








-
Studies on the taxonomy and phylogeny of freshwater fungi in Thailand, mostly focused on the northern region, and resulted in the discovery of new taxa[1−11]. A checklist of freshwater fungi of Thailand published up to the end of 2010 was provided by Zhang et al.[12] and reported 173 identified species (in 112 genera), of which 26 were new species. The number of novel fungi from freshwater habitats in Thailand increased significantly over the past years wherein Calabon et al.[8] listed an additional 129 new species, dominated by Dothideomycetes (86 species) and Sordariomycetes (40 species), between the years 2015–2020.
In freshwater habitats, Dothideomycetes is the second largest class in Ascomycota (677 species, 229 genera), next to Sordariomycetes with 823 species (in 298 genera)[10,13]. Dong et al.[7] provided a monograph of freshwater dothideomycetous taxa with a comprehensive taxonomic and phylogenetic account. Among these diverse taxa are members of Pleosporales with 391 species dominated by Dictyosporiaceae (55 species), Aigialaceae (27), Lindgomycetaceae (25), and Astrosphaeriellaceae (24). For the monotypic family Acrocalymmaceae, three species were reported from freshwater habitats: A. aquaticum[14], A. bipolare[7] and A. fici[11]. Acrocalymma aquatica, characterized by conidia having a single polar appendage, was introduced by Zhang et al.[12] collected from a freshwater stream in Chiang Mai, Thailand. This is the first species in the genus known to thrive in freshwater habitats. Acrocalymma fici, a species with flaring mucoid polar appendage, was reported by Boonmee et al.[11] from a freshwater stream in western Thailand. The species was introduced by Trakunyingcharoen et al.[15] which was collected from a terrestrial habitat on Ficus sp. The latest addition is Acrocalymma bipolare introduced by Dong et al.[7] characterized by conidia filled with oil droplets and bipolar appendages that elongate in contact with water forming filaments
The present study aims to broaden our knowledge of the taxonomy of lignicolous freshwater fungi using morpho-phylogenetic approach. Freshwater habitats in Thailand were explored to document extant and novel aquatic fungi. A new lignicolous freshwater species belonging to Acrocalymma from submerged wood in Thailand was discovered and Acrocalymma aquatica was recollected from Tak Province. A detailed description with morphological illustrations and multi-gene phylogenetic analyses to confirm the placement of new findings are provided in this paper.
-
Samples of submerged decayed wood were collected from a stream in Ban Mae Ja Wang, Tha Song Yang District, Tak Province, Thailand on 17 October 2019. Samples were incubated for five days and observed using a stereomicroscope for the presence of fruiting bodies[16]. The specimens were examined using a Motic SMZ 168 Series stereomicroscope with built-in camera for fungal fruiting bodies on the woody substrate. Micromorphological characters (e.g., ascoma, asci, ascospores) were photographed using a Nikon Eclipse 80i compound light microscope equipped with a Canon EOS 600D digital camera. Single spore isolation was used to obtain pure culture as described by Senanayake et al.[16]. Briefly, hand-sectioned ascomata were aseptically transferred from the wood to sterile glass slides containing distilled water. Asci were teased out to release spores and transferred to malt extract agar (MEA) medium. After 24 h, germinated spores were aseptically transferred into fresh MEA medium. Culture plates were incubated at 25 °C for one month and checked weekly for growth. Herbarium specimens were deposited in Mae Fah Luang University (MFLU). Living cultures were deposited at Mae Fah Luang University Culture Collection (MFLUCC). The new species was registered in Faces of Fungi (
www.facesoffungi.org )[17] and Index Fungorum databases[18]. Herbarium material of Acrocalymma fici MFLU 21–0124 was loaned from Mae Fah Luang University Fungarium, for comparison with the new taxon.DNA extraction, PCR amplification, and sequencing
-
Fungal mycelia from pure cultures grown in MEA for 30 d were scraped using a sterilized scalpel and kept in a 1.5 mL microcentrifuge tube. Genomic DNA was extracted using the Biospin Fungus Genomic DNA Extraction kit (BioFlux®, China) following the manufacturer's protocol. Polymerase chain reaction (PCR) was used to amplify four markers: the large subunit (LSU), small subunit (SSU), internal transcribed spacers (ITS) of rDNA, and the translation elongation factor 1-alpha gene (TEF1-α). The LSU was amplified using the primers LR0R and LR5[19]. For SSU and ITS, primers NS1/NS4 and ITS5/ITS4 were used[20]. TEF1-α was amplified using primers EF1-983F and EF1-2218R[21]. Polymerase chain reaction was performed in a volume of 25 μl, which contained 12.5 μl of 2× Power Taq PCR Master Mix (Bioteke Co., China), 1 μl of each primer (10 μM), 1 μl genomic DNA, and 9.5 μl deionized water. The amplification conditions used were as follows: initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 90 s, annealing at 55 °C (28S) and 48 °C (18S) for 1 min, and final extension at 72 °C for 10 min. Agarose gel electrophoresis was carried out to confirm the presence of amplicons at the expected molecular weight. PCR products were purified and sequenced with the primers mentioned above at a commercial sequencing provider (BGI, Ltd., Shenzhen, China). A BLAST search of the newly generated sequences was carried out to exclude contamination and to search for related taxa in the GenBank database (
