








-
Yams (Dioscorea spp. L.) are multiple species of the monocotyledonous family Dioscoreaceae that produce edible tubers for millions of people in tropical and sub-tropical regions, while D. alata (greater yam) was reported to be the most widely distributed yam species globally[1]. Greater yam is highly valued for its dietary carbohydrate, amino acids and essential minerals and widely grown in West Africa, Central Africa and the Caribbean. Recently, increased genomic and transcriptomic datasets have become available for Dioscorea species[2−7], which provide opportunity to conduct functional genomic studies. Although regeneration systems of greater yam were established and organogenesis directly from petiole explants was also reported[8,9], Agrobacterium-mediated transformation in yam was only reported for D. rotundata using axillary buds as explants[10].
Transient transformation in plant protoplasts provide a complementary way for gene functional analysis besides of the stable genetic transformation system, including localization of proteins, testing gene and promoter activities, validating protein-protein interactions, and analysis signal transduction[11−14]. Mesophyll protoplast-based transient expression assays are regularly used in biological studies in plants with stable protoplast isolation system, such as Arabidopsis, tobacco and rice[11,15−17], sometimes instead of studied species on their own[18−20]. Protoplasts are considered as a very reliable experimental system, as long as protoplast sources are carefully chosen[21]. Tor et al. prepared active protoplasts from immature leaves of Dioscorea yams and tested their transient gene expression[22]. Although immature leaves have fewer mucilage which is abundant in yam, it limits the quantity of tissues that can be used for protoplast preparation. Developing protoplast preparation for extended leaves in greater yam and optimizing transient transformation systems will be useful for gene function characterization.
In this study, we developed a highly repeatable and efficient protocol for mesophyll protoplast isolation and polyethylene glycol (PEG) − mediated gene transient expression in greater yam. Since greater yam leaves are rich in polysaccharide and polyphenols, the process of protoplast isolation was modified by extending the step of washing protoplasts from digested leaves and tissue culture seedlings were used as the starting materials. This protocol provides a facile tool for protein subcellular localization of an ethylene responsive transcription factor – DaERF2, which was characterized in our previous study and had a high expression level[23].
-
Tissue culture seedlings of greater yam accession Da70 were used in this study. The young seedlings were kept on MS (Murashige and Skoog) medium at 25 °C under a 12-h light/12-h dark photoperiod. The seedlings grown for 8−11 weeks after subculture were used for protoplast isolation. It is critical to obtain robust starting plant material.
Protoplast isolation
-
Protoplast isolation was carried out based on the protocols described by Yoo et al. [13] and Tor et al.[22] with modification. The second and third leaves from the top were selected and cut into 0.5−1.0 mm strips. During the cutting, the leaves were placed on 3−5 layers of filter papers soaked with 0.7 M mannitol and 0.9 to 1 g leaf strips were put into 10 ml enzyme solution. The concentration of mannitol based on the references mentioned in Tor et al. and the mannitol test in our lab, resulted in 0.7 M giving the highest protoplast yield and viability (data not shown). Different concentrations of Cellulose R-10 (0.4%, 0.8% or 1.2%, Yakult, Japan) and Macerozyme R-10 (0.5%, 1% and 1.5%, Yakult, Japan), which dissolved in the solution (0.1% BSA, 0.7 M mannitol, 20 mM KCl, 10 mM CaCl2, 20 mM MES, pH 5.8), were combined and tested for their effects on protoplast isolation under 6 h of enzymolysis in the dark (25 °C). All other chemical reagents used were ordered from Sigma-Aldrich Company (GC level, plant cell culture tested). Every enzyme combination was set as three technical repeats.
The digested leaf stripes were filtered through a nylon sieve (75 μm) by washing gently with 10 ml W5 solution (pre-cooled, 4 °C, 154 mM NaCl, 125 mM CaCl2, 5 mM KCl and 2 mM MES, pH 5.8). Since the greater yam leaves are rich in polysaccharide and polyphenols and the protoplasts tend to adhere to leaves, the digested leaves were washed with W5 solution gently when filtering. After filtering, the protoplasts released from leaves were collected by centrifugation (100× g for 10 min) twice and re-suspended in 20 ml W5 solution (pre-cooled, 4 °C). Finally, the protoplasts were subjected to centrifugation (100× g for 10 min) and re-suspended in 1 ml W5 solution (pre-cooled). Before further treatment, the isolated protoplasts hatched on ice for 30 min.
