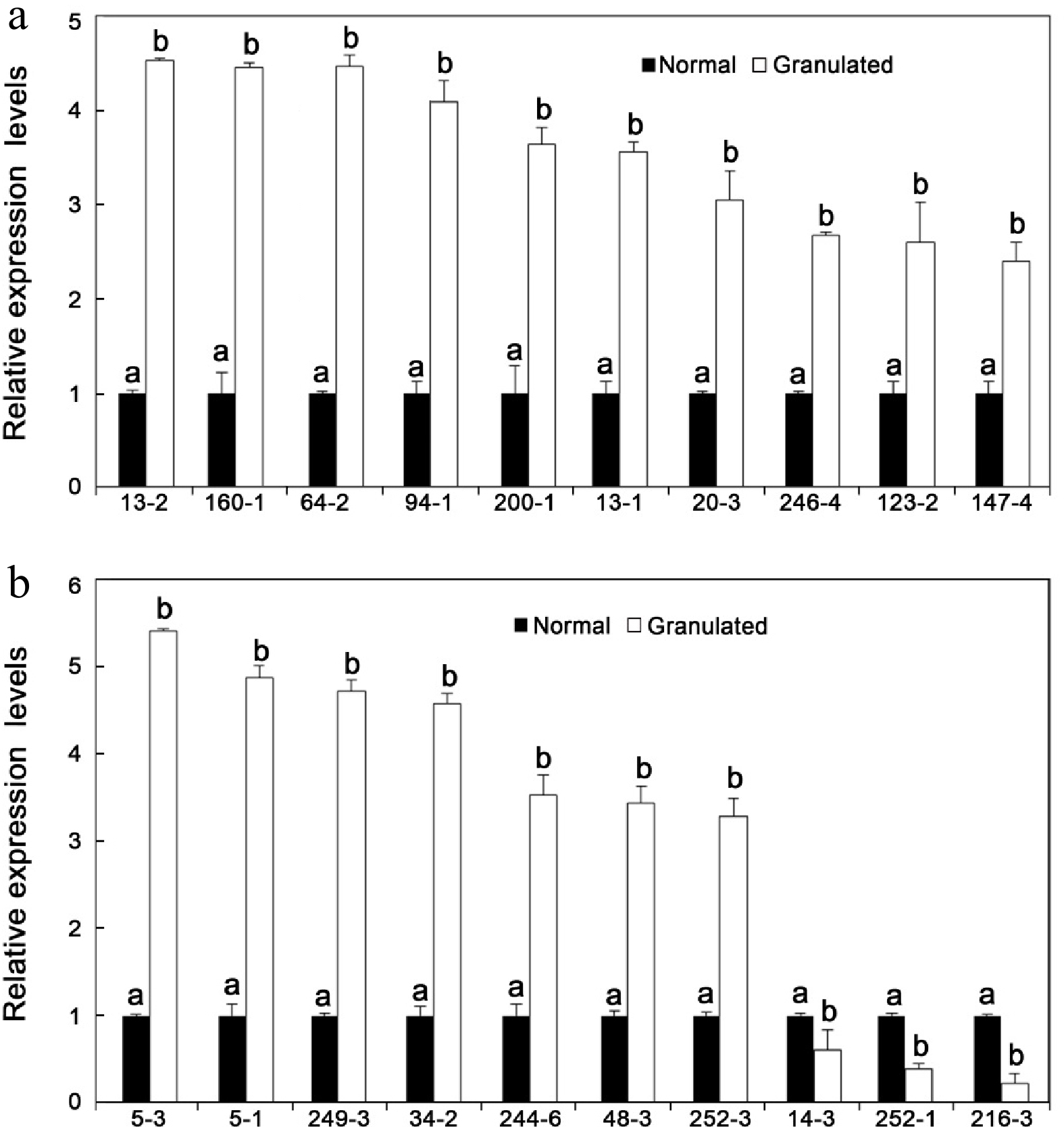
Figure 3.
Relative expression levels of transcript-derived fragments (TDFs) in C. grandis normal and granulated juice sacs. (a) Relative expression levels of genes encoding β-amylase 4 (TDF #64-2); cytokinin-O-glucosyltransferase 1 (TDF #94-1); gibberellin 20 oxidase (TDF #13-2); galactose-1-phosphate guanylyltransferases (TDF #200-1); α-galactosidase precursor (TDF #160-1); cytochrome b5 (TDF #123-2); ADP-ribosylation factor 3 (TDF #246-4); 2-oxoglutarate-dependent dioxygenase (TDF #13-1); 1,4-alpha-glucan-maltohydrolase (TDF #20-3); ethylene insensitive 3-like protein (TDF #147-4). (b) Relative expression levels of genes encoding translation initiation factor 4A2 (TDF #14-3); cytochrome P450 (TDF #48-3); auxin down-regulated-like protein (TDF #5-3); cellulose synthase (TDF #249-3); transport protein SEC31 (TDF #34-2); Ca2+-transporting ATPase (TDF #252-3); heat shock protein (TDF #252-1); senescence-associated protein (TDF #5-1); dicer-like protein 4 (TDF #244-6); cell wall-associated hydrolase (TDF #216-3). Results are shown as the mean ± SD of three independent experiments. Different letters above bars indicate significant differences at P < 0.05.
Concentrations of hormones in normal and granulated juice sacs
-
Four hormone-related genes were found to be potentially involved in the incidence of granulation in pummelo fruits: TDF #5-3 (auxin down-regulated-like protein); TDF #94-1 (cytokinin-O-glucosyltransferase 1); TDF #13-2 (gibberellin 20 oxidase); and TDF #147-4 (ethylene insensitive 3 like protein) (Table 2). Upregulation of hormone-related genes (i.e., TDFs #5-3, 94-1, and 147-4) observed in cDNA-AFLP analysis was further confirmed in qRT-PCR analysis (Fig. 3). To confirm discrepancies in hormone levels in normal and granulated juice sacs in pummelo, the contents of five hormones were determined using UPLC-MS. Contents of gibberellin 3 (GA3), gibberellin 20 (GA20), cytokinin (CK), and zeatin (ZT) were higher in normal juice sacs compared to granulated juice sacs, whereas the content of indole-3-acetic acid (IAA) was lower in normal juice sacs (Fig. 4).