www.ncbi.nlm.nih.gov/blast ).Phylogenetic analysis
-
The taxa table was assembled based on the closest matches from the BLAST search results and from recently published data by Mortimer et al.[22] and Konta et al.[23]. Sequences generated from the four loci were analyzed along with other sequences retrieved from GenBank (Table 1). Four datasets, one for each locus, were aligned with MAFFT v. 7 using the web server (
http://mafft.cbrc.jp/alignment/server [24]). Alignment was further refined manually, where necessary, using BioEdit v.7.0.9.0[25]. Aligned sequences were automatically trimmed using TrimAl v. 1.3 on the web server (http://phylemon.bioinfo.cipf.es/utilities.html ). The online tool 'ALTER'[26] was used to convert the alignment file to phylip format. Phylogenetic analysis of both individual and combined gene data was performed using maximum likelihood (ML) and Bayesian inference (BI).Table 1. Taxa used in this study for the analysis of combined SSU, LSU, and ITS rDNA sequence data and their GenBank accession numbers. The newly generated sequences are indicated with * and the ex-type strains are indicated in bold.
Taxon Strain / voucher number SSU LSU ITS TEF1-α Acrocalymma ampeli MFLUCC 20-0159 MW079341 MW063211 MW063150 – Acrocalymma ampeli NCYUCC 19-0288 MW079342 MW063212 MW063151 – Acrocalymma aquaticum MFLUCC 11-0208 JX276953 NG_042698 NR_121544 – Acrocalymma aquaticum MFLUCC 20-0124 – MT875393 MT875395 MT897894 Acrocalymma arengae MFLUCC 15–0327A ON650177 ON650673 ON650154 – Acrocalymma arengae MFLUCC 15–0327B ON650178 ON650674 ON650155 – Acrocalymma bilobatum K.L. Chen L119 – – KX034339 – Acrocalymma bilobatum* MFLUCC 20-0125 – MT875394 MT875396 MT897895 Acrocalymma bipolare MD1321 – NG_075326 – – Acrocalymma chuxiongense IFRDCC3104 – ON596248 ON595715 – Acrocalymma cycadis CBS 137972 – NG_057046 NR_137884 – Acrocalymma fici CBS 317.76 – NG_057056 NR_137953 KP170663 Acrocalymma fici MFLUCC 21-0103 – MT860429 MT864351 – Acrocalymma guizhouense CGMCC 3.20853 OM838471 OM838474 OM838410 – Acrocalymma guizhouense GZUIFR H22.028 OM838472 OM838475 OM838411 – Acrocalymma guizhouense GZUIFR H22.029 OM838473 OM838476 OM838412 – Acrocalymma hongheense HKAS 111907 MW424792 MW424777 MW424763 – Acrocalymma hongheense HKAS 111908 MW424791 MW424776 MW424762 – Acrocalymma hongheense HKAS 111909 MW424790 MW424775 MW424761 – Acrocalymma magnoliae MFLUCC 18–0545 OL331094 OK655819 OL413439 – Acrocalymma magnoliae MFLUCC 21–0204 OL331095 OK655820 OL413440 – Acrocalymma medicaginis CPC 24340 – KP170713 KP170620 – Acrocalymma medicaginis MFLUCC 17-1423 MT214387 MT214432 MT214338 – Acrocalymma medicaginis MFLUCC 17-1439 MT214388 MT214433 MT214339 – Acrocalymma pterocarpi MFLUCC 17-0926 MK347840 NG_066306 MK347732 MK360040 Acrocalymma pterocarpi MFLUCC 18–0718 OL331093 OK655818 OL413438 – Acrocalymma pterocarpi NC13-171 – LC517881 LC517880 – Acrocalymma vagum CPC 24225 – – KP170635 – Acrocalymma vagum CPC 24226 – – KP170636 – Acrocalymma walkeri UTHSC DI16-195 – LN907338 LT796832 LT797072 Acrocalymma yuxiense HKAS 111910 MW424793 MW424778 – – Ascocylindrica marina MD6011 KT252907 KT252905 – – Ascocylindrica marina MF416 MK007124 MK007123 – – Boeremia exigua CBS 431.74 EU754084 EU754183 FJ427001 KY484687 Boeremia foveata CBS 341.67 GU238203 GU237947 GU237834 KY484716 Maximum likelihood analysis was performed using RAxML v. 8 software on the CIPRES web portal[27−29] (
www.phylo.org/portal2 [30]). The GTR+GAMMA model of nucleotide evolution was used. RAxML rapid bootstrapping of 1000 replicates was performed. Bayesian inference analysis was performed using MrBayes v. 3.2.6 on XSEDE at the CIPRES webportal[31], using the parameter setting of 2 parallel runs, 4 chains, run for 1,000,000 generations at which point the standard deviation of split frequencies was below 0.01. Trees were sampled every 200th generations and all other parameters were left as default. In the phylogenetic tree, a bootstrap value of 95%–100% ML and 0.95–1.00 BYPP was considered high support. Newly generated sequences have been deposited in GenBank (Table 1). -
The combined SSU, LSU, ITS and TEF1-α gene dataset comprised of 32 taxa from Acrocalymmaceae and closely related taxa, with Boeremia exigua (CBS 431.74) and B. foveata (CBS 341.67) as the outgroup taxa (Table 1).