The protoplast yield and viability were evaluated via cell counting under a fluorescence microscope equipped with a hemocytometer. The viability of protoplasts was measured with 0.2% fluorescein diacetate (FDA) staining and determined as follows: protoplast viability (%) = (fluorescent protoplast number in view/protoplast total number in view) × 100%.
Vector construction and plasmid preparation
-
Total RNA was extracted from greater yam tuber according to the improved RNA extraction method in Palmae of Xiao et al.[24] and the first strand cDNA were synthesized according to the manufacturer’s protocols (RevertAid First Strand cDNA Synthesis Kit, Thermo Scientific). A Nanodrop ND-1000 spectrophotometer was used to test the RNA concentration and purity (Nanodrop Technologies, USA).
One ethylene responsive transcription factor (ERF) – DaERF2 (downloaded from the national center for biotechnology information website, Accession: HO811460.1) was used for the subcellular localization test, whose characters were featured in our previous research[23]. The DaERF2 gene was amplified by primers with Xba I and BamH I restriction enzyme sites (bold) introduced to their 5' ends (XbaI-ERF2-F: GCTCTAGAATGTGTGGAGGAGCTATCATATCG; BamHI-ERF2-R: CGGGATCCGAAAGAGTTGCCCACCATTG) from greater yam and inserted into the pA7 vector in fusion with a green fluoresce protein (GFP) tag at the N-terminal. The DaERF2 gene was amplified from greater yam cDNA sample under the following conditions: 95 °C for 5 min followed by 35 cycles of amplification (95 °C for 30 s, 55 °C for 30 s, 72 °C for 1.5 min) and a 10 min final extension at 72 °C.
The recombinant plasmid with DaERF2 gene cloned into the Xba I − BamH I sites of the pA7 vector, which was located on the 3' end of GFP gene, were confirmed by sequencing and designated as pA7-DaERF2-GFP. The recombinant plasmid was transformed into Escherichia coli strain − DH5α (TransGen Biotech, Beijing, China) for proliferation. The plasmid DNA was extracted via a commercial kit (Code EM121-01, Transgene, Beijing, China) to get high quality DNAs.
PEG-mediated protoplast transfection
-
PEG-mediated DNA transfection was conducted based on the protocol described by Yoo et al.[13], but optimized transfection time and PEG4000 concentration. The main transfection steps were as follows: the protoplasts were centrifuged (100× g for 10 min) and resuspended in MMg solution (0.7 M mannitol, 15 mM MgCl2, 4 mM MES); a final protoplast density 1.0 × 105 protoplasts/ml was adjusted, following by 14 ug of pA7/pA7-DaERF2-GFP plasmid DNA was mixed with 200 μl protoplasts. PEG4000 final concentration as 10%, 15%, 20% or 25% were freshly prepared in a solution (0.7 M mannitol, 100 mM CaCl2) and added into protoplasts separately. The mixtures were incubated at room temperature separately for 5, 10, 15 or 20 min and then were gently diluted with 600 μl W5 solution.
The transfect protoplasts were then centrifuged twice (100× g, 2 min) and re-suspended in 1 ml WI solution (4 mM MES, 0.4 M mannitol, 20 mM KCl, pH 5.7) and transferred into 1.5 ml tubes pre-coated with 1% BSA. The protoplast suspension was then incubated at room temperature in dark for 12−18 h to induce gene expression. Finally, we tested the transformation efficiency by confocal laser scanning microscope. Protoplast transformation rate (%) = (fluorescent protoplast number in view/protoplast total number in view) × 100%.