Table 2. Homology of differentially expressed cDNA-AFLP fragments with known gene sequences in the database using BLASTX algorithm along with their expression patterns in granulation juice sacs of C. grandis.
TDF number Size
(bp)Homologous protein Organism of origin E-value Degree of similarity (%) Genbank ID Fold change Hormone and biological regulation 48-2 292 CAAX amino terminal protease family Cucumis melo 7E-18 90 ADN33781.1 + 5-3 259 Auxin down-regulated-like protein, partial Picea sitchensis 5E-05 55 ADM77850.1 + 6-2 158 Protein embryo defective 2752 Arabidopsis thaliana 2E-12 61 NP_567830.1 0 118-3 290 Expressed protein Oryza sativa 5E-27 63 ABF95726.1 0 94-1 358 Cytokinin-O-glucosyltransferase 1 Aegilops tauschii 9E-25 48 EMT28784.1 + 103-1 211 Nitrate reductase Cucumis sativus 1E-30 89 ADK77877 + 219-5 244 ARF-GAP domain 2 Arabidopsis lyrata subsp. lyrata 5E-09 43 NP_176283.1 0 13-2 269 Gibberellin 20 oxidase Medicago truncatula 4E-42 81 AES62614.2 + 147-4 271 EIN3-like (Ethylene insensitive 3) protein C. melo 8E-41 100 BAB64345 + 195-6 197 MATE efflux family protein Theobroma cacao 2E-19 79 EOX90702.1 0 115-1 355 Transmembrane emp24 domain-containing protein p24delta9-like Crocus sativus 3E-60 98 XP_004143772.1 + 195-1 455 Multicatalytic endopeptidase complex A. thaliana 8E-69 84 CAA74030.1 + Carbohydrate and energy metabolism 134-1 192 ATP-binding protein-like A. thaliana 2E-10 75 BAB09414.1 + 249-1 311 UDP-glucosyltransferase family 1 protein Camellia sinensis 2E-25 93 ACS87991.1 + 6-1 214 Fructose-bisphosphate aldolase Lemna minor 1E-33 94 ACD10928.1 0 5-4 148 ATP synthase subunit beta Medicago truncatula 2E-13 95 XP_003627732.1 + 118-2 304 Glyceraldehyde-3-phosphate dehydrogenase, partial Vernicia fordii 4E-11 94 AFJ04516.1 0 118-1 441 Glycosyltransferase, CAZy family GT8 Populus trichocarpa 5E-75 90 XP_002312381.1 + 64-2 220 β-amylase 4 Citrus trifoliata 2E-22 69 AFQ33616 + 249-3 217 Cellulose synthase Populus tremula × Populus tremuloides 5E-12 91 AAT09895.1 + 141-3 499 Mitochondrial benzaldehyde dehydrogenase Antirrhinum majus 8E-89 80 ACM89738.1 + 160-1 174 α-galactosidase precursor Coffea arabica 2E-04 62 CAJ40777.1 + 13-1 269 2-oxoglutarate-dependent dioxygenase Populus trichocarpa 8E-48 89 XP_002330269.1 7.07 ± 0.52 200-1 389 Galactose-1-phosphate guanylyltransferases T. cacao 1E-65 83 EOY12255.1 + 20-3 220 1,4-alpha-glucan-maltohydrolase Solanum lycopersicum 1E-14 60 NP_001234052.1 + 246-3 192 ABC transporter family protein P. trichocarpa 1E-12 80 XP_002310031.2 0 221-3 123 Diacylglycerol kinase-like protein A. thaliana 6E-11 69 BAB09587.1 + 246-4 172 ADP-ribosylation factor 3 A. lyrata subsp. lyrata 4E-98 96 XP_002869315.1 0 65-1 363 Methylenetetrahydrofolate reductase family protein isoform 3 T. cacao 7E-49 72 EOY04345.1 0 Protein and nucleic acid metabolism 228-1 243 Ribonucleoside-diphosphate reductase
subunit M1A. thaliana 2E-33 89 AEC07222.1 0 251-1 188 BET1P/SFT1P-like protein 14A A. thaliana 6E-07 78 NP_191376.1 0 253-2 306 Class II aaRS and biotin synthetases
superfamily proteinA. thaliana 5E-41 83 NP_186925.4 + 130-3 193 Ribosomal protein L5 Citrullus lanatus 1E-30 96 YP_003587255.1 0 119-3 254 Nuclear transport factor 2 family protein T. cacao 1E-22 62 EOY06196.1 + 27-3 224 Glutathione S-transferase family protein T. cacao 1E-07 62 EOY27562.1 + 195-5 268 BRCA1-associated protein M. truncatula 8E-39 73 XP_003609376.2 0 54-2 169 Pre-mRNA splicing factor PRP38 family protein P.trichocarpa 2E-22 92 ERP53525.1 0 151-4 347 Ribonuclease II family protein A. thaliana 2E-51 80 NP_565418.1 + 127-1 180 Mitochondrial substrate carrier family protein isoform 2 T. cacao 2E-28 