











-
Nitrogen is both the basis of metabolism and the primary determinant of growth and yield[1]. Nitrogen is a major component of nucleic acids, proteins, chlorophyll, and other substances, and is involved in photosynthesis, carbohydrate allocation, and root formation[2, 3]. Plant nitrogen metabolism can also regulate the antioxidant system[4]. Thus, nitrogen is clearly an essential nutrient for plant growth.
Soils include inorganic nitrogen in the form of ammonium and nitrate, and organic nitrogen as amino acids, peptides, and proteins, with inorganic nitrogen are more readily absorbed by plants[5, 6]. Ammonium is a major inorganic nitrogen source in most soils, its assimilation by plants requires less energy than nitrate, and plants prefer to take up ammonium when the external nitrogen concentration is low[7, 8].
Ammonium transporters (AMTs) belong to the Ammonium transporter/Methylammonium permease/Rhesus (AMT/MEP/Rh) gene family, members of which have been identified in plants, microorganisms, and animals, indicating that ammonium transporter proteins are widely distributed in living organisms[9]. Two major groups of ammonium transporter proteins have been identified in plants: the AMT1 and AMT2 subfamilies[10]. The plant AMT2 subfamily is distantly related to the plant AMT1 subfamily[11], and in Arabidopsis, AtAMT2 is likely to play a significant role in moving ammonium[12]. Additional members of these families have also been characterized in Arabidopsis. AtAMT1;3, AtAMT1;4 and AtAMT1;5 exhibit high affinity for ammonium[13, 14], and AtAMT1;2 exhibits a relatively low affinity for ammonium[15]. A plasma membrane
$\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ The transcriptional regulation of ammonium uptake and utilization is driven by a series of transcription factors. In rice, transcription factor Indeterminate domain 10 (OsIDD10) binds to a cis-element motif present in the promoter region of OsAMT1;2 to specifically activate expression. In Arabidipsis, transcription factor Long Hypocotyles 5 (HY5) negatively regulates the expression of AtAMT1;2, an orthologous gene of OsAMT1;2[17]. Another group of plant-specific transcription factors, DNA binding with one finger (OsDOF) transcription factors, positively regulate ammonium uptake, assimilation, and significantly increase amino acid content by regulating the transcript abundance of OsAMTs[18−21]. OsMYB55, a member of the R2R3-MYB gene family, plays a positive role in amino acid metabolism by promoting the expression of OsGS1;2 and related genes[22].
A membrane-localized basic helix-loop-helix (bHLH) transcriptional factor, Glycine max Symbiotic Ammonium Transporter 1 (GmSAT1), encodes a novel regulatory gene involved in ammonium uptake during soybean root tumor development[23]. GmSAT1 is involved in the regulation of nitrogen signaling regulatory networks related to nitrogen transport and metabolism[24]. GmSAT1 activates the transcription of plasma membrane
$\text {NH}^+_4 $ $\text {NH}^+_4 $ The growth and yield of plants are highly dependent on environmental nutrient factors, including nitrogen. However, in pursuit of unilateral high yield, excessive input of nitrogen fertilizer has led to reduced nitrogen efficiency and decreased fruit quality, leading to lower agricultural production efficiency[25]. The over application of ammonium fertilizer presents a significant burden to both soil and plants[26], therefore, investigating the mechanism of ammonium utilization is an important goal in plant production[27]. Additionally, study of the tight regulation of transcription factors on nitrogen uptake can enable genetic engineering strategies to improve nutrient uptake regulation in plants[28]. In this study, we identified an ammonium-responsive MdSAT1 gene in apple and found that MdSAT1 regulates the expression of genes related to ammonium uptake. MdSAT1 can also affect root conformation and root hair development, to ultimately promote nitrogen uptake. Overall, these findings provide insight into the mechanisms by which MdSAT1 controls ammonium uptake as well as plant growth and development.
-
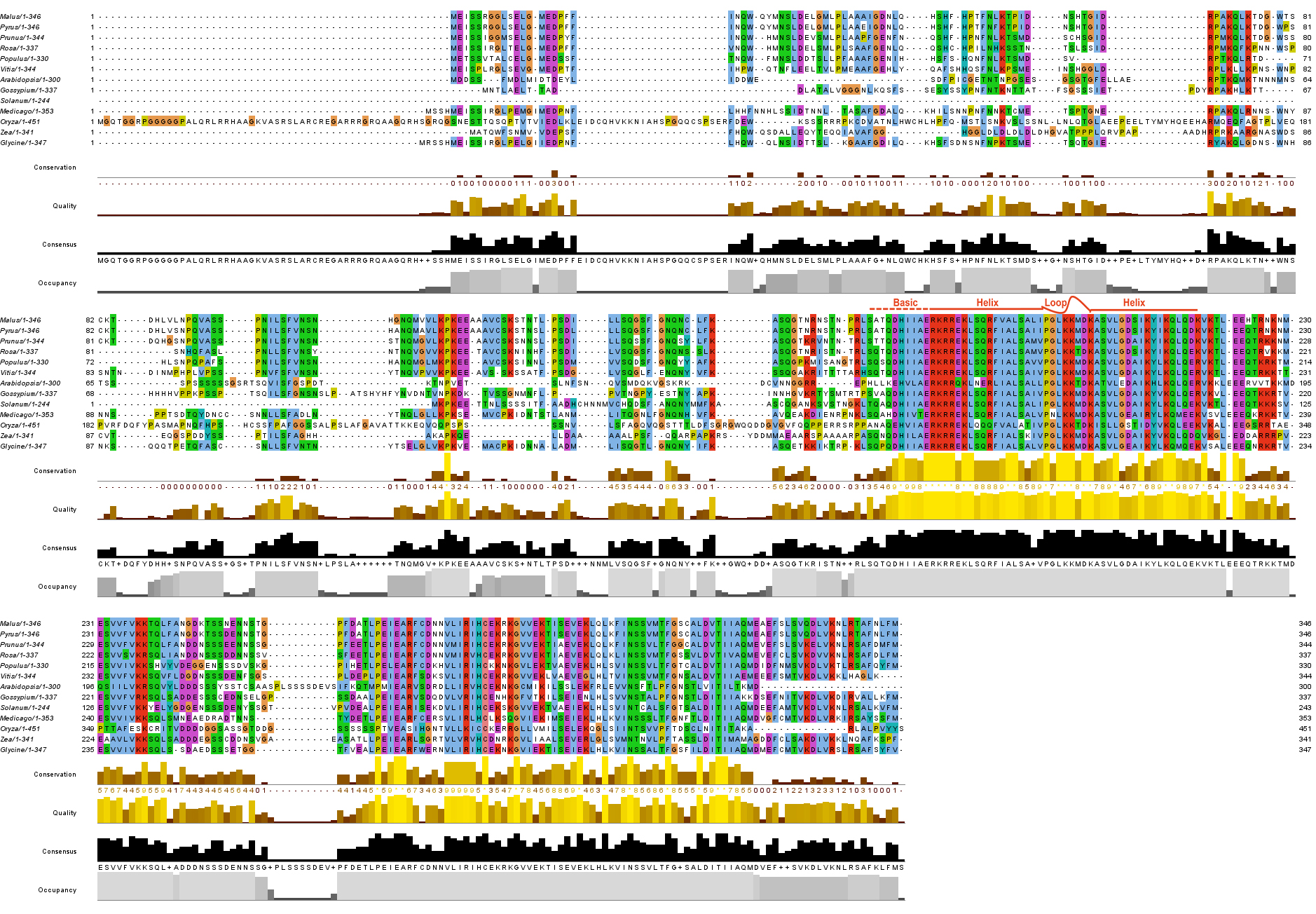
The MdSAT1 (MD10G1115500) gene was identified from the NCBI website according to the GmSAT1 sequence of soybean (Glycine max). A phylogenetic tree was constructed, and apple MdSAT1 was most closely related to pear PbSAT1 (Rosaceae) (Fig. 1a), indicating that these genes diverged recently in evolution. We compared the SAT1 protein sequences of apple with those of other plant species, and the results showed that all 13 proteins had high sequence similarity and belonged to the plant bHLH transcription factor superfamily, members of which contain a bHLH domain and an H-E-R DNA binding region (Fig. 1c; Supplemental Fig. S1). The bHLH structure domain of the MdSAT1 protein was predicted by the homology model, and the results indicated that the secondary and tertiary structures of bHLH structural domain match those of the core conserved domain (Fig. 1b, c).