-
The third and fourth leaf of greater yam plantlet were harvested and used for protoplast preparation (Fig. 1a). Testing best enzyme concentration for protoplast isolation showed that greater yam protoplast yield increased with the rise of cellulase R-10 and macerozyme R-10 concentration, and the highest production of protoplasts (3.7 × 107 protoplasts/g fresh weight (FW)) was obtained with 0.8% cellulose and 1% / 1.5% macerozyme (Table 1). When cellulose concentration increased to 1.2%, the protoplast yield decreased (3.0 × 107 to 3.4 × 107 protoplasts/g FW). Digested leave tissues were not fragile enough when cellulose concentration was low (0.4%) and protoplast viability was also not high (72%−82%). The increase of cellulose concentration (0.8%) resulted in the rise of protoplast viability (86%−92%), but 1.2% cellulose caused significant decrease of protoplast viability (Table 1). Based on the above results, the enzyme combination of 0.8% cellulase and 1% macerozyme was determined as the best for greater yam protoplast preparation, and majority mesophyll protoplasts obtained under this condition were spherical and intact (Fig. 1b & c).
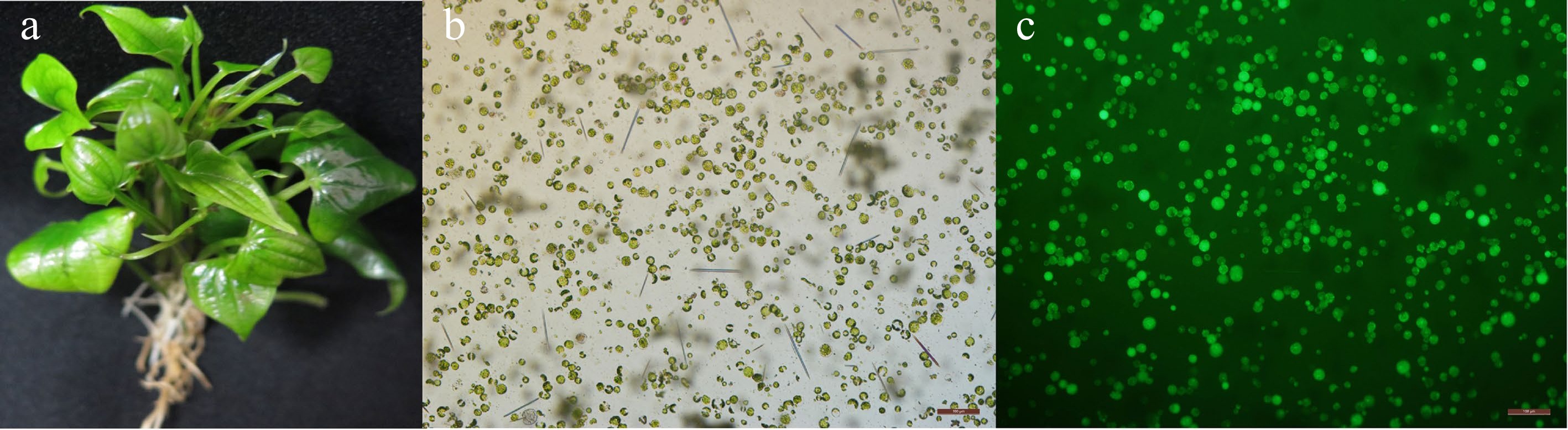
Figure 1.
Greater yam protoplast isolation and viability assay. (a) A healthy six-week-old greater yam plantlet after sub culture which is suitable for protoplast isolation. Mesophyll protoplasts were stained with FDA and observed under (b) bright light and (c) UV light. Scale bar = 100 μm.
Table 1. Effects of enzyme combination on the yield and viability of protoplasts isolated from leaves of greater yam.