The analyzed dataset, after trimming, comprised a total of 4103 characters including gaps (SSU = 987, LSU = 1,336 bp, ITS = 869, TEF1-α = 911) with 701 distinct alignment patterns and 49.47% proportion of gaps and completely undetermined characters. The ML analysis for the combined dataset provided the best scoring tree (Fig. 1) with a final ML optimization likelihood value of –11459.976 (ln). Parameters for the GTR model of the combined SSU, LSU, and ITS dataset are as follows: estimated base frequencies; A = 0.250, C = 0.250, G = 0.250, T = 0.250; substitution rates AC = 1.47340, AG = 1.83232, AT = 1.47340, CG = 1.079563, CT = 4.16678, GT = 1.000000; gamma distribution shape parameter α = 0.227521. Bayesian analysis resulted in 5001 samples of which 3751 samples were included after 1000000 generations. Phylogenetic analyses of the combined data matrix resulted in considerably high bootstrap support and well-resolved clades (Fig. 1). The tree topologies resulted from ML and BI analyses are similar. Support values for maximum likelihood (ML) above 75%, and Bayesian posterior probabilities (BYPP) greater than 0.95 are given at the nodes.

Figure 1.
Phylogenetic tree based on RAxML analyses of combined SSU, LSU, ITS, and TEF1-α sequence data. Bootstrap support values for maximum likelihood (ML) higher than 75% and Bayesian posterior probabilities (BYPP) greater than 0.95 are indicated above the nodes as ML/BYPP. The new species are represented in blue bold and type species are in bold. The tree is rooted to Boeremia foveata (CBS 341.67) and B. exigua (CBS 431.74) (Didymellaceae). Bar = 0.03 estimated number of nucleotide substitutions per site per branch
In the phylogenetic analysis (Fig. 1), Acrocalymma formed a well-supported monophyletic clade separate from the closely related family Ascocylindricaceae (100% ML, 1.00 BYPP). The novel species A. bilobatum (MFLUCC 20–0125) clustered with Acrocalymma sp. (K.L. Chen L119), which was named as A. bilobatum (K.L. Chen L119) in this study, see notes of A. bilobatum. Six distinct clades were formed: Clade I [A. bilobatum (MFLUCC 20–0125; K.L. Chen L119), A. pterocarpi (MFLUCC 17–0926; MFLUCC 18–0718; NC13-171), A. medicaginis (MFLUCC 17-1423; MFLUCC 17-1439), A. bipolare (MD1321), Acrocalymma chuxiongense (IFRDCC3104)]; Clade II [A. hongheense (HKAS 111907; HKAS 111908; HKAS 111909), Acrocalymma arengae (MFLUCC 15–0327A; MFLUCC 15–0327B), A. vagum (CPC 24225; CPC 24226), A. walkeri (UTHSC DI16-195); A. aquaticum (MFLUCC 11–0208; MFLUCC 20–0124)]; Clade III [A. ampeli (MFLUCC 20-0159; NCYUCC 19-0288)]; Clade IV [A. guizhouense (CGMCC 3.20853; GZUIFR H22.028; GZUIFR H22.029)]; Clade V [A. magnoliae (MFLUCC 18–0545; MFLUCC 21–0204)]; Clade VI [A. fici (CBS 317.76; MFLUCC 21–0103); A. yuxiense (HKAS 111910)].