61 EOY07093.1 + 15-2 349 Nuclear transcription factor Y subunit B18 M. truncatula 1E-09 68 AFK49668.1 + 15-1 392 RRNA intron-encoded homing endonuclease M. truncatula 6E-13 88 XP_003614385.1 + 20-1 280 Solute carrier family 25 member M. truncatula 6E-23 63 XP_003615848.2 0 221-1 256 TPA: heterogeneous nuclear ribonucleoprotein A3-like protein 2 isoform 1 Zea mays 1E-29 82 DAA58966.1 0.56 ± 0.06 217-5 194 Adenine nucleotide alpha hydrolases-like superfamily protein T. cacao 2E-06 96 EOY06709.1 1.86 ± 0.34 215-1 363 Transcription regulator A. thaliana 1E-06 88 NP_171710.4 + 195-4 333 PLP-dependent transferases superfamily protein A. thaliana 2E-37 60 NP_191772.1 + 21-2 210 60S ribosomal protein L24-1 T. cacao 1E-37 97 EOY23121.1 + 155-3 187 Valyl-tRNA synthetase/valine-tRNA ligase T. cacao 3E-19 75 EOY31957.1 2.23 ± 0.11 252-2 299 ACT domain-containing protein ACR8 Ricinus communis 2E-30 86 XP_002509632.1 0.79 ± 0.03 151-5 347 Exosome complex exonuclease RRP44
homolog AR. communis 5E-53 82 XP_002521738.1 + 217-2 369 Transcription initiation factor TFIID subunit A A. thaliana 1E-22 77 NP_564023.1 + 218-1 239 Chaperonin 60 alpha subunit Arachis diogoi 1E-20 86 ACA23472.1 0 31-1 293 Aspartyl protease family protein A. thaliana 3E-23 57 XP_002891474.1 + 236-3 251 Protein kinase domain-containing protein A. thaliana 3E-30 80 AEE27605.1 0 247-1 324 Importin beta-2 subunit family protein A. thaliana 2E-16 76 XP_002867489.1 + 253-1 263 Serine/threonine protein kinase TNNI3K M. truncatula 2E-09 76 XP_003601186.1 0 5-2 151 Amino acid adenylation protein Calothrix sp. PCC 6303 9.9 40 YP_007137552.1 + 119-1 290 Ricin B-like lectin EULS3 A. lyrata subsp. lyrata 4E-36 73 XP_002862306.2 + 119-2 190 Spl1-Related 2 protein A. thaliana 2E-18 76 CAB56773.1 + 128-1 286 Chloroplast elongation factor TuB (EF-TuB) Nicotiana sylvestris 7E-12 90 BAA01975.1 + 94-2 242 Clone 6F8 eukaryotic initiation factor 4A-14 gene Nicotiana benthamiana 2E-47 89 JN688263.1 + 34-2 343 Transport protein SEC31 T. cacao 2E-06 81 EOY23302.1 + 91-1 199 Phosphatase 2C family protein isoform 2 T. cacao 2E-83 87 EOY06499.1 + 160-6 271 Kinase superfamily protein isoform 1 T. cacao 3E-34 78 EOY06443.1 0 209-3 337 Ser/Thr phosphatase-containing Kelch repeat domain protein, partial N. benthamiana 1E-50 90 AFN44702.1 0 220-3 208 Pentatricopeptide repeat (PPR) superfamily protein isoform 2 T. cacao 5E -08 51 EOY04957.1 2.25 ± 0.22 197-1 398 Ubiquitin-specific protease family C19-related protein A. thaliana 9E-68 79 NP_564009.1 + 87-2 220 Tetratricopeptide repeat (TPR)-like superfamily protein isoform 1 T. cacao 3E-27 86 EOY33236.1 2.55 ± 0.06 209-5 319 Serine/threonine-protein phosphatase, partial Genlisea aurea 3E-52 88 EPS64063.1 0 194-1 474 Proteasome subunit beta type-4 Solanum nigrum 6E-73 91 ADW66147.1 0 150-9 199 Serine/threonine-protein kinase AtPK2/AtPK19 R. communis 2.8 51 XP_002528702.1 + 123-4 246 IFA binding protein Lilium longiflorum 2E-37 74 ABM68547.1 0 123-3 302 Dual specificity kinase 1 Arabidopsis thaliana 3E-27 93 AEE27635 0 26-2 324 Dephospho-CoA kinase A. thaliana 1E-43 69 NP_180318.1 0 14-3 222 Translation initiation factor 4A2 Z. mays 7E-37 97 AAD20980.1 0.65 ± 0.01 244-1 302 Translation elongation factor, partial Ammopiptanthus mongolicus 1E-05 88 AFC01200.1 + 246-2 165 Eukaryotic translation initiation factor 5
isoform 2T. cacao 3E-05 52 EOX90767.1 0 Lipid metabolism 123-1 278 Patellin-5 A. lyrata subsp. lyrata 3E-13 67 XP_002872438.1 0 141-4 209 Patellin-5 Triticum urartu 8E-27 71 EMS62735
XP_0036230.27 ± 0.02 195-3 347 Non-specific lipid-transfer protein M. truncatula 1E-05 92 596.3 + 197-3 327 Pleckstrin (PH) and lipid-binding START domains-containing protein isoform 2 T. cacao 1E-47 78 EOY34386.1 + 125-2 278 Glyoxylate/hydroxypyruvate reductase