Cellulase
R-10 (W/V)Macerozyme
R-10 (W/V)Protoplast yield
(107#/g FW)Protoplast
viability (%)0.4 0.5 2.19 ± 0.01a 72.32 ± 2.02a 0.4 1 2.28 ± 0.16a 79.27 ± 1.37b 0.4 1.5 2.97 ± 0.03b 82.31 ± 2.5b 0.8 0.5 3.46 ± 0.01cd 88.91 ± 2.45c 0.8 1 3.70 ± 0.04e 92.14 ± 0.72d 0.8 1.5 3.63 ± 0.09de 86.27 ± 2.16c 1.2 0.5 3.40 ± 0.1c 81.15 ± 1.08b 1.2 1 3.30 ± 0.13c 73.35 ± 2.2a 1.2 1.5 3.00 ± 0.02b 70.21 ± 1.41a The averages of three technical replicates ± standard error of the mean are shown. FW is abbreviation for fresh weight. The different letters indicate significant differences (p ≤ 0.05) according to the Duncan test. The digestion time in our study (6 h) was shorter than in previous research (15−18 h) (Tor et al.[22]. Testing digestion time with the optimum combination of enzymes showed that the protoplast yield increased steadily between digestion time of 3 h (3.1 × 107 protoplasts/g FW) to 9 h (4.2 × 107 protoplasts/ g FW) (Fig. 2a). However, protoplasts showed higher viability ratio when digesting time is between 6 h (95.6%) and 7 h (94.5%) than that of other digestion times (Fig. 2b).
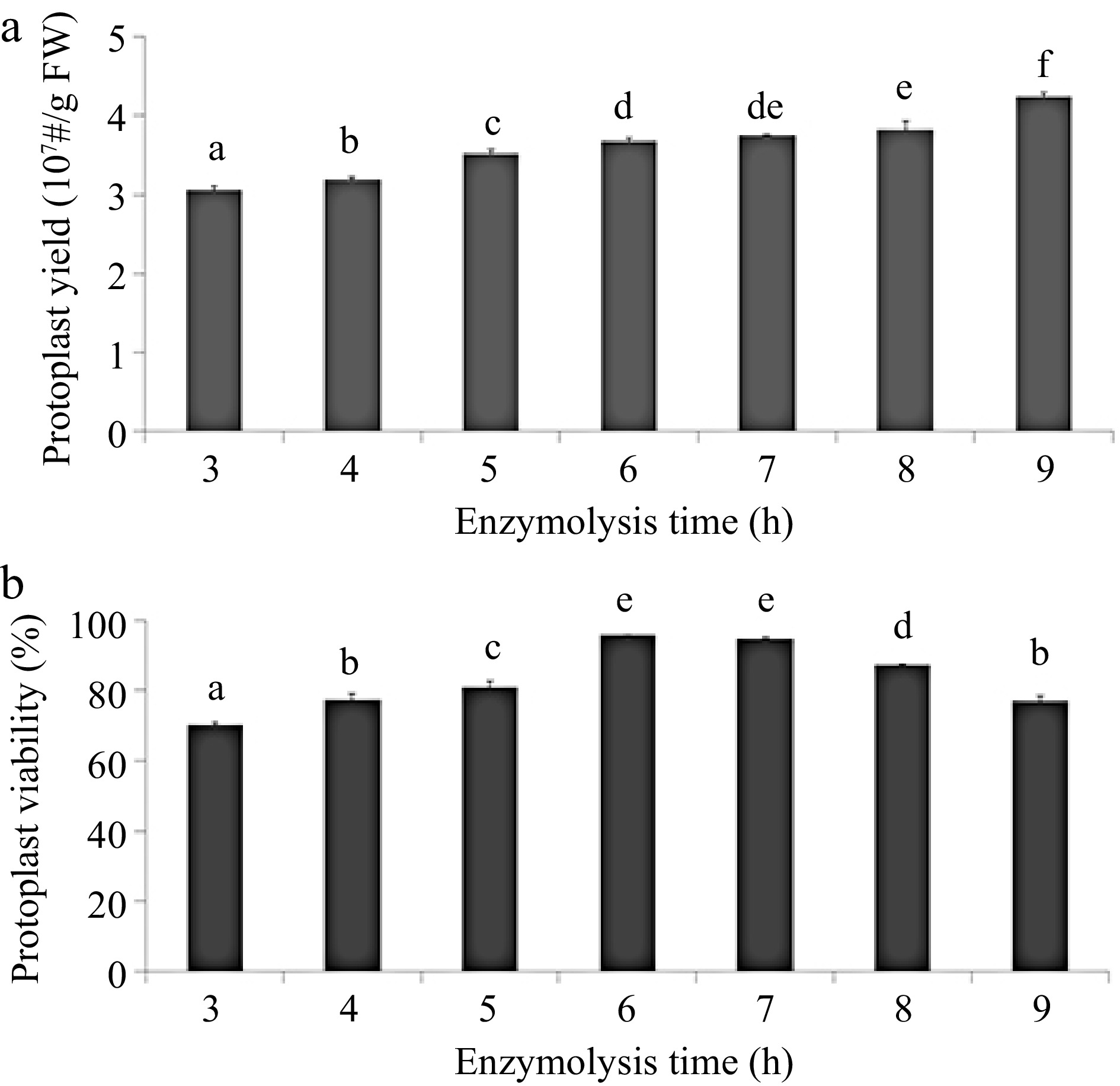
Figure 2.