Acrocalymma aquaticum Huang Zhang & K.D. Hyde, Cryptogr. Mycol. 33(3): 337 (2012)
Mycobank number: 835894; Facesoffungi number: FoF 07098, Fig. 2
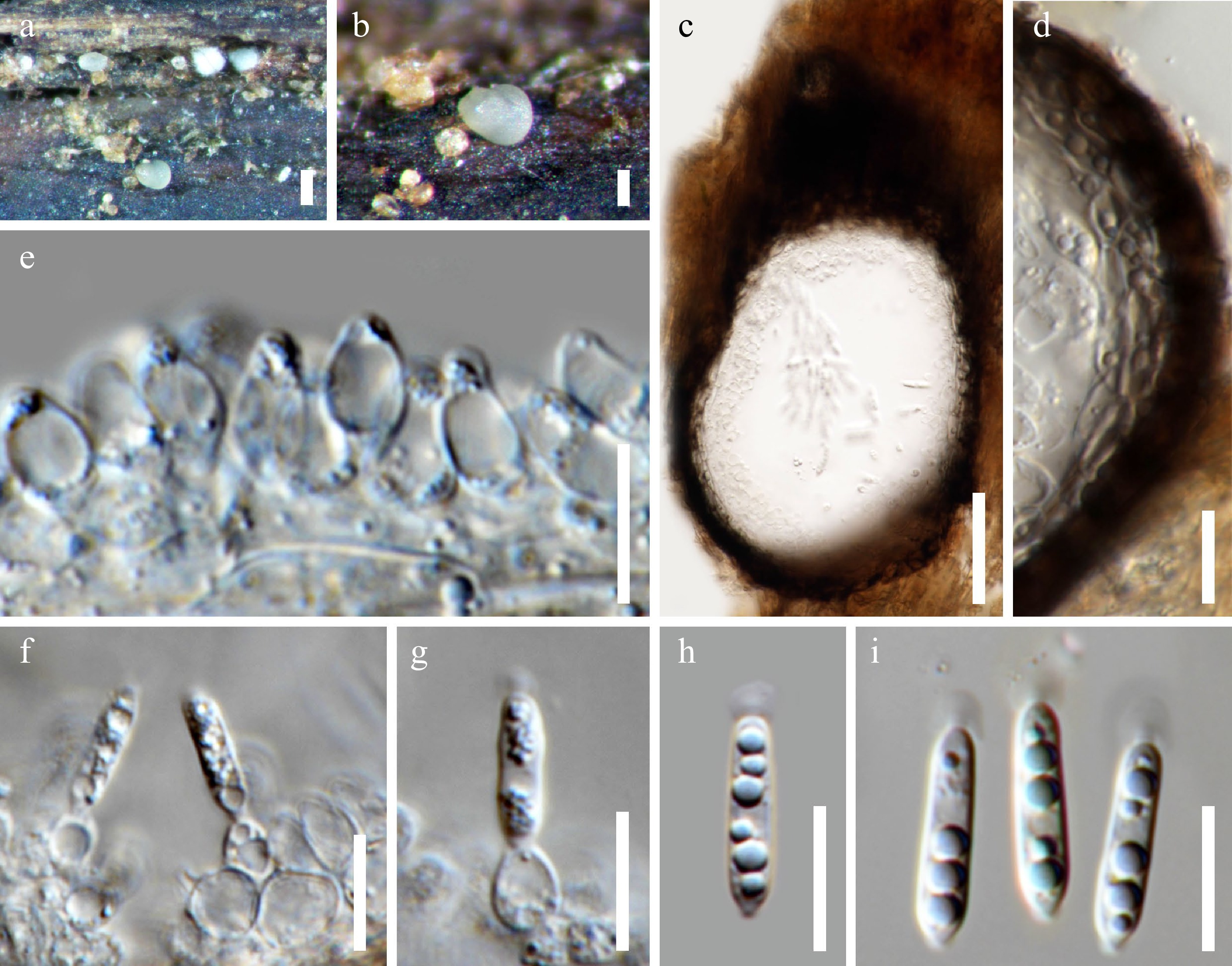
Figure 2.
Acrocalymma aquatica (MFLU 22–0114). (a), (b) Erumpent conidiomata on wood surface. (c) Vertical sections of a conidioma. (d) Section through the peridium. (e)–(g) Conidiogenous cells. (h), (i) Conidia with apical appendages. Scale bars: (a) = 200 μm, (b) = 100 μm, (c) = 50 μm, (d)–(i) = 10 μm.