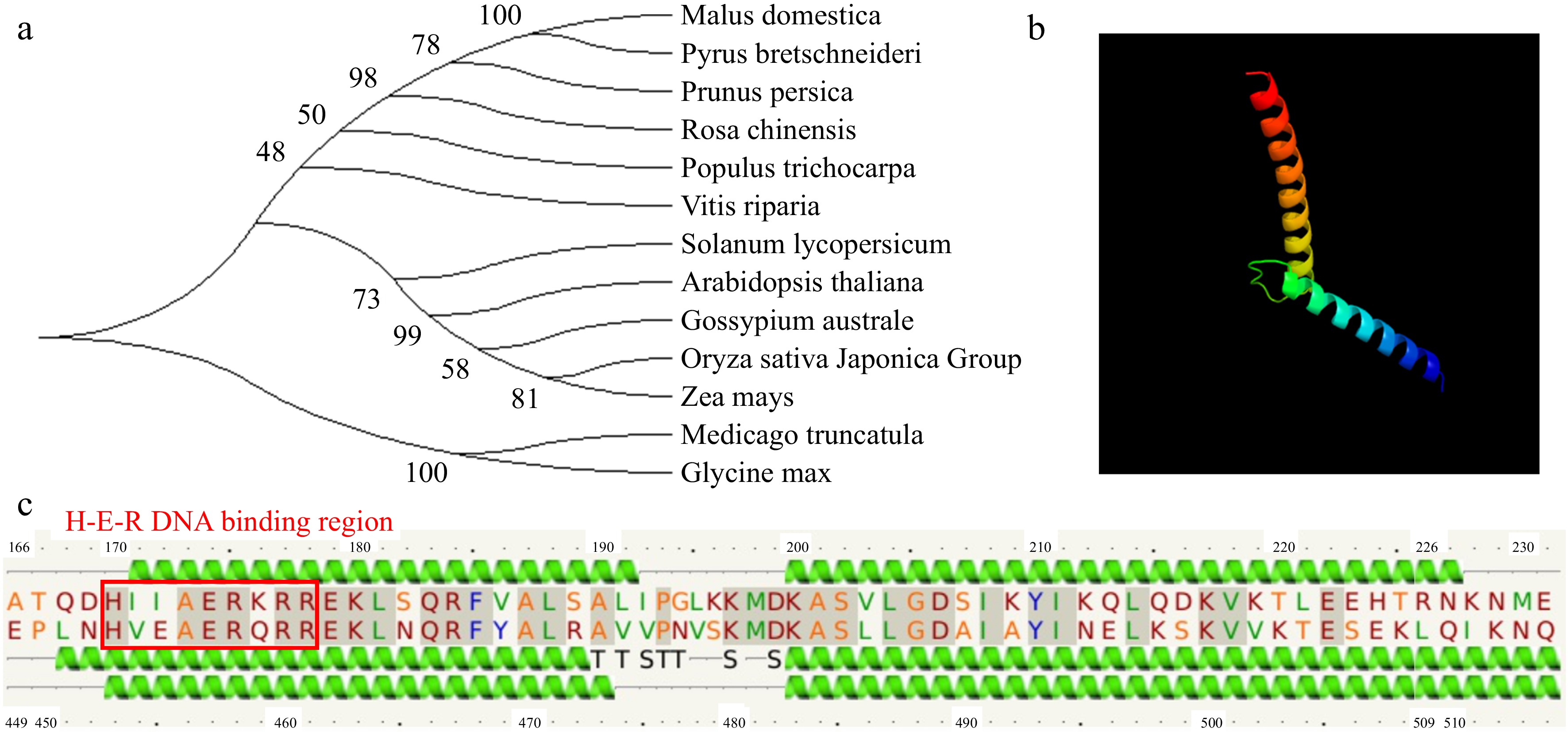
Figure 1.
Phylogenetic relationships, multiple sequence alignment, and protein structure of MdSAT1. (a) Phylogenetic tree of SAT1 sequences; the number on each branch represents the genetic distance. (b) Predicted bHLH 3D structure of MdSAT1 protein. (c) Predicted protein secondary of MdSAT1 bHLH structural domain.
MdSAT1 is an ammonium-responsive gene
-
MdSAT1 is homologous to GmSAT1, which is involved in ammonium uptake[23], and RT-PCR was used to detect the expression of MdSAT1 in response to different nitrogen forms (KCl, as a control, represents 0 N; KNO3, represents nitrate; and NH4Cl, represents ammonium). The expression of MdSAT1 was significantly induced by NH4Cl both in shoots and roots, however, the transcript level of MdSAT1 showed little change in response to nitrate (Fig. 2a, b), suggesting that MdSAT1 was specifically responsive to ammonium.
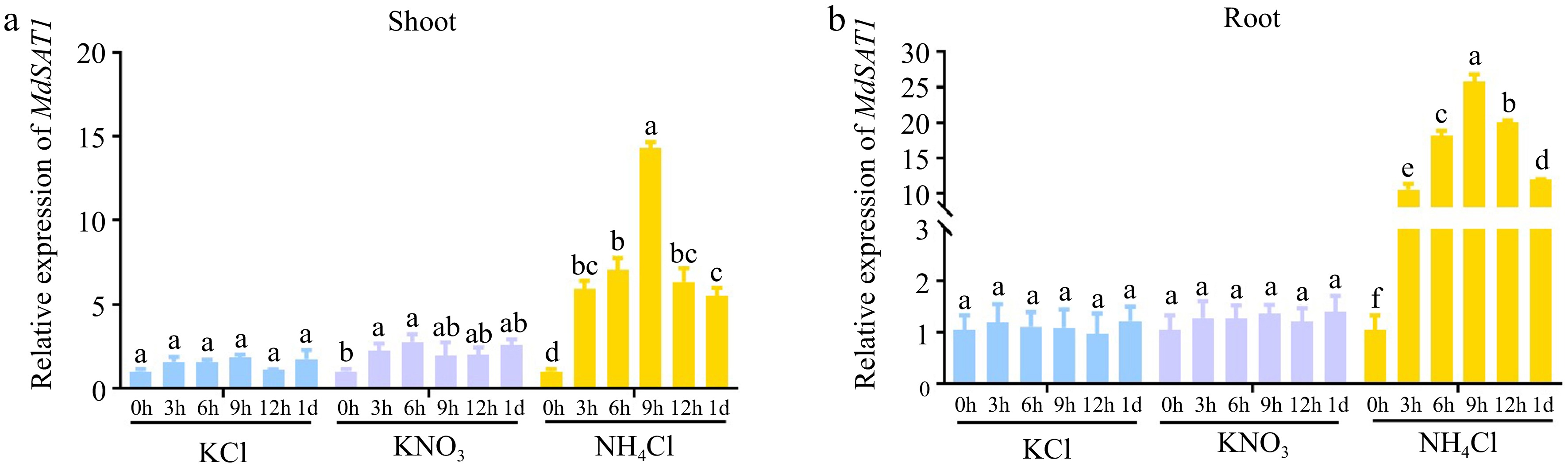
Figure 2.
Nitrogen response of MdSAT1. (a) Shoot and (b) root response of MdSAT1 to KCl (represents 0 N), KNO3 (represents nitrate), and NH4Cl (represents ammonium). Error bars represent standard deviation (n = 3). Different letters above the bars indicate significantly different values (P < 0.05).