Effects of digestion time on greater yam protoplast isolation. (a) Effects of digestion time on protoplast production. (b) Effects of digestion time on protoplast viability counted using the FDA staining assay. Different letters represent statistically significant difference at p ≤ 0.05 based on the Duncan test. Three biological replicates were used for each set of digestion time. The protoplast yield and viability were evaluated via cell counting under a fluorescence microscope equipped with a hemocytometer. The viability of protoplasts was measured with 0.2% fluorescein diacetate (FDA) staining and determined as follows: protoplast viability (%) = (fluorescent protoplast number in view/protoplast total number in view) × 100%.
Greater yam transient expression system establishment in its protoplasts
-
PEG-mediated transformation was used for greater yam mesophyll protoplasts transient transfection. To achieve higher transformation efficiency, the best PEG4000 concentration and transfection time were tested with the expression vector pA7, which has a green fluoresce protein (GFP) tag promoted by the constitutive CaMV 35S promoter on the vector. The transfection results showed that when transfection duration was set as 10 min, the transfection efficiency steadily increased from 23.5% to 48.1% between the PEG4000 concentrations of 10% to 20%, followed by a steep fall for 25% PEG4000 (Fig. 3a). When PEG4000 concentration was 20%, there was a steady increase of transfection efficiency from 5 min to 20 min PEG duration and peaked at 20 min PEG duration (Fig. 3b). Protoplast viability dropped to 80% after PEG4000 treatment for 5 min to 20 min, but it has a steep drop for 25 min transfection treatment (Fig. 3c). The highest transfection efficiency was achieved when PEG4000 concentration was 20% and transfection time was set as 20 min (Fig. 4).
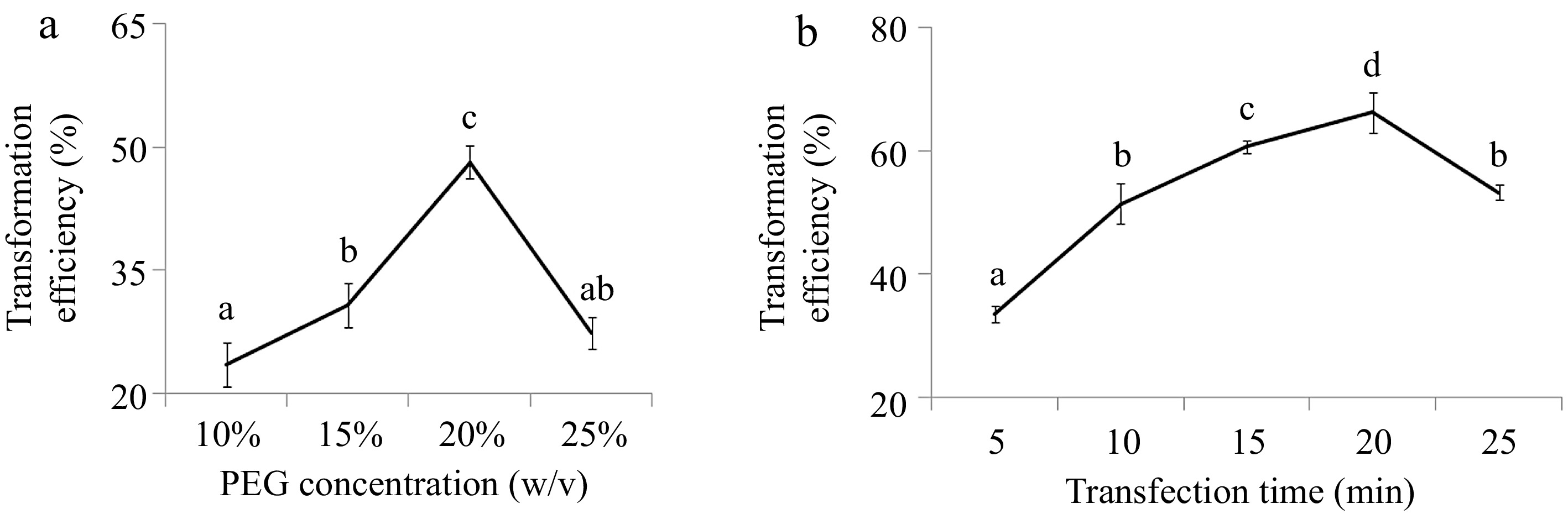
Figure 3.
Effects of PEG4000 (a) concentration and (b) transfection time on protoplast transformation efficiency in greater yam. The different letters represent significant differences (p ≤ 0.05) based on the Duncan test. Three biological replicates were used for each set of digestion time. Protoplast transformation rate (%) = (fluorescent protoplast number in view/protoplast total number in view) × 100%.

Figure 4.
Transient transformation of greater yam mesophyll protoplasts with pA7-GFP plasmid. (a) Image of GFP. (b) Bright field image of protoplasts. (c) Merged image (Scale bar = 75 μm).
Subcellular localization of GFP-fused DaERF protein in mesophyll protoplasts
-
We identified a gene member –DaERF2- belonging to ethylene responsive factor gene family in previous study[23]. DaERF2 located on the D. alata genomic contig CZHE02000125.1 and has three exons. The coding sequence (CDS) of DaERF2 is 1,140 bp. We cloned the full-length CDS of this gene in the same frame of a green fluorescence protein in plasmid pA7-GFP. Subcellular localization of this GFP-fused transcription factor protein – DaERF2 was used to test the rationality of the protoplast transient expression system. After gene expression induction in the