Saprobic on submerged wood in a freshwater stream. Sexual morph: Undetermined. Asexual morph: Conidiomata 240–270 µm diam., 100–140 µm high, dark brown or black, solitary, pycnidial, immersed to semi-immersed, unilocular, globose to subglobose, glabrous, papillate, ostiolate. Ostiole single, centrally located. Peridium 12–18 µm thick, composed of thick-walled, dark brown to black cells of textura angularis in the outer layers, becoming hyaline cells towards conidial hymenium. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 6–12 × 3.5–6 µm, hyaline, enteroblastic, phialidic, ampulliform to lageniform, with a narrow channel, determinate, formed from the inner cells of the pycnidial wall. Conidia 11.5–16.5 × 2–4 µm (x̅ = 13.9 × 2.7 µm, n = 20), hyaline, cylindrical with an obtuse apex and a narrow truncate base, straight, 0–1-septate, not constricted at the septa, thin and smooth-walled, guttulate, bearing a mucilaginous appendage (2–3 μm diam.) at the apex.
Culture characters: Conidia germinated on MEA within 24 h. Colonies on MEA reaching 4.5–5 cm diam., after 4 weeks at room temperature, colonies circular, medium dense, flat with smooth and entire margins; smoke-gray to gray in top view, reverse gray.
Material examined: THAILAND, Tak Province, Tha Song Yang, Ban Mae Ja Wang stream, on submerged wood, 17 October 2019, OD Padaruth, CC36 (MFLU 22–0114); living culture MFLUCC 20–0124.
Known distribution: THAILAND: Chiang Mai Province (Zhang et al.[14]), Tak Province (present study).
Notes: Acrocalymma aquaticum was introduced by Zhang et al.[14], which was collected from a submerged wood in a freshwater stream in Chiang Mai, Thailand. Based on the phylogenetic analysis of combined SSU, LSU, ITS, and TEF1-α sequence data, A. aquaticum (MFLUCC 20–0124) clustered with the ex-type strain of A. aquaticum (MFLUCC 11–0208) with high bootstrap support (100% ML, 1.00 BYPP). Our isolate resembles A. aquaticum in morphology and measurements of the conidiomata, conidiogenous cells, and conidia but most of the conidiomata were immersed. A comparison of the ITS nucleotides of A. aquaticum (MFLUCC 11–0208) and the new strain (MFLUCC 20–0124) revealed 1 bp (0.19%; 540 bp) nucleotide difference which indicates that the new strain is A. aquaticum[32].
Acrocalymma bilobatum M.S. Calabon, E.B.G. Jones & K.D. Hyde sp. nov.
Mycobank number: 848523; Facesoffungi number: FoF 13985; Fig. 3
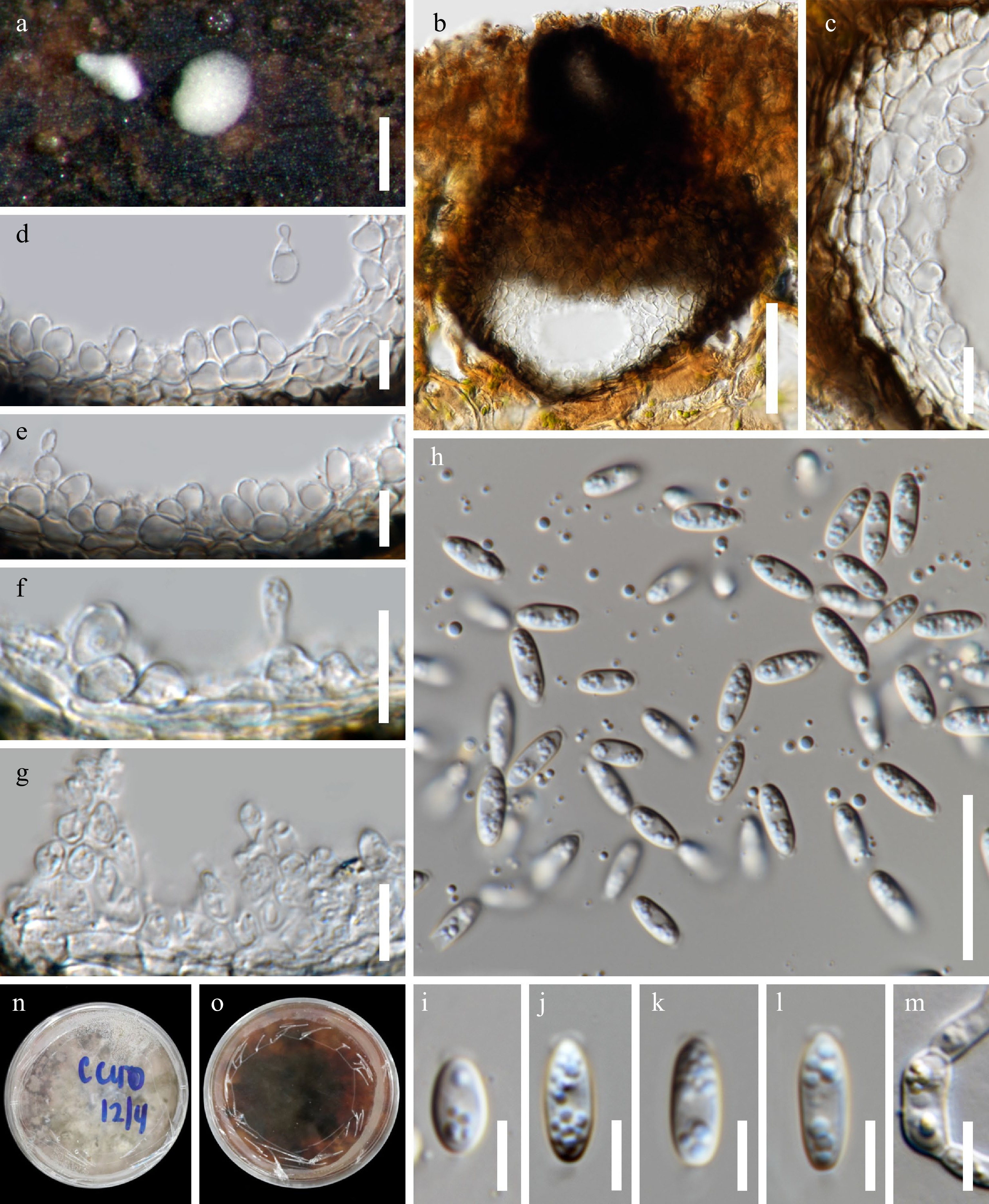
Figure 3.
Acrocalymma bilobatum (MFLU 22–0115, holotype). (a) Appearance of erumpent dark brown conidiomata on wood surface. (b) Vertical section of conidioma. (c) Section through the peridium. (d)–(g) Conidiogenous cells. (h)–(l) Conidia with appendages. (m) Germinated conidium. Colony on MEA: (n) obverse. Scale bars: (a) = 200 μm, (b) = 50 μm, (c)–(g) = 10 μm, (h) = 20 μm, (i)–(l) = 5 μm.
Etymology: derived from the bilobed polar appendages in the conidia