A HPR2A. lyrata subsp. lyrata 1E-29 70 XP_002889322.1 + Stress response and defense 48-1 263 Trehalose 6-phosphate synthase Nicotiana tabacum 3E-18 91 BAI99252.1 + 220-2 208 Transcription factor bHLH130 M. truncatula 4E-22 75 XP_003590427.1 1.41 ± 0.06 123-2 189 Cytochrome b5 N. tabacum 9E-26 80 CAA50575 + 48-3 268 Cytochrome P450 Citrus sinensis 2E-28 95 AAL24049.1 + 64-1 297 Cytochrome P450 A. thaliana 1E-23 81 NP_176086.1 + 221-2 451 Cytochrome oxidase subunit 1 Curcuma longa 1E-10 56 ABY83898.1 + 218-2 341 DNA damage-binding protein, partial M. truncatula 3E-64 67 XP003638090.1 0 252-1 353 Heat shock protein M. truncatula 6E-07 45 XP_003621962.1 0.36 ± 0.04 115-2 290 Stress responsive protein Z. mays 5E-32 64 NP_001149550.1 + 5-1 230 Senescence-associated protein Picea abies 2E-45 96 ACA04850.1 + 154-5 184 Dehydration-induced 19-like protein Gossypium hirsutum 8E-05 56 ADP30960.1 + 27-1 271 B-box zinc finger protein Bambusa oldhamii 7E-09 57 ACF35275.1 0 244-6 248 Dicer-like protein 4 A. thaliana 9E-09 63 NP_197532.3 + Cell metabolism 252-3 355 Ca2+-transporting ATPase A. thaliana 2E-04 72 NP_195479 + 130-1 240 Plasma membrane isoform 4 T. cacao 3E-29 83 EOY10146.1 0 244-2 287 Cinnamyl-alcohol dehydrogenase A. thaliana 3E-05 91 AAA99511.1 + 40-1 163 Clathrin adaptor complexes medium subunit family protein A. lyrata subsp. lyrata 3E-22 93 XP_002886592.1 0 154-3 429 RAB GTPase homolog A5A A. thaliana 1E-17 90 NP_199563.1 0.67 ± 0.01 217-6 323 Receptor-like kinase binding protein P. trichocarpa 4E-28 58 XP_002325092.1 0.87 ± 0.05 147-3 168 Calreticulin-like protein Solanum melongena 6E-22 88 BAA85118.1 3.35 ± 0.12 16-3 238 Ycf2 (chloroplast) Gossypium raimondii 3E-37 96 YP_005087735.1 + 143-4 350 Tetraspanin8 T. cacao 9E-24 69 EOY31574.1 + 216-3 443 Cell wall-associated hydrolase Vibrio cholerae 5E-39 76 ACX81677.1 0 200-2 283 Nucleic acid binding protein Z. mays 5E-43 89 NP_001152488.1 + Unknown biological processes 217-3 323 Hypothetical protein AT5G07270 A. thaliana 4E-09 59 NP_196344 0 236-1 256 Choline/ethanolamine kinase, putative Ricinus communis 4E-35 83 XP_002525542.1 + 236-2 322 Uncharacterized protein LOC8268581 R. communis 3E-31 82 XP_002530954.1 0 249-2 196 Amino acid transporter, putative R. communis 3E-03 39 XP_002531860.1 + 8-2 168 Predicted: monoacylglycerol lipase abhd6-B-like Fragaria vesca subsp. vesca 1E-18 80 XP_004303453.1 0 119-4 310 Domain of uncharacterized protein function 724 6, putative isoform 1 T. cacao 2E-22 68 EOX95351.1 0 57-1 205 Predicted: Vitis vinifera peroxidase 3-like mRNA Vitis vinifera 2E-21 91 XM_002280238.4 + 100-1 318 Hypothetical protein CICLE_v10006049mg Citrus clementina 6E-14 100 ESR32793.1 0 17-3 206 Hypothetical protein MTR_2g077840 M. truncatula 2E-3 76 XP_003596462.1 0 197-2 335 Mitochondrial protein, putative M. truncatula 3E-04 88 XP_003588355.1 0 217-4 261 Putative ATP synthetase alpha chain Oryza sativa subsp. japonica 3E-13 63 AAO72570.1 0 154-1 202 Hypothetical protein CICLE_v10022616mg Citrus clementina 7E-22 95 ESR54213.1 0 89-3 215 Hypothetical protein CICLE_v10033239mg C. clementina 3E-19 100 ESR51519.1 + TDFs: Transcript-derived fragments. Results are shown as the mean ± SD of at least three independent experiments. Fold change: 0 indicates TDFs only detected in normal juice sacs; + indicates TDFs only detected in granulated juice sacs. Relative expression ratio was obtained by analyzing gel images using PDQuest version 8.0.1 (Bio-Rad, Hercules, CA, USA). 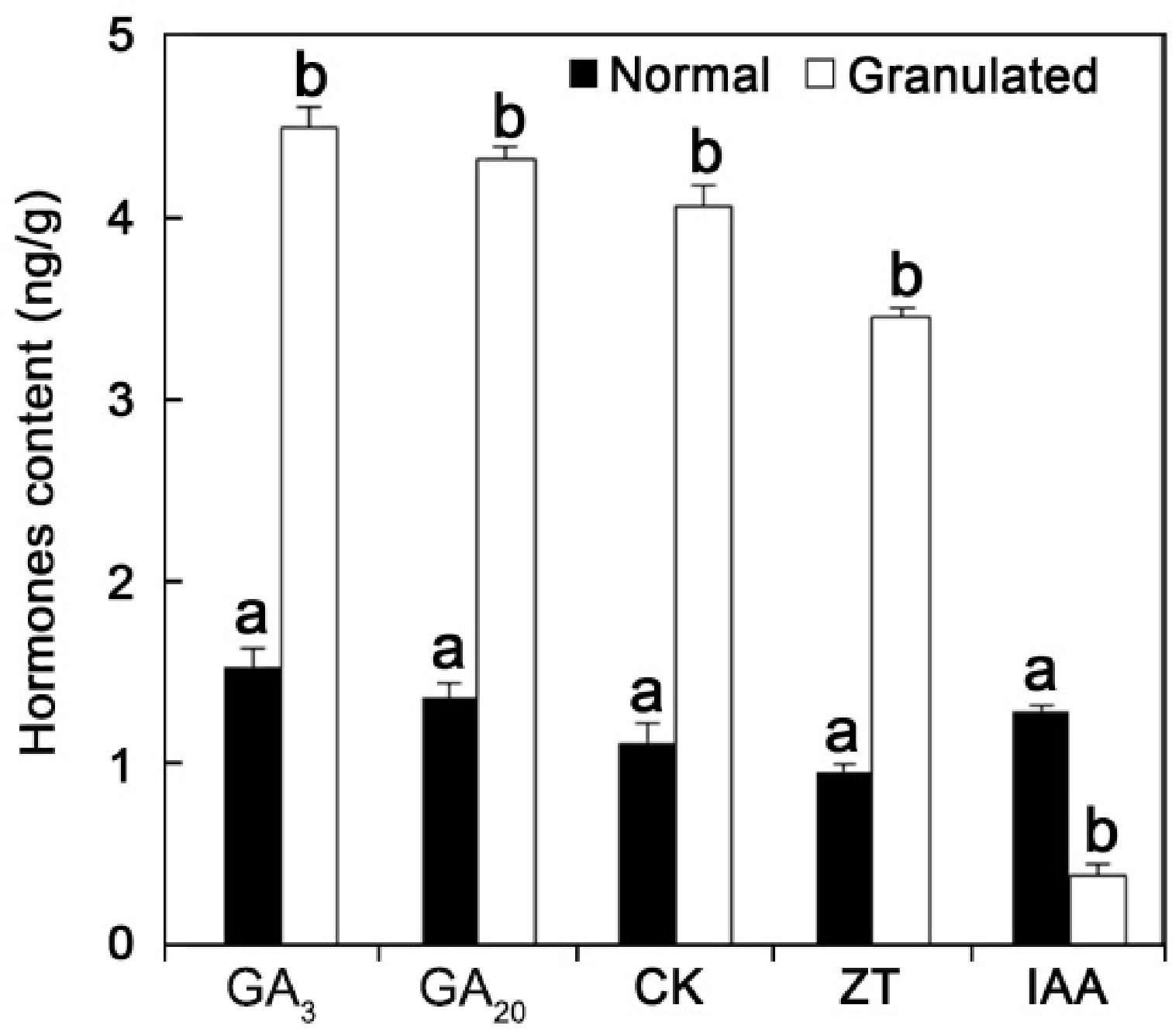
Figure 4.