dark for 12−18 h after transfection (20% PEG and 20 min treatment), strong green fluorescence was distributed intracellularly. GFP-DaERF2 protein trends to show bright signal in cell nucleus, but the signal for control vector (GFP) was observed in both nucleus and cytoplasm (Fig. 5).
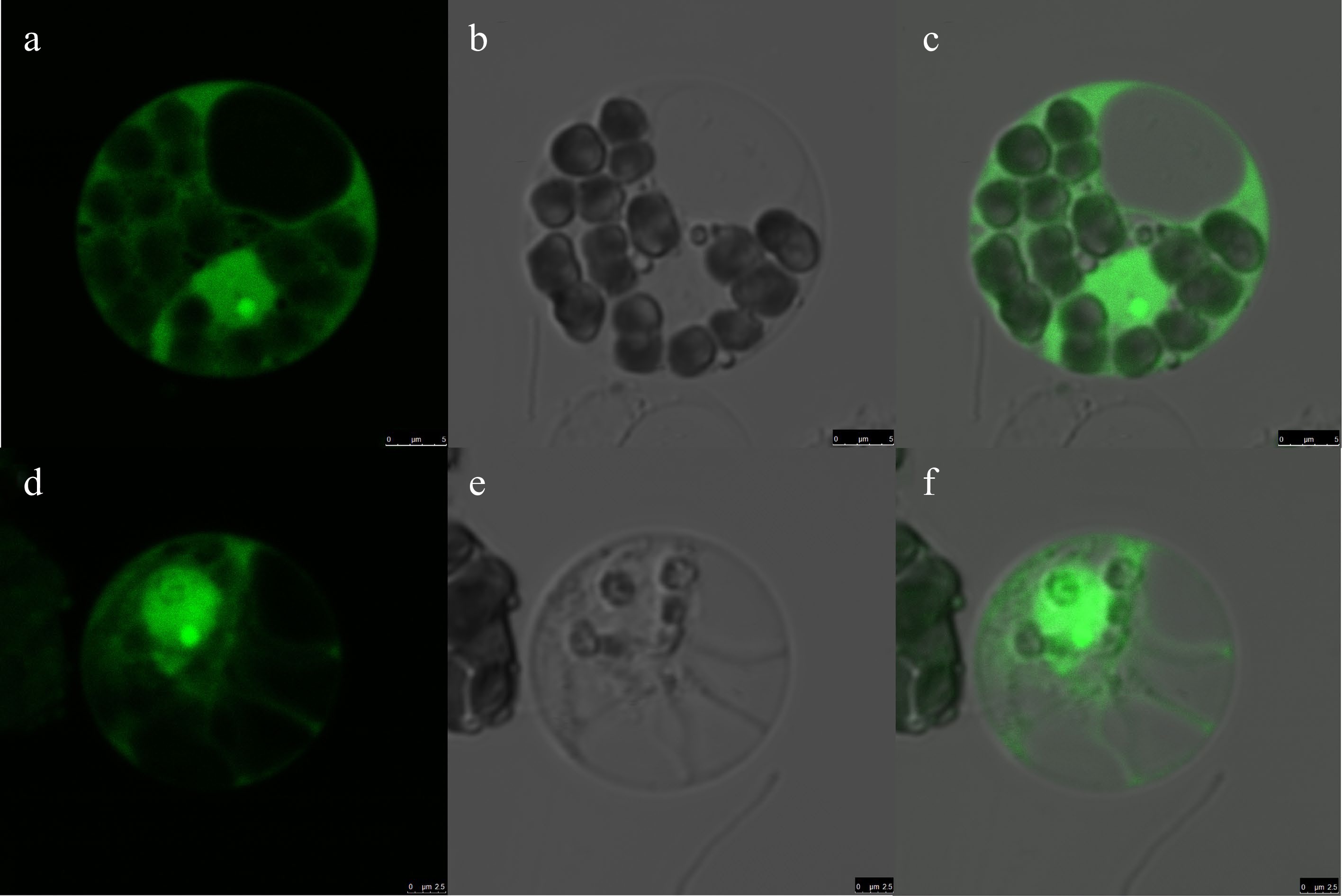
Figure 5.
Subcellular localization of DaERF1 in greater yam mesophyll protoplasts. Transient expression of GFP, showing that the GFP is distributed both in the nucleus and cytoplasm (a−c, Scale bar = 5 μm). The transient expression of GFP-fused DaERF1 protein, showing that the GFP-fused DaERF1 protein is likely localized to the nucleus (d−f, Scale bar = 2.5 μm). The laser-scanning confocal microscopy images are the (a), (d) bright field image, (b), (e) fluorescence image and (c), (f) merged image, respectively.
-
Efficient and repeatable protoplast isolation protocol is required for transient gene expression and different gene functional experiments related to transient transformation. A series of factors, such as plant condition, source tissue, the type and concentration of enzymes, are critically affecting the process of protoplast isolation. Comparing with other growth conditions, plants grown from tissue culture without thick cuticles were considered as better materials for protoplast isolation[25]. In yam, Tor et al. used young in vitro leaves (< 1.0 cm in length) which were collected from nodal segment cultures[22]. In our study, we chose tissue cultured yam seedlings as starting materials for protoplast preparation.