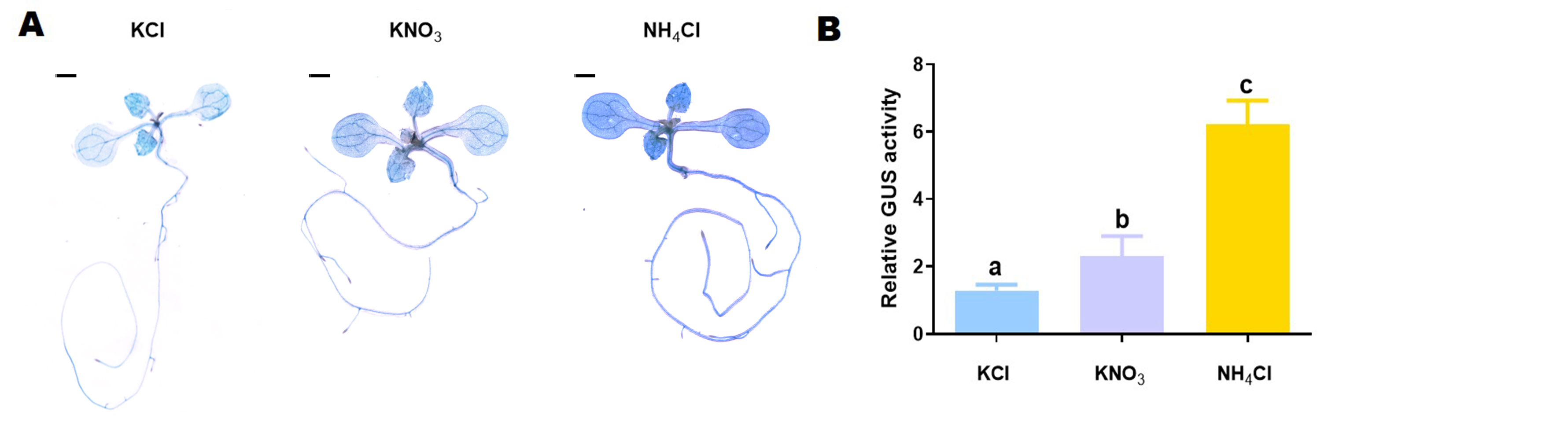
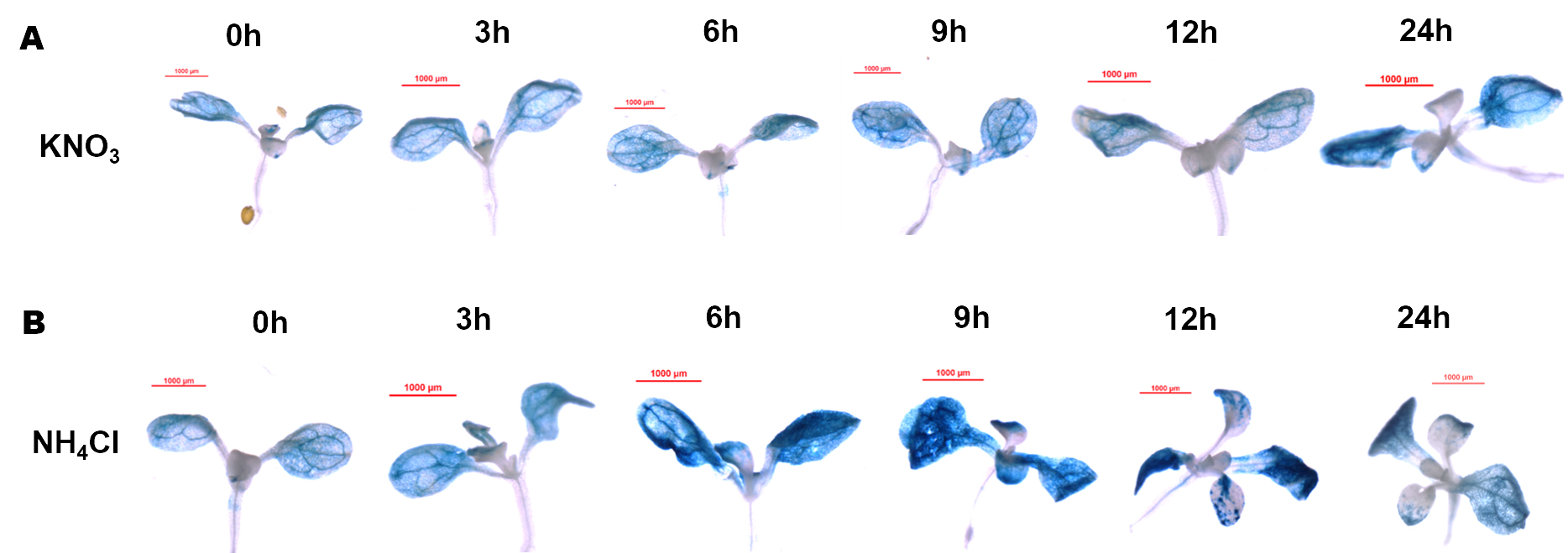
The ProMdSAT1::GUS transgenic Arabidopsis seedlings-that can be used to localize the expression location and expression intensity of MdSAT1-were treated with different forms of nitrogen. We found that GUS staining results suggested that the highest GUS activity was observed under NH4Cl treatment (Supplemental Fig. S2). With increasing time of different treatments, the expression activity of ProMdSAT1::GUS was specifically induced by NH4Cl (Supplemental Fig. S3). Taken together, these results suggest that MdSAT1 is specifically responsive to ammonium.
Overexpression of MdSAT1 regulates ammonium uptake
-
Given that MdSAT1 is an ammonium-responsive gene, we next treated MdSAT1-OE and wild type (Col) in a modified Hoagland's nutrient solution containing 0.5 mM NH4Cl (Low
$\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ 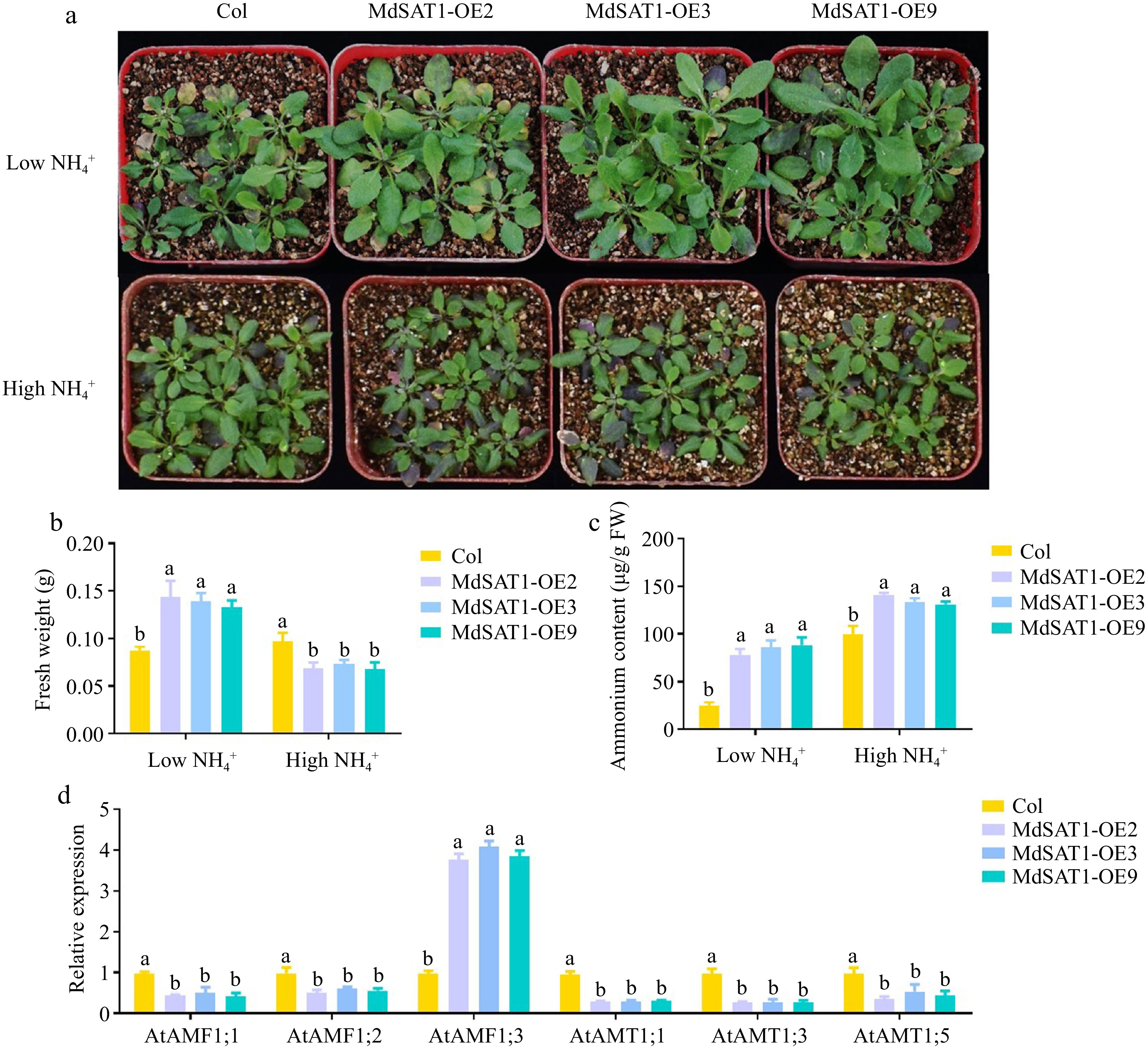
Figure 3.