Granulation led to alterations in hormone content in C. grandis juice sacs. The content of five hormones were measured by ultra-performance liquid chromatography mass-spectrometry (UPLC-MS). Normal juice sacs were considered as negative control. GA3: gibberellin 3; GA20: gibberellin 20, CK: cytokinins; ZT: zeatin; and IAA: indole-3-acetic acid. Error bars represent standard deviations calculated from three biological replicates. Different letters above bars indicate significant differences at P < 0.05.
-
Plant hormones are involved in the growth, development, ripening, and senescence of fruits. As an important regulator, hormones play a very critical role in the regulation of physiological disorders, defense, and stress responses, among other processes[17, 18]. Herein, using cDNA-AFLP technology, four hormone-related genes were found to be involved in the incidence of granulation in pummelo. qRT-PCR (Fig. 3) and UPLC-MS (Fig. 4) analyses further confirmed that the occurrence of granulation might induce changes in the hormone level in pummelo. Higher levels of GA3, GA20, CK, and ZT found in C. grandis normal juice sacs might induce increased cell division rate, and lead to granulation, whereas IAA might alter physiology of juice sacs. Taken together, these results indicated that alterations in hormone contents in C. grandis juice sacs might determine the occurrence of granulation. Our findings provide useful information about the mechanisms underlying the granulation phenomenon in C. grandis juice sacs.
As shown in Table 2, four TDFs (i.e., TDFs #48-2, 103-1, 115-1, and 195-1) in normal juice sacs and four TDFs (i.e., TDFs #6-2, 118-3, 219-5, and 195-6) in granulated juice sacs related to nutrients transformation and were identified by cDNA-AFLP. CAAX (Carboxyl-terminal three amino acids) protein is involved in the regulation of Rce1 (Ras converting enzyme) activity in cell signaling processes[19]. Nitrate reductase plays a central role in plant nitrogen acquisition by controlling nitric oxide levels[20]. Changes in the expression of genes coding for CAAX amino terminal protease (TDF #48-2) and nitrate reductase (TDF #103-1) in granulated juice sacs might be related to disrupted nitrogen absorption and utilization.
Carbohydrate and energy metabolism
-
In total, 17 differentially expressed TDFs related to carbohydrate and energy metabolism were found in pummelo juice sacs, among which five TDFs (TDFs #6-1, 65-1, 246-3, 246-4, and 118-2) were found exclusively in normal juice sacs and 12 TDFs (TDFs #221-3, 249-1, 249-3, 5-4, 118-1, 64-2, 244-2, 141-3, 160-1, 200-1, 20-3, and 134-1) were identified exclusively in granulated juice sacs (Table 2, Fig. 3). ATP synthase plays a key role in the cell by providing energy for ATP synthesis[21, 22]. In granulated juice sacs, the gene coding for ATP synthase subunit beta was upregulated, therefore energy levels are likely to be increased in granulated juice sacs. Deposition of both lignin and cellulose accompanied by juice sac granulation is widespread in harvested citrus fruit[23]. This hypothesis is further supported by the observation that ATP-binding protein-like (TDF #134-1), cellulose synthase (TDF #249-3), UDP-glucosyltransferase protein (TDF #249-1), glycosyltransferase, CAZy protein (TDF #118-1), 1, 4-alpha-glucan-maltohydrolase (TDF #20-3), β-amylase 4 (TDF #64-2), and galactose-1-phosphate guanylyltransferases (TDF #200-1) were upregulated in granulated juice sacs (Table 2, Fig. 3). Cellulose synthase belongs to the glycosyl hydrolase family which comprise enzymes that degrade complex sugars into mono- and disaccharides (glucose and cellobiose)[24]. Amylases hydrolyze starch and glycogen, and β-amylase specifically degrades amylose into maltose[25]. Researchers recently found that complex networks of pectin might be promoted by the granulation process[26]. Taken together, it is likely that major metabolic pathways related to ATP synthesis are activated in granulated juice sacs to produce more energy to meet the high demand of stressed juice sacs.
However, the observed higher mRNA levels of fructose-bisphosphate aldolase (TDF #6-1), glyceraldehyde-3-phosphate dehydrogenase (TDF #118-2), ADP-ribosylation factor 3 (TDF #246-4), and methylenetetrahydrofolate reductase protein gene (TDF #65-1) might enable higher tolerance to stressful conditions in granulation of juice sacs.