Suitable digestion time, digestion composition and mannitol concentration are important for protoplast isolation and viability maintenance. Tor et al. had established a protocol for yam protoplast isolation with enzyme mixtures for 15−18 h and a mannitol concentration of 0.7 M[22]. A good preparation yields approximately 107 protoplasts per gram fresh weight[13]. We optimized digestion time and the combinations of cellulase and macerozyme, which were widely used for protoplast extraction[13,25−29]. Our results showed that 0.8% cellulase R10 and 1% macerozyme R10 in the enzyme mixture obtained the highest yield (3.7 × 107 protoplasts/g FW) and the highest viability of protoplasts (92.1%) (Table 1). The greater yam leaf tissues in lower concentration of cellulose R-10 (0.4%) were still stable and more mechanical damage happened during protoplast isolation, while leaf tissues were fragile when cellulose R-10 concentration increased to 1.2%, both of which caused more damage for protoplast than that of suitable enzyme concentration. The maximum yield and viability of leaf mesophyll protoplasts achieved in our study were substantially higher than the results previously reported[22]. Moreover, digestion time is much shorter in our study (6–7 h), which make the process of protoplast isolation more efficient (Fig. 2).
Protoplasts are used for different assays with different numbers of viable protoplasts, such as 103–4 protoplasts for reporter enzyme assays, 104–5 protoplasts for protein labeling and immunoprecipitation or western blot analysis and 106 protoplasts for RNA extraction and microarray analysis[13]. Moreover, when transfection ratio is over 50%, the transient expression results from protoplasts will be reliable and repeatable[13]. In this study, an efficient greater yam protoplast transfection system was developed by selecting the best PEG4000 concentration and transfection time. The highest transfection efficiency (66.2%) was obtained with 20% of PEG4000 concentration for 20 min of transfection time (Fig. 3). Using the transfection protocol that we optimized, subcellular localization of DaERF2 was conducted. Our results showed that DaERF2 have been likely localized to the greater yam nucleus and the greater yam protoplast system was workable for further gene functional research in greater yam.
-
The authors confirm contribution to the paper as follows: experiment performed: Zhang J, Mei P, and Liu M; draft manuscript preparation: Xia W and Zhang J; data analysis: Wang Y and Xing D; study conception and design, draft manuscript preparation: Xia W, Huang D, and Wu W. All authors have read and approved the manuscript.
-
The ERF gene sequence used in this study is available in the national center for biotechnology information website (
www.ncbi.nlm.nih.gov ), the gene accesion number is HO811460.1 Thank you for technical help in protoplast isolation from Yunhui Hong and Yueyun Pan in Hainan University. The work was supported by 2019 Hainan basic and applied basic research program (Natural Science Field) high level talent project (2019RC157 and 2019RC116).
-
The authors declare that they have no conflict of interest.
-
Received 28 August 2023; Accepted 7 October 2023; Published online 31 October 2023
-
# These authors contributed equally: Jinlan Zhang, Pengfei Mei
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang J, Mei P, Wang Y, Liu M, Xing D, et al. 2023. Efficient mesophyll protoplast isolation and PEG-mediated transient gene expression in greater yam (Dioscorea alata). Tropical Plants 2:19 doi: 10.48130/TP-2023-0019 

Efficient mesophyll protoplast isolation and PEG-mediated transient gene expression in greater yam (Dioscorea alata)
- Received: 28 August 2023
- Accepted: 07 October 2023
- Published online: 31 October 2023
Abstract: Greater yam (Dioscorea alata) provides staple food for more than 100 million people in tropical and sub-tropical countries. Developing a transient transformation platform is critical for gene function analysis, since huge amounts of genomic and transcriptomic data for greater yam has recently become available. In this study, we determined the best enzyme combination of 0.8% cellulase and 1% macerozyme and 6 h of digestion to get greater yam protoplast output as 3.7 × 107 protoplasts per gram of fresh leaf and 92% viable protoplasts. PEG-mediated transient transformation efficiency for greater yam was 66.2% with optimized polyethylene glycol (PEG) concentration (20%) for 20 min. Using this protocol, the subcellular location of a transcription factor in greater yam - DaERF2 was specifically in nuclei. This efficient protoplast isolation and transformation protocol provides a feasible system for protein subcellular localization, and many other molecular assays in characterizing gene functions.
-
Key words:
- Greater yam /
- Protoplast isolation /
- Polyethylene glycol /
- Transient transformation