MdSAT1 regulates ammonium uptake and plant growth MdSAT1-OE and Col plants grown for four weeks under low
$\text{NH}^+_4 $ (0.5 mM NH4Cl) or high$\text{NH}^+_4 $ (5 mM NH4Cl) conditions. (a) Morphological changes, (b) fresh weight, and (c) ammonium content are presented. (d) RT-PCR analysis of AtAMF1;1, AtAMF1;2, AtAMF1;3, AtAMT1;1, AtAMT1;3 and AtAMT1;5 expression in MdSAT1-OE and Col Arabidopsis. Error bars represent the standard deviation (n = 3). Different letters above the bars indicate significantly different values (P < 0.05).To further evaluate the role of MdSAT1 in ammonium uptake, the effects of MdSAT1 on the expression of genes related to ammonium uptake were analyzed. The result showed that transcript levels of AtAMTs were not increased in MdSAT1-OE lines, however, expression of AtAMF1;3 was significantly induced in the MdSAT1-OE lines (Fig. 3d). AMF proteins promote
$\text {NH}^+_4 $ Overexpression of MdSAT1 promotes lateral root development
-
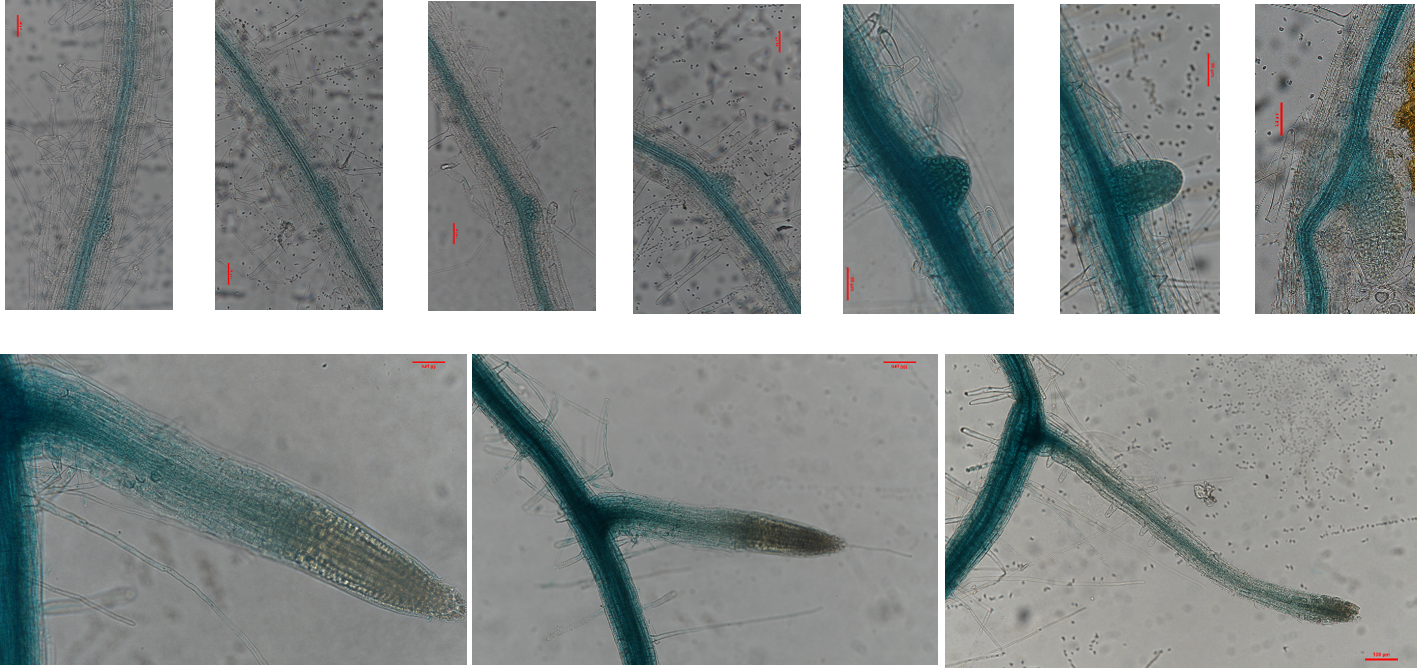
Ammonium in the soil is actively taken up by the roots, mainly by ammonium ion transporters[29]. The tissue-specific localization of MdSAT1 was detected using ProMdSAT1::GUS transgenic Arabidopsis. GUS staining results showed that MdSAT1 was differentially expressed during lateral root growth, with the highest expression observed at the time of lateral root primordium genesis (Supplemental Fig. S4). There was no significant difference of primary root length, but the lateral root numbers were significantly increased in the MdSAT1-OE lines compared with those of Col (Fig. 4a−d). These results suggest that MdSAT1 overexpression promotes lateral root growth and development.
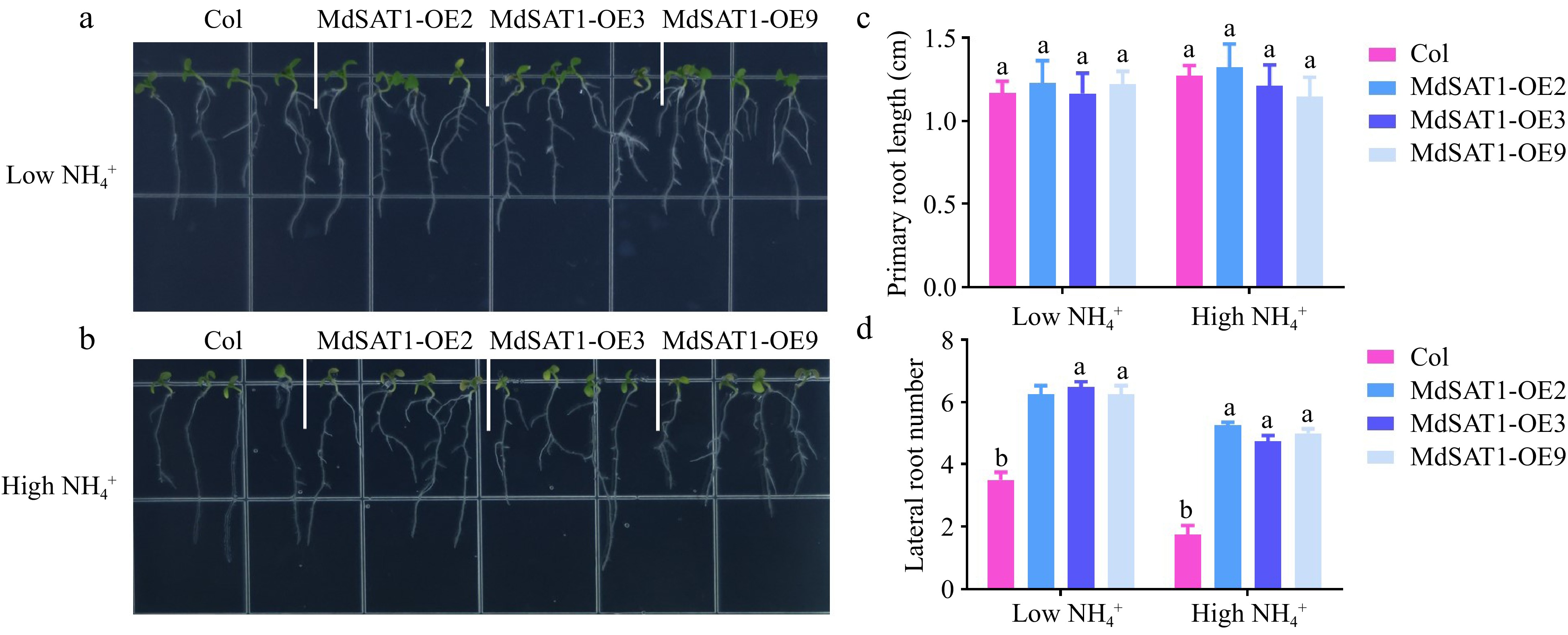
Figure 4.