Holotype: MFLU 22–0115
Saprobic on submerged wood in freshwater habitat. Sexual morph: Undetermined. Asexual morph: Conidiomata 170–275 µm diam., 135–205 µm thick/high, dark brown, pycnidial, solitary to gregarious, immersed with only the white neck visible in surface view, globose or subglobose, unilocular, glabrous and ostiolate. Ostiole cylindrical, centrally located. Peridium 12–27 µm thick, composed of thick-walled, dark brown cells of textura angularis in the outer layer, become hyaline cells of textura globulosa in the inner layer. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 4.5–11 × 3–5 μm, hyaline, heteroblastic, doliiform to ampulliform, determinate, smooth-walled, formed from the inner cells of the pycnidial wall. Conidia 7–12 × 2.5–4 µm (x̅ = 9.2 × 3.5, n = 30), hyaline, cylindric-clavate to fusiform, rounded at apex, truncate at base, straight, unicellular, thin and smooth-walled, guttulate, with rounded mucoid bilobed polar appendage (0.4–0.9 × 2–4 μm), and basal globose to hemispherical flaring mucoid appendage (0.8–1.1 × 0.1–1.4 μm).
Culture characters: Conidia germinated on MEA within 24 h. Colonies on MEA reaching 4–5 cm diam., after 4 weeks at room temperature, colonies circular, medium dense, flat with smooth and entire margins; gray to pale brown in top view, reverse dark brown to black in center and reddish in the outer region.
Material examined: THAILAND, Tak Province, Tha Song Yang, Ban Mae Ja Wang stream, on submerged wood, 17 October 2019, OD Padaruth, CC40 (MFLU 22–0115, holotype); ex-type culture MFLUCC 20–0125.
Notes: In the phylogenetic analysis, Acrocalymma bilobatum (MFLUCC 20–0125) grouped with Acrocalymma sp. (K.L. Chen L119), with high bootstrap support (96% ML, 0.98 BYPP). Comparison of morphological features of Acrocalymma sp. (K.L. Chen L119) and Acrocalymma bilobatum (MFLUCC 20–0125) was not possible because the morphology of the former is not available but comparison of the ITS nucleotides of Acrocalymma sp. (K.-L. Chen L119) and A. bilobatum (MFLUCC 20–0125) revealed two (0.37%) base pair differences, which indicates that the former is A. bilobatum. The two strains of A. bilobatum shared the same subclade with Acrocalymma chuxiongense (IFRDCC3104) and Acrocalymma bipolare (MD1321). Acrocalymma bilobatum resembles A. ampeli, A. medicaginis, and A. bipolare in the shape of conidia and appendages at both ends but differs in the sizes of conidia and appearances of the appendages. Acrocalymma bipolare has a mucoid polar appendage filled with oil droplets, which elongates in water to form filaments[7], while A. medicaginis has globose to hemispherical or helmet-shaped apical appendage and tapered, short, cylindrical to hemispherical basal appendage[33]. Acrocalymma ampeli has a flaring mucoid basal appendage with a hemispherical to bilobed apical appendages[34].