Protein and nucleic acid metabolism
-
Plants have evolved various sophisticated mechanisms for adapting to hostile environments during growth and development. Abiotic stresses demonstrably affect protein and nucleic acid metabolism in plants[27]. Studies with mutants in genes related nucleic acid metabolism revealed that nucleic acid processing, decay, and stability play a significant role in regulating gene expression at a post-transcriptional level in response to abiotic stresses in plants[28]. In plants, transcription and translation are the key steps for fine-tuning gene expression. In particular, during protein metabolism, modulation of global transcription and translation rates allows control over the production of specific proteins[29]. Differentially expressed TDFs found exclusively in granulated juice sacs included nuclear transcription factor Y subunit B18 (TDFs #15-2), transcription regulator (TDFs #215-1), transcription initiation factors TFIID (TDFs #217-2), 4A-14 (TDFs #94-2), chloroplast elongation factor TuB (TDFs #128-1), and translation elongation factor (TDFs #244-1) (Table 2), which might be related to nucleic acid accumulation. Moreover, differentially expressed mitochondrial substrate carrier family protein (TDFs #127-1), importin beta-2 subunit protein (TDFs #247-1), and transport protein SEC31 (TDFs #34-2) in granulated juice sacs suggest that protein transport might be impaired (Table 2, Fig. 3), which strengthens the hypothesis of nucleic acid accumulation in granulated juice sacs. Interestingly, all differentially expressed TDFs found exclusively in granulated juice sacs [i.e., ribonuclease II family protein (TDFs #154-4), 60S ribosomal protein L24-1 (TDFs #21-2), eukaryotic initiation factor (TDFs #94-2), translation elongation factor (TDFs #244-1), eukaryotic translation initiation factor (TDFs #246-2), and chloroplast elongation factor (TDFs #128-1)] (Table 2) could be associated with protein translation, which further indicates that protein translation might be impaired in granulated juice sacs.
Similarly, differentially expressed TDFs [i.e., phosphatase 2C family protein (TDFs #91-1), ubiquitin-specific protease family C19-related protein (TDFs #197-1), serine/threonine-protein kinase AtPK2/AtPK19 (TDFs #150-9)] related to protein phosphorylation and ubiquitination were upregulated in granulated juice sacs (Table 2), indicating that protein degradation might be increased in granulated juice sacs. Therefore, nucleic acid accumulation and protein degradation might have accelerated granulation in C. grandis juice sacs. Collectively, these findings indicate that impaired nucleic acid and protein metabolism in C. grandis juice sacs can be associated with the granulation phenotype.
Stress responses
-
Cytochromes P450s and b5 play a key role in the response to biotic and abiotic stresses in plants. Chen et al.[30] found that loss of function of the cytochrome P450 gene CYP78B5 causes giant embryos in rice. Herein, expression levels of genes encoding cytochrome P450 (TDF #48-3, 64-1), b5 (TDF #123-2), and cytochrome oxidase subunit 1 (TDF #221-2) were increased in C. grandis granulated juice sacs (Table 2, Fig. 3), which is in agreement with findings of previous studies reporting that certain cytochrome P450 genes in Arabidopsis were upregulated during biotic stresses, i.e., drought, hormone, high salinity, mechanical wounding, low temperature, herbicide (paraquat), and heavy metal (CuSO4) stress[31]. Thus, differential expression of genes coding for cytochrome P450s, b5, and cytochrome oxidase in C. grandis juice sacs might indicate an adaptation to physiological disorders.
In plants, double-stranded RNA (dsRNA) is recognized and cleaved by dsRNA-specific RNases named DCL (Dicer-like) enzymes, primarily by DCL4 and then by DCL2, producing 21- to 24-nucleotide double-stranded siRNA duplexes. Then, the antiviral silencing pathway is triggered by the presence of siRNAs, and 21-, 22-, or 24-nucleotide siRNA species mediate cleavage of mRNAs and DNA methylation in plants[32]. Expression of Dicer-like protein-coding genes might indicate that the plant's immune system was activated by biotic or abiotic stress response[33]. Herein, the gene encoding Dicer-like protein 4 was differentially and exclusively expressed in granulated juice sacs (TDF #244-6) (Table 2, Fig. 3), thus suggesting activating the immune defense system of C. grandis likely against granulation in juice sacs.
Plants under field conditions often encounter a variety of stresses, at times occurring simultaneously. Therefore, stress-responsive proteins are important effectors in plants during response to biotic or abiotic stresses[34, 35]. Under adverse conditions, many proteins have been previously found as differentially expressed in plants in response to bacterial, fungal, or viral infection, as well as to physiological disorders. Heat-shock proteins (HSPs) or the chaperone network are a major component of multiple stress-responses, and are controlled by diverse heat-shock factors which are recruited under stress conditions[34]. In the present study, differential expression of stress-responsive (TDF #115-2) and HSP (TDF #252-1) genes in granulated juice sacs may be related to a response against physiological disorders (Table 2, Fig. 3). Senescence is the final developmental stage of every plant organ, which eventually culminates in cell death. In granulated juice sacs, expression of the senescence-associated protein gene (TDF #5-1) might indicate that this altered physiological state is accompanied by accelerated senescence, dryness, hardness, and degeneration. Taken together, granulation activates stress-responsive metabolic pathways in C. grandis juice sacs, consequently increasing the expression of related genes.
Cell metabolism
-
In recent research, pectin methylesterase catalyzes the de-methylesterification of homogalacturonans and plays crucial roles in cell wall modification during plant development and fruit ripening[36]. The genes Ca2+-transporting ATPase (TDF #252-3), Ycf2 (chloroplast) (TDF #16-3), and tetraspanin 8 (TDF #143-4) (Table 2) involved in cell wall metabolism were specifically expressed in granulated juice sacs (Table 2, Fig. 3). In addition, mRNA levels of genes encoding plasma membrane isoform 4 protein (TDF #130-1), clathrin adaptor complexes medium subunit family protein (TDF #40-1), receptor-like kinase binding protein (TDF #217-6), and cell wall-associated hydrolase protein (TDF #216-3) were downregulated in granulated juice sacs. Therefore, cell wall formation or biosynthesis might be impaired in granulated juice sacs.
Conclusions
-
This work reports the first comparative investigation of normal and granulated juice sacs in pummelo (C. grandis) using the cDNA-AFLP technology. In total, 116 granulation-associated cDNA-AFLP products were identified in pummelo juice sacs. Differentially expressed TDFs were shown to be mainly involved in biological regulation and signal transduction, carbohydrate and energy metabolism, nucleic acid, protein metabolism, stress responses, and cell metabolism. Collectively, granulation in pummelo juice sacs seems to be associated with the following alterations: (1) changes in hormone levels; (2) activation of metabolic pathways related to ATP and sugar synthesis; (3) nucleic acid accumulation and increased protein degradation; (4) activation of stress-responsive metabolic pathways; (5) accelerated juice sac senescence (Fig. 5). Therefore, granulation is a complex process. The present study provides a comprehensive view into the differential responses occurring in granulated juice sacs, thus offering new insights into the adaptive mechanisms of 'Guanximiyou' pummelo (C. grandis) juice sacs at the transcriptional level during physiological distress.