MdSAT1 regulates root system conformation MdSAT1-OE and Col plants grown for seven days under low
$\text{NH}^+_4 $ (0.5 mM NH4Cl) or high$\text{NH}^+_4 $ (1.5 mM NH4Cl) conditions. (a), (b) Morphological changes, (c) primary root length and (d) lateral root number are presented. Error bars represent the standard deviation (n = 3). Different letters above the bars indicate significantly different values (P < 0.05).Overexpression of MdSAT1 regulates root hair growth and development
-
Root hairs play an important role in nutrient uptake[30], we next observed the phenotypes of root hairs in MdSAT1-OE and Col. MdSAT1 significantly increased the number and length of root hairs under low
$\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ 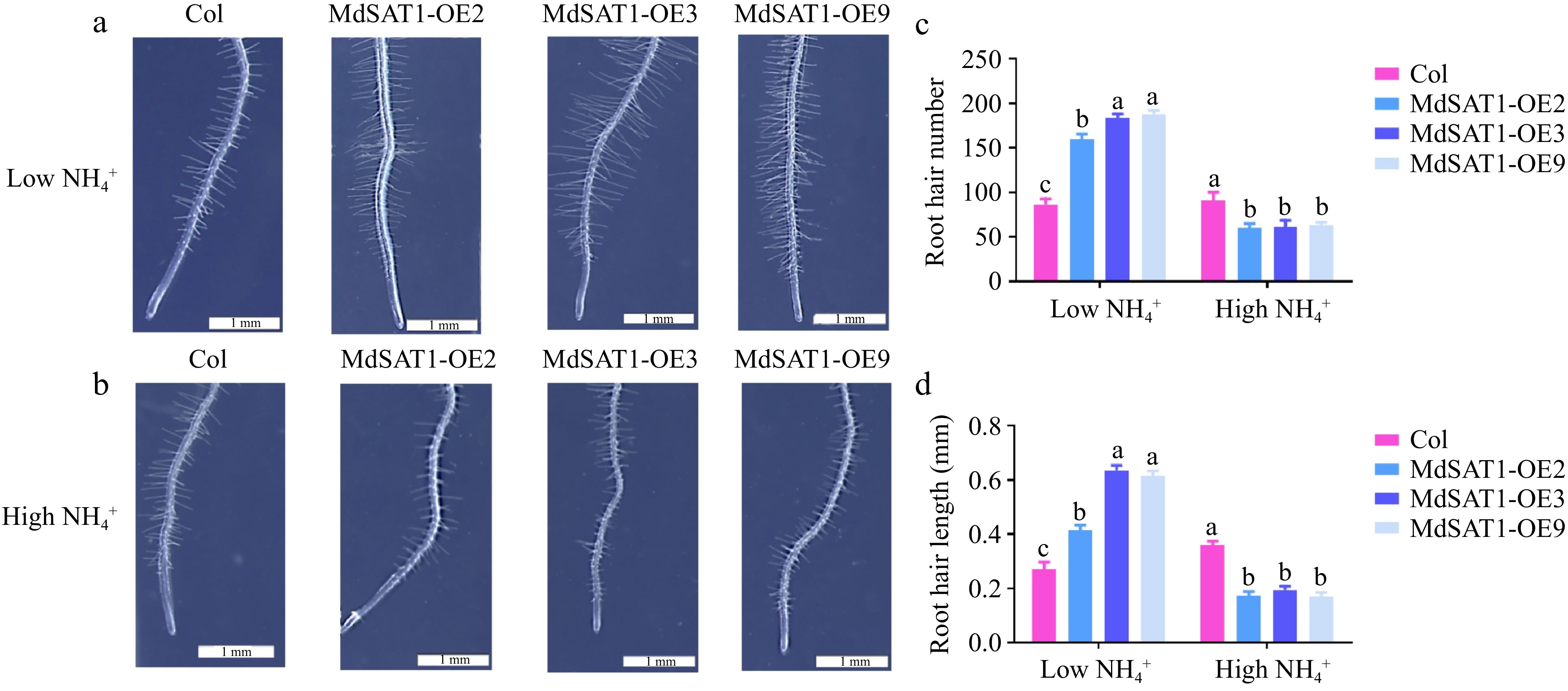
Figure 5.
MdSAT1 regulates root hair growth and development MdSAT1-OE and Col plants grown for three days under low
$\text{NH}^+_4 $ (0.5 mM NH4Cl) or high$\text{NH}^+_4 $ (1.5 mM NH4Cl) conditions. (a), (b) Morphological changes, (c) root hair number and (d) root hair length are presented. Error bars represent the standard deviation (n = 3). Different letters above the bars indicate significantly different values (P < 0.05).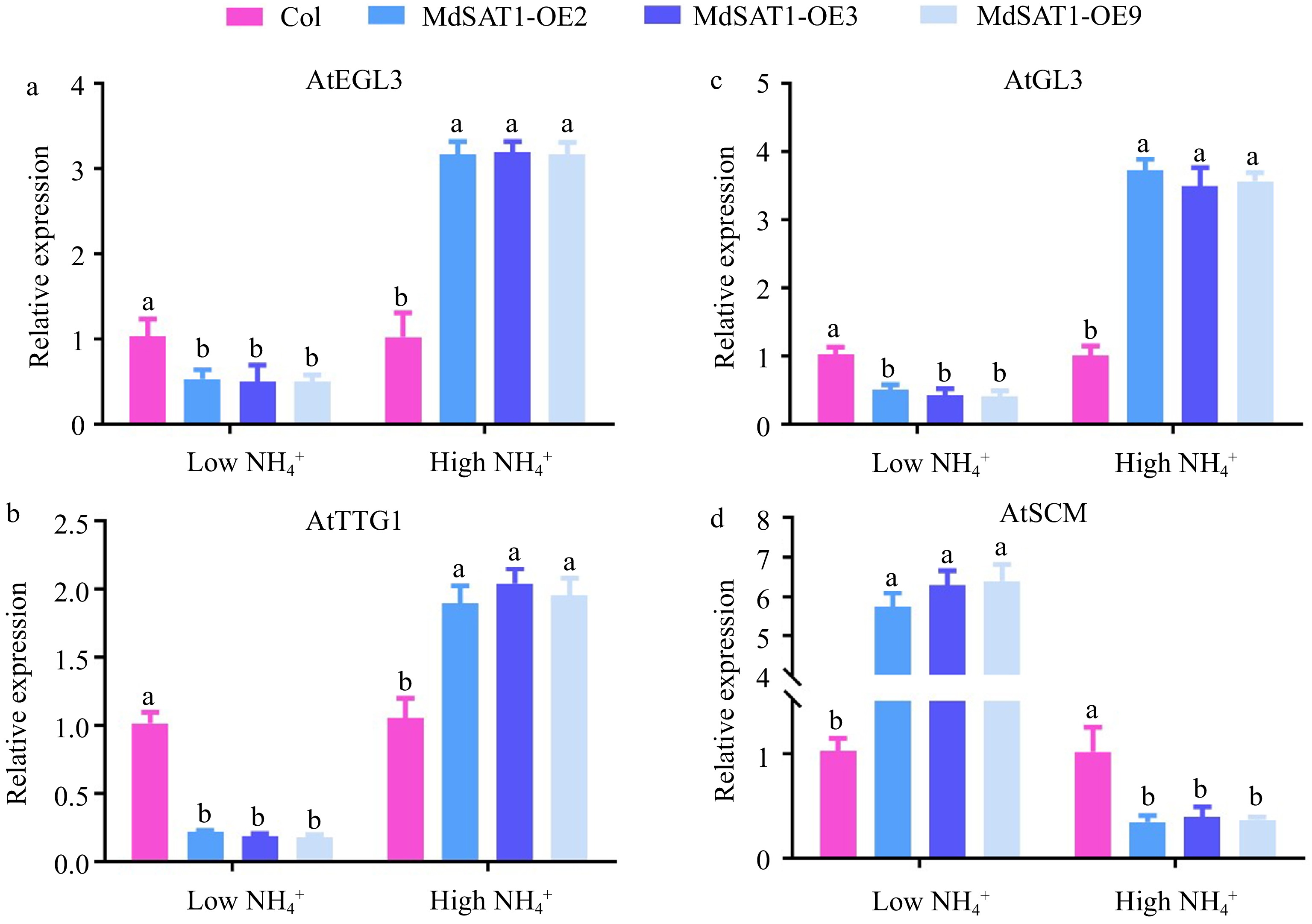
Figure 6.