Acrocalymma fici P.W. Crous & T. Trakunyingcharoen, IMA Fungus 5 (2): 405 (2014)
Mycobank number: 810838; Facesoffungi number: FoF 09155, Fig. 4

Figure 4.
Acrocalymma fici (MFLU 21–0124). (a) Appearance of conidiomata on wood surface. (b) Vertical section of conidioma. (c) Section through the peridium. (d), (e) Conidiogenous cells. (f), (g) Conidia with apical appendages. (g) Conidia stained with nigrosin. Scale bars: (a) = 500 μm, (b) = 50 μm, (c), (f), (g) = 20 μm, (d), (e) = 5 μm.
Saprobic on submerged decaying wood. Sexual morph: Undetermined. Asexual morph: Conidiomata 80–150 × 40–100 μm, white, separate to gregarious, immersed to semi-immersed, pycnidial, globose to subglobose, unilocular, glabrous, ostiolate. Peridium 40–50 μm thick, composed of thick-walled, dark brown to hyaline cells of textura angularis, become darker cells at the ostiolar region. Ostiole 40–55 μm diam., centrally located. Conidiophores reduced to conidiogenous cells or a supporting cell. Conidiogenous cells 4–10 × 2–5 µm, hyaline, enteroblastic, ampulliform to doliiform, smooth-walled. Conidia 12–15 × 2–3 (x̅ = 13.4 × 2.8 μm, n = 10), hyaline, cylindrical with sub-obtuse apex, acutely tapered at base to a small flattened central scar, 0–1-septate, not constricted at septum, smooth-walled, guttulate, with flaring mucoid apical appendage (2–5 μm diam.), visible in water mounts.
Material examined: THAILAND, Kanchanaburi, Sangkhla Buri, Liwo, on decaying wood submerged in a stream, 27 June 2019, N. Chaiwan, TFW5 (MFLU 21–0124); ex-type culture, MFLUCC 21–0103.
Notes: Acrocalymma fici (CBS 317.76) was collected from a terrestrial habitat on Ficus sp.[15] and on A. fici (MFLUCC 21–0103) on submerged decaying wood in Thailand[11]. Acrocalymma fici (MFLUCC 21–0103) resembles A. fici (CBS 317.76), but almost all the conidia are aseptate, while Trakunyingcharoen et al.[15] observed septate conidia. The differences in conidial septation may be explained by conidiomatal differences. A comparison of the ITS and TEF nucleotides of A. fici (CBS 317.76) and A. fici (MFLUCC 21–0103) revealed no nucleotide difference, which indicates that the new strain is A. fici[11,32].
-
Acrocalymmaceae, whose establishment was supported using divergence time estimates, comprises a monotypic genus Acrocalymma typified by A. medicaginis[15,35,36]. Presently, 16 species are included in this genus: A. aquatica[14], A. ampeli[34], A. arengae[23], A. cycadis[37], A. bilobatum (this study), A. bipolare[7], A. fici, A. vagum, A. walkeri[15], A. chuxiongense[38], A. guizhouense[39], A. magnoliae[40], A. medicaginis[33], A. pterocarpi[41], A. hongheense, A. yuxiense[22]. Acrocalymma species are known to thrive as pathogens (i.e., A. medicaginis, A. vagum) and saprobes (e.g., A. ampeli, A. pterocarpi) of various plant hosts (e.g., A. ampeli on Ficus ampelas, A. arengae on Arenga pinnata, A. fici on Ficus sp., A. cycadis on Cycas calcicola, A. magnoliae on Magnolia sp. and Anomianthus dulcis, A. yixienxe on Quercus glauca) in terrestrial habitats. Five species, A. arengae, A. chuxiongense, A. hongheense, A. pterocarpi, and A. walkeri are sexual morphs and the rest are asexual coelomycetous species. Mortimer et al.[22] discovered the sexual (HKAS 111909) and asexual morph (HKAS 111907; HKAS 111908) of A. hongheense collected on woody litter in China. The morphological differences of sexual and asexual morphs and comparison of habitats and localities of Acrocalymma spp. are provided by Liu & Zeng[38] and de Silva et al.[40], respectively. A key to species is provided as Table 2.