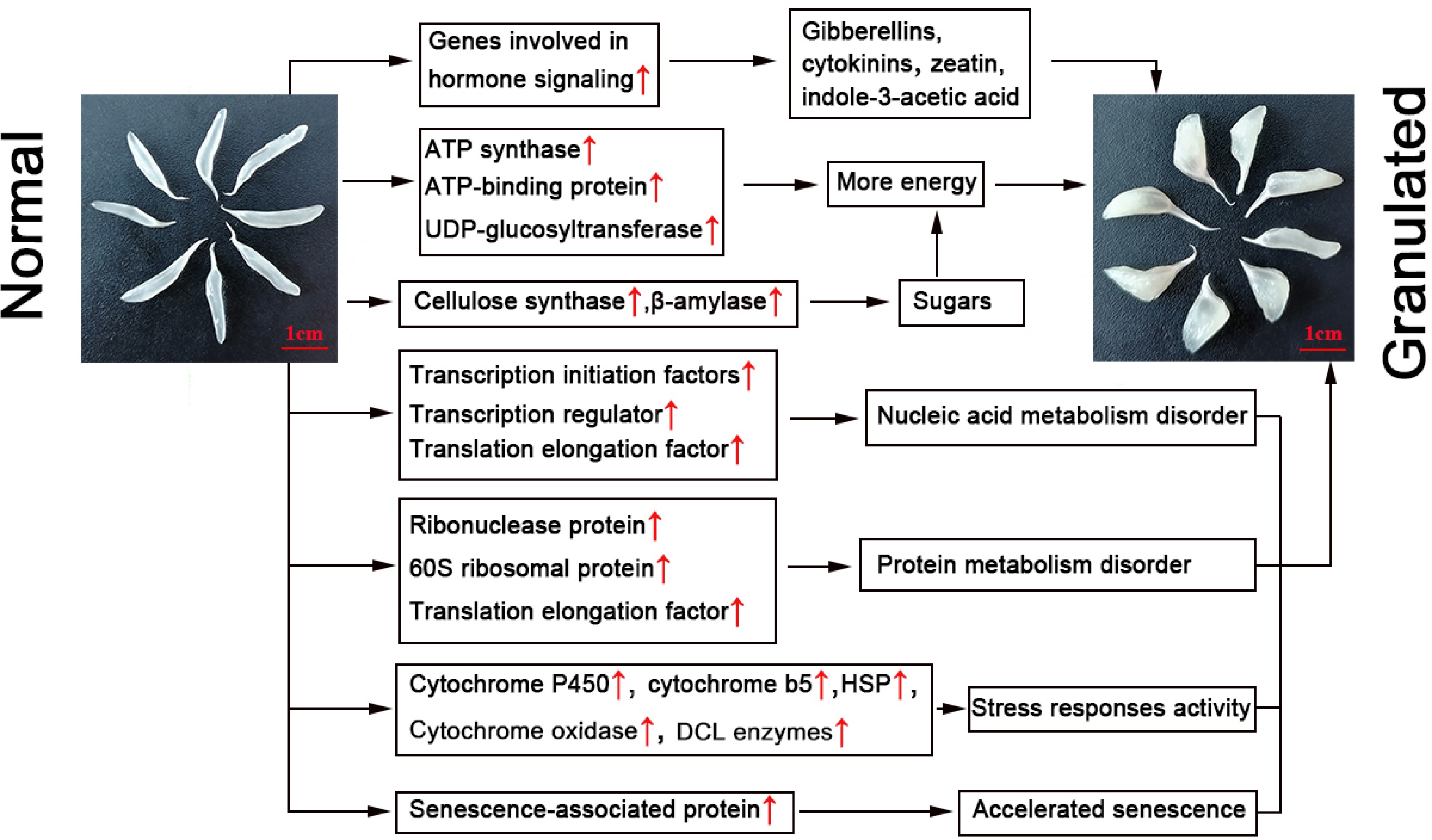
Figure 5.
Proposed regulatory network for the granulation phenomenon in C. grandis juice sacs. Red arrows indicate upregulated genes. HSP: heat-shock protein.
-
Pummelo (C. grandis) 'Guanximiyou' cultivar was used in this study. Fruits were collected from 25-year-old sour orange rootstocks in a pummelo orchard at grown at Yanban village pummelo orchard, Xiaoxi town, Pinghe county, Fujian province, China (E 24°35', N 117°31'), on single-tree replicates for all measurements on 1 October 2020. Fully mature pummelo fruits were harvested until granulation was visible. The degree of granulation was assessed according to the method of previous studies[3, 8, 14]. Normal and granulated juice sacs were collected from the same pummelo tree, a total of nine trees were sampled in the pummelo orchard. Five to ten fruit per tree were chosen from the outer of the mid-upper canopy. All the samples were immediately frozen in liquid nitrogen and stored at −80 °C until RNA isolation.
RNA extraction and cDNA synthesis
-
Normal and granulated juice sacs were ground in liquid nitrogen, and total RNA was independently isolated from samples using the RNeasy Plant Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Equal amounts of normal or granulated frozen juice sacs obtained from three pummelo units were mixed as one biological replicate, respectively. Each assay was repeated as three independent experiments, each with three biological replicates. Double-stranded cDNA synthesis was performed following the method proposed by Lu et al.[35].