MdSAT1 regulates the expression of genes related to root hair development Analysis of root hair development-related gene expression by RT-PCR under low
$\text{NH}^+_4 $ (0.5 mM NH4Cl) or high$\text{NH}^+_4 $ (1.5 mM NH4Cl) conditions: (a) AtEGL3, (b) AtTTG1, (c) AtGL3, and (d) AtSCM are presented. Error bars represent the standard deviation (n = 3). Different letters above the bars indicate significantly different values (P < 0.05).MdSAT1 promotes ROS accumulation
-
ROS plays a role in root hair development[34], so we next measured ROS content by NBT staining. The results showed that overexpression of MdSAT1 increased ROS accumulation of leaves compared with the level in Col (Fig. 7a). OFR content was promoted in the MdSAT1-OE lines, both in high and low
$\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ 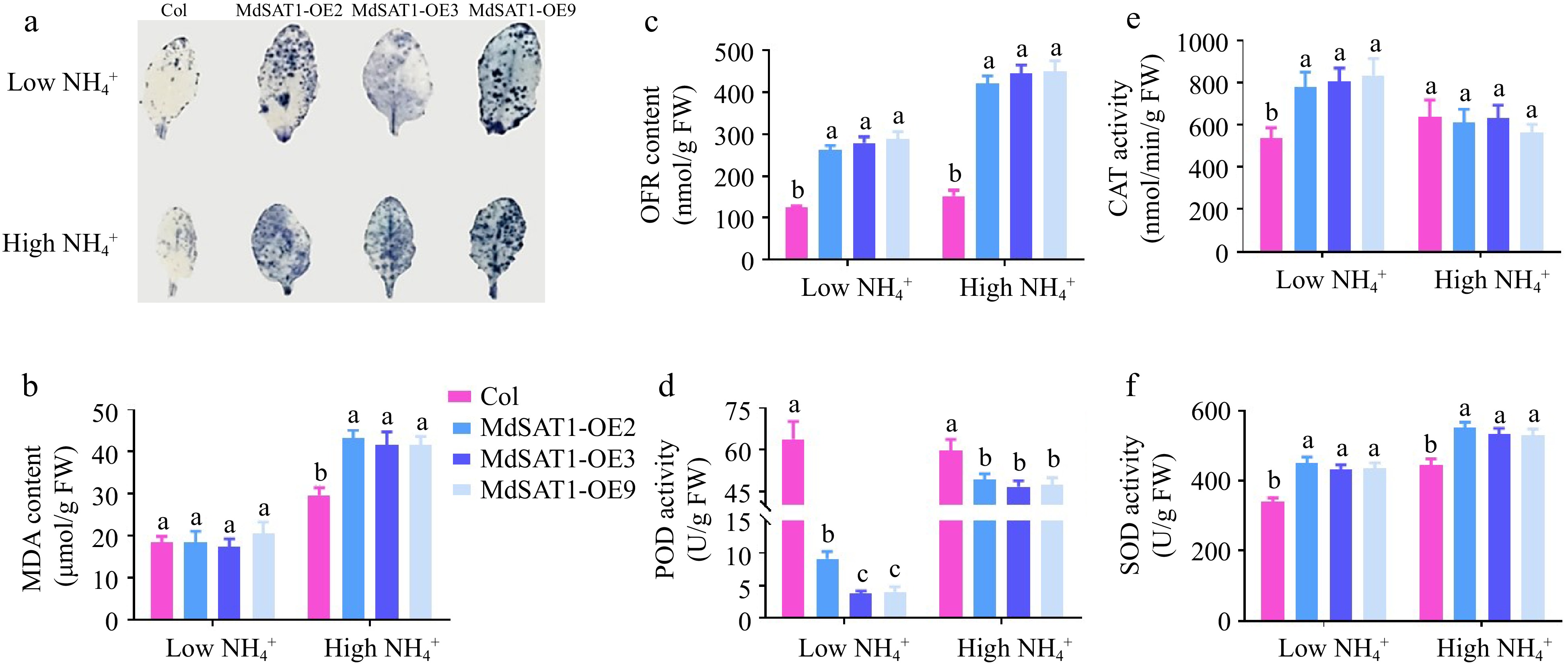
Figure 7.