Table 2. Key to species of Acrocalymma
Step Features Species 1a Sexual morph 2 1b Asexual morph 6 2a Asci, <100 μm 3 2a Asci, >100 μm 4 3a Ascospores, 17–21 × 3–5 μm A. pterocarpi 3b Ascospores, 19–22 × 4.5–5.5 μm A. walkeri 4a Ascospores, 1-septate 5 4b Ascospores, 1–3-septate A. arengae 5a Ascospores, 35–45 × 18–20 μm A. chuxiongense 5b Ascospores, 20–35 × 7–9 μm A. hongheense 6a Conidia lacks mucoid cap 7 6b Conidia with mucoid caps 8 7a Conidia, (16–)18–25(– 28) × (4.0–)4.5–6.0(–6.9) μm A. vagum 7b Conidia, 15–21 × 4–5 μm A. yuxiense 8a Conidia, mucoid caps in apex 9 8b Conidia, mucoid caps in both ends 13 9a Conidia, <20 µm 10 9b Conidia, >20 µm 12 10a Conidia <10 A. guizhouense 10b Conidia >10 11 11a Conidia, 12–17 × 3–4 µm A. cycadis 11b Conidia, 12–16 × 2.5–3 µm A. magnoliae 12a Conidia, 22–30 × 5–7 μm A. aquatica 12b Conidia, (25–)28–32(–35) × (4–)5 μm A. fici 13a Conidia <15 14 13b Conidia >15 15 14a Conidia, 7–12 × 2.5–4 µm A. bilobatum 14b Conidia, 9–12 × 3–5 µm A. bipolare 15a Conidia, 17–19 × 5.5–6.5 μm A. ampeli 15b Conidia, 11–21 × 3.5–5.0 µm A. medicaginis In freshwater habitats, four species are reported as saprobes: A. aquaticum, A. bilobatum, A. bipolare, A. fici. Three of these, except for A. bipolare, are recorded in the freshwater environments of Thailand. The amazing biodiversity of freshwater fungi in Thailand is exceptional with many novel taxa identified[42−44]. Also, it is likely that there are more species of Acrocalymma awaiting discovery as we continuously explore both terrestrial and aquatic habitats and study the different plant hosts for their fungal associates.
MS Calabon is grateful to the Mushroom Research Foundation and Department of Science and Technology – Science Education Institute (Philippines). Saranyaphat Boonmee would like to thank the Mae Fah Luang University Fund (No. 631C15001). EB Gareth Jones is supported under the Distinguished Scientist Fellowship Program (DSFP), King Saud University, Kingdom of Saudi Arabia. The researchers are grateful to Dr. J. Monkai for organizing the collection trip under her project and to OD Padaruth for collecting the freshwater samples. KD Hyde is grateful to the National Research Council of Thailand (NRCT) grant 'Total fungal diversity in a given forest area with implications towards species numbers, chemical diversity and biotechnology' (Grant No. N42A650547).
-
The authors declare that they have no conflict of interest. Kevin D. Hyde is the Editorial Board member of Studies in Fungi. He was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and his research groups.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Calabon MS, Jones EBG, Boonmee S, Li WJ, Xiao YP, et al. 2023. Acrocalymmaceae (Pleosporales) from freshwater habitats in Thailand with the introduction of Acrocalymma bilobatum sp. nov.. Studies in Fungi 8:11 doi: 10.48130/SIF-2023-0011 

Acrocalymmaceae (Pleosporales) from freshwater habitats in Thailand with the introduction of Acrocalymma bilobatum sp. nov.
- Received: 09 January 2023
- Accepted: 04 May 2023
- Published online: 09 June 2023
Abstract: An observation of lignicolous freshwater fungi in Thailand resulted in the discovery of a novel taxon and recollection of Acrocalymma aquaticum. Morphology coupled with phylogenetic analysis support the placement of the new species in Acrocalymma. Acrocalymma bilobatum sp. nov. resembles A. ampeli, A. bipolare, and A. medicaginis in conidial shape and the mucoid appendages at both ends but differs in the sizes of conidia and appearance of appendages. Phylogenetic analysis of SSU, LSU, ITS, and TEF1-α sequence data shows that A. bilobatum is closely related to A. bipolare, A. chuxiongense, A. medicaginis, and A. pterocarpi. Acrocalymma aquatica was recollected in Thailand and a detailed description and photographic documentation of its morphological characteristics is provided. The herbarium specimen of Acrocalymma fici, collected from freshwater habitats in Thailand, was reexamined to complete a review of Acrocalymmaceae species in the country.
-
Key words:
- 1 new taxon /
- Coelomycetes /
- Dothideomycetes /
- Monotypic family /
- Multi-locus analysis