cDNA-AFLP analysis
-
cDNA-AFLP analysis was performed according to the methods proposed by Xiao et al.[37]. Double-stranded cDNA was purified using an equal volume of phenol : chloroform : isoamyl alcohol (25:24:1, v/v/v). Subsequently, 500 ng of the resulting double-stranded cDNA was digested using the restriction enzymes EcoR I (10U; TaKaRa Biotechnology, China) at 37 °C for 3 h, and following Mse I (10U; TaKaRa) at 65 °C for 3 h. The resulting restricted products were ligated to AFLP adaptors (EcoR I: 5'-CTCGTAGACTGCGTACC-3', 5'-CATCTGACGCATGGTTAAP-3' and Mse I: 5'-GACGATGAGTCCTGAG-3', 5'-TACTCAGGACTCATP-3') with T4-DNA ligase (TaKaRa) and incubated overnight at 16 °C. Obtained products were pre-amplified with the corresponding pre-amplification primers: EcoR I: 5'- GACTGCGATCCAATTC-3' and Mse I: 5'-GATGAGTCCTGAGTAA-3'. A 100-fold dilution of pre-amplified products was used for the selective amplification using 256 combinations of the primers EcoR I 5'-GACTGCGATCCAATTC+MM-3' and Mse I 5'-GATGAGTCCTGAGTAA+NN-3', where MM and NN represent the following combination of nucleotides: AA, AT, AC, AG, GA, GC, GT, GG, CA, CT, CG, CC, TA, TC, TT, and TG. Final products were mixed with bromophenol blue and separated on 6% (w/v) polyacrylamide gel electrophoresis at 60 W for 3 h. Gels were silver stained to enable visualization of cDNA products. All samples in cDNA-AFLP analysis were submitted to electrophoresis at least three times independently.
Differential cDNA bands were excised, incubated in 100 µL of double-distilled H2O (ddH2O) for 10 min in a boiling water bath, then centrifuged at 10,000 rpm for 5 min. The supernatant used as template was re-amplified by PCR using the 256 combinations of the selective amplification primers. All positive amplicons were sequenced or ligated into the vector pMD18-T (TaKaRa) and further sequenced to confirm the identity of transcript-derived fragments (TDFs). Finally, differential cDNA sequences were analyzed using BLASTX and BLASTN searching engines (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ).Quantitative real-time RT-PCR (qRT-PCR)
-
qRT-PCR was performed with SYBR PrimeScript RT-PCR Kit (TaKaRa) according to the manufacturer's instructions. cDNA synthesis was performed with a mix of random primers and oligo(dT) primers provided in the kit using 500 ng of total RNA. β-actin gene served as an internal control. All qRT-PCR primers used are given in Supplemental Table S2. qPCR analysis was conducted in an ABI 7500 thermocycler (Applied Biosystems, Foster City, CA, USA). qPCR mixture consisted of 10 μL of 2× SYBR Premix Ex Taq DNA polymerase, 0.2 μL (200 nM) each of specific primer pairs, 2 μL of diluted reverse-transcribed cDNA, and 0.4 μL of ROX Dye II, in a 20 μL total reaction volume as per manufacturer's instructions. Quantification was conducted according to a previously described method[38]. Experiments were repeated at least three times independently using biological replicates.
Ultra-performance liquid chromatography mass-spectrometry (UPLC-MS) analysis
-
Hormone content in normal or granulated C. grandis juice sacs was analyzed using UPLC-MS. Approximately 100 mg of juice sac powder was weighed and transferred to a 1.5-mL centrifuge tube. Then, 500 μL of extracting solution (isopropyl alcohol : ddH2O : hydrochloric acid at a ratio of 2:1:0.002, v/v/v) and 50 μL of four standard samples were added, and tubes were slowly inverted to allow sufficient mixing at 4 °C for 30 min. Subsequently, 1,000 μL of trichloromethane was added to the mixture, and tubes were incubated at 4 °C for 30 min, followed by centrifugation at 14,000 rpm for 5 min. Supernatants were transferred to new tubes and blow-dried with nitrogen. Dried samples were redissolved in 100 μL of MeOH : H2O (1:1, v/v), filtered through a 0.1-μm membrane, and transferred to sample vials for LC-MS analysis. UPLC separation was performed using a BEH C18 column (2.1 mm × 100 mm, 1.7 μm, Waters Corporation, USA) at a flow rate of 0.3 mL min−1. The experiments were performed three times independently with biological replicates.
Statistical analysis
-
All experiments were performed with at least three replicates. Statistical analysis of data were carried out by two-way analysis of variance (ANOVA) using SPSS version 17.0 (SPSS Inc., Chicago, Illinois, US) with storage time and coating as factors. Comparison of means was performed using Duncan's multiple range test. The value of P < 0.05 or P < 0.01 represented statistical significance.
This work was supported by the National Natural Science Foundation of China (NSFC, 32002022) and Modern Agro-Industry Technology Research System (CARS-26).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 AFLP adapter and primer sequences used in this study.
- Supplemental Table S2 Degenerate primers used in this study.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang X, Guo L, Zhou R, Liu Y, Hu H, et al. 2022. cDNA-AFLP analysis reveals altered gene expression profiles involved in juice sac granulation in pummelo (Citrus grandis). Fruit Research 2:16 doi: 10.48130/FruRes-2022-0016 

cDNA-AFLP analysis reveals altered gene expression profiles involved in juice sac
- Received: 27 April 2022
- Accepted: 08 July 2022
- Published online: 31 October 2022
Abstract: Citrus fruits produced in China are often affected by granulation. Granulation is an altered physiological state of citrus fruits occurring usually before harvest but whose underlying mechanisms remain elusive. In this study, cDNA-AFLP technology enabled the identification of 116 granulation-associated genes in pummelo (C. grandis) juice sacs. Differentially expressed transcript-derived fragments (TDFs) were shown to be mainly involved in biological regulation and signal transduction, carbohydrate and energy metabolism, nucleic acid, protein metabolism, stress responses, and cell metabolism. Therefore, granulation in pummelo juice sacs seems to involve the following alterations: (1) changes in hormone levels; (2) activation of metabolic pathways related to ATP and sugar synthesis to produce more energy; (3) nucleic acid accumulation and increased protein degradation; (4) activation of stress-responsive metabolic pathways; (5) accelerated juice sac senescence. Our findings provide an overview of differential responses occurring at the transcriptional level in granulated juice sacs, thus revealing new insights into the adaptive mechanisms underlying this altered physiological state in 'Guanximiyou' pummelo (C. grandis) juice sacs.
-
Key words:
- cDNA-AFLP /
- Citrus grandis /
- Granulation /
- Juice sacs /
- Pummelo