MdSAT1 regulates ROS accumulation Col and MdSAT1-OE plants grown for four weeks under low
$\text{NH}^+_4 $ (0.5 mM NH4Cl) or high$\text{NH}^+_4 $ (5 mM NH4Cl) conditions. (a) NBT staining, (b) MDA content, (c) OFR concent, (d) POD activity, (e) CAT activity, and (f) SOD activity are presented. Error bars represent the standard deviation (n = 3). Different letters above the bars indicate significantly different values (P < 0.05). -
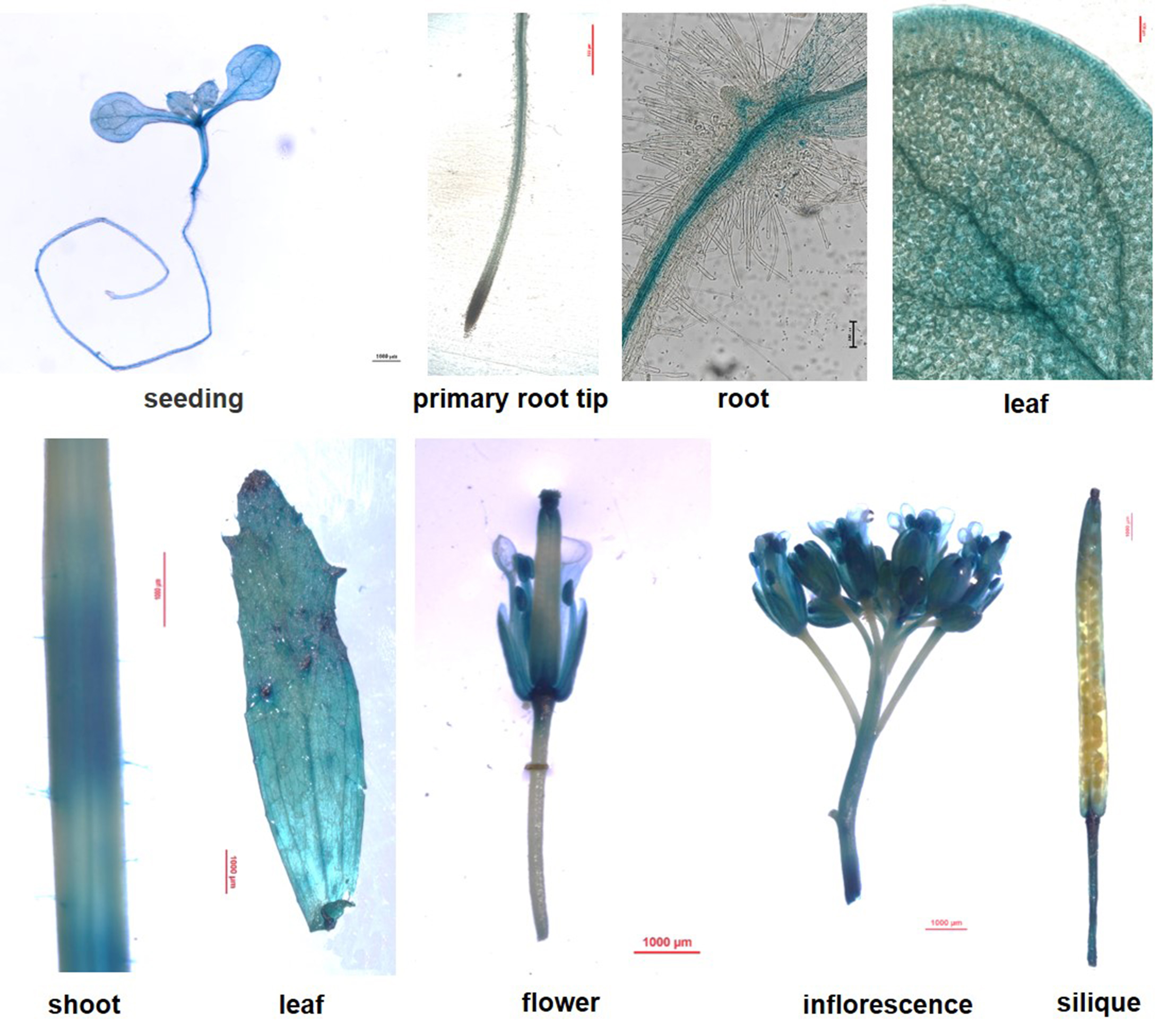
When the soil nitrogen concentration is low, plants are more likely to take up ammonium[37]. However, the absorption and utilization efficiency of nitrogen by wild-type plants (Col) is not ideal when only ammonium is applied as nitrogen fertilizer. Moreover, the growth of plants is significantly inhibited at low concentration of single ammonium fertilizer, and the use of a higher concentration of single ammonium fertilizer will cause ammonium toxicity to plants. These trends will guide the application of single ammonium as nitrogen fertilizer in agricultural production. Determination of the appropriate intermediate concentration of single ammonium for use as nitrogen fertilizer is essential to ensure the effective utilization of plants and also avoid the waste of resources and environmental pollution caused by excessive fertilization. Alternatively, combined application with nitrate nitrogen may be more appropriate. Therefore, it is important to study the law and mechanism of ammonium absorption by crops to optimize nitrogen utilization. GmSAT1 was functionally identified in soybean root nodule development[24]. In this study, phylogenetic and conserved domains analysis indicated that the MdSAT1 protein may be similar in function to GmSAT1 (Fig. 1; Supplemental Fig. S1). The function of MdSAT1 was characterized and the results showed that it is highly expressed, mainly in nutrient organs (Supplemental Fig. S5), and plays a key role in ammonium uptake and assimilation (Figs 3 & 4). MdSAT1 also regulates the accumulation of ROS and ultimately plant growth (Fig. 3a−c; Figs 4−7).
Several studies have shown that GmSAT1 is important for the symbiosis of soybean rhizobia and acts in
$\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ $\text {NH}^+_4 $ In conclusion, the results of this study showed that overexpression of MdSAT1 promotes plant growth and biomass accumulation. These findings provide theoretical guidance to resolve the mechanisms by which MdSAT1 regulates ammonium uptake and plant growth and provide a reference for future selection of superior germplasm with more efficient nitrogen uptake.
-
Apple seedlings (Malus domestica 'Royal Gala') were cultured in a plant growth chamber under 25/22 °C, 14/8 h temperature, and photoperiod. Apple group culture seedlings were grown in Murashige & Skoog (MS) medium (pH = 6.0) containing 6-Benzylamino Purine (6-BA, 0.5 mg L−1), Naphthaleneacetic Acid (NAA, 0.1 mg L−1), and Gibberellin (GA, 0.5 mg L−1) for succession every 30 d. For the nitrogen treatment experiment, 1 month-old apple seedlings were selected for rooting in ½ MS rooting medium containing 1 mg L−1 3-Indoleacetic acid (IAA). When rooting was completed, the seedlings were transferred to a nutrient bowl and cultured for about 30 d. Seedlings of uniform growth were selected and pre-treated in hydroponic conditions with ddH2O for 1 week. The seedlings were then treated with 2 mM KCl (represents 0 N), KNO3 (represents nitrate), or NH4Cl (represents ammonium), and sampled after 0, 3, 6, 9, 12, and 24 h of treatment.
Arabidopsis seeds were disinfected with 75% ethanol and 3% sodium hypochlorite, and then sown on ½ MS medium solid culture plates (15 g L−1 sucrose and 8.0 g L−1 agar powder, pH adjusted to 5.9 with 1.0 M sodium hydroxide). The plates were incubated at 4 °C with dark vernalization for 4 d. Seeds were germinated and grown at 22 °C with a 16 h/8 h light/dark cycle.
Different types of Arabidopsis (MdSAT1-OE, Col) seedlings used for gene expression analysis by RT-PCR were germinated on ½ MS medium solid culture plates for 7 d before being transplanted to vermiculite, irrigated with tap water, and then watered weekly with a modified Hoagland’s nutrient solution with either low
$\text{NH}^+_4 $ $\text{NH}^+_4 $ One or two days after germination, different types of Arabidopsis (MdSAT1-OE, Col) seedlings were transplanted and grown on low
$\text {NH}^+_4 $ $\text {NH}^+_4 $ Transgenic materials
-
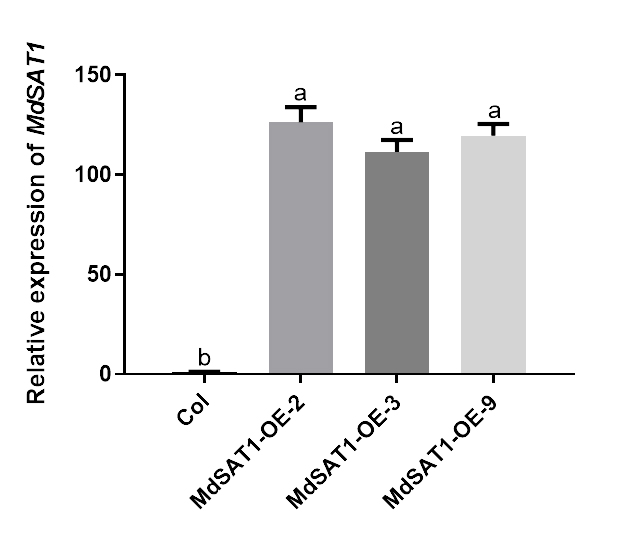
MdSAT1-OE (MD10G1115500) and ProMdSAT1::GUS Arabidopsis seeds were obtained as described[41]. We identified the transgenic plant material using M5 Exceed Lightspeed PCR mix (Mei5Bio, Beijing, China) and Omni Plant RNA Kit (tDNase I) (Tiangen, Beijing, China) for DNA and RNA extraction, respectively (Supplemental Fig. S6)[41].
Bioinformatics analysis
-
The protein sequences of SAT1 from different species were obtained using blastp at the NCBI website (
https://blast.ncbi.nlm.nih.gov/ ). The obtained sequences were used to construct a neighbor-joining phylogenetic tree with 1000 bootstrap replicates in MEGA-X[42] using the built-in ClustalW algorithm, Poisson model, and parameter settings for partial deletion (95%). The conserved domains of the SAT1 protein were predicted using Phyre2 (www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index )[43].Extraction of plant genomic RNA
-
Omni Plant RNA Kit (tDNase I) was used to extract plant RNA (Tiangen, Beijing, China). The extracted RNA was stored in tissue RNA preservation solution (RNAfollow M6100 New Cell & Molecular Biotech) for protection and stored in an ultra-low temperature refrigerator for gene expression analysis.
Real-time quantitative RT-PCR analysis of gene expression
-
From the extracted RNA, cDNA required for quantitative PCR was synthesized using the PrimeScript First Chain cDNA Synthesis Kit (Takara, Dalian, China). Reaction conditions: The RT-PCR experiments were conducted on ABl QuantStudio 3 a 96 Real-Time PCR system (Thermo Fisher, USA) using ChamQTM Universal SYBR qPCR Master Mix (Vazyme, Q711-02, Nanjing, China), using a 20-μL reaction solution.This solution was made of 10-μL of 2× ChamQ Universal SYBR qPCR Master Mix, 0.4-μL 10 uM of each primer, 2-μL 10-folddilutions of cDNA template, and DEPC-water. All the qPCR protocols were performed in triplicate on 96-well plates, along with blank (template-absent) control. The PCR program is: 95 °C, 30 s; 95 °C, 10 s, 60 °C, 30 s, 40 cycles. Apple 18S rRNA and Arabidopsis actin rRNA genes were used as controls. PCR analysis was performed using specific primer sequences designed using Primer3Plus (
http://primer3plus.com/cgi-bin/dev/primer3plus.cgi )[44] and listed in Supplemental Table S1. The RT-PCR analysis performed in triplicate, and relative gene expression was calculated using the 2−ΔΔCᴛ method.Physiological measurements
Determination of substance content
-
Ammonium, Oxygen-derived free radicals (OFR), and Malondialdehyde (MDA) levels were measured by UV spectrophotometry as described below.
Ammonium interacts with hypochlorite and phenol in a strong alkaline medium to produce the water-soluble dye indophenol blue. Indophenol blue has a characteristic absorption peak at 625 nm and the absorbance value is proportional to the ammonium nitrogen content.
OFR react with hydroxylamine hydrochloride to form
$\text {NO}^-_2 $ MDA condenses with thiobarbituric acid (TBA) to produce a red product with a maximum absorption peak at 532 nm that can be used to estimate the amount of lipid peroxide in the sample. The absorbance at 600 nm was also measured, and the difference between the absorbance at 532 nm and 600 nm was used to calculate the amount of MDA.
Determination of enzymatic activities
-
Using 0.1g of plant material, Peroxidase (POD), Catalase (CAT), and Superoxide dismutase (SOD) activities were measured using activity assay kits (Comin, Suzhou, China) based on the below principles.
POD catalyzes the oxidation of specific substrates by H2O2 and exhibits characteristic light absorption at 470 nm.
CAT catalyzes the decomposition of H2O2 by CAT with characteristic light absorption at 405 nm.
SOD can scavenge OFR, and OFR can reduce azotetrazolium to produce blue methanamine, which exhibits absorption at 560 nm.
Nitroblue tetrazolium staining
-
Nitroblue tetrazolium (NBT) staining was performed according to existing methods[45].
GUS staining and enzyme activity assay
-
Transgenic Arabidopsis (ProMdSAT1::GUS) seedlings were immersed in GUS staining buffer consisting of 1 mM 5-bromo-4-chloro-3-indolyl-β-glutamic acid, 100 mM sodium phosphate (pH 7.0), 0.1 mM EDTA, 0.5 mM ferricyanide, and 0.1% (v/v) Triton X-100 37 °C for 1 h in the dark. To quantify GUS activity, proteins were extracted from the seedlings with 1 mL of extraction buffer (50 mM Na2HPO4/NaH2PO4 [pH 7.0]), 10 mM β-mercaptoethanol, 10 mM Na2-EDTA, 0.1% (v/v) Triton X-100), and 1 mL of RIPA lysis buffer. A protein assay kit (Bio-Rad) was used to determine the total protein concentration. To measure GUS, 100 µL of the protein extract was added to 900 µL of GUS reaction buffer containing 1 mm 4-methylumbelliferone glucuronide and the mixture was incubated at 37 °C for 0, 5, 10, 15, 30, and 60 min. Then, 100 µL of the reaction mixture was added to 900 µL of the termination solution (1 M sodium carbonate). Fluorescence values were measured using a VersaFluor Spectrofluorometer (Bio-Rad) at an excitation wavelength of 365 nm and an emission wavelength of 455 nm. Arabidopsis (ProMdSAT1::GUS) seedlings (seven days-old) were pre-treated with ddH2O for two days, treated with 1 mM KCl, KNO3, or NH4Cl for different time periods, immersed in the GUS staining solution, and photographed.
Root system analysis
-
Arabidopsis taproots were observed and photographed under a body view microscope. Digimizer software was used to measure and calculate the number and length of root hairs in a 4-mm area starting 2 mm from the root tip.
Data analysis
-
All experiments were repeated independently three times, unless otherwise indicated. The data are expressed as mean and standard deviation. Data were analyzed by one-way analysis of variance, and means were compared using Duncan's multiple range test. Different letters indicate significant differences at the P < 0.05 level.
This work was supported by Shandong Province Key R&D Program, 2021CXGC010802 (Chun-Xiang You); Taishan scholar foundation, LJNY202026 (Wen-Sheng Gao); National Natural Science Foundation of China, 31972378 (Xiao-Fei Wang) and China Agriculture Research System of MOF and MARA, CARS-27 (Xiao-Fei Wang). We sincerely thank our team leader Dr. Yu-Jin Hao, who will be remembered for his great achievement and for the support and help in our work.
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Multiple sequence alignment for 13 proteins.
- Supplemental Fig. S2 GUS staining of ProMdSAT1::GUS transgenic Arabidopsis under KCl, KNO3, and NH4Cl treatments
- Supplemental Fig. S3 Dynamic changes of ProMdSAT1::GUS activity under different forms of nitrogen treatment.
- Supplemental Fig. S4 Expression of MdSAT1 varies with lateral root genesis and growth.
- Supplemental Fig. S5 Tissue expression pattern ofMdSAT1.
- Supplemental Fig. S6 The expression of MdSAT1 in transgenic Arabidopsis.
- Supplemental Table S1 Primers for qRT-PCR.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li T, Feng Z, Zhu B, Li M, Li G, et al. 2022. Functional identification of bHLH transcription factor MdSAT1 in the ammonium response. Fruit Research 2:17 doi: 10.48130/FruRes-2022-0017 

Functional identification of bHLH transcription factor MdSAT1 in the ammonium response
- Received: 05 September 2022
- Accepted: 31 October 2022
- Published online: 22 November 2022
Abstract: Plants mainly uptake inorganic nitrogen from soil as ammonium and nitrate. Less energy is required to assimilate ammonium compared to nitrates, and plants prefer to take up ammonium when the external nitrogen concentration is low. Investigating the patterns and mechanisms of ammonium absorption can help improve crop nitrogen utilization. In this study, we isolated MdSAT1, a gene of apple encoding an ammonium-responsive bHLH transcription factor. MdSAT1 promoted the growth and development of lateral roots and root hairs. Overexpression of MdSAT1 increased the transcript levels of genes related to ammonium uptake, indicating that MdSAT1 can regulate ammonium uptake and utilization at the transcriptional level. MdSAT1 also can modulate reactive oxygen species (ROS) accumulation to ultimately regulate plant growth. Taken together, these findings provide insight for us to further study the mechanisms by which MdSAT1 controls ammonium utilization as well as plant growth and development.
-
Key words:
- Ammonium /
- MdSAT1 /
- Functional identification










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}